一种高等真核生物非内含子嵌套基因NJx-1及其表达蛋白和应用
一种高等真核生物非内含子嵌套基因njx
‑
1及其表达蛋白和应用
技术领域
1.本发明属于生物工程技术领域,具体涉及一种高等真核生物非内含子嵌套基因njx
‑
1及其表达蛋白和应用。
背景技术:
2.重叠基因(overlapping gene)也叫嵌套基因(nested gene)是指两个或两个以上的基因共有一段dna序列,或是指一段dna序列成为两个或两个以上基因的组成部分。嵌套基因包括两大类:内含子嵌套基因和非内含子嵌套基因。内含子嵌套基因位于外部基因的内含子中,在高等真核生物中很常见,并且大多数被预测为编码蛋白质。非内含子嵌套基因在真核基因组中极为罕见,关于在高等真核生物(尤其是哺乳动物)中非内含子嵌套基因及其编码产物的鉴定和生物学功能的还未见报道。
3.狭颅症,又称颅缝早期融合症或颅狭窄畸形。表现有智能低下、精神活动异常、癫痫发作等。一般于生后6个月至1岁时手术。由于颅缝过早闭合,以致颅腔狭小不能适应脑的正常发育。狭颅症目前一旦确诊,没有合适的药物,只有手术;也没有早期诊断方法。
4.骨质疏松症被称为无声无息的流行病,是以单位体积内骨量减少及骨微结构改变为特征一种全身骨代谢障碍疾病,被形象地称为沉默的杀手。据统计大约30
‑
50%的女性和15
‑
30%的男性生涯患有骨质疏松,由于绝经原因,临床80%的病人为女性。预计到2020年,我国骨质疏松和低骨量患者人数将增加至2.8亿,这不但给社会带来巨大的压力,造成沉重的经济负担。目前的治疗药物主要有抑制骨吸收(抑制破骨细胞功能)和增加骨形成(增强成骨细胞功能)。遗憾的是目前具有促进骨形成作用的临床药物只有一个,甲状旁腺素(pth1
‑
34)。由于一直认为原发性骨质疏松关联的骨量减少无法恢复,因此发现促进骨形的新型分子具有重大的意义。
技术实现要素:
5.针对上述存在的问题,本技术所要解决的技术问题是提供一种高等真核生物非内含子嵌套基因njx
‑
1,满足制备用于治疗骨骼发育相关疾病的药物的使用需求。本技术所要解决的另一技术问题是提供一种上述njx
‑
1基因的表达蛋白。本技术还要解决一技术问题是提供一种上述njx
‑
1基因或蛋白的用途。
6.为了解决上述问题,本技术采用的技术方案为:
7.njx
‑
1基因在制备用于治疗骨骼相关疾病的药物中的应用;所述的njx
‑
1基因的碱基序列如seq id no.3或seq id no.4所示。
8.所述的应用,所述的骨骼相关疾病为狭颅症,骨质疏松。
9.njx
‑
1蛋白在制备用于治疗骨骼相关疾病的药物中的应用;所述的njx
‑
1蛋白的氨基酸序列如seq id no.1或seq id no.2所示,或在seq id no.1或seq id no.2所示的氨基酸序列基础上,由所述氨基酸序列中一个或多个氨基酸缺失、替换、添加或插入组成的蛋白
质和多肽,以及其中的一段氨基酸序列组成的肽。
10.所述的应用,所述的骨骼相关疾病为狭颅症,骨质疏松。
11.非内含子嵌套基因njx
‑
1,其碱基序列如seq id no.3或seq id no.4所示。
12.所述的非内含子嵌套基因njx
‑
1的表达蛋白,其氨基酸序列如seq id no.1或seq id no.2所示。
13.一种用于制备治疗狭颅症或骨质疏松药物的蛋白,在seq id no.1或seq id no.2所示的氨基酸序列基础上,由所述氨基酸序列中一个或多个氨基酸缺失、替换、添加或插入组成的蛋白质和多肽,以及其中的一段氨基酸序列组成的肽。
14.有益效果:与现有技术相比,本技术人在寻找抗骨质疏松症的药物靶标中,使用lc
‑
ms/ms从新生小鼠的颅盖细胞中找到一个15kda的未知蛋白,命名为njx
‑
1蛋白,该蛋白对应的基因序列为klf9的基因的部分片段的互补序列,为一个罕见的非内含子嵌套基因,且可以表达蛋白。本技术通过基因突变技术制备基因突变小鼠,证实基于njx
‑
1蛋白,或在本蛋白氨基酸序列基础上,由本蛋白的氨基酸序列中一个或多个氨基酸缺失、替换、添加或插入组成的蛋白质和多肽,以及其中的一段氨基酸序列组成的肽,在消除该基因影响终止njx
‑
1蛋白的表达后,该基因突变小鼠骨骼的发育发展受到极大的影响,具有明显的狭颅症,骨量减少(骨质疏松)的特点。可见通过调控njx
‑
1基因或蛋白信号通路使得该信号通路得到增强或减弱可达到对骨骼相关疾病的治疗,如但不限于狭颅症,骨质疏松等。因此,该基因及其突变可以用于狭颅症,骨质疏松等疾病的研究和药物开发。
附图说明
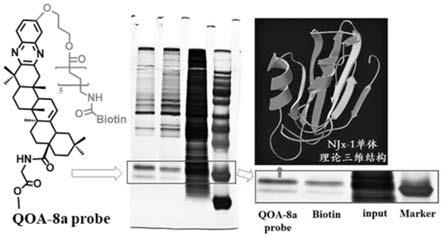
15.图1是njx
‑
1蛋白的发现过程图;通过钓蛋白和质谱分析的方式从乳小鼠头盖骨中总蛋白中发现njx
‑
1蛋白;包括qoa
‑
8a探针,电泳图;
16.图2是根据njx
‑
1蛋白的氨基酸序列转译成相应的dna序列,设计引物;进行pcr扩增,结果扩增产物与理论大小一致(a);pcr产物转化、涂平板、对平板上的单克隆菌落进行菌落pcr鉴定,结果1
‑
8号克隆均为阳性克隆(b);njx
‑
1蛋白的克隆表达的sds
‑
page和wb的结果图(c);lane m1:protein marker;lane m2:protein marker;lane 1:bsa(2.00μg);lane 2:njx
‑
1(reducing condition,2.00μg);lane 3:njx
‑
1(reducing condition);primary antibody:mouse
‑
anti
‑
his mab;
17.图3是njx
‑
1蛋白的在小鼠器官中和人源头盖骨中的分布图;b:basescope原位杂交分析人颅缝细胞中编码人njx
‑
1的rna的表达;箭头所标处表示njx
‑
1,苏木精复染细胞核;
18.图4上图是njx
‑
1基因突变小鼠制备策略,通过在klf9的外显子1上连续发生两个同义突变,获得njx
‑
1的无义突变;保留njx
‑
1 n端30个氨基酸,去除120个氨基酸;下图是基因突变小鼠;
19.图5是野生型和njx
‑1‑
ki雄性小鼠5周龄颅骨的micro
‑
ct图;a
‑
d,红色箭头表示额前缝合融合;a
‑
ii和b
‑
iv红色星号表示人字缝合融合;a
‑
iii红色箭头表示缝线混乱;b
‑
i
‑
iii图像分别显示扭曲的鼻和额骨;红色箭头表示鳞状缝线融合;星号表示牙齿错咬合;c
‑
iii和e色箭头分别表示枕孔和枕外孔;d
‑
箭头表示基底酚类与基底枕类之间的融合,星号表示碱性酚类低矿化;比例尺为1mm.f,颅骨长度、额骨、顶骨和顶骨间宽度分析;
iii,插入该蛋白的dna片段序列。其引物序列为:primer f:5
’‑
ctgttccaggggcccggatccctggccggccaacgtctgg
‑3’
;primer r:5
’‑
ctcgagtgcggccgcaagattttagcgcagggccaggtggcc
‑3’
。
30.pcr扩增,pcr产物胶回收纯化产物与载体酶切后胶回收产物经连接、转化涂平板,对平板上的单克隆菌落进行菌落pcr鉴定;结果1
‑
8号克隆均为阳性克隆(见图2a,2b)。
31.包涵体变复性纯化,主要步骤:1、大量表达和菌体高压破碎;2、包涵体制备和清洗;3、包涵体溶解和透析复性;4、组氨酸标签亲和层析填料柱上复性和纯化。
32.主要参数:1、wu破碎沉淀用2%doc+10mm dtt清洗和3%skl变性溶解:2、溶解后样品透析,透析液缓冲基础配方为:50mm tris
‑
hcl,250mm nacl,ph 7.5,5%甘油,0.5mm edta;逐渐降低skl浓度至终浓度1%;3、透析液中补加1mm gsh、0.1mm gssg和5%甘露醇,继续透析;4、逐渐降低skl浓度至终浓度0.01%;5、终止透析,样品过柱纯化。纯化后的蛋白见图2c。
33.njx
‑
1蛋白的在小鼠器官中和人源头盖骨中的分布,结果如图2所示;b:basescope原位杂交分析人颅缝细胞中编码人njx
‑
1的rna的表达;红点表示njx
‑
1,苏木精复染细胞核;结果显示njx
‑
1在人源头盖骨中存在。
34.3.njx
‑
1mrna的组织分布
35.由于njx
‑
1为未知蛋白,本实施例首先分析njx
‑
1mrna的组织分布。对于njx
‑
1的basescope rna原位杂交(高级细胞诊断)分析,小鼠胚胎(图3a)或人颅缝细胞(图3b)的8μm石蜡切片用过氧化氢预处理,然后使用rnascopeh2o2和蛋白酶regents试剂盒进行目标检索(acd,参考编号322381)和rnascopetarget retrieval regents(acd,参考编号322000)。将干燥的载玻片置于载玻片架上,并在hybez
tm
系统(acd)中与protease iii在40℃下孵育30分钟。然后,使用设计的探针(人,acd,参考编号888941;小鼠,acd,参考编号854121)根据basescope制造商的协议,使用basescope检测试剂v2
‑
red(acd,参考编号323910),阳性(人类ppib,acd,参考编号701031;小鼠ppib,acd,参考编号701071)和阴性(dapb,acd,参考编号701011)rna探针作为对照。
36.结果显示njx
‑
1的mrna在小鼠的肝,肺,骨,胰腺中含量最高;心脏,肌肉,肠,甲状腺,胃,肾,脂肪等组织中也存在(图3a)。另外在人体的颅骨中也存在(图3b)。
37.实施例2
38.1.njx
‑
1基因突变小鼠的制备
39.njx
‑
1基因敲入策略用于njx
‑
1基因突变小鼠的制备。使用同义密码子替换策略来制造njx
‑
1敲入小鼠(以下称为njx
‑1‑
ki小鼠)。klf9的157ser&158ser密码子在njx
‑
1基因第四次atg(30met)之后通过crispr/cas9系统编辑
‑
tcc>tca
‑
k1f9基因的氨基酸序列仍为丝氨酸,反义njx
‑
1基因变成了两个连续的终止密码子,终止了njx
‑
1基因的翻译(基因序列突变点见图4中小方框的表示)。
40.测序鉴定结果表明,实现了对njx
‑
1基因的突变(由于有两个连续的终止密码,保留了njx
‑
1n端30个氨基酸,去除120个氨基酸);同样,对与njx
‑
1相似的基因序列测序,结果无突变,未发现脱靶效应。经过后续f1代的繁殖,成功得到了野生型,杂合子及纯合子小鼠,经统计,符合孟德尔遗传定律。测序鉴定结果表明实现了对njx
‑
1基因的突变。新生小鼠的总体外观未显示明显异常,但体型稍小(见右图)。到16周时,约半数基因突变小鼠体重与野
生型相似(njx
‑1+/+
),半数体重明显小于野生型(统计结果基因小鼠全体体重无论雌雄均比野生型小,p<0.05)。另外一些基因小鼠的颅面出现明显的畸形,扭曲等(见图4)。
41.2.njx
‑
1基因突变小鼠颅骨发育异常
42.观察njx
‑
1基因突变的小鼠(5周龄),μct发现小鼠头部发育明显异常,主要表现为面颅变短,鼻骨歪斜(图5),严重的可以观察到明显的脑积水。μct显示njx
‑
1基因突变小鼠骨缝线融合明显,鼻骨变短,额骨变宽,与在新生婴儿中发病率极高(约1/2000)的狭颅症相似。
43.3.njx
‑
1基因突变小鼠骨发育异常,导致骨质疏松
44.对小鼠的长骨和椎骨(16周龄)进行考察。外形差别不大的基因突变小鼠(ki/ki normal weight,体重与野生型(wt/wt)相似),μct显示远股骨端(distal femur)骨小梁,皮质骨厚度均明显减少(图6);那些体重显著小于野生型的基因突变小鼠(ki/ki low weight)的骨量下降极其严重。同样,椎骨(vertebra)的骨量也显著下降。以上结果显示njx
‑
1基因对小鼠的骨发育产生重要影响。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1