一种黄芪拌种剂及其促进黄芪生长和黄芪根黄酮类化合物积累的方法与流程
本发明属于黄芪生态种植技术领域,具体涉及一种黄芪拌种剂及其促进黄芪生长和黄芪根黄酮类化合物积累的方法,采用内生菌pseudomonaspoae菌株ksc02拌种,促进干旱胁迫条件下黄芪根生物量和根冠比的增加,以及药用部位根中黄酮类活性成分芒柄花苷和毛蕊异黄酮葡萄糖苷含量的增加。
背景技术:
现有研究表明,内生菌通过合成植物生长激素吲哚乙酸(indoleaceticacid,iaa)、利用植物体内乙烯合成前体—1-氨基环丙烷-1-羧酸(1-aminocyclopropane-1-carboxylicacid,acc)作为碳源等方式,在植物生长、次生代谢物质合成积累及缓解环境胁迫等方面发挥着积极作用。针对内生菌在药用植物中的作用,国内外学者也进行了探索性研究,如已有研究发现,茅苍术内生细菌荧光假单胞菌具有促进苍术内酯合成与积累的作用;内生菌铜绿假单胞菌pseudomonasaeruginosa具有促进牛膝生长的作用;内生真菌trichodermaatroviride具有促进丹参毛状根生长及丹参酮积累的作用;内生真菌菌株gilmaniellasp.al12具有促进苍术倍半萜合成的作用;青蒿内生菌假诺卡氏菌属(pseudonocardia)菌株具有诱导青蒿素合成的作用。挖掘药用植物促生内生菌资源及其促生特征,解决目前中药材栽培中普遍存在的农药残留、药材品质下降等问题,是中药材种植业稳定、持续发展与利用的关键之一。
黄芪来源于豆科植物蒙古黄芪或膜荚黄芪的干燥根,属常用补益类大宗药材,具有补气固表、利水退肿、托毒排脓、生肌等功效。鉴于黄芪野生资源的匮乏和全民医疗保健意识的提高,目前市售黄芪多为栽培品。通过分析不同产地黄芪根内生菌群结构与组成,前期研究发现浑源黄芪细菌多样性远高于其他产地黄芪,且存在很多独特的细菌分类单元。进一步挖掘黄芪内生菌资源,应用于黄芪生态种植,对于黄芪种植产业的发展和黄芪资源持续利用具有重要意义。
合成植物激素吲哚乙酸(indoleaceticacid,iaa)是一大类植物促生菌共有特性之一,该类菌株在生长介质中含有色氨酸时,可以合成iaa并分泌至胞外,进而促进宿主植物的生长。在盐、干旱和土壤过度酸碱胁迫条件下,植物生长会受到抑制。分析胁迫条件下菌株iaa产生特征,选出促进iaa产生的胁迫因子,研究该胁迫因子作用下接种促生菌对宿主植物黄芪生长与活性成分积累特征,挖掘具有缓解环境胁迫、促进植物生长与活性成分积累的有益菌株,将有助于将该菌株应用于黄芪种植业。
技术实现要素:
本发明为了解决目前黄芪栽培、品质改善的问题,提供了一种黄芪拌种剂及其促进黄芪生长和黄芪根黄酮类化合物积累的方法,促进干旱胁迫条件下黄芪品质提高与产量提高。
本发明由如下技术方案实现的:一种黄芪拌种剂,由黄芪内生菌株草假单胞菌(pseudomonaspoae)菌株ksc02发酵制备而成,具体制备方法为:草假单胞菌(pseudomonaspoae)菌株ksc02制备成种子培养物,按照1‰接种量转接于lb液体培养基中,28℃、180rpm摇床培养过夜,培养物4℃、8000rpm离心10min,收集菌体沉淀;300mmmgcl2洗涤菌体沉淀2次后,重新悬浮菌体沉淀于mgcl2溶液中,并调整od600至1.0,制备得拌种剂,4℃保存备用。
所述黄芪内生菌株草假单胞菌(pseudomonaspoae)菌株ksc02于2017年11月21日保藏于中国微生物菌种保藏管理委员会普通微生物中心,保藏号为cgmccno.14946,菌种来源于黄芪鲜药材。该菌株为具有吲哚乙酸合成活性的内生菌株。
所述种子培养物的制备方法为:挑取保存在-80℃的草假单胞菌菌株培养物少许,在lb固体平板中划线、28℃培养24-48h,取单菌落继续划线培养1次活化菌株;挑取第二次活化菌株单菌落,转接于lb液体培养基,28℃摇床培养至对数生长晚期,即od600为1.8~2.0,获得种子培养物。
利用所述的黄芪拌种促进黄芪生长和黄芪根黄酮类化合物积累的方法,具体方法为:将黄芪种子与所述黄芪拌种剂按照湿重/g:体积/ml为3:20的比例混合后孵育黄芪种子90分钟,置于含无菌滤纸的培养皿中萌发,待两片真叶完全展开后,转移至无菌基质中光照培养4周;4周后用20%的peg6000连续浇灌幼苗3周。
本发明所述黄芪拌种用细菌菌株为申请号为201810199611.1,发明名称为《黄芪促生内生菌及其促生长方法与应用》中所记载的分离方法。
所述的黄芪促生菌在盐、干旱及酸碱胁迫条件下促进植物生长激素iaa产生;在干旱胁迫条件下黄芪生长及异黄酮活性成分积累中的应用。
本发明涉及的内生菌株:分离自黄芪道地药材—5年生浑源黄芪,应用于黄芪拌种,不存在二次污染问题;拌种促进黄芪生长及异黄酮类化合物积累应属道地药材适应栖息地干旱少雨的气候与土壤特征,属于缓解环境胁迫需要的生态适应特征,为中药材黄芪种植提供了一种绿色、无药材残留的环境友好型生物菌剂。
黄芪药材,目前以栽培为主,田间管理普遍存在喷施化学合成农药控制病虫害、施用化学合成肥料促进生长等现象,不仅容易造成农药残留量超标、质量下降,同时也造成土壤污染、板结和肥力下降等,不利于黄芪种植业的绿色发展。本发明所确定的拌种菌剂—草假单胞菌菌株ksc02,分离自5年生道地黄芪药材—浑源黄芪新鲜根部,应用于黄芪拌种,不存在二次污染的问题。此外,在甜菜、生菜、人参等经济/药用植物中的研究还发现,草假单胞菌除具有促生作用外,还具有拮抗致病性细菌、真菌生长及解磷活性。
在含7%nacl、20%peg6000及ph值为5.0的lb培养基上首先考察了盐度、干旱胁迫及酸碱度对该菌株生长及产生植物生长激素—吲哚乙酸(indoleaceticacid,iaa)的影响。在此基础上,制备菌体悬液与黄芪种子共同孵育,待幼苗生长出3~4片真叶后,20%peg6000模拟干旱胁迫处理幼苗三周。通过分析黄芪生长与活性成分含量,发现该菌株具有促进干旱胁迫条件下黄芪幼苗生长与活性成分积累的作用。
本发明涉及的黄芪拌种菌剂,还具有耐盐、耐酸碱等生物活性。浑源黄芪属道地药材,浑源属干旱、半干旱产区,产地土壤电导率介于39~44us/cm之间、ph值在8.05~8.29之间,植物生长也存在盐与碱性胁迫。耐盐碱及干旱特性,该菌株拌种显著提高了胁迫条件下黄芪产量和质量,有望开发为绿色农药,起到提高药材品质与产量、降低化学合成农药和肥料的双重作用。栽培黄芪的田间管理,亟待寻求环境友好型生长促进剂取代化学合成肥料、杀虫剂等,以应对黄芪种植中存在的上述问题,同时也有利于黄芪药用资源的持续开发利用。
附图说明
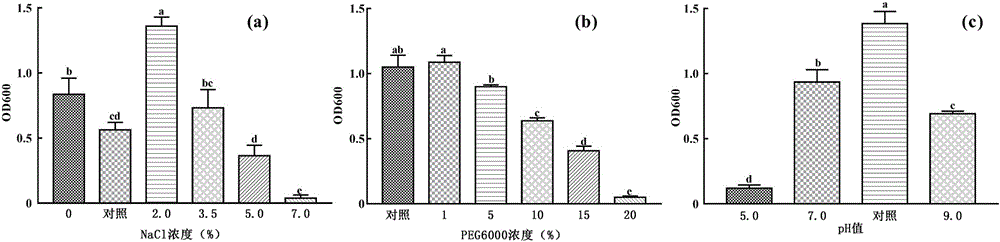
图1为胁迫条件对草假单胞菌ksc02生长(a、b、c)的影响;图中:a为盐胁迫的影响;b为干旱胁迫的影响;c为酸碱度胁迫的影响;柱状图标准偏差上的不同字母代表组间显著性差异。图2为极端胁迫条件下草假单胞菌ksc02生长(a)与iaa产生(b)特征;图中:横坐标为培养时间点,具体包括接种后5、9、13、17和30小时,a中纵坐标为od600,b中纵坐标为iaa浓度。图3为草假单胞菌ksc02拌种和20%peg6000单独处理及二者共同作用对黄芪生长的影响;图中:a为株高(厘米),b为茎叶鲜重(克),c为根鲜重(克),d为根冠比;mgcl2和ksc02分别为溶剂对照和ksc02单独处理组;20%peg6000、20%peg6000&mgcl2和20%peg6000&ksc02分别为peg6000单独处理、peg6000分别与溶剂对照和ksc02合并处理组;柱状图标准偏差上的不同字母代表组间显著性差异。图4为草假单胞菌ksc02拌种、20%peg6000单独处理及二者合并处理对黄芪幼苗根中活性成分积累的影响,图中:a为毛蕊异黄酮葡萄糖苷,b为芒柄花苷,c为黄芪皂苷ⅳ;柱状图标准偏差上的不同字母代表组间显著性差异。图5为草假单胞菌ksc02、20%peg6000单独处理及二者合并处理对黄芪根活性成分合成量的影响,图中:a为毛蕊异黄酮葡萄糖苷,b为芒柄花苷,c为黄芪皂苷ⅳ;柱状图标准偏差上的不同字母代表组间显著性差异。图6为草假单胞菌ksc02、20%peg6000单独处理及二者合并处理对黄芪幼苗成熟叶片、根脂质过氧化的影响,以丙二醛(malondialdehyde,mda)为评价指标,用于评价脂质过氧化程度;图中:a为成熟叶片,b为根;柱状图标准偏差上的不同字母代表组间显著性差异。
具体实施方式
实施例1:黄芪内生菌草假单胞菌ksc02分离与促生特征筛选
以山西浑源5年生黄芪鲜根为材料,采用组织块分离法分离黄芪内生菌。具体操作如下:将采回的鲜黄芪用自来水冲洗若干次,洗去残留在根表泥土,紧接着用含10%吐温-20的无菌水冲洗;在无菌条件下,将黄芪根切成1cm长的小段,置于75%酒精中浸泡1min,无菌水冲洗3次,放入5%次氯酸钠中浸泡10min,无菌水冲洗3次,然后放入75%酒精中漂洗40s,无菌水冲洗5次,用无菌滤纸吸去多余水分,将处理好的材料置于无菌研砵中,加入2ml生理盐水捣碎,研磨。
取100μl上清液涂布于lb固体培养基(培养基配方:蛋白胨1.0g、酵母粉0.5g、nacl1.0g、琼脂粉1.5克,溶于100ml蒸馏水中,ph7.2)平板,28℃避光培养2d。同时将最后一次无菌水洗液作为对照,若没有任何菌落长出,证明材料表面消毒彻底,分离到的菌为黄芪内生菌。
内生菌的纯化:根据菌落长出时间、形态及色泽,挑选代表性菌落转接于新的lb固体培养基平皿,进行纯化培养,经几次转接后得到菌落形态完全一致的单菌落即视为纯化后的内生菌。将纯化好的内生菌进行编号,然后转接于lb液体培养基中28℃培养至对数生长期,无菌条件下,将菌液与灭菌甘油混合均匀,甘油终浓度为20%,置于-80℃冰箱内保存。
参照文献(姜晓宇,高菊生,徐凤花,曹艳花,唐雪,张晓霞.水稻种子内生细菌多样性及其分泌植物生长素能力的测定.微生物学报,2013,53(3):269-275),定性判断草假单胞菌ksc02是否含acc脱氨酶。具体操作如下:将冰箱中保存的菌种划线接种于lb固体培养基,28℃培养过夜,选取单菌落再划线培养一次,挑取单菌落转接到adf固体平板,采用划线法连续传代5次,仍能正常生长者,说明含acc脱氨酶。
df盐培养基配方:kh2po44.0g,na2hpo46.0g,mgso4·7h2o0.2g,葡萄糖2.0g,葡萄糖酸钠2.0g,柠檬酸2.0g,(nh4)2so42.0g,组分1、组分2各0.1ml,h2o1000ml,ph7.2。其中组分1的配方为:h3bo410mg,mnso4·h2o11mg,znso4·7h2o124.6mg,cuso4·5h2o78.2mg,moo310.0mg,溶于100ml灭菌水中;组分2的配方为:feso4·7h2o100.0mg溶于10ml灭菌水中,将配置好的组分1、组分2置于4℃保存,备用。
adf培养基:将acc溶于超纯水,用无菌滤膜过滤除菌后,加入不含(nh4)2so4的df盐培养基中,其中,acc的终浓度为3.0mmol/l。在lb培养基上:草假单胞菌菌株ksc02菌落呈浅黄色,表面光滑,边缘整齐,随培养时间延长,菌落中心颜色加深,呈土黄色,lb培养基连续传代5次,产色素性能稳定;革兰氏阴性,菌体呈短杆状。草假单胞菌ksc02形态学信息见表1。
利用不含(nh4)2so4的df盐固体培养基,连续传代5次,仍能生长者,具有固氮活性。接种与培养条件与含acc脱氨酶菌株筛选方法相同。
表1草假单胞菌ksc02形态学信息
实施例2:环境胁迫对草假单胞菌ksc02生长的影响
挑取分离、纯化、鉴定并保存在-80℃的草假单胞菌菌株,首先在luria-bertani(lb)固体平板中划线、28℃培养,取单菌落继续划线培养1次,以活化菌株。挑取第二次活化菌株单菌落,转接于lb液体培养基,28℃摇床培养至对数生长晚期(od600介于1.8~2.0之间),获得种子培养物。按照1‰接种量接种上述培养物于含1%、2%、3.5%、5.0%及7%nacl的lb液体培养基中,以不添加nacl者作为阴性对照,每个浓度三个平行,摇床28℃培养过夜,转速180rpm,取部分培养物直接进行od600测定,获得菌体生长数据,。以nacl浓度为横坐标,od600为纵坐标,绘制柱状图,分析nacl浓度对菌株ksc02生长的影响。利用graphpadprism7.01软件进行组间差异显著性分析,阈值设定为p<0.05。结果见图1a。从该图可以看出,不加nacl和添加2%的nacl,od600即ksc02生物量数据显著高于对照;添加3.5%和5%nacl者,菌株生物量与对照处于同一水平;添加7%的nacl,菌体生物量数据明显低于对照,即7%nacl对菌株ksc02生长的抑制作用最强。
采用如上所述方法,制备种子培养液。按照1‰接种量接种于含1%、5%、10%、15%和20%peg6000的lb液体培养基中,以不添加peg6000者作为阴性对照,每个浓度三个平行,摇床28℃培养过夜,转速180rpm,取部分培养物直接进行od600测定,获得菌株ksc02生长数据。以peg6000浓度为横坐标,od600为纵坐标,绘制柱状图,分析干旱胁迫对菌株ksc02生长的影响,组间差异显著性分析方法同上。结果见图1b。从该图可以看出,培养液中peg6000浓度≤5%时,对菌株ksc02生长影响较小,生物量数据与对照处于同一水平;当peg6000浓度≥10%时,菌株ksc02生长明显受到抑制,其生物量数据明显低于对照,其中添加20%peg6000组,生物量数据最小,抑制作用最强。
采用如上方法,制备种子培养物。按照1‰接种量接种于ph值分别为5、7、9的lb液体培养基中,以正常的lb培养基作为对照,每个ph值三个平行,摇床28℃培养过夜,转速180rpm,取部分培养物直接进行od600测定,获得生物量数据。以ph值为横坐标,od600为纵坐标,绘制柱状图,分析酸度对菌株ksc02生长的影响,差异显著性分析方法同上。结果见图1c。从该图可以看出,实验条件下的三个ph值均引起od600的减小,其中ph5.0组降低最为明显,抑制生长作用最强。
实施例3:草假单胞菌ksc02在极端胁迫条件下生长(a)与iaa产生(b)特征
采用实施例2方法制备种子培养物。按照1‰接种量接种于含7%nacl、20%peg6000和ph为5.0已添加1mg/ml色氨酸的lb液体培养基中,以ph值为7.2、含1%nacl的正常lb培养基为对照,28℃摇床培养,转速180rpm。分别在培养5、9、13、17、30h,测定od600和od530。其中,部分培养物直接用于测定od600;同时取1.5ml培养液与等体积的salkowski显色剂(10ml0.5mol/lfecl3,500ml35%高氯酸,使用前混合,避光保存)充分混合,室温条件下避光放置30min,测定od530;以吲哚乙酸为对照品,绘制工作曲线,根据所得的线性回归方程,计算培养液中吲哚乙酸浓度。以培养时间为横坐标,od600为纵坐标,绘制散点图,分析上述极端胁迫条件对菌株ksc02生长的影响。以培养时间为横坐标,吲哚乙酸含量为纵坐标,绘制散点图,分析上述极端胁迫条件对菌株ksc02产生吲哚乙酸的影响。每个处理每个时间点三个平行。结果分别见图2a、2b。从图2a可以看出,酸胁迫对菌株ksc02生长影响最小,其生物量数据始终高于7%nacl组和20%peg6000组;此外,随着培养时间的延长,7%nacl组和ph5.0组菌株生长趋势与正常对照一致,均在培养时间为17h时生物量数据达到最大,而20%peg6000组随着培养时间的延长,生物量数据持续增大,在检测时间点生物量未达最高点,提示该菌株耐受20%peg6000能力强,活性佳。从图2b可以看出,ph5.0组与正常对照组iaa含量几乎处于同一水平,二者iaa产生模式类似,提示介质酸度对iaa产生影响较小;与对照相比,7%nacl和20%peg6000组iaa合成量显著降低,但后者随着培养时间的延长,iaa合成量明显高于7%nacl组,提示菌株ksc响应20%peg6000合成iaa活性较强。
实施例4:草假单胞菌ksc02拌种和20%peg6000单独处理及二者共同作用对黄芪生长的影响
拌种剂制备:采用实施例2方法制备种子培养物,并按照1‰接种量转接于lb液体培养基中,28℃摇床培养过夜,转速180rpm。无菌转移过夜培养物至无菌离心管中,4℃、8000rpm离心10min,收集菌体沉淀;300mol/lmgcl2洗涤菌体沉淀2次后,重新悬浮菌体沉淀于mgcl2溶液中,并调整od600至1.0,制备得拌种剂,4℃保存备用。
黄芪种子处理与萌发:首先用75%酒精振摇洗涤种子1min,而后用无菌水洗涤三次;紧接着用5%次氯酸钠溶液处理20min,无菌水洗涤5~6次;最后50℃水浴条件下孵育种子10min。待种子自然冷却至室温,按照3:20(种子质量:拌种剂体积)的用量,加入拌种剂,室温条件下共同孵育90min。挑取处理后种子于铺有无菌吸水纸的培养皿中,光照条件下萌发。光照培养条件:温度为23℃,光周期为16h(光)/8h(暗),光强度为2000~3000lux。注:按照每15g种子中加入100ml75%酒精和5%次氯酸钠的量,计算消毒剂用量。
拌种剂处理:待子叶完全展开后,转移萌发后种子于自吸水钵体中,每钵3~4粒种子。钵体基质为灭菌沙壤土与蛭石等体积混合物,前者辐射杀灭微生物,辐射剂量为10kgy;后者121℃30min灭菌。转移钵体于光照培养箱中光照培养,光周期:16h23℃(光)/8h23℃(暗),无菌水常规浇灌,光照强度1000~5000lux。无任何处理、300mol/lmgcl2处理者分别为正常对照和溶剂对照。
20%peg6000处理及材料收集:光照培养箱生长8周后,选取长势较为一致的黄芪幼苗进行胁迫处理。正常对照、溶剂对照及ksc02处理组幼苗随机分为2个亚组,一个亚组作为对照,常规浇以无菌水,另一亚组浇以20%peg6000,连续处理3周。单株收集根、茎叶。称量并记录鲜重及株高后,液氮速冻第3-5片叶子和根,-80℃保存。以茎叶鲜重代表地上部分生物量、根鲜重代表地下部分生物量,根鲜重与茎叶鲜重比值代表根冠比。
数据分析:数据以平均值±标准偏差形式表示。利用graphpadprism7.01软件进行组间差异显著性分析,阈值设定为p<0.05。有关结果见图3。从图3a可以看出,除ksc02单独处理株高显著高于对照外,其他组间均无显著差异;从图3b可以看出,ksc02单独处理引起地上部分生物量显著增加外,peg6000单独处理、peg6000&mgcl2与peg6000&ksc02共同处理均引起地上部分生物量显著降低;从图3c可以看出,除20%peg6000&ksc02共同处理组地下生物生物量显著增加外,其余组间均无显著差异;从图3d可以看出,20%peg6000&ksc02共同处理组根冠比显著高于其余组别,而ksc02单独处理导致其根冠比显著低于对照。概言之,干旱胁迫条件下,ksc02拌种处理黄芪种子具有促进黄芪药用部分根生物量及根冠比增加的作用。
实施例5:草假单胞菌ksc02拌种和20%peg6000单独处理及二者共同作用对黄芪根活性成分积累及合成量的影响
利用实施例4盆栽实验收集的根样本,进行黄芪皂苷ⅳ、毛蕊异黄酮葡萄糖苷、芒柄花苷含量测定。具体提取与含量测定,参照文献(孙欢欢,孙海峰,柴智,白云娥,高建平.碱处理在微量黄芪药材质控指标成分含量测定中的应用研究.药物分析杂志2018,38(9):1652-1660)方法进行。结果见图4。从图4a可以看出,20%peg6000与ksc02拌种共同处理组毛蕊异黄酮葡萄糖苷含量显著高于其余组别,而20%peg6000与ksc02拌种单独处理组毛蕊异黄酮葡萄糖苷含量与正常对照处于同一水平;从图4b可以看出,20%peg6000、ksc02拌种单独处理组以及二者共同处理组芒柄花苷含量均显著高于正常对照,且二者共同处理组含量略高于单独处理组;从图4c可以看出,20%peg6000、ksc02拌种单独处理组黄芪皂苷ⅳ含量与正常对照处于同一水平,二者共同处理组黄芪皂苷ⅳ含量明显低于对照。以上结果表明,干旱胁迫条件下,ksc02拌种具有促进黄芪幼苗根毛蕊异黄酮葡萄糖苷和芒柄花苷积累的作用。
鉴于20%peg6000处理可能引起细胞缩水,在活性成分合成量相同情况下,可能存在由于生物量减少而导致的活性成分含量增加;为了排除这种情况,利用实施例4盆栽实验获得的根生物量数据,乘以对应的黄芪皂苷ⅳ、毛蕊异黄酮葡萄糖苷、芒柄花苷含量数据,获得相关物质合成量数据。有关结果见图5。从图5a可以看出,20%peg6000与ksc02拌种一起处理组毛蕊异黄酮葡萄糖苷合成量远高于正常对照及其余组别;从图5b可以看出,20%peg6000、ksc02拌种单独处理组芒柄花苷含量与正常对照处于同一水平,二者共同处理组芒柄花苷合成量显著高于正常对照;从图5c可以看出,无论是20%peg6000、ksc02拌种单独处理组,还是二者共同处理组,黄芪皂苷ⅳ合成量与正常对照组处于同一水平。综上所述,ksc02拌种具有促进干旱胁迫条件下黄芪幼苗根毛蕊异黄酮葡萄糖苷和芒柄花苷积累的作用。
实施例6:草假单胞菌ksc02拌种和20%peg6000单独处理及二者共同作用对黄芪根、叶脂质过氧化水平的影响
液氮研磨实施例4盆栽实验收集的根叶材料呈粉末状,利用南京建成生物工程研究所提供的试剂盒,根据说明书推荐方法,采用分光光度法测定叶片和根中丙二醛含量。结果见图6。从图6a可以看出,20%peg6000单独处理及溶剂与peg6000共同处理组叶片mda含量与正常对照处于同一水平,而ksc02拌种单独处理及与20%peg6000共同处理组叶片mda含量显著高于正常对照,但较溶剂对照组低;从图6b可以看出,除溶剂与20%peg6000共同处理组根mda显著降低外,其余处理根mda含量与正常对照均处于同一水平。该结果提示,ksc02拌种与20%peg6000共同处理引起的黄芪根异黄酮活性成分即毛蕊异黄酮葡萄糖苷和芒柄花苷积累增加很可能与叶片组织中中等强度的脂质过氧化水平增加有关,但鉴于ksc02单独拌种与共同处理组mda处于同一水平,除抗氧化胁迫外,干旱胁迫条件下ksc02促进根生物量增加及毛蕊异黄酮葡萄糖苷和芒柄花苷积累作用还应该有别的机制存在。
- 还没有人留言评论。精彩留言会获得点赞!