基于不完备信息的多节耳材村虫化石逆向推导复原法
本发明涉及三维重构
技术领域:
,具体涉及基于不完备信息的多节耳材村虫化石逆向推导复原法。
背景技术:
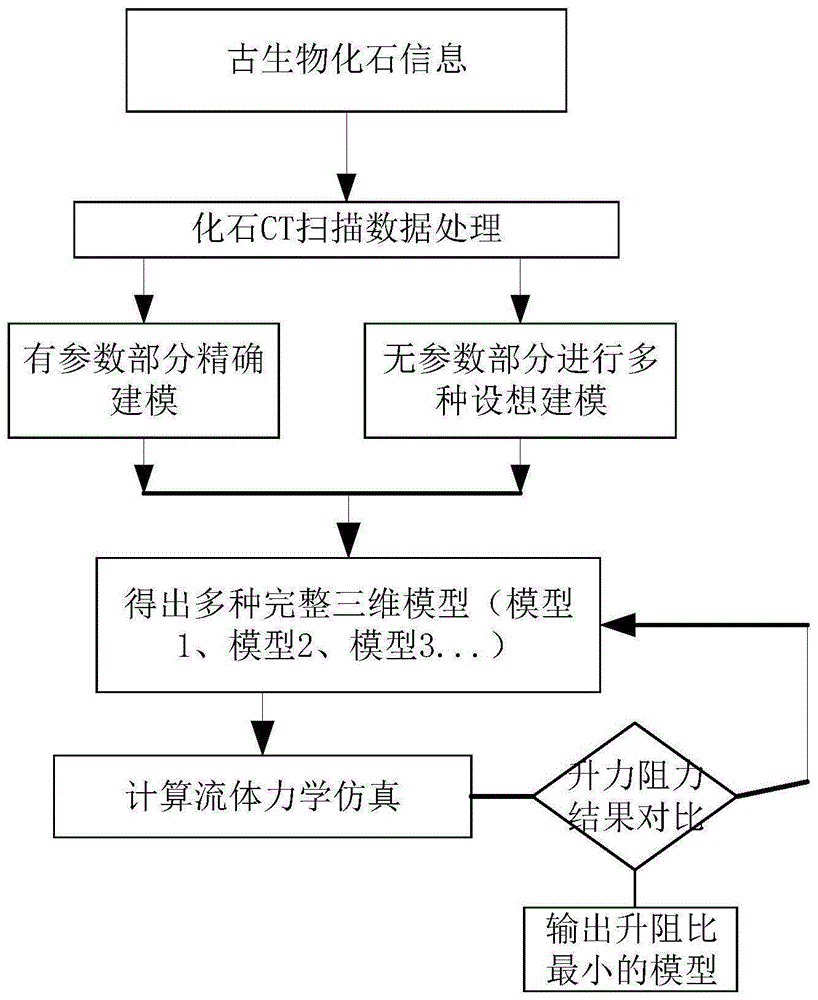
:澄江动物群是20世纪最伟大的发现之一,是寒武纪生命大爆发的直接证据,是目前地球上所发现的最古老、保存最好的一个多门类动物群,它几乎包括了现代所有的动物门类。勿容置疑,它们代表了现生动物的祖先类型,其构造特征也多为原始特征,为追溯早期动物类群的起源和研究其谱系关系提供了得天独厚的条件。三维重构技术不仅能让古生物学家对其研究对象获得更为丰富、生动的视觉效果,更为重要的是合理的三维重构模型将对仿生学的研究具有重大意义。中国云南澄江生物群的多节耳材村虫的ct扫描的原始数据不完备,导致其身体形态和壳瓣展开角度等参数不能通过化石数据获得,且目前还未有基于三维重构技术对多节耳材村虫化石进行复原的方法。技术实现要素:为了解决上述问题,本发明提供了基于不完备信息的多节耳材村虫化石逆向推导复原法,具体技术方案如下:基于不完备信息的多节耳材村虫化石逆向推导复原法,包括以下步骤:s1:根据多节耳材村虫化石的ct扫描得到原始数据,在三维建模软件中对多节耳材村虫附肢、躯干、尾叉部分进行实体建模;s2:建立多种壳瓣连接方式及展开角度和身体形态组合的多节耳材村虫完整实体模型;s3:对建立的多种完整实体模型,在计算流体力学仿真软件中进行仿真研究,每种实体模型均可得出一系列的阻力值和升力值;s4:对每个模型的升力值、阻力值进行对比分析,找出升阻比最小的对应模型,即推断该模型为多节耳材村虫的3d复原模型结果;所述升阻比为升力值与阻力值之比。优选地,所述壳瓣连接方式包括壳瓣连接状态和壳瓣分开状态。优选地,所述身体形态包括背部平直腹部凸起和背部凸起腹部平直两种状态。优选地,所述展开角度的操作具体是在最大可能展开角度和最小展开角度的范围,按n度的间隔进行展开角度建模,其中n小于最小展开角度。本发明的有益效果为:根据化石ct扫描的原始数据,利用计算机辅助设计软件对耳材村虫进行建模,由于身体形态和壳瓣展开角度等参数不能通过化石数据获得,因此对不确定参数进行多种不同形态的尝试建模,然后通过计算流体力学仿真(cfd)进行仿真研究,得出每种形态下的升力和阻力,以此推断重构的何种身体形态最为合理,最后通过实验验证该三维重构方法的有效性。本发明基于不完备信息对多节耳材村虫化石进行逆向推导复原,可以让古生物学家对其研究获得更为丰富、生动的视觉效果。本发明所涉及的三维重构方法将传统建模方法与计算流体力学仿真相结合,利用仿真结果推断建模结果的准确性,将该方法应用于古生物的3d复原,填补了耳材村虫研究上的空白。附图说明图1为本发明的流程图;图2多节耳材村虫化石的照片,其中图2(a)为多节耳材村虫化石的光学显微镜照片,图2(b)为俯视图视角的多节耳材村虫化石显微ct成像图;图2(c)为仰视图视角的多节耳材村虫化石显微ct成像图;图2(d)为侧视图视角的多节耳材村虫化石显微ct成像图;图3为耳材村虫身体轮廓线图;图4为耳材村虫身体轮廓模型;图5为附肢及尾叉轮廓线拟合图;图6为耳材村虫尾部建模效果图;其中图6(a)为短叉尾部模型图;图6(b)为长叉尾部模型图;图6(c)为尾部薄膜形成过程图;图6(d)为尾部整体模型图;图7为耳材村虫附肢建模效果图;其中图7(a)为单条腿肢模型图;图7(b)为完整腿肢模型图;图8为耳材村虫壳瓣及身体模型图;其中图8(a)为壳瓣分开状态的壳瓣模型图;图8(b)为壳瓣连接状态的壳瓣模型图;图8(c)为背部凸起腹部平直的身体模型图;图8(d)为背部平直腹部凸起的身体模型图;图9为4种完整耳材村虫模型,其中图9(a)为壳瓣分开、背部平直腹部凸起模型图;图9(b)为壳瓣分开、背部凸起腹部平直模型图;图9(c)为壳瓣连接、背部平直腹部凸起模型图;图9(d)为壳瓣连接、背部凸起腹部平直模型图;图10为壳瓣不同展开角度的模型图,其中图10(a)为壳瓣展开120度的模型图;图10(b)为壳瓣展开180度的模型图;图11为网格划分情况图;图12为四种模型在不同壳瓣展开角度的仿真结果图。具体实施方式为了更好的理解本发明,下面结合附图和具体实施例对本发明作进一步说明:如图1所示,基于不完备信息的多节耳材村虫化石逆向推导复原法,包括以下步骤:s1:根据多节耳材村虫化石的ct扫描得到原始数据,在ug三维建模软件中对多节耳材村虫附肢、躯干、尾叉部分进行实体建模;多节耳材村虫是一种具有一对外壳和鱼尾状尾部的节肢动物,发现并报道于上世纪九十年代。云南省古生物研究重点实验室研究人员使用精度达到微米级别的x射线三维成像技术(显微ct),透过厚厚的外壳和围岩,观察到了多节耳材村虫保存完好的附肢及其令人惊艳的形态细节,如图2所示。根据图2(b)、图2(c)所示的显微ct成像图,可以比较准确得到的身体形态参数有:耳材村虫身长及宽度、附肢数量与形状尺寸参数、尾叉与躯干形状尺寸参数。运用三维建模软件中的“光栅图像”功能,选择图像文件,将图2的(b)、(c)、(d)三张图片选择一个平面插入,后在光栅图片中设置插入草图的基点位置,本实施例中设置为图片的中点与原点重合,接着设置插入图片的宽度和高度。选择一个基准平面绘制草图,进入草图绘制状态,利用“艺术样条”命令,类型为“通过点”,沿着耳材村虫的轮廓往一个方向设置其余的点,终点位置设置在身体与尾部分叉交界处,最终构建的耳材村虫身体轮廓如图3所示。绘制好横截面草图后,利用“通过曲线网格”命令,主曲线选择刚绘制的耳材村虫身体轮廓线草图,交叉曲线依次选择身体轮廓框架线,如图4(a)所示。身体的其他部位也可用此方法来构建,注意保证曲面曲率的连续性,最后利用曲面缝合命令来形成实体,最终耳材村虫身体轮廓模型如图4(b)所示。与建立耳材村虫身体轮廓线的步骤一致,建立尾部和腿肢的拟合轮廓线,如图5所示。接着在尾部框线顶点处建立基准面,并且绘制截面草图,利用“扫掠”命令建立尾部模型,利用“n边曲面”命令形成尾部薄膜曲面,经过修剪后用“加厚”命令形成曲面分叉薄膜,经过修饰后利用“镜像几何体”命令建立尾部整体模型,建模效果如图6所示。同理,在轮廓线两端建立基准面,并且绘制草图,而后利用“扫掠”命令形成腿肢,如图7(a),再利用同样的方法以及阵列的方法形成其他腿肢,经过简单修饰后腿肢整体如图7(b)所示。s2:由于化石的压缩和残缺,导致耳材村虫的壳瓣形状和身体形态无法获得,因此建立多种壳瓣连接方式及展开角度和身体形态组合的多节耳材村虫完整实体模型;壳瓣连接方式包括壳瓣连接状态和壳瓣分开状态。身体形态包括背部平直腹部凸起和背部凸起腹部平直两种状态。展开角度的操作具体是在最大可能展开角度和最小展开角度的范围,按n度的间隔进行展开角度建模,其中n小于最小展开角度。在本实施例中由于耳材村虫化石是在挤压状态下保存下来的,而且残缺比较严重,导致厚度参数、身体部分轮廓形状以及壳瓣形状和展开角度均无法获得,因此对于耳材村虫完整的三维重构,无法只依赖于化石ct扫描数据,只能通过大致的轮廓形状和生物形态学进行多种形态的尝试性建模,根据对现生生物丰年虾和鲎虫的观察和生物形态学常识,对两壳瓣连接方式和身体轮廓厚度凸起形态分别作两种极限形态假设。如图8(a)所示为壳瓣连接状态和壳瓣分开状态两种不同连接方式的壳瓣模型,图8(b)为背部平直腹部凸起和背部凸起腹部平直两种不同轮廓形状的身体模型,然后将各部分进行组合,得出4种完整模型,如图9所示。根据生物学研究推断,壳瓣在耳材村虫背部是固定的,不能像鸟类翅膀一样挥动。上述研究共建立了4种完整的耳材村虫模型,但两壳瓣的展开角度仍无法确定,因此根据理论上最大可能展开角度(220度)和最小展开角度(60度)的范围,按10度的间隔,将每种模型分别建立15种不同壳瓣展开角度的模型。壳瓣展开角度定义方式:两翅膀圆弧切线的夹角。例如如图10所示为壳瓣展开120度和180度时的模型。将以上模型进行组合,共得到下表所示的60种实体模型。如下表1所示。表1建模情况表s3:对建立的多种完整实体模型,在计算流体力学仿真软件中进行仿真研究,每种实体模型均可得出一系列的阻力值和升力值;仿真是fluent中进行的,由于耳材村虫的尺寸较小,并且雷诺数相应较小,因此能够在所有模拟过程中使用层流条件减少模拟时间,同时保持良好的流体速度和压力精度。分析域是一个直径12mm、长度30mm的圆柱状空间,在下游方向上延伸的距离是模型尺寸的4倍,在横向方向上延伸的尺寸是模型尺寸的3倍,在上游方向上的延伸的尺寸是模型尺寸的3倍,网格个数约为465万个,如图11所示为网格划分情况及分析域情况。对于绕流流动,当雷诺数re<rec(临界雷诺数)时,流体状态视为层流。本实施例的仿真模型雷诺数re为:re=ρυd/μ其中:特征长度d为12mm;流体为水,密度ρ=1000kg/m3,水粘性系数;μ=0.001003kg/(ms);考虑耳材村虫可能达到的现实速度,从每秒大约一个体长(大约10mm)到每秒十个体长。因此,设定以下入口流速进行了仿真:10mm/s(大约每秒一体长)和100mm/s(每秒10体长)。带入上式计算得出雷诺数re=120,明显小于圆柱绕流的临界雷诺数2300,因此在所有模拟过程中选择层流模型laminar方程减少模拟时间,同时保持良好的流体速度和压力精度。对于边界层的处理采用了标准的近壁面函数法,离散格式选择一阶迎风模式。fluent相关设置如表1所示。表1fluent相关设置参数计算域12mm*30mm圆柱区域网格类型六面体网格流动介质水,运动粘度为0.001003kg/(ms)网格数量465万求解器三维单精度;基于压力基定常流动模型层流模型laminar方程;标准壁面函数法压力修正simple方法离散格式一阶迎风模式残差收敛0.001开始分析时还要打开阻力和升力监控曲线,打开“dragmonitor”对话框,设置阻力方向为y轴负方向,升力方向为z轴负方向,对60种模型分别做流体力学仿真,每种模型设置两种速度,0.1m/s和0.01m/s,可得出120组升力阻力值。因此四种模型在不同壳瓣展开角度的仿真结果如图12所示。s4:对每个模型的升力值、阻力值进行对比分析,找出升阻比最小的对应模型,即推断该模型为多节耳材村虫的3d复原模型结果;升阻比为升力值与阻力值之比。剔除部分异常数据,由图12分析可得:(1)每种模型的阻力不随壳瓣展开角度的变化而明显变化,基本稳定,但升力均在壳瓣展开120度时出现最大值,说明耳材村虫的壳瓣呈120度展开的固定姿态;(2)相同入口速度下,四种模型的阻力和升力数值对比,阻力值几乎相同,但最大升力均出现在两壳瓣分开、背部凸起腹部平直这个模型中,说明该模型即为最合理的耳材村虫身体形态;(3)随着入口流速的增大,各种模型的阻力和升力值均明显增大,符合流体力学的基本规律。本发明不局限于以上的具体实施方式,以上仅为本发明的较佳实施案例而已,并不用以限制本发明,凡在本发明的精神和原则之内所作的任何修改、等同替换和改进等,均应包含在本发明的保护范围之内。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1