治疗性噬菌体
本发明涉及可用于预防或治疗沙门氏菌(salmonella)感染的治疗性噬菌体、此种噬菌体的套组以及此种噬菌体的药物组合物。本发明还涉及用于干燥噬菌体溶液的方法和用于制备热稳定噬菌体的方法。
背景技术:
非伤寒沙门氏菌属(non-typhoidalsalmonellaspp.)是世界范围内食物中毒的主要原因,并且在英国,每年有33,000例人沙门氏菌病的报告。欧洲食品安全管理局(efsa)估计,11.7%报告的人沙门氏菌病病例是由消费猪肉产品引起的。英国动植物健康局(apha)的流行病学监测确定,与英国猪相关的五种最流行的沙门氏菌血清型是:鼠伤寒沙门氏菌(s.typhimurium)、s.4:5:12:i>、s.4:5:12、和德尔卑沙门氏菌(s.derby)、病牛沙门氏菌(s.derby)。德尔卑沙门氏菌、s.4,5,12:i>、和鼠伤寒沙门氏菌也在美国5种最流行的血清型之内。此外,在欧盟,鼠伤寒沙门氏菌、s.4,5,12:i>、s.4:5:12和德尔卑沙门氏菌是efsa报告的最流行的沙门氏菌血清型。
研究表明,沙门氏菌的流行和散布在断奶期间以及当市场重量的猪离开农场并被运送到屠宰场进行屠宰时特别增加,这增加了最终消费猪肉产品的躯体污染可能性。这个问题特别令人担忧,因为已经从猪中分离出多重耐药性(mdr)沙门氏菌菌株。可用于治疗猪中的沙门氏菌的抗生素种类正在减少,并且主要担心mdr的传播或人食物链中抗生素残留的存在。
技术实现要素:
显然,迫切需要抗生素的替代物来控制沙门氏菌感染。
本发明人已经分离并表征了噬菌体(靶向并杀伤特定细菌物种的细菌的天然病毒),其靶向与猪和鸡相关的多重耐药性沙门氏菌菌株。
因此,本发明提供了一种肌病毒科的噬菌体,其包含长度为200kbp或更长的环状排列的双链dna的基因组;其特征在于所述噬菌体具有以下特征中的任一个或更多个:
a)不具有穿孔素基因;和/或
b)与沙门氏菌脂多糖结合;和/或
c)仅具有一种尾纤维蛋白;
以及
d)具有与seqidno.7具有至少95%序列同一性的序列和/或与seqidno.8具有至少80%序列同一性的序列。
适当地,提供特征a)至d)中的至少两个,适当地,提供特征a)至d)中的至少三个,适当地,提供特征a)至d)中的至少四个。例如
a)和b);a)和c);a)和d);b)和c);b)和d);或c)和d),或
a)、b)和c);或a)、b)和d);或b)、c)和d);或a)、c)和d)。
优选地,所述噬菌体能够裂解3种或更多种与家禽、猪和/或牛相关的沙门氏菌菌株。例如,以下项中的任一个或更多个:a)25种鼠伤寒沙门氏菌;b)15种s.4,12:i>;c)10种s.4.5,12:i>;d)10种病牛沙门氏菌;e)10种德尔卑沙门氏菌;f)3种s.13,23:i>;g)5种肠炎沙门氏菌(s.enteritidis);h)4种婴儿沙门氏菌(s.infantis);i)3种s.ohio;和j)3种s.seftenberg。
适当地,噬菌体能够裂解4种或更多种、5种或更多种、6种或更多种、7种或更多种、8种或更多种、9种或更多种或者10种或更多种与家禽、猪和/或牛相关的沙门氏菌菌株。
所述噬菌体可以与保藏编号nctc18080701;nctc18080702;nctc18080703;nctc18080704;nctc18080705;或者nctc18080706的噬菌体中的任一种具有80%-100%的序列同一性。
这些保藏于2018年8月7日在欧洲认证细胞培养物保藏中心进行,保藏编号为18080701;18080702;18080703;18080704;18080705;或者18080706。
适当地,噬菌体可以从2018年8月7日在欧洲认证细胞培养物保藏中心保藏的保藏编号为18080701;18080702;18080703;18080704;18080705;或者18080706的噬菌体中选择。
噬菌体可以存在于套组中,该套组包含两种、三种、四种、五种、六种或更多种上述噬菌体。例如,nctc18080703和18080704。适当地,套组可以包括18080701;18080702;18080703;18080704;18080705;或18080706中的任一种或其组合,其中
表1
适当地,噬菌体可以选自spfm1、3、10、14、15、17和/或19或其组合。适当地,噬菌体可以选自spfm1至22中的任一种或更多种。适当地,噬菌体可以选自spfm1至22中的任一种或更多种,不包括spfm9和spfm11。适当地,噬菌体可以选自spfm2、4、10、14、17和/或19中的任一种或更多种。适当地,至少2种、至少3种、至少4种噬菌体可以选自spfm2、4、10、14、17和/或19。适当地,可以提供spfm4和17或者spfm10和14的组合。适当地,可以选择包括spfm4、10、14和17中任两种的组合。适当地,可以在套组中提供至少一种在ph3下稳定的噬菌体。适当地,用于套组中的噬菌体可以选自2、4或19中的至少一种或其组合。适当地,可以使用耐热降解的噬菌体。适当地,可以选择选自spfm10和17的噬菌体。适当地,可以选择spfm10-。
适当地,可以选择选自以下的三种噬菌体组合:
spfm4、10和19
spfm10、17和19
spfm2、17和19。
适当地,可以选择选自以下的四种噬菌体组合:
spfm10、14、17和19
spfm2、10、14和19
spfm2、4、10和19。
表1提供了这些噬菌体的核苷酸登录号以及根据本文所讨论的布达佩斯条约已经进行的噬菌体保藏的详细信息。
噬菌体可以是药物组合物的一部分,该药物组合物包含上述噬菌体或套组中的一种或更多种。
可替代地,噬菌体可以是动物饲料或消毒剂,其包含上述噬菌体、药物组合物或套组中的任一种。
在另一方面,上述噬菌体、套组、药物组合物或动物饲料可用于治疗方法,例如预防性治疗方法。待治疗或预防的疾病可以是家禽、猪或牛中的沙门氏菌感染。
因此,提供了根据本发明的用于治疗沙门氏菌的噬菌体。适用于在猪和/或家禽(例如鸡)中治疗沙门氏菌。
还提供了用于干燥噬菌体溶液以形成粉末的方法,所述方法包括:
a)将所述噬菌体与以下项中的任一种或更多种混合;
(i)糖;和/或
(ii)糖醇;和/或
(iii)氨基酸;和/或
(iv)聚合物;
以及
b)干燥所述混合物。
适当地,所述方法的特征包括其中:
·通过喷雾干燥进行干燥;和/或b)其中所述方法还包括由经干燥的粉末制备片剂或丸剂;
·糖是基于葡萄糖的糖,优选海藻糖。
·糖醇是甘露醇。
·氨基酸是脂肪族氨基酸,优选亮氨酸。
用于干燥方法的噬菌体溶液可以包含上述噬菌体、请求保护的套组或药物组合物中的任一种。
在另一方面,提供了一种噬菌体组合物,其包含噬菌体、糖、糖醇、氨基酸和聚合物。噬菌体、糖、糖醇、氨基酸或聚合物可以是上述记载这些中的任一种。
还提供了一种热稳定噬菌体,其包含:
a)与seqidno.1具有至少80%序列同一性的序列中的多态性1;和/或
b)与seqidno.2具有至少80%序列同一性的序列中的多态性2;和/或
c)与seqidno.2具有至少80%序列同一性的序列中的多态性3;和/或
d)与seqidno.2具有至少80%序列同一性的序列中的多态性4。
适当地,噬菌体可以具有本文所述的多态性1、2、3或4,并且与seqidno.2具有至少85%、至少90%、至少95%、至少97%和至少99%的序列同一性。
所述热稳定噬菌体可以是上述噬菌体中的任一种。
在另一方面,提供了用于制备热稳定噬菌体的方法,其包括在以下位置使噬菌体基因组的核苷酸序列突变:
a)多态性1在与seqidno.1具有至少80%的序列同一性的序列中从碱基a、g或c到t的位置;
b)多态性2在与seqidno.2具有至少80%的序列同一性的序列中从碱基a、t或c到g的位置;
c)多态性3在与seqidno.2具有至少80%的序列同一性的序列中从碱基a、g或t到c的位置;和
d)多态性4在与seqidno.2具有至少80%的序列同一性的序列中从碱基a、g或c到t的位置,
其中所述方法任选地包括将seqidno.1和/或seqidno.2引入所述噬菌体基因组的第一步骤。
适当地,噬菌体可以具有本文所述的多态性1、2、3或4,并且分别与seqidno.1或2具有至少85%、至少90%、至少95%、至少97%和至少99%的序列同一性。
下面仅以示例的方式并参考所附实施例进一步描述本发明。
具体实施方式
噬菌体
噬菌体(bacteriophage),也通常称为噬菌体(phage),是在其靶细菌中感染和复制的病毒。噬菌体可以是前噬菌体、温和/溶源性噬菌体、噬菌体样颗粒(如质粒)或裂解性噬菌体。
前噬菌体/温和/溶源性噬菌体是由双链或单链dna或rna组成的噬菌体颗粒。噬菌体基因组可以插入并整合到环形细菌dna染色体中,或作为染色体外质粒存在。这是一种潜在形式的噬菌体,其中病毒基因存在于细菌中,而不会导致细菌细胞的破坏,有时可以为细菌宿主的整体适应性提供竞争优势。
裂解性或毒性噬菌体包含与宿主细菌dna分开存在并与宿主细菌dna分开复制的病毒dna/rna。裂解性噬菌体在经感染的细胞及其膜被破坏时释放。
所述的噬菌体是裂解性噬菌体,并且是从其天然环境中分离和/或纯化的。
噬菌体的形态学
本文所述的22种噬菌体具有等长的头部和可收缩的尾部,并且被归类为尾状病毒科中的肌病毒科成员。
噬菌体的尾部从130到260nm不等。例如,140、150、160、170、180、190、200、210、220、230、240或250nm。
衣壳直径为约70nm至160nm。例如,80、90、100、110、120、130、140或150nm。
噬菌体的遗传分析
噬菌体具有线性、环状排列的双链dna基因组。基因组长度为约200-260kbp。例如,205、210、215、220、225、230、235、240、245、250或255kbp。也就是说,噬菌体被归类为巨型噬菌体。
噬菌体可以没有穿孔素基因;和/或噬菌体可以仅具有一种尾纤维蛋白;和/或噬菌体受体可以是沙门氏菌脂多糖(lps)。
噬菌体的基因组可以额外地具有以下特征中的任一个或更多个,包括这些特征的任何组合:
·仅编码一种trna;和/或
·编码多于1种的具有不同dna序列的rnap(rna聚合酶)β亚基。
也就是说,rnapβ亚基具有小于50%的序列同一性;和/或
·编码叶酸合成路径的基因,例如二氢叶酸还原酶和/或胸苷酸合酶和/
或胸苷酸激酶;和/或
·dna腺嘌呤甲基化酶基因;和/或
·自由基s-腺苷甲硫氨酸基因。
噬菌体与myrovidae家族、沙门氏菌噬菌体spn3us和segd1的其他成员共享几个核心基因和假设蛋白质的序列同一性(尽管没有核心基因与这些已知的噬菌体共享100%的序列同一性)。
几个核心序列之间的差异如下所列:
seqidno.3:内溶菌素。95%类似于spn3us和segd1。功能:噬菌体在复制周期结束时使用的酶以从内部降解细菌宿主的肽聚糖,导致细胞裂解和子代病毒体的释放。
>mfrtilfllvamttfnsnasyqttsnkhqkefvliqdqfneikplivtasrkqgvhagmlattiyresrfnknvgknksssassvvqmttgtkrsmirlygkqlnipknadlnkpkyavqlaavymkhieehltkqlkrkpstaeialgyrfgegaavamikkkspvgkrwmasyrkdaafygakmtppkaetrqlafakedhdqrvaelqkiwdtlytkispaagtllanntimkgall
seqidno.4:乙酰转移酶(acetyltransferase,gnat)家族蛋白质。95%类似于spn3us和63%类似于segd1。功能:gnat可以具有一定范围的酶功能,包括对抗生素的抗性。
>mktvaklfrssirsaefqnnvgtdtvrarrlkeflkdetysvyvvrnketwegylvatyrhgdptltchyfeefvpgafdkvglalvldhtgfeymscpaptgdveqklvekgfvlsdivwachsklfdkvitaseikiisgdayaelskddkdvilnlvsgaiceeleydvrghythigplaetviariqhanvetaiwrdgrdivgiaqmrlvapgvvelsgltvapthrkrgigrrlmgalfdfaqrhsdqvyietaadnnpanhlygnilrckqvlrtltlkrdpeliswkrvkgnradrvlkeepslpvetnprptindmfqsgmrgc
seqidno.5:n-乙酰转移酶gcn5。67%类似spn3us和99%类似segd1
>mkivfvtkpnspygkkyyadllglhnklyddqkikladlngregfnqiskleevwgegtlcacavddrgvavgfitfgytkkgqrflwvynffvdesvrsqgvghelieavenygkskgcawmqlnvlgnndraiafyerfgfrteyqdmvkel
seqidno.6:phikz样内部头蛋白。35%类似spn3us和99%类似segd1功能:类似于巨型噬菌体phikz的头蛋白。
>manfvksklaresveatgdvidglsnvepaeenidvqlaevasidgqleqldgdqetlaadterteaaidaadekiengeempeeaiahtevaqesirkrwaidrtklaresyrrgrgmtkaaqegwketlkdlfkrfvelckaviakakelklkyinvgksaqkrakayqemlrklgkqkkenisggfisklsiegdfdvansiaiakeltggkakdainklssqasesavaitktaegtataakgavdvalfgtaakklrtlpqfedgeggqkvlalpgnayvqigsktlaggqdftavaflstgdstdtkeiatpaitalasaataldaigkgfekvlqdfrsydadvekleqaaskaaaaldksndegewealrnartaadqavrnyqtlnravahvgntvisglngyigagigayekska
seqidno.7:噬菌体推定的atp依赖性dna解旋酶:41%类似segd1和99%类似spn3us
>mveiqmtytsmgvrvevpirqaerailawaeenmhapkmgkqhgrittergdayyahipslrtfifhkvfaekikhilqraaieyafqykltehhpergtpysctfdnygfsmvesdpesrfyyqnevvdaasdpnrpqtifaiqtgrgktkscmksmvkrgtrtalihrpsyvpkwlfdvcddetglrilrdevlvctgvqaiydalemgksgeldrrgikviilptvslqrflkeyintaatnpvdldtfydvlgvglvamdevhehfhlvymagimlnppasiemsatlkpgsskafiaerylerfpmeyrisipiipvvdvkalyyrlddkkfawwaskmtpynhklfegklikenlhleyanmiwdvveksflkryqpgqkclllfatvamceffteyvkdklsrdpvfhplmvakynagdsyddfiqadfsistpgkagtavdkpglvhmyistpvedqqlneqmagrprkilhnewgeidptvwlfhaynvpkhcnylnarqkslkdvvlsfriatspyvvrkshahsaassrataalhrndfskfsrkhskgvsrrrrrr
seqidno.8:hsiv家族蛋白的推定肽酶不存在于spn3us或segd1中。
mttiafdgkvlaadgqltrgsnicnlntqklficpsdeewwitgerivafgvsgditgqlalletvrlrpgykgltqssqlpakmdftflmllasgkaivggkreddvtlwwseatpplatgsgyeyalgamkmganavraveiasecdiytggeistyelr
保藏编号
包含在本发明的套组中的噬菌体可以包括保藏的噬菌体的突变体和变体。
根据下表,在本申请中提及本发明的噬菌体:
这些保藏是根据《布达佩斯条约》在nctc/ecacc进行的,遵循了根据《布达佩斯条约》运作的国际保藏机构(ida)的规则,并于2018年8月7日获得保藏。
此类保藏的噬菌体的突变体和变体保留了杀伤沙门氏菌的能力。与本发明的至少任一种保藏的噬菌体的基因组相比,突变体和变体在整个基因组中具有至少80%、85%、至少90%、至少95%、至少96%、至少97%、至少98%或至少99%的核苷酸序列同一性。
此类突变体和变体可能是从保藏的噬菌体的核酸序列添加、删除或替代核苷酸来产生的(例如1、2、3、4或5个核苷酸,任选连续核苷酸的添加、删除或替代)。
任选地,突变体或变体不包括整合酶基因和/或毒素基因。除了保留杀伤沙门氏菌的能力之外,突变体和变体可以具有对于其特征所来源的保藏噬菌体所述的任何进一步特性。
套组
噬菌体的套组可以包括本文提供的噬菌体的任何组合。噬菌体可以准备用于单独、顺序或同时施用。
可以从上述组中选择的噬菌体的所有组合考虑包括在该套组中。
套组可以具有额外的组分。例如例如,可能不是噬菌体的组分。例如,载体、赋形剂、治疗剂(例如非噬菌体剂,如抗生素)。
任选地,套组可以包含2种或更多种不同的噬菌体、3种或更多种不同的噬菌体、4种或更多种不同的噬菌体、5种或6种或更多种不同的噬菌体。
例如,套组可以包含两种噬菌体。例如,spfm10和14;spfm2和4;spfm10和17或者spfm2和10。
例如,套组可以包含三种噬菌体。例如spfm4,10,19;spfm10、17和19;或spfm2、17和19。
例如,套组可以包括四种噬菌体。例如spfm10、14、17、19;spfm2、10、14和19;或spfm2、4、10和19。
例如,套组可以包含多于四种噬菌体。例如,spfm10、14、17和19以及另一种或其他;spfm2、10、14和19以及另一种或其他;或spfm2、4、10和19以及另一种或其他。
例如,套组可以包含spfm2、4、10、14、17和19。
血清型
沙门氏菌菌株按血清分型分类。血清分型包括对核糖体rna基因拷贝的大小和数量的分析,它为每种菌株(称为血清型)建立了遗传简档。血清分型的标准协议使用pcr结合琼脂糖凝胶电泳,或者在并入荧光引物后进行毛细管分析。
本文所述的噬菌体靶向以下五种沙门氏菌血清型:a)25种鼠伤寒沙门氏菌;b)15种s.4,12:i>;c)10种s.4.5,12:i>;d)10种病牛沙门氏菌;和e)10种德尔卑沙门氏菌。这是英国的猪中五种最流行的血清型。这些血清型都表现出多重耐药性。因此,迫切需要新技术,如本文所述的噬菌体。
噬菌体的繁殖和分离
噬菌体的繁殖和分离在实施例部分中进行描述。
热稳定噬菌体
噬菌体在高温下的稳定性可能增加。本文所述的噬菌体在50℃、55℃、65℃、70℃、75℃、80℃、85℃或90℃下可以是稳定的
通过将噬菌体暴露于4℃、10℃、20℃、30℃、40℃、50℃、60℃、70℃、80℃、90℃和100℃的温度下一小时,并在聚合酶链式反应(pcr)机中孵育来测试热稳定性。为了测定噬菌体滴度,将噬菌体裂解物连续稀释10倍,并将小滴噬菌斑测定法用于具有sl1344细菌群的lb1%琼脂板上。最终噬菌体滴度以pfu/ml表示。
稳定意指当暴露于高温下时滴度损失最小。例如,在60℃、70℃、80℃或90℃下使用上述热稳定性试验,损失为0.0至3.5log10pfu/ml或更少。例如,损失为0.0-0.05、0.1、0.3、0.6、0.9、1.0、1.3、1.6、1.9、2.0、2.3、2.6、2.9、3或3.5log10pfu/m。
虽然噬菌体本身可能不稳定,但是保存溶液可以提高它们的热稳定性。因此,热稳定性也可以是指在浸入保存溶液后当暴露于高温下时滴度损失最小。具有与spfm10、14和/或17序列同一性的噬菌体是特别热稳定的。例如,噬菌体与spfm10、14或17具有80、85、90、95或100%的序列同一性。
遗传分析已经揭示了spfm10中特定的单核苷酸差异,这可能是该噬菌体稳定性的原因。这些在下面描述。
在spfm10(预计为果胶酸裂解酶)的以下序列中,在基因组的70890位有一个t而不是a(a存在于本申请中鉴定的热稳定性较差的spfm噬菌体中)。这种多态性位于以下seqidno.1号的第一行,以粗体和下划线显示。这在权利要求书中称为多态性1(从a到t)。
seqidno.1的氨基酸序列如下。从腺嘌呤到苏氨酸的snp将丝氨酸变为苏氨酸,如下面下划线所示(seqidno.9的倒数第二行)。该从丝氨酸到苏氨酸的突变可能使噬菌体更热稳定。
seqidno.9:
在另一种推定的果胶酸裂解酶中已经发现了进一步的多态性。该序列如下(seqidno.2)。在spfm10中,在以下位置处存在单核苷酸多态性:
·74291位:在spfm10中,在该位置处有g。在权利要求书中在该位置处的鸟嘌呤被称为多态性2。
·74294位:在spfm10中,在该位置处有c。在权利要求书中在该位置处的鸟嘌呤被称为多态性3。
·74295位:在spfm10中,在该位置处有g。在权利要求书中在该位置处的鸟嘌呤被称为多态性4。
在下面spfm10的seqidno.2中这些以加粗和下划线显示(参见箭头所指的第二行)。
seqidno:2(3'到5')
seqidno.2的氨基酸序列如下:
seqidno.10:
多态性2:在74291位处从腺嘌呤(如在spfm4和17中发现的)到鸟嘌呤(如在spfm10中发现的)的snp导致从亮氨酸到脯氨酸的变化(在上面ssqwynsppy(seqidno:11))。该脯氨酸的突变可以使spfm10更热稳定。
多态性3:在74294位处从腺嘌呤(如在spfm2中发现的)到胞嘧啶(如在spfm10中发现的)的snp导致从异亮氨酸到丝氨酸的变化(在上面ssqwynsppy(seqidno:12))。该脯氨酸的突变可以使spfm10更热稳定。
多态性4:在74295位处从腺嘌呤(如在spfm2中发现的)到胞嘧啶(如在spfm10中发现的)的snp导致如上面多态性3在相同位置的从异亮氨酸到丝氨酸的变化。同样,seqidno.2中在该位置处的丝氨酸可以使spfm10更热稳定。
沙门氏菌感染
沙门氏菌感染是一种影响肠道的常见细菌疾病。
该套组是兽医用的。也就是说,治疗的对象是动物。
动物可以是猪、鸡、火鸡和/或牛。
沙门氏菌感染可以是由以下血清型菌株中的任一种或更多种引起的:鼠伤寒沙门氏菌、s.4,12:i>、s.4.5,12:i>、病牛沙门氏菌和德尔卑沙门氏菌。这些菌株都表现出多重耐药性。噬菌体可以杀伤来自这些菌株中每一种的80%、85%、90%、95%或100%的分离物。
沙门氏菌感染可以是由以下血清型菌株中的任一种或更多种引起的s.13,23:i>、肠炎沙门氏菌、婴儿沙门氏菌、s.ohio、和s.seftenberg。噬菌体可以杀伤来自这些菌株中每一种的33%、100%、50%、60%或33%的分离物。
药物组合物
该组合物可以包含药学上可接受的赋形剂和/或载体。如本文所用,“药学上可接受的载体”包括任何和所有的溶剂、分散介质、包衣、表面活性剂、抗氧化剂、防腐剂(例如,抗菌剂、抗真菌剂)、等渗剂、吸收延迟剂、盐、防腐剂、药物、药物稳定剂、凝胶剂、粘合剂、赋形剂、崩解剂、润滑剂、甜味剂、调味剂、染料、类似材料及其组合,如本领域普通技术人员所知(参见,例如,remington'spharmaceuticalsciences,第18版.mackprintingcompany,1990,第1289-1329页)。
治疗方法
该噬菌体可用于治疗沙门氏菌的方法,例如,例如动物如猪、牛和家禽,例如鸡。
该组合物可以制备成各种施用途径。例如,该噬菌体或药物组合物可以添加到动物饲料中,例如猪或鸡饲料。例如,动物饲料可以包括压缩饲料,如颗粒。可以使用其他口腔组合物。可替代地,可以将噬菌体添加到喷雾剂中以对围栏或尸体进行消毒。
预防性治疗方法
该噬菌体或噬菌体的药物组合物可以在动物感染沙门氏菌之前给予动物。例如断奶期间的幼猪很容易感染。因此,在这种可能发生的情况下,可以施用预防性治疗。
治疗有效
该噬菌体通常以达到预期目的的有效剂量的方式使用。为了用于治疗或预防疾病状况,以治疗有效量施用或应用本发明的分子或其药物组合物。治疗有效量是有效改善或预防被治疗动物的症状或延长其存活时间的量。治疗有效量的确定完全在本领域技术人员的能力范围内,特别是根据本文提供的详细公开内容。
检测
基于实验室的方法目前用于检测沙门氏菌,例如elisa和基于pcr的方法,但是已经报告这些方法缺乏特异性。本申请中所述的噬菌体可用作诊断沙门氏菌菌株的替代的更具体的方法。
从猪或其他动物身上提取的用于此种诊断测试的样品可以是粪便。然后可以在噬菌体上添加标签。例如荧光标签。一旦噬菌体已经感染/结合了靶沙门氏菌,就可以检测到结合了标签的靶沙门氏菌。例如,通过从样品中洗脱剩余的噬菌体并检测沙门氏菌内结合标签或标签。然后可以评估剩余样品中存在的标签的量。这可以指示存在的细菌细胞的数量。例如,从样品中洗掉剩余的未结合噬菌体后剩余的荧光量。
序列同一性
序列同一性可以与噬菌体保藏物一致,例如与所保藏的噬菌体80%、85%、90%、95%或100%的核苷酸或氨基酸序列同一性。
可替代地,序列同一性可以参照seqidno1至10中的任一个。例如,与seqidno1至10中的任一个80%、85%、90%、95%或100%的核苷酸或氨基酸序列同一性。序列同一性是在整个序列长度上相对于seqidno号进行计算的。
多态性
单核苷酸多态性,在本文中也称为snp,代表单核苷酸的差异。例如,作为snp的结果,在一定长度的dna中核苷酸胞嘧啶(c)可以被核苷酸胸腺嘧啶(t)取代。
干燥噬菌体的方法
该噬菌体可以被干燥以提供给动物和患者更方便的施用方式。干燥也增加了噬菌体的储存寿命。例如,如果将经干燥的噬菌体添加到饲料中,则不会添加额外的水分。这还延长了产品的保质期。
在准备干燥时,可以将噬菌体浓缩至约1010pfu/ml或以上的滴度。
将噬菌体与保存溶液混合,以在干燥期间保护它们。如下所述,保存溶液可以包含糖和任选的氨基酸和聚合物。
该噬菌体可以以1:10v/v的浓度(噬菌体:混合物)与溶液混合。
糖/糖醇
保存组合物中可以使用一种或更多种糖。例如葡萄糖或基于葡萄糖的糖如甘露醇。基于葡萄糖是指包含葡萄糖的二糖或多糖。
糖可以是二糖。
保存溶液可以包含单独的糖醇或除了1种或更多种糖之外的糖醇。合适的糖醇包括甘露醇。其他6种碳糖醇也是合适的。
例如,保存溶液可以包含海藻糖、甘露醇和亮氨酸中的一种或更多种。
氨基酸
除了糖和/或糖醇之外,一种或更多种氨基酸也可以添加到保存溶液中。
氨基酸可以是非极性脂肪族氨基酸。例如,氨基酸可以是亮氨酸、异亮氨酸、甘氨酸、丙氨酸、脯氨酸、缬氨酸、蛋氨酸或苯丙氨酸。
聚合物
聚合物也可以添加到保存溶液中,以帮助包裹噬菌体并确保它们在动物肠道中不被杀伤。该聚合物还可以帮助噬菌体靶向到需要它们工作的肠道。
该聚合物可以包含衍生自丙烯酸酯和甲基丙烯酸酯的共聚物。该聚合物可以包含酸性或碱性基团,其能够使活性成分依赖于ph释放。
示例保存溶液
保存溶液可以包含海藻糖、甘露醇和亮氨酸。
例如,保存溶液可以包含5%、6%、7%、8%、9%、10%、11%、12%、13%、14%、15%、16%、17%、18%、19%或20%或更多的海藻糖,5%、6%、7%、8%、9%、10%、11%、12%、13%、14%或15%的甘露醇和0.5%、1%、2%、3%、4%或5%的亮氨酸。例如8%海藻糖、8%甘露醇、1%亮氨酸。
被称为eudragits100的聚合物(购自德国evonikhealthcare)被发现还有助于保护噬菌体。例如,向包含8%海藻糖、8%甘露醇和1%亮氨酸的保存溶液中添加2%eudragit。
干燥混合物
然后将所得的噬菌体和糖以及任选的氨基酸的溶液干燥以形成粉末,该粉末可被压实以形成片剂或丸剂。
为了干燥溶液,可以使用喷雾干燥。
用于干燥粉末的温度可以为70℃、75℃、80℃、85℃、90℃、95℃或100℃噬菌体的粉末通过干燥噬菌体产生。
该方法产生的噬菌体
因此,噬菌体/组/药物组合物或动物饲料可以额外包含上述糖、糖醇、氨基酸和/或聚合物组合中的任一种。
例如,噬菌体溶液或粉末(干燥后)可以包含上述糖、糖醇、氨基酸和/或聚合物组合中的任一种。例如,包含海藻糖、甘露醇和亮氨酸的噬菌体溶液或粉末。
在整个说明书中,除非上下文另有要求,否则术语“包括”或“包含”或如“包括”或“包含”的变体应理解为暗示该方法或试剂盒包括所述整数或整数组,但是不排除任何其他整数或整数组。
在本文中引用的每个文件、参考文献、专利申请或专利明确地通过引用整体并入在本文中,这意味着读者应该将其作为本文的一部分来理解和考虑。本文中引用的文件、参考文献、专利申请或专利不重复,在本文中仅仅是出于简明的目的。提及本文中引用的材料或所含信息不应被理解为该材料或信息是常识的一部分或在任何国家已知。
附图说明
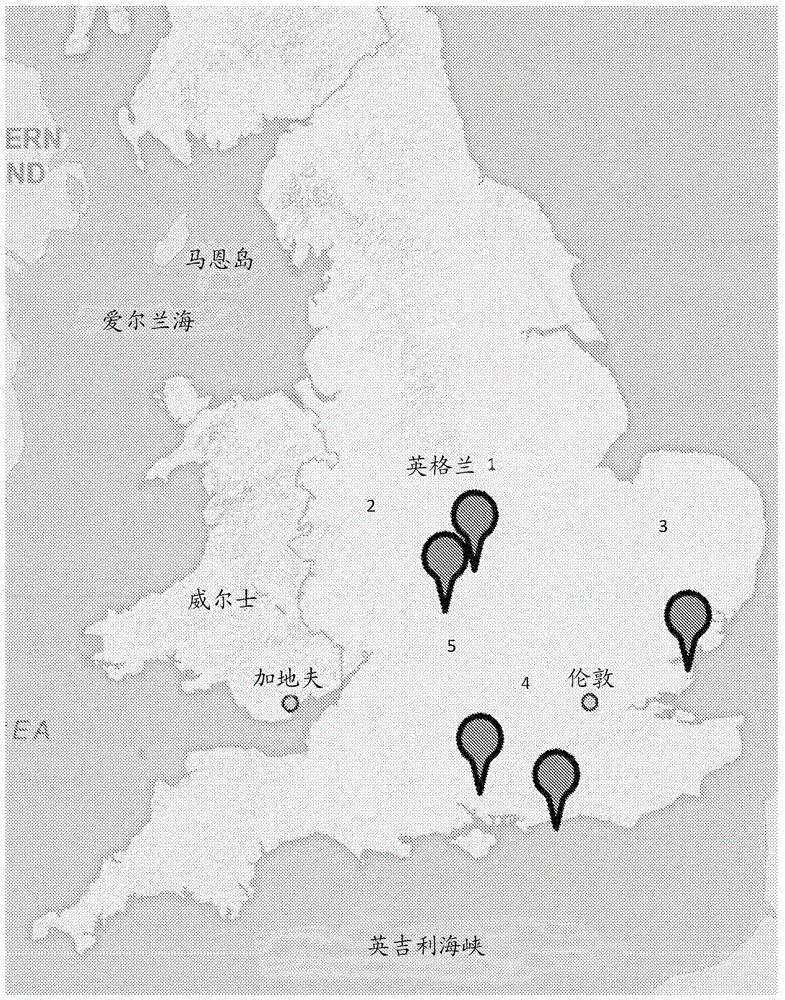
图1示出了在英国分离spfm沙门氏菌噬菌体的样品位点及其表型结构。
a.多个样品来源于分离噬菌体的英国。(1)汉普郡的野猪自然保护区(spfm1–spfm3)、(2)埃塞克斯郡的食品加工厂(spfm4–spfm11)、(3)沃里克郡沃里克的养猪场(spfm12–spfm15)、(4)莱斯特郡欣克利的养猪场(spfm16–spfm20)和(5)西苏塞克斯郡的猪自然保护区(spfm21–spfm22)。英国地图来源于
图2示出了21种spfm沙门氏菌噬菌体的宿主范围分析及其接种效率。a.宿主范围分析基于spfm噬菌体的裂解谱,针对多重耐抗生素菌株:25种鼠伤寒沙门氏菌、15种s.4,12:i>、10种s.4,5,12:i>、10种病牛沙门氏菌和10种德尔卑沙门氏菌,并且完全裂解分别地通过橙色条、红色条、绿色条、蓝色条和紫色条(从左至右)给出。所有菌株是从猪中分离的。浅橙色条(左侧栏)显示浑浊的清液,浅紫色(右侧条)显示无感染。b.通过主成分分析法分析了spfm噬菌体对11种沙门氏菌分离株的接种效率和给出的数据。该双标图代表了方差最大(总方差为59.3%)的两种主要成分,来自五种猪相关沙门氏菌血清型的两个代表性分离株的spfm噬菌体(标记为黑色环)的eop和噬菌体繁殖宿主鼠伤寒沙门氏菌sl1344(总共11种菌株)的eop。噬菌体的颜色取决于它们的eop,其中红色(左边)的噬菌体在所有分离株上显示相同的eop,绿色(右边)的噬菌体在特定菌株上具有更高的eop,并且位置更接近单个菌株(黑色箭头和文字)。对于宿主范围分析(a)和eop分析(b),对所有菌株进行了三个生物学重复,所提供的数据是所有三个重复的平均值。
图3.spfm噬菌体的一个代表性实例并且给出噬菌体spfm1的基因组。内部黑色环表示gc含量,外部环表示预测的开放阅读框,刻度单位为碱基对。属于不同类别的功能基因被相应地着色:红色是结构蛋白,绿色是参与dna复制和转录的基因,黄色是包装基因,橙色是裂解基因,紫色突出显示额外基因,并且trna被标记为蓝色(从最右侧栏的第二项)。未标记的orf代表假设的蛋白质。
图4.基于辅助基因的存在和缺失通过分级聚类分析产生的的21种spfm沙门氏菌噬菌体的谱系图。基于46个辅助基因的存在和缺失数据,计算噬菌体之间相异度的量度jaccard距离,并分级聚类。单位0表示存在相同的基因,1表示变异最大。
图5.a.ncbi数据库中158种全基因组测序沙门氏菌噬菌体的成对平均核苷酸同一性(ani)值的热图,包括本研究中的21种spfm噬菌体。数值范围从0(0%)ani到1(100%ani),并且分别用紫色到绿色和黄色编码,后者代表高度相似的噬菌体簇,分支代表不同的簇。spfm噬菌体在热图上都聚集在一起,并且簇设置在右上方(实心黄色框,由白色箭头指示)。
图6.系统基因组树显示了21种spfm噬菌体和簇内两种相似的肠道细菌噬菌体segd1和spn3us之间的关系。该树是从所有23种测序噬菌体共有的188个核心基因的密码子比对中获得的单核苷酸多态性(snps)构建的,该树是按比例绘制的。
图7.沙门氏菌噬菌体在不同温度(a)和ph(b)下的稳定性。噬菌体在不同的温度和ph下孵育一小时,并且在其繁殖宿主上滴定。噬菌体spfm2的稳定性以蓝色b显示,spfm4以紫色p显示,spfm10以红色r显示,spfm14以绿色g显示,spfm17以橙色o显示,spfm19以银色s显示(沿图的顶部显示顺序的颜色参考)。给出的数据是三个生物学重复的平均值,每个重复有三个技术重复,误差条代表平均值的标准误差(sem)。另参见表4。
图8.1、2、3和4种噬菌体混合物的不同组合在降低体外mdrs01160-12鼠伤寒沙门氏菌上的功效。用不同的噬菌体混合物(a-f)以100的moi感染细菌培养物,并在6小时内每小时测定细菌计数(cfu/ml)。给出的数据是三个生物学重复的平均值,每个重复有三个技术重复,误差条代表sem。
图9.噬菌体混合物对幼虫感染模型中的mdrs01160-12鼠伤寒沙门氏菌的功效。测试的噬菌体混合物为:spfm10-spfm14的2-噬菌体(红线/条(r));spfm4-spfm10-spfm19的3-噬菌体(蓝线/条(b));和4-噬菌体spfm2-spfm10-spfm14-spfm19(绿线/条(g))。用沙门氏菌以105cfu/幼虫时感染幼虫,或者在感染前1小时用噬菌体混合物进行预防性治疗p-1至p-3(a、c和e),或者同时施用噬菌体和沙门氏coi-1至coi-3(b、d和f)。(a)和(b)表示幼虫存活超过72小时。橙色线表示在组c-1中未感染的健康幼虫,灰色线表示在组c-2中仅施用pbs的幼虫,黑色线表示在组c-6中仅感染沙门氏菌的幼虫组c-6的存活。方框图(c)和(d)(颜色从左到右顺序为黑色、红色、蓝色、绿色)表示了不同幼虫治疗组中沙门氏菌计数的变化,黑色条表示仅感染沙门氏菌的幼虫计数。方框图(e)和(f)(颜色从左到右顺序为红色、蓝色、绿色)表示了用于预防性和共感染治疗的噬菌体混合物的噬菌体计数。误差条表示图中显示了仅感染沙门氏菌且用噬菌体治疗的幼虫之间的sem和统计学显著差异(*p≤0.05,**p≤0.01和***p≤0.001)。
图10.噬菌体对从幼虫感染模型中分离的沙门氏菌菌落的成斑率。幼虫感染mdrs01160-12鼠伤寒沙门氏菌菌株,并且用2-噬菌体(spfm10和spfm14)、3-噬菌体(spfm4、spfm10和spfm19)或4-噬菌体混合物(spfm2、spfm10、spfm14和spfm19)治疗。如果72小时后回收沙门氏菌,则挑选菌落,并且对于每组中的10个菌落,对混合物中的噬菌体评估噬菌体成斑率。然后将成斑率与野生型菌株上噬菌体的成斑率比较。红色误差条代表用2-噬菌体混合物治疗幼虫后挑选的菌落,蓝色误差条代表3-噬菌体混合物,绿色误差条代表4-噬菌体混合物。平均成斑率由三次生物复制显示,误差条代表sem。
图11提供了表a。当与噬菌体spn3us中带注释和预测的rnap比较时,所有spfm噬菌体中存在的病毒(v)rnap和非病毒(nv)rnap亚基。
图12提供了表b。用于构建图5中热图的所有沙门氏菌噬菌体基因组及其在热图中顺序的列表。
实施例
现在将仅通过示例的方式并参考以下实验来说明本发明的方面和实施方案。
数据给出本研究中分离出21种沙门氏菌噬菌体,称为spfm噬菌体的表征。针对从英国农场爆发中分离出的流行的和相关的循环多重耐药性英国猪沙门氏菌菌株,测试噬菌体杀伤活性。测定了所有spfm噬菌体的接种效率和宿主范围。他们也被测序,通过系统基因组分析进行研究,与ncbi数据库中所有以前测序的沙门氏菌噬菌体进行比较,并鉴定出其阳性选择基因。这是第一项对靶向英国猪相关沙门氏菌菌株的大量噬菌体进行全面表征和测序的研究。
实验程序
细菌菌株和培养条件
本研究总共使用了68种沙门氏菌菌株。肠道沙门氏菌亚种肠道鼠伤寒沙门氏菌血清型sl1344(登记号fq312003)被用作参考菌株。剩下的67种肠道沙门氏菌亚种肠道菌株是在2012年至2015年间英国威布里奇的动物和植物健康机构(apha)从英国的猪中分离出的肠道毒株。这些菌株的血清型为:22鼠伤寒沙门氏菌;15s.4,12:i>;10s.4,5,12:i>;10病牛沙门氏菌;和10德尔卑沙门氏菌。所有菌株对以下抗生素中的至少一种有耐药性:萘啶酸、四环素、新霉素、氨苄西林、呋喃唑酮、头孢他啶、磺胺甲噁唑、甲氧苄啶、氯霉素、阿米卡霉素、阿莫西林/克拉维酸、庆大霉素、链霉素、复方磺胺、头孢噻肟、安普霉素和环丙沙星。
所有沙门氏菌菌株保存在-80℃的50%甘油肉汤(英国abtekbiologicalsltd.)中。菌株通常在木糖赖氨酸脱氧胆酸盐(xld)琼脂(oxoid,英国)上在37℃下培养18小时,然后在nzcym肉汤(英国melfordbiolaboratoriesltd)中以100rpm在37℃下再培养18小时。
噬菌体的分离、纯化和繁殖
所有样品都是从2015年7月至12月在英国的五个地理位置采集的。总共15个样品采集自汉普郡的自然野猪保护区;10个样品采集自埃塞克斯的食品加工厂;15个样品采集自沃里克郡的精加工养猪场;18个样品采集自莱斯特郡的小猪和精猪的养猪场;并且7个样品采集自西苏塞克斯的野生猪自然保护区(图1a)。
使用相同的富集程序加工所有样品,将1ml或1克样品与9mlnzcym肉汤混合,并添加100μl指数生长的沙门氏菌培养物。为了使噬菌体分离最大化,将相同的样品等分并分别用12种不同的mdr沙门氏菌宿主菌株富集。富集物在37℃下振荡孵育12小时(100rpm),然后样品在室温下以4000×g离心15分钟。上清液通过0.22μm孔径的注射器过滤器(millipore,英国)过滤,并且过滤后的样品储存在4℃下,直到进一步使用。通过小液滴噬菌斑测定法(mazzocco等,2009)筛选滤液中的噬菌体。简而言之,在luria-bertani(lb)1%(w/v)琼脂板(fisherscientific,英国)上点样10μl富集的样品,nzcym0.5%(w/v)琼脂作为顶层,与100μl指数生长的沙门氏菌培养物混合。将板在37℃孵育18小时,并通过清除或噬菌体斑的存在来检查噬菌体裂解。
为了噬菌体纯化,用1μl环挑选单个噬菌斑,与500μl含明胶(100mmnacl,8mmmgs04.7h2o,50mmtris-cl和0.01%(w/v)明胶)的sm缓冲液混合,并以21,000×g离心10分钟。所得上清液用于下一轮通过双琼脂覆盖噬菌斑测定法(kropinski等,2009)的噬菌体纯化,并且该过程重复7次以产生克隆噬菌体储备物。
通过在nzcym肉汤中于37℃下振荡(100rpm)6小时,将感染了107pfu/ml噬菌体的沙门氏菌的sl1344(107cfu/ml)的指数生长液体培养物混合,来制备增加体积的纯化噬菌体裂解物。将噬菌体培养物以4,200xg离心15分钟,用0.22μm孔径的过滤器过滤上清液,并将噬菌体裂解物在4℃下储存。为了测定噬菌体滴度,将噬菌体裂解物连续稀释10倍,并在1%lb琼脂板上使用小液滴噬菌斑测定法(mazzocco等,2009)。最终噬菌体滴度以pfu/ml表示。
透射电子显微术
在透射电子显微术(tem)分析之前,通过以21,000×g离心1小时浓缩噬菌体裂解物,将沉淀物用0.1m乙酸铵(fisherscientific,英国)重悬,以21,000×g离心1小时,并用0.1m乙酸铵溶液重悬。高浓度噬菌体(1011pfu/ml)用1%乙酸铀酰(w/v)负染色10秒,并应用于3mm碳涂覆的铜网格(琼脂科学有限公司,英国)。用80kv的eol1220(joelukltd,英国)ran检测噬菌体,并通过带有分析软件(奥林巴斯软件成像解决方案,德国)的sismegaviewiii相机获得图像(ackermann2009)。tem分析由英国莱斯特大学核心生物技术服务部的aliali博士和natalieallcock进行。
噬菌体宿主范围分析和成斑率(efficiencyofplating)
通过小滴噬菌斑测定法(mazzocco等,2009)在不同的肠道沙门氏菌亚种肠道血清型上确定单个噬菌体的宿主范围并在37℃下孵育18小时。℃检查板是否有细菌裂解(通过清除或噬菌斑)或没有感染,并从三个生物学重复中观察到平均观察结果,每个重复有三个技术重复。
成斑率(eop)在来自以下5种沙门氏菌血清型的两株代表性mdr菌株上进行:鼠伤寒沙门氏菌;s.4,12:i>;s.4.5,12:i>;病牛沙门氏菌;和德尔卑沙门氏菌。在噬菌体繁殖宿主s1344上也建立了eop。对于eop,使用小滴噬菌斑测定法,并且将噬菌体裂解物连续稀释10倍并点在细菌菌苔上(kutter,2009)。平均pfu/mleop值是根据三个生物学重复进行的,每个重复有三个技术重复。对所有噬菌体在使用rbase软件包(rcoreteam,2017)中的prcomp函数筛选的所有菌株上的eop值进行了主成分分析,并通过ggplot2软件包中的autoplot函数绘制为双标图(wickham,2009)。
噬菌体dna提取,测序和注释
采用改良的苯酚-氯仿-异戊基方法,使用高滴度噬菌体裂解物(1010pfu/ml)来提取dna(nale等,2016)。提取后,将最终dna沉淀物溶解在5mmtrishcl中,使用qubit荧光计和qubit双链hs试剂盒(thermoscientific,英国)进行定量,并通过illuminamiseq平台进行测序。nexteraxt文库是根据制造商的说明书使用1ng输入dna制备的。在使用spadesv3.9.1(bankevich等,2012)进行组装之前,使用sicklev1.33(joshi和fass,2011)对读取进行修剪。使用prokkav1.11(seemann2014)用从所有噬菌体蛋白质构建的定制数据库(2017年12月)和hmmscan进行注释,以鉴定pvog(grazziotin等,2017)。表1列出了分配给噬菌体基因组的登录号。将噬菌体基因组上载到欧洲核苷酸档案馆(ena)时出错,并且噬菌体名称不正确。即使这样,表1中列出的登录号也是正确的,并且允许区分噬菌体。ena当前正在解决该问题,并且可以通过该链接http://s3.climb.ac.uk/sinfo/spfm_genome.tar.gz找到具有正确名称的正确文件。
计算沙门氏菌噬菌体基因组之间的ani
从该研究中新测序的两个噬菌体和先前研究中测序的噬菌体中收集了158株来自沙门氏菌感染噬菌体的基因组。使用pyani软件包中的blastn比对选项(pritchard等,2016),在噬菌体基因组的所有成对组合之间测量了由(goris等,2007)定义的平均核苷酸同一性,并使用heatmaply在交互式热图中绘制(galili等,2017)。
蛋白质直系同源物聚类
使用在get_homologs.pl脚本(vinuesa和contreras-moreira,2015)中执行的cog三角形算法(kristensen等,2010)采用blastpe-value阈值1e-5将prokka注释的噬菌体蛋白质序列聚类到直系同源物组中。每个噬菌体基因组之间的jaccard距离基于基因的存在/不存在,该基因的存在/不存在由噬菌体蛋白质序列的cog聚类确定。使用基础rdist函数计算jaccard距离,使用基础rhclust函数根据距离构建树状图,并使用r中的ggdendro软件包绘制树状图(vries和ripley,2016)。ortholog聚类数据显示在补充表4中,如在thankiam、brownn、millardad、clokiemrj.genomiccharacterizationofjumbosalmonellaphagesthateffectivelytargetunitedkingdompig-associatedsalmonellaserotypes.frontmicrobiol.2019;10:1491.2019年7月2日发表,doi:10.3389/fmicb.2019.01491.中发表的。
假设基于核心基因的系统基因组树
用musclev3.8.31(edgar,nar32(5))和pal2nal.pl脚本(suyama等,2006),使用包含汉城病毒属噬菌体基因组蛋白质序列的cog来构建相应核苷酸序列的密码子识别比对。来自密码子识别核苷酸比对的snp是使用snp位点等提取的(page等,2016),并使用fasttreev.2.1.10sse3(price等,2010)并用核苷酸进化的广义可逆模型将其用于构建系统发育树。
测量阳性选择
使用hyphy软件包在每个直系同源直系同源簇的每个密码子上进行阳性选择测试(pond等,2005)。特别地,来自musclev3.8.31的每个直系同源直系同源簇的蛋白质比对用于指导构建来自每个簇的核苷酸序列的密码子识别比对。使用hyphy软件包中执行的gard算法(pond等,2006)检测每个密码子识别比对中的重组断裂点,并在每个断裂点的任一侧构建分区的比对和树。使用hyphy软件包中执行的bayesianfubar算法(murrell等,2013)对这些分区在每个位点的阳性选择进行了测试,该算法返回每个位点处于阳性选择下的后验概率(即,dn/ds大于1)。由于样品大小太小,无法准确评价单个密码子的dn/ds,因此后验概率值与dn/ds值一起给出。该假设检验足够灵敏以检测阳性选择,psrf/n有效比保持低于0.006,使阳性选择的后验概率收敛于一个解决方案。
结果
几项体外和体内研究已经表明,噬菌体可以显著减少食品餐具和猪中的沙门氏菌属的数量,并且因此可以为减少猪中的沙门氏菌属提供可行的选择来提高食品安全性。然而,到目前为止,并不是所有已发表的研究都从感染性和测序的方面充分表征了它们的噬菌体。测序对于治疗中使用的噬菌体是有用的,因为它可以证实噬菌体是专性裂解的,它们不携带毒素基因,它们不属于已知的促进宿主抗生素抗性基因传播并且有助于合理优化噬菌体治疗混合物的特定噬菌体属(例如felixo1病毒属)。本发明人已经表征了从环境源分离的21种新的裂解性巨型spfm沙门氏菌噬菌体,其再次靶向通常与猪相关的沙门氏菌属。所有spfm噬菌体对从猪中分离的临床相关英国mdr沙门氏菌菌株都具有广泛的宿主范围和高eop值,并且可用于噬菌体治疗,可以有助于提高食品安全性。
噬菌体的分离,噬菌斑形态和繁殖
在2015年7月至2015年12月之间,从食品加工厂、英国野生和家养猪和野猪中收集了用于噬菌体分离的样品。筛选了65个样品,分离出21种噬菌体,分别命名为spfm1至spfm22。由于其基因组序列不完整,将噬菌体spfm18排除在分析之外。在五种不同的沙门氏菌肠道血清型鼠伤寒沙门氏菌菌株上分离了15种spfm噬菌体,其余的7种在沙门氏菌肠道血清型4,12:i>菌株上分离。三种噬菌体源自野猪粪便,八种源自食品加工厂,十种源自从沃里克郡、欣克利和西萨塞克斯郡的农场获得的精猪和幼猪粪便(图1a和表1)。
在噬菌斑形态方面,噬菌体spfm9、spfm10和spfm11是在鼠伤寒沙门氏菌菌株上分离的和产生清楚的约1mm的直径噬菌斑,并且剩下的18种噬菌体产生直径约0.5mm的清楚噬菌斑。所有噬菌体传播在鼠伤寒沙门氏菌sl1344上繁殖以产生高滴度(1010噬菌斑形成单位(pfu)/ml)的原液。这样的主要动机是原始分离菌株自发地释放了噬菌体,这使下游的表征复杂化,而从未观察到从sl1344释放噬菌体。
表1.分离的噬菌体及其相应的分离宿主、英国来源和基因组信息的总结。
a样品来源编号与图1相关:(1)汉普郡的野猪自然保护区、(2)埃塞克斯郡的食品加工厂、(3)沃里克郡沃里克的养猪场、(4)莱斯特郡欣克利的养猪场、(5)西苏塞克斯郡的猪的自然保护区。
b通过该链接可以找到所有基因组序列:http://s3.climb.ac.uk/sinfo/spfm_genome.tar.gz。
噬菌体形态
tem分析显示,这21种噬菌体具有等长的头部和可收缩的尾巴,因此归类为caudovirales中肌病毒科家族的成员(图1b)。噬菌体spfm1、spfm2、spfm6、spfm7、spfm10、spfm12、spfm14、spfm15、spfm16、spfm17、spfm20和spfm22的尾巴长度为160±20nm,衣壳直径为100±15nm。噬菌体spfm3、spfm4、spfm5、spfm8、spfm9、spfm11、spfm19和spfm21的尾巴长度为200±20nm,衣壳直径为105±15nm。噬菌体spfm13的尾巴最长,为140±10nm,衣壳直径为135±7nm。
宿主范围分析
在从猪中分离的67种mdr肠道沙门氏菌亚种肠道菌株上测试了spfm噬菌体的功效。全部67种菌株代表了五大英国猪相关的沙门氏菌血清型,即鼠伤寒沙门氏菌;s.4,12:i>;s.4.5,12:i>;病牛沙门氏菌;和德尔卑沙门氏菌。所有噬菌体具有广泛的活性裂解谱,每株均裂解超过80%的菌株(图2a)。噬菌体spfm1、spfm3、spfm10、spfm14、spfm15、spfm17和spfm19可以裂解100%筛选的沙门氏菌菌株。噬菌体spfm2、spfm7、spfm21和spfm22裂解67/68种菌株(99%)。相反,spfm9和spfm11分别仅裂解84%和81%的菌株,即使在裂解细菌的地方,在至少一种菌株上裂解物是混浊的。
成斑率(eop)
为了确定在临床上相关菌株上噬菌体感染的效率,从而决定最终应该组成噬菌体混合物的噬菌体,在沙门氏菌血清型鼠伤寒沙门氏菌;s.4,12:i>;s.4.5,12:i>;病牛沙门氏菌;和德尔卑沙门氏菌的两种代表性mdr猪菌株上进行对spfm噬菌体的eop。噬菌体在其繁殖宿主鼠伤寒沙门氏菌sl1344上的eop被包括作为对照。为了鉴定eop数据集中的模式,使用了主成分分析(pca),这显示噬菌体聚集类成两个独特的组(图2b);(i)在所有菌株中均具有相似的eop的噬菌体;以及(ii)在特定沙门氏菌菌株和血清型中具有较高eop的噬菌体。通过t检验证实了,噬菌体spfm4、spfm5、spfm8、spfm9、spfm12、spfm13、spfm14、spfm15、spfm16、spfm17和spfm20形成的组(i)及其eop在所有五种血清型之间均无显著差异。剩余的10种噬菌体形成组(ii)。为了扩大组(ii),噬菌体spfm10对病牛沙门氏菌分离物a具有更高的eop;噬菌体spfm1,spfm11和spfm12在所有病牛沙门氏菌菌株上具有更高的eop。噬菌体spfm19和spfm21在s.4,12:i>菌株上均具有较高的eop;spfm3在德尔卑沙门氏菌分离物b上具有较高的eop;spfm22在s.4,5,12:i>分离物a上具有较高的eop;以及噬菌体spfm6和spfm7二者在德尔卑沙门氏菌分离物上具有较高的eop。
spfm噬菌体的基因组表征
使用illuminamiseq平台对spfm噬菌体组进行测序。所有21种噬菌体具有233kb至242kb的线性、环状排列的dsdna基因组,并且编码258至307个编码序列(cds)(表1)。由于所有基因组均大于200kb,因此它们被归类为巨型噬菌体,并且由于缺乏已知的溶原相关基因而被预测为可溶的。
尽管spfm基因组大小不同,但是它们都编码一种trna,平均基因组含量为48.5%(表1),平均基因长度为0.860±0.010kb,基因密度为1.074/kb,基因编码区域占其基因组的93%。由于噬菌体的结构和含量在遗传上相似,spfm1的代表性基因组图显示在图3中。大多数预测的基因编码没有已知功能的蛋白质,假定的作用只能分配给约30%的基因。可通过与其他噬菌体的同源性识别的基因包括那些编码结构蛋白的基因,例如主要衣壳蛋白、尾纤维蛋白和尾鞘蛋白。编码包装蛋白终止酶的基因也被鉴定,如噬菌体裂解酶也被鉴定。可以鉴定出编码参与dna复制和转录的产物的几个基因,例如内氧核糖核酸酶、解旋酶、推定的核酸酶sbccdd亚基、推定的核糖核酸酶h和六种rnapβ(β/β')多亚基。编码rnap多亚基的基因的长度从0.240至4.206kb不同,并且所有六种亚基与沙门氏菌spn3us(登录号:jn641803.1),欧文氏菌噬菌体vb(登录号:kx397364.1)和cronobacter噬菌体cr5(登录号:jx094500)的平均核苷酸同一性(ani)为99%%、80%和55%((lee等2016)。
在所有spfm噬菌体基因组中发现了其他基因,这有可能减轻它们在感染期间对其细菌宿主的依赖。特别感兴趣的基因包括二氢叶酸还原酶、胸苷酸合酶和胸苷酸激酶,所有这些都被预测用于叶酸合成和参与在噬菌体感染期间增强宿主代谢的自由基s-腺苷甲硫氨酸(sam)基因(lee等,2011)。此外,在spfm噬菌体中发现了dna腺嘌呤甲基化酶基因,该基因可以抵御宿主的限制性修饰系统。
分离的spfm噬菌体的层次聚类分析
使用成对的局部比对用核苷酸blast(ncbi,1988)对基因组进行了比较,并且观察到所有spfm噬菌体在遗传上彼此非常相似,ani约为95%。为了深入了解基因组内的变异,根据46个共享辅助基因的存在或不存在,建立了树状图(图4)。层次聚类分析鉴定了噬菌体spfm5、spfm15、spfm17、spfm19、spfm21彼此最相似。噬菌体spfm2、spfm6、spfm9、spfm12和spfm14组成了第二组,并且共享相同的辅助基因和核心基因。剩余的11种噬菌体证实了辅助基因之间的变异,并且最大的差异在于噬菌体spfm1、spfm13、spfm16与spfm20之间。与其他spfm噬菌体相比,所有这些的基因组大小也不同(表1)。
spfm噬菌体与先前测序的沙门氏菌噬菌体的比较
为了确定spfm噬菌体与先前保藏在genbank中(直到2017年12月)的沙门氏菌噬菌体的相似程度,针对158个完全测序的沙门氏菌噬菌体基因组进行了全盘比较分析(图5)。用于分析的所有沙门氏菌噬菌体的基因组大小范围为约33至240kb,表b中列出了用于分析的所有噬菌体。聚类分析鉴定了21个不同的组,其中一个聚类定义为与该聚类的其他成员共享>50%ani的噬菌体。所有21种spfm噬菌体均与在不同研究中分离出的噬菌体spn3us(登录号:jn641803.1)(lee等,2011)和segd1(登录号:ku726251.1)组在一起。噬菌体spn3us和segd1的基因组也为约240kb,与所有spfm噬菌体具有约95%-97%的ani%。噬菌体spn3us和segd1是spn3us病毒属的一部分(adriaenssens等,2017),现在已更新为汉城病毒属。这是基于目前95%以上的ani的标准,spfm噬菌体也归类为该属。汉城病毒属簇中的噬菌体被归类为phikz样噬菌体。phikz样噬菌体的显著特征是它们都包含两种多亚基rna聚合酶。第一多亚基被包装在衣壳中作为“病毒体”rnap,期在噬菌体转录程序中转录早期基因,第二“非病毒体”rnap在其基因组中负责转录晚期基因。
汉城病毒属噬菌体簇
为了确定spfm噬菌体与沙门氏菌噬菌体spn3us和segd1的亲缘关系如何,基于共享的188个核心基因内的单核苷酸多态性(snp)构建了系统发育分析(图6)(数据已在补充表3中提供,如在thankiam,brownn,millardad,clokiemrj.genomiccharacterizationofjumbosalmonellaphagesthateffectivelytargetunitedkingdompig-associatedsalmonellaserotypes.frontmicrobiol.2019;10:1491.2019年7月2日发表doi:10.3389/fmicb.2019.01491/中发表的)。这显示噬菌体spfm5、spfm9、spfm10和spfm11共享相同的snp。根据核心基因中snp进行的噬菌体聚类与基于辅助基因存在或不存在的分层聚类分析不同。其他噬菌体(除spfm1之外)归为一组,其核心基因变化很小或几乎没有变化。spfm1核心基因的snp导致该噬菌体与其他20种spfm噬菌体归为独立的分枝。噬菌体spn3us和segd1形成与spfm噬菌体不同的更紧密的次分枝(图4)。
阳性选择下的噬菌体基因
为了确定哪些基因处于进化选择压力下,在汉城病毒属簇中鉴定出阳性选择下的基因。这是通过确定21种spfm噬菌体、spn3us和segd1的核心直系同源直系同源物的成对比较的非同义与同义取代率/之比(dn/ds)来进行的(补充表5,如在thankiam,brownn,millardad,clokiemrj.genomiccharacterizationofjumbosalmonellaphagesthateffectivelytargetunitedkingdompig-associatedsalmonellaserotypes.frontmicrobiol.2019;10:1491.2019年7月2日发表doi:10.3389/fmicb.2019.01491/中发表的)。表2中显示的数据说明了在阳性选择下的预测基因及其直系同源簇编号的大于0.900的后验概率值。分析预测33个基因处于阳性选择下。这些基因中的22个是推定的毒粒结构蛋白,一个是推定的内脱氧核糖核酸酶rusa,两个编码胸苷酸合酶,并且八个是假设的蛋白。对于阳性选择下的推定的毒粒结构蛋白,使用hhpred(zimmermann等,2017)确定结构蛋白是否参与噬菌体尾巴或衣壳组装。该程序以超过75%的概率预测它们是推定的基板楔形蛋白,因此可能参与噬菌体尾部组装。另外,直系同源簇编号201(与补充表5中的直系同源簇编号有关)对c-末端果胶酸裂合酶结构域具有98.26%的概率命中,这也是噬菌体尾纤维形成的一部分。并非所有阳性选择下的基因都可以分配功能,但是可以预测以下直系同源簇编号106、108、129、130和211的推定蛋白可能是推定的毒粒结构蛋白,因为在基因组上,它们的位置与其他推定的结构蛋白接近。类似地,具有直系同源簇编92的假设蛋白质位于靠近dna复制和转录基因聚类的位置,因此可能参与该功能。
表2.在汉城病毒噬菌体簇中阳性选择下的基因,包括噬菌体spn3us、segd1和21spfm噬菌体。
a使用工具hhpred(zimmermann等2017)重新注释蛋白质序列,并且呈现了超过75%可能性的注释。
b样品量太小,无法准确评价单个密码子的dn/ds,但是假设检验(后验概率)足够灵敏以检测到阳性选择。
本发明人已经表征了21种新的沙门氏菌噬菌体。所有噬菌体均可感染从猪中分离出的代表性mdr菌株。鉴定了七种有毒的噬菌体,它们可以感染100%的测试的沙门氏菌分离物。这些噬菌体的高感染性的可能解释可能是因为噬菌体感染鼠伤寒沙门氏菌,它们还可以感染其单相变体,这些变体在遗传上密切相关(moreno等,2013)。噬菌体可能使用相同的细菌受体来附着这些血清型的菌株。基于它们的宿主范围和成斑率,这七种噬菌体似乎是治疗用途的理想选择。
两种噬菌体spfm9和spfm11仅可以感染约80%筛选菌株。另外,两种噬菌体在五分之一的菌株上产生浑浊的清除,这可能表明潜在的溶源性,因为浑浊的清除通常是温带噬菌体的特征(gallet等,2011)。尽管序列分析证实了两种噬菌体没有已知的溶菌模块,并且可能是裂解噬菌体,因为只有约30%的基因具有已知功能,可能存在未知的溶菌模块。关于为什么观察到浑浊清除的可能解释可能是筛选的细菌菌株可能部分地对噬菌体具有抗性,因此仅感染了亚细胞群,从而导致浑浊清除(bull等,2014)。替代的解释可能是噬菌体在这些菌株中诱导了前噬菌体,这可能已经产生了浑浊的清除(campoy等,2006)。尽管观察到浑浊的清除,但是spfm9和spfm11能够在筛选出的11株用于eop分析的菌株上感染并复制,这表明它们是溶菌性感染。由于潜在的不完全裂解以及对噬菌体的抗性或诱导,噬菌体spfm9和spfm11将不是治疗的良好候选物(chan等,2013;abedon等,2017)。
为了进一步表征spfm噬菌体并缩小其具有理想的治疗使用特性,进行了eop分析(mirzaei和nilsson,2015)。数据显示了一组噬菌体在来自五种不同血清型的所有代表性菌株中的eop均无差异,并且由于菌株之间的感染性也无差异,因此这些噬菌体似乎是治疗应用的理想候选者。特别是噬菌体spfm14、spfm15和spfm17是良好的候选者,因为它们可以感染来自主要英国沙门氏菌血清型的所有代表性菌株,并且在筛选出的菌株上具有较高的eop。
所有分离的spfm噬菌体的基因组大于233kb,并且显著增加ncbi上约170个巨型噬菌体基因组的数量和多样性(yuan和gao,2017)。与其他沙门氏菌噬菌体相比,spfm噬菌体的基因组大小也更大,例如基因组大小为162,910bp的肌病毒vb_salm_sj_3(wall等,2010;zhang等,2010;saez等,2011;zhang等,2014)和基因组大小为48,110bp的短尾病毒uab_78(bardina等,2016)。巨型噬菌体很少分离,并且不能通过常规方法频繁分离,这可能会偏向于较小的基因组大小的噬菌体(hillyard等,2016;saad等,2018;salmond和fineran,2015;serwer等,2009)。然而,它可以被认为,迄今为止所述的总噬菌体,大约2%是巨型的,所以它可能是巨型噬菌体,是真正稀少的,如前所述,并且实际上没有充分表现出来。
即使使用不同的沙门氏菌菌株进行富集并从各种环境来源收集样品,所有spfm噬菌体在遗传上也彼此相似(jurczak-kurek等,2016)。spfm噬菌体确实在snp上有所不同,snp可能存在于与宿主相互作用的蛋白质中。这可能影响噬菌体与细胞表面的附着动力学,导致宿主特异性的改变(switt等,2013)。这潜在地可以解释为什么在spfm噬菌体之间观察到宿主范围和eop的差异。用90%的遗传相同的假单胞菌(pseudomonas)噬菌体描述了类似的观察结果,同样snp导致噬菌体之间的表型差异(ceyssens等,2011)。应指出,噬菌体spfm5、spfm9、spfm10和spfm11共享相同的snp,这可能表明它们代表具有snp的克隆。snp可能是由繁殖宿主诱导的,这不同于最初分离噬菌体的宿主。
将spfm噬菌体与所有测序的沙门氏菌噬菌体进行比较,并将它们与其他已知的巨型沙门氏菌噬菌体spn3us(lee等,2011)和segd1聚类,它们分别在华南地区从鸡粪便和在韩国分离得到。非常有趣的是,spfm噬菌体在英国分离但聚集并且在遗传上类似于在不同大陆分离的噬菌体。类似地,从美国和欧洲的不同国家分离出87%遗传相似的假单胞菌噬菌体(ceyssens等,2011)。
大基因组大小的巨型噬菌体允许携带不存在于较小基因组大小噬菌体17的许多基因),例如所有spfm噬菌体具有的六个rnapβ亚基。噬菌体spn3us的多个rnap亚基在最近的出版物中已被广泛研究,2017),并且与phikz样噬菌体的rnapβ亚基非常相似。通过构建噬菌体基因的琥珀色突变体,最近在噬菌体spn3us中预测了另外三个rnap亚基。所有三个预测亚基也在spfm噬菌体中鉴定出:nvrnapβ’、vrnapβ存在于c末端和vrnapβ’存在于c末端(表a)。此外,预测的三个rnap亚基是噬菌体之间共享的核心基因的一部分。存在多个rnapβ亚基与其他测序的巨型噬菌体一致,例如在溶珊瑚弧菌(v.coralliilyticus)噬菌体bonaishi、青枯雷尔氏菌(ralstoniasolanacearum)噬菌体rp12和φ8rp31中已通过生物信息学鉴定的七个rnapβ亚基。总体而言,巨型噬菌体中额外基因的存在可以减少它们对与噬菌体生命周期相关的必需蛋白质的细菌宿主的依赖性,因此可以帮助扩大噬菌体宿主范围。这可以解释为什么所有spfm噬菌体可以感染多种临床上相关的沙门氏菌分离物,这可以使spfm噬菌体成为噬菌体治疗的理想候选者。
在汉城病毒属中,从噬菌体共有的核心基因中鉴定出阳性选择下的基因。阳性选择下的基因包括宿主相互作用的蛋白质,例如两个推定的毒粒结构蛋白,预计它们是基板楔形蛋白,并且参与尾纤维的形成,两者均残余噬菌体与细菌细胞的结合。这些宿主相互作用的蛋白质必须适应不同的细菌宿主,这可以解释为什么它们处于阳性选择下。这些结果还可以合理解释spfm噬菌体在宿主范围和eop中观察到的差异。其他噬菌体研究也已经鉴定宿主相互作用的蛋白质处于阳性选择下,例如编码底板蛋白并且可能参与宿主特异性的基因gp6。需要进一步的遗传和突变研究来表征阳性选择下的噬菌体基因以了解其重要性。
本发明人的研究已经描述并表征了可以裂解的通常与英国猪有关的沙门氏菌菌株的遗传相似的21种裂解性巨型噬菌体。全面的宿主范围分析和eop鉴定出许多噬菌体,它们是噬菌体治疗以提高食品安全性的理想候选者。
下文所述的工作鉴定出在体外和体内最大程度降低沙门氏菌的最佳噬菌体或噬菌体混合物。
方法和材料
细菌菌株和培养条件
这项研究中使用了两种肠道沙门氏菌亚种肠道血清型鼠伤寒沙门氏菌菌株:sl1344(登录号fq312003),实验室参考菌株,和2012年由动植物卫生局(weybridge,英国)从猪中分离出的s01160-12噬菌体dt193型菌株(card等,2016)。s01160-12是一种多抗生素抗性菌株,对四环素、新霉素、氨苄青霉素、磺胺甲噁唑、氯霉素、庆大霉素、链霉素、复方磺酰胺和阿普霉素有抗性。两种沙门氏菌分离物均常规在37℃下在木糖赖氨酸脱氧胆酸盐(xld)琼脂(oxoid,英国)上培养18小时,并且在nzcym肉汤中接种液体培养物(melfordbiolaboratoriesltd,英国),并在37℃下以100rpm培养18小时。
噬菌体繁殖和滴定
繁殖了噬菌体spfm2、spfm4、spfm10、spfm14,spfm17和spfm19。将在nzcym肉汤中培养的沙门氏菌sl1344的简要指数生长液体培养物(107cfu/ml/)用107噬菌斑形成单位(pfu)/ml噬菌体感染,并在37℃下以100rpm孵育6小时。将噬菌体培养物在4,200xg下离心15分钟,用0.22μm过滤器过滤上清液,并将噬菌体裂解物储存在4℃下。为了确定噬菌体的滴度,将噬菌体裂解物连续稀释10倍,并在具有sl1344细菌菌苔的lb1%琼脂板上使用小滴噬菌斑测定法(mazzocco等,2009)。最终噬菌体滴度表示为pfu/ml。
一步生长测定
将sl1344的指数培养物以0.2的光密度(od600)和107cfu/ml的细胞密度与噬菌体混合,感染复数为0.01,即细菌细胞与噬菌体之比为1:0.01。使噬菌体在37℃下吸附5分钟,并通过以4200×g离心10分钟除去未结合的噬菌体。将沉淀物用nzcym肉汤重悬,并在37℃下以100rpm摇动孵育。每10分钟取等分试样1.5小时,并使用小滴噬菌斑测定法确定pfu/ml计数(mazzocco等,2009)。进行了三个生物学重复,每次重复具有三个技术重复。
噬菌体受体分析
sangryeolryu为鼠伤寒沙门氏菌sl1344菌株的鞭毛产生基因(flgk)缺失提供了帮助;编码维生素b12的基因摄取外膜蛋白(butb);以及与lps相关的o抗原产生相关的基因(rfal)。使用λ红色重组方法构建突变菌株(shin等,2012)。为了确定噬菌体是否使用这三种中的任一种作为其宿主受体,将噬菌体裂解物稀释10倍,并使用小滴噬菌斑测定法(mazzocco等,2009)铺板在突变菌株上。进行了三个生物学重复,每次重复具有三个技术重复。
温度和ph稳定性测定
通过将噬菌体暴露于4℃、10℃、20℃、30℃、40℃、50℃、60℃、70℃、80℃、90℃和100℃的温度范围一小时来测试噬菌体裂解物的热稳定性。将噬菌体在聚合酶链反应(pcr)机器中孵育。对于ph研究,将100μl噬菌体裂解物添加到900μl的sm缓冲液中,该缓冲液的ph范围调整在1至14。为了确定暴露于不同温度和ph后的噬菌体滴度,请将噬菌体裂解物连续稀释10倍,并在具有有sl1344细菌菌苔的lb1%琼脂板上使用小滴噬菌斑测定法%(mazzocco等2009)。最终噬菌体滴度表示为pfu/ml,并进行了三个生物学重复,具有三个技术重复。
用噬菌体混合物的体外细菌杀伤测定
单独用噬菌体和由以相等体积混合的两种、三种和四种噬菌体组成的噬菌体混合物进行杀伤测定。对于不同的杀伤测定,将噬菌体混合物s01160-12组合培养物培养至光密度(od600)为0.2,此时以100的moi添加噬菌体。此外,有两个对照:一个是细菌对照,仅包含sl1344,另一个是噬菌体对照,仅包含噬菌体。将对照和测试样品的等分试样均在37℃下以100rpm孵育,并在0、1、2、3、4、5和6小时的时间点取等分试样。对于每个时间点,在lb1%琼脂板上测定细菌浓度(cfu/ml),并通过小滴噬菌斑测定法(mazzocco等,2009)测定噬菌体浓度(pfu/ml)。对于cfu/ml和pfu/ml,进行了三个技术重复,并将板在37℃下孵育16小时。每种杀伤测定进行了三个生物学重复。使用graphpadprism6绘制杀伤测定图,并使用配对t检验确定数据的显著性,其中p≤0.5被认为是显著的。
测试噬菌体混合物在体内大蜡螟(galleriamellonella)感染模型中的功效
蜡螟的制备
幼虫购自livefoodukltd.(rooksbridge,英国),在4℃下储存并在5天内使用。对于所有体内实验,选择重约0.25至0.30g的幼虫,并用蘸有70%乙醇的棉拭子表面消毒。
用噬菌体混合物处理的感染沙门氏菌的大蜡螟
对于所有幼虫噬菌体疗法研究,使用先前描述的方法(nale,chutia等,2016)。通过经口途径将最终浓度为105cfu的沙门氏菌菌株s01160-12和最终浓度为107pfu的噬菌体混合物施用到幼虫。通过首先培养包含109cfu/ml的s01160-12的液体培养物来制备菌株s01160-12,然后将其以4,200xg离心10分钟。丢弃上清液,将沉淀物重悬于0.1m磷酸盐缓冲盐水(pbs)中,并以4,200×g离心10分钟。再次丢弃上清液,将沉淀物重悬于pbs中,并在pbs中稀释10倍至105cfu/ml。使用hamilton注射泵通过经口途径向幼虫施用10μl细菌培养物和噬菌体混合物。噬菌体混合物在感染前1小时进行预防性施用,或与沙门氏菌同时施用(所有方案详见表3)。之后,将幼虫在37℃下孵育3天。每24小时监测一次幼虫的存活率并处死幼虫以确定cfu和pfu计数。为此,将幼虫在-20℃下储存3小时,进行解剖以除去其血淋巴,将其重悬于1mlpbs中并涡旋30秒(nale等,2016)。对于pfu计数,将样品以5,000xg离心5分钟。然后将滤液稀释10倍并通过小滴噬菌斑测定法滴定(mazzocco等2009)。对于cfu计数,将混悬液稀释10倍,并在xld琼脂板上进行点样测试。使用5μl环挑选暴露于噬菌体混合物后回收的沙门氏菌菌落,划线到xld琼脂板,并在xld琼脂上重新划线三次。将菌落接种到nzcym肉汤中,并在37℃以100rpm培养2小时。然后将培养物用于制作菌苔,以通过点检确定其对噬菌体的敏感性(mazzocco等,2009)。发现浓度为108pfu/ml的噬菌体。为了确定噬菌体在分离的菌落上的铺板效率,将噬菌体裂解物稀释10倍并使用小滴噬菌斑测定法(mazzocco等2009)进行铺板。
对于所有处理组,总共使用了60只幼虫,并评估了20只幼虫的存活率,并每24小时进行解剖。如果幼虫对触摸无反应,并且颜色从浅棕色变为黑色,则认为它们死亡。类似地,对于对照组,使用60只幼虫并每24小时处死20只幼虫,以确定其血淋巴中的cfu和pfu计数。所有实验中包括的对照幼虫为:健康幼虫;通过经口途径施用107pfu噬菌体的幼虫;通过经口途径施用105cfus01160-12的幼虫;和施用pbs的幼虫(表3)。每24小时监测所有对照的存活,并解剖其胃肠道的cfu和pfu计数。
表3.评价噬菌体治疗对处理感染沙门氏菌的幼虫的有效性的实验设置。
1每组中使用60只幼虫用于研究。每24小时检查20只幼虫的存活,并剔除以计算pfu和cfu计数。
2噬菌体混合物为:2-噬菌体spfm10-spfm14;3-噬菌体spfm4-spfm10-spfm19;和4-噬菌体spfm2-spfm10-spfm14-spfm19。
存活曲线
对于所有实验,进行了三个生物学重复,并采用kapad-meier方法使用graphpadprism6版(graphpadsoftwareinc,美国)绘制了存活数据。使用log-rank(mantel-cox)检验评估生存率差异。
生物信息学分析
噬菌体先前已保藏到ena中,它们的登录号为spfm2(lr535921)、spfm4(lr535902)、spfm10(lr535908)、spfm14(lr535912)、spfm17(lr535914)和spfm19(lr535916)。也可以通过该链接http://s3.climb.ac.uk/sinfo/spfm_genome.tar.gz找到带注释的噬菌体基因组。对于snp分析,可以考虑高于90%且p值较低的频率,但是当频率降低时,还可以鉴定snp。
结果
噬菌体裂解量
对六种spfm噬菌体进行噬菌体生长曲线。噬菌体spfm10和spfm17的裂解量为175和162pfu/每细胞,潜伏期短为20分钟。spfm2、spfm4、spfm14和spfm19的裂解量分别为每个单元165、162、162和153pfu/每细胞,潜伏期更长,为30分钟(表4)。
表4.spfm噬菌体的裂解量。
1平均结果来自三个生物学重复,每个重复具有三个技术重复。
噬菌体的温度和ph稳定性
为了确定噬菌体spfm2、spfm4、spfm10、spfm14、spfm17和spfm19对热的敏感性,将噬菌体暴露于4℃、10℃、20℃、30℃、40℃、50℃、60℃、70℃、80℃和90℃的温度1小时(图7)。所有噬菌体在50℃下稳定,与在4℃下储存的噬菌体相比,噬菌体滴度没有下降。当将噬菌体在60℃下孵育时,仅噬菌体spfm10和spfm17存活,滴度分别平均下降0.0和6.5log10pfu/ml。在70℃下噬菌体spfm17的滴度下降7.5log10pfu/ml,而在80℃下噬菌体不能存活。相比之下,spfm10更稳定,在60℃、70℃和80℃下滴度没有显著下降,但是在90℃下,噬菌体滴度降低了4.5log10pfu/ml,在100℃下没有回收到活的噬菌体。为了验证噬菌体spfm10的热稳定性,使用两种不同的温度控制设备(加热块和水浴)孵育噬菌体,并将其与使用pcr机孵育噬菌体时收集的原始数据进行比较。对于所有三种机器,结果都是相同的,进一步证实了spfm10的热稳定性。
喷雾干燥spfm噬菌体
喷雾干燥之前,使用截断值为10kda的50mlamicon超浓缩器(millipore,英国)浓缩噬菌体,将15ml噬菌体裂解液添加到浓缩器中,并以5,000xg的转速旋转20分钟。丢弃流出物,并将另外的15ml噬菌体裂解物添加到浓缩器中。滴定收集在过滤器上方的浓缩噬菌体,如果滴度为约1010,则用于喷雾干燥实验。
为了进行噬菌体喷雾干燥,将溶解于ph7.5的超纯水中的200ml体积的4%海藻糖(oxoid,英国)赋形剂溶液与0.2ml滴度为约5x1010pfu/ml的浓缩噬菌体以/混合。使用实验室规模的带有进行雾化的双流体喷嘴的labplant喷雾干燥器(英国)将赋形剂噬菌体溶液喷雾干燥,喷嘴的孔口直径为0.5mm。所有运行均使用3ml/min的恒定进料速度,雾化气流为6l/min。将干燥入口温度加热到80℃、85℃、90℃或100℃,相应的出口温度从40℃至60℃变化。使干粉状噬菌体通过旋风分离器,收集在100ml玻璃瓶中,并在4℃下储存直至使用。为了确定噬菌体滴度,将0.05g干粉状噬菌体悬浮于500μl噬菌体混悬液缓冲液中,稀释10倍并通过噬菌斑测定法进行滴定。
噬菌体在用于喷雾干燥的不同入口温度下的温度稳定性。
喷雾干燥spfm噬菌体
用糖赋形剂海藻糖将所有六种噬菌体喷雾干燥。在喷雾干燥期间,噬菌体暴露于热应力,因为使用加热的空气将噬菌体-海藻糖液体干燥成粉末。为了蒸发液体,使用了高入口干燥温度(可以手动设置),噬菌体-海藻糖溶液仅暴露于其中数毫秒。随后,收集粉末并将其暴露于较低的出口温度,该温度取决于设置的入口温度。在该研究中,使噬菌体经受80℃、85℃、90℃和100℃的不同入口干燥温度,以优化干燥过程,从而使噬菌体滴度减少最小化。
无论干燥期间使用的入口温度如何,spfm2、spfm4、spfm17和spfm19的噬菌体滴度损失至少为1log10pfu/g。当入口温度降到80℃时,spfm2和spfm4更稳定,但是对于spfm4,在所有入口温度下,噬菌体滴度损失是显著的,为3.00至3.16log10pfu/g。spfm17和spfm19的噬菌体滴度损失最低,入口温度分别为85℃和90℃。噬菌体spfm10和spfm14是这6种中最稳定的,当使用80℃作为入口温度时,噬菌体的最小滴度损失为0.02和0.35log10pfu/g。此外,当spfm14的入口温度升高时注意到滴度损失,但是spfm10噬菌体的滴度损失仍低于0.2log10pfu/g。对于所有噬菌体,回收的干粉状噬菌体的最终产量始终较低,为0.2-0.5g,因此,对于所有测试的入口温度,噬菌体的回收率为5%-13%。
该研究中使用的所有六种spfm噬菌体在遗传上彼此95%相似,但是在表型上,它们具有不同的特征,如从它们的个体以及在体外和体内对鼠伤寒沙门氏菌的组合杀伤活性中观察到。为了进一步调查为什么噬菌体组合spfm10-spfm14;spfm4、spfm10和spfm19是最好的,通过确定噬菌体的温度稳定性对所有六种噬菌体进行了表型分析。与先前表征的噬菌体一致,spfm2、spfm4、spfm14和spfm19在高达50℃的温度下仍稳定。当噬菌体暴露于较高的温度时,可能发生对噬菌体蛋白质的不可逆的损害,这导致随后的噬菌体感染性丧失。有趣的是,尽管有这种噬菌体spfm10和spfm17都是热稳定的,并且在高达70℃的温度下保持活性。尽管使用噬菌体spfm17,但是在高于50℃的温度下孵育后,噬菌体滴度显著降低。与噬菌体spfm10相比,在高达80℃的温度下噬菌体滴度没有下降,甚至在暴露于90℃一小时后仍保留了55%以上的噬菌体活性。噬菌体spfm10在极端温度下具有增强的热稳定性,并且比报道的大肠杆菌噬菌体vb_eco4m-7更稳定,该大肠杆菌在暴露于95℃5分钟后仅保留了20.5%的噬菌体活性。已经假设在噬菌体衣壳内二硫键交联的形成可能涉及在极端温度下稳定噬菌体,这已被rna噬菌体pp7证实。噬菌体spfm10和spfm17的衣壳中的二硫键交联是否有助于增加热稳定性尚不清楚。
此外,spfm10的热稳定性与其在喷雾干燥期间增加的稳定性之间可能存在相关性。噬菌体spfm10是天然非常有弹性的,当与赋形剂海藻糖组合时,噬菌体在暴露于100℃的最高入口温度℃时仍保持活性。令人惊讶的是,噬菌体spfm14尽管不是特别热稳定,但是在喷雾干燥期间非常稳定。相反,可以预测噬菌体spfm17可以承受喷雾干燥,因为它是天然热稳定性,但是与spm2、spfm4和spm19相似,在最高测试入口温度下高达3log10pfu/g减少。该研究强调,喷雾干燥期间的噬菌体稳定性在很大程度上取决于所使用的噬菌体,即使所测试的噬菌体在遗传上相似并且都是肌病毒,因此具有相同的形态。先前假设,肌病毒由于其脆弱的病毒体结构而不能很好地喷雾干燥,因为它们的长尾巴可能会收缩,或者它们的衣壳可能会在压力下分离。这可以解释为什么噬菌体spm2、spfm4、spfm17和spm19出现滴度损失,对于金黄色葡萄球菌噬菌体romulus也观察了滴度损失,注意到2.5log10减少。该假设的例外是巨型噬菌体spfm10、spfm14和假单胞菌噬菌体kz。
将噬菌体暴露于ph范围,以确定它们对ph的敏感性(图7b)。所有噬菌体在ph4至12是稳定的,噬菌体滴度没有显著损失。在ph3下,spfm10、spfm14和spfm17的噬菌体滴度分别降低了5.44、3.12和2.51log10pfu/ml。然而,噬菌体spfm2、spfm4和spfm19在ph3下是稳定的,噬菌体滴度没有变化。所有噬菌体在ph1、2、13和14下都不稳定,并且没有回收噬菌斑。
鉴定噬菌体受体
在sl1344的不同受体突变菌株上筛选成斑率,以确定噬菌体用于结合沙门氏菌的细菌受体(表5)。将噬菌体在如下菌株上进行测试:在编码维生素b12摄取外膜蛋白的基因(sl1344δbutb)中有缺失和在生产鞭毛基因(sl1344δflgk)中有缺失。与野生型sl1344相比,两种菌株的成斑率均无显著变化。然而在菌株sl1344δrfal(其在参与lps相关o-抗原生产基因中有缺失)上,所有噬菌体都不能感染并且成斑率为0。在该数据基础上,我们预计所有六种噬菌体使用将o抗原lps作为宿主受体。
表5.野生型和受体突变菌株sl1344的沙门氏菌噬菌体滴度。
六种spfm噬菌体的体外感染动力学
对单独噬菌体spfm2、spfm4、spfm10、spfm14、spfm17和spfm19以及两种、三种和四种混合物组合进行了测试,以确定引起的多种抗生素抗性鼠伤寒沙门氏菌菌株的最高减少的最佳组合。在六个小时的时间过程中评估了杀伤情况,并将其与未感染的对照进行了比较。图8中显示的数据是在感染复数(moi)为100下进行的杀伤测试。
单独噬菌体混悬液的细菌杀伤杀伤测定
在moi100下,当噬菌体spfm2、spfm4、spfm10、spfm14、spfm17和spfm19单独添加时,它们在感染后1小时引起显著减少(p≤0.05)。然而,与未感染的培养物相比,在感染后2小时观察到最大细菌细胞裂解,分别减少了1.77、2.36、2.15、2.43、2.32和1.52log10cfu/ml(图8a)。特别地,噬菌体phispfm2、phispfm4、phispfm5和phispfm6的减少率非常显著(p≤0.01)。6小时后,保持细菌计数减少的最有效噬菌体为phispfm2、phispfm4、phispfm5和phispfm6(p≤0.05),分别为0.71、0.80、0.83和0.85log10cfu/ml。
用两种噬菌体混合物的细菌杀伤杀伤测定
测试了15种不同的两种噬菌体混合物,对于所有组合与未感染的培养物相比,感染后1小时鼠伤寒沙门氏菌减少1和3log10cfu/ml(图8b和8℃)。与单独噬菌体的杀伤测定相似,噬菌体组合spfm10-spfm14、spfm2-spfm4、spfm10-spfm17和spfm2-spfm10在2小时后观察到最大统计学显著减少(p≤0.001),分别为4.43,4.46,4.34and4.36log10cfu/ml。此外,在2小时后,噬菌体混合物spfm2-spfm14、spfm2-spfm6、spfm10-spfm19和spfm2-spfm19引起细菌减少2.36、2.60、2.64和3.29log10cfu/ml(p≤0.001)。感染后6小时,噬菌体spfm10-spfm14和spfm2-spfm10保持显著的鼠伤寒沙门氏菌减少(p≤0.05),分别为1.97和2.25log10cfu/ml。与未感染的对照相比,其他12种两噬菌体组合在6小时后效率较低,并且维持0.5至1log10cfu/ml的细菌减少。
用三种噬菌体混合物的细菌杀伤测定杀伤
与单独噬菌体和两种噬菌体混合物相比,测试的十一种三种噬菌体组合在感染后1小时清除细菌液体培养物更快速,(p≤0.01),约2.5log10cfu/ml(图8d和8e)。2小时后,杀伤还增强,最有效的三种噬菌体混合物为spfm4-spfm10-spfm19、spfm10-spfm17-spfm19和spfm2-spfm17-spfm19,分别引起2.60,3.50and3.53log10cfu/ml减少(p≤0.001)。感染后6小时,被测试的十一种三种噬菌体组合中的两种将裂解延长:spfm10-spfm17-spfm19和spfm2-spfm10-spfm19(p≤0.05),为1.10和1.70log10cfu/ml。
用四种噬菌体混合物的细菌杀伤测定杀伤
对于所有四种测试的四种噬菌体混合物,在感染后1小时,观察到了快速细菌裂解引起鼠伤寒沙门氏菌1.5-2.5log10cfu/ml减少(p≤0.01)。在这四个组合,三个组合:spfm10-spfm14-spfm17-spfm19、spfm2-spfm10-spfm14-spfm19和spfm2-spfm4-spfm10-spfm19在感染后2小时减少细菌计数上最有效地,分别为3.51、3.40和3.38log10cfu/ml(图8f)。2小时后,第四种噬菌体组合spfm2-spfm10-spfm17-spfm19导致细菌密度减少2.61log10cfu/ml。与三种噬菌体混合物相比,6小时后,细菌灭活与spfm2-spfm10-spfm14-spfm19和spfm2-spfm4-spfm10-spfm19的组合是一致的,与未感染的细菌对照相比分别为1.58和1.61log10cfu/ml。
噬菌体混合物在体内的感染动力学
为了确定是否体外观察到的功效结果转变为噬菌体混合物的体内功效,在大蜡螟感染模型中测试了spfm10和spfm14(两种噬菌体);spfm4、spfm10和spfm19(三噬菌体);spfm2、spfm10、spfm14和spfm19(四噬菌体)。幼虫感染了mdr鼠伤寒沙门氏菌s01160-12菌株。评估了噬菌体混合物的预防性处理活性和向沙门氏菌同时施用噬菌体(图3)。噬菌体混合物以moi100施用,与体外杀伤测定研究一致。
噬菌体混合物的预防性处理
噬菌体混合物进行预防性施用,在感染前1小时,以确定它们在处理沙门氏菌感染的功效(图9和9c)。24小时后,所有经噬菌体施用的幼虫的平均存活率为90%,而仅感染沙门氏菌的对照幼虫c-6的存活率为81%。经2-噬菌体(p-1)、3-噬菌体(p-2)和4-噬菌体(p-3)混合物预防性处理的受感染幼虫的平均细菌计数分别为2log10、3.3log10和3.8log10cfu/幼虫(表3)。对照组c-6组的平均细菌计数更高,为4.6log10cfu/幼虫。48小时后,c-6组的沙门氏菌计数为约4.8log10cfu/幼虫,存活率降至59%。观察到幼虫组p-1和p-3的沙门氏菌计数进一步降低了约3.3log10cfu/幼虫,两组的12/20幼虫中均未检出沙门氏菌。比较组p-2的沙门氏菌减少1.3log10cfu/幼虫,6/20的幼虫中未检出沙门氏菌。p-1、p-2和p-3的存活率分别为95%%、81%和78%。72小时后,c-6组的细菌计数进一步增加至5.7log10cfu/幼虫,存活率降至3%。p1、p2和p3组的存活率分别显著提高90%%、68%和62%。噬菌体组中细菌载量的额外减少反映了更高的幼虫存活率。对于p-1,沙门氏菌平均计数为1log10cfu/幼虫,对于p-2为2.4log10cfu/幼虫,对于p-3为2.0log10cfu/幼虫。此外,从p-1组的14/20幼虫、p-2组的10/20和p-3组的13/20中没有回收沙门氏菌,这突出说明了这些幼虫中的感染的完全清除。
噬菌体混合物的共感染
将噬菌体混合物和沙门氏菌同时经口施用到幼虫并且在72小时每24小时监测一次(图9b和9d)。24小时后,经2-噬菌体(coi-1)、3-噬菌体(coi-2)和4噬菌体(coi-3)混合物感染的幼虫的存活率超过约88%,而c-6的存活率为81%(表3)。在幼虫组coi-1和coi-2中,观察到沙门氏菌计数降低了约2.2log10cfu/幼虫,在coi-3组中降低了2.8log10cfu/幼虫。48小时后,c-6组中60%的幼虫存活,而coi-1、coi-2和coi-3组中分别84%%、72%和78%的幼虫存活。幼虫组中,coi-1组的沙门氏菌平均计数为2.4log10cfu/幼虫,coi-2为2.8log10cfu/幼虫,coi-3为1.9log10cfu/幼虫,而c-6组中沙门氏菌平均计数为4.8log10cfu/幼虫。此外,在两组coi-2和coi-3中,从12/20幼虫中没有分离沙门氏菌,并且在coi-1组中,仅从3/20幼虫中没有分离沙门氏菌。72小时后,与c-6组中幼虫的平均计数相比,所有噬菌体混合物导致沙门氏菌计数降低了约4log10cfu/幼虫。coi-2和coi-3组的幼虫平均存活率为65%,在coi-1组中,72%的幼虫存活%,而c-6组中仅感染沙门氏菌的幼虫3%存活%。在coi-1、coi-2和coi-3组中分别从13/20、16/20和12/20幼虫中没有分离沙门氏菌,但是c-6组的所有幼虫中分离出沙门氏菌。
对照幼虫组
72小时后,对照幼虫组c-1、c-2、c-3、c-4和c-5的存活率为100%,并且在这些组中均未检测到沙门氏菌。仅从c-3、c-4和c-5组中回收噬菌体和在72小时内,每24小时进行噬菌体计数(图9e和9f)。对于预防和共感染研究,对于对照组c-3,从24小时到48小时的总噬菌体计数增加了1-log。对于对照组c-4和c-5,没有观察到噬菌体滴度的显著增加,并且总噬菌体滴度在72小时内保持恒定。
在幼虫模型中暴露于噬菌体混合物后回收的沙门氏菌菌落的敏感性
72小时后,从幼虫中回收168个沙门氏菌菌落,暴露于2-噬菌体混合物后56个菌落,3-噬菌体混合物回收56个菌落,并且4-噬菌体混合物回收56个菌落。测试了这些菌落对它们各自混合物内的单独噬菌体的敏感性。所有菌落对它们所暴露的混合物内的单独噬菌体保持敏感,这基于是否观察到了清晰的裂解区。选择每个噬菌体处理的10个10个沙门氏菌菌落,测定混合物内的单独噬菌体的成斑率并与野生型鼠伤寒沙门氏菌s01160-12菌株比较(图10)。通常,所有成斑率为1.0左右,表明噬菌体在该体内模型中仍然有效。
对于一些筛选的菌落,观察到成斑率的差异,例如,对于暴露于2-噬菌体混合物后分离出的3/10菌落,spfm14的成斑率降低了10%-19%。反之,4/10菌落上spfm10的效率提高了10%-26%。对于暴露于3噬菌体混合物后分离出的菌落,spfm4和spfm10在3/10菌落上spfm19的成斑率降低了约15%。然而,在4/10菌落上,spfm4和spfm10的成斑率提高了10%以上。在暴露于4-噬菌体混合物的菌落上,在3/10菌落上,spfm10和spfm19的活性降低了约15%,但是在所有10菌落上spfm14一致地具有更高的成斑率。
六种spfm噬菌体之间的遗传差异
基于平均核苷酸同一性,分离出的沙门氏菌噬菌体彼此95%相似。进行了单核苷酸多态性(snp)分析,并将热稳定性噬菌体spfm10与spfm2、spfm4、spfm14、spfm17和spfm19比较。snp数据列于表6中,并且在噬菌体之间总共鉴定出很少的snp。在所有五种噬菌体中鉴定出的一致snp位于70,890位,该基因的一部分被注释为假设蛋白。这是噬菌体spfm10和spfm14之间唯一识别出的snp差异。反之,当将spfm10与spfm2、spfm4、spfm17和spfm19比较时,分别在噬菌体基因中鉴定出总共3、3、5和2个snp。所有鉴定出的snp都在假设的蛋白中,期望spfm10与spfm17在基因组57985位的snp差异注释为染色体分区蛋白smc。此外,将噬菌体与两种巨型噬菌体spn3us和segd1比较。鉴定出大约超过6900个snp,并且在所有带注释的基因中至少存在一个snp。
表6.将噬菌体spfm10与spfm2、spfm4、spfm14、spfm17和spfm19比较以鉴定噬菌体之间的snp
沙门氏菌属是一个主要的动物和人食品安全问题,并且由于mdr沙门氏菌菌株引起的感染增加,需要抗生素替代物来应对这种感染。噬菌体可以提供另一种选择。21种巨型噬菌体被认定是可以杀伤英国猪相关的流行血清型的代表菌株。基于它们广泛的宿主范围,从集合中选择六种噬菌体,如本文中鉴定和如本文讨论保藏的。所有六种噬菌体均具有大的裂解量和短的潜伏期,这是有毒噬菌体的特征之一,非常适合治疗。大的裂解量也增加了噬菌体与其靶病原体之间接触的可能性,这对于感染至关重要。此外,大的裂解量可能意味着噬菌体消灭细菌的速度快于其复制速度,这可能潜在地降低选择噬菌体抗性细菌的风险。已分离出的其他巨型噬菌体具有相似的短潜伏期,小于40分钟,每个受感染细胞的裂解量超过140个噬菌体,其包括klebseilla噬菌体vb_klem-rak2(
所有六种噬菌体在遗传学上彼此95%相似%,并且与相同的细菌受体结合,但是ph和温度稳定性数据突出表明,噬菌体之间存在表型差异。ph稳定性数据显示六种噬菌体中三种在ph3时噬菌体滴度显著降低。此外,尽管所有噬菌体均在50℃稳定,与其他表征的噬菌体一致,只有噬菌体spfm10和spfm17在暴露于70℃后仍能保持活性。然而,噬菌体spfm17在高于50℃的温度下孵育后,噬菌体滴度显著降低。与噬菌体spfm10相比,直至80℃,甚至在暴露于90℃1小时后,噬菌体滴度也没有下降,并且保持超过55%的噬菌体活性。这是非常不寻常的,因为通常在噬菌体暴露于高温时它可能导致对噬菌体蛋白的不可逆转破坏,随后导致噬菌体感染性的丧失(ahmadi等2017)。spfm10的天然热稳定性很罕见,并且比报道的大肠杆菌噬菌体vb_eco4m-7显著更稳定,在95℃下暴露5分钟后仅保留了20.5%的噬菌体活性(jurczak-kurek等2016)。在噬菌体衣壳内,二硫键交联的形成可以在极端温度期间参与稳定噬菌体。噬菌体spfm10和spfm17衣壳中的二硫键交联可能有助于提高热稳定性。
用于治疗的理想噬菌体或噬菌体混合物是能有效消除靶细菌物种的多个亚组并且能够延迟噬菌体抗性突变体的出现的那些噬菌体或噬菌体混合物。在体外测试了spfm噬菌体组合:单独、两种、三种和四种噬菌体,并且确定哪种组合可以快速裂解代表性鼠伤寒沙门氏菌mdr分离物(agency2014)。单独测试时,所有噬菌体仅在1小时后就导致细菌数量减少1log10cfu/ml。沙门氏菌在1小时后立即重新生长,并且6小时时,噬菌体感染培养物的沙门氏菌计数与未感染的对照相似,两者之间均无显著差异。这些结果与以前用单独噬菌体对沙门氏菌进行的杀伤测定一致。
测试了两种、三种和四种噬菌体组合的噬菌体混合物,并且2小时后细菌裂解提高了约3至4log10cfu/ml减少,鼠伤寒沙门氏菌的重新生长不太突出。体外测试了总共36种不同的噬菌体组合。噬菌体混合物spfm10-spfm14和spfm2-spfm10对裂解鼠伤寒沙门氏菌最有效,并且是用于噬菌体治疗的理想候选物。类似地,在以前的研究中,三种(bardina等,2012)、六种(albino等,2014)和十种(zhang等,2010)的噬菌体混合物可能使沙门氏菌数量减少约2log10cfu/ml。这与本研究中最好的两种噬菌体混合物观察到的4log10cfu/ml减少不同。该体外数据突出表明,两种巨型噬菌体混合物有效地快速裂解沙门氏菌,并且两个组合能够裂解100%的%英国顶级猪血清型相关的筛选的代表性分离物。有趣的是,两种混合物还包含天然热稳定性噬菌体spfm10,这可能表明它比其他五种噬菌体更具毒性。关于为什么用三种或四种噬菌体混合物不能改善细菌杀伤的可能解释是,当噬菌体以等体积混合时,将噬菌体稀释以达到最终的moi,并且可以稀释噬菌体的毒性。此外,由于噬菌体竞争沙门氏菌细胞上的相同受体位点,因此在混合物中一种噬菌体可能会胜过其他噬菌体。胜过其他噬菌体的噬菌体可能是繁殖速度更快(潜伏期短且裂解量大)的噬菌体,并且由于混合物中其他噬菌体的竞争能力较弱,因此它可能降低混合物的整体功效。评估了不同噬菌体的组合对于确定在减少靶病原体上的最佳混合是很重要的。
在体内幼虫感染模型中测试了噬菌体混合物清除mdr沙门氏菌菌株的功效,以与噬菌体体外和体内功效比较。测试了三种混合物:spfm10-spfm14;spfm4-spfm10-spfm19;和spfm2-spfm10-spfm14-spfm19,并且预防性施用和共感染。与体外数据相似,噬菌体混合物spfm10-spfm14通过两种处理在清除感染上最佳,它显著提高了幼虫的存活,并且72小时后从超过65%的幼虫中没有回收沙门氏菌。结果提供了进一步的证据,表明这种混合物对于治疗是可能是理想。在其他幼虫研究中已经观察到了类似的噬菌体混合物清除靶细菌的功效,例如24小时后,在感染铜绿假单胞菌(pseudomonasaeruginosa)前2小时以100的moi施用6种噬菌体混合物能够使幼虫的存活提高80%(beeton等2015)。另一项独立研究表明,当感染有毒的铜绿假单胞菌菌株后,不同的六种混合物将幼虫的存活提高30%(forti等2018)。尽管三种和四种噬菌体混合物的体外功效较低,但是两种混合物均能提高幼虫的存活和沙门氏菌清除,但三种噬菌体混合物在共同感染时更有效。结果突出表明体外杀伤数据在体内不总是相关,并且在幼虫感染模型中筛选噬菌体混合物的功效可能是评估噬菌体混合物毒性的一种改进方法(seed和dennis2009;nale,chutia等2016);abbasifar等,2014年;manohar,nachimuthu和lopes,2018年)。据我们所知,这是在幼虫感染模型中评估巨型噬菌体对mdr鼠伤寒沙门氏菌的活性的第一项研究和使用巨型噬菌体的第一项研究。
在幼虫感染模型中暴露于三种不同噬菌体混合物后分离出的菌落仍然对具有该混合物的单独噬菌体敏感,这表明我们没有分离出任何抗噬菌体性的沙门氏菌细胞。此外,噬菌体在回收的菌落上仍然具有高的成斑率,并且与野生型菌株相比可以以相同的程度感染。
由于六种spfm噬菌体之间存在表型差异,因此进行了snp分析以鉴定噬菌体之间的遗传差异。当将热稳定性噬菌体spfm10与其他五种spfm噬菌体比较时,它们之间的snps非常少。在所有五种噬菌体中,假设蛋白在基因组70890位上有snp,并且该snp存在于一种处于阳性选择且自然易于突变的蛋白中。hhpred的进一步分析预测该基因已命中c-末端果胶裂解酶结构域(98%概率),并且可能参与噬菌体尾纤维的形成。果胶裂解酶结构域功能预计切割糖苷键,并且可能参与降解沙门氏菌表面的多糖,因此在噬菌体宿主范围中可能起重要作用。此外,该snp可能在热稳定性中发挥作用,这是spfm10与spfm14之间的唯一区别,后者的噬菌体仅在50℃下稳定,而spfm10的热稳定性直到90℃。spfm10与噬菌体spfm2、spfm4、spfm17和spfm19之间的其他snp在另一种预测的果胶酸裂解酶蛋白中得到了鉴定,该蛋白也表明处于阳性选择下。该蛋白质可能是宿主相互作用基因,它必须易于突变。当与spfm10比较时,仅在spfm17中在染色体分区蛋白和假设蛋白中又鉴定出另外两个snp。spfm17还可耐受高达70℃的高温,这两个snp的添加可能潜在地在稳定化中发挥作用,因为它们是噬菌体所特有的。再次需要进一步的诱变分析以确定两个snp的存在对热稳定性的重要性。
下文描述的工作描述了噬菌体对与英国鸡相关的顶级血清型的感染性。
实验程序
细菌菌株和培养条件
在该研究中,总共检查了18种沙门氏菌菌株。所有肠道沙门氏菌亚种肠道菌株是由英国韦布里奇的动植物卫生局(apha)从2015到2017之间从英国的鸡中分离出的(表7)。从这些菌株的血清型为:3种s.13,23:i>;5种肠炎沙门氏菌;4种婴儿沙门氏菌;3种s.ohio和3种s.seftenberg。
表7.鸡沙门氏菌菌株。
噬菌体的分离,纯化和繁殖
通过将感染了107pfu/ml噬菌体的沙门氏菌sl1344(107cfu/ml)的指数生长液体培养物在nzcym肉汤中于37℃摇动(100rpm)混合6小时,来制备增加体积的纯化噬菌体裂解物。将噬菌体培养物以4,200xg离心15分钟,用0.22μm孔径的过滤器过滤上清液,并将噬菌体裂解物在4℃下储存。为了确定噬菌体滴度,将噬菌体裂解物连续稀释10倍,并在lb1%琼脂板上使用小滴噬菌斑测定法(mazzocco等,2009)。最终噬菌体滴度表示为pfu/ml。
噬菌体宿主范围分析和成斑率
通过小滴噬菌斑测定法(mazzocco等,2009)在不同的肠道沙门氏菌亚种肠道血清型上确定单独噬菌体的宿主范围,并在37℃孵育18小时。检查板,通过清除或噬菌斑的细菌裂解或无感染,并且从三个生物学重复中注意到平均观察结果,每个重复具有三个技术重复。
宿主范围分析
在从鸡中分离出的18种肠道沙门氏菌亚种肠道菌株上测试了spfm2、4、10、14、17和19噬菌体的功效。所有18种菌株代表与英国顶级猪相关的沙门氏菌血清型,即s.13,23:i>;肠炎沙门氏菌;婴儿沙门氏菌;s.ohio和s.seftenberg。所有噬菌体可以感染所有肠炎沙门氏菌菌株。噬菌体spfm2、spfm14和spfm19在所有婴儿沙门氏菌上产生浑浊的澄清,并且不可以感染s.seftenberg血清型的代表性菌株。噬菌体spfm4和spfm17可以裂解所有婴儿沙门氏菌,并且只可以裂解1/3的s.seftenberg菌株。
表8.沙门氏菌鸡分离物中spfm噬菌体的宿主范围,并且呈现的数据是三个重复的平均值。菌株编号与表7中列出的编号有关。
尽管已经参照特定的实施例具体示出和描述了本发明,但是本领域技术人员将理解,在不脱离本发明的范围的情况下,可以在形式和细节上进行各种改变。
序列表
<110>莱斯特大学
<120>治疗性噬菌体
<130>p193346.wo.01
<150>1815483.1
<151>2018-09-24
<160>12
<170>patentinversion3.5
<210>1
<211>903
<212>dna
<213>phikz样巨型噬菌体
<400>1
tacagctaatagaatttgacctttgttgtcggtccggtctgaaatctaaggtaactttag60
atagcaatgctgttgggatctgtttcacaattgggtcatcgtggcccccttggataacat120
tgtgatggtccattgtgttgctgcatgctcctgttctggtggcttcgcatattttgcata180
gttatagcgtagcaccgccatttcccgtgacttttccaataccccgacggctaacatgtc240
ccgctgaaaggttactgtcctgggccgggtgtccttgaataagcaccgctgaccgtagca300
ccgatgaaaccgtgtgactgtttgcttcttaaataagaattggtacggctcgacttacca360
aattaaccgaaattacggaccttggttcgaggttgtaacaaggtatttaagcaaaagttt420
cctgcgtaagacaagtaaggactgtgccactgggcaaatccctgctgcaccttgcttgtt480
atggtcgttccagaccggaccccttgtctactgatgccaaagggcggtgcctcgcagaga540
tggtgattggtcgcagcctgtaaattatttctgccaatgctcatacaccatgcaaacgga600
gctgagcctctgatgtcaatacccgtgtcgtggatggattcgtcgccaccaataaaagtt660
ctgccgctcaccgtgttgtgcaagcgttcgaacaaggcgatgcgcccgcagcattctgca720
aaaccgctgaatggctgcgcgaggccgccatcaggttggctaagttcgcgttgtaacaaa780
cggttgccaatactgttgcggtgggggaccatgatagcgtcgttgagtggtctgagtggt840
agcatgtggtgaagctggttatcgcgcagccatcaggtgcaatagcttgaccaagaatcg900
att903
<210>2
<211>900
<212>dna
<213>phikz样巨型噬菌体
<400>2
tacggataatgtaatgtgaaacgtgttgccttcggtgtccgcgagctatcgtagctttag60
atagcgttctgcggcccgttgcggtggtaactacaattacgtggcccttgcggagaccgt120
tgggaaggcccactgcgatgtagcatgcttctgttgcgacaccttttgttgtggtgaatg180
gcaataacctatcgacgccaatttctgccactcgcacagaagttgtggggatggcgggtt240
cccaagaaaaatctgtggcccggaccgggggtctttgacaatatgccgctgaccgtgcga300
cctatgaaaccttgtcattgcggtcgtctcaataagtgattgtgccttgacttgccggag360
caccacttatacaagtcaaggcggcaaccacggaccgtatttaagtaaatgtttttggca420
taagacaaatacaggctattgagctattatttgcgatcgggcgtcaagtaaatattggtc480
ccctactacatgccggtcctaccgttacctatacatggcccgacccgatgacgtgcattg540
gttgccgcgtgccactgctttttgccaatgcttatgcaccacgcgaacggcataaacaag600
ctgatgtttaagacctgcagaatgcaaccttcgccattgtgactcaacatagagctgcca660
ctcaccaaattgtggaagcgcgcggacatgttgatgtcacccgtcaaaggcgcgaaactg720
ctgtaaggccagccaggttagctaagccaatgacgggttgttagacgtaagaaacgccga780
tacttattgtgcaggtcagtcaccatgttgtcaggtggaatgggactcagcggcagcacc840
tcattgatgccatggtgatgttatgcgaggtaacgccatgaacttgagcaagacggaatt900
<210>3
<211>240
<212>prt
<213>肌病毒科
<400>3
metpheargthrileleupheleuleuvalalametthrthrpheasn
151015
serasnalasertyrglnthrthrserasnlyshisglnlysgluphe
202530
valleuileglnaspglnpheasngluilelysproleuilevalthr
354045
alaserarglysglnglyvalhisalaglymetleualathrthrile
505560
tyrarggluserargpheasnlysasnvalglylysasnlysserser
65707580
seralaserservalvalglnmetthrthrglythrlysargsermet
859095
ileargleutyrglylysglnleuasnileprolysasnalaaspleu
100105110
asnlysprolystyralavalglnleualaalavaltyrmetlyshis
115120125
ileglugluhisleuthrlysglnleulysarglysproserthrala
130135140
gluilealaleuglytyrargpheglygluglyalaalavalalamet
145150155160
ilelyslyslysserprovalglylysargtrpmetalasertyrarg
165170175
lysaspalaalaphetyrglyalalysmetthrproprolysalaglu
180185190
thrargglnleualaphealalysgluasphisaspglnargvalala
195200205
gluleuglnlysiletrpaspthrleutyrthrlysileserproala
210215220
alaglythrleuleualaasnasnthrilemetlysglyalaleuleu
225230235240
<210>4
<211>333
<212>prt
<213>肌病毒科
<400>4
metlysthrvalalalysleupheargserserileargseralaglu
151015
pheglnasnasnvalglythraspthrvalargalaargargleulys
202530
glupheleulysaspgluthrtyrservaltyrvalvalargasnlys
354045
gluthrtrpgluglytyrleuvalalathrtyrarghisglyasppro
505560
thrleuthrcyshistyrpheglugluphevalproglyalapheasp
65707580
lysvalglyleualaleuvalleuasphisthrglypheglutyrmet
859095
sercysproalaprothrglyaspvalgluglnlysleuvalglulys
100105110
glyphevalleuseraspilevaltrpalacyshisserlysleuphe
115120125
asplysvalilethralasergluilelysileileserglyaspala
130135140
tyralagluleuserlysaspasplysaspvalileleuasnleuval
145150155160
serglyalailecysglugluleuglutyraspvalargglyhistyr
165170175
thrhisileglyproleualagluthrvalilealaargileglnhis
180185190
alaasnvalgluthralailetrpargaspglyargaspilevalgly
195200205
ilealaglnmetargleuvalalaproglyvalvalgluleusergly
210215220
leuthrvalalaprothrhisarglysargglyileglyargargleu
225230235240
metglyalaleupheaspphealaglnarghisseraspglnvaltyr
245250255
ilegluthralaalaaspasnasnproalaasnhisleutyrglyasn
260265270
ileleuargcyslysglnvalleuargthrleuthrleulysargasp
275280285
progluleuilesertrplysargvallysglyasnargalaasparg
290295300
valleulysglugluproserleuprovalgluthrasnproargpro
305310315320
thrileasnaspmetpheglnserglymetargglycys
325330
<210>5
<211>154
<212>prt
<213>肌病毒科
<400>5
metlysilevalphevalthrlysproasnserprotyrglylyslys
151015
tyrtyralaaspleuleuglyleuhisasnlysleutyraspaspgln
202530
lysilelysleualaaspleuasnglyarggluglypheasnglnile
354045
serlysleuglugluvaltrpglygluglythrleucysalacysala
505560
valaspaspargglyvalalavalglypheilethrpheglytyrthr
65707580
lyslysglyglnargpheleutrpvaltyrasnphephevalaspglu
859095
servalargserglnglyvalglyhisgluleuileglualavalglu
100105110
asntyrglylysserlysglycysalatrpmetglnleuasnvalleu
115120125
glyasnasnaspargalailealaphetyrgluargpheglyphearg
130135140
thrglutyrglnaspmetvallysgluleu
145150
<210>6
<211>425
<212>prt
<213>肌病毒科
<400>6
metalaasnphevallysserlysleualaarggluservalgluala
151015
thrglyaspvalileaspglyleuserasnvalgluproalagluglu
202530
asnileaspvalglnleualagluvalalaserileaspglyglnleu
354045
gluglnleuaspglyaspglngluthrleualaalaaspthrgluarg
505560
thrglualaalaileaspalaalaaspglulysilegluasnglyglu
65707580
glumetprogluglualailealahisthrgluvalalaglngluser
859095
ilearglysargtrpalaileaspargthrlysleualaarggluser
100105110
tyrargargglyargglymetthrlysalaalaglngluglytrplys
115120125
gluthrleulysaspleuphelysargphevalgluleucyslysala
130135140
valilealalysalalysgluleulysleulystyrileasnvalgly
145150155160
lysseralaglnlysargalalysalatyrglnglumetleuarglys
165170175
leuglylysglnlyslysgluasnileserglyglypheileserlys
180185190
leuserilegluglyasppheaspvalalaasnserilealaileala
195200205
lysgluleuthrglyglylysalalysaspalaileasnlysleuser
210215220
serglnalasergluseralavalalailethrlysthralaglugly
225230235240
thralathralaalalysglyalavalaspvalalaleupheglythr
245250255
alaalalyslysleuargthrleuproglnphegluaspglyglugly
260265270
glyglnlysvalleualaleuproglyasnalatyrvalglnilegly
275280285
serlysthrleualaglyglyglnaspphethralavalalapheleu
290295300
serthrglyaspserthraspthrlysgluilealathrproalaile
305310315320
thralaleualaseralaalathralaleuaspalaileglylysgly
325330335
pheglulysvalleuglnasppheargsertyraspalaaspvalglu
340345350
lysleugluglnalaalaserlysalaalaalaalaleuasplysser
355360365
asnaspgluglyglutrpglualaleuargasnalaargthralaala
370375380
aspglnalavalargasntyrglnthrleuasnargalavalalahis
385390395400
valglyasnthrvalileserglyleuasnglytyrileglyalagly
405410415
ileglyalatyrglulysserlysala
420425
<210>7
<211>568
<212>prt
<213>肌病毒科
<400>7
metvalgluileglnmetthrtyrthrsermetglyvalargvalglu
151015
valproileargglnalagluargalaileleualatrpalagluglu
202530
asnmethisalaprolysmetglylysglnhisglyargilethrthr
354045
gluargglyaspalatyrtyralahisileproserleuargthrphe
505560
ilephehislysvalphealaglulysilelyshisileleuglnarg
65707580
alaalaileglutyralapheglntyrlysleuthrgluhishispro
859095
gluargglythrprotyrsercysthrpheaspasntyrglypheser
100105110
metvalgluseraspprogluserargphetyrtyrglnasngluval
115120125
valaspalaalaseraspproasnargproglnthrilephealaile
130135140
glnthrglyargglylysthrlyssercysmetlyssermetvallys
145150155160
argglythrargthralaleuilehisargprosertyrvalprolys
165170175
trpleupheaspvalcysaspaspgluthrglyleuargileleuarg
180185190
aspgluvalleuvalcysthrglyvalglnalailetyraspalaleu
195200205
glumetglylysserglygluleuaspargargglyilelysvalile
210215220
ileleuprothrvalserleuglnargpheleulysglutyrileasn
225230235240
thralaalathrasnprovalaspleuaspthrphetyraspvalleu
245250255
glyvalglyleuvalalametaspgluvalhisgluhisphehisleu
260265270
valtyrmetalaglyilemetleuasnproproalaserileglumet
275280285
seralathrleulysproglyserserlysalapheilealagluarg
290295300
tyrleugluargpheprometglutyrargileserileproileile
305310315320
provalvalaspvallysalaleutyrtyrargleuaspasplyslys
325330335
phealatrptrpalaserlysmetthrprotyrasnhislysleuphe
340345350
gluglylysleuilelysgluasnleuhisleuglutyralaasnmet
355360365
iletrpaspvalvalglulysserpheleulysargtyrglnprogly
370375380
glnlyscysleuleuleuphealathrvalalametcysgluphephe
385390395400
thrglutyrvallysasplysleuserargaspprovalphehispro
405410415
leumetvalalalystyrasnalaglyaspsertyraspasppheile
420425430
glnalaasppheserileserthrproglylysalaglythralaval
435440445
asplysproglyleuvalhismettyrileserthrprovalgluasp
450455460
glnglnleuasngluglnmetalaglyargproarglysileleuhis
465470475480
asnglutrpglygluileaspprothrvaltrpleuphehisalatyr
485490495
asnvalprolyshiscysasntyrleuasnalaargglnlysserleu
500505510
lysaspvalvalleuserpheargilealathrserprotyrvalval
515520525
arglysserhisalahisseralaalaserserargalathralaala
530535540
leuhisargasnasppheserlyspheserarglyshisserlysgly
545550555560
valserargargargargargarg
565
<210>8
<211>162
<212>prt
<213>肌病毒科
<400>8
metthrthrilealapheaspglylysvalleualaalaaspglygln
151015
leuthrargglyserasnilecysasnleuasnthrglnlysleuphe
202530
ilecysproseraspgluglutrptrpilethrglygluargileval
354045
alapheglyvalserglyaspilethrglyglnleualaleuleuglu
505560
thrvalargleuargproglytyrlysglyleuthrglnsersergln
65707580
leuproalalysmetaspphethrpheleumetleuleualasergly
859095
lysalailevalglyglylysarggluaspaspvalthrleutrptrp
100105110
serglualathrproproleualathrglyserglytyrglutyrala
115120125
leuglyalametlysmetglyalaasnalavalargalavalgluile
130135140
alaserglucysaspiletyrthrglyglygluileserthrtyrglu
145150155160
leuarg
<210>9
<211>300
<212>prt
<213>phikz样巨型噬菌体
<400>9
metserileileleuasntrplysglnglnproglyglnthrleuasp
151015
serilegluiletyrargtyraspasnproargglnservalasnpro
202530
valalaproglygluproilevalthrleuproglyasnthrthrthr
354045
tyrgluasplysthrthrglualatyrlysthrtyrglntyrargile
505560
valalavallysglythrglulysvalmetglyleuproilevalgln
65707580
glyasppheprometthrglyproglyproglngluleuilearggly
859095
asptrphisargglytyrpheglythrleuthrasngluglupheile
100105110
leuasnhisalagluleuasnglyleuileglypheasnalatrpasn
115120125
glnalaprothrleuphehislysphevalphelysglyargileleu
130135140
pheileproaspthrvalthrargleuglythrthrtrpasnglugln
145150155160
tyrglnglnglyleualatrpglythraspasptyrglyphepropro
165170175
argservalserthrthrasnglnargargthrpheasnlysaspgly
180185190
tyrglutyrvalvalargleuproargleuglyasptyrsertyrgly
195200205
hisserthrtyrleuserserglyglytyrpheglnaspglyglutrp
210215220
hisasnthrphealaserleupheargtyralaglyvalvalargarg
225230235240
pheglyaspleuprothrargserglyglyserprothraspserser
245250255
alathrleuphealaasnglytyraspasnalathrprotrptyrtyr
260265270
argserasnserproaspserprosertyrthrthrserthrasnser
275280285
alaservalvalhisvalilegluleuvalleuser
290295300
<210>10
<211>299
<212>prt
<213>phikz样巨型噬菌体
<400>10
metproilethrleuhisphealaglnarglysproglnalaleuasp
151015
serilegluiletyrarglysthrproglyasnalathrileaspval
202530
asnalaproglythrproleualathrleuproglyaspalathrser
354045
tyrgluaspasnalavalgluasnasnthrthrtyrargtyrtrpile
505560
alaalavallysaspglygluargvalpheasnthrprothralagln
65707580
glyphepheleuaspthrglyproglyproglnlysleuleutyrgly
859095
asptrphisalaglytyrpheglythrvalthrproalagluleuphe
100105110
thrasnthrgluleuasnglyleuvalvalasnmetpheserserala
115120125
valglyalatrphislyspheiletyrlysasnargileleuphemet
130135140
seraspasnserileileasnalaserproglnpheiletyrasngln
145150155160
glymetmettyrglyglnaspglyasnglytyrvalproglytrpala
165170175
thralaargasnglnargargthrvalthrlysasnglytyrglutyr
180185190
valvalargleuprotyrleupheasptyrlysphetrpthrsertyr
195200205
valglyserglyasnthrgluleutyrleuaspglyglutrppheasn
210215220
thrphealaargleutyrasntyrserglyglnpheproargpheasp
225230235240
aspileprovalglyproileaspservalthralaglnglnserala
245250255
phephealaalametasnasnthrserserglntrptyrasnserpro
260265270
protyrprogluserprosertrpserasntyrglythrthrthrile
275280285
argserilealavalleugluleuvalleupro
290295
<210>11
<211>10
<212>prt
<213>phikz样巨型噬菌体
<400>11
serserglntrptyrasnserproprotyr
1510
<210>12
<211>10
<212>prt
<213>phikz样巨型噬菌体
<400>12
serserglntrptyrasnserproprotyr
1510
pct/ro/134表
- 还没有人留言评论。精彩留言会获得点赞!