一种提高苹果矮化效果和开花性能的方法与流程
1.本发明属于果树栽培领域,具体涉及一种提高苹果矮化效果和开花性能的方法。
背景技术:
2.矮化密植栽培是实现苹果现代化生产的基本要求,其中促进幼树产生一定数量的有效分枝是矮化密植栽培的关键技术之一。目前,国内外一般采用外源激素处理和人工修剪的方法调控苹果分枝(张庆伟等,2011)。带分枝的优质大苗可以直接进行定植,缩短了果园建成的时间;另外,优质大苗定植后,第二年即可开花结果,3~5年即可达到可观的产量。因此,枝组分布均匀、矮化效果好、开花性能佳是苹果现代化生产的需求。
3.苹果主要通过嫁接繁殖,对于我国苹果种植品种较为单一的富士系,其苗木的分枝性主要取决于砧木和激素处理等。本团队于2010年发现一株山定子(malus baccata borkh.)砧木突变体,其基部萌生出具有多分枝变异性状的新稍。经多年的生物学观察,该突变体与野生型相比,节间短、茎粗、叶片颜色红,分枝数量为野生型的10倍以上,该多分枝砧木资源也许能够解决以上存在的生产问题。
技术实现要素:
4.本发明针对上述现有技术存在的不足,提供一种提高苹果矮化效果和开花性能的方法。
5.具体技术方案如下:
6.一种提高苹果矮化效果和开花性能的方法,其将苹果多分枝芽变作为中间砧,以富士系苹果作为接穗。
7.其中,所述的苹果多分枝芽变型为山定子(malus baccata borkh.)具有多分枝性的自然突变植株或通过生物手段获得的其复制体。具体地,为植物新品种大棠博来,品种权号为20210256。
8.所述的生物手段可以是组培或嫁接等方式。
9.多分枝芽变(multi-branched bud,mb)是2010年在青岛市农业科学研究院北宅基地发现的一株山定子(malus baccata borkh.)自然突变体,具有多分枝性和矮化性等。2011年春,分别取mb与其野生型(wt)枝条接穗,以芽接的方式嫁接于一年生平邑甜茶,嫁接高度为30cm,分枝性状稳定遗传。2015年,观测到mb的开花情况。本发明团队于2019年为该突变体申请了植物新品种权,并于2021年授权,新品种名称为大棠博来,品种权号为20210256。
10.实验证明,mb作为中间砧,与富士系亲和性良好,枝组分配较为均匀,具有一定的矮化效果和提高开花的性能。
11.进一步,以平邑甜茶作为基砧。
12.进一步,本发明方法包括如下步骤:
13.(1)春季,将苹果多分枝芽变以芽接的方式嫁接于一年生平邑甜茶;
14.(2)当年秋,将富士系以芽接的方式嫁接于苹果多分枝芽变。
15.具体地,步骤(1)中,嫁接高度为30~50cm。
16.具体地,步骤(2)中,嫁接高度为保留苹果多分枝芽变30~50cm。
17.具体地,成苗后,移栽定植。
18.进一步,所述的富士系苹果为唐木田或烟富3。
19.本发明的有益效果如下:
20.本发明将mb(多分枝芽变)作为中间砧,与富士系亲和性良好,可以降低树体的生长势,使树体达到矮化效果,田间枝组发育较为均匀,枝组分布情况优于平邑甜茶,有一定的早花效果,具有较高的应用价值。本发明将为加快分子标记辅助育种进程,解决苹果生产中存在的现实问题提供重要的科技支撑。
附图说明
21.图1为实施例1中苹果多分枝芽变对树高的影响;
22.图2为实施例1中苹果多分枝芽变对节间长度和茎的粗度的影响;
23.图3为实施例1中苹果多分枝芽变的休眠芽特征;
24.图4为实施例1中苹果多分枝芽变的分枝特性;
25.图5为实施例1中苹果多分枝芽变的花特征;
26.图6为实施例1中苹果多分枝芽变的果实特征;
27.图7为实施例2中苹果多分枝芽变的叶片和茎的结构观察;
28.图8为实施例3中的苹果多分枝芽变的叶片叶绿素含量与野生型的对比;
29.图9为实施例3中的苹果多分枝芽变的叶片失水率与野生型的对比;
30.图10为实施例4中mb作为中间砧的树体植株的田间表现。
具体实施方式
31.以下结合实例对本发明的原理和特征进行描述,所举实例只用于解释本发明,并非用于限定本发明的范围。
32.具体实施方式中,使用的多分枝芽变(multi-branched bud,mb)生物材料为植物新品种大棠博来,品种权号为20210256。该生物材料保存于青岛市农业科学研究院,公众可向青岛市农业科学研究院获得,青岛市农业科学研究院保证在申请日起二十年内向公众发放该生物材料。该生物材料在如下文献中公开:
33.tan m,li g,chen x,et al.role of cytokinin,strigolactone,and auxin export on outgrowth of axillary buds in apple[j].other,2019,10.
[0034]
ge h,li g,wan s,et al.whole genome re-sequencing and transcriptome reveal an alteration in hormone signal transduction in a more-branching mutant of apple[j].gene,2022,818(6):146214.
[0035]
实施例1mb生物学特征观察
[0036]
2014~2017连续三年观察树形,枝干节间长度、茎粗度、当年生分枝数量,叶片大小、形状等,花(花量、花色、花瓣形状和花的育性),果实大小、形状和果色。卷尺测定株高,游标卡尺测定茎粗、节间长度、叶片大小和果实大小,量角器测定分枝角度。在花发育的不
同时期,用游标卡尺测定花丝和柱头长度(未剥除花托)。
[0037]
多分枝芽变对株高的影响如图1所示,多分枝芽变(mb)的一年生株高均值为138.7cm,为其野生型(wt)的80%,两者差异达显著水平。
[0038]
多分枝芽变对节间的影响如图2所示,多分枝芽变(mb)的茎直径为1.2cm,为wt茎粗的1.3倍,两者差异极显著;茎的节间长度为1.0cm,为wt的节间长度的二分之一左右,两者差异达到极显著水平。
[0039]
多分枝休眠芽特征如图3所示,多分枝芽变的休眠芽鳞片有绒毛;wt的叶缘无绒毛,休眠芽鳞片光滑,同样在休眠芽的着生位置上可以看出,wt的节间长度比mb的长,验证了一年生枝节间长度状态。
[0040]
多分枝芽变的分枝特性如图4所示,mb的一年生枝上的当年分枝数量均值为13.1个,为wt分枝数量的8倍,两者差异达到极显著水平;同时,mb的分枝与主干的角度能达到70
°
,野生型的分枝相对较为直立,分枝角度在30
°
左右,两者差异达到极显著水平;多分枝芽变的侧枝长度能达到30cm,是野生型的2倍以上,两者差异达到极显著水平(图4,a、b)。多分枝芽变的侧枝着生点较小,侧枝比主枝粗大,与主枝的连接部位极小,碰触易折断(图4,c)。
[0041]
多分枝芽变的花特征如图5所示,mb的花量较大,花瓣皱缩、淡粉,雌蕊发育弯曲,不能正常突出雄蕊群,花药肥大;wt的花量适中,花瓣平整、颜色深粉,雌蕊发育完整,能够正常坐果。
[0042]
多分枝芽变的果实特征如图6所示,mb在正常开花10d后,会出现二次开花现象,同时,果实在授粉20d变黄脱落;wt能够正常坐果,果型圆,果实较大(相对于海棠资源),红色,有棱,没有二次开花现象。
[0043]
实施例2mb叶片和茎结构观察
[0044]
实验方法如下:
[0045]
1、石蜡切片方法如下:
[0046]
(1)取材:从mb和wt一年生新稍顶端向下取第4片叶片,在叶中部靠近主脉处剪取0.5cm
×
0.5cm小块,用标准固定液(faa)固定后,茎的取样于茎中部切取0.5cm长茎段。
[0047]
(2)固定:固定48h以上。
[0048]
(3)脱水:30%
‑‑
50%
‑‑
70%(每步60分钟)
‑‑
85%
‑‑
95%
‑‑
100%
‑‑
100%(每步30min)。
[0049]
(4)透明:转入1/2二甲苯+1/2无水酒精混合液20ml透明1h,纯二甲苯20ml透明1h,再用纯二甲苯20ml透明40min,并回收各液。
[0050]
(5)浸蜡:
[0051]
①
将材料分别放入25%、50%、75%的石蜡-二甲苯溶液中,每级30分钟;该过程在高于石蜡熔点3℃的温箱中进行;
[0052]
②
将材料放入溶解的纯蜡中,时间为30min,该过程再重复两次;在高于石蜡熔点3℃的温箱中进行。
[0053]
(6)包埋:
[0054]
①
将溶解好的石蜡在加入预先折好的纸船中,石蜡溶解后应过滤后使用,以免含杂质影响切片;
[0055]
②
材料放入纸船,并摆好在蜡中的位置,如果包埋的组织块较多,应进行编号;
[0056]
③
将纸船放置在冷水上加速冷却过程;
[0057]
(7)切片:
[0058]
①
修整:用刀片修成方形或长方形蜡块;
[0059]
②
以少许热蜡液,将蜡块底部粘附于小木块上,以少许石蜡融化在蜡块边缘,加强粘附的强度;
[0060]
③
木块粘附于切片机上,调整切片机,使蜡块切面与切片刀刃平行;
[0061]
④
切成连续的蜡带,切片刀的锐利度、蜡块的硬度都会影响切片质量,可用热水或冷水适当改变石蜡的硬度,也可在冰箱中放置一段时间;
[0062]
⑤
分割蜡带,分开的蜡片放在45℃温水上,使蜡片展开。
[0063]
(8)粘片与烤片:
[0064]
①
可用粘片剂防止蜡片滑脱,但粘附剂通常容易被染色而使切片背景有颜色,影响观察,实验表明,载玻片洗的足够干净,一般不会滑片;
[0065]
②
用载玻片捞取展开后的蜡片,调整蜡片的位置使位于载玻片中央,自然干燥。
[0066]
(9)脱蜡与醇化:
[0067]
①
将切片放入纯二甲苯溶液中10min,使石蜡完全溶解,共2次;
[0068]
②
溶去石蜡的切片依次放入75%、50%、25%二甲苯-乙醇溶液中,每级10min;
[0069]
③
再将切片依次放入100%、95%、90%、80%、70%、60%、50%的乙醇溶液中进行醇化,每级10min。
[0070]
(10)染色:
[0071]
①
将切片放入番红染液中30分钟,番红染色后,将切片依次放入50%、60%、70%、80%的乙醇溶液中分级洗脱,每级10min;
[0072]
②
洗脱后的切片放入固绿染液中染色1min。
[0073]
(11)脱水、透明与封片:
[0074]
①
染色后的切片依次放入90%、90%、95%、100%乙醇溶液中分级洗脱,每级10min。
[0075]
②
洗脱后的切片依次放入25%、50%、75%、100%、100%二甲苯溶液中透明,每级10min,出现浑浊表示脱水不彻底,需重来。
[0076]
③
滴1~2滴中性树胶,将洁净的盖玻片倾斜放下,封片,镜检,选择好的贴上标签,永久性切片制作完成。
[0077]
2、解剖结构观察:
[0078]
在显微镜下观察切片,并拍照。采用e-ruler测量叶片厚度及上表皮、栅栏组织、海绵组织和下表皮厚度;观察茎的表皮、韧皮部和木质部。
[0079]
3、叶片表皮的扫描电镜观察:
[0080]
扫描电镜叶片取样方法同石蜡切片叶片的取样方法,放入2.5%戊二醛进行固定,经不同梯度的乙醇脱水,临界点干燥后,置镀膜仪上喷金,在kyky2008b扫描电镜下观察拍照。用e-ruler软件测量气孔大小,并根据单位面积内气孔个数计算气孔密度。
[0081]
实验结果如下:
[0082]
多分枝芽变的叶片特征如图7所示,多分枝芽变(mb)的完全展开叶片(功能叶)不
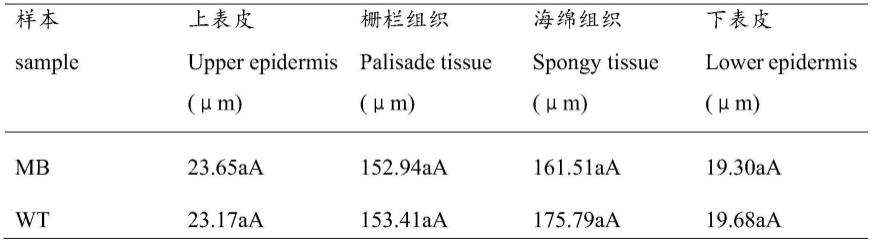
平整,早期叶片红色,花青苷着色程度高,后期转为浅绿,叶缘较为椭圆,叶片有绒毛;野生型的叶片较为平整,早期为红色,花青苷着色程度低,后期转为深绿,叶缘为锐锯齿状,绒毛无或较少(图7,a)。对多分枝芽变及野生型的叶片和茎解剖结构观察,结果表明:多分枝芽变的叶片栅栏组织2~3层,排列较为紧密,海绵组织稀疏,内有气孔开放细胞,其中上表皮、栅栏组织、海绵组织和下表皮厚度与野生型相比,差异不显著(见表1)。wt的栅栏组织内积累的淀粉粒较多,排列更为紧密(图7,b)。
[0083]
表1多分枝芽变的叶片解剖结构分析
[0084][0085]
多分枝芽变的叶片正面绒毛有绒毛,野生型未观察到或有极少的绒毛;多分枝芽变的叶片反面有气孔,气孔形状较圆,气孔开度与野生型相比略大(图7,c)通过观察多分枝芽变和野生型的一年生茎横切面,两者上表皮厚度差异不显著,但韧皮部、木质部和髓质的横截半径分别达到极显著差异;mb的韧皮部厚度为502.65μm,几乎是wt的2倍;mb的木质部半径高达995.24μm,是野生型的木质部半径的2倍以上;髓质的半径也几乎达到了wt的2倍(见表2)。
[0086]
表2多分枝芽变的茎横切解剖结构分析
[0087][0088]
实施例3mb叶片生理指标的测定
[0089]
实验方法如下:
[0090]
1、叶绿素a、叶绿素b和类胡萝卜素含量的测定:
[0091]
在5月中旬取2组叶片(每组5~7片叶片),分别为mb、wt的完全展开叶片,从茎尖往下数第5片叶,置于冰盒带回实验室。各组叶片用不锈钢打孔器打成1cm2的圆片,混匀,用分析天平准确称取0.5g左右,分别放入50ml锥形瓶中,加20ml蒸馏水,少量caco3粉末(防止脱镁),用高速电动匀浆机制成匀浆,定容至25ml,准确量取2.5ml,加入到50ml的试剂瓶中,再加入22.5ml无水丙酮,加塞,暗中提取光合色素30min,滤纸过滤,取滤液,即为90%丙酮光合色素提取液。
[0092]
90%丙酮提取液中叶绿素a、叶绿素b和类胡萝卜素浓度的测定:用90%丙酮作空白,用7200可见分光光度计测定提取液在440nm、644nm和662nm处的吸光度值(a),代入公式
(1)~(4)计算提取液中叶绿素a、叶绿素b和类胡萝卜素的浓度。
[0093]
(1)叶绿素a浓度(mg/l):
[0094]
ca=9.78
×
a662-0.99
×
a644
[0095]
(2)叶绿素b浓度(mg/l):
[0096]
cb=21.43
×
a644-4.65
×
a662
[0097]
(3)叶绿素总浓度(mg/l):
[0098]
ca+b=ca+cb=5.13
×
a662-20.44
×
a644
[0099]
(4)类胡萝卜素浓度(mg/l):
[0100]
ck=4.7
×
a440-0.27
×
ca+b
[0101]
=4.7
×
a440-(1.38
×
a662+5.48
×
a644)
[0102]
叶片中光合色素含量的计算:以下式分别计算叶片中叶绿素a、叶绿素b和类胡萝卜素的含量(mg/g鲜重)。
[0103]
x=25
×
103×c×
10/w=0.25
×
c/w
[0104]
式中:x-叶片中某种光合色素含量(mg/g);c-提取液中某种光合色素浓度(me/l);w-分析样品的鲜重(g)。
[0105]
结果见图8。
[0106]
2、叶片失水率的测定:
[0107]
分别取mb和wt完全展开叶片(茎尖向下第3~5片),每个样本取5片叶子,立即置于冰盒带回实验室。分别称取重量后,将叶片置于阴凉处,平放,每隔2h测定一下叶片重量,计算叶片失水率。
[0108]
结果见图9。
[0109]
3、光合指标的测定:
[0110]
用便携式光合测定系统(li-6400t,li,usa)测定,于9:30~11:00测定成熟健壮叶片的光合作用参数:净光合速率(net photosynthetic efficiency,pn)、气孔导度(stomatal conductance,gs)、蒸腾速率(transpiration rate,tr)和胞间co2浓度(intercellular co
2 concentration,ci)。开放式气路,co2浓度为400μmol
·
mol-1
,内置红蓝光源,设定光合有效辐射(par)为1000μmol
·
m-2
·
s-1
,5个生物学重复,每个重复测不同植株的叶片。
[0111]
结果见表4。
[0112]
实验结果如下:
[0113]
如图8所示,mb的叶绿素a、b和类胡萝卜素含量均低于wt的相关含量,差异分别达到显著性;同时根据spad值测定,mb的spad值为33.1,低于wt的39.4,两者差异达到极显著水平(见表3)。
[0114]
表3苹果多分枝芽变的spad值
[0115]
[0116]
注:**表示0.01水平的显著差异。以下相同。
[0117]
如图9所示,通过wt与mb叶片失水率测定,结果表明:叶片失水率随着时间的推移,逐渐升高;mb与wt的相比,在相同时间点的叶片失水率高;在44h后,mb的叶片失水率达到60%,与wt相比达到极显著水平。
[0118]
如表4所示,mb与wt的光合作用结果显示:mb的净光合作用和蒸腾速率仅有wt的一半,mb的气孔导度为18.8,仅为wt的三分之一,以上指标两者之间差异分别达到显著水平。
[0119]
表4多分枝芽变的光合作用分析
[0120][0121]
注:*表示0.05水平的显著差异。以下相同。
[0122]
实施例4mb作为中间砧对接穗生长及田间表现的影响
[0123]
实验方法如下:
[0124]
2016年春,分别将苹果多分枝芽变mb及其野生型wt以芽接的方式嫁接于一年生平邑甜茶,嫁接高度为30cm;当年秋,分别将唐木田或烟富3号以芽接的方式嫁接于mb或wt上,嫁接高度以保留mb或wt 30cm。砧穗组合分别为:唐木田/mb/p、唐木田/wt/p、yf3/mb/p和yf3/wt/p。(注:砧穗组合为:接穗/中间砧/基砧;p为平邑甜茶;yf3为烟富3)。上述砧穗组合成苗后,将其移栽定植于城阳农场,定植株行距分别为1.5米
×
2米。每年观察植株生长状况,连续观察3年。
[0125]
表5为唐木田不同砧穗组合树体生长特性调查,除前述砧穗组合外,还与同一农场同年生的其他组合:唐木田/cg80/p、唐木田/07qz1-5、唐木田/qz1*luo2-2进行了对比。(注:砧穗组合为:接穗/中间砧/基砧;或接穗/自根砧;p为平邑甜茶;07qz1-5为青砧1号后代(半矮化);qz1*luo2-2为青砧1号杂交后代(半矮化))。
[0126]
表5唐木田不同砧穗组合树体生长特性调查(城阳农场,2019,3年生)
[0127]
[0128]
如表5所示,不同砧木对相同接穗唐木田的生长发育有影响。中间砧mb株高均值最低,为1.94m,与中间砧wt、自根砧07qz1-5、07qz1*luo2-2差异达到极显著水平;与中间砧cg80差异不显著,矮化效果较好。中间砧mb的枝干比均值为0.48,大于wt的0.45,小于07qz1*luo2-2的0.56,分别与wt、07qz1-5、07qz1*luo2-2差异不显著,枝组分布较好;cg80的枝干比最大,均值为0.62,与wt、mb等差异达到显著水平,枝组分布最差。中间砧mb的分枝数量均值为22.67,高于cg80的13.4,低于07qz1-5的24.60,与07qz1-5、07qz1*luo2-2差异不显著,与cg80、wt差异达到显著水平。统计2019年开花结果率,结果表明:mb为中间砧的唐木田在第3年就可开花,但wt并没有开花结果的迹象,mb的早花性优于wt。但2019年的开花结果率,以唐木田/qz1*luo2-2最高,为72.73。
[0129]
表6为烟富3不同砧穗组合树体生长特性调查,除前述砧穗组合外,还与同一农场同年生的其他组合:yf3/gm256、yf3/b9进行了对比。(注:砧穗组合为:接穗/中间砧/基砧;或接穗/自根砧;p为平邑甜茶;yf3为烟富3)。
[0130]
表6 yf3不同砧穗组合树体生长特性调查(城阳农场,2019,3年生)
[0131][0132]
如表6所示,砧穗组合yf3/mb/p的株高与yf3/b9相似,均为190cm,mb具有一定的矮化效果,与b9相当。wt的株高最大,均值能达到218.16cm,与mb、gm256、b9差异达到极显著水平。行内和行间冠径均以yf3/wt/p最大,与mb、gm256、b9分别达到极显著水平。yf3/gm256的侧枝数量最多,均值能达到25.40个/株,而wt为中间砧的树体侧枝数量最少,均值为21.56,分别与mb、gm256达到显著差异。2年生树的开花数量以yf3/b9为最佳,均值为4.85个/株,分别与mb、gm256、wt达到极显著差异。
[0133]
苹果多分枝芽变(mb)的田间表现如图10所示,图10中,mb代表mb作为中间砧的植株(砧穗组合yf3/mb/p)。mb为中间砧的树体植株较矮,与b9、qz2(青砧2号)相似;m2(美2)的树体高大,长势旺盛。mb为中间砧的树体侧枝数量优于b9和m2,与qz2相似。总体mb在田间表现较好,具有矮化性,枝组分布较均匀。
[0134]
砧穗间的相互作用,对树体最明显的表观影响即为树冠形态、枝类组成、生物量及树体开花结果习性的改变。砧穗组合红富士/八楞海棠比红富士/山定子组合的萌芽率高4.3,短枝率高3.0%,成枝率低3.0%(王丽荣等,2007)。大量研究结果表明矮化砧木作为中间砧或基砧均可影响接穗品种地上部的生长,不同矮化砧木的矮化效果有所差异,主要表现为树高、冠径、干周、枝量及叶片生长量偏少,中短枝的单枝鲜重、比枝重、单枝叶数、单枝叶面积、单枝叶鲜重等偏大,各种形态的改变使矮砧树体的树冠变小(姜林1998;王继世1987;李登科1993)。12年生矮化中间砧国光苹果树的单株枝量比乔化砧苹果树少29.2%,单位面积的枝量比乔化树多44.8%,短枝多48.5%(张国岐1994)。sh38作中间砧在江苏地
区可强烈抑制苹果树体生长,在抑制树高、新梢生长量方面效果显著优于m26、sh28、sh10等(徐继忠,2002)。
[0135]
砧木对树体生长的影响,因不同砧穗组合差异不同,砧木的长度也会影响砧木与接穗间的相互作用(樊秀芳和史联让,2001)。万述伟等(2016)认为“m26”、“b9”、“cg80”、“mark”和“崂山柰子”等苹果矮化中间砧对苹果品种“嘎拉”的顶芽形成早晚,分枝角度大小,1年生枝条长度、粗度、节间长度,枝量与枝类构成具有明显影响,从而影响嫁接树的株高、树干横断面积、树冠大小和成花能力。从矮化中间砧对接穗品种前期生长发育的影响来看,“m26”、“b9”、“cg80”作中间砧在抑制“嘎拉”苹果树体生长、促进成花等方面的综合表现优于“mark”,更优于“平邑甜茶”。姜淑冬(2010)认为梨树矮化中间砧的矮化效果与矮化砧木的长度成正相关。m26为红富士苹果中间砧木,其露出地面越高,对树体生长的抑制作用越明显,露出地面过低会导致矮化效果不理想,露出地面的最适宜高度为10-15cm(秦立者等,2008)。研究表明矮化中间砧入土深度为15cm时,苹果树体生长势强,细根数量多,有利于树体生长发育(任雪菲等,2013)。也有研究认为矮化中间砧对大龄苹果树的影响较小,杨廷祯等(2010)研究了二十年生的红富士苹果树,其中间砧木采用sh1,结果表明sh1中间砧长度及sh1中间砧入土深度与树体的生长无明显相关性。mb作为中间砧,可以降低树体的生长势,使树体达到矮化效果,枝组分布情况优于平邑甜茶。
[0136]
实施例5苹果多分枝芽变(mb)砧穗组合对富士品质的影响
[0137]
2014年春,分别将苹果多分枝芽变mb及其野生型wt以芽接的方式嫁接于一年生平邑甜茶,嫁接高度为30cm;当年秋,将烟富3以芽接的方式嫁接于mb或wt上,嫁接高度以保留mb或wt 30cm,砧穗组合分别为烟富3/mb/p、烟富3/wt/p。上述砧穗组合成苗后,将其定植于北宅基地,定植株行距分别为1.5米
×
2米。2019年,测试砧穗组合对富士品质的影响,并与同基地、同年生的砧穗组合烟富3/p、烟富3/m26/p、烟富3/qz2进行对比(注:p为平邑甜茶,qz2为青砧2号)。结果见表7、8。
[0138]
表7不同砧木对烟富3果实品质影响(北宅基地,2019,5年生)
[0139][0140]
表8不同砧木对富士果实品质影响(北宅基地,2019,5年生)
[0141][0142]
如表7、8所示,不同砧木对富士果实品质有影响。单果重以p为自根砧的富士果实最大,均值达到272.26g;m26次之,均值为265.50g;wt为中间砧的富士苹果单果重最小,仅有191.65,可能与当年wt结果过多所致。纵横比以p和qz2为佳,能达到0.8以上。可溶性固形物以wt最高,均值达到16.33;mb次之,均值为15.88,两者差异不显著。可溶性固形物最低的p为基砧的富士,均值仅14.25,与wt、mb差异显著。果实硬度以qz2为基砧的富士最大,mb次之,m26最小,其中qz2分别与p、m26、wt相比,达到显著差异水平。
[0143]
液相色谱检测结果对于果实中矿质元素含量统计分析,结果表明:不同砧木对富士果实中ca、cu、fe、mg、mn、zn和总酚含量的影响差异不显著,se含量以p和wt最高,均值为0.03μg/g,其中wt与mb的0.02μg/g相比达到显著差异水平;以p为砧木的富士果实可滴定酸含量最高,均值为2.15mg/g,分别与mb、m26、wt、qz2相比达到显著差异水平;vc以wt为中间砧的富士含量最高,均值达58.05μg/g,分别与mb、wt、qz2相比差异不显著;vc以m26为中间砧的富士含量最低,仅18.75μg/g,与p相比差异不显著,分别与mb、wt、qz2相比差异达到显著水平。
[0144]
同样的,对于mb的不同株系(生长特性稍有差别),对富士果实品质也有影响,如表9所示。就单果重和纵横径比而言,mb不同株系与wt相比,均没有显著差异。果实硬度以mb-2的最大,均值达到4.02,与mb-1、mb-3和wt相比,分别达到显著差异水平。可溶性固形物,wt均值为12.00,小于mb-1的13.2,大于mb-2的11.4,分别与两者相比差异不显著;但mb-1与mb-2相比,达到显著差异水平。风味以wt为佳,mb-1次之,mb-2最差。
[0145]
表9不同砧木对富士果实品质影响(北宅基地,2020,6年生)
[0146][0147]
[0148]
砧木作为疏导系统,影响了果实激素水平或矿质营养的运输与积累。果实的糖酸含量及组成对果实品质有着重要影响(dandekar et al.2004)。gao等(1992)研究表明,不同砧木对氮素的不同形式反应不同,矮化砧木和半矮化砧木的木质部赤霉素与细胞分裂素类激素受氮素影响明显,从而影响嫁接苹果果实的糖酸含量和矿质元素积累。烟富6/a1和烟富6/a1d的各单糖含量均较高;烟富3则以a1d、m26和平邑甜茶为砧木的组合各单糖含量较高,a1的砧穗组合最低,这可能与砧穗互作机理有关,不同砧木对不同接穗所产生的影响不同。无论是烟富6还是烟富3可溶性固形物含量均以a1d和m26砧穗组合最高;可滴定酸含量也以m26砧穗组合最高,烟富6/a1d和烟富3/a1组合最低(解贝贝等,2013)。本发明实验表明,不同砧木对富士果实品质有一定的影响,其中矿质元素se含量、可滴定酸和总酚含量上存在显著差异,在硬度、可溶性固形物和其他矿质元素中并没有表现显著差异性。
[0149]
对于该砧木的应用前景,虽然该多分枝芽变本身扦插性能较弱,组培苗也较难获得,下一步我们可以通过杂交育种对其改良。其作为中间砧,具有一定的矮化和早花效果,田间枝组发育较为均匀,也具有一定的应用价值。
[0150]
以上所述仅为本发明的较佳实施例,并不用以限制本发明,凡在本发明的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1