一种藻类垂向非均匀水体的遥感反射比校正方法与流程
本发明涉及一种藻类垂向非均匀水体的遥感反射比校正方法。
背景技术:
我国内陆湖泊中的浮游植物主要包括:硅藻门、绿藻门、蓝藻门、隐藻门、金藻门、裸藻门、甲藻门和黄藻门(余涛,2010)。每种藻类都有各自对生存环境的要求。对于富营养化湖泊(如巢湖、太湖等)而言,水体中的浮游植物门类主要为蓝藻门、绿藻门和硅藻门,通常达90%以上,尤其是夏季基本为蓝藻门(成芳,2010;姜霞等,2010)。藻类在水体中运动形态主要包括悬浮、上浮和游泳。我国巢湖、太湖中的绿藻门和硅藻门的藻类主要以悬浮形态存在,改变其空间位置则完全依赖于水动力条件(胡鸿钧和魏心印,2006)。与只能靠风速、水动力等外界条件实现垂向迁移的绿藻和硅藻不同,蓝藻独特的生理结构和浮力调节能力是其成为优势种群的必要条件。
蓝藻改变其垂向位置的主要动力来自于自身的浮力以及水动力条件。蓝藻的浮力调节能力来自于细胞内的气囊(伪空泡/伪空胞),具有伪空胞的蓝藻根据水体中光照、营养盐的垂向分布情况,改变藻细胞浮力大小,使得它们在水体中上下运动(成慧敏等,2006;安强等,2012)。伪空胞结构有利于蓝藻依据其自身的生理节律和生理状态,以及外环境变化,通过上浮或下沉来选择最佳的生长和生存空间(susanaandphlips,1992)。湖泊中水深方向的紊流作用是推动蓝藻垂向迁移的主要外力。当地的气候、气象条件以及出/入湖河流都是影响湖泊垂向水动力条件的因素(cullenandeppley,1981)。当风速大于临界风速时,风引起的波浪、扰动等共同作用使得藻颗粒在水体中上下混合(朱永春和蔡启铭,1997;杨正健,2010;孔繁翔和宋立荣,2011)。但当风速较小(<3m/s)、水动力趋缓的环境下,蓝藻自身的浮力调节能力是其进行上浮或下沉的主要动力来源,蓝藻细胞的趋光性促使其迅速地上浮至水体表面形成蓝藻聚集体(水华)(齐雨藻等,1997;唐汇娟等,2003)。
日常遥感监测发现,藻华面积会在短时间内(几天甚至数小时内)发生较大的变化。例如,goci数据监测中国东海藻华面积发现,受潮汐或洋流的影响一天之内藻华面积会产生大于100%的误差(louandhu,2014)。事实上,富营养化湖泊中的藻华暴发现象,往往不是短时间内生物量的急剧增加,而是水体内已经富集的藻颗粒的垂向非均匀分布(caoetal.,2006)。也就是说,外界水动力或环境因子的变化使得藻颗粒在水体中上下迁移,改变了藻类的垂向分布结构,从而引起表面上看上去的藻华暴发或消失(beaveretal.,2013;blottièreetal.,2013;ndongetal.,2014)。藻类垂向结构的变化使得只监测水表面藻华不能反映整个水体的富营养化状况。而且,这会影响水体光学参数遥感反演的精度(stramskaandstramski,2005)和色素生物量的估计(silulwaneetal.,2010)。
由此可见,作为富营养化湖泊的优势藻种——蓝藻因其独特的生理结构(伪空胞)以及外环境水动力条件,通过上浮或下沉来选择其最佳的生长和生存空间,客观上造成了藻类在水体中的垂向不均匀分布,客观上与传统湖泊水色遥感的垂向均一的假设不符。
遥感探测到的信号不仅包括水体表层的信息,还反映了一定深度内水下光场的结构,遥感反射比对水体光学组分的垂向非均匀分布具有一定的响应。目前,传统的生物光学模型和水色参数的现场测量及使用大都假设水体垂向均一,不考虑水体中藻类的垂向非均匀分布。实际上,不考虑水色参数的垂向非均匀分布会在水体遥感反射比及水色参数反演算法中引起很大误差(ballestero,1999;nanuandrobertson,1993;sathyendranathandplatt,1989)。研究表明,一类水体中不考虑叶绿素a浓度的垂向非均匀分布,可以对遥感反射比造成大于5%的误差,甚至可能超过70%。近表层水体的水体光学特性变化越大,并且衰减系数越小时,下层水体对遥感反射比的影响越大(stramskaandstramski,2005;xiuetal.,2008)。藻类垂向非均匀分布影响了遥感反射比光谱的大小及光谱形状(kutseretal.,2008)。在i类水体中,叶绿素a浓度分布曲线为高斯分布情况下,chla的高斯峰值减少了蓝光波段的遥感反射比,增加了绿光波段的遥感反射比(席颖等,2010)。在赤潮多发区,水柱内平均叶绿素a一定的情况下,垂向非均匀条件下的遥感反射比要高于垂向均匀的情况,叶绿素a浓度垂向结构参数的变化主要影响绿光波段,对蓝、红波段影响可忽略(王云飞和贺明霞,2010)。
sathyendranath等(1989)研究表明,在假设叶绿素a浓度垂向分布均一条件下,光学深度zp和真光层ze内平均叶绿素a的相对误差百分比最大可超过100%。传统的基于蓝绿波段比值的叶绿素a浓度反演算法可引起7倍的高估(stramskaandstramski,2005)。在蓝藻水华暴发的水体,叶绿素浓度的垂向分布会造成红/近红外波段比值的叶绿素a反演算法2–6倍的高估(kutseretal.,2008)。马孟枭等(2014)利用辐射传输模拟软件hydrolight分析了藻类垂向分布呈高斯函数时藻类垂向非均匀分布对波段比值叶绿素a算法的影响,发现靠近水体表面的藻类垂向非均匀分布使得算法失效。此外,国内外一些学者利用统计回归方法(hidalgo-gonzálezandalvarez-borrego,2001;
水体中水色参数的组成、垂向分布、水体固有光学特性等共同影响了藻类垂向非均匀分布对遥感反射比的影响程度。传统的水质参数反演算法忽略了藻类垂向非均匀分布,往往通过表层几十厘米的混合样或者单一水层的数据建立算法。藻类垂向非均匀分布影响了水体遥感反射比,进而会引起基于垂向均一假设得到反演产品的高估或低估。在不同的像元或不同采样点,这种影响是不一致的,因此加剧了这种影响的不确定性。在富营养化湖泊,无机颗粒物和cdom含量较高,来源复杂,与叶绿素a不共变。因此,在研究藻类垂向分布时,如何控制悬浮物和cdom两个变量也增加了研究的难度。
参考文献
ballestero,d.1999.remotesensingofverticallystructuredphytoplanktonpigments.top.meteor.oceanogr,6(1):14-23.
beaver,j.r.,d.a.casamatta,t.l.east,k.e.havens,a.j.rodusky,r.t.james,c.e.tausz,andk.m.buccier.2013.extremeweathereventsinfluencethephytoplanktoncommunitystructureinalargelowlandsubtropicallake(lakeokeechobee,florida,usa).hydrobiologia,709(1):213-226.
bengio,y.,andy.grandvalet.2003.nounbiasedestimatorofthevarianceofk-foldcross-validation.journalofmachinelearningresearch,5(3):1089-1105.
binding,c.,j.jerome,r.bukata,andw.booty.2008.spectralabsorptionpropertiesofdissolvedandparticulatematterinlakeerie.remotesensingofenvironment,112(4):1702-1711.
blondeau-patissier,d.,v.e.brando,k.oubelkheir,a.g.dekker,l.a.clementson,andp.daniel.2009.bio-opticalvariabilityoftheabsorptionandscatteringpropertiesofthequeenslandinshoreandreefwaters,australia.journalofgeophysicalresearch,114(c5):1289-1301.
blottière,l.,m.rossi,f.madricardo,andf.d.hulot.2013.modelingtheroleofwindandwarmingonmicrocystisaeruginosabloomsinshallowlakeswithdifferenttrophicstatus.theoreticalecology,7(1):35-52.
cao,h.,f.kong,l.luo,x.shi,z.yang,x.zhang,andy.tao.2006.effectsofwindandwind-inducedwavesonverticalphytoplanktondistributionandsurfacebloomsofmicrocystisaeruginosainlaketaihu.journaloffreshwaterecology,21(2):231-238.
cullen,j.j.,andr.w.eppley.1981.chlorophyllmaximumlayersofthesoutherncaliforniabightandpossiblemechanismsoftheirformationandmaintenance.oceanologicaacta,4(1):23-32.
hidalgo-gonzález,r.m.,ands.alvarez-borrego.2001.chlorophyllprofilesandthewatercolumnstructureinthegulfofcalifornia.oceanologicaacta,24(1):19-28.
kutser,t.,l.metsamaa,anda.g.dekker.2008.influenceoftheverticaldistributionofcyanobacteriainthewatercolumnontheremotesensingsignal.estuarine,coastalandshelfscience,78(4):649-654.
lou,x.,andc.hu.2014.diurnalchangesofaharmfulalgalbloomintheeastchinasea:observationsfromgoci.remotesensingofenvironment,140:562-572.
nanu,l.,andc.robertson.1993.theeffectofsuspendedsedimentdepthdistributiononcoastalwaterspectralreflectance:theoreticalsimulation.internationaljournalofremotesensing,14(2):225-239.
ndong,m.,d.bird,t.nguyen-quang,m.l.deboutray,a.zamyadi,b.vincon-leite,b.j.lemaire,m.prevost,ands.dorner.2014.estimatingtheriskofcyanobacterialoccurrenceusinganindexintegratingmeteorologicalfactors:applicationtodrinkingwaterproduction.waterresearch,56:98-108.
pitarch,j.,d.odermatt,m.kawka,anda.wüest.2014.retrievalofverticalparticleconcentrationprofilesbyopticalremotesensing:amodelstudy.opticsexpress,22(103):a947-a959.
susana,a.,ande.j.phlips.1992.lightabsorptionbycyanobacteria:implicationsofthecolonialgrowthform.limnology&oceanography,37(2):434-441.
sathyendranath,s.,andt.platt.1989.remotesensingofoceanchlorophyll:consequenceofnonuniformpigmentprofile.appliedoptics,28(3):490-495.
sathyendranath,s.,t.platt,c.m.caverhill,r.e.warnock,andm.r.lewis.1989.remotesensingofoceanicprimaryproduction:computationsusingaspectralmodel.deepsearesearchparta.oceanographicresearchpapers,36(3):431-453.
silulwane,n.f.,a.j.richardson,f.a.shillington,andb.a.mitchell-innes.2010.identificationandclassificationofverticalchlorophyllpatternsinthebenguelaupwellingsystemandangola-benguelafrontusinganartificialneuralnetwork.southafricanjournalofmarinescience,23(1):37-51.
stramska,m.,andd.stramski.2005.effectsofanonuniformverticalprofileofchlorophyllconcentrationonremote-sensingreflectanceoftheocean.appliedoptics,44(9):1735-1747.
susana,a.,ande.j.phlips.1992.lightabsorptionbycyanobacteria:implicationsofthecolonialgrowthform.limnology&oceanography,37(2):434-441.
安强,龙天渝,刘春静等.雷诺数对藻类垂向分布特性的影响[j].湖泊科学,2012,05:717-722.
成芳.太湖水体富营养化与水生生物群落结构的研究[d].2010,苏州:苏州大学.
成慧敏,邱保胜.蓝藻的伪空泡及其对蓝藻在水体中垂直分布的调节[j].植物生理学通讯,2006,05:974-980.
冯春晶.基于人工神经网络的海中叶绿素浓度垂直分布特征研究[d].2004,青岛:中国海洋胡鸿钧,魏心印.中国淡水藻类——系统、分类及生态[m].2006,北京:科学出版社.
姜霞,王书航,钟立香等.巢湖藻类生物量季节性变化特征[j].环境科学,2010,09:2056-2062.
孔繁翔,宋立荣.蓝藻水华形成过程及其环境特征研究[m].2011,北京:科学出版社.
刘斌.一类水域中从海面反射比反演叶绿素浓度垂直分布初探[d].2009,中国海洋大学.
马孟枭,张玉超,钱新等.巢湖水体组分垂向分布特征及其对水下光场的影响[j].环境科学,2014,05:1698-1707.
齐雨藻,黄长江,钟彦等.甲藻塔玛亚历山大藻昼夜垂直迁移特性的研究[j].海洋与湖沼,1997,05:458-467.
唐汇娟,谢平,陈非洲.微囊藻的昼夜垂直变化及其迁移[j].中山大学学报(自然科学版),2003,s2:236-239.
王云飞,贺明霞.东海赤潮多发区非均匀叶绿素浓度剖面对遥感反射比的影响[j].中国海洋大学学报(自然科学版),2010,10:100-104.
席颖,杜克平,张丽华等.叶绿素浓度垂直不均一分布对于分层水体表观光学特性的影响[j].光谱学与光谱分析,2010,02:489-494.
杨正健.基于藻类垂直迁移的香溪河水华暴发模型及三峡水库调控方案研究[d].2010,宜昌:三峡大学.
余涛.巢湖浮游植物群落结构研究[d].2010,合肥:安徽大学.
朱永春,蔡启铭.风场对藻类在太湖中迁移影响的动力学研究[j].湖泊科学,1997,02:152-158.
技术实现要素:
本发明的目的在于提供一种藻类垂向非均匀水体的遥感反射比校正方法,把藻类垂向非均匀水体的遥感反射比等效为具有相同平均藻类浓度的垂向均一水体的遥感反射比,消除传统水质参数反演模型在应用中的误差。
本发明的上述目的通过独立权利要求的技术特征实现,从属权利要求以另选或有利的方式发展独立权利要求的技术特征。
为达成上述目的,本发明所采用的技术方案如下:
一种藻类垂向非均匀水体的遥感反射比校正方法,包括:建立藻类不同垂向分布类型的δrrs与藻类垂向分布函数的结构参数之间的定量关系式;构建不同藻类垂向分布类型的遥感反射比校正算法,把垂向非均匀水体的遥感反射比等效为垂向均匀水体的遥感反射比。
作为进一步的示例,前述方法的具体实现包括:
1)建立藻类不同垂向分布类型的δrrs与藻类垂向分布的结构参数之间的定量关系;
其中,所述δrrs指藻类不同垂向分布类型水体的垂向非均匀条件下的遥感反射比rrs-v与对应的垂向均匀条件下的遥感反射比rrs-h的相对误差;
2)建立藻类垂向非均匀水体的遥感反射比校正算法;
根据步骤1)建立的定量关系,构建不同藻类垂向分布类型的遥感反射比校正算法,将垂向非均匀水体的遥感反射比等效为垂向均匀水体的遥感反射比。
基于前述步骤和方法,将遥感反射比校正算法应用至实测高光谱数据,基于前述的反演算法方法,对不同藻类垂向分布类型的遥感反射比校正后应用于反演算法,以评价基于实测数据的遥感反射比校正算法的效果。
由以上本发明的技术方案可知,本发明的藻类垂向非均匀水体的遥感反射比校正方法,基于水体光学遥感基本原理,提出藻类垂向非均匀水体的遥感反射比校正方法,通过藻类垂向非均匀水体的遥感反射比与藻类垂向均匀水体的遥感反射比、及结构参数之间的定量关系,来实现对藻类垂向非均匀水体的遥感反射比校正,改变了传统湖泊水色遥感基于藻类垂向均一分布的假设前提,更加准确的获取水体遥感反射比这一关键参数。藻类垂向非均匀水体的遥感反射比校正,消除了藻类垂向非均匀分布对遥感反射比的光谱形状和大小的影响,定量揭示水体遥感反射比对藻类垂向分布的响应,有助于提高水质参数反演算法的精度,为利用卫星遥感手段反演巢湖蓝藻总量提供理论基础,为富营养化湖泊的水环境管理和决策提供重要的科研支持。
应当理解,前述构思以及在下面更加详细地描述的额外构思的所有组合只要在这样的构思不相互矛盾的情况下都可以被视为本公开的发明主题的一部分。另外,所要求保护的主题的所有组合都被视为本公开的发明主题的一部分。
结合附图从下面的描述中可以更加全面地理解本发明教导的前述和其他方面、实施例和特征。本发明的其他附加方面例如示例性实施方式的特征和/或有益效果将在下面的描述中显见,或通过根据本发明教导的具体实施方式的实践中得知。
附图说明
附图不意在按比例绘制。在附图中,在各个图中示出的每个相同或近似相同的组成部分可以用相同的标号表示。为了清晰起见,在每个图中,并非每个组成部分均被标记。现在,将通过例子并参考附图来描述本发明的各个方面的实施例,其中:
图1是δrrs(λ)与叶绿素a结构参数关系式的系数。
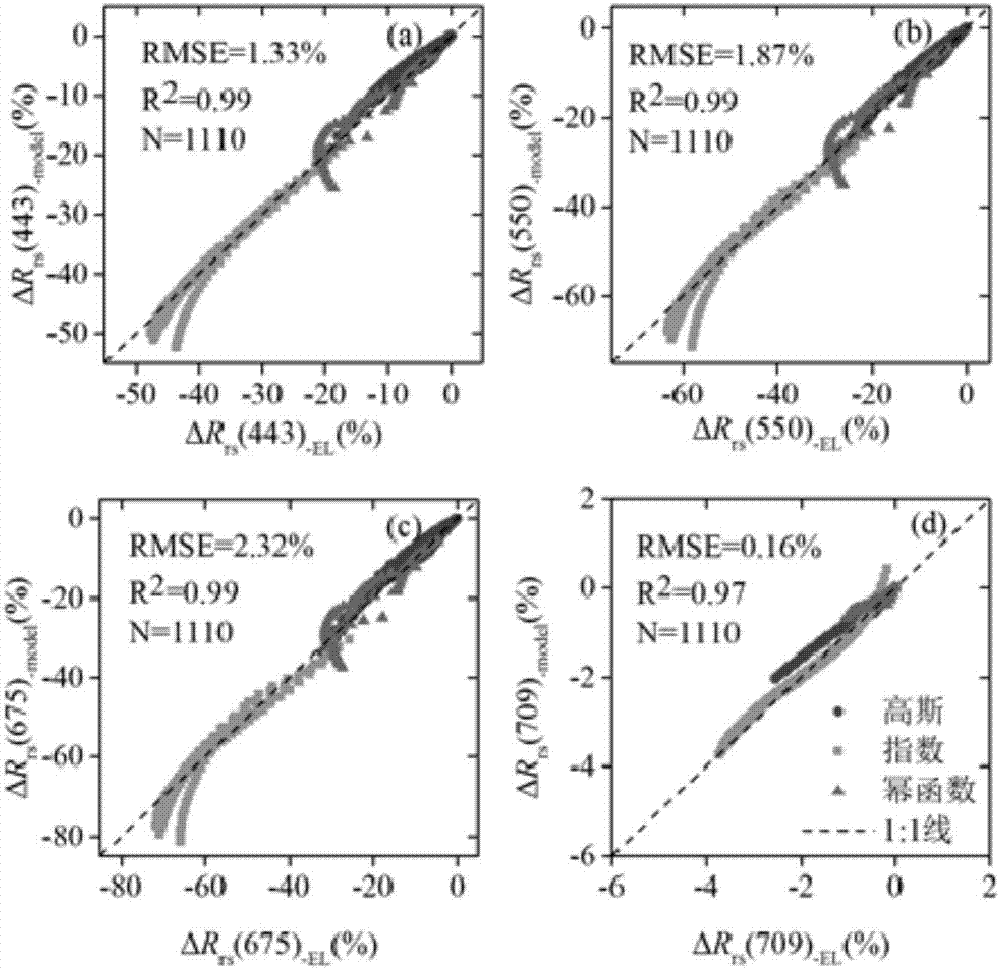
图2是在443、550、675、710nm波段处模型计算得到的δrrs与模拟数据的δrrs的散点图。
图3是遥感反射比校正算法应用于实测高光谱数据流程图。
图4是(a)不同类型下的实测叶绿素a浓度chla-insitu与反演得到的叶绿素a浓度chla-model的散点图;(b)不同类型下的遥感反射比校正前(δ-model)、后(δ-corrected)的反演得到的叶绿素a浓度与实测叶绿素a浓度的相对误差百分比曲线。
图5是spim和ag(440)变化对δrrs影响的敏感性分析图。
前述图示1-5中,作为英文形式表达的各坐标、标识或其他表示,均为本领域所公知的,并不在本例中再做赘述。
具体实施方式
为了更了解本发明的技术内容,特举具体实施例并配合所附图式说明如下。
在本公开中参照附图来描述本发明的各方面,附图中示出了许多说明的实施例。本公开的实施例不必定义在包括本发明的所有方面。应当理解,上面介绍的多种构思和实施例,以及下面更加详细地描述的那些构思和实施方式可以以很多方式中任意一种来实施,这是因为本发明所公开的构思和实施例并不限于任何实施方式。另外,本发明公开的一些方面可以单独使用,或者与本发明公开的其他方面的任何适当组合来使用。
本实施例基于实测高光谱数据对藻类垂向非均匀水体的遥感反射比进行校正,实现方式如下:根据实测叶绿素a浓度垂向分布数据的藻类垂向分布类型及结构参数,设置模拟方案,利用ecolight辐射传输模拟手段,模拟大规模数据集。然后,建立不同藻类垂向分布类型的δrrs与其结构参数之间的关系,把藻类垂向非均匀水体的遥感反射比等效为具有相同平均叶绿素a浓度的垂向均一水体的遥感反射比。建立不同藻类垂向分布类型的遥感反射比校正算法;利用预留的ecolight模拟数据进行了验证;最后,利用实测高光谱数据对算法进行验证。
作为示例性的描述,下面结合附图所示,对前述方法的实施进行具体说明。
步骤1、基于ecolight模拟数据集,建立藻类不同垂向分布类型的δrrs与藻类垂向分布的结构参数之间的定量关系;
本实施例中采用叶绿素a垂向分布类型指代藻类垂向分布类型,建立叶绿素a不同垂向分布类型的δrrs与叶绿素a垂向分布的结构参数之间的定量关系;
根据三种类型的叶绿素a垂向非均匀分布(公式1-3,fi;pi为结构参数)的rrs-v(λ)与垂向均一的rrs-h(λ),分别计算δrrs(λ)(公式4),其中i=1、2、3分别代表高斯、指数、幂函数类型);
f2(z)=m1×exp(m2×z),p2=[m1,m2](2)
式中z为水深;c0为叶绿素背景浓度值;σ为标准偏差;h为高斯分布类型的结构参数;m1、m2为指数分布类型的结构参数;n1、n2为幂函数类型的结构参数。
式中rrs-v-i(λ)指在波长λ处,藻类垂向分布类型i的垂向非均匀条件下的水体遥感反射比,rrs-h-i(λ)指在波长λ处,藻类垂向分布类型i的垂向均匀条件下的水体遥感反射比,i=1,2,3分别指高斯、指数和幂函数分布类型。
通过分析不同叶绿素a垂向分布类型下的结构参数对δrrs(λ)的影响,建立400-900nm间隔5nm的δrrs-i(λ)与chla(z)结构参数pi的经验关系模型(公式5-7),每种类型的系数如图1;
高斯类型:
指数类型:δrrs-2(λ)=b1(λ)ln(m1)+b2(λ)m2+b3(λ)(6)
幂函数类型:δrrs-3(λ)=c1(λ)n1+c2(λ)n2+c3(λ)(7)
式中,δrrs-i(λ)指在波长λ处,垂向非均匀条件下的遥感反射比与对应的垂向均匀条件下的遥感反射比的相对误差;i=1,2,3分别指高斯、指数、幂函数分布类型下,δrrs(λ)的数值。
2、建立不同藻类垂向分布类型的遥感反射比校正算法
在已知chla(z)垂向结构参数的情况下,根据前两步的关系式,得到δrrs-i(λ),把叶绿素a垂向非均匀的rrs-v-i(λ)校正为叶绿素a垂向均一的等效rrs-c-i(λ);根据chla(z)参数结构pi和叶绿素a垂向非均匀的遥感反射比,最终形成遥感反射比的校正算法,校正算法如公式(8)所示:i=1、2、3。
3、利用预留的ecolight模拟数据进行验证
利用以上关系模型,利用随机预留的模拟数据的1/4进行验证,通过4个波段的ecolight模拟数据计算得到的δrrs-el与根据chla(z)结构参数计算得到的δrrs-model的散点图比较(图2),可以发现验证结果较好,r2在0.97以上,rmse(log)小于3%。
δrrs-el根据预留模拟数据的4个波段的rrs-v与rrs-h的值,根据公式(4),计算得到。
δrrs-mode由每种藻类垂向分布类型的结构参数分别带入公式(5-7),计算得到。
在没有进行藻类垂向异质水体的遥感反射比校正时,δrrs<5%的数据只占41.2%,δrrs>50%的数据占10.8%。进行遥感反射比校正之后,δrrs<5%的数据占91.1%,只有1.0%的数据δrrs>10%。高斯、指数、幂指数的平均误差分别为1.6%、2.8%、3.7%,rmse(log)为0.011、0.018、0.024。因此,遥感反射比校正模型对三种藻类垂向分布类型的模拟数据校正效果较好。
4、获取基于实测高光谱数据的遥感反射比校正结果
1)建立藻类垂向均一条件下的遥感反射比与叶绿素a的反演算法;
利用藻类垂向均一类型的数据建立的波段比值的叶绿素a浓度反演算法如下:
x=rrs(709)/rrs(675),
chla-model=13.49x2+107.95x-80.85.(9)
2)把藻类垂向非均一条件下(高斯、指数、幂函数)的遥感反射比带入上述算法,计算得到的叶绿素a浓度是用传统方法反演的结果chla-model;
3)把利用遥感反射比校正模型校正后的rrs-c(λ)带入上述算法,计算得到的叶绿素a浓度为校正后的叶绿素a浓度(chla-corrected)(图3)。把chla-model、chla-corrected与实测的叶绿素a浓度chla-insitu进行比较,计算他们的r2、rmse(log)相对误差(re=(chla-model/corrected-chla-insitu)/chla-insitu*100%),验证遥感反射比校正模型的有效性。
比较实测的均匀水体叶绿素a浓度chla-insitu和模型计算得到的叶绿素a浓度chla-model发现,两者的r2为0.64,相对误差re为2.7%,rmse(log)为0.07。藻类垂向非均匀水体(高斯、指数、幂函数)的chla-insitu和chla-corrected的平均相对误差re从校正前的53.5%,下降到了校正后的14.8%(图4)。这与前面的结论一致,即藻类垂向非均匀分布会引起根据垂向均一数据建立的叶绿素a浓度反演结果的高估,遥感反射比的校正可以明显地降低这种高估的情况。
接下来,进行了无机悬浮物spim和cdom吸收系数ag(440)变化对δrrs影响的敏感性分析(图5)。当chla(z)垂向分布呈高斯曲线(c0=10,h=36,σ=0.6)时,chla(z)对遥感反射比的影响程度随spim的增加而减弱,当spim从10mg/l增加到50mg/l时,443、550、675nm的δrrs大约下降5%,rrs(709)/rrs(675)大约下降8%。cdom对蓝、绿波段的遥感反射比的影响小于5%,对红光和近红外波段几乎没有影响。在第4部分利用ecolight模拟数据集时,巢湖野外测量的spim和cdom的平均值作为不变的背景值。当spim和cdom的变化较大时,会改变藻类垂向非均匀分布对遥感反射比的影响程度。因此,选取spim和cdom的平均值也是校正模型应用到野外测量数据时的误差来源之一。
虽然本发明已以较佳实施例揭露如上,然其并非用以限定本发明。本发明所属技术领域中具有通常知识者,在不脱离本发明的精神和范围内,当可作各种的更动与润饰。因此,本发明的保护范围当视权利要求书所界定者为准。
- 还没有人留言评论。精彩留言会获得点赞!