用于信使RNA体外转录的改进方法与流程
用于信使rna体外转录的改进方法
1.相关申请的交叉引用
2.本技术要求2020年2月18日提交的美国临时申请序列号62/978,180的优先权,该临时申请的公开内容据此以引用的方式并入。
3.序列表
4.本说明书参考了序列表(于2020年2月18日以电子形式提交的名为mrt-2121usp1_sl的.txt文件)。.txt文件创建于该日期,并且大小为24,675字节。该序列的全部内容以引用的方式并入本文。
背景技术:
5.mrna疗法对于治疗各种疾病变得越来越重要。据报道,t7和sp6rna聚合酶在mrna的体外合成期间都产生异常终止转录物(nam等人,1988,the journal of biological chemistry,263:34,pp 18123-18127;lee等人,nucleic acids research 2010,1-9)。在基于体外合成的mrna的治疗组合物中,这种异常终止转录物的存在会影响其安全性和有效性。
6.已知特别是由t7 rna聚合酶产生的mrna转录物被比所需转录物更长和更短的rna污染,例如由于失控转录产生比模板序列更长的延长的转录物。转录物的这些非模板化的延长的部分可以与rna分子本身或另一个rna分子退火,以形成分子内或分子间rna双链体(gholamalipour等人2018,nucleic acids research,46:18pp 9253-9263)。rna双链体可以是高度免疫原性的(mu等人2018,nucleic acids research,46:5239-5249)。使用标准实验室方案不能有效地从体外转录的(ivt)mrna中去除rna双链体杂质。最有效的纯化方法被认为是离子对反相高效液相色谱法(hplc)。然而,这种方法不可成规模使用,需要使用有毒试剂,并且对于许多实验室来说过于昂贵(等人2019,molecular therapy:nucleic acids,15:26-35)。在含乙醇的缓冲液中,双链rna与纤维素的选择性结合最近被确定为从ivt mrna中去除rna双链杂质的可成规模使用的方法,尽管这种方法导致rna产量的显著降低(等人2019,同上)。
7.sp6 rna聚合酶已被用作t7 rna聚合酶的替代物。然而,当sp6 rna聚合酶用于体外转录时,不完整的mrna转录物仍然是个问题。以前曾报道过sp6 rna聚合酶在rrnb t1终止子中的两个信号(上游和下游信号)处停止转录,并且信号区的改变影响终止效率(kwon&kang 1999,the journal of biological chemistry,274:41pp 29149-29155)。本发明人发现rrnb t1样终止信号经常存在于用于mrna体外转录的模板dna序列中。另外,他们发现sp6 rna聚合酶也可以发生失控转录。
8.wo 2017/009376提供了从环状dna产生rna的方法,其中环状dna模板序列包括rna聚合酶启动子序列,然后是编码自切割核酶的序列,然后是rna聚合酶终止子序列元件。本技术中包括的数据证实对于从包含自切割核酶和两个或四个终止子序列的线性化dna质粒进行的体外转录,终止效率可达到约95%。这种量级的终止效率不足以用于生产治疗性mrnas的商业规模过程。
9.wo 2012/170443提供了从环状dna模板产生rna的方法,其中噬菌体启动子可操作地连接到编码感兴趣的rna多核苷酸的序列,该序列可操作地连接到多终止子结构域。多终止子结构域包含至少三个选自i类和ii类终止信号的终止信号。i类终止信号(以phi噬菌体t7终止子为例,也称为t7 phi终止子)编码rna序列,该序列可以形成稳定的茎环结构,然后是一连串6个u残基。ii类终止信号(以人类前甲状旁腺激素原(pth)基因为例)编码6个u残基的中断序列,但缺乏明显的茎环结构。rrnb t1终止信号是ii类终止信号。像在wo 2017/009376中一样,在wo 2012/170443的实施例中测试的dna模板包括在编码感兴趣的rna多核苷酸的序列和由两个终止子(i类)、两个ptg终止子(ii类)和一个pbr322终止子(i类)组成的多重终止子结构域之间编码自切割核酶的序列。
10.du等人(2009,biotechnol.biogen.,104(6):1189-1196)认为终止子的大尺寸(100bp)和低效率是有问题的,并且试图通过依次包括1-3个水疱性口炎病毒(vsv)ii类终止信号(tatctgttagtttttttc)(每个信号由8个碱基对分开)来提高从环状dna模板转录期间的终止效率。他们发现,当使用单个vsv终止信号时,终止效率仅为53%-62%。当使用2-3个vsv终止子时,终止效率增加到65%-75%。
11.因此,需要改进的体外转录方法,其产生没有过早终止的转录物和双链mrna的全长mrna转录物。
技术实现要素:
12.本发明通过提供制备优化的dna序列作为mrna体外转录的模板的方法来满足这种需求。优化这些dna序列以避免rna聚合酶过早终止转录。另外,本发明还提供了用于制备优化的dna序列的方法,该序列在其3’末端包括一个或多个终止信号。终止信号减少或防止失控转录,并且因此使用这些优化的dna序列使双链mrna转录物的形成最小化。
13.在一方面,本发明涉及用于制备编码蛋白质的优化的dna序列作为体外转录模板的方法,所述方法包括:(a)提供包含蛋白质编码序列的dna序列;(b)确定dna序列中终止信号的存在,其中终止信号具有以下核酸序列:5
’‑
x1atctx2tx
3-3’(seq id no:1),其中x1、x2和x3独立地选自a、c、t或g;以及(c)如果存在一个或多个终止信号,通过用其它三个核酸中的任何一个替换所述一个或多个终止信号的位置2、3、4、5和7中任何一个位置的一个或多个核酸来修饰dna序列,以产生优化的dna序列,其中,如果需要,选择一个或多个替换核酸以保留由蛋白质编码序列编码的蛋白质的氨基酸序列。
14.在一些实施方案中,步骤b和c由计算机执行。
15.在一些实施方案中,该dna序列进一步包含编码5’utr的第一核酸序列和/或编码3’utr的第二核酸序列。
16.在一些实施方案中,dna序列中紧邻终止信号的3’的5个核苷酸不包含3个或更多个t核苷酸。
17.在一些实施方案中,该方法进一步包括相对于编码相同蛋白质序列的野生型dna序列修饰dna序列的步骤,以优化:(a)与mrna加工和稳定性相关的元件;和/或(b)与翻译或蛋白质折叠相关的元件;其中修饰在产生优化的dna序列之前进行。与mrna加工或稳定性相关的元件可以包括隐蔽剪接位点、mrna二级结构、mrna的稳定自由能、重复序列和rna不稳
定基序。与翻译或蛋白质折叠相关的元件可以包括密码子使用偏好性、密码子适应性、内部chi位点、核糖体结合位点、过早终止聚a位点、shine-dalgarno序列、密密码子背景、密码子-反密码子相互作用和翻译暂停位点。
18.在一些实施方案中,该方法进一步包括合成优化的dna序列的步骤。该方法可以进一步包括将合成的优化的dna序列插入用于在体外转录中使用的核酸载体。核酸载体可以包含可操作地连接到优化的dna序列的rna聚合酶启动子,任选地,其中rna聚合酶是sp6 rna聚合酶或t7rna聚合酶。在一些实施方案中,核酸载体是质粒。在体外转录之前,可以将质粒线性化。
19.在一些实施方案中,该方法进一步包括在体外转录中使用合成的优化的dna序列来合成mrna。mrna可以由sp6 rna聚合酶合成。sp6 rna聚合酶可以是天然存在的sp6 rna聚合酶或重组sp6聚合酶。重组sp6聚合酶可以包含标签(例如his标签)。在一些实施方案中,mrna由t7 rna聚合酶合成。
20.在一些实施方案中,该方法进一步包括对合成的mrna加帽和/或加尾的单独步骤。在一些实施方案中,加帽和加尾发生在体外转录期间。
21.在一些实施方案中,mrna在反应混合物中合成,所述反应混合物包含每种ntp浓度范围为1-10mm的ntp、浓度范围为0.01-0.5mg/ml的dna模板和浓度范围为0.01-0.1mg/ml的sp6 rna聚合酶。例如,反应混合物可以包含每种ntp浓度为5mm的ntp、浓度为0.1mg/ml的dna模板,和浓度为0.05mg/ml的sp6 rna聚合酶。ntp可以是天然存在的ntp,或者可以包括经修饰的ntp。
22.在一些实施方案中,mrna可以在37℃-56℃的温度范围内合成。
23.在一些实施方案中,提供了包含指令的计算机程序,当该程序由计算机执行时,所述指令使计算机(a)接收包含蛋白质编码序列的dna序列,并且(b)执行上述方法的步骤b和c,用于制备编码蛋白质的优化的dna序列,作为本发明体外转录的模板。本发明还提供了一种计算机可读数据载体,其上存储有本发明的计算机程序。本发明另外提供了一种承载本发明的计算机程序的数据载体信号。本发明另外提供了一种数据处理系统,其包括用于实施制备编码蛋白质的优化dna序列的方法的工具,所述dna序列作为本发明的体外转录的模板。
24.在另一方面,本发明涉及一种制备编码蛋白质的优化的dna序列作为体外转录模板的方法,所述方法包括:(a)提供编码蛋白质的dna序列;以及(b)在dna序列的3’末端添加一个或多个终止信号以提供优化的dna序列,其中一个或多个终止信号包含以下核酸序列:5
’‑
x1atctx2tx
3-3’(seq id no:1),其中x1、x2和x3独立地选自a、c、t或g。
25.在一些实施方案中,终止信号包含核酸序列5
’‑
x1atctgtt-3’(seq id no:2)。
26.在一些实施方案中,x1是t。在一些实施方案中,x1是c。
27.在一些实施方案中,终止信号选自5’ttttatctgttttttt-3’(seq id no:3)、5’ttttatctgttttttttt-3’(seq id no:4)、5’cgttttatctgttttttt-3’(seq id no:5)、5’cgttccatctgttttttt-3’(seq id no:6)、5’cgttttatctgtttgttt-3’(seq id no:7)、5’cgttttatctgtttgttt-3’(seq id no:8)或5’cgttttatctgttgtttt-3’(seq id no:9)。
28.在一些实施方案中,将两个或更多个、三个或更多个、四个或更多个终止信号添加至dna序列的3’末端。
29.在一些实施方案中,编码蛋白质的dna序列可以进一步包含编码5’utr的第一核酸序列和/或编码3’utr的第二核酸序列。dna序列可以进一步包含或不包含编码聚a尾的第三核酸序列。
30.在一些实施方案中,编码蛋白质的dna序列不进一步包含编码核酶的dna序列。
31.在一些实施方案中,编码蛋白质的dna序列中紧邻终止信号3’的5个核苷酸不包含3个或更多个t核苷酸。
32.在一些实施方案中,dna序列包括一个以上的终止信号,并且所述终止信号相隔10个碱基对或更少个碱基对,例如相隔5-10个碱基对。
33.在一些实施方案中,优化的dna序列包含以下序列:(a)5
’‑
x1atctx2tx
3-(zn)-x4atctx5tx
6-3’(seq id no:10)或(b)5
’‑
x1atctx2tx
3-(zn)-x4atctx5tx
6-(zm)-x7atctx8tx
9-3’(seq id no:11),其中x1、x2、x3、x4、x5、x6、x7、x8和x9独立地选自a、c、t或g,zn代表n个核苷酸的间隔序列,并且zm代表m个核苷酸的间隔序列,每个核苷酸独立地选自a、c、t或g,并且其中n和/或m独立地为10或更小。在一些实施方案中,n是5、6、7、8、9或10和/或m是5、6、7、8、9、10。在一些实施方案中,z是t。
34.在一些实施方案中,该方法进一步包括相对于编码相同蛋白质序列的野生型dna序列修饰dna序列的步骤,以优化:(a)与mrna加工和稳定性相关的元件;和/或(b)与翻译或蛋白质折叠相关的元件;其中修饰在产生优化的dna序列之前进行。与mrna加工或稳定性相关的元件可以包括隐蔽剪接位点、mrna二级结构、mrna的稳定自由能、重复序列和rna不稳定基序。与翻译或蛋白质折叠相关的元件可以包括密码子使用偏好性、密码子适应性、内部chi位点、核糖体结合位点、过早终止聚a位点、shine-dalgarno序列、密密码子背景、密码子-反密码子相互作用和翻译暂停位点。
35.在一些实施方案中,该方法可以进一步包括将优化的dna序列插入用于在体外转录中使用的核酸载体。
36.在另一方面,本发明涉及用于在体外转录中使用的dna序列,以5’到3’的顺序包含:(a)5’utr;(b)蛋白质编码序列;(c)3’utr;(d)任选的编码聚a尾的核酸序列;和(e)终止信号;其中终止信号包含下列核酸序列:5
’‑
x1atctx2tx
3-3’(seq id no:1),其中x1、x2和x3独立地选自a、c、t或g。在一些实施方案中。
37.在一些实施方案中,x1是t。在一些实施方案中,x1是c。
38.在一些实施方案中,dna序列的终止信号选自5’ttttatctgttttttt-3’(seq id no:3)、5’ttttatctgttttttttt-3’(seq id no:4)、5’cgttttatctgttttttt-3’(seq id no:5)、5’cgttccatctgttttttt-3’(seq id no:6)、5’cgttttatctgtttgttt-3’(seq id no:7)、5’cgttttatctgtttgttt-3’(seq id no:8)或5’cgttttatctgttgtttt-3’(seq id no:9)。
39.在一些实施方案中,dna序列可以包含一个以上的终止信号,例如两个或更多个、三个或更多个、四个或更多个终止信号。在一些实施方案中,终止信号相隔10个碱基对或更少个碱基对,例如相隔5-10个碱基对。
40.在一些实施方案中,dna序列包含以下序列:(a)5
’‑
x1atctx2tx
3-(zn)-x4atctx5tx
6-3’(seq id no:10)或(b)5
’‑
x1atctx2tx
3-(zn)-x4atctx5tx
6-(zm)-x7atctx8tx
9-3’(seq id no:11),其中x1、x2、x3、x4、x5、x6、x7、x8和x9独立地选自a、c、t或g,zn代表n个核苷酸的间隔序列,并且zm代表m个核苷酸的间隔序列,每个核苷酸独立地选自a、c、t或g,并且
其中n和/或m独立地为10或更小。在一些实施方案中,n为5、6、7、8、9或10和/或m为5、6、7、8、9或10。在一些实施方案中,z是t。
41.在一些实施方案中,终止信号不存在于dna序列的5’utr、蛋白质编码序列和3’utr。
42.在一些实施方案中,编码蛋白质的dna序列不进一步包含编码核酶的dna序列。
43.在一些实施方案中,相对于编码相同蛋白质序列的野生型dna序列修饰dna序列,以优化:(a)与mrna加工和稳定性相关的元件;和/或(b)与翻译或蛋白质折叠相关的元件。
44.在一些实施方案中,本发明进一步提供了包含本发明dna序列的核酸载体。核酸载体可以包含可操作地连接到优化的dna序列的rna聚合酶启动子,任选地,其中rna聚合酶是sp6 rna聚合酶或t7 rna聚合酶。在一些实施方案中,核酸载体是质粒。
45.在一些实施方案中,本发明还提供了用于在体外转录中使用的试剂盒,其包含本发明的dna序列或核酸载体。该试剂盒可以进一步包含ntp和rna。
46.另一方面,本发明涉及产生mrna的方法,所述方法包括将本发明的核酸载体添加到包含ntp和rna聚合酶的反应混合物中,其中rna聚合酶将dna序列转录成mrna转录物。核酸载体可以是质粒,其在体外转录之前可以线性化或可以不线性化。rna聚合酶可以是sp6 rna聚合酶。sp6 rna聚合酶可以是天然存在的sp6 rna聚合酶或重组sp6rna聚合酶。重组sp6 rna聚合酶可以包含标签(例如his标签)。替代地,rna聚合酶可以是t7 rna聚合酶。
47.在一些实施方案中,产生mrna的方法进一步包括对合成的mrna加帽和/或加尾的单独步骤。在一些实施方案中,加帽和加尾发生在体外转录期间。
48.在一些实施方案中,mrna在反应混合物中合成,所述反应混合物包含每种ntp浓度范围为1-10mm的ntp、浓度范围为0.01-0.5mg/ml的dna模板和浓度范围为0.01-0.1mg/ml的sp6 rna聚合酶。例如,反应混合物可以包含每种ntp浓度为5mm的ntp、浓度为0.1mg/ml的dna模板,和浓度为0.05mg/ml的sp6 rna聚合酶。ntp可以是天然存在的ntp,或者可以包括经修饰的ntp。
49.在一些实施方案中,mrna可以在37℃-56℃的温度范围内,例如在50℃-52℃下合成。
50.在一些实施方案中,用于产生mrna的方法可导致至少80%、至少85%、至少90%、至少95%的mrna转录物在终止信号处终止。终止位点可以通过(i)消化所述mrna以产生小于100个核苷酸大小的3’末端片段,和(ii)通过液相色谱分析所述3’末端片段来确定。终止位点可以通过rna测序来确定。
51.在一些实施方案中,rna聚合酶是t7 rna聚合酶,并且其中mrna转录物基本上不含rna双链体。相对于对照,mrna转录物可以含有不可检测水平的rna双链体。可以用特异性结合dsrna的抗体检测rna双链体。
52.可以将本文所述的任何方面或实施方案与如本文所公开的任何其他方面或实施方案组合。尽管已经结合本公开的具体实施方式描述了本公开,但是以上描述旨在说明而不是限制本公开的范围,本公开的范围由所附权利要求的范围限定。其他方面、优点和修改在以下权利要求的范围内。
53.本文援引的专利和科学文献建立了本领域技术人员可获得的知识。本文引用的所有美国专利和已公开或未公开的美国专利申请均以引用的方式并入本文。本文引用的所有
已公开的外国专利和专利申请均据此以引用的方式并入。本文引用的所有其他已公开的参考文献、文件、手稿和科学文献据此以引用的方式并入。
54.从附图以及下面的详细描述(包括实施例和权利要求书),本发明的其他特征和优点将变得显而易见。
附图说明
55.当结合附图时,根据下面的详细描述,将更清楚地理解上述以及另外的特征。然而,附图仅用于说明性目的,而不是限制。
56.图1,部分i是显示用sp6 rna聚合酶合成的mrna-1的毛细管电泳图谱的电泳图。
57.图2是通过毛细管电泳对mrna-1和用sp6 rna聚合酶合成的在tatctgtt终止信号序列中具有点突变的mrna-1变体的总rna进行定量分析产生的数字凝胶图像。
58.图3是显示在用sp6 rna聚合酶或t7 rna聚合酶制备的mrna样品中检测到的dsrna量的斑点杂交图像。用鼠单克隆抗体j2确定dsrna的存在,使用辣根过氧化物缀合的抗鼠igg抗体进行检测。用sp6 rna聚合酶制备的样品中可能存在的任何dsrna都低于检测下限(llod)。用t7 rna聚合酶制备的样品中dsrna的量超过25ng。
59.图4提供了sp6 mrna转录物3’末端的分析结果。使用rna酶h消化由sp6 rna聚合酶转录的mrna,并且通过液相色谱质谱(lc/ms)分析3’末端消化产物(图4a),并根据质谱确定的片段大小鉴定片段(图4b)。
60.图5比较了sp6 rna聚合酶(上图)和t7 rna聚合酶(下图)mrna转录物的非模板延长。通过lc/ms(图5a)和rna测序(图5b)确定模板转录后添加到mrna转录物3’末端的额外核苷酸的数量。
61.图6提供了显示用sp6 rna聚合酶从线性化质粒合成的mrna-12的毛细管电泳图谱(部分i)的电泳图谱。该质粒或者是未经修饰的(图6a),或者是通过在编码mrna转录物的dna序列的3’末端添加一个(图6b)或两个(图6c)rrnb终止t1信号而修饰的。
62.图7提供了显示用sp6 rna聚合酶从超螺旋(非线性化)质粒合成的mrna-12的毛细管电泳图谱(部分i)的电泳图。该质粒或者是未经修饰的(图7a),或者是通过在编码mrna转录物的dna序列的3’末端添加一个(图7b)或两个(图7c)rrnb终止t1信号而修饰的。
63.图8提供了显示在37℃(图8a)或50℃(图8b)下用sp6 rna聚合酶从超螺旋(非线性化)质粒合成的mrna-12的毛细管电泳图谱(部分i)的电泳图。通过在编码mrna转录物的dna序列的3’末端添加两个rrnb终止t1信号来修饰质粒。
64.图9提供了电泳谱,显示了在37℃(图9a、9c、9e、9g)或50℃(图9b、9d、9f、9h)用sp6 rna聚合酶从超螺旋(非线性)质粒合成的mrna-12产生的毛细管电泳图谱的电泳图。该质粒或者是未经修饰的(图9a、9b),或者是通过在编码mrna转录物的dna序列的3’末端添加一个(图9c、9d)、两个(图9e、9f)或三个(图9g、9h)rrnb终止t1信号而修饰的。
65.图10比较了从线性化的未修饰质粒(不包含终止序列)转录的mrna-12和从超螺旋质粒转录的mrna-12表达的蛋白质水平,所述超螺旋质粒通过在编码mrna转录物的dna序列的3’末端添加三个rrnb终止t1信号而被修饰。
66.定义
67.为了使本发明更容易理解,首先在下文定义了某些术语。以下术语和其他术语的
其他定义阐述在整个说明书中。
68.除非上下文另有明确说明,否则在本说明书和所附权利要求书中使用的单数形式“一个”、“一种”和“所述”包括复数指代。
69.除非特别声明或根据上下文显而易见,否则如本文所用,术语“或”应理解为是包含性的并且涵盖“或”与“和”两者。
70.如本文所用的术语“例如”和“即”仅作为实例使用,而不意欲进行限制,并且不应解释为仅指代本说明书中明确列举的那些条目。
71.术语“或更多个”、“至少”、“超过”等,例如,“至少一个”应理解为包括但不限于至少1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149或150、200、300、400、500、600、700、800、900、1000、2000、3000、4000、5000个或超过所陈述值。还包括其间的任何较大数目或分数。
72.相反,术语“不超过”包括小于所陈述值的每个值。例如,“不超过100个核苷酸”包括100、99、98、97、96、95、94、93、92、91、90、89、88、87、86、85、84、83、82、81、80、79、78、77、76、75、74、73、72、71、70、69、68、67、66、65、64、63、62、61、60、59、58、57、56、55、54、53、52、51、50、49、48、47、46、45、44、43、42、41、40、39、38、37、36、35、34、33、32、31、30、29、28、27、26、25、24、23、22、21、20、19、18、17、16、15、14、13、12、11、10、9、8、7、6、5、4、3、2、1和0个核苷酸。还包括其间的任何较小数目或分数。
73.术语“多个”、“至少两个”、“两个或更多个”、“至少第二”等应理解为包括但不限于至少2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149或150、200、300、400、500、600、700、800、900、1000、2000、3000、4000、5000个或更多个。还包括其间的任何较大数目或分数。
74.在整个说明书中,词语“包括”或者诸如“包含”或“含有”的变化形式将被理解为意味着包括所陈述的要素、整体或步骤、或者要素、整体或步骤的组,但不排除任何其他要素、整体或步骤、或者要素、整体或步骤的组。
75.除非特别声明或根据上下文显而易见,否则如本文所用,术语“约”应理解为在本领域的正常公差范围内,例如在平均值的2个标准偏差内。“约”可以理解为在所陈述值的10%、9%、8%、7%、6%、5%、4%、3%、2%、1%、0.5%、0.1%、0.05%、0.01%或0.001%内。除非上下文中另有说明,否则本文提供的所有数值反映了技术人员可以理解的正常波动。
76.如本文所用,术语“异常终止转录物”或“预终止转录物”等是比由dna模板编码的全长mrna分子更短的任何转录物,其由rna聚合酶以不依赖序列的方式从模板dna中过早释放产生。在一些实施方案中,异常终止转录物可以小于从靶dna分子转录的全长mrna分子长度的90%,例如,小于全长mrna分子长度的80%、70%、60%、50%、40%、30%、20%、10%、5%、1%。
77.如本文所用,术语“批次”是指一次合成的(例如,在同一制造周期期间根据单个制造订单所生产的)mrna的数量或量。批次可以是指在通过将酶的单个等分试样和/或dna模板的单个等分试样用于在一组条件下连续合成而发生的一个反应中合成的mrna的量。在一些实施方案中,批次将包括由其中随着反应进行并非补充和/或添加所有试剂和/或组分的反应生产的mrna。术语“批次”将不表示在不同时间合成的被组合以达到所需量的mrna。
78.如本文所用,术语“密码子优化”和“密码子优化的”是指编码肽、多肽或蛋白质的天然存在的或野生型核酸的密码子组成的修饰,这不改变其氨基酸序列,从而改善所述核酸的蛋白质表达。可以进行对天然存在的或野生型核酸的此类修饰以达到尽可能高的g/c含量,调整密码子使用以避免稀有或速率限制密码子,以去除不稳定的核酸序列或基序和/或以消除暂停位点或终止子信号。
79.如本文所用,术语“递送”涵盖局部递送和全身递送。例如,mrna的递送涵盖其中将mrna递送至靶组织并且编码的蛋白质在靶组织内表达且保留的情况(也称为“局部分布”或“局部递送”),以及其中将mrna递送至靶组织并且编码的蛋白质表达且分泌到患者的循环系统(例如,血清)内,并且全身分布且被其他组织吸收的情况(也称为“全身分布”或“全身递送”)。
80.如本文所用,术语“药物”、“药物治疗”、“治疗剂”、“活性剂”、“治疗化合物”、“组合物”或“化合物”可互换使用,并且是指可用于治疗或预防疾病、病症或身体功能障碍的任何化学实体、药剂、药物、生物制品、植物制剂等。药物可以包含已知的和潜在的治疗化合物。通过使用本领域普通技术人员已知的筛选方法进行筛选,可以确定药物是治疗性的。“已知的治疗化合物”、“药物”或“药物治疗”是指已经显示出(例如通过动物试验或先前向人施用的经验)在这种治疗中有效的治疗化合物。“治疗方案”涉及包括本文公开的“药物”、“药物治疗”、“治疗剂”、“活性剂”、“治疗化合物”、“组合物”或“化合物”的治疗,和/或包括受试者行为矫正的治疗,和/或包括手术手段的治疗。
81.如本文所用,术语“包封”或语法等同形式是指将mrna分子限制在纳米颗粒内的过程。将所需mrna掺入纳米颗粒内的过程经常称为“加载”。示例性方法描述于lasic等人,febs lett.,312:255-258,1992中,该文献以引用方式并入本文。掺入纳米颗粒的核酸可以完全或部分地位于纳米颗粒的内部空间中、双层膜内(对于脂质体纳米颗粒)或与纳米颗粒的外表面结合。
82.如本文所用,核酸序列的“表达”是指以下事件中的一个或多个:(1)从dna序列生产rna模板(例如,通过转录);(2)rna转录物的加工(例如,通过剪接、编辑、5'帽形成和/或3'末端形成);(3)将rna翻译成多肽或蛋白质;以及/或(4)多肽或蛋白质的翻译后修饰。在本专利申请中,术语“表达”和“产生”以及语法等价物可互换使用。
83.如本文所用,“全长mrna”是当使用特定的测定(例如凝胶电泳和使用uv和uv吸收光谱的检测)以及通过毛细管电泳分离时进行表征。编码全长多肽的mrna分子的长度是从
靶dna转录的全长mrna分子的长度的至少50%,例如,从靶dna转录的全长mrna分子的长度的至少60%、70%、80%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.01%、99.05%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%、99.9%。
84.如本文所用,术语“改进”、“增加”或“减少”或语法等同项表示相对于基线测量值,诸如在本文所述治疗开始之前同一个体的测量值,或在没有本文所述治疗的情况下对照受试者(或多个对照受试者)的测量值的值。“对照受试者”是患有与所治受试者相同形式的疾病,其年龄与所治受试者大约相同的受试者。
85.如本文使用的,术语“杂质”是指在限定量的液体、气体或固体内,与目标材料或化合物的化学组成不同的物质。杂质也被称为污染物。
86.如本文所用,术语“体外”是指在人工环境中,例如在试管或反应容器中,在细胞培养物中等,而不是在多细胞生物体内发生的事件。
87.如本文所用,术语“体内”是指在多细胞生物体诸如人和非人动物内发生的事件。在基于细胞的系统的上下文中,所述术语可用于指在活细胞内发生的事件(与例如体外系统相反)。
88.如本文所用,术语“分离的”是指(1)与最初生产时(无论是天然的和/或在实验环境中)与其相关联的至少一些组分分离,和/或(2)由人工生产、制备和/或制造的物质和/或实体。
89.如本文所用,术语“信使rna(mrna)”是指编码至少一种多肽的多核苷酸。如本文所用,mrna涵盖经修饰的和未经修饰的rna二者。mrna可以含有一个或多个编码和非编码区。mrna可以从天然来源纯化,使用重组表达系统来产生以及任选地纯化、体外转录或化学合成。在适当的情况下,例如在化学合成分子的情况下,mrna可以包含核苷类似物,诸如具有化学修饰的碱基或糖、骨架修饰等的类似物。除非另有说明,否则mrna序列的显示方向为5’至3’。
90.mrna通常被认为是将信息从dna传递到核糖体的rna类型。mrna的存在通常非常短暂,并且包括加工和翻译,然后是降解。通常,在真核生物体中,mrna加工包括在n(5
′
)末端添加一个“帽”,和在c(3
′
)末端添加一个“尾巴”。典型的帽是7-甲基鸟苷帽,其是通过5'-5'-三磷酸键与第一个转录的核苷酸连接的鸟苷。帽的存在对于提供对存在于大多数真核细胞中的核酸酶的抗性很重要。尾巴通常是聚腺苷酸化事件,由此聚a部分被添加到mrna分子的3'末端。这种“尾巴”的存在用于保护mrna免受核酸外切酶降解。信使rna通常被核糖体翻译成组成蛋白质的一系列氨基酸。
91.如本文所用,术语“核酸”在其最广泛的意义上是指掺入或可以掺入多核苷酸链中的任何化合物和/或物质。在一些实施方案中,核酸是由磷酸二酯键掺入或可以由磷酸二酯键掺入多核苷酸链的化合物和/或物质。在一些实施方案中,“核酸”是指单独的核酸残基(例如,核苷酸和/或核苷)。在一些实施方案中,“核酸”是指包含单个核酸残基的多核苷酸链。在一些实施方案中,“核酸”涵盖rna以及单链和/或双链dna和/或cdna。此外,术语“核酸”、“dna”、“rna”和/或类似术语包括核酸类似物,即具有除磷酸二酯骨架以外的类似物。除非另外指明,否则核酸序列以5'至3'方向呈现。
92.如本文所用,术语“过早终止”是指在全长dna模板被转录之前转录的终止。过早终
止是由dna模板中终止信号的存在引起的,并导致比全长mrna更短的mrna转录物(“过早终止的转录物”或“截短的mrna转录物”)。终止信号的实例包括大肠杆菌rrnb终止子t1信号(共有序列:atctgtt)及其变体,如本文所述。
93.如本文所用,术语“失控转录”是指在mrna转录物的末端非模板化添加核酸。如本文所述,rna聚合酶在遇到转录终止信号后,继续以非模板介导的方式延长mrna转录物。添加的序列在本文中被称为“失控”或“失控序列”。在一些实施方案中,失控序列可能能够自我退火或与模板化mrna转录物的部分退火,以形成双链或双链体rna。
94.如本文所用,术语“短聚体”用于特指过早终止的短mrna寡核苷酸,也称为短终止rna转录物,其是体外转录反应期间不完全mrna转录的产物。在说明书中,短聚体、过早终止的mrna、预终止的mrna或短终止的mrna转录物可互换使用。
95.如本文所用,术语“基本上”是指表现出全部或接近全部范围或程度的目标特征或特性的定性条件。生物学领域的普通技术人员将理解,生物学和化学现象很少(如果曾经有)完成和/或继续完成或达到或避免绝对结果。因此,在本文中使用术语“基本上”来捕获许多生物学和化学现象中固有的潜在完整性缺失。
96.如本文所用,术语“模板dna”(或“dna模板”)通常涉及包含核酸序列的dna分子,所述核酸序列编码待通过体外转录合成的mrna转录物。模板dna用作体外转录的模板,以产生由模板dna编码的mrna转录物。模板dna包含体外转录所必需的所有元件,特别是用于结合dna依赖性rna聚合酶的启动子元件,诸如例如t3、t7和sp6 rna聚合酶,其可操作地连接到编码所需mrna转录物的dna序列上。此外,模板dna可包含编码mrna转录物的dna序列的5’和/或3’引物结合位点,以例如通过pcr或dna测序确定编码mrna转录物的dna序列的身份。本发明上下文中的“模板dna”可以是线性或环状dna分子。如本文所用,术语“模板dna”可指dna载体,诸如质粒dna,其包含编码所需mrna转录物的核酸序列。
97.除非另有定义,否则本文所用的所有技术和科学术语具有与本技术所属领域的普通技术人员通常所理解的以及在本技术所属领域中常用的含义相同的含义;此类技术以引用的方式以其整体并入。如果发生冲突,将以本说明书(包括定义)为准。
具体实施方式
98.dna模板序列中包括共有基序tatctgtt的终止信号的偶然存在可导致sp6和t7 rna聚合酶的体外转录的过早终止,导致mrna转录物的异质群体,其中所需全长mrna转录物的产量显著降低。本发明人鉴定了共有终止信号tatctgtt的位置1、6或8的单点突变足以防止体外转录的过早终止。本发明人还发现,先前鉴定的共有基序tatctgtt的此类变体经常存在于用于在体外转录中使用的密码子优化的dna模板序列中。此外,以前的工作表明,紧靠共有基序tatctgtt的3’的富含t的序列是转录终止所必需的(kwon&kang 1999,the journal of biological chemistry,274:41,pp 29149-29155),但本发明人证实这不是终止信号的必需元件。本发明人的发现使得筛选终止信号并有效地将它们从此类dna模板序列中去除成为可能。
99.因此,在一方面,本发明涉及一种制备编码蛋白质的优化的dna序列作为体外转录的模板的方法,所述方法包括:(a)提供包含蛋白质编码序列的dna序列;(b)确定dna序列中终止信号的存在,其中终止信号具有如下核酸序列:5
’‑
x1atctx2tx
3-3’,其中x1、x2和x3独立
地选自a、c、t或g;以及(c)如果存在一个或多个终止信号,通过用其它三个核酸中的任何一个替换所述一个或多个终止信号的位置2、3、4、5和7中任何一个位置的一个或多个核酸来修饰dna序列,以产生优化的dna序列,其中,如果需要,选择一个或多个替换核酸以保留由蛋白质编码序列编码的蛋白质的氨基酸序列。
100.与t7 rna聚合酶相比,sp6 rna聚合酶合成的mrna具有显著减少的异常终止转录物(所谓的“短聚体”),因此特别适用于mrna的大规模体外合成(参见wo 2018/157153)。另外,本发明人在此证实,与t7rna聚合酶不同,sp6 rna聚合酶合成的mrna转录物不形成分子内或分子间双链体,并且因此基本上不含双链体mrna。
101.本发明人发现,当使用sp6 rna聚合酶或t7 rna聚合酶时,在体外合成期间发生mrna转录物的非模板延长(失控转录)。由于各种原因,mrna转录物末端存在失控序列可能是有问题的。例如,由于批次与批次之间的差异,这增加了所得mrna制备的异质性,因此使质量控制更加困难。失控还可能将与mrna加工和稳定性相关的不想要的元件引入mrna转录物中。另外,至少对于使用t7 rna聚合酶的体外转录过程,失控转录导致rna双链体的形成。为了改进通过体外合成产生mrna的现有方法,本发明提供了以下方法和dna序列,其中在dna模板的3’末端添加了一个或多个终止信号,以防止mrna转录物的非模板延长。本发明人惊奇地发现,一个或多个终止信号的添加可以很有效地终止rna聚合酶对相应修饰的dna模板的转录,以至于不再需要在体外转录之前线性化包含dna模板的质粒。去除线性化步骤(其通常涉及与限制性内切酶一起孵育)可导致mrna生产成本的显著节约,尤其是在大规模生产药物产品时。在wo 2017/009376和wo 2012/170443中,环状质粒被用作通过体外合成生产rna的模板。然而,dna模板序列包括编码自切割核酶的序列和编码多个终止信号的序列。本发明人首次证实,在mrna合成期间,通过仅添加终止子序列从环状dna模板进行体外转录,可以实现超过90%的终止效率。
102.因此,在另一个方面,本发明提供了制备编码蛋白质的优化的dna序列作为体外转录模板的方法,所述方法包括:(a)提供编码蛋白质的dna序列;以及(b)在dna序列的3’末端添加一个或多个终止信号以提供优化的dna序列,其中一个或多个终止信号包含以下核酸序列:5
’‑
x1atctx2tx
3-3’,其中x1、x2和x3独立地选自a、c、t或g。本发明还提供了用于在体外转录中使用的dna序列,以5’至3’顺序包含:(a)5’utr;(b)蛋白质编码序列;(c)3’utr;(d)任选的编码聚a尾的核酸序列;和(e)终止信号;其中终止信号包含以下核酸序列:5
’‑
x1atctx2tx
3-3’,其中x1、x2和x3独立地选自a、c、t或g。此外,本发明提供了包含dna序列的核酸载体,通常可操作地连接到rna聚合酶启动子,以及这些核酸载体在mrna生产方法中的用途,其中rna聚合酶将dna序列转录成mrna转录物。
103.在以下部分中详细描述了本发明的各个方面。部分的使用并不意在限制本发明。每个部分可以适用于本发明的任何方面。
104.dna模板
105.本发明中可以使用各种核酸模板。通常,可以使用带有双链sp6启动子序列的完全双链或大部分单链的dna模板。
106.在一些实施方案中,将合成的优化的dna序列插入用于在体外转录中使用的核酸载体。在一些实施方案中,核酸载体是质粒。术语“质粒”或“质粒核酸载体”是指环状核酸分子,优选指人工核酸分子。本发明上下文中的质粒dna适用于掺入或包含所需的核酸序列,
诸如包含编码rna的序列和/或编码至少一种蛋白质、多肽或肽的开放阅读框的核酸序列。此类质粒dna构建体/载体可以是表达载体、克隆载体、转移载体等。质粒dna通常包含对应于(编码)所需mrna转录物的序列或其部分,诸如对应于mrna的开放阅读框和5
’‑
和/或3’utr的序列。在一些实施方案中,对应于所需mrna转录物的序列也可以在3’utr之后编码聚a酸尾,使得多聚腺苷酸尾包括在mrna转录物中。更典型地,在本发明的上下文中,对应于所需mrna转录物的序列由5’/3’utr和开放阅读框组成。在本发明的后一个实施方案中,在体外转录期间由dna质粒合成的mrna转录物不含聚a尾,并且需要mrna转录物的合成后加工以添加聚a尾。
107.表达载体可用于在称为rna体外转录的过程中产生表达产物,诸如rna,例如mrna。例如,表达载体可以包含载体序列段的rna体外转录所需的序列,诸如启动子序列,例如,rna聚合酶启动子序列,例如t3、t7或sp6 rna聚合酶启动子序列。
108.克隆载体通常是含有克隆位点的载体,克隆位点可用于将核酸序列掺入(插入)到载体中。克隆载体可以是例如质粒载体或噬菌体载体。转移载体可以是适用于将核酸分子转移到细胞或生物体中的载体,例如病毒载体。适用于本发明的质粒dna载体通常包含多克隆位点、rna聚合酶启动子序列、任选的选择标记物(诸如抗生素抗性因子)和适用于载体增殖的序列(诸如复制起点)。特别合适的是质粒dna载体或表达载体,其包含dna依赖性rna聚合酶诸如t3、t7和sp6的启动子。用于实施本发明的合适质粒包括,例如,puc19和pbr322。
109.线性化的质粒dna(经由一种或多种限制酶线性化)、线性化的基因组dna片段(通过限制酶和/或物理手段)、pcr产物和/或合成的dna寡核苷酸可以用作模板,用于与sp6/t7 rna聚合酶进行体外转录,条件是它们在待转录的dna序列的上游(并且以正确的方向)含有双链sp6启动子,或者与t7 rna聚合酶行体外转录,条件是它们在待转录的dna序列的上游(并且以正确的方向)含有双链t7启动子。
110.在一些实施方案中,线性化dna模板具有平端。
111.在本发明的一个特定实施方案中,对于体外转录不需要线性化质粒dna。具体地,本发明首次使得使用sp6/t7 rna聚合酶从环状核酸载体诸如质粒dna(通常是超螺旋的)产生mrna转录物用于体外转录成为可能。
112.在一些实施方案中,dna模板包括5'和/或3'非翻译区。在一些实施方案中,5’非翻译区包括一个或多个影响mrna的稳定性或翻译的元件,例如铁应答元件。在一些实施方案中,5’非翻译区的长度可以在约50和500个核苷酸之间。
113.在一些实施方案中,3’非翻译区包括一个或多个聚腺苷酸化信号,影响mrna在细胞中定位稳定性的蛋白质结合位点,或一个或多个mirna结合位点。在一些实施方案中,3’非翻译区的长度可以在50和500个核苷酸之间或更长。
114.示例性的3’utr序列和/或5’utr序列可以源自稳定的mrna分子(例如,球蛋白、肌动蛋白、gapdh、微管蛋白、组蛋白和柠檬酸循环酶),以增加有义mrna分子的稳定性。例如,5’utr序列可以包括cmv立即早期1(ie1)基因的部分序列或其片段,以提高核酸酶抗性和/或提高多核苷酸的半衰期。还考虑了在多核苷酸(例如,mrna)的3’末端或非翻译区包含编码人类生长激素(hgh)或其片段的序列,以进一步稳定该多核苷酸。通常,这些修饰相对于其未修饰的对应物改善了多核苷酸的稳定性和/或药代动力学性质(例如半衰期),并且包括例如为改善此类多核苷酸对体内核酸酶消化的抗性而进行的修饰。
115.序列优化
116.本发明的一个方面涉及去除dna模板中的终止子序列以制备优化的dna序列。该方法包括尤其是确定dna序列中终止信号的存在的步骤,以及如果存在一个或多个终止信号,则通过将所述一种或多种终止信号的位置2、3、4、5和7中任何一个位置的一个或多个核酸用其它三个核酸中的任何一个替换来修饰dna序列,以产生优化的dna序列,其中,如果需要,选择一个或多个替换核酸以保留由蛋白质编码序列编码的蛋白质的氨基酸序列。终止信号可以在dna序列的任何地方检测到(例如在编码蛋白质编码序列的区域内,在编码5’非翻译区的区域内和/或在编码3’非翻译区的区域内)。上述步骤可以由计算机执行。适用于检测dna序列中特定核酸序列(例如,本发明的终止信号)的存在并且鉴定保留由蛋白质编码序列编码的蛋白质的氨基酸序列的核酸取代的计算机程序是本领域众所周知的。
117.待转录的dna序列可以进一步优化,以促进更有效的转录和/或翻译。例如,可以针对顺式调节元件(例如tata盒、终止信号和蛋白质结合位点)、人工重组位点、chi位点、cpg二核苷酸含量、阴性cpg岛、gc含量、聚合酶滑脱位点和/或其他与转录相关的元件优化dna序列;可以针对隐蔽剪接位点、mrna二级结构、mrna的稳定自由能、重复序列、rna不稳定性基序和/或与mrna加工和稳定性相关的其他元件优化dna序列;可以针对密码子使用偏好、密码子适应性、内部chi位点、核糖体结合位点(例如ires)、过早终止聚a位点、shine-dalgarno(sd)序列和/或与翻译相关的其他元件优化dna序列;和/或可以针对密密码子背景、密码子-反密码子相互作用、翻译暂停位点和/或其他与蛋白质折叠相关的元件方面进行优化dna序列。本领域已知的优化方法可用于本发明,例如geneoptimizer by thermofisher and optimumgene
tm
,其在us 20110081708中有所描述,其内容以引用的方式整体并入本文。
118.在一些实施方案中,使用密码子优化算法来修饰dna序列,以促进更有效的转录和/或翻译。在一些实施方案中,密码子优化算法确定dna序列中终止信号的存在。在一些实施方案中,密码子优化算法通过替换一个或多个核酸来修饰dna序列,并且如果需要,可以选择一个或多个替换核酸来保留由蛋白质编码序列编码的蛋白质的氨基酸序列。
119.在一个特定实施方案中,密码子优化算法确定dna序列中终止信号的存在,其中终止信号具有以下核酸序列:5
’‑
x1atctx2tx
3-3’,其中x1、x2和x3独立地选自a、c、t或g;以及如果存在一个或多个终止信号,通过用其它三个核酸中的任何一个替换所述一个或多个终止信号的位置2、3、4、5和7中任何一个位置的一个或多个核酸来修饰dna序列,以产生优化的dna序列,其中,如果需要,选择一个或多个替换核酸以保留由蛋白质编码序列编码的蛋白质的氨基酸序列。
120.密码子优化算法通过最大化密码子适应指数(cai)产生序列。cai是密码子使用偏好的数值分数,用于测量序列与参考基因集的偏差。在一些实施方案中,参考组的基因是哺乳动物基因。在一个特定实施方案中,参照组的基因是人类基因。cai通常是根据感兴趣的蛋白质密码子序列中所有密码子的使用频率的偏好来计算的。在第一步中,密码子优化算法反复修饰输入蛋白质密码子序列以获得具有最佳cai的第一输出序列。在第二步中,分析第一输出序列中已知在转录或翻译水平对基因表达有负面影响的序列元件的存在。这包括本文所述的终止信号。如果鉴定了此类序列元件,密码子优化算法修饰第一输出序列以去除它们,从而产生第二输出序列。在同一个或随后的步骤中,还分析第一或第二输出序列的
一个或多个下列参数:gc含量、编码的mrna转录物的稳定自由能以及框外起始密码子的存在。如果需要,修饰第一或第二输出序列以优化这些参数中的一个或多个。例如,任何框外起始密码子可以通过适当的密码子取代来去除。具有较低gc含量的输出序列通常比具有较高gc含量的输出序列具有更负的自由能值。自由能的最负值被认为导致最结构化,因此得到最稳定的mrna转录物。因此,在一些实施方案中,该算法通过进一步的密码子取代来增加第一或第二输出序列的gc含量。
121.终止信号的靶向插入
122.本发明的另一方面涉及在编码感兴趣的蛋白质(例如治疗性蛋白质)的dna序列的3’末端包含一个或多个终止信号,以制备优化的dna序列作为体外转录的模板。在编码mrna转录物的dna序列的3’末端靶向插入一个或多个终止信号(例如两个或三个终止信号)可以避免在体外转录之前对编码模板的质粒进行线性化的需要。因此,在一方面,本发明涉及用于在体外转录中使用的dna序列,以5’至3’顺序包含:
[0123]-5’utr;
[0124]-蛋白质编码序列;
[0125]-3’utr;
[0126]-任选的编码聚a尾的核酸序列;和
[0127]-终止信号。
[0128]
根据本发明,终止信号包含以下核酸序列:5
’‑
x1atctx2tx
3-3’,其中x1、x2和x3独立地选自a、c、t或g。在一个实施方案中,终止信号包含核酸序列5
’‑
x1atctgtt-3’。x1可以是t或c。合适的末端可以选自5
’‑
ttttatctgttttttt-3’、5
’‑
ttttatctgttttttttt-3’、5
’‑
cgttttatctgttttttt-3’、5
’‑
cgttccatctgttttttt-3’、5
’‑
cgttttatctgtttgttt-3’、5
’‑
cgttttatctgtttgttt-3’或5
’‑
cgttttatctgttgtttt-3’。
[0129]
通常,dna序列包含一个以上的终止信号,例如两个或更多个、三个或更多个、或四个或更多个终止信号。发明人已经表明,为了发生有效的终止,终止信号可以相隔10个碱基对或更少个碱基对,例如相隔5-10个碱基对。在一些实施方案中,dna序列在长度为30个核苷酸的核苷酸序列内包含两个终止信号(例如5
’‑
x1atctx2tx
3-3’,其中x1、x2和x3独立地选自a、c、t或g)。因此,在一些实施方案中,用于本发明的dna序列在其3’末端包含以下序列:5
’‑
x1atctx2tx
3-(zn)-x4atctx5tx
6-3’,其中x1、x2、x3、x4、x5和x6独立地选自a、c、t或g并且zn代表n个核苷酸的间隔序列,每个核苷酸独立地选自a、c、t或g,并且其中n为10或更小。例如,n可以是5、6、7、8、9或10。z可以是t。在一些实施方案中,dna序列包含以下序列:ttttatctgtttttttttttttatctgttttttttt(seq id no:12)。在其他实施方案中,dna序列在长度为50个核苷酸的核苷酸序列内包含三个终止信号(例如5
’‑
x1atctx2tx
3-3’,其中x1、x2和x3独立地选自a、c、t或g)。因此,在一些实施方案中,用于本发明的dna序列在其3’末端包含以下序列:5
’‑
x1atctx2tx
3-(zn)-x4atctx5tx
6-(zm)-x7atctx8tx
9-3’,其中x1、x2、x3、x4、x5、x6、x7、x8和x9独立地选自a、c、t或g,zn代表n个核苷酸的间隔序列,并且zm代表m个核苷酸的间隔序列,每个核苷酸独立地选自a、c、t或g,并且其中n和/或m独立地为10或更小。例如,n可以是5、6、7、8、9或10。m可以是5、6、7、8、9或10。z可以是t。在一个具体实施方案中,dna序列在其3’末端包含以下序列:ttttatctgtttttttttttttatctgtttttttttttttatctgttttttttt(seq id no:13)。
[0130]
如本文所示,在dna序列的3’末端依次具有两个终止信号可导致体外转录的有效终止。本技术的实施例进一步证实,当在编码mrna转录物的dna序列的3’末端存在两个以上拷贝的终止信号时,可以达到接近100%的正确终止的mrna转录物的产率。具体而言,在dna序列的3’末端依次添加三个或更多个终止信号可以产生100%的终止。当在37℃下进行体外转录时,已经进行了这种观察。
[0131]
此外,本发明人已经表明,序列中间隔10个核苷酸(例如t)或更少的两个或三个(或更多个)终止子信号(例如tatctgtt)的最小终止序列足以在dna序列的末端有效终止体外转录。因此,用于本发明的dna序列不包含任何其他终止信号和/或序列。具有本发明最小终止序列的dna序列可以产生正确终止的mrna转录物,而不需要3’末端的核酶序列或替代性的终止信号。因此,在一些实施方案中,dna序列在其3’末端不包含编码核酶的其他序列。另外或可替代地,dna序列不包含i类终止信号。事实上,除了本文公开的最小终止子序列之外,不需要其它终止信号来实现体外转录的终止。
[0132]
根据本发明,终止信号不存在于dna序列的5’utr、蛋白质编码序列和3’utr,以避免在rna聚合酶到达dna序列的3’末端之前体外转录的过早终止。
[0133]
本文还提供了制备前述段落中描述的dna序列的方法。该方法包括:(a)提供编码蛋白质的dna序列;以及(b)在dna序列的3’末端添加一个或多个终止信号以提供dna序列,其中一个或多个终止信号包含以下核酸序列:5
’‑
x1atctx2tx
3-3’(seq id no:1),其中x1、x2和x3独立地选自a、c、t或g。在一些实施方案中,添加在dna序列3’末端的终止信号包含以下序列:ttttatctgtttttttttt(seq id no:14)。
[0134]
本技术的实施例证实,对于线性和超螺旋dna模板,添加两个或更多个终止信号导致在体外转录期间mrna转录物的不期望的延长减少。因此,在一些实施方案中,将两个或多个、三个或更多个、四个或更多个终止信号被添加至dna序列的3’末端。在一些实施方案中,添加到3’末端的终止序列包含以下或由以下组成:长度为30个核苷酸的核苷酸序列内的两个终止信号(例如5
’‑
x1atctx2tx
3-3’,其中x1、x2和x3独立地选自a、c、t或g)。在一些实施方案中,添加到3’末端的终止序列包含以下序列或由以下序列组成:5
’‑
x1atctx2tx
3-(zn)-x4atctx5tx
6-3’,其中x1、x2、x3、x4、x5和x6独立地选自a、c、t或g,zn代表n个核苷酸的间隔序列,每个核苷酸独立地选自a、c、t或g,并且其中n为10或更小。在一些实施方案中,终止序列包含以下序列或由以下序列组成:ttttatctgtttttttttttttatctgttttttttt(seq id no:12)。
[0135]
在一些实施方案中,添加到3’末端的终止序列包含以下或由以下组成:长度为50个核苷酸的核苷酸序列内的三个终止信号(例如5
’‑
x1atctx2tx
3-3’,其中x1、x2和x3独立地选自a、c、t或g)。在一些实施方案中,将三个终止信号添加至dna序列的3’末端。在一些实施方案中,添加到3’末端的终止序列包含以下序列或由以下序列组成:5
’‑
x1atctx2tx
3-(zn)-x4atctx5tx
6-(zm)-x7atctx8tx
9-3’,其中x1、x2、x3、x4、x5、x6、x7、x8和x9独立地选自a、c、t或g,zn代表n个核苷酸的间隔序列,并且zm代表m个核苷酸的间隔序列,每个核苷酸独立地选自a、c、t或g,并且其中n和/或m独立地为10或更小。在一些实施方案中,终止序列包含以下序列或由以下序列组成:ttttatctgtttttttttttttatctgtttttttttttttatctgttttttttt(seq id no:13)。
[0136]
sp6 rna聚合酶
[0137]
sp6 rna聚合酶是对sp6启动子序列具有高度序列特异性的dna依赖性rna聚合酶。通常,这种聚合酶催化其启动子下游的单链dna或双链dna上rna的5
′→3′
体外合成;它将天然核糖核苷酸和/或修饰的核糖核苷酸掺入聚合的转录物中。
[0138]
噬菌体sp6 rna聚合酶的序列最初被描述(genbank:y00105.1)为具有以下氨基酸序列:
[0139]
mqdlhaiqlqleeemfnggirrfeadqqrqiaagsesdtawnrrllseliapmaegiqaykeeyegkkgrapralaflqcvenevaayitmkvvmdmlntdatlqaiamsvaeriedqvrfskleghaakyfekvkkslkasrtksyrhahnvavvaeksvaekdadfdrweawpketqlqigttlleilegsvfyngepvfmramrtyggktiyylqtsesvgqwisafkehvaqlspayapcvipprpwrtpfnggfhtekvasrirlvkgnrehvrkltqkqmpkvykainalqntqwqinkdvlavieevirldlgygvpsfkplidkenkpanpvpvefqhlrgrelkemlspeqwqqfinwkgecarlytaetkrgsksaavvrmvgqarkysafesiyfvyamdsrsrvyvqsstlspqsndlgkallrftegrpvngvealkwfcinganlwgwdkktfdvrvsnvldeefqdmcrdiaadpltftqwakadapyeflawcfeyaqyldlvdegradefrthlpvhqdgscsgiqhysamlrdevgakavnlkpsdapqdiygavaqvvikknalymdaddattftsgsvtlsgtelramasawdsigitrsltkkpvmtlpygstrltcresvidyivdleekeaqkavaegrtankvhpfeddrqdyltpgaaynymtaliwpsisevvkapivamkmirqlarfaakrneglmytlptgfileqkimatemlrvrtclmgdikmslqvetdivdeaammgaaapnfvhghdashliltvcelvdkgvtsiavihdsfgthadntltlrvalkgqmvamyidgnalqklleehevrwmvdtgievpeqgefdlneimdseyvfa(seq id no:15)。
[0140]
适用于本发明的sp6 rna聚合酶可以是具有与噬菌体sp6 rna聚合酶基本上相同的聚合酶活性的任何酶。因此,在一些实施方案中,适用于本发明的sp6 rna聚合酶可以从seq id no:15.例如,合适的sp6 rna聚合酶可以包含一个或多个氨基酸置换、缺失或添加。在一些实施方案中,合适的sp6 rna聚合酶具有与seq id no:15约99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、89%、88%、87%、86%、85%、84%、83%、82%、81%、80%、75%、70%、65%或60%一致或同源的氨基酸序列。在一些实施方案中,合适的sp6 rna聚合酶可以是(从n-末端、c-末端或内部)截短的蛋白质,但保留聚合酶活性。在一些实施方案中,合适的sp6 rna聚合酶是融合蛋白。
[0141]
在一些实施方案中,sp6 rna聚合酶由具有以下核苷酸序列的基因编码:atgcaagatttacacgctatccagcttcaattagaagaagagatgtttaatggtggcattcgtcgcttcgaagcagatcaacaacgccagattgcagcaggtagcgagagcgacacagcatggaaccgccgcctgttgtcagaacttattgcacctatggctgaaggcattcaggcttataaagaagagtacgaaggtaagaaaggtcgtgcacctcgcgcattggctttcttacaatgtgtagaaaatgaagttgcagcatacatcactatgaaagttgttatggatatgctgaatacggatgctacccttcaggctattgcaatgagtgtagcagaacgcattgaagaccaagtgcgcttttctaagctagaaggtcacgccgctaaatactttgagaaggttaagaagtcactcaaggctagccgtactaagtcatatcgtcacgctcataacgtagctgtagttgctgaaaaatcagttgcagaaaaggacgcggactttgaccgttgggaggcgtggccaaaagaaactcaattgcagattggtactaccttgcttgaaatcttagaaggtagcgttttctataatggtgaacctgtatttatgcgtgctatgcgcacttatggcggaaagactatttactacttacaaacttctgaaagtgtaggccagtggattagcgcattcaaagagcacgtagcgcaattaagcccagcttatgccccttgcgtaatccctcctcgtccttggagaactccatttaatggagggttccatactgagaaggtagctagccgtatccgtcttgtaaaaggtaaccgtgagcatgtacgcaagttgactcaaaagcaaatgccaaaggtttataaggctatcaacgcattacaaaatacacaatggcaaatcaacaaggatgtattagcagttattgaagaagtaatccgcttagaccttggttatggtgtaccttccttcaagccactgattgacaa
ggagaacaagccagctaacccggtacctgttgaattccaacacctgcgcggtcgtgaactgaaagagatgctatcacctgagcagtggcaacaattcattaactggaaaggcgaatgcgcgcgcctatataccgcagaaactaagcgcggttcaaagtccgccgccgttgttcgcatggtaggacaggcccgtaaatatagcgcctttgaatccatttacttcgtgtacgcaatggatagccgcagccgtgtctatgtgcaatctagcacgctctctccgcagtctaacgacttaggtaaggcattactccgctttaccgagggacgccctgtgaatggcgtagaagcgcttaaatggttctgcatcaatggtgctaacctttggggatgggacaagaaaacttttgatgtgcgcgtgtctaacgtattagatgaggaattccaagatatgtgtcgagacatcgccgcagaccctctcacattcacccaatgggctaaagctgatgcaccttatgaattcctcgcttggtgctttgagtatgctcaataccttgatttggtggatgaaggaagggccgacgaattccgcactcacctaccagtacatcaggacgggtcttgttcaggcattcagcactatagtgctatgcttcgcgacgaagtaggggccaaagctgttaacctgaaaccctccgatgcaccgcaggatatctatggggcggtggcgcaagtggttatcaagaagaatgcgctatatatggatgcggacgatgcaaccacgtttacttctggtagcgtcacgctgtccggtacagaactgcgagcaatggctagcgcatgggatagtattggtattacccgtagcttaaccaaaaagcccgtgatgaccttgccatatggttctactcgcttaacttgccgtgaatctgtgattgattacatcgtagacttagaggaaaaagaggcgcagaaggcagtagcagaagggcggacggcaaacaaggtacatccttttgaagacgatcgtcaagattacttgactccgggcgcagcttacaactacatgacggcactaatctggccttctatttctgaagtagttaaggcaccgatagtagctatgaagatgatacgccagcttgcacgctttgcagcgaaacgtaatgaaggcctgatgtacaccctgcctactggcttcatcttagaacagaagatcatggcaaccgagatgctacgcgtgcgtacctgtctgatgggtgatatcaagatgtcccttcaggttgaaacggatatcgtagatgaagccgctatgatgggagcagcagcacctaatttcgtacacggtcatgacgcaagtcaccttatccttaccgtatgtgaattggtagacaagggcgtaactagtatcgctgtaatccacgactcttttggtactcatgcagacaacaccctcactcttagagtggcacttaaagggcagatggttgcaatgtatattgatggtaatgcgcttcagaaactactggaggagcatgaagtgcgctggatggttgatacaggtatcgaagtacctgagcaaggggagttcgaccttaacgaaatcatggattctgaatacgtatttgcctaa(seq id no:16)。
[0142]
适用于本发明的编码sp6 rna聚合酶的合适基因可以与seq id no:16具有约99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、89%、88%、87%、86%、85%、84%、83%、82%、81%或80%的相同或同源。
[0143]
适用于本发明的sp6 rna聚合酶可以是可商购产品,例如来自ambion、new england biolabs(neb)、promega和roche。sp6可以根据本文所述的seq id no:15或seq id no:15的变体的氨基酸序列从商业或非商业来源订购和/或定制设计。sp6 rna聚合酶可以是标准保真聚合酶,或者可以是高保真/高效/高容量的聚合酶,其已经被修饰以促进rna聚合酶活性,例如,sp6 rna聚合酶基因中的突变或sp6 rna聚合酶本身的翻译后修饰。此类经修饰的sp6的实例包括来自ambion的sp6 rna聚合酶-plus
tm
、来自neb的hiscribe sp6、和来自promega的ribomax
tm
和系统。
[0144]
在一些实施方案中,该sp6 rna聚合酶是热稳定的。在一个特定实施方案中,用于本发明的sp6 rna聚合酶的氨基酸序列含有相对于野生型sp6聚合酶的一个或多个突变,这使得该酶在37℃至56℃的温度范围内具有活性。在一些实施方案中,用于本发明的sp6 rna聚合酶在50℃-52℃的最佳温度下发挥作用。在其它实施方案中,用于本发明的sp6rna聚合酶在50℃下具有至少60分钟的半衰期。例如,用于本发明的特别合适的sp6 rna聚合酶在50℃下具有60分钟至120分钟(例如70分钟至100分钟或80分钟至90分钟)的半衰期。
[0145]
在一些实施方案中,合适的sp6 rna聚合酶是融合蛋白。例如,sp6rna聚合酶可以
包括一个或多个促进酶的分离、纯化或溶解度的标签。合适的标签可以位于n-末端、c-末端和/或内部。合适标签的非限制性实例包括钙调素结合蛋白(cbp);肝片吸虫8-kda抗原(fh8);flag标签肽;谷胱甘肽-s-转移酶(gst);组氨酸标签(例如六组氨酸标签(his6));麦芽糖结合蛋白(mbp);n利用质a(nusa);小泛素相关修饰物(sumo)融合标签;链霉亲和素结合肽(strep);串联亲和纯化(tap);和硫氧还蛋白(trxa)。在本发明中可以使用其他标签。这些和其他融合标签已有描述,例如costa等人frontiers in microbiology 5(2014):63和pct/us16/57044,它们的内容以引用的方式整体并入本文。在一些实施方案中,his标签位于sp6的n末端。
[0146]
sp6启动子
[0147]
任何可以被sp6 rna聚合酶识别的启动子都可以用于本发明。通常,sp6启动子包含5
′
atttaggtgacactatag-3
′
(seq id no:17)。已发现和/或创建了sp6启动子的变体以优化sp6与其启动子的识别和/或结合。非限制性变体包括但不限于:5
′‑
atttaggggacactatagaagag-3
′
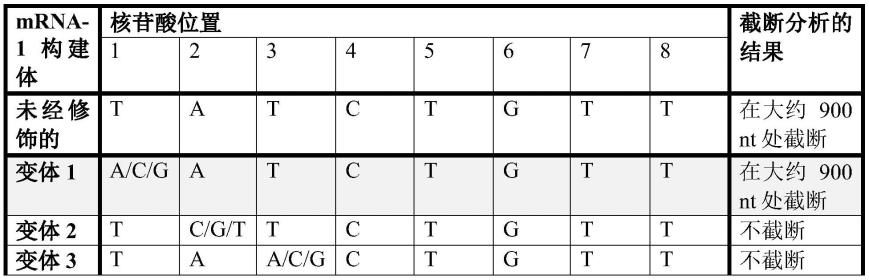
;5
′‑
atttaggggacactatagaagg-3
′
;5
′‑
atttaggggacactatagaaggg-3
′
;5
′‑
atttaggtgacactatagaa-3
′
;5
′‑
atttaggtgacactatagaaga-3
′
;5
′‑
atttaggtgacactatagaagag-3
′
;5
′‑
atttaggtgacactatagaagg-3
′
;5
′‑
atttaggtgacactatagaaggg-3
′
;5
′‑
atttaggtgacactatagaagng-3
′
;和5
′‑
catacgatttaggtgacactatag-3
′
(seq id no:18至seq id no:27)。
[0148]
另外,适用于本发明的sp6启动子可以与seq id no:18至seq id no:27中的任一者具有约95%、90%、85%、80%m、75%或70%的一致或同源。此外,适用于本发明的sp6启动子可以包括本文所述的任何启动子序列的一个或多个另外的核苷酸5’和/或3’。
[0149]
t7 rna聚合酶
[0150]
t7 rna聚合酶是对t7启动子序列具有高度序列特异性的dna依赖性rna聚合酶。通常,这种聚合酶催化其启动子下游的单链dna或双链dna上rna的5
′→3′
体外合成;它将天然核糖核苷酸和/或修饰的核糖核苷酸掺入聚合的转录物中。
[0151]
在一些实施方案中,该t7 rna聚合酶是热稳定的。在一个特定实施方案中,用于本发明的t7 rna聚合酶的氨基酸序列含有相对于野生型t7聚合酶的一个或多个突变,这使得该酶在37℃至56℃的温度范围内具有活性。合适的rna聚合酶的一个实例是来自neb的rna聚合酶。在一些实施方案中,用于本发明的t7 rna聚合酶在50℃-52℃的最佳温度下发挥作用。在其它实施方案中,用于本发明的t7 rna聚合酶在50℃下具有至少60分钟的半衰期。例如,用于本发明的特别合适的t7 rna聚合酶在50℃下具有60分钟至120分钟(例如70分钟至100分钟或80分钟至90分钟)的半衰期。
[0152]
t7启动子
[0153]
任何可以被t7 rna聚合酶识别的启动子都可以用于本发明。通常,t7启动子包含5
′‑
taatacgactcactatag-3
′
(seq id no:28)。
[0154]
mrna合成
[0155]
可以根据多种已知方法中的任一种合成根据本发明的mrna。各种方法描述于已公布的美国申请第us 2018/0258423号中,并且可用于实践本发明,所有这些方法通过引用并入本文。例如,可以经由体外转录(ivt)来合成根据本发明的mrna。简而言之,ivt通常使用线性或环状dna模板进行,所述模板包含启动子,三磷酸核糖核苷酸库,可能包含dtt和镁离
子的缓冲系统以及适当的rna聚合酶(例如,t3、t7或sp6 rna聚合酶),dna酶i,焦磷酸酶和/或rna酶抑制剂。确切条件将根据具体应用而改变。
[0156]
在一些实施方案中,合适的模板序列是编码蛋白质、多肽或肽的dna序列。在一些实施方案中,针对在人细胞中的有效表达的合适的模板序列进行密码子优化。密码子优化通常包括修饰编码肽、多肽或蛋白质的天然存在的或野生型核酸序列,以达到尽可能高的g/c含量,以调节密码子使用以避免稀有或限速密码子,以去除不稳定的核酸序列或基序和/或以消除暂停位点或终止子序列,而不改变mrna编码的肽、多肽或蛋白质的氨基酸序列。在一些实施方案中,合适的蛋白质编码序列是天然存在的或野生型序列。在一些实施方案中,合适的蛋白质编码序列编码在其氨基酸序列中含有一个或多个突变的蛋白质、多肽或肽。
[0157]
本文公开的方法可用于mrna的大规模生产。在一些实施方案中,根据本发明的方法在单批次中合成至少100mg、150mg、200mg、300mg、400mg、500mg、600mg、700mg、800mg、900mg、1g、5g、10g、25g、50g、75g、100g、250g、500g、750g、1kg、5kg、10kg、50kg、100kg、1000kg或更多mrna。在一些实施方案中,根据本发明的方法在单批次中合成至少1kg、10kg或100kg。如本文所用,术语“批次”是指一次合成的数量或量的mrna,例如,根据单一制造环境产生。批次可以指在一个反应中合成的mrna的量,该反应通过单等分的酶和/或单等分的dna模板发生,用于在一组条件下连续合成。在单批次中合成的mrna不包括在不同时间合成的mrna,它们组合起来以达到所需的量。通常,反应混合物包括rna聚合酶、dna模板和rna聚合酶反应缓冲液(其可包括核糖核苷酸或可能需要添加核糖核苷酸)。dna模板可以是线性的,尽管在本发明的上下文中更典型的是环形的。
[0158]
根据本发明,通常1-100mg的rna聚合酶用于产生约1克(g)mrna。在一些实施方案中,约1-90mg、1-80mg、1-60mg、1-50mg、1-40mg、10-100mg、10-80mg、10-60mg、10-50mg的rna聚合酶用于产生约1克mrna。在一些实施方案中,约5-20mg rna聚合酶用于产生约1克mrna。在一些实施方案中,约0.5至2克rna聚合酶用于产生约100克mrna。在一些实施方案中,约5至20克rna聚合酶用于约1千克mrna。在一些实施方案中,至少5mg rna聚合酶用于产生至少1克mrna。在一些实施方案中,至少500mg rna聚合酶用于产生至少100克mrna。在一些实施方案中,至少5克rna聚合酶用于产生至少1千克mrna。在一些实施方案中,约10mg、20mg、30mg、40mg、50mg、60mg、70mg、80mg、90mg或100mg质粒dna用于产生每克mrna。在一些实施方案中,约10-30mg质粒dna用于产生约1克mrna。在一些实施方案中,约1至3克质粒dna用于产生约100克mrna。在一些实施方案中,约10至30克质粒dna用于约1千克mrna。在一些实施方案中,至少10mg质粒dna用于产生至少1克mrna。在一些实施方案中,至少1克质粒dna用于产生至少100克mrna。在一些实施方案中,至少10克质粒dna用于产生至少1千克mrna。
[0159]
在一些实施方案中,反应混合物中rna聚合酶的浓度可以为约1至100nm、1至90nm、1至80nm、1至70nm、1至60nm、1至50nm、1至40nm、1至30nm、1至20nm或约1至10nm。在某些实施方案中,rna聚合酶的浓度为约10至50nm、20至50nm或30至50nm。可以使用浓度为100至10000单位/ml的rna聚合酶,可以使用例如浓度为100至9000单位/ml、100至8000单位/ml、100至7000单位/ml、100至6000单位/ml、100至5000单位/ml、100至1000单位/ml、200至2000单位/ml、500至1000单位/ml、500至2000单位/ml、500至3000单位/ml、500至4000单位/ml、500至5000单位/ml、500至6000单位/ml、1000至7500单位/ml和2500至5000单位/ml。
[0160]
反应混合物中每种核糖核苷酸(例如atp、utp、gtp和ctp)的浓度在约0.1mm和约10mm,例如,在约1mm至约10mm之间、在约2mm至约10mm之间、在约3mm至约10mm之间、在约1mm至约8mm之间、在约1mm至约6mm之间、在约3mm至约10mm之间、在约3mm至约8mm之间、在约3mm至约6mm之间、在约4mm至约5mm之间。在一些实施方案中,每种核糖核苷酸在反应混合物中为约5mm。在一些实施方案中,反应中使用的rntp(例如,atp、gtp、ctp和utp的组合)的总浓度范围在1mm和40mm之间。在一些实施方案中,反应中使用的rntp(例如,atp、gtp、ctp和utp的组合)的总浓度范围在1mm和30mm之间,或在1mm和28mm之间,或在1mm和25mm之间,或在1mm和20mm之间。在一些实施方案中,总rntp浓度小于30mm。在一些实施方案中,总rntp浓度小于25mm。在一些实施方案中,总rntp浓度小于20mm。在一些实施方案中,总rntp浓度小于15mm。在一些实施方案中,总rntp浓度小于10mm。
[0161]
在一个特定实施方案中,基于编码给定mrna转录物的核酸序列中每种核酸的频率来优化反应混合物中每种rntp的浓度。具体地,这种序列优化的反应混合物包含一定比例的四种rntp(例如atp、gtp、ctp和utp)的每一种,其对应于mrna转录物中这四种核酸(a、g、c和u)的比例。
[0162]
在一些实施方案中,在体外转录开始之前,将起始核苷酸添加到反应混合物中。起始核苷酸是对应于mrna转录物的第一核苷酸(+1位置)的核苷酸。可以特别添加起始核苷酸以增加rna聚合酶的起始速率。起始核苷酸可以是核苷单磷酸、核苷二磷酸、核苷三磷酸。起始核苷酸可以是单核苷酸、二核苷酸或三核苷酸。在mrna转录物的第一核苷酸是g的实施方案中,起始核苷酸通常是gtp或gmp。在一个具体实施方案中,起始核苷酸是帽类似物。cap类似物可以选自由以下组成的组:g[5']ppp[5']g、m7g[5']ppp[5']g、m
32,2,7
g[5']ppp[5']g、m
27,3
’‑og[5']ppp[5']g(3'-arca)、m
27,2
’‑ogpppg(2'-arca)、m
27,2
’‑
o gppspg d1(β-s-arca d1)和m
27,2
’‑ogppspg d2(β-s-arca d2)。
[0163]
在具体实施方案中,rna转录物的第一个核苷酸是g,起始核苷酸是g的帽类似物,并且相应的rntp是gtp。在此类实施方案中,与gtp相比,反应混合物中存在过量的cap类似物。在一些实施方案中,添加的cap类似物的初始浓度在约1mm至约20mm、约1mm至约17.5mm、约1mm至约15mm、约1mm至约12.5mm、约1mm至约10mm、约1mm至约7.5mm、约1mm至约5mm或约1mm至约2.5mm的范围内。
[0164]
更典型地,在本发明的上下文中,仅在mrna转录物已经合成后(例如在合成后加工步骤中)将帽结构诸如帽类似物添加到在体外转录期间获得的mrna转录物中。通常,在此类实施方案中,在添加帽结构之前,mrna转录物首先被纯化(例如,通过切向流过滤)。
[0165]
rna聚合酶反应缓冲液通常包括盐/缓冲剂,例如tris、hepes、硫酸铵、碳酸氢钠、柠檬酸钠、乙酸钠、磷酸钾、磷酸钠、氯化钠和氯化镁。
[0166]
反应混合物的ph可以在约6至8.5、6.5至8.0、7.0至7.5之间,并且在一些实施方案中,ph为7.5。
[0167]
将dna模板(例如如上所述的,其量/浓度足以提供所需量的rna)、rna聚合酶反应缓冲液和rna聚合酶混合以形成反应混合物。将反应混合物在约37℃和约56℃之间孵育30分钟至6小时,例如约60分钟至约90分钟。在一些实施方案中,孵育发生在大约37℃至约42℃。在其它实施方案中,孵育发生在约43℃至约56℃,例如约50℃至约52℃。如本文所证实,通过在编码感兴趣的mrna转录物的dna序列的末端包括一个或多个本文所述的终止信号,
并在约50℃至约52℃的温度下用包括dna序列的模板进行反应,可以显著增加在体外转录反应中获得的精确终止的mrna转录物的产量。
[0168]
在一些实施方案中,在约37℃至约42℃下,在合适的rna聚合酶反应缓冲液(最终反应混合物ph为约7.5)中的约5mm ntp、约0.05mg/ml rna聚合酶和约0.1mg/ml dna模板孵育60至90分钟。在其他实施方案中,在约50℃至约52℃下,在合适的rna聚合酶反应缓冲液(最终反应混合物ph为约7.5)中的约5mm ntp、约0.05mg/ml rna聚合酶和约0.1mg/ml dna模板孵育60至90分钟。
[0169]
在一些实施方案中,反应混合物含有双链dna模板,其具有rna聚合酶特异性启动子、rna聚合酶、rna酶抑制剂、焦磷酸酶、29mm ntp、10mm dtt和反应缓冲液(当稀释10倍时为800mm hepes、20mm亚精胺、250mm mgcl2,ph 7.7),并且其量足以(qs)达到不含rna酶的水的所需反应体积;然后将该反应混合物在37℃下孵育60分钟。然后通过添加dna酶i和dna酶i缓冲液(当在10x时为100mm tris-hcl、5mm mgcl2和25mm cacl
2 ph7.6)来淬灭聚合酶反应,以促进用于纯化的制剂中的双链dna模板的消化。该实施方案已显示足以产生100克的mrna。
[0170]
在一些实施方案中,反应混合物包括浓度为1-10mm的ntp、浓度为0.01-0.5mg/ml的dna模板和浓度为0.01-0.1mg/ml的rna聚合酶,例如反应混合物包含浓度为5mm的ntp、浓度为0.1mg/ml的dna模板和浓度为0.05mg/ml的rna聚合酶。
[0171]
核苷酸
[0172]
根据本发明,各种天然存在的或经修饰的核苷可用于产生mrna。在一些实施方案中,用天然核苷(即腺苷、鸟苷、胞苷、尿苷)合成根据本发明的mrna转录物。在其他实施方案中,根据本发明的mrna转录物用天然核苷(例如腺苷、鸟苷、胞苷、尿苷)和下列的一种或多种合成:核苷类似物(例如2-氨基腺苷、2-硫代胸苷、肌苷、吡咯并嘧啶、3-甲基腺苷、5-甲基胞苷、c-5丙炔基-胞苷、c-5丙炔基-尿苷、2-氨基腺苷、c5-溴尿苷、c5-氟尿苷、c5-碘尿苷、c5-丙炔基-尿苷、c5-丙炔基-胞苷、c5-甲基胞苷、2-氨基腺苷、7-脱氮腺苷、7-脱氮鸟苷、8-氧代腺苷、8-氧代鸟苷、o(6)-甲基鸟苷、假尿苷(例如n-1-甲基-假尿苷)、2-硫代尿苷和2-硫代胞苷);化学修饰的碱基;生物修饰的碱基(例如甲基化的碱基);插入的碱基;修饰的糖(例如2
’‑
氟核糖、核糖、2
’‑
脱氧核糖、阿拉伯糖和己糖);和/或修饰的磷酸基团(例如,硫代磷酸酯和5
′‑
n-亚磷酰胺键)。
[0173]
在一些实施方案中,mrna包含一种或多种非标准核苷酸残基。非标准核苷酸残基可以包括,例如,5-甲基胞苷(“5mc”)、假尿苷(“u”)和/或2-硫代尿苷(“2su”)。参见,例如,美国专利号8,278,036或wo2011012316关于此类残基及其掺入mrna的讨论。mrna可以是rna,其被定义为其中25%的u残基是2-硫代-尿苷且25%的c残基是5-甲基胞苷。关于使用rna的教导公开于美国专利公开us20120195936和国际公开wo2011012316中,这两个专利据此以引用的方式整体并入。非标准核苷酸残基的存在可使得mrna比具有相同序列但仅含有标准残基的对照mrna更稳定和/或免疫原性更小。在另外的实施方案中,mrna可以包含一个或多个选自以下的非标准核苷酸残基:异胞嘧啶、假异胞嘧啶、5-溴尿嘧啶、5-丙炔基尿嘧啶、6-氨基嘌呤、2-氨基嘌呤、肌苷、二氨基嘌呤和2-氯-6-氨基嘌呤胞嘧啶,以及这些修饰和其他核碱基修饰的组合。一些实施方案可以另外包括对呋喃糖环或核碱基的另外的修饰。另外的修饰可以包括,例如,糖修饰或取代(例如,2'-o-烷基修饰、锁核酸(lna)中的一
者或多者)。在一些实施方案中,rna可与另外的多核苷酸和/或肽多核苷酸(pna)复合或杂交。在糖修饰是2'-o-烷基修饰的一些实施方案中,这种修饰可以包括但不限于2'-脱氧-2'-氟修饰、2'-o-甲基修饰、2'-o-甲氧基乙基修饰和2'-脱氧修饰。在一些实施方案中,这些修饰中的任一者可以存在于0-100%的核苷酸中——例如,多于0%、1%、10%、25%、50%、75%、85%、90%、95%或100%的组成核苷酸单独或组合。
[0174]
合成的mrna
[0175]
本发明提供了高质量的体外合成的mrna。例如,本发明提供了合成mrna的一致性/同质性。具体而言,本发明的组合物包括多个基本上全长的mrna分子。例如,至少81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%的mrna分子是全长mrna分子。这种组合物被认为是“富含”全长mrna分子的。在一些实施方案中,根据本发明合成的mrna基本上是全长的。与通过现有技术方法(即包括使用根据本发明的优化的dna序列的方法)产生的组合物相比,本发明的组合物具有更高百分比的全长mrna分子。
[0176]
在本发明的一些实施方案中,制备组合物或批次时没有特异性去除非全长mrna分子的mrna分子的步骤(即异常终止转录物或终止转录物,或过早终止的转录物)。
[0177]
在一些实施方案中,通过本发明合成的mrna分子的长度大于500、600、700、800、900、1000、2000、3000、4000、5000、10,000或更多个核苷酸;在本发明中还包括具有介于两者之间的任何长度的mrna。
[0178]
合成后加工
[0179]
通常,可以在合成后添加5'帽和/或3'尾。帽的存在对于提供对存在于大多数真核细胞中的核酸酶的抗性很重要。“尾”的存在用于保护mrna免受核酸外切酶降解。
[0180]
通常按如下方式添加5’帽:首先,rna末端磷酸酶从5’核苷酸除去一个末端磷酸基团,剩下两个末端磷酸酯;然后通过鸟苷酸转移酶将三磷酸鸟苷(gtp)添加到末端磷酸酯中,产生5
’5’
5三磷酸酯键;并且然后用甲基转移酶将鸟嘌呤的7-氮甲基化。帽结构的实例包括但不限于tom7g(5’)ppp(5’)(2’omeg)、m7g(5’)ppp(5’)(2’omea)、m7(3’omeg)(5’)ppp(5’)(2’omeg)、m7(3’omeg)(5’)ppp(5’)(2’omea)、m7g(5')ppp(5'(a,g(5')ppp(5')a和g(5')ppp(5')g。在一个具体实施方案中,帽结构是m7g(5’)ppp(5’)(2’omeg)。另外的帽结构描述于2017年2月27日提交的公开的美国申请号us 2016/0032356和美国临时申请62/464,327中,所述申请以引用的方式并入本文。
[0181]
通常,尾结构包括poly(a)和/或poly(c)尾。mrna的3'末端的多聚-a或多聚-c尾通常包含分别地至少50个腺苷或胞嘧啶核苷酸、至少150个腺苷或胞嘧啶核苷酸、至少200个腺苷或胞嘧啶核苷酸、至少250个腺苷或胞嘧啶核苷酸、至少300个腺苷或胞嘧啶核苷酸、至少350个腺苷或胞嘧啶核苷酸、至少400个腺苷或胞嘧啶核苷酸、至少450个腺苷或胞嘧啶核苷酸、至少500个腺苷或胞嘧啶核苷酸、至少550个腺苷或胞嘧啶核苷酸、至少600个腺苷或胞嘧啶核苷酸、至少650个腺苷或胞嘧啶核苷酸、至少700个腺苷或胞嘧啶核苷酸、至少750个腺苷或胞嘧啶核苷酸、至少800个腺苷或胞嘧啶核苷酸、至少850个腺苷或胞嘧啶核苷酸、至少900个腺苷或胞嘧啶核苷酸、至少950个腺苷或胞嘧啶核苷酸、或至少1kb腺苷或胞嘧啶核苷酸。在一些实施方案中,多聚-a或多聚-c尾可分别为约10至800个腺苷或胞嘧啶核苷酸(例如约10至200个腺苷或胞嘧啶核苷酸、约10至300个腺苷或胞嘧啶核苷酸、约10至400个
腺苷或胞嘧啶核苷酸、约10至500个腺苷或胞嘧啶核苷酸、约10至550个腺苷或胞嘧啶核苷酸、约10至600个腺苷或胞嘧啶核苷酸、约50至600个腺苷或胞嘧啶核苷酸、约100至600个腺苷或胞嘧啶核苷酸、约150至600个腺苷或胞嘧啶核苷酸、约200至600个腺苷或胞嘧啶核苷酸、约250至600个腺苷或胞嘧啶核苷酸、约300至600个腺苷或胞嘧啶核苷酸、约350至600个腺苷或胞嘧啶核苷酸、约400至600个腺苷或胞嘧啶核苷酸、约450至600个腺苷或胞嘧啶核苷酸、约500至600个腺苷或胞嘧啶核苷酸、约10至150个腺苷或胞嘧啶核苷酸、约10至100个腺苷或胞嘧啶核苷酸、约20至70个腺苷或胞嘧啶核苷酸或约20至60个腺苷或胞嘧啶核苷酸)。在一些实施方案中,尾结构包括具有本文所述的各种长度的多聚(a)和多聚(c)尾的组合。在一些实施方案中,尾结构包括至少50%、55%、65%、70%、75%、80%、85%、90%、92%、94%、95%、96%、97%、98%或99%的腺苷核苷酸。在一些实施方案中,尾结构包括至少50%、55%、65%、70%、75%、80%、85%、90%、92%、94%、95%、96%、97%、98%或99%的胞嘧啶核苷酸。
[0182]
如本文所述,5'帽和/或3'尾的添加有助于检测体外合成过程中产生的异常终止转录物,因为没有加帽和/或加尾,那些过早无效的mrna转录物的大小可能太小而无法被检测到。因此,在一些实施方案中,在测试mrna的纯度(例如,存在于mrna中的异常终止转录物的水平)之前,将5'帽和/或3'尾添加至合成的mrna。在一些实施方案中,在如本文所述纯化mrna之前将5'帽和/或3'尾添加至合成的mrna。在其他实施方案中,在如本文所述纯化mrna之后将5'帽和/或3'尾添加至合成的mrna。
[0183]
mrna的纯化
[0184]
根据本发明合成的mrna无需进一步纯化即可使用。具体而言,根据本发明合成的mrna可以在不存在除去短聚体的步骤的情况下使用。在一些实施方案中,可以进一步纯化根据本发明合成的mrna。可以使用各种方法来纯化根据本发明合成的mrna。例如,可以使用离心、过滤和/或色谱法来进行mrna的纯化。在一些实施方案中,合成的mrna通过乙醇沉淀或过滤或色谱、或者凝胶纯化或任何其他合适的方式来纯化。在一些实施方案中,mrna通过hplc纯化。在一些实施方案中,mrna在本领域的技术人员熟知的标准苯酚:氯仿:异戊醇溶液中提取。在一些实施方案中,mrna使用切向流过滤来纯化。合适的纯化方法包括在us2016/0040154、us 2015/0376220、于2018年2月27日提交的题为“methods for purification of messenger rna”的pct申请pct/us18/19978和于2018年2月27日提交的题为“methods for purification of messenger rna”的pct申请pct/us18/19954、于2018年11月8日提交的美国临时申请号62/757,612以及于2019年8月26日提交的美国临时申请号62/891,781中描述的那些方法,这些申请的全部内容以引用的方式并入本文并可以用于实践本发明。
[0185]
在一些实施方案中,mrna在加帽和/或加尾之前纯化。在一些实施方案中,mrna在加帽之前纯化。在一些实施方案中,mrna在加尾之前纯化。在一些实施方案中,mrna在加帽和加尾之后纯化。在一些实施方案中,mrna在加帽和加尾之前和之后纯化。
[0186]
在一些实施方案中,mrna在加帽和加尾之前或之后或者在加帽和加尾之前和之后都通过离心纯化。
[0187]
在一些实施方案中,mrna在加帽和加尾之前或之后或者在加帽和加尾之前和之后都通过过滤纯化。
[0188]
在一些实施方案中,mrna在加帽和加尾之前或之后或者在加帽和加尾之前和之后都通过切向流过滤(tff)纯化。在一些实施方案中,mrna可以进行进一步纯化,包括透析、透滤和/或超滤。
[0189]
在一些实施方案中,mrna在加帽和加尾之前或之后或者在加帽和加尾之前和之后都通过色谱纯化。
[0190]
mrna的沉淀
[0191]
不纯制剂中的mrna,诸如体外合成反应混合物,可使用缓冲液和合适的条件沉淀,如2018年11月8日提交的美国临时申请号62/757,612或2019年8月26日提交的美国临时申请号62/891,781中所述,并可用于实践本发明,然后使用本领域已知的各种纯化方法。如本文所用,术语“沉淀”(或其任何语法等同物)是指溶液中不溶性物质(例如固体)的形成。当与mrna结合使用时,术语“沉淀”是指在液体中mrna的不溶或固体形式的形成。
[0192]
通常,mrna沉淀涉及变性条件。如本文所用,术语“变性条件”是指可以导致对mrna的天然确认的破坏的任何化学或物理条件。由于分子的天然构象通常是最水溶性的,因此破坏分子的二级和三级结构可能导致溶解度发生变化并且可能导致mrna从溶液中沉淀出来。
[0193]
例如,从不纯的制剂中沉淀mrna的合适方法涉及用变性剂处理不纯的制剂,使得mrna沉淀。适合用于本发明的示例性变性试剂包括但不限于氯化锂、氯化钠、氯化钾、氯化胍、硫氰酸胍、异硫氰酸胍、乙酸铵及其组合。合适的试剂可以以固体形式或以溶液形式提供。
[0194]
在一些实施方案中,在变性缓冲液中使用胍盐来沉淀mrna。作为非限制性实例,胍盐可以包括氯化胍、硫氰酸胍或异硫氰酸胍。硫氰酸胍(gcsn),也称为胍硫氰酸盐,可用于沉淀mrna。胍盐诸如硫氰酸胍盐的使用浓度可以高于通常用于变性反应的浓度,产生基本上不含蛋白质污染物的mrna。在一些实施方案中,适用于mrna沉淀的溶液含有浓度大于4m的胍硫氰酸盐。
[0195]
在一个典型的实施方案中,对含有mrna转录物的体外转录反应混合物和/或由加帽和/或加尾反应产生的混合物(包括加帽和加尾的mrna转录物)进行纯化,该纯化过程包括添加变性剂诸如胍盐(例如硫氰酸胍),然后添加沉淀剂(例如100%乙醇)以使mrna从溶液中沉淀出来。将得到的mrna悬浮液添加至切向流过滤(tff)柱。
[0196]
除了变性试剂之外,用于mrna沉淀的合适溶液还可以包含盐、表面活性剂和/或缓冲剂。例如,合适的溶液还可以包括月桂酰肌氨酸钠和/或柠檬酸钠。在一些实施方案中,适用于mrna沉淀的缓冲液包含约5mm柠檬酸钠。在一些实施方案中,适用于mrna沉淀的缓冲液包含约10mm柠檬酸钠。在一些实施方案中,适用于mrna沉淀的缓冲液包含约20mm柠檬酸钠。在一些实施方案中,适用于mrna沉淀的缓冲液包含约25mm柠檬酸钠。在一些实施方案中,适用于mrna沉淀的缓冲液包含约30mm柠檬酸钠。在一些实施方案中,适用于mrna沉淀的缓冲液包含约50mm柠檬酸钠。
[0197]
在一些实施方案中,适用于mrna沉淀的缓冲液包含表面活性剂,诸如n-月桂基肌氨酸(十二烷基肌氨酸钠)。在一些实施方案中,适用于mrna沉淀的缓冲液包含约0.01%n-月桂基肌氨酸。在一些实施方案中,适用于mrna沉淀的缓冲液包含约0.05%n-月桂基肌氨酸。在一些实施方案中,适用于mrna沉淀的缓冲液包含约0.1%n-月桂基肌氨酸。在一些实
施方案中,适用于mrna沉淀的缓冲液包含约0.5%n-月桂基肌氨酸。在一些实施方案中,适用于mrna沉淀的缓冲液包含1%n-月桂基肌氨酸。在一些实施方案中,适用于mrna沉淀的缓冲液包含约1.5%n-月桂基肌氨酸。在一些实施方案中,适用于mrna沉淀的缓冲液包含约2%、约2.5%或约5%n-月桂基肌氨酸。
[0198]
在一些实施方案中,用于mrna沉淀的合适溶液包含还原剂。在一些实施方案中,还原剂选自二硫苏糖醇(dtt)、β-巯基乙醇(b-me)、三(2-羧乙基)膦(tcep)、三(3-羟丙基)膦(thpp)、二硫赤藓糖醇(dte)和二硫丁胺(dtba)。在一些实施方案中,还原剂是二硫苏糖醇(dtt)。
[0199]
在一些实施方案中,dtt以大于1mm且高达约200mm的最终浓度存在。在一些实施方案中,dtt以2.5mm至100mm之间的最终浓度存在。在一些实施方案中,dtt以5mm至50mm之间的最终浓度存在。在一些实施方案中,dtt以约20mm的最终浓度存在。
[0200]
当存在或不存在还原剂时,即使在低浓度的变性剂下也可能发生蛋白质变性。高浓度gscn和高浓度dtt在变性溶液中的组合用于沉淀含有杂质的mrna,产生纯的且基本上不含蛋白质污染物的mrna。沉淀在缓冲液中的mrna可以通过过滤器进行处理。在一些实施方案中,在使用包含约5m gscn和约10mm dtt的缓冲液过滤后的单次沉淀之后的洗脱液具有高质量和纯度,没有可检测的蛋白质杂质。另外,该方法在大范围的mrna处理量下是可重复的,在约1克、或约10克、或约100克、或约500克、或约1000克或更多的mrna的范围内,而不会阻碍流体流过过滤器。
[0201]
在一些实施方案中,用于沉淀步骤的缓冲液进一步包含醇。在一些实施方案中,沉淀在以下条件下进行:mrna、变性缓冲液(包含gscn和还原剂,例如dtt)和醇(例如100%乙醇)以以下体积比存在:1:(5):(3)。在一些实施方案中,沉淀在以下条件下进行:mrna、变性缓冲液和醇(例如100%乙醇)以以下体积比存在:1:(3.5):(2.1)。在一些实施方案中,沉淀在以下条件下进行:mrna、变性缓冲液和醇(例如100%乙醇)以以下体积比存在:1:(4):(2)。在一些实施方案中,沉淀在以下条件下进行:mrna、变性缓冲液和醇(例如100%乙醇)以以下体积比存在:1:(2.8):(1.9)。在一些实施方案中,沉淀在以下条件下进行:mrna、变性缓冲液和醇(例如100%乙醇)以以下体积比存在:1:(2.3):(1.7)。在一些实施方案中,沉淀在以下条件下进行:mrna、变性缓冲液和醇(例如100%乙醇)以以下体积比存在:1:(2.1):(1.5)。
[0202]
在一些实施方案中,希望将不纯的制剂与本文所述的一种或多种变性试剂在允许大量mrna沉淀的所需温度下孵育一段时间。例如,可以将不纯的制剂和变性剂的混合物在室温或环境温度下孵育一段时间。在一些实施方案中,合适的孵育时间是等于或大于约2、3、4、5、6、7、8、9、10、15、20、25、30、40、50或60分钟的时段。在一些实施方案中,合适的孵育时间是等于或小于约60、55、50、45、40、35、30、25、20、15、10、9、8、7、6或5分钟的时段。在一些实施方案中,将混合物在室温下孵育约5分钟。通常,“室温”或“环境温度”是指范围为约20℃-25℃,例如约20℃、21℃、22℃、23℃、24℃或25℃的温度。在一些实施方案中,也可以将不纯的制剂和变性剂的混合物在高于室温(例如,在约30℃-37℃,或者特别是在约30℃、31℃、32℃、33℃、34℃、35℃、36℃或37℃)下或在低于室温(例如,约15℃-20℃,或者特别是在约15℃、16℃、17℃、18℃、19℃或20℃)下孵育。可以基于孵育温度来调节孵育时间。通常,较高的孵育温度需要较短的孵育时间。
[0203]
替代地或另外地,可以使用溶剂来促进mrna沉淀。合适的示例性溶剂包括但不限于异丙醇、丙酮、甲基乙酮、甲基异丁酮、乙醇、甲醇、地那铵(denatonium)及其组合。例如,可以将溶剂(例如,100%乙醇)与变性试剂一起或者在添加变性试剂和如本文所述的孵育之后添加到不纯的制剂中,以进一步增强和/或加速mrna沉淀。通常,在添加合适的溶剂(例如,100%乙醇)之后,可以将混合物在室温下再孵育一个时间段。通常,合适的孵育时间段为或大于约2、3、4、5、6、7、8、9、10、15、20、25、30、40、50或60分钟。在一些实施方案中,合适的孵育期为等于或小于约60、55、50、45、40、35、30、25、20、15、10、9、8、7、6或5分钟的时段。通常,将混合物在室温下孵育约5分钟。在适当调节孵育时间的情况下可以使用高于或低于室温的温度。替代地,可在4℃或-20℃下孵育沉淀。
[0204]
在一些实施方案中,纯化mrna的方法不含醇。因此,在一些实施方案中,悬浮液中沉淀mrna包含一种或多种两亲性聚合物来代替醇(例如,100%乙醇)。许多两亲性聚合物是本领域已知的。在一些实施方案中,两亲性聚合物包括普朗尼克、聚乙烯吡咯烷酮、聚乙烯醇、聚乙二醇(peg)或其组合。在一些实施方案中,两亲性聚合物选自以下中的一种或多种:peg三乙二醇、四乙二醇、peg 200、peg 300、peg 400、peg 600、peg 1,000、peg 1,500、peg 2,000、peg 3,000、peg 3,350、peg 4,000、peg 6,000、peg 8,000、peg 10,000、peg 20,000、peg 35,000和peg 40,000或其组合。
[0205]
在一些实施方案中,两亲性聚合物包含两种或更多种分子量peg聚合物的混合物被使用。例如,在一些实施方案中,两种、三种、四种、五种、六种、七种、八种、九种、十种、十一种或十二种分子量peg聚合物包含两亲性聚合物。因此,在一些实施方案中,peg溶液包含一种或多种peg聚合物的混合物。在一些实施方案中,peg聚合物的混合物包含具有不同分子量的聚合物。在一些实施方案中,悬浮液中沉淀mrna包含peg聚合物。本领域认识到各种类型的peg聚合物,其中一些具有不同的几何构型。peg聚合物包括例如具有直链构型、支链构型、y形构型或多臂构型的peg聚合物。在一些实施方案中,peg在包含一种或多种具有不同几何构型的peg的悬浮液中。在一些实施方案中,可以使用peg-6000沉淀mrna来实现沉淀mrna。在一些实施方案中,可以使用peg-400沉淀mrna来实现沉淀mrna。
[0206]
在其他实施方案中,纯化mrna的无醇方法包括用三甘醇(teg)沉淀mrna。在一些实施方案中,可以使用三乙二醇单甲醚(mteg)沉淀mrna来实现沉淀mrna。在一些实施方案中,可以使用叔丁基-teg-o-丙酸酯沉淀mrna来实现沉淀mrna。在一些实施方案中,可以使用teg-二甲基丙烯酸酯沉淀mrna来实现沉淀mrna。在一些实施方案中,可以使用teg-二甲基醚沉淀mrna来实现沉淀mrna。在一些实施方案中,可以使用teg-二乙烯基醚沉淀mrna来实现沉淀mrna。在一些实施方案中,可以使用teg-单丁基醚沉淀mrna来实现沉淀mrna。在一些实施方案中,可以使用teg-甲基醚甲基丙烯酸酯沉淀mrna来实现沉淀mrna。在一些实施方案中,可以使用teg-单癸基醚沉淀mrna来实现沉淀mrna。在一些实施方案中,可以使用teg-二苯甲酸酯沉淀mrna来实现沉淀mrna。这些基于peg或teg的试剂中的任何一种均可以与gscn组合使用来沉淀mrna。根据本发明产生的纯化mrna的示例性无乙醇方法使用gscn和mteg的组合来沉淀mrna。
[0207]
在一些实施方案中,在悬浮液中沉淀mrna包含peg聚合物,其中peg聚合物包含peg修饰的脂质。在一些实施方案中,peg修饰的脂质是1,2-二肉豆蔻酰基-sn-甘油、甲氧基聚乙二醇(dmg-peg-2k)。在一些实施方案中,peg修饰的脂质是dopa-peg缀合物。在一些实施
方案中,peg修饰的脂质是泊洛沙姆-peg缀合物。在一些实施方案中,peg修饰的脂质包含dotap。在一些实施方案中,peg修饰的脂质包含胆固醇。
[0208]
在一些实施方案中,在包含任何前述peg或teg试剂的悬浮液中沉淀mrna。在一些实施方案中,peg或teg在悬浮液中为约10%至约100%重量/体积浓度。例如,在一些实施方案中,peg或teg以约5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、100%的重量/体积浓度以及其间的任何值存在于悬浮液中。
[0209]
在一些实施方案中,在悬浮液中沉淀mrna包含约0.1至约5.0的peg或teg与总mrna悬浮液体积的体积/体积比。例如,在一些实施方案中,peg或teg以约0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9、1.0、1.25、1.5、1.75、2.0、2.25、2.5、2.75、3.0、3.25、3.5、3.75、4.0、4.25、4.5、4.75、5.0的体积/体积比存在于mrna悬浮液中。
[0210]
在一些实施方案中,用于mrna沉淀的反应体积包含(i)gscn和(ii)peg或teg。
[0211]
mrna的表征
[0212]
全长的、异常终止的和/或过早终止的mrna转录物可以使用本领域可用的任何方法进行检测和定量。在一些实施方案中,合成的mrna分子使用印迹、毛细管电泳、色谱、荧光、凝胶电泳、hplc、银染、光谱、紫外(uv)或uplc或者它们的组合来检测。本领域已知的其他检测方法包括在本发明中。在一些实施方案中,合成的mrna分子使用紫外吸收光谱来检测,通过毛细管电泳来分离。在一些实施方案中,在凝胶电泳(“乙二醛凝胶电泳”)之前,首先通过乙二醛染料使mrna变性。在一些实施方案中,在加帽或加尾之前表征合成的mrna。在一些实施方案中,在加帽和加尾之后表征合成的mrna。
[0213]
在一些实施方案中,除全长mrna以外,通过本文公开的方法产生的mrna包含小于10%、小于9%、小于8%、小于7%、小于6%、小于5%、小于4%、小于3%、小于2%、小于1%、小于0.5%、小于0.1%的杂质。杂质包括ivt污染物,例如蛋白质、酶、游离核苷酸和/或短聚体(shortmer)。
[0214]
在一些实施方案中,根据本发明产生的mrna基本上不含短聚体或异常终止转录物。具体而言,根据本发明产生的mrna包含通过毛细管电泳或乙二醛凝胶电泳检测不到水平的短聚体或异常终止转录物。如本文所用,术语“短聚体”或“异常终止转录物”是指小于全长的任何转录物。在一些实施方案中,“短聚体”或“异常终止转录物”为小于100个核苷酸的长度,小于90个、小于80个、小于70个、小于60个、小于50个、小于40个、小于30个、小于20个或小于10个核苷酸的长度。在一些实施方案中,在添加5'-帽和/或3'-聚a尾巴之后,检测或定量短聚体。
[0215]
延长的mrna转录物
[0216]
在一些实施方案中,由本文公开的方法产生的mrna转录物的至少75%、至少80%、至少85%、至少90%、至少95%在终止信号处终止。如本文所用的“在终止信号处终止”指终止信号3’末端的10个核苷酸、9个核苷酸、8个核苷酸、7个核苷酸、6个核苷酸、5个核苷酸、4个核苷酸、3个核苷酸、2个核苷酸、1个核苷酸或0个核苷酸内转录的终止。
[0217]
为了检测失控转录,可以消化mrna转录物以产生短的3’末端片段,使用液相色谱质谱(“消化lc/ms”)对其进行分析。合适的3’末端片段的大小小于100个核苷酸,例如,小于90个核苷酸、80个核苷酸、70个核苷酸、60个核苷酸、50个核苷酸、40个核苷酸。所需长度的
3’末端片段可以通过提供探针寡核苷酸来产生,所述探针寡核苷酸与模板化mrna转录物的3’末端特异性杂交,从而形成dna/rna杂交体。探针寡核苷酸可以结合在模板化mrna转录物的3’末端的约5至约20个核苷酸内。然后用rna酶h消化dna/rna杂交体,以产生所需长度的3’末端片段。可以使用rna测序来分析3’末端片段,以确定失控序列的存在和长度。用于rna测序的合适方法是纳米孔测序。
[0218]
合适的探针寡核苷酸是经修饰的rna-dna缺口寡核苷酸(通常也称为缺口多聚体(gapmer))。典型的缺口多聚体设计由5
’‑
翼和随后的8至12个脱氧核酸单体的缺口组成,所述脱氧核酸单体可以是天然核酸或在磷基团中含有硫离子(ps键),然后是3
’‑
翼。这种rna-dna-rna样构型通常含有被修饰的rna核苷酸,例如通过含有2
’‑
o-甲基核糖。本文公开rna-dna缺口寡核苷酸偏离了这种标准设计,因为它们具有仅3-5个脱氧核糖核酸单体(通常为4个脱氧核糖核酸单体)的较短缺口。这使得rna酶h消化能够精确定位于模板化mrna转录物的3’末端。为了确保与mrna转录物精确退火,rna-dna缺口寡核苷酸为10-20个核苷酸长,例如约15-18个核苷酸。包含经修饰的rna核苷酸的5’和3’翼序列不具有相同的长度。在一些实施方案中,5’翼(例如具有4-6个核苷酸的长度)比3’翼(其例如具有7-10个核苷酸的长度)短。在其他实施方案中,5’翼(例如具有7-10个核苷酸的长度)比3’翼(其例如具有4-6个核苷酸的长度)长。
[0219]
蛋白质表达
[0220]
用t7 rna聚合酶合成mrna转录物通常被比所需转录物更长或更短的rna污染(参见wo 2018/157153)。延长的序列被认为是通过在终止信号之后,在模板编码的mrna转录物的末端非模板化添加核苷酸而产生的。另外的核苷酸通常被称为“失控”。当失控的3’末端具有足够的互补性以分别结合自身或第二mrna分子以形成可延伸的分子内或分子间双链体时,可发生进一步延伸(gholamalipour等人2018,nucleic acids research 46:18,pp 9253-9263)。当双链rna(dsrna)进入细胞时,它被感知为病毒入侵者。这导致dsrna依赖酶的激活,诸如寡腺苷酸合成酶(oas)、rna特异性腺苷脱氨酶(adar)和rna激活蛋白激酶(pkr),导致蛋白质合成抑制(等人2019,molecular therapy:nucleic acids,15:26-35)。
[0221]
本技术的实施例证实了根据本发明的mrna合成防止了来自线性和超螺旋dna模板的mrna转录物的不期望的延长。根据本发明合成的mrna(包括使用t7 rna聚合酶合成的mrna,其3’末端没有不希望的延长)基本上不含dsrna,这可以用对dsrna特异的单克隆抗体来确定(例如,通过使用斑点印迹测定)。因此,当向受试者施用时,它不激活dsrna依赖性酶。因此,根据本发明合成的mrna导致更有效的蛋白质翻译。
[0222]
在一些实施方案中,相对于使用现有技术方法,特别是使用t7或t3rna聚合酶合成的相同量的mrna,根据本发明合成的mrna一旦转染到细胞中,就导致蛋白质表达增加,例如增加至少2倍、3倍、4倍、5倍、10倍、20倍、30倍、40倍、50倍、100倍、500倍、1000倍或更多。
[0223]
在一些实施方案中,相对于使用现有技术方法,特别是使用t7或t3rna合成的相同量的mrna,根据本发明合成的mrna一旦转染到细胞中,就导致由mrna编码的蛋白质活性增加,例如增加至少2倍、3倍、4倍、5倍、10倍、20倍、30倍、40倍、50倍、100倍、500倍、1000倍或更多。
[0224]
使用本发明可以合成任何mrna。在一些实施方案中,mrna编码一种或多种天然存
在的肽。在一些实施方案中,mrna编码一种或多种修饰的或非天然的肽。
[0225]
在一些实施方案中,mrna编码细胞内蛋白质。在一些实施方案中,mrna编码胞质蛋白质。在一些实施方案中,mrna编码与肌动蛋白细胞骨架相关的蛋白质。在一些实施方案中,mrna编码与质膜相关的蛋白质。在一些具体实施方案中,mrna编码跨膜蛋白。在一些具体实施方案中,mrna编码离子通道蛋白。在一些实施方案中,mrna编码核周蛋白。在一些实施方案中,mrna编码核蛋白。在一些具体实施方案中,mrna编码转录因子。在一些实施方案中,mrna编码伴侣蛋白。在一些实施方案中,mrna编码细胞内酶(例如,编码与尿素循环或溶酶体贮积代谢障碍相关的酶的mrna)。在一些实施方案中,mrna编码参与细胞代谢、dna修复、转录和/或翻译的蛋白质。在一些实施方案中,mrna编码细胞外蛋白质。在一些实施方案中,mrna编码与细胞外基质相关的蛋白质。在一些实施方案中,mrna编码分泌的蛋白质。在具体实施方案中,本发明的组合物和方法中使用的mrna可以用于表达被一种或多种靶细胞排泄或分泌到周围的细胞外液中的功能性蛋白质或酶(例如,编码激素和/或神经递质的mrna)。
[0226]
本发明提供了用于制备富含编码感兴趣的肽或多肽的全长mrna分子的治疗组合物的方法,所述治疗组合物用于递送至或治疗受试者,例如人类受试者或人类受试者的细胞或经处理并递送至人类受试者的细胞。
[0227]
因此,在某些实施方案中,本发明提供了用于制备富含编码肽或多肽的全长mrna的治疗组合物的方法,该治疗组合物用于递送至或治疗受试者的肺或肺细胞。在某些实施方案中,本发明提供了用于制备富含编码囊性纤维化跨膜传导调节因子(cftr)蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码atp结合盒亚家族a成员3蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码动力蛋白轴突中间链1蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码动力蛋白轴突重链5(dnah5)蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码α-1-抗胰蛋白酶蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码叉头盒p3(foxp3)蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码一种或多种表面活性蛋白的全长mrna的治疗组合物的方法,所述表面活性蛋白例如表面活性a蛋白、表面活性b蛋白、表面活性c蛋白和表面活性d蛋白中的一种或多种。
[0228]
在某些实施方案中,本发明提供了用于制备富含编码肽或多肽的全长mrna的治疗组合物的方法,该治疗组合物用于递送至或治疗受试者的肝脏或肝细胞。此类肽和多肽可包括与尿素循环紊乱相关的、与溶酶体贮积紊乱相关的、与糖原贮积紊乱相关的、与氨基酸代谢紊乱相关的、与脂质代谢或纤维化紊乱相关的、与甲基丙二酸血症相关或与任何其他代谢紊乱相关的那些,对于所述代谢紊乱,以富集的全长mrna递送至或治疗肝脏或肝细胞提供治疗有益效果。
[0229]
在某些实施方案中,本发明提供了用于制备富含编码与尿素循环障碍相关的蛋白质的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码鸟氨酸转氨甲酰酶(otc)蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码精氨琥珀酸合成酶1蛋白的全长mrna的治疗组合物的方法。在某
些实施方案中,本发明提供了用于制备富含编码氨基甲酰磷酸合成酶i蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码精氨琥珀酸裂合酶蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码精氨酸酶蛋白的全长mrna的治疗组合物的方法。
[0230]
在某些实施方案中,本发明提供了用于制备富含编码与溶酶体贮积障碍相关的蛋白质的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码α半乳糖苷酶蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码葡萄糖脑苷脂酶蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码异氰酸酯-2-硫酸酯酶蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码艾杜糖苷酸酶蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码n-乙酰基-α-d-葡糖胺苷酶蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码乙酰肝素n-硫酸酯酶蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码半乳糖胺-6硫酸酯酶蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码β-半乳糖苷酶蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码溶酶体脂肪酶蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码芳基硫酸酯酶b(n-乙酰半乳糖胺-4-硫酸酯酶)蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码转录因子eb(tfeb)的全长mrna的治疗组合物的方法。
[0231]
在某些实施方案中,本发明提供了用于制备富含编码与糖原贮积障碍相关的蛋白质的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码酸性α-葡糖苷酶蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码葡萄糖-6-磷酸酶(g6pc)蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码肝糖原磷酸化酶蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码肌肉磷酸甘油酸突变酶蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码糖原解支化酶的全长mrna的治疗组合物的方法。
[0232]
在某些实施方案中,本发明提供了用于制备富含编码与氨基酸代谢相关的蛋白质的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码苯丙氨酸羟化酶的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码戊二酰-coa脱氢酶的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码丙酰基-coa羧化酶的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码草酸酶丙氨酸-乙醛酸氨基转移酶的全长mrna的治疗组合物的方法。
[0233]
在某些实施方案中,本发明提供了用于制备富含编码与脂质代谢或纤维化障碍相关的蛋白质的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码mtor抑制剂的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码atp酶磷脂转运8b1(atp8b1)蛋白的全长mrna的治疗组合物的方法。在某
些实施方案中,本发明提供了用于制备富含编码一种或多种nf-κb抑制剂的全长mrna的治疗组合物的方法,所述抑制剂诸如i-κbα、干扰素相关的发育调节剂1(ifrd1)和sirtuin 1(sirt1)中的一种或多种。在某些实施方案中,本发明提供了用于制备富含编码ppar-γ蛋白或活性变体的全长mrna的治疗组合物的方法。
[0234]
在某些实施方案中,本发明提供了用于制备富含编码与甲基丙二酸血症相关的蛋白质的全长mrna的治疗组合物的方法。例如,在某些实施方案中,本发明提供了用于制备富含编码甲基丙二酰辅酶a突变酶蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码甲基丙二酰辅酶a差向异构酶蛋白的全长mrna的治疗组合物的方法。
[0235]
在某些实施方案中,本发明提供了用于制备富含全长mrna的治疗组合物的方法,对于该治疗组合物,递送至或治疗肝脏可以提供治疗有益效果。在某些实施方案中,本发明提供了用于制备富含编码atp7b蛋白(也被称为wilson病蛋白)的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码胆色素原脱氨酶的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码一种或多种凝结酶的全长mrna的治疗组合物的方法,所述凝结酶诸如因子viii、因子ix、因子vii和因子x。在某些实施方案中,本发明提供了用于制备富含编码人血色素沉着病(hfe)蛋白的全长mrna的治疗组合物的方法。
[0236]
在某些实施方案中,本发明提供了用于制备富含编码肽或多肽的全长mrna的治疗组合物的方法,该治疗组合物用于递送至或治疗受试者的心血管系统或心血管细胞。在某些实施方案中,本发明提供了用于制备富含编码血管内皮生长因子a蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码松弛素蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码骨形态发生蛋白9蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码骨形态发生蛋白2受体蛋白的全长mrna的治疗组合物的方法。
[0237]
在某些实施方案中,本发明提供了用于制备富含编码肽或多肽的全长mrna的治疗组合物的方法,该治疗组合物用于递送至或治疗受试者的肌肉或肌肉细胞。在某些实施方案中,本发明提供了用于制备富含编码肌营养不良蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码人源线粒体蛋白(frataxin)的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码肽或多肽的全长mrna的治疗组合物的方法,该治疗组合物用于递送至或治疗受试者的心肌或心肌细胞。在某些实施方案中,本发明提供了用于制备富含编码调节肌肉组织或肌肉细胞中钾通道和钠通道中的一者或两者的蛋白质的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码调节肌肉组织或肌肉细胞中kv7.1通道的蛋白质的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码调节肌肉组织或肌肉细胞中nav1.5通道的蛋白质的全长mrna的治疗组合物的方法。
[0238]
在某些实施方案中,本发明提供了用于制备富含编码肽或多肽的全长mrna的治疗组合物的方法,该治疗组合物用于递送至或治疗受试者的神经系统或神经系统细胞。例如,在某些实施方案中,本发明提供了用于制备富含编码存活运动神经元1蛋白的全长mrna的治疗组合物的方法。例如,在某些实施方案中,本发明提供了用于制备富含编码存活运动神
经元2蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码人源线粒体蛋白(frataxin)的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码atp结合盒亚家族d成员1(abcd1)蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码cln3蛋白的全长mrna的治疗组合物的方法。
[0239]
在某些实施方案中,本发明提供了用于制备富含编码肽或多肽的全长mrna的治疗组合物的方法,该治疗组合物用于递送至或治疗受试者的血液或骨髓或者血细胞或骨髓细胞。在某些实施方案中,本发明提供了用于制备富含编码β球蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码布鲁顿酪氨酸激酶蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码一种或多种凝结酶的全长mrna的治疗组合物的方法,所述凝结酶诸如因子viii、因子ix、因子vii和因子x。
[0240]
在某些实施方案中,本发明提供了用于制备富含编码肽或多肽的全长mrna的治疗组合物的方法,该治疗组合物用于递送至或治疗受试者的肾脏或肾脏细胞。在某些实施方案中,本发明提供了用于制备富含编码iv型胶原α5链(col4a5)蛋白的全长mrna的治疗组合物的方法。
[0241]
在某些实施方案中,本发明提供了用于制备富含编码肽或多肽的全长mrna的治疗组合物的方法,该治疗组合物用于递送至或治疗受试者的眼或眼细胞。在某些实施方案中,本发明提供了用于制备富含编码atp结合盒亚家族a成员4(abca4)蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码视黄醛壳素蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码视网膜色素上皮特异性65kda(rpe65)蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码290kda的中心体蛋白(cep290)的全长mrna的治疗组合物的方法。
[0242]
在某些实施方案中,本发明提供了用于制备富含编码肽或多肽的全长mrna的治疗组合物的方法,该治疗组合物用于向受试者或受试者的细胞递送疫苗或用疫苗治疗。例如,在某些实施方案中,本发明提供了用于制备富含编码来自诸如病毒之类的传染源的抗原的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码来自流感病毒的抗原的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码来自呼吸道合胞病毒的抗原的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码来自狂犬病病毒的抗原的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码来自巨细胞病毒的抗原的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码来自轮状病毒的抗原的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码来自肝炎病毒的抗原的全长mrna的治疗组合物的方法,所述肝炎病毒诸如甲型肝炎病毒、乙型肝炎病毒或丙型肝炎病毒。在某些实施方案中,本发明提供了用于制备富含编码来自人乳头瘤病毒的抗原的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码来自单纯疱疹病毒的抗原的全长mrna的治疗组合物的方法,所述单纯疱疹病毒诸如单纯疱疹病毒1或单纯疱疹病毒2。在某些实施方案中,本发明提供了用
于制备富含编码来自人免疫缺陷病毒的抗原的全长mrna的治疗组合物的方法,所述人免疫缺陷病毒诸如人免疫缺陷病毒1型或人免疫缺陷病毒2型。在某些实施方案中,本发明提供了用于制备富含编码来自人间质肺炎病毒的抗原的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码来自人副流感病毒的抗原的全长mrna的治疗组合物的方法,所述人副流感病毒诸如人副流感病毒1型、人副流感病毒2型或人副流感病毒3型。在某些实施方案中,本发明提供了用于制备富含编码来自疟疾病毒的抗原的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码来自寨卡病毒的抗原的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码来自基孔肯雅病毒的抗原的全长mrna的治疗组合物的方法。
[0243]
在某些实施方案中,本发明提供了用于制备富含全长mrna的治疗组合物的方法,该mrna编码与受试者的癌症相关的抗原或从受试者的癌细胞中鉴定的抗原。在某些实施方案中,本发明提供了用于制备富含全长mrna的治疗组合物的方法,该mrna编码从受试者自身的癌细胞确定的抗原,即提供个性化的癌症疫苗。在某些实施方案中,本发明提供了用于制备富含全长mrna的治疗组合物的方法,该mrna编码从突变kras基因表达的抗原。
[0244]
在某些实施方案中,本发明提供了用于制备富含编码抗体的全长mrna的治疗组合物的方法。在某些实施方案中,抗体可以是双特异性抗体。在某些实施方案中,抗体可以是融合蛋白的一部分。在某些实施方案中,本发明提供了用于制备富含编码针对ox40的抗体的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码针对vegf的抗体的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码针对组织坏死因子α的抗体的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码针对cd3的抗体的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码针对cd19的抗体的全长mrna的治疗组合物的方法。
[0245]
在某些实施方案中,本发明提供了用于制备富含编码免疫调节剂的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码白介素12的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码白介素23的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码白介素36γ的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含全长mrna的治疗组合物的方法,该mrna编码一种或多种干扰素基因(sting)蛋白刺激物的组成型活性变体。
[0246]
在某些实施方案中,本发明提供了用于制备富含编码核酸内切酶的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含全长mrna的治疗组合物的方法,该mrna编码rna指导的dna核酸内切酶蛋白,诸如cas 9蛋白。在某些实施方案中,本发明提供了用于制备富含编码大范围核酸酶蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码转录激活因子样效应子核酸酶蛋白的全长mrna的治疗组合物的方法。在某些实施方案中,本发明提供了用于制备富含编码锌指核酸酶蛋白的全长mrna的治疗组合物的方法。
[0247]
脂质纳米颗粒
[0248]
根据本发明合成的mrna可以使用任何方法配制和递送用于体内蛋白质生产。在一
些实施方案中,mrna被包封在转移媒介物诸如纳米颗粒中。其中,这种包封的一个目的通常是保护核酸免受可能含有降解核酸的酶或化学物质和/或导致核酸快速排泄的系统或受体的环境的影响。因此,在一些实施方案中,合适的递送媒介物能够增强其中包含的mrna的稳定性和/或促进mrna递送至靶细胞或组织。在一些实施方案中,纳米颗粒可以是基于脂质的纳米颗粒,例如包括脂质体或者基于聚合物的纳米颗粒。在一些实施方案中,纳米颗粒可以具有小于大约40-100nm的直径。纳米颗粒可以包括至少1μg、10μg、100μg、1mg、10mg、100mg、1g或更多的mrna。
[0249]
在一些实施方案中,转移媒介物是脂质体囊泡,或促进核酸转移至靶细胞和组织的其他工具。合适的转移媒介物包括但不限于脂质体、纳米脂质体、含神经酰胺的纳米脂质体、蛋白脂质体、纳米颗粒、磷酸钙-硅酸盐纳米颗粒、磷酸钙纳米颗粒、二氧化硅纳米颗粒、纳米晶体颗粒、半导体纳米颗粒、聚(d-精氨酸)、纳米树枝状聚合物、基于淀粉的递送系统、胶束、乳液、囊泡、质粒、病毒、磷酸钙核苷酸、适体、肽和其他媒介标签。还考虑使用生物纳米胶囊和其他病毒衣壳蛋白组装件作为合适的转移媒介物。(hum.gene ther.2008年9月;19(9):887-95)。
[0250]
脂质体可以包括一种或多种阳离子脂质、一种或多种非阳离子脂质、一种或多种基于甾醇的脂质和/或一种或多种peg修饰的脂质。脂质体可以包括三种或多种不同的脂质组分,一种不同的脂质组分是基于甾醇的阳离子脂质。在一些实施方案中,基于甾醇的阳离子脂质是咪唑胆固醇酯或“ice”脂质(参见wo 2011/068810,其通过引用整体并入)。在一些实施方案中,基于甾醇的阳离子脂质构成脂质纳米颗粒(例如脂质体)中总脂质的不超过70%(例如不超过65%和60%)。
[0251]
合适的脂质的实例包括,例如,磷脂酰化合物(例如磷脂酰甘油、磷脂酰胆碱、磷脂酰丝氨酸、磷脂酰乙醇胺、鞘脂类、脑苷脂和神经节苷脂)。
[0252]
阳离子脂质的非限制性实例包括c12-200、mc3、dlindma、dlinkc2dma、ckk-e12、ice(基于咪唑的)、hgt5000、hgt5001、of-02、dodac、ddab、dmrie、dospa、dogs、dodap、dodma和dmdma、dodac、dlendma、dmrie、clindma、cplindma、dmoba、docarbdap、dlindap、dlincarbdap、dlincdap、klin-k-dma、dlin-k-xtc2-dma和hgt4003或其组合。
[0253]
非阳离子脂质的非限制性实例包括神经酰胺;脑磷脂;脑苷脂;二酰基甘油;1,2-二棕榈酰基-sn-甘油-3-磷酰甘油钠盐(dppg);1,2-二硬脂酰基-sn-甘油-3-磷酸乙醇胺(dspe);1,2-二硬脂酰基-sn-甘油-3-磷酸胆碱(dspc);1,2-二棕榈酰基-sn-甘油-3-磷酸胆碱(dppc);1,2-二油基-sn-甘油-3-磷酸乙醇胺(dope);1,2-二油酰基-sn-甘油-3-磷酸乙醇胺(depe),1,2-二油基-sn-甘油-3-磷脂酰胆碱(dopc);1,2-二棕榈酰基-sn-甘油-3-磷酸乙醇胺(dppe);1,2-二肉豆蔻酰基-sn-甘油-3-磷酸乙醇胺(dmpe);和1,2-二油基-sn-甘油-3-磷酸-(1'-rac-甘油)(dopg)、1-棕榈酰基-2-油基-磷脂酰乙醇胺(pope);1-棕榈酰基-2-油基-sn-甘油-3-磷酸胆碱(popc);1-硬脂酰基-2-油基-磷脂酰乙醇胺(sope);鞘磷脂;或它们的组合。
[0254]
在一些实施方案中,peg修饰的脂质可以是长度为最多5kda的聚(乙二醇)链,其共价连接至具有一条或多条c
6-c
20
长度的烷基链的脂质。peg修饰的脂质的非限制性实例包括dmg-peg、dmg-peg2k、c8-peg、dog peg、神经酰胺peg和dspe-peg,或其组合。
[0255]
还考虑使用聚合物作为转移媒介物,无论是单独使用还是与其他转移媒介物组合
使用。合适的聚合物可包括例如聚丙烯酸酯、聚氰基丙烯酸烷基酯、聚丙交酯、聚丙交酯-聚乙交酯共聚物、聚己内酯、葡聚糖、白蛋白、明胶、海藻酸盐、胶原、壳聚糖、环糊精和聚乙烯亚胺。基于聚合物的纳米颗粒可以包括聚乙烯亚胺(pei),例如支化的pei。
[0256]
通常,根据本发明的脂质体的脂质部分由3或4种脂质组分组成。根据本发明的4组分脂质体通常具有以下脂质组分:阳离子脂质(通常为可离子化的阳离子脂质,诸如ckk-e12或基于环状氨基酸的脂质)、非阳离子脂质(例如dope或depe)、基于胆固醇的脂质(例如胆固醇)和peg修饰的脂质(例如dmg-peg2k)。根据本发明的3组分脂质体通常具有以下脂质组分:基于甾醇的脂质(例如ice或其他基于咪唑的胆固醇衍生物)、非阳离子脂质(例如dope或depe)和peg修饰的脂质(例如dmg-peg2k)。
[0257]
在以下中的一个或多个中描述了与本发明有关的附加教导:wo 2011/068810、wo 2012/075040、us 15/294,249,us 62/421,021和us 15/809,680,以及申请人于2017年2月27日提交的题为“methods for purification of messenger rna”、“novel codon-optimized cftr sequence”和“methods for purification of messenger rna”的相关申请,这些申请的全部内容以引用的方式整体并入。
[0258]
用于本发明的组合物中的脂质体转移媒介物可通过本领域目前已知的各种技术制备。例如,多层囊泡(mlv)可根据常规技术制备,例如通过将脂质溶解在合适的溶剂中,将所选择的脂质沉积在合适的器皿或容器的内壁上,然后蒸发溶剂以在容器的内部上留下薄膜或喷雾干燥。然后可用涡旋运动将水相添加至器皿中,这导致mlv的形成。然后可通过多层囊泡的均质化、超声处理或挤出来形成单层囊泡(ulv)。另外,可通过清洁剂去除技术形成单层囊泡。
[0259]
各种方法在公开的美国申请号us 2011/0244026、公开的美国申请号us 2016/0038432、公开的美国申请号us 2018/0153822、公开的美国申请号us 2018/0125989和2019年7月23日提交的美国临时申请号62/877,597中有所描述,并且可以被用于实践本发明,所有这些申请均以引用的方式并入本文。如本文所用,方法a是指通过将mrna与脂质混合物混合而无需首先将脂质预先形成为脂质纳米颗粒来包封mrna的常规方法,如us 2016/0038432中所述。如本文所用,方法b是指通过将预先形成的脂质纳米颗粒与mrna混合来包封信使rna(mrna)的过程,如us 2018/0153822中所述。
[0260]
将所需mrna掺入脂质体内的过程经常称为“加载”。示例性方法在lasic等人febs lett.,312:255-258,1992中有所描述,该文献以引用的方式并入本文。掺入脂质体的mrna可完全或部分地位于脂质体的内部空间中、脂质体的双层膜内、或与脂质体膜的外表面结合。将mrna掺入脂质体内在本文中也称为“包封”,其中核酸完全包含在脂质体的内部空间内。将mrna掺入转移媒介物(例如脂质体)中的目的经常是保护核酸免受环境侵害,所述环境可能含有降解核酸的酶或化学物质和/或促使核酸快速排泄的系统或受体。因此,在一些实施方案中,合适的递送媒介物能够增强其中包含的mrna的稳定性和/或促进其递送至靶细胞或组织。
[0261]
药物组合物
[0262]
通过组合本文所述的各种方法以提供通过本文所述的方法准确转录为模板的优化的dna序列,提供了高质量的mrna转录物,其基本上不含dsrna以及污染的短聚体和长聚体序列。使用本文描述的纯化方法(特别是本文所述的用于纯化mrna的基于沉淀的、无乙醇
的方法)有效回收此类mrna转录物,产生具有相同优异性质的高纯度mrna转录物。通过将预先形成的脂质纳米颗粒与纯化的mrna混合(例如,通过使用如上所述的方法b)来包封这些mrna转录物可以产生异常高的包封效率(例如,大于90%)。组合这些不同加工步骤的最终结果是一种药物产品,其在将mrna递送至靶细胞以实现mrna编码的肽、多肽或蛋白质的最大表达方面极其有效。
[0263]
因此,在一些实施方案中,本发明提供了一种制备药物组合物的方法,包括以下步骤:
[0264]
a)提供包含蛋白质编码序列的dna序列;
[0265]
b)通过以下方式优化dna序列:
[0266]
i)确定dna序列中终止信号的存在,其中终止信号具有以下核酸序列:5
’‑
x1atctx2tx
3-3’,其中x1、x2和x3独立地选自a、c、t或g,并且如果存在一个或多个终止信号,通过用其它三个核酸中的任何一个替换所述一个或多个终止信号的位置2、3、4、5和7中任何一个位置的一个或多个核酸来修饰dna序列,以产生优化的dna序列,其中,如果需要,选择一个或多个替换核酸以保留由蛋白质编码序列编码的蛋白质的氨基酸序列,和/或
[0267]
ii)在蛋白质编码序列的3’末端添加一个或多个终止信号,其中一个或多个终止信号包含以下核酸序列:x1atctx2tx
3-3’,其中x1、x2和x3独立地选自a、c、t或g;
[0268]
c)从步骤(b)的优化的dna模板通过体外转录合成mrna;
[0269]
d)从步骤(c)的制剂中沉淀mrna;
[0270]
e)通过切向流过滤纯化包含步骤(d)的经沉淀的mrna的不纯制剂;
[0271]
f)将步骤(e)的纯化mrna包封在脂质体中,所述脂质体包含一种或多种阳离子脂质、一种或多种非阳离子脂质、一种或多种基于甾醇的脂质和一种或多种peg修饰的脂质。
[0272]
在一些实施方案中,该方法包括在步骤(e)之后进行的单独的加帽和加尾反应。在这些实施方案中,在加帽和加尾反应后重复步骤(d)和(e)。在一些实施方案中,纯化不纯制剂涉及无乙醇方法。在一些实施方案中,包封涉及将纯化的mrna与预先形成的脂质纳米颗粒混合。在一些实施方案中,步骤(f)之后是配制步骤。配制步骤可以涉及缓冲液交换。在一些实施方案中,配制步骤涉及冻干包封mrna的脂质体。
[0273]
尽管与本文描述的那些类似或等同的方法和材料可以用于本发明的实践或测试中,但下面对合适的方法和材料进行描述。本文提及的所有出版物、专利申请、专利和其他参考文献以引用的方式整体并入本文。本文引用的参考文献不被视为所要求保护的发明的现有技术。另外,所述材料、方法和实施例仅仅是说明性的,并且不旨在进行限制。
[0274]
实施例
[0275]
实施例1:使用t7和sp6 rna聚合酶的mrna合成的示例性实验设计
[0276]
该实施例说明了基于t7和sp6 rna聚合酶的mrna合成、转染及其表征的示例性条件。
[0277]
信使rna材料
[0278]
用限制性内切酶将具有编码感兴趣的蛋白质编码序列的dna序列的质粒线性化并纯化,所述dna序列与rna聚合酶启动子可操作地连接。由纯化的和线性化的质粒通过体外转录合成mrna转录物。t7转录反应由1x t7转录缓冲液(80mm hepes ph 8.0、2mm亚精胺和25mm mgcl2,最终ph 7.7),10mm dtt,atp、gtp、ctp和utp各7.25mm、rna酶抑制剂、焦磷酸酶
和t7 rna聚合酶组成。sp6反应包括5mm的每种ntp、约0.05mg/ml的sp6 rna聚合酶dna和约0.1mg/ml的模板dna;转录缓冲液的其他组分各不相同。反应在37℃下进行60至90分钟(除非另有说明)。添加dna酶i以终止反应,并在37℃下再孵育15分钟。按照制造商的建议,使用qiagen rna maxi柱纯化体外转录的mrna。
[0279]
用部分gtp(1.25mm)处理来自上述体外转录步骤的纯化mrna转录物,将s-腺苷甲硫氨酸、rna酶抑制剂、2
’‑
o-甲基转移酶和鸟苷酸转移酶与反应缓冲液(10x,500mm tris-hcl(ph 7.5),60mm kcl,10mm mgcl2)混合在一起。将合并的溶液在37℃下孵育一段时间持续30至90分钟。完成后,添加atp(2.0mm)、聚a聚合酶和加尾反应缓冲液(10x,500mm tris-hcl(ph 7.5)、2.5m nacl、100mm mgcl2)并将总反应混合物在37℃下进一步孵育持续一段时间20至45分钟。完成后,将最终的反应混合物淬灭并相应地纯化。
[0280]
琼脂糖凝胶电泳:
[0281]
使用在50ml tae缓冲液中的0.5g琼脂糖制备1%琼脂糖凝胶。用2x乙二醛凝胶加载染料或2x甲酰胺凝胶加载染料处理1至2g rna,加载到琼脂糖凝胶上,并在130v下运行30或60分钟。
[0282]
毛细管电泳(ce)
[0283]
标准灵敏度rna分析试剂盒(15nt)购自advanced analytical,并用于具有12个毛细管阵列的片段分析仪(advanced analytical)上的毛细管电泳运行。凝胶引发后,将300ng总rna与稀释标记物以1∶11(rna∶标记物)的比例混合,并将24l加载到96孔板的每个孔中。通过混合2μl的标准灵敏度rna梯度与22μl稀释标记物混合制备分子量指示梯度。样品注入电压为5.0kv,时间为4秒,样品分离电压为8.0kv,时间为40.0分钟。通过prosize 2软件(advanced analytical)处理每个样品的基于荧光的电泳图,产生样品中存在的片段表格形式的大小(bp)和丰度(ng/μl)。
[0284]
消化液相色谱质谱(lc/ms)
[0285]
探针寡核苷酸与mrna转录物退火,然后用rna酶h和虾碱性磷酸酶消化。消化反应由1x rna酶h缓冲液(neb)、rnase h(neb)、虾碱性磷酸酶(neb)、退火的mrna和探针寡核苷酸组成。消化反应在37℃下进行40分钟。
[0286]
用uhplc-qtof系统(agilent)进行mrna片段的分析。流动相a由100mm六氟异丙醇和8.6mm三乙胺组成,ph 8.3,并且流动相b是100%meoh。所有分析均使用agilent infinitylab c18 2.1x 100mm柱,流速为0.5ml/min,温度为50℃。在经12分钟使用自5%至23%梯度的流动相b,随后使用2min用50%流动相b进行洗涤步骤,以洗脱rna片段。所有质谱都是以负离子模式在400-3200m/z的扫描范围内获得的,ms设置如下:干燥气流,13l/min;气体温度,350℃;喷雾器压力,10psi;毛细管电压,3750v。使用masshunter数据采集软件(agilent)采集样品数据。
[0287]
rna测序
[0288]
设计一种寡核苷酸,其包含10-15个碱基的3’条形码序列,随后是25-40as的聚a段,在3’和5’末端都有磷酸基团,以允许分别连接到mrna转录物并防止自身连接。首先,用rsap处理mrna转录物以去除它们的5’磷酸基团,从而防止寡核苷酸连接到mrna转录物的5’末端。使用t4 rna连接酶,将寡核苷酸连接到mrna转录物上,产生ho-mrna转录物-条形码-聚a-po4构建体。第二rsap处理步骤从上述构建体中除去3’磷酸,为纳米孔测序做准备
(minion,oxford nanopore)。
[0289]
对于纳米孔测序,对ho-mrna转录物-条形码-聚a-oh构建体进行退火并连接到测序接头上,并在加载到纳米孔细胞芯片中之前根据制造商的方案被锚定。一旦上样,样品以3’到5’的方向通过纳米孔。测序完成后,解析含有条形码的部分的读段,并收集和分析该区域后的碱基。
[0290]
实施例2:rrnb终止子t1信号的存在导致mrna转录物的过早终止
[0291]
该实施例说明了在感兴趣的蛋白质编码序列的密码子优化的dna序列中终止信号的意外存在如何导致体外转录的过早终止,从而导致mrna转录物的异质群体,其中只有一部分包括全长蛋白质编码序列。
[0292]
如实施例1中所述,使用sp6 rna聚合酶将具有可操作地连接到rna聚合酶启动子的编码感兴趣的密码子优化的蛋白质编码序列(mrna-1)的dna序列的质粒用于mrna转录物的体外转录。通过毛细管电泳(ce)评估mrna转录物的大小(图1)。全长mrna-1转录物的长度约为1900个核苷酸。大约45%的mrna-1转录物是长度约900个核苷酸的截短转录物(图1)。
[0293]
已报道大肠杆菌rrnb终止子t1信号共有序列tatctgtt的存在导致sp6和t7 rna聚合酶两者的暂停或终止(kwon&kang 1999,the journal of biological chemistry,274:41,pp 29149-29155;sohn&kang,2005,pnas,102:1,pp.75-80)。对mrna-1蛋白编码序列的分析发现,它包括起始于核苷酸796的共有rrnb终止子t1信号。
[0294]
在野生型rrnb终止子t1信号中,共有序列tatctgtt紧接着是gtttgtcgtg序列。当评估变体rrnb1终止子t1信号序列的终止效率时,kwon&kang(同上)在除了一个测试的变体终止子t1信号之外的所有测试的变体终止子t1信号中,在tatgtctt共有序列的紧接3’的5个核苷酸中包括至少三个t碱基。当该区域被删除时,观察到0%的终止效率,表明该下游富含t的序列是在rrnb终止子t1信号处终止所必需的。然而,mrna-1中的共有rrnb终止子t1信号后面没有下游富含t的序列(相反,它存在于以下序列中:5
’‑
gctatctgttcatca-3’,seq id no:29),证明这不是终止子序列的必需元件。
[0295]
该实施例证实,mrna构建体中rrnb终止子t1信号共有序列的存在可导致mrna转录物的过早终止,甚至在缺乏富含t的序列的情况下,导致所需全长mrna转录物的产量显著降低。
[0296]
实施例3:rrnb终止t1信号的变体也导致mrna转录物的过早终止
[0297]
该实施例证实了在相对于共有序列的位置1、6或8具有点突变的rrnb终止子t1信号的存在也导致体外转录期间的过早终止。
[0298]
产生来自实施例2的mrna-1蛋白编码序列的变体,其中tatctgtt共有终止信号序列在单个位置突变,以确定rrnb终止t1信号中的哪些核苷酸对于转录的终止是必需的(见下表1)。
[0299]
使用sp6 rna聚合酶将变体用于体外转录。如实施例1所述,通过毛细管电泳确定mrna转录物的大小。在大约1900个核苷酸处观察到的条带代表全长mrna-1构建体。在大约900个核苷酸处观察到一条带,其中mrna转录物由于在变体终止信号处过早终止而被截短。该实验的结果显示在通过ce由mrna转录物的定量产生的数字凝胶图像中(图2),并总结在表1中。
[0300]
表1:截断分析的结构设计和结果
[0301][0302][0303]
如果相对于共有序列,位置1、6或8处的核苷酸身份已经改变,mrna-1变体仍然产生截短的mrna转录物。然而,如果位置2、3、4、5或7的核苷酸已经突变,则没有观察到截短。因此,当在具有蛋白质编码序列的dna序列中筛选终止信号时,应该考虑共有序列和在位置1、6和8处具有点突变的序列变体,以避免体外转录的过早终止。
[0304]
先前已知,如果tatctgtt共有终止信号序列第四位的c被g替换,则不会发生终止(sohn&kang,2005,pnas,102:1,pp.75-80)。如果位置2、3、4、5或7上的任何残基被该位置上非天然存在的另一个核苷酸替换,则没有观察到截短,这一发现为去除终止信号而不改变dna序列编码的蛋白质编码序列提供了更大的灵活性。
[0305]
实施例4:rrnb终止子t1信号导致mrna转录物的过早终止
[0306]
该实施例证实了通过计算机筛选rrnb终止子t1信号,可以预测使用sp6 rna聚合酶的体外转录产生的mrna转录物中的过早终止位点。
[0307]
在计算机中筛选具有密码子优化的蛋白质编码序列的各种dna序列,以确定大肠杆菌rrnb终止子t1信号共有序列tatctgtt或在位置1、6或8处不同于该序列的变体序列的存在,如实施例3中所确定的。进行该分析以预测当聚合酶在这些终止信号处过早终止时将产生的截短的mrna转录物的大小(见下表2)。
[0308]
为了测试计算机预测的准确性,使用sp6/t7 rna聚合酶体外转录密码子优化的蛋白质编码序列,并通过毛细管电泳(ce)确定mrna转录物的实际大小,如实施例1中所述。将任何截短的mrna转录物的实际大小与通过计算机分析预测的大小进行比较(见表2)。
[0309]
表2:截短的mrna转录物大小的预测值和实验确定值
[0310][0311]
在所有鉴定的终止子信号中观察到mrna转录物的截短。基于rrnb终止子t1信号的鉴定的截短的mrna转录物的预测大小与实验确定的截短的mrna转录物的大小成正相关。
[0312]
实施例5:由sp6 rna聚合酶产生的mrna转录物不包含双链rna
[0313]
本实施例证实了sp6 rna聚合酶的mrna转录物确实包含rna双链体,这不同于t7 rna聚合酶合成的mrna转录物。
[0314]
用t7 rna聚合酶合成的mrna转录物通常被比所需转录物更长或更短的rna污染(参见wo 2018/157153)。非常短的转录物通常被称为“短聚体”,并且通常必须通过大量纯化体外转录的mrna来去除。
[0315]
延长的序列被认为是通过在终止信号之后,在模板编码的mrna转录物的末端非模板化添加核苷酸而产生的。另外的核苷酸通常被称为“失控”。当失控的3’末端具有足够的互补性以分别结合自身或第二mrna分子以形成可延伸的分子内或分子间双链体时,可发生进一步延伸(gholamalipour等人2018,nucleic acids research 46:18,pp 9253-9263)。
[0316]
使用sp6 rna聚合酶或t7 rna聚合酶从四种不同的模板质粒通过体外转录产生mrna转录物,如实施例1所述。每个模板质粒编码一个编码相同蛋白质的mrna转录物。rna双链体的存在通过用抗dsrna单克隆抗体j2进行的斑点印迹分析来检测,如等人2019,molecular therapy:nucleic acids,15:26-35中所述。
[0317]
将2μl每份体外转录的mrna样品(相当于每点200ng总mrna)点在带正电荷的尼龙膜上。以每点2ng和25ng点样dsrna的对照样品。为了检测dsrna,将膜与抗dsrna鼠单克隆抗体j2一起孵育。使用与辣根过氧化物酶偶联的抗小鼠igg抗体进行检测。产生的斑点印迹如图3所示。从该图可以看出,在用sp6 rna聚合酶合成的mrna转录物中没有检测到双链rna,而在用t7 rna聚合酶制备的样品中检测到大量的双链rna。
[0318]
sp6 rna聚合酶合成的mrna转录物不形成分子内或分子间双链体。
[0319]
实施例6:由t7 rna聚合酶和sp6 rna聚合酶产生的mrna转录物通过非模板延长而延伸
[0320]
该实施例证实了由t7和sp6 rna聚合酶合成的mrna转录物被失控转录延长。
[0321]
使用sp6 rna聚合酶进行体外转录避免了通常使用t7 rna聚合酶观察到的mrna转录物中短聚体的形成(参见wo 2018/157153)。由sp6 rna聚合酶合成的mrna转录物通常在大小上显得更均匀。
[0322]
为了确定sp6 rna聚合酶,如t7 rna聚合酶,在遇到终止信号后是否继续以非模板介导的方式延长mrna转录物(失控转录),设计了一组探针寡核苷酸,其结合到由模板序列编码的mrna转录物的3’末端。探针寡核苷酸是由integrated dna technologies(coralville,ia)合成rna-dna缺口寡核苷酸。它们的序列和糖修饰如表3所示。2
’‑
o-甲基核糖修饰的rna核苷酸在相应的碱基前用“m”表示,并且dna核苷酸用斜体表示。
[0323]
表3:探针寡核苷酸
[0324][0325]
添加rna酶h来消化dna/rna杂交体,只留下模板序列3’末端的mrna转录物片段。mrna转录物的3’末端消化产物的大小通过液相色谱质谱(lc/ms)确定,如实施例1所述。在测试的六种寡核苷酸中,探针寡核苷酸#1产生最长的预期的3’消化产物(caucaagcu),并被选择用于进一步的实验。
[0326]
结果示于图4a中。如果sp6 mrna转录物终止于模板序列的末端(即其中没有失控延长),则用探针寡核苷酸#1获得9个核苷酸的3’消化产物(caucaagcu)。如实施例1中所述,通过质谱法证实了该消化产物的身份。质谱分析的结果示于图4b中。在mrna产物发生失控延长的地方获得了更长的3’消化产物。
[0327]
用t7 rna聚合酶重复该实验。图5a中比较了sp6和t7 rna聚合酶转录的mrna的3’消化产物的lc/ms分析结果。还通过对t7 rna聚合酶和sp6 rna聚合酶mrna转录物进行测序确定了通过失控延长添加到3’末端的碱基数量(图5b)。相对于由t7 rna聚合酶合成的mrna转录物,由sp6 rna聚合酶合成的mrna转录物具有更短的失控序列,但是还产生了更低百分比的没有另外失控序列的mrna转录物。
[0328]
这些数据证实,当同时使用sp6 rna聚合酶和t7 rna聚合酶时,体外合成的mrna转录物发生非模板延长。
[0329]
实施例7:在3’末端包含终止信号防止了由线性化质粒合成的mrna转录物的不希望的延长
[0330]
该实施例证实了在编码mrna转录物的dna序列的3’末端添加一个或多个终止信号减少了由线性化质粒转录的mrna的不期望的延长。
[0331]
使用标准分子生物学方法,通过插入质粒,将编码mrna转录物(mrna-12)的dna序列可操作地连接到sp6 rna聚合酶启动子上。将得到的质粒用于体外转录(经历或未经历预先线性化)。通过用序列特异性限制酶在转录起始位点下游880bp处切割质粒来进行线性化。如图6a所示,如实施例1所述,通过毛细管电泳确定线性化的质粒产生单一的879nt长的mrna转录物。
[0332]
为了确定终止序列的插入是否能导致dna序列末端转录的有效终止,制备了两种
修饰的质粒。质粒1在编码mrna转录物的dna序列的3’末端包括一个单一的rrnb终止t1信号(ttttatctgtttttttttt)。质粒2在dna序列的3’末端含有两个拷贝的相同终止信号(ttttatctgtttttttttttttatctgttttttttt)。
[0333]
将未经修饰的质粒和修饰的质粒1和2线性化,并用作用于使用sp6rna聚合酶的体外转录的模板,并通过毛细管电泳测定mrna转录物的大小,如实施例1中所述。
[0334]
如图6b所示,质粒1产生了796nt长的稍短的mrna转录物,表明终止发生在新添加的终止序列处。然而,终止信号在停止聚合酶方面并不完全有效,正如接近第一个峰的第二个峰所证明的那样,这表明存在相对大量的情况,其中rna聚合酶不直接在终止信号处终止,而是在终止转录之前继续转录一小段距离。对于从质粒2产生的mrna转录物来说,该第二个峰是不可见的(见图6c),这表明包含仅相隔10个碱基对两个串联的终止信号有效地防止了mrna转录物的不期望的延长。
[0335]
实施例8:在3’末端包含终止信号防止了由超螺旋质粒dna合成的mrna转录物的不希望的延长
[0336]
该实施例证实,在编码mrna转录物的dna序列的3’末端添加一个或多个终止信号,消除了在体外转录之前线性化环状核酸载体的需要。
[0337]
为了确定如果终止信号包括在编码mrna转录物的dna序列的3’末端,是否需要线性化,在体外转录之前,重复实施例7的实验,而不线性化质粒。当将超螺旋质粒dna用于未经修饰质粒的体外转录时,在所得mrna转录物的毛细管电泳期间可以看到多个新峰(图7a)。最大峰(代表总mrna转录物的约55%)对应于长度为约3126nt至约3230nt的mrna转录物。对质粒的核苷酸序列的更仔细的检查鉴定了编码mrna转录物的dna测序下游的终止信号(catctatt)。基于核苷酸序列分析,mrna转录物的预测大小预计为3306nt,并且因此与在约3126nt至约3230nt观察到的峰大小成正相关。这一观察表明,rna聚合酶继续转录超螺旋质粒dna,直到它遇到已经存在于质粒骨架中的终止信号,此时转录被终止,顺便证实了在编码所需mrna转录物的dna序列的末端包含终止信号应消除在体外转录之前对质粒线性化的需要。对应于甚至更大转录物的多个较小峰的存在表明,在反应终止之前反应混合物中至少一部分rna聚合酶多次转录质粒模板。相反,当质粒1用作模板时,终止信号tatctgtt的存在导致更有效的转录终止,约70%的mrna转录物具有约792nt的大小(图7b)。质粒2含有两个串联的tatctgtt终止信号,仅相隔10个碱基对,正确终止的mrna转录物的百分比进一步提高到约95%(图7c)。
[0338]
该实施例证实,在编码mrna转录物的dna序列的3’末端存在一个或多个终止信号,消除了对包含模板的质粒进行线性化的需要,因为mrna合成主要终止于dna序列的末端。鉴于mrna转录物的长度,终止信号的存在似乎也阻止了失控转录。具体而言,包含仅由10个碱基对分隔的两个共有终止信号可以导致转录的高效终止,防止失控转录并消除质粒线性化的需要。
[0339]
实施例9:体外转录在高于37℃的温度下改善了终止
[0340]
该实施例证实,如果体外转录反应在高于37℃的温度下进行,则在一个或多个终止信号处发生终止的可能性更高。
[0341]
为了确定正确终止的mrna转录物的百分比可否进一步提高,用质粒2重复实施例8中描述的实验,但是在不同的温度下。使用sp6 rna聚合酶,并且除了温度之外,反应条件与
实施例1中描述的相同,超螺旋质粒2被用作体外转录反应的模板。同样如实施例1中所述,通过毛细管电泳确定所得mrna转录物的大小。
[0342]
通过在置于加热器中的eppendorf管中进行体外转录反应来控制反应温度。在先前使用的37℃温度下,从超螺旋质粒2获得的92%-95%的mrna转录物被正确终止,如图8a所示。随着进行体外转录反应的温度增加到43℃、50℃或55℃,正确终止的质粒的百分比进一步增加。正确终止的mrna转录物的产量在50℃达到峰值。如图8b所示,在该温度下,从质粒2获得的99.7%的mrna转录物被正确终止。在50℃下仅观察到极小的降解。
[0343]
使用具有一个或多个终止信号的超螺旋质粒,并在高于37℃的温度下进行体外转录反应,得到了最大化正确终止的mrna转录物产量而无需质粒线性化的反应条件。
[0344]
实施例10:在3’末端包含两个以上的终止信号导致在37℃正确终止的mrna转录物
[0345]
该实施例证实,当在37℃下进行体外转录时,如果在编码mrna转录物的dna序列的3’末端存在超过两个拷贝的终止信号,可以达到接近100%的正确终止的mrna转录物的产量。这使得在常规条件下进行体外转录反应成为可能,而不需要在进行反应之前线性化环状核酸载体。
[0346]
为了研究在模板dna质粒中插入两个以上终止信号的效果,制备了编码mrna-12的进一步修饰的质粒。如实施例7中所述制备质粒1和2。质粒3在编码mrna转录物的dna序列的3’末端含有三个拷贝的rrnb终止t1信号,产生以下终止子序列:
[0347]
ttttatctgtttttttttttttatctgtttttttttttttatctgttttttttt。
[0348]
质粒1和2中的终止子序列直接插入在mrna-12的蛋白质编码区之后,而质粒3中的终止子序列插入在3’utr区之后。因此,当质粒3用作体外转录的dna模板时,正确终止的转录物(约880个核苷酸长)比使用质粒1或2时产生的转录物(约780个核苷酸长)更长。
[0349]
对于未经修饰的质粒和经修饰的质粒1、2和3,重复实施例9的实验(dna质粒未线性化,在37℃和50℃进行体外转录)。通过在置于热循环仪中的eppendorf管中进行体外转录反应来控制反应温度。
[0350]
与实施例8非常相似,在37℃下观察到的未经修饰质粒的体外转录的最大峰对应于长度约3119nt的mrna转录物(图9a)。这再次表明rna聚合酶继续转录超螺旋质粒dna,直到其遇到已经存在于质粒骨架中的终止信号,此时转录终止。当在50℃下进行体外转录时,观察到降解,导致对应于长度约为3099nt的mrna转录物的较小峰(相对于37℃转录反应的等效峰)(图9b)。
[0351]
当质粒1用作模板时,终止信号的存在导致更有效的转录终止,对于在37℃和50℃下进行的转录反应,分别约62%和约74%的mrna转录物具有对应于全长mrna-12转录物的大小(图(9c和9d)。对于质粒2,其包括串联的两个终止信号,正确终止的mrna-12转录物的百分比进一步增加,对于在37℃和50℃下进行的转录反应,分别达到约90%和约93%(图(9e和9f))。对于质粒3,其包括串联的三个终止信号,正确终止的转录物的比例进一步增加,对于在37℃下进行的转录反应,达到接近100%的产率,对于50℃下进行的转录反应,达到》99.0%的产率(图9g和9h)。至于未经修饰的质粒,对于使用质粒1、2或3作为dna模板在50℃下进行的转录反应,观察到mrna转录物的显著降解。在该实验中观察到在50℃下进行的反应的显著降解,而在实施例9中描述的实验中没有观察到这一事实,这表明实施例9中的反应温度可能没有始终保持在50℃下(例如,因为eppendorf管的一部分暴露于温度比加
热块本身低得多的环境空气中)。这表明可能需要优化反应条件,以在高于37℃的温度下将降解降至最低。
[0352]
该实施例进一步证实,增加编码mrna转录物的dna序列3’末端终止信号的数量可以避免包含模板的质粒线性化的需要,因为mrna合成主要终止于dna序列的末端。该实施例还表明,当一个或两个终止信号被串联时,正确终止的mrna转录物的产量可以通过在较高温度下进行转录反应来提高,尽管必须注意使mrna降解最小化。包含两个以上串联的终止信号使得正确终止的mrna转录物的产量接近100%,表明当转录反应在37℃下进行时,对于此类dna质粒模板,终止效率可以最大化。
[0353]
实施例11:从具有3’终止信号的超螺旋质粒dna转录的mrna在体外有效表达
[0354]
该实施例证实,对于从具有3’终止信号的超螺旋质粒dna转录的mrna和从线性化质粒转录的mrna可以获得相当水平的蛋白质表达。因此,该实施例提供了进一步的证据,即包含3’终止信号消除了在体外转录之前线性化环状核酸载体的需要。
[0355]
确定mrna-12转录物以及等效mrna-12转录物的蛋白质表达水平,所述mrna-12转录物是通过由如实施例10中所述的超螺旋质粒3(在编码mrna转录物的dna序列的3’末端含有三个拷贝的rrnb终止t1信号)的体外转录制备的,以及所述等效mrna-12转录物是通过由线性化的未修饰质粒(不含有终止信号)的体外转录制备的。
[0356]
使用非细胞翻译系统(cfts)评估蛋白质表达水平。cfts是以高通量方式筛选mrna构建体表达的有用工具,不需要维持细胞培养或使用转染剂。cfts的核心组分是从hela细胞产生的细胞质提取物,hela细胞含有表达蛋白质所需的必要机制(mikami等人2005,protein expression and purification,46,348-357)。通过调节辅助反应组分,主要是mg
2+
和k
+
水平,针对感兴趣的蛋白质优化蛋白质表达。该实施例中使用的cfts反应条件和组分已被优化用于表达由mrna-12转录物编码的蛋白质。
[0357]
为每种mrna-12转录物制备两种单独的cfts反应混合物。cfts反应混合物在65μl反应体积中含有325个fmol mrna-12转录物、40%(v/v)hela细胞质提取物(20mg/ml总蛋白)、27mm hepes(ph 7.5)、140mm koac、1.2mm mg(oac)2、16mm kcl、rna酶抑制剂(1u/μl)、1mm dtt、1.2mm atp、125μm gtp、30μm氨基酸混合物、300μm亚精胺、18mm肌酸磷酸酶、60μg/ml肌酸激酶和90μg/ml小牛肝trna。
[0358]
将反应混合物在25℃下孵育两小时。此后,将反应混合物储存在-80℃下,直到通过elisa确定蛋白质表达水平。该分析的结果如图10所示。图10显示,与从线性化的未经修饰质粒转录的mrna-12相比,从超螺旋质粒3转录的mrna-12可获得相当或甚至略高的蛋白质表达水平。
[0359]
这些数据证实了发明人的发现,即终止序列的添加有效地终止了rna聚合酶对相应的经修饰的dna模板的转录,因此不再需要在体外转录之前线性化包含dna模板的质粒。因此,由具有3’终止信号的超螺旋dna模板产生的mrna可以替换现有的mrna生产方法中由线性化质粒提供的mrna。
[0360]
质粒线性化步骤通常涉及用限制性内切酶孵育。因此,去除该步骤可以导致mrna生产中相当大的成本节约,特别是当大规模产生mrna作为药物产品时。
[0361]
等效物
[0362]
本领域的技术人员将认识到,或仅仅使用常规实验就能够确定本文描述的本发明
的具体实施方案的许多等同形式。本发明的范围并非旨在限于以上说明书,而是如以下权利要求书中所述。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1