AG型果胶在延寿抗衰中的应用
ag型果胶在延寿抗衰中的应用
技术领域
1.本发明属于生物医药技术领域,具体涉及ag型果胶在延寿抗衰中的应用。
背景技术:
2.衰老是一切多细胞生物随着时间的推移而自发的必然过程,伴随机体和组织的各级水平出现有害的改变,并表现出功能、适应性和抵抗力的减退,是一种生理和病理性的变化过程,具有多层次、多原因的特点。在衰老的过程中,生物体多种生理功能逐渐退化,机体应对外界应激的抵抗力逐渐下降,导致衰老相关疾病逐步增加,最终生物体走向死亡。衰老是导致许多疾病的高风险因素,包括神经退行性疾病、心血管疾病、糖尿病和癌症等。干预衰老可以改善衰老相关功能性退化,增加衰老机体抵抗环境应激的能力,延长健康寿命。
3.目前,已经报道的抗衰老药物有雷帕霉素、二甲双胍、阿卡波糖、锂等,它们在酵母、秀丽线虫、果蝇、小鼠等多种模式动物中表现出延寿抗衰以及预防衰老相关性疾病的功效。多糖是植物中含量较高的一种生物活性物质,然而多糖结构复杂、功能繁多,多糖的结构影响着其功效的发挥。已有研究表明,茯苓、黄芪、枸杞、柴胡、香菇、银耳、灵芝等中药中提取的多糖,具有免疫调节、抗辐射、抗凝血、降血糖、降血脂等功效。目前还不明确具有何种结构特征的多糖在延寿抗衰方面起着关键作用,它们通过何种途径抵抗衰老也仍不清楚。
技术实现要素:
4.本发明提供了ag型果胶在延寿抗衰中的应用,通过抑制tor信号通路,增强转录因子foxo/daf-16和nrf2/skn-1的活性,达到延寿抗衰的效果;同时该ag型果胶通过其侧链结构中的阿拉伯聚糖结构域发挥作用。
5.为解决上述技术问题,本发明提供了以下技术方案:
6.本发明提供ag型果胶在制备延寿抗衰药物中的应用,通过所述ag型果胶侧链结构中的阿拉伯聚糖结构域发挥作用。
7.本发明提供ag型果胶在制备靶向tor信号通路药物中的应用,通过所述ag型果胶侧链结构中的阿拉伯聚糖结构域发挥作用。
8.本发明提供ag型果胶在制备促进转录因子daf-16和skn-1下游靶基因表达水平提高药物中的应用,通过所述ag型果胶侧链结构中的阿拉伯聚糖结构域发挥作用。
9.优选的,所述ag型果胶来源于人参和/或三七。
10.优选的,所述ag型果胶为wgpa-1ag、pnpa-2a、wgpa-1ag-g、pnpa-2a-g、wgpa-1ag-exoa、pnpa-2a-exoa中的任意一种或多种。
11.优选的,所述ag型果胶以细胞内吞依赖的方式发挥作用。
12.优选的,所述ag型果胶浓度为0.05~1.0mg/ml。
13.与现有技术相比,本发明具有如下有益效果:
14.(1)本发明所述ag型果胶通过抑制tor信号通路,增强转录因子foxo/daf-16和
nrf2/skn-1的活性,达到延寿抗衰的效果。
15.(2)本发明利用rna干扰技术,探究出了ag型果胶以细胞内吞依赖的方式达到延寿抗衰的作用。
16.(3)本发明通过部分酸水解和果胶侧链酶对ag型果胶的降解,探究出ag型果胶侧链结构中的阿拉伯聚糖结构域是延寿抗衰的关键结构,为筛选植物多糖中具有抗衰老活性的多糖结构单元提供了新的策略,并且有利于多糖在延寿抗衰保健品和功能性食品领域的开发应用。
附图说明
17.图1人参或三七ag型果胶经四种果胶侧链酶酶解以及部分酸水解作用的产物分子量分布。
18.图2人参或三七ag型果胶经果胶侧链酶单酶解或双酶解作用的产物分子量分布。
19.图3人参或三七ag型果胶对秀丽线虫daf-16细胞核定位的影响(a:nuc为全入核、inter为部分入核、cyt为不入核,b:p值由chi-test计算,***p《0.001)。
20.图4人参或三七ag型果胶对秀丽线虫skn-1b/c在肠道细胞核积累的影响(a:high表示高入核、medium表示中度入核、low表示低入核,b:p值由chi-test计算,***p《0.001)。
21.图5人参或三七ag型果胶对daf-16和skn-1靶基因的mrna表达水平的影响(p值由student's t-test计算,**p《0.01;***p《0.001,未标注“*”即表示无显著性差异p≥0.05)。
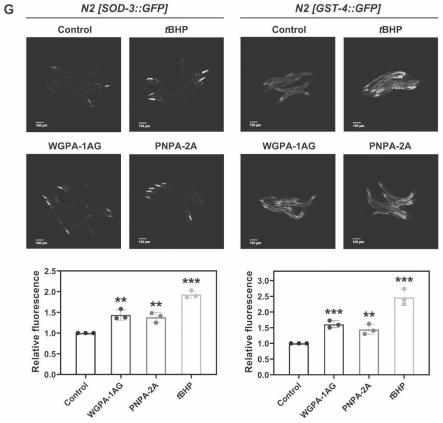
22.图6人参或三七ag型果胶对线虫sod-3和gst-4蛋白表达水平的影响(p值由student's t-test计算,**p《0.01;***p《0.001)。
具体实施方式
23.本发明提供了阿拉伯半乳聚糖(ag)型果胶在制备延寿抗衰药物中的应用,通过ag型果胶侧链结构中的阿拉伯聚糖结构域发挥作用。本发明所述ag型果胶通过靶向tor信号通路,增强转录因子forkhead box o(foxo)/daf-16和nf-e2相关因子2(nrf2)/skinhead-1(skn-1)的活性,达到延寿抗衰的效果。
24.在本发明中,所述ag型果胶来源于人参和/或三七。本发明来源人参和/或三七的ag型果胶的制备方法为:(1)将人参水提总多糖(wgp)或三七水提总多糖(pnp)分别上样于deae-纤维素柱,先用蒸馏水洗脱获得人参淀粉样葡聚糖(wgpn)和三七淀粉样葡聚糖(pnpn),再用0.5m氯化钠溶液洗脱获得人参总果胶(wgpa)和三七总果胶(pnpa)。(2)将总果胶wgpa和pnpa分别再次上样于deae-纤维素柱,先利用蒸馏水洗脱除去样品中的中性多糖,再利用0.1m、0.2m和0.3m氯化钠溶液进行梯度洗脱,获得电荷分布均一的果胶级分。(3)利用sepharose cl-6b凝胶柱,分别对这些电荷分布均一的果胶级分进行纯化,通过用0.15m氯化钠溶液洗脱,最终获得分子量分布均一的ag型果胶级分:人参ag型果胶-1(wgpa-1ag)、人参ag型果胶-2(wgpa-2ag)、三七ag型果胶-1(pnpa-1a)、三七ag型果胶-2(pnpa-2a)。本发明所述人参水提总多糖和三七水提总多糖需利用deae-纤维素柱分离纯化。本发明所述wgpa-1ag、wgpa-2ag、pnpa-1a、pnpa-2a经过透析除盐、减压浓缩和冻干获得。
25.在本发明中,所述ag型果胶为wgpa-1ag、pnpa-2a、wgpa-1ag-g、pnpa-2a-g、wgpa-1ag-exoa、pnpa-2a-exoa中的任意一种或多种。本发明所述wgpa-1ag-g和pnpa-2a-g是所述
wgpa-1ag和pnpa-2a经过聚半乳糖内切酶和半乳糖外切酶的酶解,去除其ag侧链中部分半乳糖结构后的产物;所述wgpa-1ag-exoa、pnpa-2a-exoa是所述wgpa-1ag和pnpa-2a经过阿拉伯糖外切酶的酶解,去除其ag侧链阿拉伯糖结构中的分支、末端阿拉伯糖后的产物。
26.在本发明中,所述ag型果胶依赖于细胞内吞方式发挥作用。本发明通过rnai抑制线虫内吞关键基因rab-5和rab-7表达,明显降低了wgpa-1ag和pnpa-2a对线虫寿命的延长活性,探究出ag型果胶是依赖于细胞内吞方式发挥作用的。
27.在本发明中,所述ag型果胶浓度为0.05~1.0mg/ml,优选为0.08~0.5mg/ml,更优选为0.1mg/ml。
28.在本发明中,若无特殊说明,所有的原料组分均为本领域技术人员熟知的市售商品。
29.下面将结合本发明中的实施例,对本发明中的技术方案进行清楚、完整地描述。显然,所描述的实施例仅仅是本发明一部分实施例,而不是全部的实施例。基于本发明中的实施例,本领域普通技术人员在没有做出创造性劳动前提下所获得的所有其他实施例,都属于本发明保护的范围。
30.实施例1人参或三七中ag型果胶的制备
31.(1)将人参水提总多糖和三七水提总多糖分别上样于deae-纤维素柱,先用蒸馏水洗脱获得wgpn和pnpn,再用0.5m氯化钠溶液洗脱获得wgpa和pnpa。
32.(2)将总果胶wgpa和pnpa分别再次上样于deae-纤维素柱,先利用蒸馏水洗脱除去样品中的中性多糖,再利用0.1m、0.2m和0.3m氯化钠溶液进行梯度洗脱,获得电荷分布均一的果胶级分。
33.(3)利用sepharose cl-6b凝胶柱,分别对这些电荷分布均一的果胶级分进行纯化,通过用0.15m氯化钠溶液洗脱,分别获得分子量分布均一的ag型果胶级分wgpa-1ag、wgpa-2ag、pnpa-1a、pnpa-2a。
34.(4)上述步骤中的所有多糖级分均分别经过透析除盐、减压浓缩和冻干处理后获得。
35.实施例2人参或三七中ag型果胶的结构修饰及相应级分的制备
36.(1)整体去除ag型果胶的ag侧链
37.a:取5mg实施例1制备的wgpa-1ag和pnpa-2a溶解在1ml乙酸钠缓冲液(100mm,ph4.0)中,为了尽可能降解大部分的ag侧链结构,按2u/ml的加酶量,将聚阿拉伯糖内切酶(endo-1,5-α-arabinanase,e.c.3.2.1.99)、阿拉伯糖外切酶(α-arabinofuranosidase,e.c.3.2.1.55)、聚半乳糖内切酶(endo-1,4-β-galactanase,e.c.3.2.1.89)、半乳糖外切酶(β-galactosidase,e.c.3.2.1.23)四种果胶侧链酶(爱尔兰megazyme)全部加入样品中。加酶后的样品在40℃下反应12小时,100℃加热10分钟将酶灭活,再重复酶解一次,得果胶侧链酶水解产物。
38.为了尽可能完全去除ag型果胶中的侧链结构,称取10mg实施例1制备的wgpa-1ag和pnpa-2a溶解在1ml蒸馏水中,加入等体积的0.2m三氟乙酸(tfa),100℃反应12小时,得酸水解产物。
39.b:检测果胶侧链酶水解产物和酸水解产物分子量分布的变化情况。所用仪器为:shimadzu高效凝胶液相系统(hpgpc,输液泵:lc-20ai;示差折光检测器:rid-20a;色谱柱:
tsk-gel g3000pw
xl
、7.8
×
300mm,日本岛津)。结果见图1。
40.c:将果胶侧链酶水解产物和酸水解产物离心,取上清,加入3倍体积的无水乙醇(75%醇沉),反复震荡数次后离心取沉淀,以此方法除去样品中的寡糖和单糖。加入蒸馏水将沉淀复溶,除盐冻干,获得酶解和酸水解后样品中剩余的多糖部分。其中果胶侧链酶水解级分为wgpa-1ag-a-g和pnpa-2a-a-g;酸水解级分为wgpa-1ag-tfa和pnpa-2a-tfa。
41.d:通过pmp柱前衍生化和shimadzu高效液相色谱系统(hplc,输液泵:lc-20at;紫外检测器:spd-20a;反相c18色谱柱:cosmosil5c
18-paq、4.6
×
250mm,日本岛津)分析上述所有酶解、酸水解级分的单糖组成。结果见表1。
42.从图1可以看出,wgpa-1ag和pnpa-2a经四种果胶侧链酶水解后,大部分高分子量的果胶结构(保留时间在9-12分钟)被降解为低分子量的寡糖和单糖,这些低聚物在保留时间14-16分钟处被洗脱出来,但仍有少量的高分子量果胶结构得以保留,出峰时间在11分钟左右。部分酸水解后的果胶样品几乎不含有高分子量果胶结构,洗脱图谱在保留时间12-14分钟处出现一个连续不对称的宽峰,推测其为带有不同聚合度的阿拉伯糖及半乳糖侧链结构的中分子量rg-i片段。
43.表1人参和三七ag型果胶酶解或酸水解级分的单糖组成
[0044][0045]
从表1可以看出,wgpa-1ag-a-g和pnpa-2a-a-g的单糖组成均以半乳糖醛酸和鼠李糖为主,另外样品中还含有约40%的半乳糖及少量的阿拉伯糖,表明使用四种果胶侧链酶水解可除去人参或三七ag型果胶侧链中的大部分阿拉伯糖结构,但还有一部分半乳糖结构连接在rg-i主链上。与果胶侧链酶水解级分相比,wgpa-1ag-tfa和pnpa-2a-tfa中的半乳糖含量明显下降,半乳糖醛酸和鼠李糖的含量可达70%以上,说明通过部分酸水解法可除去人参或三七ag型果胶中的更多ag侧链结构。通过单糖组成的结果计算可知,四种果胶侧链酶的组合酶解使人参或三七ag型果胶中约75%的ag侧链结构被降解,而tfa水解可除去ag型果胶中》90%的ag侧链结构。
[0046]
(2)分别去除ag型果胶中的不同侧链结构域
[0047]
a:与第(1)部分步骤a中区别在于加入的酶为聚阿拉伯糖内切酶和阿拉伯糖外切酶,降解侧链中的大部分阿拉伯糖结构,其余步骤均相同。获得的多糖级分为wgpa-1ag-a和pnpa-2a-a。
[0048]
b:与第(1)部分步骤a中区别在于加入的酶为聚半乳糖内切酶和半乳糖外切酶,降
解侧链中β-d-1,4-半乳糖结构,其余步骤均相同。获得的多糖级分为wgpa-1ag-g和pnpa-2a-g。
[0049]
c:与第(1)部分步骤a中区别在于加入的酶为聚阿拉伯糖内切酶,降解侧链中的阿拉伯聚糖结构域,其余步骤均相同。获得的多糖级分为wgpa-1ag-endoa和pnpa-2a-endoa。
[0050]
d:与第(1)部分步骤a中区别在于加入的酶为阿拉伯糖外切酶,降解侧链结构中的分支及末端阿拉伯糖,其余步骤均相同。获得的多糖级分为wgpa-1ag-exoa和pnpa-2a-exoa。
[0051]
通过第(1)部分步骤b~d的方法检测各酶解产物分子量分布的变化和各多糖级分的单糖组成。结果见图2和表2。
[0052]
从图2可以看出,无论是经过两种果胶侧链酶的组合酶解,还是单独加入聚阿拉伯糖内切酶/阿拉伯糖外切酶进行酶解,样品的分子量均降低。单糖组成结果表明,wgpa-1ag-a和pnpa-2a-a中仅剩余少量的阿拉伯糖(《10%),表明使用聚阿拉伯糖内切酶和阿拉伯糖外切酶的组合酶解法除去了果胶样品中大部分的阿拉伯糖结构;使用聚半乳糖内切酶和半乳糖外切酶的组合酶解法获得的级分wgpa-1ag-g和pnpa-2a-g中半乳糖的含量与未酶解原样相比有一定程度的下降,但它们仍含有一定量的半乳糖;聚阿拉伯糖内切酶水解级分wgpa-1ag-endoa、pnpa-2a-endoa和阿拉伯糖外切酶水解级分wgpa-1ag-exoa和pnpa-2a-exoa中的阿拉伯糖含量与原样相比均有所下降。
[0053]
表2人参或三七ag型果胶各酶解级分的单糖组成
[0054][0055]
从表2可以看出,两种阿拉伯糖苷酶水解除去了人参或三七ag型果胶中90%以上的阿拉伯糖结构;两种半乳糖苷酶水解除去了约30-40%左右的半乳糖结构。而根据酶的特性可知,聚阿拉伯糖内切酶主要除去人参或三七ag型果胶中的阿拉伯聚糖结构域,而阿拉伯糖外切酶主要除去ag型果胶中的分支以及末端阿拉伯糖。
[0056]
实施例3人参或三七中纯化的各多糖级分对秀丽线虫寿命及氧化压力耐受性的影响
[0057]
1、不同浓度各人参多糖级分对秀丽线虫氧化压力耐受性的影响
[0058]
(1)配制含9.125mm的叔丁基过氧化氢(tbhp)(sigma购买)的固体线虫生长培养基(ngm),在超净工作台中吹干待用。
[0059]
(2)用蒸馏水将各多糖级分(各多糖级分种类见表3,实施例1制备得到)配制成5mg/ml的溶液,在超净工作台中使用0.22μm过滤器抽滤,再与大肠杆菌op50菌液分别按1:99、1:49、1:9、1:4混合,使各多糖级分的终浓度为0.05mg/ml、0.1mg/ml、0.5mg/ml、1.0mg/ml。将分别含有四个浓度的各多糖级分的混合菌液均匀铺在步骤(1)配制的ngm培养基上,在超净工作台中过夜吹干,即得四个不同浓度各多糖级分的实验组ngm培养基。对照组为将各多糖溶液替换为等体积的蒸馏水,再按上述步骤铺菌、晾干,即得对照组培养基。
[0060]
(3)将day1时期野生型成虫(n2 bristol,来源caenorhabditis genetics center(cgc))分别转移至步骤(2)制备的培养基上培养,温度20℃。并每小时统计一次线虫的死亡数目,直至线虫全部死亡。
[0061]
(4)线虫死亡的鉴定标准:用铂丝轻轻拨动线虫,若线虫没有反应,即认为其死亡。线虫钻入培养基底部、爬至培养基边缘以及wormbag的情况均视为丢失,不计入数据统计。所有寿命实验至少重复三次,结果以平均寿命
±
sem表示,显著性差异通过log-rank检验计算。
[0062]
表3不同浓度人参各多糖级分对秀丽线虫氧化压力耐受性的影响
[0063][0064][0065]
从表3可以看出,以上人参各多糖级分在0.1mg/ml的浓度条件下均表现出良好的
抗氧化活性。
[0066]
2、人参或三七各多糖级分对秀丽线虫寿命的影响
[0067]
(1)用蒸馏水将各多糖级分(各多糖级分种类见表4,实施例1制备得到)配制成1mg/ml的溶液,在超净工作台中使用0.22μm过滤器抽滤,再与大肠杆菌op50菌液分别按1:9混合,使各多糖级分的终浓度为0.1mg/ml。将配制好的各多糖级分菌液均匀铺在线虫生长培养基(ngm)上,在超净工作台中过夜吹干,即得四个不同浓度各多糖级分的实验组ngm培养基。对照组为将各多糖溶液替换为等体积的蒸馏水,再按上述步骤铺菌、晾干,即得对照组培养基。
[0068]
(2)同期化处理的野生型线虫在实验组和对照组的培养基上培养,温度20℃。记录每日死亡的线虫数目,并将剩余存活的线虫转入新鲜ngm培养基中,直至线虫全部死亡。
[0069]
(3)线虫死亡的鉴定标准:同该实施例第1部分步骤(4)。
[0070]
表4人参或三七各多糖级分对秀丽线虫寿命的影响
[0071][0072]
3、人参或三七各多糖级分对秀丽线虫氧化压力耐受性的影响
[0073]
与该实施例步骤1的区别在于多糖级分种类(多糖级分种类见表5,实施例1制备得到)、多糖级分的终浓度为0.1mg/ml,其余步骤均相同。
[0074]
表5人参或三七各多糖级分对秀丽线虫氧化压力耐受性的影响
[0075][0076][0077]
从表4-5可以看出,人参或三七水提总多糖wgp和pnp具有延寿抗衰作用,且从两种总多糖中分离得到的总果胶wgpa和pnpa是主要的延寿作用组分,并进一步发现从wgpa和pnpa中分离得到的电荷、分子量分布均一的ag型果胶wgpa-1ag、pnpa-1a和pnpa-2a是人参或三七多糖中发挥延寿抗衰作用的关键组分。
[0078]
4、人参或三七各ag果胶级分对秀丽线虫寿命的影响
[0079]
与该实施例步骤2的区别在于ag果胶级分种类(ag果胶级分种类见表6,实施例1和实施例2制备得到),其余步骤均相同。
[0080]
表6人参或三七各果胶级分对秀丽线虫寿命的影响
[0081]
[0082][0083]
注:a代表的p值为与对照组比较求出的值,b和c代表的p值为分别与wgpa-1ag或pnpa-2a比较求出的值。
[0084]
5、人参或三七各ag果胶级分对秀丽线虫氧化压力耐受性的影响
[0085]
与该实施例步骤3的区别在于ag果胶级分种类(ag果胶级分种类见表7,实施例1和实施例2制备得到),其余步骤均相同。
[0086]
表7人参或三七各ag果胶级分对秀丽线虫氧化压力耐受性的影响
[0087]
[0088]
[0089][0090]
注:a代表的p值为与对照组比较求出的值,b和c代表的p值为分别与wgpa-1ag或pnpa-2a比较求出的值。
[0091]
从表6-7可以看出,通过四种果胶侧链酶的酶解和酸水解去除ag侧链结构的wgpa-1ag-a-g、pnpa-2a-a-g、wgpa-1ag-tfa和pnpa-2a-tfa,其对线虫寿命的延长作用以及对氧化胁迫耐受能力的增强作用明显低于wgpa-1ag和pnpa-2a。而且几乎完全去除ag侧链结构的wgpa-1ag-tfa和pnpa-2a-tfa不具有延寿抗衰的作用。这说明,ag侧链结构主要决定了wgpa-1ag和pnpa-2a的延寿抗衰作用。
[0092]
wgpa-1ag-a和pnpa-2a-a是wgpa-1ag和pnpa-2a经过聚阿拉伯糖内切酶和阿拉伯糖外切酶的酶解,去除其ag侧链中阿拉伯糖结构后的产物。wgpa-1ag-g和pnpa-2a-g是wgpa-1ag和pnpa-2a经过聚半乳糖内切酶和半乳糖外切酶的酶解,去除其ag侧链中部分半乳糖结构后的产物。与wgpa-1ag和pnpa-2a的作用相比,wgpa-1ag-a和pnpa-2a-a对线虫寿命的延长作用及其对氧化胁迫耐受能力的增强作用明显下降。但是wgpa-1ag-g、pnpa-2a-g延长线虫寿命及提高其抗氧化能力的效果与wgpa-1ag和pnpa-2a无明显差异。这说明阿拉伯糖结构是wgpa-1ag和pnpa-2a的延寿抗衰作用的关键结构。
[0093]
wgpa-1ag-endoa和pnpa-2a-endoa是wgpa-1ag和pnpa-2a经过聚阿拉伯糖内切酶的酶解,去除其ag侧链阿拉伯糖结构中的阿拉伯聚糖结构域后的产物。wgpa-1ag-exoa和pnpa-2a-exoa是wgpa-1ag和pnpa-2a经过阿拉伯糖外切酶的酶解,去除其ag侧链阿拉伯糖结构中的阿拉伯糖分支、末端后的产物。与wgpa-1ag和pnpa-2a的作用相比,wgpa-1ag-endoa和pnpa-2a-endoa对线虫寿命的延长作用及其对氧化胁迫耐受能力的增强作用明显下降,而wgpa-1ag-exoa和pnpa-2a-exoa对线虫的延寿抗衰作用与wgpa-1ag和pnpa-2a的作用无明显差异。这说明阿拉伯聚糖结构域是wgpa-1ag和pnpa-2a的延寿抗衰作用的关键结构。
[0094]
6、人参或三七ag型果胶延寿抗衰依赖于tor信号通路的研究
[0095]
将wgpa-1ag、pnpa-2a分别喂食tor信号通路的功能缺失突变品系线虫rsks-1(ok1255)(rb1206,来源于caenorhabditis genetics center(cgc))和iis信号通路的功能缺失突变品系线虫daf-2(e1370)(cb1370,来源于caenorhabditis genetics center(cgc)),然后进行常规寿命和氧化胁迫耐受实验(实验步骤同该实施例步骤2和3)。
[0096]
将wgpa-1ag和pnpa-2a分别喂食daf-16(mu86)(cf1038,来源于caenorhabditis genetics center(cgc))和skn-1(zu135)(lg340,来源于caenorhabditis genetics center(cgc))突变体线虫,再进行常规寿命和氧化应激耐受实验(实验步骤同该实施例步骤2和3)。
[0097]
表8人参或三七ag型果胶对突变体秀丽线虫寿命的影响
[0098][0099][0100]
从表8可以看出,wgpa-1ag、pnpa-2a不能延长rsks-1(ok1255)的寿命和提高其对氧化胁迫的耐受能力,但是可以显著延长daf-2(e1370)的寿命,这些结果表明,人参、三七ag型果胶(wgpa-1ag、pnpa-2a)的延寿抗衰作用依赖于tor信号通路,不依赖于iis信号通路。
[0101]
从表9可以看出,wgpa-1ag、pnpa-2a几乎不能延长daf-16(mu86)和skn-1(zu135)突变线虫的寿命,也几乎不能提高他们的抗氧化能力。这表明wgpa-1ag和pnpa-2a的延寿抗
衰作用依赖于转录因子daf-16和skn-1。
[0102]
7、人参或三七ag型果胶延寿抗衰依赖于细胞内吞的研究
[0103]
(1)rab-5和rab-7rnai条件下的寿命检测实验:用蒸馏水将各ag型果胶分别配制成1mg/ml的溶液,在超净工作台中使用0.22μm滤器抽滤,再与含有相应dsrna质粒(rab-5和rab-7)的大肠杆菌ht115菌液按1:9混合(对照组混合等体积蒸馏水),使各ag型果胶的终浓度均为0.1mg/ml。将含有各ag型果胶的菌液分别均匀铺在已吹干的rnai培养基上,在超净工作台中过夜吹干,即可作为实验组培养基使用。铺菌和晾干的过程应全程避光。将同期化处理的野生型线虫(n2)在普通的实验组和对照组ngm培养基上培养至l4期,再转入相应的实验组/对照组rnai培养基中,该天记为day0。从第二天开始,每组分别随机挑取约90条线虫转入相应的60mm对照组/实验组rnai培养基,每盘30-45条,并每日转板。对线虫死亡数目的数据统计与常规寿命实验一致。(数据统计同该实施例步骤2)。
[0104]
(2)抗氧化胁迫耐受实验:与上述步骤(1)的区别在于rnai培养基和对照组培养基中加入9.125mm的叔丁基过氧化氢,其余均相同。
[0105]
表10人参或三七ag型果胶在rnai(rab-5和rab-7)条件下对线虫寿命的影响
[0106][0107]
表11人参或三七ag型果胶在rnai(rab-5和rab-7)条件下对线虫抗氧化胁迫能力的影响
[0108][0109]
从表10-11可以看出,通过rnai抑制线虫内吞关键基因rab-5和rab-7的表达后,wgpa-1ag、pnpa-2a不再延长野生型n2线虫的寿命,也不能再提高其抗氧化胁迫能力。说明wgpa-1ag、pnpa-2a的延寿抗衰作用依赖于细胞内吞作用。
[0110]
实施例4转录因子的核定位分析ag型果胶延寿抗衰的机制
[0111]
(1)对转基因线虫daf-16::gfp(tj356,zls356,来源caenorhabditis genetics center(cgc))和skn-1::gfp(ld1,ldls7,来源caenorhabditis genetics center(cgc))喂食wgpa-1ag、pnpa-2a作为实验组,未喂食的作为对照组。喂食步骤同实施例3步骤2。
[0112]
(2)用1%甲醛固定实验组和对照组的转基因线虫daf-16::gfp(tj356),用2mm左旋咪唑固定实验组和对照组的转基因线虫skn-1b/c::gfp(ld1)。然后在激光共聚焦显微镜(德国carl zeiss)下观察gfp表达情况,并在同一曝光条件下进行拍照。对于daf-16::gfp和肠道中skn-1b/c::gfp在细胞核中的定位模式定义如下:“nuc”或“high”是指在所有细胞核中观察到较强的gfp信号。“inter”或“medium”是指在所有细胞核中观察到较弱的gfp信号,或仅在线虫头部或尾部观察到高水平的gfp信号。“cyt”或“low”表示在所有细胞核中几乎都检测不到gfp信号。
[0113]
由图3和图4可知,与对照组相比,不管是喂食wgpa-1ag还是喂食pnpa-2a后,daf-16::gfp的“nuc”模式、以及skn-1b/c::gfp的“high”模式出现的比例都有明显的增加。说明
喂食wgpa-1ag、pnpa-2a明显促进了线虫转录因子daf-16和skn-1的细胞核定位。
[0114]
实施例5rna提取和实时荧光定量pcr(rt-qpcr)分析ag型果胶延寿抗衰的机制
[0115]
(1)对野生型n2线虫喂食wgpa-1ag、pnpa-2a作为实验组,未喂食的作为对照组。喂食步骤同实施例3步骤2。
[0116]
(2)将分别喂食wgpa-1ag、pnpa-2a的野生型n2线虫,以及对照组n2线虫(果胶替换为蒸馏水)培养至成虫。使用trizol试剂盒(日本takara)提取上述线虫的总rna,并使用逆转录酶(美国promega)获得cdna。rt-qpcr反应是通过利用预混tb green premix ex taq酶混合物(日本takara),于quantstudio 3实时荧光定量pcr仪(美国thermo)进行的。act-1作为内参基因。
[0117]
如图5所示,通过rt-qpcr检测了daf-16靶基因sod-3、mtl-1、lys-7、hsp-16.2,以及skn-1靶基因gcs-1、gst-4、gst-7、gst-10的mrna表达水平。结果表明,与对照组相比,喂食了wgpa-1ag和pnpa-2a后,野生型线虫中daf-16靶基因sod-3和lys-7,以及skn-1靶基因gcs-1、gst-4、gst-7、gst-10的mrna表达水平明显提高。这说明人参、三七ag型果胶wgpa-1ag和pnpa-2a激活了转录因子daf-16和skn-1的转录活性。
[0118]
实施例6 sod-3和gst-4蛋白表达量检测分析ag型果胶延寿抗衰的机制
[0119]
(1)对转基因线虫sod-3::gfp(cf1553,muis84,来源caenorhabditis genetics center(cgc))、gst-4::gfp(cl2166,dvis19,来源caenorhabditis genetics center(cgc))喂食wgpa-1ag、pnpa-2a作为实验组,未喂食的作为对照组。喂食步骤同实施例3步骤2。
[0120]
(2)在上述两种转基因品系线虫的对照组中取出一部分线虫,在9.125mm叔丁基过氧化氢处理的条件下培养2小时,作为阳性对照组。用1%甲醛固定实验组和对照组的转基因线虫sod-3::gfp(cf1553)、gst-4::gfp(cl2166)。然后在激光共聚焦显微镜(德国carlzeiss)下观察gfp表达情况,并在同一曝光条件下进行拍照。
[0121]
(3)使用多功能酶标仪(瑞士tecan)对各实验组/对照组sod-3::gfp和gst-4::gfp的荧光表达进行定量分析,激发光的波长为488nm,发射光的波长为535nm。
[0122]
如图6所示,结合荧光蛋白细胞定位技术并利用多功能酶标仪,对没有喂食果胶的对照组线虫,和喂食了wgpa-1ag、pnpa-2a的实验组线虫sod-3::gfp和gst-4::gfp荧光蛋白进行了定量分析。结果表明,与对照组相比,在喂食wgpa-1ag和pnpa-2a后,线虫中sod-3和gst-4的蛋白表达水平明显提高。进一步证明人参、三七ag型果胶wgpa-1ag和pnpa-2a激活了转录因子daf-16和skn-1的转录活性,使其下游靶基因的表达水平明显提高。
[0123]
以上所述仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1