DNA的合成的制作方法
领域
本发明涉及用于合成dna、rna、蛋白质及类似分子的改进的方法,尤其是dna的无细胞酶促合成,优选以大的规模。
背景
脱氧核糖核酸(dna)的扩增可以通过使用基于细胞的方法来进行,例如通过在发酵罐中培养增殖待扩增的dna的细菌。也已经描述了用于从起始模板扩增dna的无细胞酶促方法,包括聚合酶链反应和链置换反应。
过去,已使用基于微孔板和自动控制的移液管的装置添加所需的反应成分来实施测试规模的dna扩增。这样的装置和方法适用于生产少量的dna分子以用于测试目的,但不提供足够的用于其他目的的量。特定核酸和蛋白质的大规模扩增和生产大部分是通过基于细胞的方法进行的。这样的方法通常有效地生产非常大体积的产物,但其设置是昂贵的。
也有很多可用的装置,其特别适应于使用热循环(thermocyclic)方法引起聚合酶链反应(pcr)来扩增dna样品。这些装置理想地适于该反应,但其是不灵活的,不能适应于实施其他反应。这种装置的实例公开在us8,163,489中。
已知使用化学合成(如亚磷酰胺法)的大规模dna合成,但其并非没有缺点。反应通常必须在有机溶剂中进行,这些有机溶剂中的许多是有毒的或者有害的。化学合成的另一缺点是,其并非完全有效率,因为在每次添加核苷酸之后,至少2%的生长中的寡核苷酸链被封端,导致产率损失。因此,被合成的核苷酸链的总产率损失随着向序列中添加的每个核苷酸而增加。化学合成寡核苷酸的这种固有的无效率性最终将能够有效生产的寡核苷酸的长度限制在具有50个核酸残基或以下的寡核苷酸。
到目前为止,还未常规地研究将生物催化剂,例如聚合酶,用于工业规模地生产dna产物,且反应在很大程度上被限制于微升规模的体积。使用dna的酶促合成的放大方法已被证明是有问题的,尤其是dna产物令人失望的产率。
因此,存在对能够用于以显著的规模合成dna及类似分子的方法的需要。
概述
本发明涉及涉及用于无细胞生产dna的方法。该方法可以允许与目前的涉及封闭的分批过程的方法相比增强的dna生产。这显著增加生产率,同时降低合成dna的成本,尤其是以大的规模。本发明一般性地涉及扩增dna的等温方法,其不要求通过加热和冷却使温度循环。
因此,提供了一种用于合成dna的无细胞方法,包括使dna模板与至少一种聚合酶在一种或多种引物和核苷酸的存在下、在促进所述模板扩增的条件下接触,以形成反应混合物,所述模板扩增是通过另一条链的链置换复制来置换经复制的链,其中以受控的方式向反应混合物供给另外的核苷酸(furthernucleotides)。
因此,无细胞方法涉及模板经由链置换复制的扩增。提供了一种用于合成dna的无细胞方法,包括使dna模板与至少一种聚合酶在一种或多种引物和核苷酸的存在下、在促进所述模板通过链置换复制而扩增的条件下接触,以形成反应混合物,其中以受控的方式向反应混合物供给另外的核苷酸。
因此,还提供了一种用于合成dna的无细胞方法,包括使dna模板与至少一种聚合酶在核苷酸的存在下、在促进所述模板扩增的条件下接触,以形成反应混合物,所述模板扩增是通过另一条链的链置换复制来置换经复制的链,其中以受控的方式向反应混合物供给另外的核苷酸。
因此,无细胞方法涉及模板经由链置换复制的扩增。提供了一种用于合成dna的无细胞方法,包括使dna模板与至少一种聚合酶在核苷酸的存在下、在促进所述模板通过链置换复制而扩增的条件下接触,以形成反应混合物,其中以受控的方式向反应混合物供给另外的核苷酸。
因此,向反应混合物受控地供给另外的核苷酸是有利的,因为这种合理的控制策略能够允许改进dna的合成速率和/或改进dna产率。这些改进是与其中在开始时向反应混合物供给所有核苷酸的类似的反应混合物进行比较。另外的核苷酸的受控供给可以允许增加生产率和/或聚合酶的持续合成能力。
或者,提供了一种用于合成dna的无细胞方法,包括使dna模板与至少一种聚合酶在一种或多种引物、核苷酸和金属阳离子的存在下、在促进所述模板扩增的条件下接触,以形成反应混合物,所述模板扩增是通过另一条链的链置换复制来置换经复制的链,其中以受控的方式向反应混合物供给另外的金属阳离子。
因此,无细胞方法涉及模板经由链置换复制的扩增。提供了一种用于合成dna的无细胞方法,包括使dna模板与至少一种聚合酶在一种或多种引物、核苷酸和金属阳离子的存在下、在促进所述模板通过链置换复制而扩增的条件下接触,以形成反应混合物,其中以受控的方式向反应混合物供给另外的金属离子。
或者,提供了一种用于合成dna的无细胞方法,包括使dna模板与至少一种聚合酶在核苷酸和金属阳离子的存在下、在促进所述模板扩增的条件下接触,以形成反应混合物,所述模板扩增是通过另一条链的链置换复制来置换经复制的链,其中以受控的方式向反应混合物供给另外的金属阳离子。
因此,无细胞方法涉及模板经由链置换复制的扩增。提供了一种用于合成dna的无细胞方法,包括使dna模板与至少一种聚合酶在核苷酸和金属阳离子的存在下、在促进所述模板通过链置换复制而扩增的条件下接触,以形成反应混合物,其中以受控的方式向反应混合物供给另外的金属离子。
因此,向反应混合物受控地供给另外的金属阳离子是有利的,因为这种合理的控制策略能够允许改进dna的合成速率。这些改进是与其中在开始时向反应混合物供给所有金属阳离子的类似的反应混合物进行比较。另外的金属阳离子的受控供给可以允许增加生产率和/或聚合酶的持续合成能力。
还提供了一种用于合成dna的无细胞方法,包括使dna模板与至少一种聚合酶在一种或多种引物、核苷酸和金属阳离子的存在下、在促进所述模板扩增的条件下接触,以形成反应混合物,所述模板扩增是通过另一条链的链置换复制来置换经复制的链,其中以受控的方式向反应混合物供给另外的核苷酸和金属阳离子。
因此,无细胞方法涉及模板经由链置换复制的扩增。提供了一种用于合成dna的无细胞方法,包括使dna模板与至少一种聚合酶在一种或多种引物、核苷酸和金属离子的存在下、在促进所述模板通过链置换复制而扩增的条件下接触,以形成反应混合物,其中以受控的方式向反应混合物供给另外的核苷酸和另外的金属阳离子。
还提供了一种用于合成dna的无细胞方法,包括使dna模板与至少一种聚合酶在核苷酸和金属阳离子的存在下、在促进所述模板扩增的条件下接触,以形成反应混合物,所述模板扩增是通过另一条链的链置换复制来置换经复制的链,其中以受控的方式向反应混合物供给另外的核苷酸和金属阳离子。
因此,无细胞方法涉及模板经由链置换复制的扩增。提供了一种用于合成dna的无细胞方法,包括使dna模板与至少一种聚合酶在核苷酸和金属离子的存在下、在促进所述模板通过链置换复制而扩增的条件下接触,以形成反应混合物,其中以受控的方式向反应混合物供给另外的核苷酸和另外的金属阳离子。
另外的核苷酸和另外的金属阳离子可以一起或单独供给。优选的是,核苷酸和金属离子在被供给至反应混合物之前不混合在一起。另外的核苷酸和另外的金属阳离子可以在各自被需要时彼此独立地添加至反应混合物。
因此,向反应混合物受控地供给另外的核苷酸和另外的金属阳离子是有利的,因为这种合理的控制策略能够允许改进dna的合成速率和/或改进的dna产率。这些改进是与其中在开始时向反应混合物供给所有核苷酸和金属阳离子的类似的反应混合物进行比较。另外的核苷酸的受控供给可以允许增加生产率和/或聚合酶的持续合成能力。
根据本发明的每个若干另外的方面,提供了被配置为(configuredto)实施根据本发明的第一、第二和第三方面的方法的装置。该装置可以用于实施根据本发明的任一方面的方法。
根据本发明的另一方面,dna模板可以包含一个或多个加工酶靶位点。可以使反应混合物在任何时间与一种或多种加工酶接触。可以以受控的方式向所述反应混合物提供所述酶。优选地,加工酶是原核端粒酶(protelomerase)。
独立地,本发明的任一方面的任选特征可以是:dna模板可以是环状的。所述dna模板的链置换扩增可以通过滚环扩增(rca)来进行。dna聚合酶可以是phi29或其变体。dna的扩增可以是等温扩增。一种或多种引物可以是随机引物。合成的dna可以包含多联体,所述多联体包含由dna模板扩增的dna序列的串联单元。dna模板可以是封闭的线状dna;优选地,将dna模板在变性条件下孵育,以形成封闭的环状的单链dna。dna模板可以包含至少一个加工酶靶序列,优选重组酶靶序列或原核端粒酶靶序列。dna模板可以包含表达盒,所述表达盒包含有效地与目标编码序列连接的真核生物启动子,以及任选的真核转录终止序列。表达盒可以在任一侧侧接(flank)有加工酶靶序列,优选重组酶靶序列或原核端粒酶靶序列。扩增的dna在适当时可以与重组酶或原核端粒酶接触。
可以合成的dna的量等于或高于1克每升反应混合物。
可以以多个等份向反应混合物供给另外的核苷酸和/或另外的金属离子。可以在整个过程中以有规律的间隔,任选至少每30分钟,供给另外的核苷酸和/或另外的金属离子。可以向反应混合物供给至少3、4、5、6、7、8、9或10份。可以以连续的方式向反应混合物供给另外的核苷酸和/或另外的金属离子。另外的金属离子和/或另外的核苷酸可以任选地通过使用泵而由外部来源进料至反应混合物,或者通过使用渗透泵供给至反应混合物。
核苷酸或另外的核苷酸可以包含生物上无活性的核苷酸;任选地,通过活化(优选通过化学或物理方式)而将这些生物上无活性的核苷酸供给至反应混合物。生物上无活性的核苷酸可以是通过光解而活化的笼锁的(caged)核苷酸。
可以响应于与反应混合物中dna的浓度相关的信号而供给另外的核苷酸和/或另外的金属离子和/或加工酶,其可以任选地通过测量反应混合物或其可能产生信号的部分的压力的差别来监测。可以通过测量由去除和返回反应混合物的一部分的循环泵任选地在压力传感器上产生的压力差来产生指示dna浓度的信号。
金属阳离子或另外的金属阳离子可以包含一种或多种选自下列的金属:mg2+、be2+、ca2+、sr2+、li+、na+、k+、mn2+或zn2+,优选为mg2+。另外的金属阳离子可以与另外的核苷酸一起供给。反应混合物中的金属阳离子和核苷酸之间的比例可以维持在约3:1。本文所讨论的金属离子的浓度,尤其是游离金属离子浓度,涉及所关注的特定金属阳离子(即,mg2+),而不是反应混合物中所有种类的金属离子的浓度。
反应混合物可以含有至少一种核苷酸。然后可以添加一种或多种另外的核苷酸。核苷酸或另外的核苷酸为脱氧核苷三磷酸(dntp)或其衍生物或修饰的形式。核苷酸或另外的核苷酸是脱氧腺苷三磷酸(datp)、脱氧鸟苷三磷酸(dgtp)、脱氧胞苷三磷酸(dctp)、脱氧胸苷三磷酸(dttp)及其衍生物中的一种或多种。核苷酸或另外的核苷酸以游离酸、其盐或其螯合物中的一种或多种的形式提供。盐或螯合物可以包括一种或多种以下金属离子:mg2+、be2+、ca2+、sr2+、li+、na+、k+、mn2+或zn2+。反应混合物中核苷酸的浓度可以保持在0.001mm至6mm,任选的为约3mm。
方法可以是分批法或连续流法。另外任选的特征描述在从属权利要求中。
另外的优点描述在下文中。
附图的简要说明
在下文中,会参考示例性实施方式和附图来进一步描述本发明,其中:
图1是可用于根据本发明的实施方式的合成装置的透视图。为了易于理解,以及使主要部件更可视,在该图中省略了某些部件和连接管路;
图2是图1的装置的示意图;
图3是可用于根据本发明的实施方式的反应容器的横截面图;
图4是显示仅用水的对照反应的差别压力数据(方块)与由使用phi29dna聚合酶的标准dna扩增反应产生的数据(菱形)随时间变化(如表1所示1)的数据对比的图。压力差的单位是随意的,且时间以分钟为单位;
图5是压力差相对于通过
图6是dna扩增反应中共价封闭的线状dna产物(2.6k碱基和2k碱基)的电泳凝胶的照片,其中起始的dntp浓度为2mm、4mm和8mm;
图7是对于以下反应,差别压力测量相对于时间(分钟)的绘图:2mmdntp的起始浓度(未添加另外的dntp)(圆圈)、2mmdntp进料和另外的2mmdntp(三角形)、起始时4mmdntp(未添加另外的dntp)(菱形)和2mmdntp进料和三次2mm浓度的另外的dntp(叉)。箭头指明每个反应中dntp添加的时间;
图8是对于以下反应,起始反应速率(差别压力单位/min)相对于镁离子与dntp的比例的绘图:起始浓度为2mmdntp的进料和另外3次添加2mmdntp;
图9描绘了protlx-n3x2cal09hadna模板;
图10描绘了protlxb5x4cal09hadna模板;
图11描绘了protlx-kn3x2ovadna模板;
图12描绘了protlx-kn3x2luxdna模板;和
图13描绘了对于以下反应,差别压力测量相对于时间(分钟)的绘图:初始含有2mmdntp和7.5mmmg2+,在指明的点(300分钟、600分钟、900分钟、1080分钟、1230分钟和1440分钟–白色三角形)受控地添加(“进料”)核苷酸和mg2+,其中添加2mmdntp和足量的mg2+以在第一次进料后维持3:1的mg2+:dntp比例),如所指明地(方块)在1050分钟受控地向其添加原核端粒酶(2μmteln)。箭头指明每次dntp添加随后立即添加mg2+以及添加teln的时间。
详细描述
本发明涉及用于合成dna的无细胞方法。本发明的方法可以允许高生产量地合成dna。
根据本发明所合成的脱氧核糖核酸(dna)可以是任何dna分子。dna可以是单链的或双链的。dna可以是线状的。dna可以被加工,以形成环,尤其是微环、单链闭环、双链闭环、双链开环或封闭的线性双链dna。dna可以被允许形成或被加工而形成特定的二级结构,例如但不限于发夹环(茎环)、不完美型发夹环、假结体或各种类型的双螺旋(a-dna、b-dna或z-dna)中的任一种。dna还可以形成发夹。
所合成的dna可以是任何适当的长度。使用本发明的方法,高达或超过77千碱基(kb或k碱基)的长度是可能的。更具体的,根据该方法可以合成的dna的长度可以是高达60千碱基、或高达50千碱基、或高达40千碱基、或高达30千碱基的量级。优选地,所合成的dna可以是100碱基至60千碱基、200碱基至20千碱基,更优选200碱基至15千碱基,最优选2千碱基至15千碱基。
根据本发明的方法所合成的dna的量可以超过10g/l反应混合物。优选地,所合成的dna的量为高达10、9、8、7、6或5g/l。所合成的dna的优选的量为0.2至5g/l,优选为1至5g/l。所生产的dna的量可以被描述为工业或商业的量、以大的规模、或大规模生产。通过该方法生产的dna在质量方面,即dna长度和序列方面可以是均一的。因此,该方法可以适于大规模合成dna。
在目前可用的合成dna的技术中,大部分是小的或基于实验室的规模,反应以微升的体积而发生。放大pcr的努力仍然导致很差的dna产率,例如约20-40mg每升反应混合物(vandalia),因为反应的发生仅用起始量的核苷酸,而未供给另外的核苷酸,且产物的形成抑制该反应。
dna模板可以是任何适当的模板。dna模板可以是单链的(ss)或双链的(ds)。dna模板可以是线性的或环状的。双链的dna模板可以是开环的dsdna、闭环的dsdna、开放的线性dsdna或封闭的线性dsdna。单链的dna模板可以是线性的ssdna或闭环的ssdna。后面的dna模板可以通过使封闭的线性dsdna变性来生产。封闭的线性dna,即线性ds共价封闭的dna分子,通常包含共价封闭的末端,即发夹末端,该处不存在互补的dna链之间的碱基对。发夹环使互补的dna链的末端结合在一起。该环本身可以含有互补的序列,特别是如果该环包含原核端粒酶靶序列的一部分时。封闭的线性dsdna可以是质粒。
dna模板可以包含任何序列,无论是天然起源的或人工的。dna模板可以包含至少一种加工酶靶序列,例如一个、两个、三个、四个或更多个加工酶靶点。这样的靶序列允许dna在合成后任选地被进一步加工。加工酶是识别其靶位点并加工dna的酶。加工酶靶序列可以是用于限制性酶的靶序列。限制性酶,即限制性内切酶,与靶序列结合并在特定位点断裂。加工酶靶序列可以是重组酶的靶标。重组酶定向地催化对每种重组酶特异的短的(30–40核苷酸)靶位点序列之间的dna交换反应。重组酶的实例包括cre重组酶(以loxp作为靶序列)和flp重组酶(具有短的翻转酶识别靶标(frt)位点)。加工酶靶序列可以是位点特异性整合酶例如phic31整合酶的靶标。加工酶靶序列可以是原核端粒酶的靶标。原核端粒酶靶序列是任何dna序列,其在dna模板中的存在允许其通过原核端粒酶的酶活性而转变成封闭的线性dna。换句话说,原核端粒酶靶序列是双链dna通过原核端粒酶的断裂和再连接以形成共价封闭的线性dna所需要的。通常,原核端粒酶靶序列包含任何回文序列,即任何具有二重旋转对称的双链dna序列,在本文中也被描述为反向重复。反向重复的长度根据具体的生物体而不同。回文或反向重复可以是完美的或不完美的。原核端粒酶靶序列优选包含至少14个碱基对长度的双链回文(反向重复)序列。适当的原核端粒酶靶向位点是本领域已知的,且在ep2,391,731中有讨论,其通过引用并入本文。适当的原核端粒酶可以是来自大肠杆菌噬菌体n15的teln。优选的是,加工酶靶位点是原核端粒酶靶向位点。
加工酶靶序列可以是用于rna聚合酶的靶序列,使得dna成为用于多肽合成的模板。在这种情况下,加工酶靶向位点是启动子,优选真核生物启动子。
该方法可以包括另外的使反应混合物与加工酶接触的步骤。可以使反应混合物在任何时间与加工酶接触。优选地,一旦dna合成完成就使反应混合物与加工酶接触。加工酶可以是rna聚合酶、重组酶、限制性内切酶或原核端粒酶。优选地,加工酶是原核端粒酶。
本发明中可用的原核端粒酶是任何能够使包含原核端粒酶靶点的模板断裂和重新连接以生产共价封闭的线性dna分子的多肽。因此,原核端粒酶具有dna断裂和连接功能。具有原核端粒酶型活性的酶还被描述为端粒解离酶(例如在伯氏疏螺旋体(borreliaburgdorferi)中)。原核端粒酶的典型底物是环状双链dna。如果该dna含有原核端粒酶靶位点,则酶能够在该位点切割dna并且将末端连接以产生线性双链的共价封闭的dna分子。上文讨论了对原核端粒酶靶位点的要求。还如上文所概述的,可以使用本领域中描述的任何适当的测定方法来测定给定的多肽催化从包含原核端粒酶靶为点的模板生产封闭的线性dna的能力。
dna的加工可能要求使用至少一种原核端粒酶。本发明的方法可以包括使用超过一种原核端粒酶,例如两种、三种、四种、五种或更多种不同的原核端粒酶。适当的原核端粒酶的实例包括来自噬菌体的那些,例如来自海水盐单胞菌(halomonasquamarina)的phihap-1、来自小肠炎耶尔森菌(yersiniaenterolytica)的py54、来自产酸克雷伯菌(klebsiellaoxytoca)的phik02和来自弧菌属(vibriosp.)的vp882,和来自大肠杆菌的n15(seqidno:15),或其任何变体。噬菌体n15原核端粒酶或其变体的使用是尤其优选的。该酶也被称为teln。这些酶进一步描述在wo2012/017210中,其通过引用并入本文。
因此,该方法可以在包括加工酶的dna模板上实施,且可以以受控的方式向反应混合物供给加工酶。可以在任何适当的时间向反应混合物供给加工酶;这可以是响应于信号。可以进行加工酶的一次或多次添加。优选的是,以不连续地向反应混合物添加的多个等份来供给加工酶。可以向反应混合物添加1、2、3、4、5、6、7、8、9、10或更多等份的加工酶。恰好在需要一份另外的核苷酸和/或另外的金属离子之前添加加工酶可以是有利的。或者,加工酶可以与一份另外的核苷酸一起添加或在一份另外的核苷酸添加至反应混合物之后添加。可以响应于反应混合物中的dna已达到期望的浓度的信号来添加加工酶。可以如本文所述地检测该dna浓度。
dna模板可以包含表达盒,该表达盒包含有效地与编码(enclosing)目标蛋白质的序列连接的真核生物启动子和任选的真核转录终止序列、由其组成或者基本上由其组成。任选地,表达盒可以是最小盒,其缺少一种或多种细菌或载体序列,通常选自:(i)细菌复制起点;(ii)细菌选择标记物(例如抗生素抗性基因),和(iii)未甲基化的cpg基序。“启动子”是启动和调节多核苷酸转录的核苷酸序列。启动子可以包括诱导型启动子(其中有效地与启动子连接的多核苷酸序列的表达是被分析物、辅因子、调节蛋白质等诱导的)、阻抑型启动子(其中有效地与启动子连接的多核苷酸序列的表达是被分析物、辅因子、调节蛋白质等所抑制的)和组成型启动子。术语“启动子”或“控制元件”意图包括全长启动子区域和这些区域的功能性(如控制转录或翻译)节段。“有效地连接”是指元件的安排,其中所述的成分被设置以使其实施其通常的功能。因此,给定的有效地与核酸序列连接的启动子在存在适当的酶时能够引起该序列的表达。启动子不需要与该序列紧邻,只要它起作用以指导其表达即可。因此,例如,在启动子序列和核酸序列之间可以存在介于其间的未翻译的但转录了的序列,且启动子序列仍然可以被看做是与编码序列“有效地连接”。因此,术语“有效地连接”旨在涵盖启动子元件和目标dna序列的任何间隔和定向,其在由通过转录复合物识别出启动子元件时允许启动目标dna序列的转录。
dna模板可以是任何适当的长度。特别地,dna模板可以是高达60千碱基、或高达50千碱基、或高达40千碱基、或高达30千碱基。优选地,dna模板可以是100碱基至60千碱基、200碱基至20千碱基,更优选200碱基至15千碱基,最优选2千碱基至15千碱基。
可以通过本领域已知的任何方法以足以用于该方法的量来提供dna模板。例如,模板可以通过pcr来生产。
dna模板的全部或所选的部分可以以该方法扩增。
dna模板可以包含用于表达的dna序列。dna可以用于在细胞(即在体外或体内转染的细胞)中表达,或者可以用于在无细胞体系(即蛋白质合成)中表达。用于表达的dna序列可以用于治疗目的,即dna疫苗的基因治疗。用于表达的序列可以是基因,且所述基因可以编码dna疫苗、治疗性蛋白质等。dna序列可以包含被转录成活性rna形式(即小干扰rna分子(sirna))的序列。
dna模板与至少一种聚合酶接触。可以使用一种、两种、三种、四种或五种不同的聚合酶。聚合酶可以是任何适当的聚合酶,使得其合成dna的聚合物。聚合酶可以是dna聚合酶。可以使用任何dna聚合酶,包括任何可商购的dna聚合酶。可以使用两种、三种、四种、五种或更多种不同的dna聚合酶,例如一种提供校读功能,一种或多种其他的不提供。可以使用具有不同机理的dna聚合酶,如链置换型聚合酶和通过其他方法复制dna的dna聚合酶。不具有链置换活性的dna聚合酶的适当的实例是t4dna聚合酶。
聚合酶可以是高度稳定的,从而其活性不会因为在方法条件下延长的孵育而显著降低。因此,酶优选在包括但不限于温度和ph的一系列方法条件下具有长的寿命。还优选聚合酶具有一个或多个适于制造过程的特性。聚合酶优选具有高的保真度,例如通过具有校读活性。此外,优选聚合酶显示高的持续合成能力、高的链置换活性以及对于dntp和dna的低km。聚合酶可以能够使用环状的和/或线性的dna作为模板。聚合酶可以能够使用dsdna或ssdna作为模板。优选聚合酶不显示与其校读活性不相关的dna核酸外切酶活性。
本领域技术人员能够确定给定的聚合酶与下列的可商购的聚合酶显示的性质相比是否显示上文所定义的特性:phi29(newenglandbiolabs,inc.,ipswich,ma,us)、deep
链置换型聚合酶是优选的。优选的链置换型聚合酶是phi29、deepvent和bstdna聚合酶i或其中任一种的变体。“链置换”描述聚合酶在合成过程中在遇到双链dna的区域时置换互补链的能力。因此,模板通过置换互补链和合成新的互补链而扩增。因此,在链置换复制过程中,新复制的链会被置换,为聚合酶让路,以复制另外的互补链。当引物或单链模板的3’游离末端退火至模板上的互补序列(二者均为引发事件)时,扩增反应引发。当dna合成进行时,且如果其遇到退火至模板的另外的引物或其他链,则聚合酶置换它并继续其链延伸。链置换产生新合成的单链dna,其可以用作更多引发事件的模板。新合成的dna的引发导致超支化,以及产物的高产率。应当理解,链置换扩增方法不同于基于pcr的方法之处在于,变性的循环对于有效的dna扩增不是关键的,因为双链dna不是连续合成新dna链的障碍。链置换扩增可以仅要求初始的一轮加热,以使初始的模板(如果其是双链的)变性,以允许引物退火至引物结合位点(如果使用的话)。在这之后,扩增可以被描述为等温的,因为不要求进一步的加热或冷却。相反,pcr方法在扩增方法过程中要求变性(即,将温度升高至94摄氏度或更高)的循环,以解链双链dna并提供新的单链模板。在链置换过程中,聚合酶会置换已合成的dna的链。此外,其会使用新合成的dna作为模板,确保dna的快速扩增。
本发明的方法中所用的链置换聚合酶优选具有至少20kb、更优选至少30kb、至少50kb或至少70kb或更高的持续合成能力。在一个实施方式中,链置换dna聚合酶具有与phi29dna聚合酶相当或更高的持续合成能力。
因此,链置换复制是优选的。在链置换复制过程中,模板通过置换已复制的链而扩增,该已复制的链已通过聚合酶的作用而合成,其反过来又置换另一个链,该另一个链可以是双链模板的原始互补链或新合成的互补链,后者是通过聚合酶对退火至模板的在先引物的作用而合成的。因此,模板的扩增可以通过另一个链的链置换复制来置换已复制的链而发生。该方法可以被描述为链置换扩增或链置换复制。
优选的链置换复制方法是环介导的等温扩增或lamp。lamp通常使用4-6个识别模板dna的6-8个不同区域的引物。简单地说,链置换dna聚合酶引发合成,且引物中的2个形成环结构以有利于随后的扩增轮次。含有目标dna的正义和反义链的序列的内引物引发lamp。以下由外引物引发的链置换dna合成释放单链dna。这充当用于由杂交至靶标的另一末端的第二内和外引物引发的dna合成的模板,这产生茎环dna结构。在随后的lamp循环中,一个内引物杂交至产物上的环,并引发置换dna合成,产生起始的茎环dna和具有长度为两倍的茎的新的茎环dna。
优选的链置换复制方法是滚环扩增(rca)。术语rca描述rca型聚合酶围绕环状dna模板链连续进展并同时延伸杂交引物的能力。这导致形成具有扩增的dna的多个重复的线性单链产物。环状模板的序列(单一单位)在线性产物中多次重复。对于环状模板,链置换扩增的初始产物是单链的多联体,其是正义的或反义的,这取决于模板的极性。这些线性单链产物充当多次杂交、引物延伸和链置换事件的基础,导致形成多联体双链dna产物,其同样包含扩增的dna的多个重复。因此,在多联体双链dna产物中存在每个扩增的“单一单位”dna的多个副本。特别优选将rca聚合酶用于本发明的方法。rca型链置换复制方法的产物可能需要加工,以释放单一单位dna。如果需要dna的单一单位,则这是期望的。
为了允许扩增,还使dna模板与一种或多种引物接触。引物可以是非特异性的(即,序列是随机的)或可以对dna模板内包含的一个或多个序列是特异性的。如果引物是随机序列,则它们允许在dna模板上任一位点处的非特异性的引发。这允许通过多个引发反应从每个模板链的高效扩增。随机引物的实例为六聚物、七聚物、八聚物、九聚物、十聚物或长度更长的序列,例如长度为12、15、18、20或30个核苷酸。随机引物的长度可以是6至30个、8至30个或12至30个核苷酸。随机引物通常以寡核苷酸的混合物的形式提供,这些寡核苷酸是dna模板中例如六聚物、七聚物、八聚物或九聚物的所有可能的组合的代表。
在一个实施方式中,引物或者引物中的一种或多种是特异性的。这意味着它们具有与期望从其引发扩增的dna模板的序列互补的序列。在该实施方式中,可以使用一对引物来特异性地扩增dna模板中位于两个引物结合位点之间的部分。或者,可以使用单一的特异性引物。
在一个实施方式中,dna模板包括一个或多个原核端粒酶靶序列。这样的靶序列在性质上是回文的。在该实施方式中,可以使用与原核端粒酶结合位点内的回文序列结合的特异性引物,并且因此可以在dna模板的两个链上均引发扩增。这样的方法详细描述在wo2012/017210中。因此,单一引物对于扩增是足够的。
引物可以不被标记,或者可以包含一个或多个标记,例如放射性核素或荧光染料。引物还可以包含化学修饰的核苷酸。例如,引物可以被封端,以防止dna合成的引发,直至封端被移除,即,通过化学或物理方式。通常可以基于温度考虑来选择引物长度/序列,即,能够在扩增步骤中使用的温度下与模板结合。
dna模板与聚合酶和一种或多种引物的接触可以在促进引物退火至dna模板的条件下进行。条件包括允许引物杂交的单链dna的存在。条件还包括允许引物退火至模板的温度和缓冲剂。可以基于引物的性质来选择适当的退火/杂交条件。本发明中所用的优选的退火条件的实例包括缓冲剂:30mmtris-hclph7.5,20mmkcl,8mmmgcl2。退火可以在使用加热而变性之后通过逐步冷却至期望的反应温度而进行。
然而,使用链置换复制的扩增还可以在没有引物的条件下进行,并且因此不要求发生杂交和引物延伸。相反,单链dna模板通过形成具有可用于延伸的游离3’末端的发夹而自引发。扩增的其余步骤保持相同。
还使dna模板和聚合酶与核苷酸接触。dna模板、聚合酶和核苷酸的组合形成反应混合物。反应混合物还可以包含一种或多种引物。反应混合物还可以独立地包括一种或多种金属阳离子。
核苷酸是核酸的单体或单一单位,且核苷酸由含氮碱基、五个碳的糖(核糖或脱氧核糖)和至少一个磷酸盐基团组成。可以使用任何适当的核苷酸。
核苷酸可以以游离酸、其盐或螯合物,或者游离酸和/或盐或螯合物的混合物的形式存在。
核苷酸可以以一价金属离子核苷酸盐或二价金属离子核苷酸盐的形式存在。盐可以包括二价金属离子的盐,例如但不限于镁(mg2+)、锰(mn2+)、钙(ca2+)、铍(be2+)、锌(zn2+)和锶(sr2+),或者一价金属离子的盐,包括但不限于锂(li+)、钠(na+)或钾(k+)。
含氮碱基可以是腺嘌呤(a)、鸟嘌呤(g)、胸腺嘧啶(t)、胞嘧啶(c)和尿嘧啶(u)。含氮碱基还可以是修饰的碱基,例如5-甲基胞嘧啶(m5c)、假尿苷(ψ)、二氢尿苷(d)、肌苷(i)和7-甲基鸟苷(m7g)。
优选五个碳的糖是脱氧核糖,从而核苷酸是脱氧核苷酸。
核苷酸可以是脱氧核苷三磷酸(记为dntp)的形式。这是本发明优选的实施方式。适当的dntp可以包括datp(脱氧腺苷三磷酸)、dgtp(脱氧鸟苷三磷酸)、dttp(脱氧胸苷三磷酸)、dutp(脱氧尿苷三磷酸)、dctp(脱氧胞苷三磷酸)、ditp(脱氧肌苷三磷酸)、dxtp(脱氧黄苷三磷酸)及其衍生物和修饰的形式。优选dntp包含datp、dgtp、dttp或dctp或其修饰的形式或衍生物中的一种或多种。优选使用datp、dgtp、dttp和dctp或其修饰的形式的混合物。
核苷酸可以是溶液的形式或者以冻干的形式提供。优选核苷酸的溶液。
核苷酸可以包含修饰的核苷酸,并且这些修饰的核苷酸可以是生物上无活性的形式。因此,修饰的核苷酸可以是生物上无活性的核苷酸。生物上无活性的核苷酸可以具有保护核苷酸的基团(例如3′碳氧或末端磷酸盐氧)的可移除部分。生物上无活性核苷酸可以是笼锁的核苷酸或闭锁的(blocked)核苷酸。适当的部分包括但不限于可光活化的(笼锁)基团,包括α-羧基-2-硝基苄基(cnb)、1-(2-硝基苯基)乙基(npe)、4,5-二甲氧基-2-硝基苄基(dmnb)、1-(4,5-二甲氧基-2-硝基苯基)乙基(dmnpe)和5-羧基甲氧基-2-硝基苄基(cmnb)(molecularprobes)。这些基团通过经由末端磷酸盐氧而连接。该部分可以通过任何适当的方式而除去。例如,如果该部分是光不稳定的,则通过紫外光对该部分进行闪光光解导致生物活性核苷酸在光照位点处的快速和高度局部化的释放。该部分可以是热不稳定的,并且因此可以通过热来释放生物活性核苷酸。如果使用热来使核苷酸生物活化,则必须考虑聚合酶的温度要求,且这在本领域技术人员的能力范围内。例如,末端磷酸盐可以被笼锁或闭锁部分酯化。
可以以一种或多种适当的碱基的混合物的形式提供核苷酸,优选腺嘌呤(a)、鸟嘌呤(g)、胸腺嘧啶(t)、胞嘧啶(c)中的一种或多种。在合成dna的方法中使用两种、三种或者优选所有四种核苷酸(a、g、t和c)。
核苷酸可以均为天然核苷酸(即,未修饰的),它们可以是修饰的核苷酸,其与天然核苷酸类似地起作用,且是生物活性的(即,lna核苷酸–锁核酸),它们可以是修饰的且生物上无活性的,或者它们可以是未修饰的和修饰的核苷酸的混合物,和/或生物活性的和生物上无活性的核苷酸的混合物。每种类型(即,碱基)的核苷酸可以以一种或多种形式提供,即,未修饰的和修饰的,或者生物活性的和生物上无活性的。
在本发明的一个方面,反应混合物中包括核苷酸且控制向反应混合物供给另外的核苷酸。根据该方面,反应混合物中存在的核苷酸的起始浓度可以是低于8mm、低于7mm、低于6mm、低于5mm、低于4mm、低于3mm、低于2mm或低于1mm。如此添加核苷酸以形成反应混合物,使得反应混合物中核苷酸的起始浓度是8mm或更低,优选5mm或更低,更优选4mm或更低。起始浓度是指反应开始时反应混合物中核苷酸(所有碱基)的总浓度。
反应混合物中存在的核苷酸的起始浓度可以足以合成至少0.3g/l的dna。优选起始浓度足以合成0.5g/ldna,更优选0.6g/l或0.65g/ldna。给出的体积是反应混合物的体积。核苷酸的起始浓度不会足以由反应混合物合成期望量的dna,即,其可能不足以达到最终的产率。核苷酸的起始浓度可以基于反应混合物中一种或多种聚合酶来确定。
可选地或者此外,核苷酸的起始浓度可以被定义为聚合酶的浓度与核苷酸的浓度之间的比例。在开始时,核苷酸分子的数目与每个聚合酶之间的比例可以是高达500,000:1、高达300,000:1或高达200,000:1或者为5,000:1至100,000:1,优选20,000:1至90,000:1,更优选30,000:1至80,000:1,更优选40,000:1至60,000:1。核苷酸分子的数目与每个聚合酶之间的比例优选为高于5,000:1。在一个优选的方面,在开始时,核苷酸:聚合酶的比例为50,000:1。
在促进所述模板扩增的条件下孵育反应混合物。优选链置换扩增。优选地,该条件促进所述模板通过另一个链的链置换复制来置换已复制的链而扩增。条件包括使用任何允许dna扩增的温度,通常为20至90摄氏度的范围。优选的温度范围可以是约20至约40或者约25至约35摄氏度。对于lamp扩增,优选的温度是约50至约70摄氏度。
通常,基于具体的聚合酶具有最佳活性的温度来选择适当的温度。该信息是普遍可得的,且形成本领域技术人员通常知识的一部分。例如,当使用phi29dna聚合酶时,适当的温度范围会是约25至约35摄氏度,优选约30摄氏度。本领域技术人员会常规地能够识别根据本发明的方法用于有效扩增的适当温度。例如,可以在一定的温度范围内进行该方法,并且可以监测扩增的dna的产率,以识别对于给定聚合酶最佳的温度范围。扩增可以在恒定的温度下进行,且优选该方法是等温的。由于链置换扩增是优选的,所以不要求改变温度以分离dna链。因此,该方法可以是等温方法。
促进dna模板扩增的其他条件包括适当的缓冲剂/ph和对于酶性能和稳定性所需的其他因素的存在。适当的条件包括用于提供本领域已知的聚合酶的活性的任何条件。
例如,反应混合物的ph可以为3至10,优选5至8或约7,例如约7.5。可以通过使用一种或多种缓冲剂使ph保持在该范围内。这样的缓冲剂包括但不限于mes、bis-tris、ada、aces、pipes、mobs、mops、mopso、bis-tris丙烷、bes、tes、hepes、dipso、tapso、trizma、heppso、popso、tea、epps、tricine、gly-gly、bicine、hepbs、taps、ampd、tabs、ampso、ches、capso、amp、caps、cabs、磷酸盐、柠檬酸-磷酸氢钠、柠檬酸-柠檬酸钠、乙酸钠-乙酸、咪唑和碳酸钠-碳酸氢钠。
尽管使用热的应用(暴露于95℃下数分钟)来使双链dna变性,但也可以使用更适用于dna合成的其他方法。通过暴露于高或低ph环境或者在阳离子不存在或以非常低浓度存在例如在去离子水中,可以很容易使双链dna变性。聚合酶要求短的寡核苷酸引物序列结合至dna模板的单链区域以引发其复制。该相互作用的稳定性以及因此dna扩增的效率可以尤其受金属阳离子浓度的影响,尤其是二价阳离子例如mg2+离子,其可以被看做该方法的不可缺少的一部分。
反应混合物还可以包含金属离子。反应混合物还可以包含金属的盐,例如但不限于二价金属离子的盐:镁(mg2+)、锰(mn2+)、钙(ca2+)、铍(be2+)、锌(zn2+)和锶(sr2+),或者一价金属离子的盐,包括但不限于锂(li+)、钠(na+)或钾(k+)。盐可以包括氯化物、乙酸盐和硫酸盐。可以包括的其他盐为铵盐,尤其是硫酸铵。本领域技术人员会知晓,以金属离子盐的形式供给核苷酸会影响反应混合物中金属离子的浓度,并且如果需要的话能够考虑该另外的金属离子来源。反应混合物中金属离子的总浓度优选地包括金属离子的所有来源。
反应混合物中还可以包括去污剂。适当的去污剂的实例包括tritonx-100、tween20和其中任一种的衍生物。反应混合物中还可以包括稳定剂。可以使用任何适当的稳定剂,尤其是牛血清白蛋白(bsa)和其他稳定化蛋白质。还可以通过添加使dna松弛和使模板变性更容易的试剂来改进反应条件。这样的试剂包括例如二甲基亚砜(dmso)、甲酰胺、甘油和甜菜碱。反应混合物中还可以包括dna浓缩剂。这样的试剂包括例如聚乙二醇或阳离子脂质或阳离子聚合物。
应当理解,本领域技术人员能够基于其通常的知识,使用这些另外的成分和条件来修改并优化用于本发明的方法的扩增和孵育条件。类似地,可以基于本领域先前的实例来选择特定试剂的具体条件,并基于通常的知识进一步优化。作为实例,可以优化反应混合物中存在的聚合酶的量。这可以涉及在dna合成的过程中向反应混合物进一步添加聚合酶。作为另一个实例,可以优化dna模板的量。这可以涉及在dna合成的过程中向反应混合物进一步添加dna模板。聚合酶和/或dna模板的进一步供给可以是连续的或不连续的,优选不连续的。
作为实例,本领域中基于rca的方法中所用的适当的反应缓冲剂是50mmtrishcl、ph7.5、10mmmgcl2、20mm(nh4)2so4、5%甘油、0.2mmbsa、1mmdntp。本发明的rca扩增中所用的优选的反应缓冲剂是30mmtris-hcl、ph7.4、30mmkcl、7.5mmmgc12、10mm(nh4)2so4、4mmdtt、2mmdntp。该缓冲剂尤其适合于与phi29rca聚合酶一起使用。
反应混合物还可以包含使用一种或多种另外的蛋白质。dna模板可以在至少一种焦磷酸酶例如酵母无机焦磷酸酶的存在下扩增。可以使用两种、三种、四种、五种或更多种不同的焦磷酸酶。这些酶能够使在链复制过程中通过聚合酶由dntp产生的焦磷酸盐降解。焦磷酸盐在反应中的累积能够导致dna聚合酶的抑制,并降低dna扩增的速度和效率。焦磷酸酶可以将焦磷酸盐分解成非抑制性的磷酸盐。用于本发明的方法的适当的焦磷酸酶的实例是酿酒酵母(saccharomycescerevisiae)焦磷酸酶,其可商购自newenglandbiolabs,inc.。
本发明的方法中可以使用任何单链结合蛋白质(ssbp),以稳定单链dna。ssbp是活细胞的基本成分,且参与所有涉及ssdna的过程,例如dna复制、修复和重组。在这些过程中,ssbp与短暂形成的ssdna结合,并有助于稳定ssdna结构。用于本发明的方法的适当的ssbp的实例是t4基因32蛋白质,其可商购自newenglandbiolabs,inc.。
如上文所述,可以在促进dna模板扩增的条件下孵育反应混合物。扩增优选为链置换复制。反应混合物包含核苷酸,其优选为先前讨论的起始浓度或起始比例(与聚合酶的比例)。反应混合物可以进一步包含金属离子。
根据本发明的一个方面,为了合成dna,要求供给另外的核苷酸。根据本发明的第一方面,控制核苷酸向反应混合物的供给。因此,在方法开始时,不向反应混合物添加dna合成所要求的所有核苷酸。可以在任何适当的时间向反应混合物供给或提供另外的核苷酸。向反应混合物供给另外的核苷酸的时机和/或向反应混合物供给的核苷酸的量是受控的或有指导的。因此,反应本身可以通过精确供给核苷酸来控制。该控制可以是基于酶动力学的理论计算,或者该控制可以是基于反应混合物的可测量的参数,从而在其被聚合酶要求时供给另外的核苷酸。
一旦反应混合物已在促进dna模板扩增的条件下形成和孵育后,就可以供给另外的核苷酸。可以在扩增已开始或已被引发后供给另外的核苷酸。在一个实施方式中,在提供起始的核苷酸之后至少20分钟、至少30分钟或至少40分钟开始进一步供给核苷酸。然而,可以在反应开始时开始供给,尤其是如果核苷酸的供给为连续时。
在一个实施方式中,反应混合物中核苷酸的浓度或量在方法过程中不超过较高的阈值。控制核苷酸的供给,从而核苷酸的浓度或量不超过该较高的阈值。该较高的阈值可以是反应混合物中8mm,或7mm或6mm或5mm或4mm或3mm或2mm核苷酸。优选该较高的阈值是反应混合物中约4mm核苷酸。类似地,在一个实施方式中,反应混合物中核苷酸的浓度或量在方法过程中可以不下降至低于较低的阈值。控制核苷酸的供给,从而核苷酸的浓度或量不下降至低于该较低的阈值。该较低的阈值可以是反应混合物中0.1μm、0.5μm、1μm、5μm或10μm核苷酸。优选在方法过程中使反应混合物中核苷酸的浓度保持在较高的和较低的阈值之间。因此,可以使反应混合物中核苷酸的浓度保持在0.0001mm至8mm,优选为0.001mm至6mm,最优选为0.01mm至5mm。可以通过控制向反应混合物供给另外的核苷酸来保持核苷酸浓度。在一个实施方式中,可以通过离子色谱来估计反应混合物中核苷酸的浓度。或者,如本文所讨论的,可以通过估计所合成的dna的量来计算核苷酸的大致浓度。
或者,可以与反应混合物中存在的核苷酸和聚合酶的量之间的比例相关地描述较高和较低的阈值。因此,较高的阈值可以是500,000:1(核苷酸:聚合酶)、200,000:1、150,000:1、100,000:1或80,000:1。较低的阈值可以是1,000:1、1,500:1或2,000:1。因此,可以使核苷酸与聚合酶的比例保持在1,000:1至500,000:1,优选2,000:1至450,000:1,更优选5,000:1至400,000:1,最优选10,000:1至100,000:1。最优选将该比例保持在40,000:1至70,000:1、在约50,000:1。
在一个实施方式中,以一次或多次不连续的添加,即分份,向反应混合物供给另外的核苷酸。任何添加次数或份数均在该实施方式的范围内。因此,可以向反应混合物添加1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20或更多次,即20-25、25-30、35-40次添加或份数。因此,可以使用1-50、1-40、1-30或1-20等份来供给核苷酸。一等份可以是方法中要求的或添加的核苷酸总量的一部分。该等份中核苷酸的体积和/或浓度可以根据方法的要求而变化。一等份可以包含少量的核苷酸,即,该量足以确保在反应混合物中存在特定浓度或量的核苷酸。可以以规律的间隔(即,每30分钟、每60分钟、每90分钟、每120分钟、每180分钟或每240分钟或任何其他适当的时间段)向反应混合物分份供给核苷酸。可以以不规律的间隔向反应混合物分份供给核苷酸。例如,每次供给核苷酸之间的时间段可以在开始时较短,然后随着方法进行而增加,或者相反。可以提供各等份以保持反应混合物中恒定的核苷酸浓度,或者可以提供各等份以补充核苷酸浓度至起始浓度。在优选的实施方式中,供给核苷酸,从而反应混合物中的浓度不超过8mm,或7mm或6mm或5mm或4mm。在优选的实施方式中,以这样的方式供给核苷酸,以确保反应混合物中核苷酸的浓度或量落在较高和较低阈值之间。
优选地,该等份的提供不仅是为了替换被取走用于分析或测试的反应混合物的体积,而是因为反应要求另外的核苷酸。优选地,反应混合物的测试是非侵入性的,且直至反应完成,不从反应混合物抽取任何物质。
可以响应于对反应混合物中更多核苷酸的需要而供给另外的核苷酸。这可以通过若干参数来确定,包括但不限于dna合成速率、所合成的dna的浓度或者反应混合物的粘度或体积。可以从所合成的dna的浓度随时间的变化来计算dna合成速率。当dna合成速率开始下降,可以供给另外的核苷酸。或者,可以通过下文进一步讨论的方法监测所合成的dna的浓度。当dna的浓度增加至阈值水平,可以供给另外的核苷酸。可以测量反应混合物的粘度,并且粘度的增加可以指示需要另外的核苷酸。可以用粘度计测量反应的粘度。
在一个实施方式中,连续地向反应混合物供给另外的核苷酸,即,以连续的方式将它们添加至反应混合物。或者说,可以滴加或持续地供给核苷酸。根据该实施方式供给另外的核苷酸可以是以足够小的量,从而核苷酸的供给与聚合酶对另外的核苷酸的需求相匹配。在该实施方式中,核苷酸的持续供给保持核苷酸在反应混合物中大约恒定的浓度。优选地,向反应混合物持续供给另外的核苷酸确保反应混合物中核苷酸的浓度保持在较高和较低阈值之间。
可以以物理方式向反应混合物提供另外的核苷酸,即,它们在反应混合物之外。因此,该供给可以开始于外部来源,例如储存器。该储存器可以连接至反应混合物。核苷酸可以被泵送至反应混合物。因此,核苷酸和反应混合物是物理上分开的,且另外的核苷酸的供给是核苷酸的物理供给。或者,可以经由渗透泵来供给核苷酸。渗透泵提供用于受控的和连续的核苷酸供给的方法。
或者,可以通过活化生物上无活性的核苷酸(例如先前讨论的那些)而向反应混合物提供核苷酸。为了向反应混合物供给核苷酸,通过适当的方式活化生物上无活性的核苷酸。因此,另外的核苷酸可以包含任何适当的生物上无活性的核苷酸。生物上无活性的核苷酸可以在开始时包括在反应混合物中,但仅通过受控的活化供给至反应混合物。或者,可以分份或连续向反应混合物物理添加生物上无活性的核苷酸,并且随后将其活化以向反应混合物供给核苷酸。活化可以是物理的(即,热或光)或化学的,且通过活化的控制来控制核苷酸的供给。
可以控制向反应混合物供给核苷酸,直至完成dna的合成,例如一旦dna模板耗尽,且不可能再有进一步的并入。或者,一旦已合成了预期的或期望的dna产率,则反应可以完成。这样的产率可以是基于所添加的核苷酸的量。一旦已合成了需要量的dna,例如1g每升反应混合物,2g/l、3g/l、4g/l、5g/l或者高达或超过10g/l,则dna的合成可以完成。一旦dna合成的速率已降低,且进一步供给核苷酸不增加该速率,反应混合物用光了另一种成分(例如但不限于引物),聚合酶被灭活,无机焦磷酸酶被灭活,不再有单链模板,所需的辅因子例如金属阳离子被用尽,反应混合物的粘度达到特定的阈值,反应混合物的体积达到一定的阈值或者聚合酶被抑制,则dna的合成可以完成。
反应的产率涉及所合成的dna的量。由根据本发明的方法预期的产率可以超过10克/升反应混合物,或者10、9、8、7、6、5、4、3、2或1g/l。本发明改进了酶促合成dna的可能的产率。本发明的一个目标是改进无细胞酶促dna合成方法的产率,从而能够以成本有效的途径大规模合成dna。本发明允许使用被聚合酶催化的酶促方法以工业规模经济地制造/合成dna。本发明的方法允许有效地将核苷酸整合到dna产物中。认为本发明的方法会允许反应混合物被放大至数升,包括数十升。改进的产率、dna合成速率、生产率或持续合成能力可以与其中在开始时供给所有核苷酸的相同的反应混合物相当。认为可以通过控制向反应混合物供给另外的核苷酸来改进收率,如实施例中所证明的。
dna合成的速率涉及方法中每分钟所合成的dna的量或浓度。在实施例中会看到,dna合成的速率随着方法进行而逐步变慢。
通过使用本发明的一种或多种方法,可以增加方法的生产率,或者每单位供给的核苷酸的dna产出的速率。
在一个实施方式中,本发明涉及用于增强dna的合成的方法。该增强可以与除了包括核苷酸在内的所有需要的成分均在开始时添加之外相同的反应混合物相当。未向该对照反应混合物进行进一步的添加。
在一个方面,提供了一种用于合成dna的无细胞方法,包括使dna模板与至少一种聚合酶在一种或多种引物、核苷酸和金属阳离子的存在下在促进所述模板扩增的条件下接触,以形成反应混合物,所述模板扩增是通过另一条链的链置换复制来置换经复制的链,其中以受控的方式向反应混合物供给另外的金属阳离子。
或者说,扩增通过模板的链置换复制而发生。因此,本发明提供了一种用于合成dna的无细胞方法,包括使dna模板与至少一种聚合酶在一种或多种引物、核苷酸和金属阳离子的存在下在促进所述模板通过链置换复制而扩增的条件下接触,以形成反应混合物,其中以受控的方式向反应混合物供给另外的核苷酸和另外的金属阳离子。
优选地,通过以受控的方式向反应混合物供给另外的阳离子来控制用于合成dna的无细胞方法的反应速率。
根据本发明的该方面,可以控制向反应混合物供给金属阳离子。认为这种控制改进dna合成的速率,尤其在大规模方法中。在开始时,反应混合物中可以存在一种或多种金属阳离子。因此,反应混合物还可以包含一种或多种金属阳离子,其优选以盐的形式供给。可以控制向反应混合物供给另外的金属阳离子。可以供给一种或多种金属阳离子。适当的金属阳离子包括二价金属离子:镁(mg2+)、锰(mn2+)、钙(ca2+)、铍(be2+)、锌(zn2+)和锶(sr2+),或者一价金属离子,包括但不限于锂(li+)、钠(na+)或钾(k+)。金属阳离子优选以盐的形式供给。盐可以包括氯化物、乙酸盐和硫酸盐,且优选以氯化物盐的形式供给金属阳离子。可以包括的其他的盐是铵盐,尤其是硫酸铵。优选金属阳离子是镁或锰,最优选为镁。在最优选的实施方式中,金属阳离子以氯化镁的形式提供。
根据本发明的一个方面,为了合成dna,需要供给另外的金属阳离子。根据本发明的第二方面,控制向反应混合物供给金属阳离子。因此,不在方法的开始向反应混合物添加合成dna所需的所有金属阳离子。可以在任何适当的时间向反应混合物供给或提供另外的金属阳离子。向反应混合物供给另外的金属阳离子的时机和/或向反应混合物供给金属离子的量是受控的或有指导的。因此,反应本身可以通过精确供给金属离子来控制。该控制可以是基于酶动力学的理论计算,或者该控制可以是基于反应混合物的可测量的参数,从而在其被要求时供给另外的金属离子。可以与另外的核苷酸的受控供给平行地或相关地控制另外的金属阳离子的供给。供给可以被独立地控制或一起控制。
一旦反应混合物已在促进dna模板扩增的条件下形成或孵育后,就可以供给另外的金属阳离子。可以在扩增已开始或已被引发后供给另外的金属阳离子。在一个实施方式中,在提供起始浓度的金属阳离子之后至少20分钟、至少30分钟或至少40分钟开始进一步供给金属阳离子。然而,可以在反应开始时开始供给,尤其是如果金属阳离子的供给为连续时。
在一个实施方式中,反应混合物中游离的或未结合的金属阳离子的浓度或量在方法过程中不超过较高的阈值。控制金属阳离子的供给,从而游离的或未结合的金属阳离子的浓度或量不超过最高的阈值。该最高的阈值可以是反应混合物中4mm、3mm或2mm。优选该最高的阈值为反应混合物中约3mm。类似地,在一个实施方式中,反应混合物中游离金属阳离子的浓度或量在方法过程中可以不下降至低于最低的阈值。控制金属阳离子的供给,从而游离金属阳离子的浓度或量不下降至低于该最低的阈值。该最低的阈值可以是反应混合物中的0.03μm、0.15μm、0.3μm、1.5μm或3μm。优选在方法过程中使反应混合物中游离金属阳离子的浓度保持在最高的和最低的阈值之间。因此,可以使反应混合物中游离金属阳离子的浓度保持在0.03μm至4mm,或0.5μm至3mm或1μm至2mm或10μm至1mm。优选使反应混合物中游离金属阳离子的浓度保持在1mm至1.5mm。
或者,最高的阈值可以是反应混合物中的10mm、8mm、6mm、4mm、3mm或2mm。优选最高的阈值是反应混合物中的约3mm。类似地,在一个实施方式中,反应混合物中游离金属阳离子的浓度或量在方法过程中可以不下降至低于最低的阈值。控制金属阳离子的供给,从而游离金属阳离子的浓度或量不下降至低于该最低的阈值。该最低的阈值可以是反应混合物中的0.03μm、0.15μm、0.3μm、1.5μm、3μm和5μm。优选在方法过程中使反应混合物中游离金属阳离子的浓度保持在最高的和最低的阈值之间。因此,可以使反应混合物中游离金属阳离子的浓度保持在0.03μm至10mm,或0.15μm至8mm或0.3μm至6mm或1.5μm至3mm。优选使反应混合物中游离金属阳离子的浓度保持在0.15μm至5μm。
可以通过控制向反应混合物供给另外的金属离子来保持游离金属阳离子的浓度。在一个实施方式中,可以通过原子吸收光谱、离子色谱和本领域中已知的其他方法来估计反应混合物中游离金属离子的浓度。或者,如本文所讨论的,可以通过估计所合成的dna的量来计算游离金属阳离子的大致浓度。为了避免疑问,金属阳离子,例如镁离子,可以与反应混合物中不同的物质结合,包括但不限于焦磷酸盐、磷酸盐和dna。这些离子是结合的,而不是游离的。
在一个实施方式中,以一次或多次不连续的添加,即分份,向反应混合物供给另外的金属阳离子。任何添加次数或份数均在该实施方式的范围内。因此,可以向反应混合物添加1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20或更多次,即20-25、25-30、35-40次添加或份数。因此,可以使用1-50、1-40、1-30或1-20等份来供给金属阳离子。一等份可以是方法中要求的或添加的金属阳离子总量的一部分。该等份中金属阳离子的体积和/或浓度可以根据方法的要求而变化。一等份可以包含少量的金属阳离子,即,该量足以确保在反应混合物中存在特定浓度或量的金属阳离子。可以以规律的间隔(即,每30分钟、每60分钟、每90分钟、每120分钟、每180分钟或每240分钟或任何其他适当的时间段)向反应混合物分份供给金属阳离子。可以以不规律的间隔向反应混合物分份供给金属阳离子。例如,每次供给金属阳离子之间的时间段可以在开始时较短,然后随着方法进行而增加,或者相反。可以提供各等份以保持反应混合物中恒定的金属阳离子浓度,或者可以提供各等份以补充金属阳离子浓度至起始浓度,优选最佳的游离金属离子浓度。
在优选的实施方式中,以这样的方式供给金属阳离子,以确保反应混合物中游离金属阳离子的浓度或量落在最高和最低的阈值之间。
可以响应于反应混合物中对其的需要而供给另外的金属阳离子。如先前所讨论的,这可以通过若干参数来确定。
在一个实施方式中,连续地向反应混合物供给另外的金属阳离子。
可以以物理方式向反应混合物提供另外的金属阳离子,即,它们在反应混合物之外。
优选也控制另外的核苷酸的供给。
在另一方面,提供了用于合成dna的无细胞方法,包括使dna模板与至少一种聚合酶在一种或多种引物、核苷酸和金属阳离子的存在下在促进所述模板扩增的条件下接触,以形成反应混合物,所述模板扩增是通过另一条链的链置换复制来置换经复制的链,其中以受控的方式向反应混合物供给另外的核苷酸和金属阳离子。
还提供了一种用于合成dna的无细胞方法,包括使dna模板与至少一种聚合酶在核苷酸和金属阳离子的存在下在促进所述模板扩增的条件下接触,以形成反应混合物,其中以受控的方式向反应混合物供给另外的核苷酸和另外的金属阳离子。
可以一起或分开地供给另外的核苷酸和另外的金属离子。另外的核苷酸或另外的金属阳离子的受控供给可以如先前与这些物质相关地讨论的来实现。
金属阳离子或另外的金属阳离子的供给可以平行于核苷酸的供给,或者可以与核苷酸的供给不同时。
优选地,另外的金属阳离子可以与核苷酸的供给一起供给至反应混合物,尤其是当金属阳离子为一价时。因此,金属阳离子和核苷酸可以以混合物的形式供给。因此,金属阳离子和核苷酸的供给可以是组合的,尽管优选使它们保持分开。
另外的金属阳离子和/或另外的核苷酸的供给可以是如对这些物质各自单独所述的。
在本发明的优选实施方式中,供给金属阳离子和核苷酸,从而金属阳离子:核苷酸的比例(量)为9:1至1:9,或6:1至1:6或3:1或1:3,或1:1。优选反应混合物中这些量之间的比例为约3:1至1:3,优选约3:1。当金属阳离子是二价金属阳离子例如但不限于镁(mg2+)、锰(mn2+)、钙(ca2+)、铍(be2+)、锌(zn2+)和锶(sr2+)时,这些比例是尤其优选的。
认为保持金属阳离子:核苷酸优选镁阳离子的比例对于控制dna合成的速率可能是重要的。还可以如先前所述地保持核苷酸:聚合酶的比例。因此,通过控制核苷酸和金属阳离子的供给而控制这两个比例,可以控制dna的合成。
与本发明的任何方面相关,如果以金属阳离子的盐的形式供给核苷酸,则供给至反应混合物的这些盐中的金属阳离子的浓度会包括这些金属阳离子。本领域技术人员会知晓,以金属阳离子盐的形式供给核苷酸会影响反应混合物中金属阳离子的浓度,并且能够考虑该另外的量。
三磷酸核苷(atp、gtp、ctp和ttp),由于其化学性质,如果使用高浓度以增加dna产物产率,具有以多种方式干扰dna扩增的可能性。高浓度可以被视为反应混合物中高于4mm的dntp浓度。dntp的磷酸盐基团能够与一价和二价金属阳离子相互作用,且对mg2+具有最强的亲合力。例如,atp对mg2+的亲合力比na+高700倍以上,且比li+高将近400倍。
在扩增期间,聚合酶从引入到生长的dna链中的核苷酸释放焦磷酸盐。焦磷酸盐对mg2+离子的结合亲合力类似于atp,因此该过程不释放游离的mg2+离子。在扩增期间使用高起始浓度的核苷酸的后果会是降低游离mg2+离子的水平。由于聚合酶催化活性可能需要mg2+离子,通过与磷酸盐或磷酸盐基团的相互作用而造成的亚理想的水平对有效的扩增可能是有害的。然而,相反地,已知高浓度的mg2+离子影响某些dna聚合酶的保真度。因此,mg2+离子的反应浓度可能被认为对dna产率和质量是关键的。
焦磷酸盐分解可以被酶催化,且可能期望从焦磷酸盐释放结合的mg2+离子。然而,mg2+离子与po43-离子结合以形成基本上不溶的mg3(po4)2(20℃下在水中的溶解度为0.01mm)。
mg2+离子也会与合成的dna结合。
在任何dna聚合酶催化的反应中在任何特定的时间点时的游离mg2+离子的水平会取决于起始浓度、能够结合阳离子的分子的总浓度和反应的ph。由于方法处于平衡,所以总是可能存在一些游离的mg2+。在一个实施方式中,通过控制优选以镁盐的形式向反应混合物供给游离的mg2+离子来控制游离mg2+的水平。
能够通过滚环和链置换机理扩增环状dna模板的聚合酶能够生产模板的长的线性多联体重复。这样的滚环链置换聚合酶是优选的。例如,据报道,phi29dna聚合酶能够扩增环状dna模板,以生产长度为高达77千碱基的线性重复。这些非常长的分子的行为类似于聚合物,且能够通过增加粘度而显著改变反应的流变学性质。粘度的这种增加可以通过压力的变化而测量,该压力的变化可以与所合成的dna的浓度直接相关。这样的测量可以连续进行或者在设定的时间进行,以提供信号,优选在线信号,其可以用于指示反应的状态。可以响应于压力信号来控制另外的核苷酸的供给。可以响应于指示反应混合物的参数的信号进行核苷酸的这种供给,以增加产率和/或反应混合物的生产率。
相对于基于荧光化学试剂例如syto染料的结合的测定方法,估计dna浓度的压差测量具有重要的优势。与仅与双链dna结合的这些试剂不同,压差测量在含有单链形式的样品中不给出错误的dna估计。
在一个实施方式中,响应于信号来控制核苷酸的供给。信号可以是任何适当的信号,且可以是在线信号。优选地,信号与dna的浓度相关,尤其是基于对反应混合物中dna的总浓度的估计。或者,信号可以与反应混合物的体积相关。类似地,可以响应于信号来控制另外的金属离子的供给。信号可以相同或不同。
优选的信号是与反应混合物的粘度相关的信号。反应混合物的粘度可以提供对反应混合物中dna浓度的估计。可以通过包括粘度计在内的任何适当的方式测量粘度。优选通过压力的变化测量粘度。可以通过压力传感器、压力变换器、压力传送器、压力指示器、压力表或压力计来测量压差。在一个实施方式中,可以使用去除并替换反应混合物的一部分的循环泵来产生压差。优选地,以诸如本文指示的那些非侵入方式来实时监测反应。
本发明的方法可以以分批反应来实施。在该实施方式中,反应混合物存在于反应器中。另外的核苷酸和/或另外的金属阳离子的供给可以被进料到反应器中。因此,可以在分期分批反应器中实施该方法。或者,可以在连续流反应器中实施该方法。这样的反应器在本领域技术人员的知识范围内。下文给出尤其优选的形式。
在一个实施方式中,以一次或多次不连续的添加,即分份,向反应混合物供给一种或多种加工酶。任何添加次数或份数均在该实施方式的范围内。因此,可以向反应混合物添加1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20或更多次,即20-25、25-30、35-40次添加或份数。因此,可以使用1-50、1-40、1-30或1-20等份来供给加工酶。一等份可以是方法中要求的或添加的加工酶总量的一部分。该等份中加工酶的体积和/或浓度可以根据方法的要求而变化。一等份可以包含少量的加工酶,即,该量足以确保在反应混合物中存在特定浓度或量的加工酶。可以以规律的间隔(即,每30分钟、每60分钟、每90分钟、每120分钟、每180分钟或每240分钟或任何其他适当的时间段)向反应混合物分份供给加工酶。可以以不规律的间隔向反应混合物分份供给加工酶。例如,每次供给加工酶之间的时间段可以在开始时较长,然后随着方法进行而减少,或者相反。
可以响应于反应混合物中对其的需要而供给加工酶。如先前所讨论的,这可以通过若干参数来确定,包括反应混合物中dna的浓度和/或dna合成的速率。
可以以物理方式向反应混合物提供加工酶,即,它们在反应混合物之外。这些酶可以被容纳在单独的储存器中,并且以受控的方式释放进入反应混合物。
优选地,加工酶具有单一的转换,例如原核端粒酶,因为控制加工酶的量直接控制被酶加工的dna的量。加工酶可以与另外的核苷酸一起添加,或者在另外的核苷酸之后添加。如果还需要另外的金属离子,则可以优选将其在添加加工酶之后添加,从而在添加另外的金属离子之前发生dna的加工。
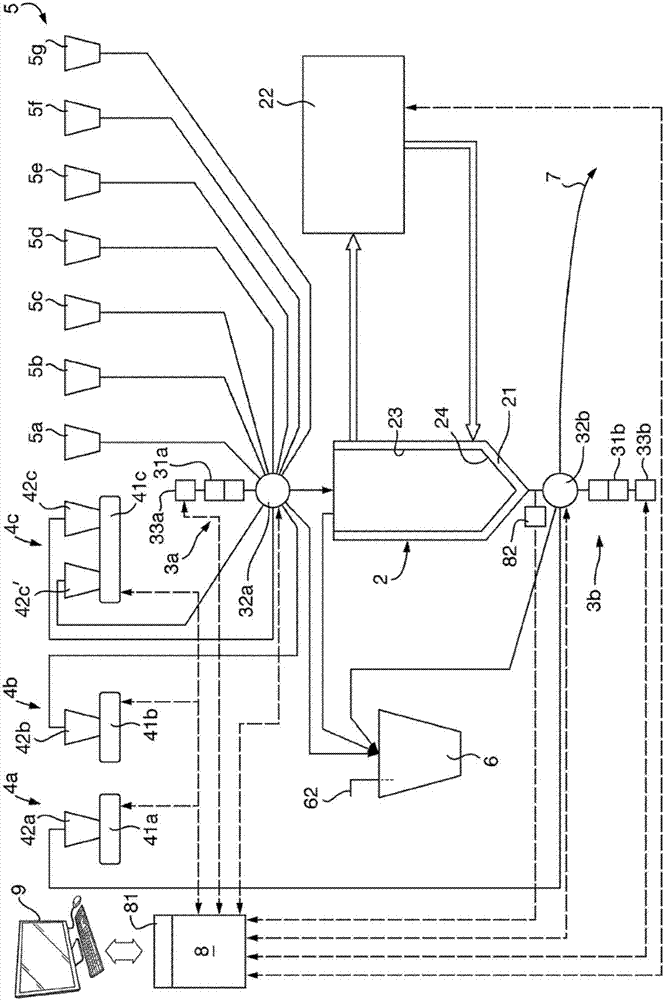
可以在任何适当的装置上实施该方法。现在将描述适于进行该方法的具体装置。合成装置1构建在独立的底架11上,且提供有可打开的前盖12以允许接近主要部件。前盖12特意由透明材料制成,但可以是有色的,以保护反应成分不接受过量的光水平。其还用于限制污染物进入。装置提供有可外部接触到的电源开关和功率输入13,例如,在装置的侧面上。此外,存在输入/输出端口14以允许装置与通用计算机执行的实验室自动化软件(例如由nationalinstrumentscorporationoftexas,usa供应的labviewtm)通讯和/或被该软件控制。在从装置的前方可视的位置提供操作指示灯15,其指示装置是否在操作中。
合成装置的中心部件是夹套反应容器2,其体积可以为超过30ml,例如50ml、75或120ml。反应容器的体积可以低于300ml。可以根据装置的预期用途和预期在一个批次中制备的产物的量或者连续方法中的生产速率来选择反应容器的体积。显然,反应容器的大小设定了一个批次中能够生产的产物的量的上限。
反应容器的宽高比可以是不超过8:1,或不超过7:1、6:1或5:1。如果反应容器过高,则其变得更难通过反应混合物的抽取和返回而引起内容物的充分混合。如果反应容器过宽,则其变得更难确保整个反应容器内一致的温度,尤其当具有较小的反应体积时。混合物的搅拌能够有助于在整个混合物内获得期望的温度,尤其是如果反应混合物被加热至95℃时。具有上述界限内的宽高比的反应容器可以适应于不同量的反应成分。
可用于本发明的实施方式的反应容器2示于图3中的横截面图中。反应容器2包含被夹套25包围的内容器23。温度控制流体循环经过内容器23和夹套25之间的空间26,以控制内容器23的内容物的温度。提供下部开口27和上部开口28以分别连接内容器至选择阀门32b、32a。
内容器23可以具有主要部分,其具有圆筒形式,该圆筒的直径d1为例如20至50mm,优选25至35mm;和渐缩部分24,其连接圆筒部分至下部开口27。内容器的总高度h1可以是150mm至300mm,期望为200mm至250mm。渐缩部分24的高度可以是5至20mm,优选10至15mm。渐缩部分的侧壁相对于内容器的轴线的角度可以是30至60°,优选为约45°。下部开口27的直径d2可以是5至10mm。
反应容器可以具有非恒定的横截面,例如在其整个高度内向下逐渐变细或者具有中央凸出部分。
可以通过使温度控制流体经过外部夹套流至反应容器来控制反应容器的温度。可以通过再循环温度控制装置22将温度控制流体通过入口2a来供给并通过出口2b来移除。再循环温度控制装置通过加热和/或冷却流体来控制温度控制流体的温度。可以使反应容器保持在恒定的温度,例如尽管发生吸热或放热反应,或者可以控制反应容器遵守期望的温度分布。温度控制流体可以是水或油。可以经由提供在装置的底架上的开口16来提供温度控制流体。适当的再循环温度控制装置是美国宾夕法尼亚的julabousa,inc.制造的prestoa30温度控制系统。在本发明的实施方式中,再循环温度控制装置22包含欧姆加热器和/或peltier装置。在一个实施方式中,反应容器的温度可控制到4°至95℃的温度,精确到0.01℃。
可以提供用于合成反应中要用的反应成分的温度受控的储存器4a-4c。反应成分容纳在安装在温度控制装置41a-41c中的注射器42a-42c(如一次性聚丙烯注射器)中。该实施方式中的温度控制装置包括导电材料例如铝的模块,其具有孔,以紧密地接受注射器42a-42c。如果需要的话,可以提供导热性糊以增加模块与注射器之间的导热性。加热/冷却装置,如欧姆加热器或peltier装置,与导热性模块连接,以控制其温度且因此控制储存器42a-42c中储存的反应成分的温度。
可以使用温度控制装置41a-41c以加热或者冷却储存的反应成分。期望将一些反应成分保持在低温,例如约4℃,以防止其降解。引物可以被储存在升高的温度下,以便防止和/或降低引物二聚化(diming)。还可以加热储存器以使模板和引物在添加至反应容器之前变性和退火。像反应容器的主要温度控制那样,可以使温度受控的储存器保持在恒定的温度或遵守期望的温度分布,例如,就在添加至反应容器之前加热反应成分。在一个实施方式中,温度受控的储存器的温度可独立地控制到4℃至95℃,精确到0.01℃。
可以提供一个或多个在线加热器(未示出)以在添加至反应容器之前提高诸如引物的反应成分的温度,例如至95℃。储存器4a-c、5a-g和选择阀门32c之间和/或选择阀门32a和反应容器2之间的管路上提供加热器。
在该具体情况下,温度受控的储存器之一4c包括两个与单一的温度控制模块41c为热接触的储存器注射器42c和42c’。在需要将两种反应成分保持在相同的温度时,或者在对期望的反应需要较大体积的单一反应成分时,这是有用的。
装置还包括多个未提供有温度控制装置的另外的储存器5a-5g。在该实施方式中,有七个这样的储存器,在其他实施方式中,提供更多更少。另外的储存器5a-5g也可以包括注射器,例如玻璃或聚丙烯注射器,其在为空时可以被移除或处理掉。注射器的使用是有利的,因为储存器的体积随着反应成分被抽取而自动地减小。用可折叠的袋或管可以达到类似的效果。如果使用刚性容器,则期望在任何通向大气的通气孔中提供过滤器以防止污染物进入。
方便的是,提供可旋转的支架51以安装储存器5。如果需要大量的反应成分,则装置可以提供有连接至外部储存器的开口。期望的是,规划该装置以使连接储存器至选择阀门的所需的管路的长度最小化。管路可以从储存器拆卸,然后可被连接至与清洗流体或水的供给相连的歧管,以进行冲洗。以这样的方式,整个装置可以被现场清洗冲洗,即,无需被拆卸。
储存器4a-c和5a-g的容量可以是3至10ml,例如约5ml。对于例如缓冲溶液,可以提供一个或多个更大的储存器,其容量为10至500ml,例如约250ml。
可以通过往复式注射泵3a向反应容器供给反应成分。该泵被称为供给泵。这包括注射器31a,例如玻璃或聚丙烯注射器,其由螺线管33a驱动并连接至可控的选择阀门32a,以选择性地连接螺线管至每个储存器和反应容器。为了向反应容器添加受控量的反应成分,首先使用选择阀门32a来连接泵注射器至相关的储存器,然后启动螺线管以将相关量的反应成分抽入注射器。然后,使用选择阀门32a以连接注射器31a至反应容器2,且螺线管反转以驱动注射器内容物进入反应容器。通过该设置可以顺序地向反应容器迅速地添加各种量的期望的反应成分,而不要求多个注射泵和多个进入反应容器的入口。然而,如果期望同时向反应容器添加多种反应成分,则可以提供多个供给泵3a。如果必须保持某些反应成分在其添加至反应容器之前完全分离,则这可以是期望的。
此外,可以使用第二往复式注射泵3b来搅动反应容器的内容物以及在反应结束时或者在反应过程中为取样目的而去除反应混合物或产物。该泵被称为抽取泵且包含由螺线管33b驱动的注射器31b。第二注射泵3b经由第二选择阀门32b连接至反应容器底部的出口。为了搅动反应容器的内容物,使用第二选择阀门32b来连接注射器31b至反应容器,同时从反应容器抽取一定量的反应混合物然后将其返回。可以控制抽取的反应混合物的量以及将其返回的速率,以控制产生的搅动的程度。可以与反应成分的添加相关地在整个反应过程中连续地、定期地、或者在特定时间产生这样的搅动。
可选地或者此外,第二往复式注射泵3b还可以连接至储存器4a并且可以用于将储存器中储存的反应成分供给至反应容器2。
压力传感器82可以提供在与反应容器2和选择阀门32b之间或者选择阀门32b和抽取泵3b之间的管路连接的短突出物上。在从反应容器2抽取物质和向其返回物质的过程中,由传感器82测量的压力指示被转移的混合物的粘度:压力越高,则粘度越高。尤其是,在材料的周期性抽取和返回过程中,压力变化的幅度指示反应混合物的粘度。压力和粘度之间的确切关系取决于多个因素,包括材料抽取和返回的速率、所用管路的直径和反应容器的几何形状。可以通过理论或校正来确定该关系。在一个实施方式中,确切的关系不是必要的,且可以将来自压力传感器82的信号简单地用于检测反应混合物中的变化,而无需知道其确切粘度。在粘度由于产物形成而增加的反应中,来自压力传感器的信号可以用于控制反应。通过将泵激活以基于压力信号以适当的速率或以适当的量从一个或多个储存器递送一种或多种适当的试剂,可以增加产物产率。在其他情况下,压力信号可以指示反应的完成或者反应中的阶段,并且因此被用于引发产物的收集或者添加成分以用于下一阶段。
来自压力传感器82的信号可以用于控制从反应容器抽取和/或返回混合物的速率,以确保混合物中的dna物质不被剪切力破坏。可以控制抽取和/或返回混合物的速率,以确保不超过压力极限。压力极限可以是粘度依赖性的。
为了从反应容器去除物质,例如反应产物,首先通过第二选择阀门32b和被操作以抽出注射器的活塞的螺线管将抽取泵3b连接至反应容器,以便从反应容器2抽取物质。然后通过第二选择泵和被激活以通过出口7喷出收集的物质的螺线管33b将抽取泵连接至出口7。
可以将出口7连接至例如用于反应产物的容器、样品容器、测量装置或任何其他的装置。对于已被移除用于测量目的的样品,还可以在测量之后返回至反应容器.
抽取泵3b可以具有比供给泵3a更大的容量,从而例如整个反应容器可以在一次泵送操作中被清空。为了帮助通过用大体积的清洗溶液和/或去离子水冲洗整个系统来清洗装置,抽取泵3b的这种大容量也是期望的。为此,装置可以提供有另外的大储存器以容纳足量的清洗溶液和去离子水。
可以使用单一的泵来供给反应成分、搅动反应混合物和抽取反应混合物和/或产物。
供给泵3a和/或抽取泵3b可以包括另一形式的泵,例如蠕动泵。
装置还包括废物容器6,其经通气孔62连接至大气,该通气孔62可以提供有单向阀。废物容器6与反应容器2的上部连接,以便为其提供通气孔并防止反应容器2中的压力积累。还提供管路以将废物容器6与选择阀门32a和32b连接,以便允许倾倒来自反应容器的产物以及任何未使用的和不期望的反应成分。废物容器还可以用于在现场清洗和冲洗过程中接受清洗和冲洗流体,以及从管路的填装(priming)接受少量的反应成分,以确保精确地向反应容器递送期望的量。
可以省略废物容器6。在这种情况下,期望向反应容器2提供安全阀以及提供从选择阀门32a、32b至外部排放的连接。
装置的操作被控制器8控制,其与温度控制装置41a-c、选择注射器32a-b、螺线管33a-b、温度控制装置22以及压力传感器82为电连接,以便控制装置的这些部件。控制器8还包括界面81,其允许连接至用于整体控制待进行的方法的外部计算机9。
控制器8可以被预先编程,以实施特定的常用例行程序,例如填装、取样、收集、冲洗和清洗。在一个实施方式中,控制器8简单地将命令从外部计算机9传递至装置的不同部件。所提及的例行程序可以在外部计算机或手动控制下实施。
装置的填装可以包括从每个待用的储存器抽取反应成分,从而储存器和选择阀门之间的管路被各反应成分填充。为了增加递送至反应容器的反应成分的量精确度并降低被污染的可能性,这是期望的。在一个实施方式中,反应混合物的取样包括在反应过程中移除反应成分的预定量至外部容器或传感器。在某些情况下,将样品返回至反应容器是可能的。收集可以包括在分批方法结束时或者在连续方法的适当时间抽取部分或所有的反应产物。冲洗可以包括用纯水冲洗用过的管路和储存器。装置的清洗可以包括用清洗溶液冲洗管路和储存器。
在本发明的实施方式中,将dna模板、至少一种聚合酶、一种或多种引物和核苷酸添加至反应容器,以形成反应混合物。从反应容器抽取反应混合物的一部分或一份并将其返回至反应容器,以搅动反应混合物并确保其混合。在反应结束时,从反应容器抽取dna。在一个实施方式中,从反应容器的下部(优选反应容器最低点处)提供的开口抽取反应混合物。在一个实施方式,被移除的反应混合物的部分占整个反应混合物的50%或以下,期望的是40%或以下,优选30%或以下。在一个实施方式中,被移除的反应混合物的部分占整个反应混合物的5%或更多,期望的是10%或更多,优选20%或更多。
在一个实施方式中,反应混合物的起始体积为10ml至10升,优选10至100ml、100ml至1000ml或1至10升。
将参考若干非限制性实施例来描述本发明。
实施例
材料和方法
使用作用于含有原核端粒酶teln的靶点的ds环状模板的phi29dna聚合酶来进行以下实施例中进行的dna扩增。phi29聚合酶产生环状模板(多联体)的长的线性重复,其可以用原核端粒酶teln(touchlightgenetics)加工成期望的封闭的线性dna产物。通过phi29dna聚合酶产生多联体dna由于其长的长度(据报道为高达77千碱基)而导致反应粘度增加,同时用原核端粒酶teln加工双链多联体dna由于其低得多的长度(在给出的实施例中为2至3千碱基)而导致粘度降低。
在实施例1和3中,在如本文所述并参考图1至3的装置中生产dna。
实施例1:通过压差测量进行反应中产生的dna的在线定量
首先,将装置清洗并净化。将管从10个试剂储存器(42a、42b、42c’、42c、5a至5g)断开,并经由鲁尔(luer)接头再次连接至从含有10%次氯酸钠溶液的120ml储存器进料的10位歧管。类似地,10%次氯酸钠溶液的60ml储存器从选择阀门32b连接至出口7。通过控制选择阀门32a和32b的位置以及螺线管33a和33b分别对5ml玻璃注射器31a和31b的作用,包括反应容器2本身的整个系统完全被次氯酸钠溶液填充,没有死体积。通过每个管抽取最少5ml的溶液。通过对热循环装置22的控制作用,将反应容器(120ml容量)加热至50℃,且使系统保持在该状态30分钟。该操作确保所有进料管和反应容器本身中的所有受污染的dna被完全破坏。
类似地,通过使用适当的程序来控制上文所述的选择阀门和螺线管,通过将次氯酸钠溶液分配到废物容器6而将整个系统清空。在用去离子水置换储存器中的次氯酸钠之后,重复上述过程5次,确保从整个系统完全去除任何残余的次氯酸钠,使得其准备就绪。
然后将管从歧管断开,再次连接至包含具有鲁尔锁定接头的无菌一次性聚丙烯注射器的10个试剂储存器(5ml至20ml容量)。如下文所示,这些注射器以2ml的最小体积含有单独的反应成分。否则,在活塞被完全压下时它们被排空。储存器5d由两个平行的60ml注射器组成,以给出120ml的去离子水总容量。在含有试剂的注射器中,在填充后,通过手动压下活塞直至液体恰好离开注射器出口,排出每个注射器筒中的空气。在该操作过程中,注射器出口在注射器活塞上方保持垂直,以允许排出筒中所有的空气。
表1
通过温度控制装置41c,使分别在储存器42c和42c’中的酶试剂phi29dna聚合酶和焦磷酸酶保持在4℃的稳定条件下。所有其他试剂保持在室温下。
为了实现向反应容器2精确地分配试剂,通过顺序地抽取非常少量的试剂并分配至废物容器6来填装试剂储存器与选择阀门32a和32b之间的管。
为了使dna模板变性并结合寡核苷酸引物,经由选择阀门32a(通过螺线管33a对注射器31a的作用)将来自储存器5a、5b和5c的试剂分配进入封闭的空的5ml注射器(储存器42b)。通过温度控制装置41b来控制该储存器的温度。使用对41b的控制作用,升高其温度至95℃3分钟,然后冷却至30℃。然后经由选择阀门32a将储存器42b中组合的反应成分分配进入反应容器2。以表1中所示的顺序经由选择阀门32a顺序地分配其余的5种反应成分,从而反应容器2中的最终体积为100ml。通过对热循环装置22的控制作用,将反应容器2的温度设定在30℃。不将反应成分暴露于高于设定温度2℃以上的表面温度下。
通过对螺线管33b和随后对混合注射器31b的移动的控制作用来实现反应成分的混合。控制注射器作用,以从反应容器2抽取5ml的反应成分并将其返回至反应容器2。混合是不连续的,其中注射器以29分钟的间隔激活1分钟,抽取/分配速率为55ml/min。
通过真空和压力传感器(modelpx409-2.5cgusbh,omegaengineeringlimited,manchester,m445bd,uk)来测量反应容器2与注射器31b之间的管线中的压力变化。该传感器测量-2.5至+2.5psi的压力差,且经由t型管使用1/8英寸直径的ptfe管线的短节来连接。实时监测来自传感器的输出信号,并将其登录在计算机上,该计算机使用生产商提供的软件来控制反应器。混合注射器的往复式作用产生在正和负值(混合注射器的拉/推冲击)之间振荡的压力传感器输出。对于在每个混合时间点记录的每分钟的压力数据,对于20秒的3组计算最小和最大压力读数。对于每一组,从最大值减去最小值,并计算平均值。在所有其他变量(例如注射器速度、管线材料、内径和长度以及选择阀门的尺寸和几何形状)固定的情况下,峰高的任何变化可以仅由于反应成分的粘度而变化。在压力差计算中,并未补偿可能归因于环境温度和压力波动的非常小的压力变化。用于分析的第一数据点是在大约45分钟之后,以确保系统已被完全平衡。
图4清晰地显示了压力差随着phi29dna聚合酶催化的dna扩增反应的时间而增加。还使用qubittmbrdsdna荧光计测定(lifetechnologies,paisley,uk.)直接分析了从该反应所取的样品中的dna。该测定仅测量双链dna。用原核端粒酶teln来加工反应中产生的多联体dna以产生三种共价封闭的线性dna分子。用peg8000和nacl分别以5%和2m的最终浓度使经加工的dna沉淀。使沉淀的dna在14,000g下离心20分钟,并再次悬浮在等体积的不含核酸酶的去离子水中。
图5清晰地显示,反应过程中压力差信号的增加与dna浓度的增加直接相关。压力和所测量的dna之间的直接线性关系还表明,在该反应期间内,dna以双链形式占主导地位地存在。
实施例2:起始dntp浓度对其整合入dna的效率的影响
在该实验中,在50ml聚丙烯离心管中进行dna扩增反应(5ml体积)。以表2中所示的顺序将成分顺序地添加至管,以给出所示的最终浓度。测试了2mm、4mm和8mm的起始dntp浓度对dna扩增的作用。
表2
以最小的搅动将所有的管在轨道式孵育器中在30℃下孵育61.5小时的时间。该时间段对应于供给有2mmdntp的dna扩增反应完成所需时间的大约10倍。在反应结束时,用原核端粒酶teln加工在每一条件下产生的多联体dna,以产生两种不同的共价封闭的线性dna分子。用peg8000和nacl分别以5%和2m的最终浓度使经加工的dna沉淀。使沉淀的dna在14,000g下离心20分钟,并再次悬浮在5ml不含核酸酶的去离子水中。将该dna产物的样品在去离子水中稀释50倍并加热至75℃1分钟。将10μl该稀释的dna样品与2μl上样染料(noveljuice,newmarketscientificltd.,uk.)混合并将10μl样品施加到0.8%琼脂糖凝胶,并在标准电泳条件下跑胶,直至dna成分被清晰地分开(图6)。
图6中的电泳凝胶清晰地显示了在起始dntp浓度为8mm时两种共价封闭的线性dna产物的产物产率的显著下降。由于反应进行的时间长,所以这表明对dna扩增的非常强的抑制效果。还使用260nm处的吸光度测量而测量了在反应结束时所取的样品中的dna,且结果示于表3中。
表3:在反应结束时总加工的dna的浓度从260nm处的吸光度测量来计算并表示为从不同水平的dntp的最大理论dna产率的百分数。
表3证实了图6中所示的电泳结果,即超过4mm的起始dntp浓度会显著降低phi29dna聚合酶催化的反应中dna的产率。还很显然的是,采用8mm的起始dntp浓度,产物纯度也有显著的降低,其证据是两个主要产物带周围的弥散(图6)。
观察到的由增加dntp的起始浓度导致的dna产率的下降可以归因于对反应的多个影响。这些包括但不限于:酶的底物抑制;li+或核苷酸对dna链相互作用(主要是引物模板相互作用)的影响和/或mg2+与dntp的结合导致的可用于酶活性的游离mg2+水平。dna扩增的抑制可以是由单一因素或者以直接或间接方式起作用的多个因素引起的。清楚的是,不能通过简单地增加dntp的起始浓度而将反应中dna的产率增加至工业生产中认为可接受的水平。
实施例3:将dntp进料至phi29dna聚合酶催化的dna扩增反应的作用
进行实验以比较用另外量的dntp来补充phi29催化的dna扩增反应的作用。如实施例1所述地进行反应,但由260nm处的吸光度测量来计算dna浓度。在以下反应之间进行比较:用2mmdntp且未添加另外的dntp进行的反应(反应1),用2mmdntp且在530min进料另外的2mmdntp进行的反应(反应2),从开始时用4mmdntp进行的反应(未添加另外的dntp)以及用2mmdntp且三次进料2mm浓度的另外的dntp(在500、1430和4340min)进行的反应。对进料任何添加的dntp的触发是压力差别信号的稳定化。这激活从储存器添加适当体积的dntp溶液,使得反应器中dntp的浓度增加2mm。
对于这些反应条件中的每一个,压力差别对时间的作图如图7所示。
如果需要的话,从图7中所示的数据,在dntp添加之前、之间和之后计算初始速率。这些示于表4中。此外,在每个反应结束时,估计dna的最终产率并从所添加的dntp的浓度计算为最大理论产率的百分数。数据示于表5中。
表4:从差别压力测量估计的反应1至4的dna扩增初始速率
表5:从260nm处的吸光度测量计算反应1至4的最终dna产率
表4清晰地显示,增加dntp的浓度显著降低dna被phi29dna聚合酶扩增的速率。对反应速率的影响是累积的,且不取决于何时添加另外的dntp。
在实施例2中显示,起始dntp浓度为8mm的反应仅产生最大理论产率的15%,表明极慢的反应。在表5中,其中以2mm的等份添加8mmdntp的反应(反应4)达到的dna产率为最大理论产率的54%,这是高度显著的改进。尽管逐步进料dntp似乎导致反应速率的渐进式下降,但总速率更快,且比在反应开始时引入高浓度的dntp的产率更高。
表4还显示,在进料dntp的过程中,mg2+离子与dntp磷酸盐基团的摩尔比(mg2+/po43-比例)显著下降,且与观察到的反应速率的下降一致。这在图8中示出。
因此,进料dntp能够在酶催化反应中将dna生产增加至工业可接受的水平。然而,为了达到更高的反应速率,似乎需要通过与dntp一起进料mg2+离子而将mg2+离子与dntppo43-基团的摩尔比保持在最佳的水平。
实施例4:镁离子浓度对将dntp整合入dna中的效率的影响
在该实验中,在50ml聚丙烯离心管中进行dna扩增反应(5ml体积)。将成分预热至30℃并以表6中示出的顺序依序添加至管中,以给出所指示的最终浓度。2.24mm、4.52mm、9mm和11.24mm的mg2+起始浓度与3mm的起始dntp浓度组合分别给出0.25、0.5、1和1.25的mg2+/po43-比例。将管在30℃下孵育19小时,随后添加另外等份的dntp和mgcl2,以增加dntp的反应浓度至6mm且增加mg2+浓度至4.48mm、9.04mm、18mm和22.48mm,以保持mg2+/po43-比例为0.25、0.5、1和1.25。允许反应再进行24小时,此时收集dna产物并估计其浓度。用足量的原核端粒酶teln消化dna并用6%peg8000使dna在含有500mmnacl、100mmmgcl2的缓冲剂中沉淀。将dna再次悬浮在4ml的体积中,并使用260nm吸光度测量对dna进行定量。
表6.反应混合物的成分和添加顺序
表7:保持固定的mg2+/po43-比例的dntp进料反应中的dna产率
表7中的结果显示dntp进料反应中不同的mg2+/po43-比例对43小时后dna产率的影响。0.25的mg2+/po43-比例在该时间后达到较低的dna产率表明由mg2+离子的浓度不足导致的较慢的反应速率或反应的抑制。不论哪一种方式,数据表明,对于用于工业用途的有效dna扩增反应,存在需要保持的最佳的mg2+/po43-比例。数据表明,最佳的比例可以在0.5至1.25之间。数据还表明,如果将dntp进料至反应,则添加另外的mg2+离子以保持最佳的mg2+/po43-比例(理想地为1)允许向dna的有效转化。
实施例5:原核端粒酶teln添加对phi29dna聚合酶催化的用多份dntp进料的dna扩增反应的作用
进行实验以比较用另外量的dntp和原核端粒酶teln补充phi29催化的dna扩增反应对质粒protlx-kn3x2lux的作用(图13)。反应基本上如实施例1所述地进行,但用含有12mmtris-hcl(ph7.4)、90mmnacl、0.1mmedta、1mmdtt、50%甘油中的25μmteln溶液且保持在4℃的储存器42b以及含有去离子水中的100mmmgcl2溶液的储存器5g。模板protlx-k_n3x2lux的起始浓度为5μg/ml。
在反应的分批阶段之后,在300min、600min、900min、1080min、1230min和1440min的所选的时间间隔,通过注射器31a向反应容器2添加各份dntp,以达到2mm的浓度增加。在同一时间间隔且在添加dntp之后立即添加各份mgcl2,以达到3:1的[mg2+]:[dntp]比例。在1050min时,将原核端粒酶teln分配到反应容器2中,给出2μm的最终浓度。
在反应的初始分批阶段(高达300min)之后,在与多联体dna合成对应的差别压力测量存在稳定的增长。在1050min分钟向反应添加原核端粒酶teln之后,在压力改变的速率方面有3.6倍的增加,对应于dna合成的显著增加。现提出,在反应的dntp/mg2+进料阶段期间单次添加teln导致产生封闭的线性dna,其可以用作用于进一步的dna扩增的模板。在该点之后进一步进料mg2+/dntp对于确保观察到的增加的dna合成速率是关键性的。
- 还没有人留言评论。精彩留言会获得点赞!