用于治疗HBV的手段和方法与流程
本发明涉及用于针对乙型肝炎病毒(hbv)治疗或疫苗接种的改进的重组疫苗接种载体以及包含所述重组疫苗接种载体的药物组合物或疫苗。本发明还涉及用于在针对hbv疫苗接种的方法中使用的重组疫苗接种载体,以及用于针对hbv疫苗接种的试剂盒。
背景技术:
超过2.4亿人患有慢性乙型肝炎病毒(hbv)感染。已经批准的治疗选择是核苷(酸)类似物和干扰素α。核苷(酸)类似物控制但不治愈乙型肝炎,并需要昂贵的长期治疗,这可能与耐药病毒的出现有关。干扰素α治疗受到副作用的限制,且仅对15-20%的患者有治愈作用。
该病毒分为四种主要血清型(adr、adw、ayr、ayw),它们根据其包膜蛋白上存在的抗原表位诱导差异性抗体应答,并且根据基因组的总体核苷酸序列变化分为(至少)8种基因型(a-i)。所述基因型具有独特的地理分布并用于追踪病毒的进化和传播。基因型之间的差异影响疾病的严重程度,病程和并发症的可能性,以及对治疗和可能的疫苗接种的应答。此外,存在亚基因型,例如a1-5。在中欧和美国,主要的基因型是a2。然而,只有1%的感染人群携带a2亚基因型,而大多数患者携带b、c或d基因型的hbv。已经表明,与其它(亚)基因型相比,常规hbv疫苗对与包含在疫苗中的hbv抗原相同的(亚)基因型的hbv提供更好的保护。
已充分确定的是,宿主适应性免疫系统对于有效的hbv控制至关重要。针对hbv小表面抗原(hbsag)的中和性抗体阻止病毒扩散至未感染的肝细胞,并且从hbsag至抗-hbs的血清转化代表了hbv感染的临床终点(rehermann和nascimben,natrevimmunol2005;5:215-29)。清除病毒的患者产生强的多克隆和多特异性cd8+和cd4+t细胞应答,而慢性感染与抗病毒t细胞的消耗和进行性功能障碍相关(rehermann等,jexpmed1995;181:1047-58)。临床观察发现,从hbv免疫供体获得骨髓的骨髓移植受者体内的慢性hbv感染消退,这为t细胞控制感染提供了强有力的支持(ilan等,gastroenterology1993;104:1818-21)。基于这些观察,设计治疗性疫苗接种来活化内源性hbv特异性t细胞应答。然而,在慢性感染患者中,许多临床尝试仅对抗-hbv免疫应答具有短暂作用并且不能控制hbv(kutscher等,microbbiotechnol2012;5:270-82)。持续接触高水平的循环病毒抗原似乎是免疫疗法的主要障碍,因为它们诱导耐受性和效应细胞功能障碍。
此外,治疗性疫苗接种与抗病毒治疗(控制病毒血症但对循环抗原水平没有影响)的结合并未改善疫苗功效和临床疗效(michel等,jhepatol2011;54:1286-96)。
为了促进新的免疫治疗策略的发展,已经开发了hbv转基因小鼠(hbvtg),其是垂直传播的慢性hbv感染模型(guidotti等,jvirol1995;69:6158-69)。hbvtg小鼠在肝细胞中复制hbv,产生hbcag、hbsag和乙型肝炎e抗原(hbeag)并将感染性病毒释放到血液中(guidotti等,jvirol1995;69:6158-69)。hbv抗原的表达在出生时开始,小鼠对hbv编码的蛋白质具有免疫耐受性(shimizu等,jimmunol1998;161:4520-9),但是这种耐受性可以被破坏(buchmann等,vaccine2013;31:1197-203)。
修饰的安卡拉牛痘病毒(mva)与痘病毒科正痘病毒属成员牛痘病毒有关。通过在安卡拉牛痘病毒株(cva)的鸡胚成纤维细胞上进行516次连续传代,产生了mva(综述参见mayr,a等,infection3,6-14(1975))。作为这些长期传代的结果,所得到的mva病毒的基因组有约31000个碱基的基因组序列缺失,因此其被描述为高度限制复制成禽类细胞的宿主细胞(meyer,h.等,j.gen.virol.72,1031-1038(1991))。在多种动物模型中显示,得到的mva显著无毒(mayr,a.&danner,k.,dev.biol.stand.41:225-34(1978)),但仍然产生针对痘病毒的保护性免疫应答。因此,这种mva菌株已经在临床试验中作为疫苗进行了测试,以预防人类天花疾病(mayr等,zbl.bakt.hyg.i,abt.org.b167,375-390(1987);stickl等,dtsch.med.wschr.99,2386-2392(1974))。这些研究涉及超过120,000人,包括高风险患者,并且证明,与牛痘基疫苗相比,mva毒力降低且耐受性良好,但仍能诱导良好的特异性免疫应答。
在随后的几十年中,mva被设计用作用于重组基因表达的病毒载体或作为重组疫苗(sutter,g.等,vaccine12:1032-40(1994))。已经描述了用于开发疫苗或药物的mva株。参见美国专利no.6,761,893和no.6,193,752。这些株能够在不同的非人类细胞和细胞系,特别是鸡胚成纤维细胞(cef)中繁殖复制,但不能在已知允许复制其他已知牛痘病毒株的人细胞系中显著繁殖复制。
基于以上所述,本领域需要可用于针对hbv的治疗性疫苗接种的改进的手段和方法。这些手段和方法应平行诱导针对多种hbv亚型的中和性抗体应答和多特异性t细胞应答。
技术实现要素:
本发明涉及用于针对乙型肝炎病毒(hbv)治疗或疫苗接种的新型重组疫苗接种载体以及包含所述mva的药物组合物或疫苗。本发明还涉及一种使用重组mva针对hbv的疫苗接种方法。设想本发明的手段和方法在受体中针对hbv诱导抗体应答以及t细胞应答,并且进一步设想所诱导的免疫应答对多种hbv基因型和血清型有效。
本文所用的“重组疫苗接种载体”是指减毒病毒或细菌。在本文中,“载体”是指用作载体(carrier)的病毒或细菌。通常,这种减毒病毒或细菌用于将编码抗原的核酸引入受试者的细胞。
本发明的重组疫苗接种载体可以是减毒沙门氏菌(salmonella)菌株。然而,可选地,可以使用其它原核微生物,比如大肠杆菌(escherichiacoli)、志贺氏菌(shigella)、耶尔森氏菌(yersinia)、乳酸杆菌(lactobacillus)、分枝杆菌(mycobacteria)、李斯特菌(listeria)或弧菌(vibrio)的减毒菌株。合适的微生物菌株的实例包括鼠伤寒沙门氏菌(salmonellatyphimurium)、伤寒沙门氏菌(salmonellatyphi)、都柏林沙门氏菌(salmonelladublin)、肠炎沙门氏菌(salmonellaenteretidis)、弗氏志贺氏菌(shigellaflexeneri)、索氏志贺氏菌(shigellasonnel)。减毒沙门氏菌菌株是得到充分表征的粘膜疫苗载体中的一种。减毒但仍具有侵袭性的重组沙门氏菌菌株已被用作口服疫苗载体,以携带几种病原体的保护性表位进入粘膜相关淋巴组织,从而诱导针对载体和外源抗原的粘膜、系统和ctl免疫应答。
此外,本发明的重组疫苗接种载体可以是减毒沙门氏菌菌株、cmv-基载体、vsv-基载体、腺病毒载体或麻疹载体。
本发明的重组疫苗接种载体可以是病毒载体,其可以是具有感染性的病毒颗粒,其也是用于将基因导入细胞的载体。本领域技术人员熟知病毒疫苗接种载体。本发明特别考虑的是痘病毒载体。重组痘病毒可以是通过标准基因工程方法产生的痘病毒。在一个优选的实施方案中,本发明的重组疫苗接种载体是mva。
mva特别适合作为载体系统。已经证实了mva在针对例如小痘病毒的疫苗接种后在短时间内有效诱导体液和细胞免疫应答的效力。mva的安全性已建立。甚至mva在免疫受损的受试者中诱导免疫应答的效力也是已知的。
然而,除了mva用作hbv疫苗确实具有的优点之外,hbv疫苗最大的挑战是它既能够诱导体液也能够诱导细胞免疫应答,并且该免疫应答优选地针对hbv的多种基因型和/或血清型。载体的选择、疫苗接种方案和hbv抗原都可能有助于疫苗接种的有效性。
本发明涵盖使用来自不同hbv基因型和/或血清型的hbv抗原的组合。与仅包含来自单一hbv基因型的抗原的疫苗相比,这种组合不仅导致针对多种hbv株的广泛免疫应答,而且导致针对每种hbv基因型的更强的免疫应答。事实上,使用本发明的手段和方法,发明人甚至能够克服具有低、中和甚至高抗原水平的受试者的免疫耐受性。
由于若干原因,基于mva的疫苗是有利的。例如,优选的mva株mva-f6(sutter和staib,2003.curr.drugtargetsinfect.disord.3:263-271)在原代鸡胚成纤维细胞(cef)中生长良好,但不在人细胞中复制。在人细胞中,表达病毒基因以及工程化转基因,但不产生感染性病毒。mva的受限宿主范围可以解释包括人在内的广范围哺乳动物物种体内观察到的非毒性表型。在许多毒性研究中已经表明mva是安全的。本领域已经描述了重组mva的构建、生产和使用,例如在wo97/02355,sutter和staib,2003(同上)和wo2003/008533中。
本发明设想了表达来自乙型肝炎病毒(hbv)的一种或多种抗原的重组疫苗接种载体,优选修饰的安卡拉牛痘病毒(mva)。所述一种或多种抗原可以选自来自hbv的包膜蛋白(hbs-抗原)、来自hbv的核心蛋白(hbc-抗原)或来自hbv的聚合酶的rt结构域。所述包膜蛋白可以是例如hbvadw血清型的包膜蛋白,比如hbva基因型adw血清型、比如hbva2基因型adw2血清型的包膜蛋白。所述核心蛋白可以是例如hbvayw血清型的核心蛋白,比如hbvd基因型ayw血清型的核心蛋白。
本发明的重组疫苗接种载体,优选mva可以表达超过一种包膜蛋白,其中所述超过一种包膜蛋白可以来自hbv的不同基因型或血清型。例如,本发明的mva可表达hbva基因型adw血清型的包膜蛋白以及c基因型的包膜蛋白。类似地,本发明的mva可以表达超过一种核心蛋白,其中所述超过一种核心蛋白可以来自hbv的不同基因型或血清型。例如,本发明的mva可以表达hbvd基因型ayw血清型的核心蛋白以及c基因型的核心蛋白。
本发明的重组疫苗接种载体,优选mva也可表达来自hbv的聚合酶或所述hbv聚合酶的rt结构域。该聚合酶可以是任何基因型或血清型。所述聚合酶优选可以为来自hbvd基因型的聚合酶。
本发明的重组疫苗接种载体,优选mva可以表达(a)来自乙型肝炎病毒adw血清型的包膜蛋白(hbs-抗原),其中所述包膜蛋白优选为来自乙型肝炎病毒a基因型adw血清型的大包膜或小包膜蛋白,优选为小包膜蛋白;和(b)来自乙型肝炎病毒ayw血清型的核心蛋白(hbc-抗原),其中所述核心蛋白优选来自乙型肝炎病毒d基因型ayw血清型;和以下中的至少一个:(c)来自乙型肝炎病毒的包膜蛋白(hbs-抗原),其与seqidno:1所示的氨基酸序列具有至少90%的序列同一性,(d)来自乙型肝炎病毒的核心蛋白(hbc-抗原),其与seqidno:2所示的氨基酸序列具有至少90%的序列同一性,和/或(e)来自乙型肝炎病毒的聚合酶的rt结构域,其与seqidno:3所示的氨基酸序列具有至少90%的序列同一性。可以理解,本发明的mva可表达三种,优选四种或最优选五种hbv抗原,比如抗原(a)、(b)、(c),抗原(a)、(b)、(d),或抗原(a)、(b)、(e),抗原(a)、(b)、(c)、(d),抗原(a)、(b)、(c)、(e),抗原(a)、(b)、(d)、(e),或抗原(a)、(b)、(c)、(d)、(e)。
如本文所用,hbv的“包膜蛋白”、“hbs-抗原”或“hbsag”是指乙型肝炎病毒(hbv)的表面抗原。所述“包膜蛋白”、“hbs-抗原”或“hbsag”可能与三种变体即由不同的mrna翻译而来的小、中和大包膜蛋白中的任何一种有关。所有三种变体的共同之处在于位于s基因的主亲水区(mhr)内的密码子位置124至147处的“a”决定簇表位。这种“a”决定簇是急性乙型肝炎初始免疫应答过程中抗-hbs抗体的主要靶标之一。可选地,术语“包膜蛋白”、“hbs-抗原”或“hbsag”可涉及所述包膜蛋白的免疫原性片段。包膜蛋白的免疫原性片段涉及来源于n-末端和/或c-末端缩短的,即缺少n-末端和/或c-末端氨基酸中的至少一个的任何全长包膜蛋白的蛋白质或肽。这种片段包含包膜蛋白的一级序列的优选至少70、优选至少80、优选至少90、优选至少100、更优选至少125、最优选至少150个连续氨基酸,并且这种片段通常是免疫原性的。通常,与全长蛋白相比,这种免疫原性片段至少包含对应于所述小包膜蛋白的氨基酸位置的氨基酸99至168。
如本文所用,hbv的“核心蛋白”或“hbc-抗原”是指核衣壳的结构蛋白。全长核心蛋白长度为183个氨基酸,并由装配结构域(氨基酸1至149)和核酸结合结构域(氨基酸150至183)组成。34个残基的核酸结合结构域是非常碱性的,具有17个精氨酸,与其功能一致。核心蛋白在溶液中是二聚体的,这些二聚体自组装成二十面体衣壳。应该理解,仅包含氨基酸1至149的截短的核心蛋白也能够形成衣壳。任选地,术语“核心蛋白”或“hbc-抗原”可涉及所述核心蛋白的免疫原性片段。核心蛋白的免疫原性片段涉及来源于n-末端和/或c-末端缩短,即缺少n-末端和/或c-末端氨基酸中的至少一个的任何全长核心蛋白的蛋白质或肽。这种片段包含核心蛋白的一级序列的优选至少100、更优选125、最优选149个或更多个连续氨基酸,并且这种片段通常是免疫原性的。通常,与全长核心蛋白相比,这种免疫原性片段至少包含对应于seqidno:11所示序列位置的氨基酸18至143。一个典型实例是由seqidno:11所示的全长核心蛋白的氨基酸1至149组成的核心蛋白片段。
如本文所用,“免疫原性”是指特定物质,比如抗原或表位在人体或动物体内引起免疫应答的能力。换言之,免疫原性是诱导体液和/或细胞介导的免疫应答的能力。抗原引发免疫应答的能力被称为免疫原性,其可以是体液和/或细胞介导的免疫应答。不受理论束缚,认为来自hbv的任何天然存在的hbs-抗原、hbc-抗原或聚合酶是免疫原性的。此外,与天然存在的序列相比,认为来自hbv的天然存在的hbs-抗原、hbc-抗原或聚合酶的许多变体中的一个或多个氨基酸交换、缺失或插入。作为示例性实例,天然存在的hbs-抗原的免疫原性变体是hbs-抗原,其中“a”决定簇表位已被另一血清型的hbs-抗原的“a”决定簇替换。通常,天然存在的hbs-抗原、hbc-抗原或聚合酶的免疫原性变体可与天然存在的hbs-抗原、hbc-抗原或聚合酶的氨基酸序列具有90%的序列同一性。
本发明的重组疫苗接种载体,优选mva包含来自乙型肝炎病毒adw血清型的包膜蛋白。这种包膜蛋白可以是小、中或大包膜蛋白,其中小或大包膜蛋白是优选的,小包膜蛋白是最优选的。优选地,所述包膜蛋白为a基因型,adw血清型(a/adw),优选地为hbva2/adw2。然而,任何具有hbvadw的天然存在的包膜蛋白的免疫原性的天然存在的或工程化的包膜蛋白均适用于本发明。优选的包膜蛋白包含序列,所述序列与seqidno:08所示的氨基酸序列具有至少约90%、优选地至少约91%、优选地至少约92%、优选地至少约93%、优选地至少约94%、优选地至少约95%、优选地至少约96%、优选地至少约97%、优选地至少约98%、优选地至少约99%、优选地约100%的序列同一性,或者优选地包含seqidno:08所示的氨基酸序列或由seqidno:08所示的氨基酸序列组成。所述包膜蛋白在n-或c-末端还可以具有另外的氨基酸。优选地,所述包膜蛋白在n-和/或c-末端具有30或更少、优选地25或更少、优选地20或更少、优选地15或更少、优选地10或更少、优选地5或更少、优选地4或更少、优选地3或更少、优选地2或更少、优选地1或更少的另外的氨基酸。优选地,这些氨基酸为kim等2011,plosone,6(4):e18556中描述的自切割位点的片段,比如p2a或t2a。因此,任选地,所述包膜蛋白在n-末端包含另外的脯氨酸和/或在其c-末端包含另外的序列gsgatnfsllkqagdveenpg(seqidno:09)。因此,所述包膜蛋白可以具有seqidno:10所示的序列。
本发明的重组疫苗接种载体,优选mva包含来自乙型肝炎病毒ayw血清型的核心蛋白。优选地所述核心蛋白为hbvd基因型/ayw。任何具有hbvayw的天然存在的核心蛋白的免疫原性的天然存在的或工程化的包膜蛋白均适用于本发明。在优选的实施方案中,所述核心蛋白是由全长蛋白的氨基酸1至149组成的全长核心蛋白的片段。优选的核心蛋白包含序列,所述序列与seqidno:11所示的氨基酸序列具有至少约90%、优选地至少约91%、优选地至少约92%、优选地至少约93%、优选地至少约94%、优选地至少约95%、优选地至少约96%、优选地至少约97%、优选地至少约98%、优选地至少约99%、优选地约100%的序列同一性,或者优选地包含seqidno:11所示的氨基酸序列或由seqidno:11所示的氨基酸序列组成。所述核心蛋白在n-或c-末端还可以具有另外的氨基酸。优选地,所述核心蛋白在n-和/或c-末端具有30或更少、优选地25或更少、优选地20或更少、优选地15或更少、优选地10或更少、优选地5或更少、优选地4或更少、优选地3或更少、优选地2或更少、优选地1或更少的另外的氨基酸。优选地,这些氨基酸为自切割位点的片段,比如p2a或t2a。因此,任选地,所述核心蛋白在n-末端包含另外的脯氨酸和/或在其c-末端包含另外的seqidno:12所示的序列。认为由全长氨基酸1至149组成且具有如本文所述的c末端序列的核心蛋白片段仍然具有组装成衣壳的能力,并且认为另外的c-末端序列不会干扰衣壳形成。
本发明的重组疫苗接种载体,优选mva可以包含来自乙型肝炎病毒的免疫原性包膜蛋白(hbs-抗原),所述免疫原性包膜蛋白与seqidno:01所示的氨基酸序列具有至少约90%、优选地至少约91%、优选地至少约92%、优选地至少约93%、优选地至少约94%、优选地至少约95%、优选地至少约96%、优选地至少约97%、优选地至少约98%、优选地至少约99%、优选地约100%的序列同一性。seqidno:01是c基因型株的大包膜蛋白的共有序列,其根据代表hbv株世界分布的500个hbv序列的比对产生。与seqidno:04所示的氨基酸序列具有至少约90%序列同一性、与seqidno:1具有99%序列同一性的免疫原性包膜蛋白的示例性实例对应于这样的序列,该序列来源于具有genbank登录号abv02797(2007年9月12日的版本abv02797.1gi:157057635)的hbv的推定前-s蛋白,其中通过交换2个氨基酸将a-决定簇(抗-hbs抗体的最重要的靶序列)修饰为ayw(用于形成针对ayw血清型的抗体)。与seqidno:04具有基本上相同序列的hbv-株的存在确保了基本过程(折叠/加工/呈递)对于编码的蛋白质是有功能的。因此,本发明还设想所述与seqidno:1所示氨基酸序列具有至少约90%的序列同一性的免疫原性包膜蛋白是这样一种包膜蛋白,其与seqidno:04所示的氨基酸序列具有至少约90%、优选地至少约91%、优选地至少约92%、优选地至少约93%、优选地至少约94%、优选地至少约95%、优选地至少约96%、优选地至少约97%、优选地至少约98%、优选地至少约99%、优选地约100%的序列同一性。此外,c基因型株的包膜蛋白的共有序列与b基因型株的包膜蛋白的共有序列高度相似。由于免疫原性包膜蛋白是基于c基因型株的包膜蛋白的共有序列的,其同时非常类似于b基因型株的包膜蛋白的共有序列,因此该免疫原性包膜蛋白能够针对至少b基因型和c基因型株的广谱而诱导免疫应答。应当理解,任选地,所述与seqidno:1所示的氨基酸序列具有至少约90%的序列同一性的免疫原性包膜蛋白也指该免疫原性包膜蛋白的免疫原性片段,其与seqidno:1的相应的片段具有至少90%、优选地至少约91%、优选地至少约92%、优选地至少约93%、优选地至少约94%、优选地至少约95%、优选地至少约96%、优选地至少约97%、优选地至少约98%、优选地至少约99%、优选地约100%的序列同一性。所述包膜蛋白在n-或c-末端还可以具有另外的氨基酸。优选地,所述包膜蛋白在n-和/或c-末端具有30或更少、优选地25或更少、优选地20或更少、优选地15或更少、优选地10或更少、优选地5或更少、优选地4或更少、优选地3或更少、优选地2或更少、优选地1或更少的另外的氨基酸。优选地,这些氨基酸为自切割位点的片段,比如p2a或t2a。因此,任选地,所述包膜蛋白在n-末端包含另外的脯氨酸和/或在其c-末端包含另外的序列gsgegrgslltcgdveenpg(seqidno:13)。因此,所述包膜蛋白可以具有seqidno:14所示的序列。
本发明的重组疫苗接种载体,优选mva还可包含来自乙型肝炎病毒的免疫原性核心蛋白(hbc-抗原),所述免疫原性核心蛋白与seqidno:2所示的氨基酸序列具有至少约90%、优选地至少约91%、优选地至少约92%、优选地至少约93%、优选地至少约94%、优选地至少约95%、优选地至少约96%、优选地至少约97%、优选地至少约98%、优选地至少约99%、优选地约100%的序列同一性。seqidno:2是c基因型株的核心蛋白的共有序列,其根据代表hbv株世界分布的500个hbv序列的比对产生。与seqidno:05所示氨基酸序列具有至少约90%序列同一性、与seqidno:2具有100%序列同一性的免疫原性核心蛋白的示例性实例基本上与这样的序列相同,该序列来源于具有genbank登录号np_647607(2015年7月9日的版本np_647607.1gi:21326588)的hbv的衣壳蛋白。因此,本发明还设想所述与seqidno:2所示氨基酸序列具有至少约90%的序列同一性的免疫原性核心蛋白是这样一种包膜蛋白,其与seqidno:05所示的氨基酸序列具有至少约90%、优选地至少约91%、优选地至少约92%、优选地至少约93%、优选地至少约94%、优选地至少约95%、优选地至少约96%、优选地至少约97%、优选地至少约98%、优选地至少约99%、优选地约100%的序列同一性。此外,c基因型株的共有序列与b基因型株的共有序列相同。由于免疫原性包膜蛋白是基于c基因型株的包膜蛋白共有序列的,其与b基因型株的包膜蛋白的共有序列相同,因此该免疫原性包膜蛋白能够针对至少b基因型和c基因型株的广谱而诱导免疫应答。应当理解,任选地,所述与seqidno:2所示的氨基酸序列具有至少约90%的序列同一性的免疫原性核心蛋白也指该免疫原性核心蛋白的免疫原性片段,其与seqidno:2的相应的片段具有至少90%、优选地至少约91%、优选地至少约92%、优选地至少约93%、优选地至少约94%、优选地至少约95%、优选地至少约96%、优选地至少约97%、优选地至少约98%、优选地至少约99%、优选地约100%的序列同一性。作为示例性实例,这种免疫原性片段可以是与seqidno:2的氨基酸1至149具有90%序列同一性的片段。所述核心蛋白在n-或c-末端还可以具有另外的氨基酸。优选地,所述核心蛋白在n-和/或c-末端具有30或更少、优选地25或更少、优选地20或更少、优选地15或更少、优选地10或更少、优选地5或更少、优选地4或更少、优选地3或更少、优选地2或更少、优选地1或更少的另外的氨基酸。优选地,这些氨基酸为自切割位点的片段,比如p2a或t2a。因此,任选地,所述核心蛋白在n-末端包含另外的脯氨酸。任选地,所述核心蛋白还可进一步在其c-末端包含seqidno:09或13所示的另外的序列。因此,所述核心蛋白可以具有seqidno:15所示的序列。
本发明的重组疫苗接种载体,优选mva还可包含来自乙型肝炎病毒的聚合酶的免疫原性rt结构域,所述聚合酶的免疫原性rt结构域与seqidno:3所示的氨基酸序列具有至少约90%、优选地至少约91%、优选地至少约92%、优选地至少约93%、优选地至少约94%、优选地至少约95%、优选地至少约96%、优选地至少约97%、优选地至少约98%、优选地至少约99%、优选地约100%的序列同一性。seqidno:3是a、b、c和d基因型株的rt结构域的共有序列,其根据代表hbv株世界分布的500个hbv序列的比对产生。聚合酶的这种免疫原性rt结构域的示例性实例是具有seqidno:6所示氨基酸序列的蛋白,seqidno:6与seqidno:3具有97%的序列同一性。seqidno:6对应于一种聚合酶的rt结构域的氨基酸序列,所述聚合酶来源于具有genbank登录号afy09280(2013年1月31日的版本afy09280.1gi:425891330)的hbv的部分聚合酶。由于seqidno:6是与所产生的a、b、c和d基因型的共有序列具有高度相似性的聚合酶的天然存在的rt结构域,因此,seqidno:6是一个典型实例。具有这一序列的hbv-株的存在确保了基本过程(折叠/加工/呈递)对于编码的rt-结构域是有功能的。因此,本发明还设想所述与seqidno:3所示氨基酸序列具有至少约90%的序列同一性的免疫原性rt结构域是这样一种包膜蛋白,其与seqidno:06所示的氨基酸序列具有至少约90%、优选地至少约91%、优选地至少约92%、优选地至少约93%、优选地至少约94%、优选地至少约95%、优选地至少约96%、优选地至少约97%、优选地至少约98%、优选地至少约99%、优选地约100%的序列同一性。由于所述聚合酶的rt结构域包含若干高度保守的表位和由于包含在本发明的mva中的所述聚合酶的rt结构域是基于a、b、c和d基因型株的rt结构域共有序列的,因此,聚合酶的这种免疫原性rt结构域能够针对至少a、b、c和d基因型株的广谱而诱导免疫应答。本发明还设想所述聚合酶的rt结构域可包含在全长聚合酶中。因此,全长聚合酶或包含所述免疫原性rt结构域的截短的全长聚合酶也涵盖在本发明的范围内。应当理解,任选地,所述与seqidno:3所示的氨基酸序列具有至少约90%的序列同一性的聚合酶的免疫原性rt结构域也指该免疫原性rt结构域的免疫原性片段,其与seqidno:3的相应的片段具有至少约90%、优选地至少约91%、优选地至少约92%、优选地至少约93%、优选地至少约94%、优选地至少约95%、优选地至少约96%、优选地至少约97%、优选地至少约98%、优选地至少约99%、优选地约100%的序列同一性。所述聚合酶的rt结构域在n-或c-末端还可以具有另外的氨基酸。优选地,所述rt结构域在n-和/或c-末端具有30或更少、优选地25或更少、优选地20或更少、优选地15或更少、优选地10或更少、优选地5或更少、优选地4或更少、优选地3或更少、优选地2或更少、优选地1或更少的另外的氨基酸。优选地,这些氨基酸为自切割位点的片段,比如p2a或t2a。因此,任选地,所述rt结构域在n-末端包含另外的脯氨酸。任选地,所述核心蛋白还可进一步在其c-末端包含seqidno:13所示的另外的序列。因此,所述rt结构域可以具有seqidno:16所示的序列。
如本文所述的重组疫苗接种载体,优选mva,还可以包含或进一步表达cd70分子。优选地,所述cd70是人cd70。
cd70是cd27的配体,其也被称为cd27l。它是一种ii型跨膜蛋白并在高度活化的淋巴细胞(如在t-和b-细胞淋巴瘤中)上表达。此外,它在肾细胞癌上表达并被评估为肿瘤靶标。cd70配体表达通常被严格调控,然而当cd27l在b细胞上组成型表达时,会形成广泛和有效的记忆样t细胞库(arens,r.等,2004jexpmed199(11)1595-605)。在慢性病毒感染中,cd70信号传导可能与结果有关。
cd27/cd70是以其t细胞形状特性而众所周知的肿瘤坏死因子受体/肿瘤坏死因子(“tnfr/tnf”)超家族的成员。在cd27/cd70中,该家族包括cd30/cd30l、cd40/cd40l、ox40/ox40l、4-1bb/4-1bbl、gitr/gitrl和fas/fasl。已经在广泛的传染病模型中研究了cd70,以及其他比如ox40l和4-1bbl对原发性和继发性t-细胞应答的作用[hendrick等,“cd27isrequiredforgenerationandlong-termmaintenanceoft-cellimmunity,”natureimmunol.1(5):433-440(2000);matter等,“virus-inducedpolyclonalb-cellactivationimprovesprotectivectlmemoryviaretainedcd27expressiononmemoryctl,”eur.j.immunol.35(11):3229-3239(2005);a.schildknecht等,“primingofcd8+t-cellresponsesbypathogenstypicallydependsoncd70-mediatedinteractionswithdendriticcells,”eur.j.immunol.37(3):716-728(2007)]。有趣的是,通过组合的tlr/cd40刺激可以诱导共刺激分子(包括树突细胞(“dc”)上的cd70等)的上调[sanchez等,“combinedtlr/cd40stimulationmediatespotentcellularimmunitybyregulatingdendriticcellexpressionofcd70invivo,”j.immunol.178(3):1564-1572(2007)]。
因此,本发明还考虑cd70与seqidno:26所示的氨基酸序列具有至少约90%、优选地至少约91%、优选地至少约92%、优选地至少约93%、优选地至少约94%、优选地至少约95%、优选地至少约96%、优选地至少约97%、优选地至少约98%、优选地至少约99%、优选地约100%的序列同一性。
本发明的重组疫苗接种载体,优选mva针对广泛的hbv谱的治疗性疫苗接种进行了优化,并包含来自若干常见hbv基因型的抗原序列。因此该mva能够诱导针对不同基因型和血清型的hbv的免疫应答。本发明的发明人惊讶地发现,与仅包含hbvd/ayw抗原的mva疫苗相比,来自hbva/adw的hbs-抗原与来自hbvd/ayw的hbc-抗原的组合不仅诱导针对a和d基因型的hbv的广泛免疫应答,而且还诱导针对hbvd/ayw的甚至更强的t细胞应答。此外,本发明的mva还可表达基于c基因型hbv株的共有序列的hbs-抗原或hbc-抗原。所述c基因型hbv株的这些共有序列还与b基因型株的共有序列高度相似或甚至相同。这意味着包含这些序列中的一个或两个的本发明的mva将进一步诱导针对至少b基因型和c基因型株的广泛免疫应答。另外,本发明的mva可以表达聚合酶的rt结构域,所述聚合酶的rt结构域基于a、b、c和d基因型株的共有序列并包含高度保守的抗原。因此,将该rt结构域引入mva将进一步促进诱导针对至少a、b、c和d基因型的hbv的广泛免疫应答。因此,本发明的mva是针对至少a、b、c和d基因型的mbv的有效疫苗。
术语“抗原”是指含有一个或多个刺激宿主免疫系统以产生细胞抗原特异性免疫应答或体液抗体应答的表位的分子。抗原可以包括蛋白质、多肽、抗原蛋白片段等。此外,所述抗原可以来自任何已知的病毒、细菌、寄生虫、朊病毒、植物、原生动物或真菌,并且可以是完整的生物体。所述术语还包括肿瘤抗原。合成抗原,比如多表位、侧翼(flanking)表位以及其他重组或合成来源的抗原也包括在本申请中。在优选的实施方案中,本发明中的抗原是多肽或蛋白质。
关于术语“表位”,术语“抗原”是指(较长的)序列,特别是(较长的)氨基酸序列或蛋白质序列,而短语“抗原表位”或“抗原的表位”涵盖来自较长序列的一段较短的序列。因此,术语“抗原”涵盖表位。所述术语“抗原”还包括如本文所述的蛋白质、多肽和抗原蛋白片段的变体。而且,所述术语“抗原”涵盖与天然序列相同的序列以及对天然序列的修饰,例如缺失、添加,插入和置换。优选地,抗原变体与参考抗原(即所述抗原的来源抗原)具有至少约50%、至少约60%或65%、至少约70%或75%、至少约80%、81%、82%、83%、84%、85%、86%、87%、88%或89%,更典型地至少约90%、91%、92%、93%或94%,甚至更典型至少约95%、96%、97%、98%或99%,最典型的至少约99%的氨基酸同一性。
表位,在本文中也称为“抗原表位”,形成仍然在宿主中引起免疫应答的抗原的一部分。然而,表位不限于其来源抗原的确切序列。因此,术语“表位”涵盖与天然序列相同的序列以及对天然序列的修饰,例如缺失、添加、插入和置换。优选地,表位变体与参考表位(即所述表位的来源表位)具有至少约50%、至少约60%或65%、至少约70%或75%、至少约80%、81%、82%、83%、84%、85%、86%、87%、88%或89%,更典型地至少约90%、91%、92%、93%或94%,甚至更典型至少约95%、96%、97%、98%或99%,最典型的至少约99%的氨基酸同一性。
用于确定两条核酸和氨基酸之间的序列同一性的技术是本领域已知的。可以通过确定两条或更多条序列(多核苷酸或氨基酸)的“同一性百分比”来比较它们。两条序列(无论是核酸还是氨基酸序列)的同一性百分比是两个比对序列之间的精确匹配的数量除以较短序列的长度并乘以100。
关于本文所述的抗原或表位的“氨基酸序列同一性百分比(%)”定义为在进行序列比对并引入间隔(如果需要)以实现最大序列同一性百分比,而不考虑任何保守性置换作为序列同一性的一部分之后,候选序列中与参考序列(即所述抗原或表位的来源抗原或表位)中的氨基酸残基相同的氨基酸残基的百分比。为了确定氨基酸序列同一性百分比的比对可以以本领域技术范围内的各种方式来实现,例如,使用公共可用的计算机软件比如blast、align或megalign(dnastar)软件。本领域技术人员可以确定用于测量比对的合适参数,包括在被比较的序列的全长上实现最大比对的任何算法。
加以必要修改,上述情况同样适用于“核苷酸序列同一性百分比(%)”。
例如,通过smith和waterman,(1981),advancesinappliedmathematics2:482-489的局部同源算法来提供核酸序列的适当比对。可通过使用由dayhoff,atlasofproteinsequencesandstructure,m.o.dayhoffed.,5suppl.3:353-358,nationalbiomedicalresearchfoundation,washington,d.c.,usa研发,并且由gribskov(1986),nucl.acidsres.14(6):6745-6763标准化的评分矩阵对氨基酸序列应用此算法。geneticscomputergroup(madison,wis.)在“bestfit”应用程序中提供了此算法用于测定序列的同一性百分比的示例性实现方式。wisconsinsequenceanalysispackageprogrammanual,版本8(1995)(可得自geneticscomputergroup,madison,wis.)中描述了此方法的缺省参数。在本发明的上下文中建立同一性百分比的优选方法为使用由爱丁堡大学(universityofedinburgh)拥有版权、由johnf.collins和shanes.sturrok研发并且由intelligenetics,inc.(mountainview,calif)分销的mpsrch程序包。由此程序包套装可采用史密斯-沃特曼算法(smith-watermanalgorithm),其中在评分表中使用缺省参数(例如间隔开口罚分为12、间隔延伸罚分为一并且间隔为六)。产生“匹配”值的数据反映“序列同一性”。其它适合的用于计算序列之间的同一性或相似性百分比的程序通常为本领域中已知的,例如另一比对程序为与缺省参数一起使用的blast。例如,可使用采用以下缺省参数的blastn和blastp:遗传密码=标准;过滤器=无;链=两者;截止值=60;预期=10;矩阵=blosum62;描述=50个序列;排序依据=高得分;数据库=非冗余,genbank+embl+ddbj+pdb+genbankcds翻译+swiss蛋白质+spupdate+pir。这些程序的细节可见于以下网址:http://www.ncbi.nlm.gov/cgi-bin/blast。
编码本发明的mva中的hbv抗原的核酸还可包含在单独的表达盒中,或者优选地全部一起包含在单个表达盒中。本文所用术语“表达盒”涵盖了能够定向表达合适宿主细胞中特定核苷酸序列的dna和rna序列。一般而言,所述表达盒包括与目的多核苷酸可操作地连接的启动子,其任选的与终止信号和/或其他调控元件可操作地连接。所述表达盒可包括转录调控核苷酸序列。表达盒还可以包括核苷酸序列适当翻译所需的序列。所述表达盒还可以是天然存在的,但以用于异源表达的重组形式获得的表达盒。编码区通常编码目标蛋白。包含目标多核苷酸序列的表达盒可以是嵌合的,意指其组分中的至少一个相对于该表达盒的其它组分中的至少一个是异源的。通常,本文的表达盒在mva基因组中不是天然存在的(即,异源的或外源的或外来的)并且能够在感染的细胞中转录。
表达盒中的核苷酸序列的表达可处于组成型启动子或诱导型启动子的控制下,后者仅当宿主细胞暴露在一些特定的外部刺激下时才启动转录。在本发明的表达盒中,所述启动子优选为痘病毒启动子。这种痘病毒启动子可以是天然存在的启动子或合成启动子。作为示例性实例,所述痘病毒启动子是pr7.5启动子、杂合早期/晚期启动子、prs启动子、合成或天然早期或晚期启动子(比如wo2010/102822或wo2005/054484中描述的启动子之一),或牛痘病毒ati启动子。例如,所述痘病毒启动子是p7.5(seqidno:17)(endo等,jgenvirol.1991mar;72(pt3):699-703)。然而,优选的启动子是比p7.5更强的启动子,例如在us2011/0064769中描述的具有seqidno:18所示的序列的启动子ph5。
优选地,编码所述抗原或其表位的核酸序列是密码子优化的。“密码子优化的”核酸序列是指含有被所需宿主细胞(优选人宿主细胞)优选的密码子替换的密码子的核酸序列。将核酸序列转化为具有相同翻译多肽序列但具有可选密码子使用(特别是使用靶生物体的最常见密码子)的密码子优化的核酸序列。产生抗原的密码子优化的核酸序列的方法通常包括:识别天然存在的抗原序列中通常与靶生物体中的高表达基因无关的密码子并用已知广泛应用于靶生物体的基因表达中的密码子替代它们。与在所需宿主细胞中的天然存在的序列相比,密码子优化的核酸序列可以表现出改进的表达。技术人员可以检测密码子优化序列相比于非优化序列是否会诱导蛋白质产量的提高。
密码子优化避免了对期望的宿主使用稀有密码子,因为稀有密码子可阻断或减少编码蛋白质的表达。此外,优选地避免了可以引入用于期望宿主的核酸信号的置换。此信号包括但不限于剪接信号、终止信号和起始信号。优选地,根据所用载体的类型避免以下序列基序,如在许多其它载体中不需要避免的牛痘病毒早期转录终止信号、内部tata-盒、chi-位点和核糖体进入位点;富含-at和富含-gc的序列段;are、ins和crs序列元件;重复序列和rna二级结构;(隐藏的)剪接供体位点和接受体位点,和分支点;和牛痘早期转录终止信号:(tttttnt)。
用于密码子优化的技术是本领域中已知。用不同核苷酸置换核苷酸是指用其他核苷酸技术或人工替代核苷酸。优选地,置换的核苷酸不改变编码的氨基酸序列。可通过如下方式实施置换:识别两个同源核苷酸序列中编码相同氨基酸的密码子并改变所述两个同源核苷酸序列中的一条序列中的密码子,使得该密码子仍编码相同的氨基酸。所述改变可以在一个、两个或全部同源核苷酸序列中进行。
本发明设想所有hbv抗原优选地翻译为一条包含若干抗原的单一多肽链。在该多肽链上,抗原序列之间优选通过自切割位点序列隔开。因此,所述包含若干抗原的多肽链在翻译后将被切割成多条多肽链,其中所述多条多肽链中的每一条可包含单一hbv抗原。这种方法的优点是所有hbv抗原都以约等摩尔水平表达。对于包含若干抗原的多肽链,一种优选的排列是-从n-端至c-端-来自hbva/adw的包膜蛋白、p2a位点、来自hbvd/ayw的核心蛋白、p2a位点、如本文所述的来自乙型肝炎病毒的聚合酶的免疫原性rt结构域、t2a位点、如本文所述的来自乙型肝炎的免疫原性包膜蛋白(hbs-抗原)、t2a位点、如本文所述的来自乙型肝炎的免疫原性核心蛋白(hbs-抗原)。在优选的实施方案中,所述包含若干抗原的多肽链包含seqidno:07所示的序列。如图16b所示,两种不同的包膜蛋白将位于细胞膜中,并可能作为亚病毒颗粒分泌。这些亚病毒颗粒可包含来自不同hbv基因型的两种包膜蛋白,并且可以被抗原呈递细胞摄取,这可以增加诱导的免疫应答。所述核心颗粒可形成空衣壳,其中所述空衣壳可类似地包含来自不同hbv基因型的核心蛋白。这样的一个衣壳将触发针对多种hbv基因型的免疫应答。所述聚合酶将在蛋白酶体中降解并在hla背景下呈递。本文所述的排列进一步具有以下优点:大部分仅部分加工的蛋白,即其中自切割位点未被切割的蛋白将被并入分泌的病毒样颗粒中,所述病毒样颗粒将假定增加免疫应答,特别是增强并拓宽适应性免疫应答(参见图18)。
本发明涵盖一种重组mva,其包含并入mva基因组中多个插入位点的hbv基因。“多个插入位点”意指编码hbv抗原的hbv基因可以分别插入mva基因组中相同或一个或多个不同的插入位点,其中,优选相同的插入位点。不同的含义是,例如,第一个插入位点与第二个插入位点不同。
作为示例性实例,可以将所述hbv基因插入mva的基因间区域(igr)。这样的igr可以选自igr07/08、igr44/45、igr64/65、igr88/89、igr136/137和igr148/149。然而,优选将所述hbv基因插入mva的天然存在的缺失位点i、ii、iii、iv、v或vi中,其中优选地,将至少一个或优选全部hbv基因插入天然存在的缺失位点iii。作为示例性实例,如本文所述,所有hbv基因可包含在单个开放阅读框的单一表达盒中,其中所述单一表达盒插入天然存在的缺失位点iii。
所述重组mva病毒可以通过本领域已知的常规方法产生。获得重组痘病毒的方法或将外源编码序列插入痘病毒基因组的方法是本领域技术人员熟知的。例如,方法描述在以下参考文献中:molecularcloning,alaboratorymanual.第二版.由j.sambrook,e.f.fritsch和t.maniatis编.coldspringharborlaboratorypress.2003:描述了标准分子生物学技术的技术,例如dna克隆、dna和rna分离、蛋白质印迹分析、rt-pcr和pcr扩增技术。virologymethodsmanual.由brianwjmahy和hillarokangro编.academicpress.1996:描述了处理和操作病毒的技术。molecularvirology:apracticalapproach.由ajdavison和rmelliott编.thepracticalapproachseries.irlpressatoxforduniversitypress.oxford1993.第九章:expressionofgenesbyvacciniavirusvectors。currentprotocolsinmolecularbiology.出版商:johnwiley和soninc.1998.第16章,第iv部分:expressionofproteinsinmammaliancellsusingvacciniaviralvector:描述了mva的处理、操作和基因工程的技术和知识。
为了产生根据本发明的重组痘病毒,可以使用不同的方法。可将待插入到病毒中的所述dna序列置于其中已经插入了与痘病毒的一段dna同源的dna的大肠杆菌质粒构建体中。另外,所述待插入的dna序列可以连接到启动子上。所述启动子-基因连接可以位于质粒构建体中,使得启动子-基因连接的两端均为与位于包含非必需基因座的痘病毒dna的区域侧翼的dna序列同源的dna。所得到的质粒构建体可以通过在大肠杆菌内繁殖进行扩增并分离。所述含有待插入的dna基因序列的分离的质粒可以被转染到细胞培养物,例如鸡胚成纤维细胞(cef)中,连同用痘病毒感染该培养物。质粒中的同源痘病毒dna与所述病毒基因组之间的重组分别可产生一种由外源dna序列存在而修饰的痘病毒。
可用痘病毒感染合适的细胞培养物的细胞,如cef细胞。随后,可以用包含优选地处于痘病毒表达控制元件的转录控制下的外源基因的第一质粒载体转染所述感染的细胞。如上所说明,所述质粒载体还包含能够引导外源序列插入痘病毒基因组的选定部分的序列。任选地,所述质粒载体还含有包含可操作地连接于痘病毒启动子的标记和/或选择基因的盒。合适的标记或选择基因是例如编码绿色荧光蛋白、β-半乳糖苷酶、新霉素-磷酸核糖基转移酶或其它标记的基因。选择盒或标记盒的使用使对产生的重组痘病毒的鉴定和分离简化。然而,重组痘病毒也可以通过pcr技术来识别。随后,其它细胞可用如上所述获得的重组痘病毒感染,并且用包含第二外来基因的第二载体转染。在这个情况下,这个基因可被引入痘病毒基因组的不同插入位点中,所述第二载体也在引导所述第二外来基因整合至痘病毒的基因组中的痘病毒同源性序列方面不同。在已发生同源性重组之后,可分离所述包含两个或更多个外来基因的重组病毒。为了将另外的外来基因引入重组病毒中,可通过使用在先前感染步骤中分离的重组病毒,以及通过使用包含用于转染的其它外来基因的另一载体来重复感染和转染步骤。
可选地,如上所述的感染和转染步骤可互换,即适合细胞可首先用包含外来基因的质粒载体转染,然后用痘病毒感染。作为另一替代方案,也可以将每一外来基因引入不同病毒中,用所有所得重组病毒共同感染细胞,并筛选包括所有外来基因的重组体。第三替代方案是在体外连接dna基因组和外来序列,并使用辅助病毒重建重组牛痘病毒dna基因组。第四替代方案是在大肠杆菌或另一细菌物种中,在克隆为细菌人工染色体(bac)的牛痘病毒基因组与侧接有dna序列的线性外来序列之间的同源性重组,所述dna序列与侧接所述牛痘病毒基因组中的所需整合位点的序列同源。
由于根据本发明的重组mva病毒的复制高度受限以及因此高度减毒,所以它是用于治疗广泛范围的哺乳动物(包括人以及甚至免疫损害的人)的理想候选物。此外,由于mva可以方便地在cef细胞中培养,因此基于mva的疫苗的生产或多或少独立于常规疫苗的现有生产能力。因此,本发明的基于mva的疫苗可以在短时间内大量生产。此外,本发明的mva载体疫苗可以递送多种hbv抗原并因此允许同时诱导高水平的体液和细胞免疫。因此,本发明还提供了用于在包括人在内的活动物体中诱导免疫应答的药物组合物和疫苗。
优选地,所述疫苗包含浓度范围为104至109tcid50/ml、优选地浓度范围为105至5×108tcid50/ml、更优选地浓度范围为106至108tcid50/ml、和最优选地浓度范围为107至108tcid50/ml的mva病毒。
用于人的优选疫苗接种剂量包括106至109tcid50,最优选地为106tcid50或l07tcid50或108tcid50的剂量。
所述药物组合物通常可包含一种或多种药学上可接受的和/或批准的如载体、抗生素、防腐剂、佐剂、稀释剂和/或稳定剂的添加剂。这种辅助物质可为水、盐水、甘油、乙醇、湿润剂或乳化剂、ph缓冲物质等。合适的载体通常是代谢缓慢的大分子,诸如蛋白质、多糖、聚乳酸、聚乙醇酸、聚合氨基酸、氨基酸共聚物、脂质聚集体等。
术语“药学上可接受的”指不干扰根据本发明的mva的生物学活性的有效性的无毒材料。载体的特征取决于施用途径。药物组合物可以进一步含有增强活性或用于治疗的其它试剂。这些另外的因子和/或试剂可以包含在药物组合物中以用于根据本发明的免疫方法,以产生协同作用或使副作用最小化。可以在“remington′spharmaceuticalsciences”,(muckpublishinggompany,easton,pa,最新版)中找到用于配制和施用根据本发明的mva的技术。
对于疫苗的制备,可将根据本发明的重组mva转变成生理上可接受的形式。这可基于在制备用于针对天花进行疫苗接种的痘病毒疫苗方面的经验来进行(如stickl,h.等,[1974]dtsch.med.wschr.99,2386-2392所描述)。
例如,纯化的病毒可以5×108tcid50/ml的效价于约10mmtris、140mmnacl(ph7.4)中配制来储存在-80℃下。对于疫苗注射物(shot)的制备,例如可在安瓿(优选玻璃安瓿)中在2%蛋白胨和1%人白蛋白存在下将102-108个病毒颗粒于100ml磷酸盐缓冲盐水(pbs)中冻干。可选地,所述疫苗注射物可通过逐步冷冻干燥病毒来以制剂形式产生。该制剂可含有适于体内施用的另外的添加剂,比如甘露醇、右旋糖酐、糖、甘氨酸、乳糖或聚乙烯吡咯烷酮或其它助剂,比如抗氧化剂或惰性气体、稳定剂或重组蛋白(例如人血清白蛋白)。然后将玻璃安瓿密封,并且可在4℃与室温之间储存数月。然而,在不存在需要时,优选将安瓿储存在-20℃以下的温度下。
对于疫苗接种或治疗,可将所述冻干物溶解于水性溶液优选是生理盐水或tris缓冲液中,并且全身性或局部施用,即胃肠外、皮下、静脉内、肌肉内、或为熟练从业者所知的任何其它施用途径。施用模式、施用剂量和次数可由本领域技术人员以已知方式进行优化。
应该理解,优选的疫苗或药物组合物包含如本文所述的本发明的重组mva。还应理解,本发明所述的重组mva可用于优选地针对hbv的治疗或疫苗接种,优选治疗性疫苗接种。
本发明的重组mva可以有利地用于制备用于治疗和/或预防诸如感染病或乙型肝炎的病理状况的药物或疫苗。
本发明进一步设想了一种针对乙型肝炎的疫苗接种方法。所述疫苗接种方法可以是治疗性疫苗接种,即用于治疗疾病的方法。在本发明的疫苗接种方法中,将本文所述的表达来自hbvadw的包膜蛋白的mva病毒和本文所述的表达来自hbvayw的核心蛋白的mva病毒施用于受试者。所述受试者可以是本文定义的任何受试者,优选人类受试者。优选地,所述受试者需要所述施用。在本发明的疫苗接种方法中,所述来自hbvadw的包膜蛋白和所述来自hbvayw的核心蛋白可以由两种不同的mva表达,其中mva中的每种表达包膜蛋白或核心蛋白。在这种情况下,必须将两种mva都施用于受试者。然而,所述来自hbvadw的包膜蛋白和所述来自hbvayw的核心蛋白也可以由同一mva表达。在这种情况下,仅有一种mva必须施用于受试者。应该理解,后一种实施方案是优选的。
此外,本发明涵盖施用于受试者的表达如本文所述的来自hbvadw的包膜蛋白并进一步表达cd70分子的mva病毒和表达如本文所述的来自hbvayw的核心蛋白并进一步表达cd70分子的mva病毒。所述受试者可以是本文定义的任何受试者,优选人类受试者。优选地,所述受试者需要所述施用。此外,优选地,所述受试者为需要治愈性治疗的慢性乙型肝炎患者。在本发明的疫苗接种方法中,所述来自hbvadw的包膜蛋白和cd70分子以及来自hbvayw的核心蛋白和cd70分子可以由两种不同的mva表达,其中mva中的每种表达包膜蛋白和cd70或核心蛋白和cd70。在这种情况下,必须将两种mva都施用于受试者。然而,所述来自hbvadw的包膜蛋白和来自hbvayw的核心蛋白以及cd70分子也可以由同一mva表达。在这种情况下,仅有一种mva必须施用于受试者。应该理解,后一种实施方案是优选的。
还应理解的是,本发明的疫苗接种方法包括将本发明的mva施用于受试者,并且优选地,所述mva为有效剂量。
本发明的疫苗接种方法可以包括至少两个疫苗接种步骤。在此,免疫应答由初次(prime)/加强方案诱导,其中“游离”蛋白(比如来自hbva/adw的包膜蛋白和/或来自hbvd/ayw的核心蛋白)用于初次疫苗接种,其中将一种或多种本发明的重组mva用于至少一次加强疫苗接种。
因此,在第一步骤(初次免疫)中,可以将“蛋白质疫苗”施用于受试者。如本文所用的术语“蛋白质疫苗”是指包含来自hbva/adw的包膜蛋白和/或来自hbvd/ayw的核心蛋白、优选包含所述包膜蛋白和所述核心蛋白的组合物。优选地,包膜蛋白和核心蛋白都为“游离”蛋白,这意味着它们优选不包含在病毒颗粒中。所述包膜蛋白和核心蛋白可以存在于两种单独施用或彼此组合施用的组合物中。所述包膜蛋白和核心蛋白也可包含在单一组合物中。所述包含在“蛋白质疫苗”中的包膜蛋白或核心蛋白可以由微生物,例如由细菌或真菌细胞重组产生。
应该理解,优选地,所述“蛋白质疫苗”基本上不含病毒颗粒,特别是基本上不含mva。在本文中“基本上不含”是指所述蛋白质疫苗包含小于103tcid50/ml、优选小于102tcid50/ml、优选小于101tcid50/ml、优选小于100tcid50/ml、优选小于10-1tcid50/ml、优选小于10-2tcid50/ml。所述蛋白质疫苗可以进一步包含合适的佐剂。如本文所用,“佐剂”是指增强、提高或加强宿主对抗原或其片段的(抗体和/或细胞介导的)免疫应答的物质。技术人员已知合适的佐剂。优选的佐剂选自聚[二(对丙酸钠苯氧基)]磷腈(pcep)、免疫刺激性寡核苷酸、toll样受体(tlr)激动剂、皂苷或它们的组合,其中所述tlr激动剂优选为tlr3激动剂、tlr4激动剂、tlr7激动剂、tlr8激动剂或tlr9激动剂,并且其中所述免疫刺激性寡核苷酸优选为聚i/c、聚iclc(聚i/c的稳定化形式)、cpg、rig-i配体、sting配体、环状di-amp、环状di-cmp、环状di-gmp、tlr7激动剂、tlr8激动剂、cta1dd或dmlt或它们的组合。然而,本发明的发明人已经发现,如果蛋白质疫苗包含无铝佐剂,例如cpg佐剂或pcep,则本发明的疫苗接种方法将诱导更强的t细胞应答。这一发现令人惊讶,因为包含hbv抗原的常规疫苗,如engerix-b通常包含含铝佐剂(如氢氧化铝)。因此,本文所述的蛋白质疫苗优选包含cpg佐剂或pcep或两者。本发明进一步提供了病毒载体,优选mva病毒载体,其中佐剂是环状di-amp。
在进一步的步骤(加强免疫)中,将如本文所述的一种或多种重组mva施用于受试者。所述一种或多种重组mva可以包含在本文所述的药物组合物或疫苗中。
通常,所述重组mva在初次接种后至少约1天、优选地至少约5天、优选地至少约1周、优选地约1周至约8周、优选地约2周至约5周、优选地约3周至约4周施用。
本发明还涵盖该疫苗接种方案包括在初次疫苗接种后两次或更多次的加强疫苗接种。在一个优选的实施方案中,将蛋白质疫苗用于初次疫苗接种和第一次加强疫苗接种。本发明还设想,可以进行采用该蛋白质疫苗的第二、第三或进一步的加强疫苗接种步骤。在用蛋白质疫苗加强疫苗接种后,通过施用如本文所述的一种或多种重组mva进行加强免疫接种。本发明还涵盖第一次施用重组mva之后可以进行第二次、第三次或另外的施用(进一步的加强疫苗接种)。
在这种情况下,采用所述蛋白质疫苗的第一次加强疫苗接种在先前疫苗接种步骤后至少约1天、优选地至少约5天、优选地至少约1周、优选地约1周至约8周、优选地约2周至约5周、优选地约3周至约4周进行。任何采用重组mva或采用蛋白质疫苗的随后的加强疫苗接种均在先前疫苗接种后至少约1天、优选地至少约5天、优选地至少约1周、优选地约1周至约8周、优选地约2周至约5周、优选地约3周至约4周进行。
本发明的疫苗接种方法可以进一步包括在初次-加强疫苗接种之前用sirna和/或shrna治疗受试者。设想这种sirna或shrna靶向hbv基因,由此降低hbv感染受试者中被靶向的hbv基因的表达。优选地,所述sirna和/或shrna靶向编码被靶向的hbv基因的rna的3′区域。在优选的实施方案中,所述编码被靶向的hbv基因的rna的被靶向的3′区域位于rna的polya尾部的上游(即,所述靶向的区域是对于所述polya尾部的5′,或换言之在5′至3′的方向,首先是所述靶向的区域,然后是所述polya尾部)。如果使用sirna,设想靶向至少一个hbv基因。然而,在一个优选实施方案中,靶向全部hbv基因。可将一种sirna或shrna用于靶向hbv基因。然而,也可将超过一种sirna和/或shrna用于靶向hbv基因。可以设想,这种采用sirna和/或shrna的治疗将被靶向的基因的表达(即翻译)降低至少约1%、5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%或85%,优选地至少约90%。这种采用sirna和/或shrna对受试者的治疗可以在初次-加强疫苗接种开始之前1天至20周、1周至19周、2周至18周、3周至17周、4周至16周、5周至15周、6周至14周、7周至13周,优选地约8周进行。本发明人惊讶地发现,在初次-加强疫苗接种开始之前的这种采用sirna和/或shrna的治疗使初次-加强疫苗接种后产生的t细胞应答增加。
根据本发明的免疫方法将利用治疗有效量的所述蛋白质疫苗或所述重组mva。治疗有效剂量进一步是指足以导致症状改善,如治疗、治愈、预防或改善这些病症的化合物/成分的量。在优选的实施方案中,所述免疫可以是预防性和/或治疗性的。用于影响免疫应答的本发明的有效剂量取决于许多不同因素而变,包括抗原或疫苗的类型、施用手段、靶位点,受试者是人还是动物,以及治疗是预防性还是治疗性的。本文公开了重组mva的优选剂量。
本发明提供了用于免疫受试者动物(包括鸟)的方法。优选地,所述动物是哺乳动物,包括大鼠、兔、猪、小鼠和人,包括向受试者施用mva的剂量。最优选地,所述受试者是人。在一个实施方案中,所述受试者是成人。
施用可以通过技术人员确定的任何施用途径来完成。优选地,施用为肠胃外、肠内粘膜,优选地为肌肉内、静脉内、皮下(如通过刮擦或注射)、鼻(如通过吸入)或口服。优选的施用模式是肠胃外或粘膜。
此外,本发明提供了肌肉内施用,并且其中施用包括佐剂的施用。优选地,所述佐剂包含环状di-amp。
另外,本发明还涵盖皮下或肌肉内施用,并且其中所述施用包括佐剂的施用。优选地,所述佐剂包含聚i/c或rig-i配体。
本发明还考虑如上所述的疫苗接种方法,其中所述疫苗接种载体不是mva而是一种或多种表达与mva相同的抗原的沙门氏菌菌株。因此,本文所述的疫苗接种方法采用沙门氏菌菌株的疫苗接种方法加以适当修改而应用。优选地,这种疫苗接种方法为粘膜疫苗接种方法,其中所述蛋白质疫苗优选地采用cta1dd或dmlt辅佐。
本发明还涵盖本文方法中公开的用于在这些方法中使用的任何化合物。例如,本发明涵盖用于在本文公开的疫苗接种方法中使用的所述重组mva。
本发明还考虑包含本发明的重组疫苗接种载体中的一种或多种的药物组合物或疫苗。如上文所提及的,所述“疫苗”可用于预防或治疗受试者中的病理状态。该术语涵盖亚单位疫苗,即含有抗原的疫苗组合物,所述抗原与该抗原在自然界中相关的完整生物体是分开的和不连续的,以及涵盖组合物,所述组合物含有本发明的重组痘病毒(尤其是携带所述抗原和/或其表位的重组痘病毒)。所述疫苗可以包含或不包含一种或多种增强活性成分的免疫活性的其他组分。疫苗可另外包含药物组合物的其他典型组分。本发明的疫苗优选用于人和/或兽医用途。
本发明的疫苗或组合物,如包含mva病毒的疫苗,通常可以包括一种或多种药学上可接受的载体、添加剂、抗生素、防腐剂、佐剂、稀释剂和/或稳定剂。这种辅助物质可以是水、盐水、甘油、乙醇、湿润剂或乳化剂、ph缓冲物质等。合适的载体通常是代谢缓慢的大分子,诸如蛋白质、多糖、聚乳酸、聚乙醇酸、聚合氨基酸、氨基酸共聚物、脂质聚集体等。也可以将如us2011/0052627第7栏中所述的其他载体加入到所述组合物中。
本发明还涵盖包含至少两个用于初次/加强免疫的小瓶/容器的试剂盒,其中至少一个小瓶/容器包含用于第一次接种(初次接种)的本文所述的蛋白质疫苗和任选地用于进一步接种的另外的小瓶(加强接种),其中优选地所述另外的小瓶包含本发明的重组疫苗接种载体。所述试剂盒还包含至少一个另外的小瓶/容器,其包含如本文所述的重组疫苗接种载体用于至少另一次接种(“加强接种”)。所述试剂盒可进一步包含将所述重组疫苗接种载体施用于受试者的说明书。
本发明考虑所述优选的受试者是人。所述说明书可指示将如本文所定义的蛋白质疫苗和/或重组疫苗接种载体在特定时间点(如在先前施用后至少4周、至少6周、至少8周)以多个(即2、3、4、5、6个等)剂量施用于受试者。优选地,所述说明书指示所述蛋白质疫苗和/或重组疫苗接种载体以至少3或至少4个剂量施用。
本发明考虑的是本文所述的试剂盒,其中包含在试剂盒中的疫苗适用于肠胃外施用。在这样的试剂盒中,所述蛋白质疫苗可以优选地采用pcep和/或cpg疫苗辅佐,并且所述重组疫苗接种载体优选地可以是本发明的mva。
本发明还考虑本文所述的试剂盒,其中包含在试剂盒中的疫苗适用于粘膜施用。在这种试剂盒中,所述蛋白质疫苗可优选地采用cta1dd、dmlt、pcep、c-di-amp、c-di-cmp、c-digmp或它们的组合辅佐,并且所述重组疫苗接种载体可优选地为沙门氏菌菌株、cmv-基载体、vsv-基载体、腺病毒载体或麻疹载体,优选地为本发明的沙门氏菌菌株。
另外,本发明还考虑本文所述的试剂盒,其中包含在试剂盒中的疫苗适合于粘膜施用。在这种试剂盒中,所述蛋白质疫苗可优选地采用cta1dd、dmlt、pcep、polyi/c、rig-i-配体、c-di-amp、c-di-cmp、c-digmp或它们的组合辅佐,并且所述重组疫苗接种载体可优选地为本发明的沙门氏菌菌株,或优选地为沙门氏菌菌株、cmv-基载体、vsv-基载体、腺病毒载体或麻疹载体。
本发明进一步提供一种试剂盒,其中所述蛋白质组合物适用于肌肉内施用。优选地,所述组合物包含至少一种为环状di-amp的佐剂。
本发明还考虑了一种试剂盒,其中所述蛋白质组合物适用于皮下或肌肉内施用。优选地,所述组合物包含至少一种为聚i/c的佐剂。
本发明还提供了一种包含本文所述的重组mva的(宿主)细胞。允许痘病毒的细胞的实例包括但不限于,cos、hek-293、bhk、cho、tm4、cvi、vero-76、hela、mdck、brl3a和nih/3t3细胞。对于mva,优选的细胞是cef和bhk细胞。其它细胞系是本领域普通技术人员已知的。可以通过磷酸钙转染、电穿孔、感染以及本领域已知的和在标准实验室手册如currentprotocolsinmolecularbiologyjohnwiley&sons,inc.newyork中描述的其它方法来实现痘病毒构建体向细胞的导入。
本发明还提供了感染有重组痘病毒的(宿主)细胞。因此,所述(宿主)细胞可由mva病毒载体感染,并且用包含待插入基因的其他载体(例如质粒载体)转染,所述基因优选地在mva或痘病毒表达控制元件或启动子(如合成的prs启动子)的转录控制下。如上所述,所述质粒载体包含能够引导所述异源序列插入痘病毒基因组的选定部分的序列,例如侧接天然存在的缺失位点或基因间区域之一的序列。
如本文所用,术语“约”或“大约”是指在给定值或范围的20%偏差内,比如在10%偏差内或5%内。
附图说明
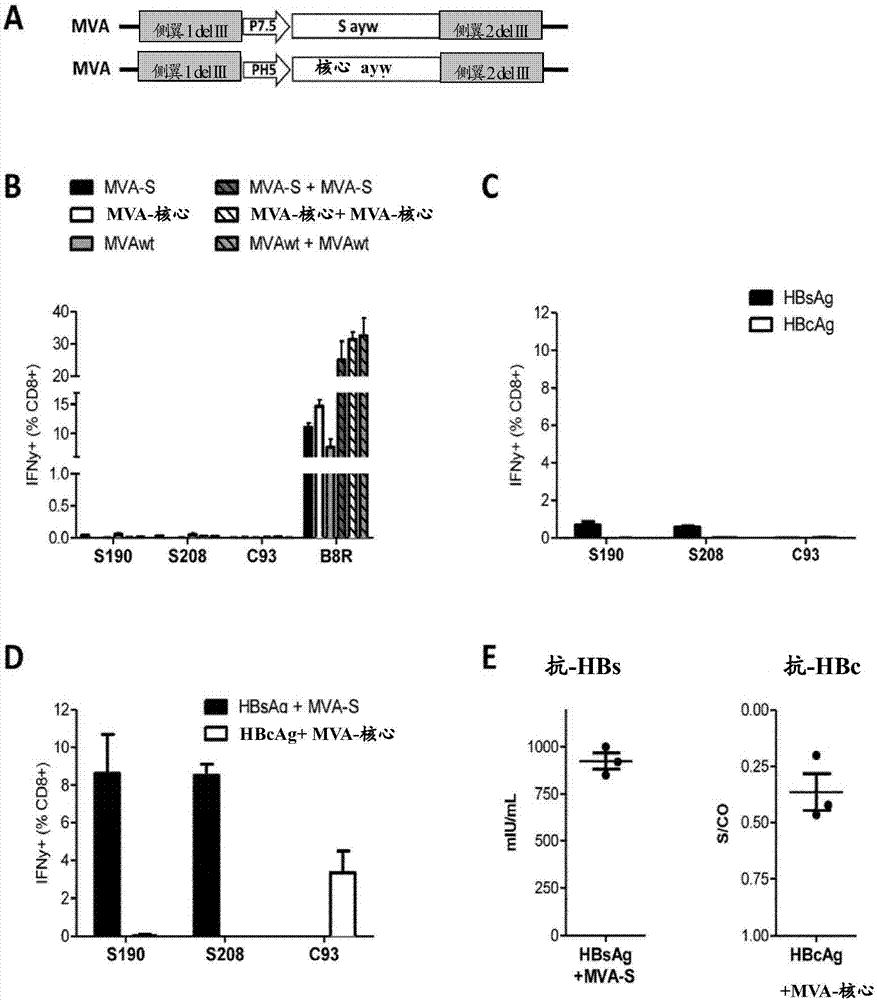
图1:蛋白质-初次/mva-加强疫苗接种是高度免疫原性的.(a)将hbvayw亚型的s和核心开放阅读框插入mva基因组的缺失iii(deliii),分别在痘病毒启动子p7.5和ph5的控制下。(b)给野生型小鼠(n=3)接种mva-s(1×108i.u.)、mva-核心(1×108i.u.)或mvawt(1×108i.u.)一次(第0天;填充柱)或两次(第0天和21天;条纹柱)。第8天(初次后)或第27天(加强后),用源自hbsag(s190和s208)-或hbcag(c93)的肽刺激脾细胞并通过细胞内细胞因子染色分析ifnγ表达。(c)给c57bl/6小鼠接种12μg重组hbsag或hbcag。cpg用作佐剂。第8天,用源自hbsag或hbcag的肽刺激脾细胞并通过ics分析ifnγ表达。(d)c57bl/6小鼠初次接种12μg重组的、cpg辅佐的hbcag或hbsag,并且第21天用mva-核心(1×108i.u.)或mva-s(1×108i.u.)加强。第27天,用源自hbsag或hbcag的肽刺激脾细胞并通过ics分析ifnγ表达。通过免疫测定分析血清中的抗-hbs(中间小图)或通过竞争elisa分析血清中的抗-hbc(右边小图)。示出的产生ifnγ的t细胞的频率是背景扣除的。s/co:信号与临界值之比;i.u.:感染单位。
图2:疫苗接种诱导的hbv特异性抗体-和cd4+t细胞应答与抗原血症成负相关.(a-d)采用cpg辅佐的hbsag或hbcag免疫低、中和高抗原血症的hbvtg小鼠(n=4)。第21天,将小鼠分别用mva-s(1×108i.u.)或mva-核心(1×108i.u.)加强。第27天(加强后的第6天),分析血清中的(a)抗-hbs和(b)抗-hbc抗体。用hbsag刺激低-抗原血症hbvtg小鼠的(c)脾细胞和(d)肝相关淋巴细胞并通过细胞内细胞因子染色分析表达ifnγ的cd4+t细胞。示出的产生ifnγ的t细胞的频率是背景扣除的。s/co:信号与临界值之比;neg.=阴性;i.u.:感染单位。
图3:疫苗接种诱导的hbv特异性cd8+t细胞频率与抗原血症成负相关.采用12μgcpg辅佐的hbsag或hbcag免疫低、中和高抗原血症的hbvtg小鼠(n=4)。第21天,将小鼠用mva-s(1×108i.u.)或mva-核心(1×108i.u.)加强。加强后第27天,用hbsag(s109和s208)-或hbcag(c93)-特异性肽刺激脾细胞(a)和肝相关淋巴细胞(b)并通过细胞内细胞因子染色分析ifnγ表达。上面和中间的小图示出了示例性动物,底部的小图给出了扣除背景后产生ifnγ的t细胞的频率。i.u.:感染单位。
图4:疫苗接种诱导的hbv特异性cd8+t细胞的功能。采用cpg辅佐的hbsag免疫野生型小鼠以及低和中抗原血症的hbvtg小鼠。第21天,将小鼠用mva-s(1×108i.u.)加强。加强后第6天,(a)分析源自s-特异性脾的cd8+t细胞的cd107a和ifnγ表达。(b)分析源自s-特异性脾或肝的cd8+t细胞经肽s190刺激后的ifnγ-、il-2-和tnfa-表达。(c)s190特异性cd8+t细胞的多聚体染色以及cd127和klrg-1表面表达的共染色。i.u.:感染单位。
图5:异种疫苗接种破坏了高抗原血症hbvtg小鼠中的t细胞耐受性。(a)将sayw和adw亚型的开放阅读框置于mva基因组的缺失iii(deliii)中,在强痘病毒启动子ph5的控制下。(b)至(d)在第0和14天,给高抗原血症hbvtg小鼠(n>4)接种与所示佐剂一起的16μghbsag(ayw或adw亚型)和16μghbcag(ayw亚型)。第28天,将小鼠用mva-ph5-s(5×107i.u.;ayw或adw亚型)和mva-核心(5×107i.u.)加强。加强后的第6天,(b)将脾细胞(左边小图)和肝相关淋巴细胞(lal,右边小图)分离并用肽s109和s208(若示出,adw亚型)或c93刺激,然后通过细胞内细胞因子染色分析ifnγ表达。(c)用hbsag特异性的15-mer肽的c-末端库(覆盖氨基酸145至226,ayw亚型)刺激脾细胞和lal并通过ics分析表达ifnγ的cd4+t细胞。(d)对于tnfα和il-2表达,分析表达ifnγ的s208特异性cd8+t细胞的代表性facs图。产生ifnγ的t细胞频率的平均值±sem以背景扣除示出。ns:不显著。*p<0.05,**p<0.01studentst-检验;i.u.:感染单位。
图6:异种疫苗接种在高抗原血症hbvtg小鼠中诱导血清转化。(a)至(d)在第0和14天,给高抗原血症hbvtg小鼠(n>4)接种cpg和pcep辅佐的16μghbcag(ayw亚型)和16μghbsag(如所示,ayw或adw亚型)。第28天,将小鼠用mva-ph5-s(5×107i.u.;ayw或adw亚型)和mva-核心(5×107i.u.)加强。作为对照,在第0和14天,给5只高抗原血症hbvtg小鼠接种佐剂cpg和pcep,并随后用1×108mva-wt加强。加强后的第6天,分析血清中(a)抗-hbs或(b)抗-hbc抗体的存在。(c)分析来自hbvtg小鼠的抗-hbs阳性血清的ayw亚型hbsag的中和能力。(d)示出了疫苗接种前后的血清hbsag水平。示出了平均值±sem。ns:不显著。s/co:信号与临界值之比。**p<0.01studentst-检验。
图7:mva载体的hbv抗原表达.(a)和(b)用mva-s、mva-核心或mvawt(10个moi)感染鼠nih-3t3细胞。感染后16h,(a)通过蛋白质印迹法分析总细胞裂解物的hbsag和hbcag表达。(b)通过hbsag-特异性elisa测定了上清液中分泌的hbsag。s/co:信号与临界值之比。
图8:cpg辅佐的hbsag的疫苗接种.用12μg含有cpg作为佐剂的hbsag免疫高抗原血症hbvtg小鼠。第21天,将小鼠用mva-s加强。初次免疫后的第0、27、35、42、49和56天,分析血清的hbsag和抗-hbs的水平。s/co:信号与临界值之比。
图9:用于蛋白质初次疫苗接种的佐剂的比较.(a)至(c)第0天,给野生型小鼠接种与所示佐剂一起的16μghbsag(ayw亚型)和16μghbcag(ayw亚型)。第28天,将小鼠用mva-ph5-s(5×107i.u.;ayw亚型)和mva-核心(5×107i.u.)加强。加强后的第6天,分析血清中(a)抗-hbs和(b)抗-hbc的存在。(c)加强后的第6天,采用hbsag(s190和s208adw)-或hbcag(c93)-特异性肽刺激后,通过ics分析脾细胞。柱示出了背景扣除后对ifnγ染色阳性的cd8+细胞的百分比(平均值±sem)。i.u.:感染单位。
图10:hbvtg小鼠的分组.
根据小鼠的抗原水平对小鼠分组,在hbvtg小鼠的情况下,所述抗原水平与血清中测定的病毒效价密切相关。
图11:用于刺激t细胞的肽.
表中给出了用于确定由鼠kb/kdmhs-i等位基因限制的cd8+t细胞应答的肽序列。s库用于广泛确定小鼠和人的cd4+和cd8+t细胞应答。
图12:实施例4的疫苗接种方案的图示.
在第0和14天,给高抗原血症hbvtg小鼠(≥4)接种与所示佐剂一起的16μghbsag(ayw或adw亚型)和16μghbcag(ayw亚型)。第28天,将小鼠用mva-ph5-s(5×107i.u.;ayw或adw亚型)和mva-核心(5×107i.u.)加强。加强后的第6天,(b)将脾细胞(左边小图)和肝相关淋巴细胞(lal,右边小图)分离并用肽s109和s208(若示出,adw亚型)或c93刺激,然后通过细胞内细胞因子染色(ics)分析ifnγ表达。
图13:hbv特异性cd8+t细胞的多聚体与细胞内细胞因子染色的相关性.用12μg含有cpg作为佐剂的hbsag免疫hbvtg小鼠。第21天,将小鼠用mva-s加强。在第28天通过s190多聚体染色或在用肽s190离体再刺激后通过ics检测hbv特异性t细胞应答。
图14:血清免疫复合物的检测.用cpg辅佐的hbsag免疫hbvtg小鼠.第21天,将小鼠用mva-s(1×108i.u.)加强。加强后的第6天,分析血清的沉淀的免疫复合物中的抗-hbs、hbsag和hbsag。i.u.:感染单位;nd:不可检测。
图15:不同疫苗佐剂的比较.用在不同佐剂制剂中复合的hbcag和hbsag免疫野生型ch57bi/6小鼠。1:cpg加氢氧化铝(alumnhydroxide);2:聚磷腈pcep加明矾(alum);3:pcep加cpg加铝(alumn);4:仅cpg;5:仅pcep;6:cpg加pcep。第28天,将全部动物用表达核心和sadw亚型的mva加强。图15a-b示出了28(仅蛋白质初次免疫)和34天(蛋白质初次免疫加mva加强)后抗-hbs(图15a)和抗-hbc(图15b)抗体应答。图15c-d示出了初次免疫后(图15c)和所述mva加强后(图15d),针对s和核心表位的cd8+t细胞应答。图15e示出了中和分析,其中在感染之前将hbvayw亚型与所示的血清稀释液一起孵育,并且在4、7和10天后测量被感染细胞的hbsag分泌。
图16:多抗原的开放阅读框.图16a描绘了由seqidno:07所示的多抗原多肽链的结构。图16b示意性地描绘了包含hbsa/adw和c/ayw抗原的亚病毒颗粒的形成。图16c示意性地描绘了包含hbcd/ayw和c/ayw抗原的空衣壳的形成。
图17:源自seqidno:07所示的多抗原多肽链的完全加工的蛋白及其命运(fate)的示意图.
图18:源自seqidno:07所示的多抗原多肽链的部分未加工的蛋白及其预期命运(fate)的示意图.大多数所述部分未加工的蛋白由于并入分泌的病毒样颗粒/细丝而被认为增加免疫应答(尤其是增强和扩大适应性免疫应答)。
图19:包含c-末端突出端(overhang)的hbva2/adw2的小包膜蛋白的氨基酸序列(seqidno:10).加下划线的序列对应于不含c-末端突出端的hbva2/adw2的小包膜蛋白的氨基酸序列(seqidno:08)。所述c-末端突出端是对应于seqidno:09的p2a切割片段。
图20:包含n-和c-末端突出端的hbvd/ayw的核心蛋白片段1-149的氨基酸序列(seqidno:12).加下划线的序列对应于不合n-和c-末端突出端的hbvd/ayw的核心蛋白片段1-149的氨基酸序列(seqidno:11)。所述c-末端突出端是对应于seqidno:09的p2a切割片段。
图21:包含n-和c-末端突出端的hbv聚合酶的rt结构域的氨基酸序列(seqidno:16).加下划线的序列对应于不合n-和c-末端突出端的hbv聚合酶的rt结构域的氨基酸序列(seqidno:06)。所述c-末端突出端是对应于seqidno:13的t2a切割片段。
图22:包含n-和c-末端突出端的hbvc/ayw的大包膜蛋白的氨基酸序列(seqidno:14).加下划线的序列对应于不含n-和c-末端突出端的hbvc/ayw的大包膜蛋白的氨基酸序列(seqidno:04)。所述c-末端突出端是对应于seqidno:13的t2a切割片段。
图23:包含n-末端突出端的hbvc/ayw的核心蛋白的氨基酸序列(seqidno:15).加下划线的序列对应于不含n-末端突出端的hbvc/ayw的核心蛋白的氨基酸序列(seqidno:05)。
图24:hbv聚合酶的rt结构域的共有序列的氨基酸序列(seqidno:03).
图25:c基因型hbv株的大包膜蛋白的共有序列的氨基酸序列(seqidno:01).
图26:c基因型hbv株的核心蛋白的共有序列的氨基酸序列(seqidno:02).
图27:rnai和治疗性疫苗接种的组合.(a)实验设置。hbvxfs转基因小鼠接受hbv特异性sirna(sihbv)、无关sirna(sineg)或未经处理。八周后,全部小鼠接受蛋白质初次免疫-用hbv核心和表面抗原(hbcag和hbsag)进行mva加强治疗性免疫。(b)所述hbv特异性sirna/shrna设计的示意图。
图28:hbv抗原水平和抗体应答以及cd8+t细胞应答.(a)血清hbeag和hbeag水平的动力学。(b)在处死时间点(第91天,第13周(w))时小鼠血清中的抗-hbs抗体。(c)初次-加强免疫接种后在肝和脾中测量的cd8+t细胞应答。
图29:最佳mva剂量的评估.
根据血清hbeag水平对低及中抗原血症的hbvtg小鼠进行分组。(a)以两周的间隔,用15μgc-di-amp辅佐的颗粒状hbcag免疫hbvtg小鼠组(n=3-4)两次。第28天,将小鼠用4种不同剂量的mva-核心(分别为3×106、1×107、3×107、1×108pfu)加强。分析第0天和第35天(加强后第7天)小鼠的血清中hbsag、hbeag、抗-hbs和抗-hbc抗体(b)和alt水平(c)。(d)第35天,分离hbvtg小鼠的脾细胞和肝相关淋巴细胞,用来自mva的肽b8r或来自hbcag的肽c93刺激,并通过细胞内细胞因子染色分析表达ifnγ的cd8+t细胞。示出的产生ifnγ的t细胞的频率是背景扣除的。i.m.-肌肉内免疫;s/co:信号与临界值之比;pfu-噬斑形成单位;u-单位;iu-国际单位。
图30:c-di-amp作为用于蛋白质初次免疫(priming)的佐剂的评估.
根据血清hbeag水平对中及高抗原血症的hbvtg小鼠进行分组。(a)以两周的间隔,用c-di-amp辅佐的15μg颗粒状hbsag和15μghbcag的混合物,或cpg/pcep的组合免疫hbvtg小鼠组(n=7)。第28天,将小鼠用108mva-s/核心加强。用c-di-amp注射两次并且用“空”mva(mvawt)加强的hbvtg小鼠(n=4)用作对照。分析第0天和第34天(加强后第6天)小鼠的血清中alt水平(b)、hbsag、hbeag、抗-hbs和抗-hbc抗体(c-d)。(e)第34天,分离hbvtg小鼠(n=4)的脾细胞和肝相关淋巴细胞,用来自hbcag的肽c93或来自hbsag的肽s208刺激,并通过细胞内细胞因子染色分析表达ifnγ的cd8+t细胞。示出的产生ifnγ的t细胞的频率是背景扣除的。i.m.-肌肉内免疫;s/co:信号与临界值之比;pfu-噬斑形成单位;iu-国际单位。
图31:对于各种佐剂:c-di-amp、聚-ic和rig-i配体,最佳递送途径的评估.
根据血清hbeag水平对低及中抗原血症的hbvtg小鼠进行分组。(a)以两周的间隔,用c-di-amp、聚-ic或rig-i配体辅佐的15μg颗粒状hbsag和15μghbcag的混合物免疫hbvtg小鼠组(n=5)。第28天,将小鼠用6×107mva-s/核心加强。免疫接种仅通过肌肉内途径进行,或皮下施用蛋白质初次免疫,然后腹膜内加强。分析第0天和第34天(加强后第6天)小鼠的血清中hbsag、hbeag、抗-hbs和抗-hbc抗体(a)。实验中每周监测hbvtg小鼠的体重(b)。(c)第34天,分离hbvtg小鼠的脾细胞和肝相关淋巴细胞,用来自mva的肽b8r、来自hbsag的肽s208或来自hbcag的肽c93刺激。然后通过细胞内细胞因子染色分析细胞中表达ifnγ的cd8+t细胞。示出的产生ifnγ的t细胞的频率是背景扣除的。i.m.、s.c.、i.p.-分别为肌肉内、皮下、腹膜内免疫;s/co:信号与临界值之比;pfu-噬斑形成单位;iu-国际单位。
图32:c57bl/6小鼠中新mva构建体(mvahbvvac)的评估.
(a)mva-s/核心和mva-hbvvac的示意图。b)来自产生所示mva克隆的细胞的裂解物的蛋白质印迹分析。使用多克隆抗体对非糖基化和糖基化的s进行染色。(c)用c-di-amp辅佐的20μg颗粒状hbsag和20μghbcag的混合物初次免疫c57bl/6小鼠组(n=5)一次。两周后,将小鼠用6×107mva-s/核心或6×107mva-hbvvac加强,从而表达hbsag、hbcag和hbv聚合酶的rt结构域。(d)分析第21天(加强后第7天)小鼠的血清中抗-hbs和抗-hbc抗体。(e)第21天,分离脾细胞并用来自mva的肽b8r,hbsag-、hbcag-和hbvrt-特异性肽以及肽库刺激。来自卵白蛋白的肽siinfekl作为阴性对照。然后通过细胞内细胞因子染色分析细胞中的表达ifnγ的cd8+t细胞。红色箭头表示阳性rt-特异性cd8+t细胞应答。i.m.-肌肉内免疫;s/co:信号与临界值之比;pfu-噬斑形成单位;iu-国际单位。
图33:进一步表达cd70的重组疫苗接种载体(rmva)的构建体的核苷酸序列(seqidno:27).
所述构建体的不同结构域如下所示:deliii-侧翼序列1、
图34:在c57bl/6小鼠中表达cd70的mva的评估.
(a)mva核心和mva核心-cd70的示意图。两种载体都表达hbv核心d基因型序列和gfp以使纯化更容易。mva核心-cd70还表达cd70基因。(b)用20μgpcep和cpg辅佐的颗粒状hbcag肌肉内初次免疫c57bl/6小鼠组(n=6-7)一次。三周后,将小鼠通过腹膜内注射108i.u.mva核心或108mva核心-cd70或108mva-野生型(mvawt)进行加强。(c)第35天,分离脾细胞并用来自mva的肽b8r或核心-肽c93刺激。来自卵白蛋白的肽siinfekl用作阴性对照。然后通过细胞内细胞因子染色分析细胞中的表达ifnγ的cd8+t细胞。每组的数据以平均值±sd给出。点表示单个小鼠中测定的值。i.u.-感染单位。
图35:hbv转基因小鼠中表达cd70的mva的评估.
(a)mva核心和mva核心-cd70的示意图。两种载体都表达hbv核心d基因型序列和gfp以使纯化更容易。mva核心-cd70还表达cd70基因。(b)用20μgpcep和cpg辅佐的颗粒状hbcag肌肉内初次免疫携带1.3倍超长(overlength)基因组的转基因小鼠组(n=5-6)一次,三周后,将该小鼠组通过腹膜内注射108i.u.mva核心或108mva核心-cd70进行加强。用108mva-野生型(mvawt)相应地处理的2只小鼠用作对照。(c)第35天,分离肝相关淋巴细胞(lal)并用来自mva的肽b8r或核心-肽c93刺激。未刺激的细胞用作阴性对照。在细胞内细胞因子染色ifnγ(红色)和il-2(蓝色)后通过流式细胞术分析细胞。示出了三种代表性动物的facs图。
实施例
小鼠和免疫接种
c57bl/6野生型(wt)和hbv-转基因小鼠(毒株hbv1.3.32(guidotti等,jvirol1995;69:6158-69)(hbvd基因型,ayw亚型),由f.chisari,thescrippsinstitute,lajolla,ca,usa友情提供)是根据公共机构的指导从特定无病原体条件下的内部繁殖中衍生出来的。对于蛋白质疫苗接种,用在50μlpbs中与31.91μg的合成硫代磷酸化cpgodn1668和/或25或50μg聚[二(对丙酸钠苯氧基)]磷腈(pcep)混合的重组酵母hbsag或大肠杆菌(e.coli)hbcag(applatvijasbiomedictnas,riga,latvia)皮下地免疫小鼠。对于mva疫苗接种,使小鼠腹腔内地接种500μlpbs中的1×108感染单位的相应的重组mva。
细胞内细胞因子染色、多聚体染色和脱粒分析
如前所述分离脾细胞和肝相关淋巴细胞(lal)(stross等,hepatology2012;56:873-83)并在1mg/ml布雷菲德菌素a(brefeldina)(sigma-aldrich,taufkirchen,germany)存在下,用h2-kb-或h-2db-限制性肽(图10)(jptpeptidetechnologies,berlin,germany)或重组hbsag(由rheinbiotech-dynavax,dusseldorf,germany友情提供)刺激5h。将细胞用溴化乙锭单叠氮溴(invitrogen,karlsruhe,germany)进行活/死染色并用抗-cd16/cd32-fc-block(bdbiosciences,heidelberg,germany)封闭。表面标志物用pb缀合的抗cd-8a和pe缀合的抗-cd4(ebiosciences,eching,germany)染色。采用cytofix/cytoperm试剂盒(bdbiosciences,heidelberg,germany),根据制造商的建议,使用fitc抗-ifnγ(xmg1.2)、pe-cy7抗-tnfa和apc抗-il-2(ebiosciences,eching,germany)进行细胞内细胞因子染色(ics)。
对于脱粒分析,在莫能菌素(monensin)、布雷菲德菌素a、fitc缀合的抗-cd107a抗体和apc缀合的抗-cd107b抗体存在下用肽刺激脾细胞5h,然后,采用cytofix/cytoperm试剂盒(bdbiosciences,heidelberg,germany),根据制造商的建议,采用percp-cy5.5ifnγ(ebiosciences,eching,germany)进行表面pacificbluecd8a染色和ics。
对于多聚体染色,将脾细胞和lal用pe缀合的s190(vwlsviwm,seqidno:19)或mvab8r(tsykfesv,seqidno:20)多聚体染色20分钟,然后用pacificbluecd8a、fitcklrg1和apccd127(ebiosciences,eching,germany)在抗-fc受体抗体(克隆2.4g2)存在下染色20分钟。通过facs分析在afacscantoii(bdbiosciences,heidelberg,germany)上获得数据并采用flowjo软件(treestar,ashland,usa)分析。
血清学分析
采用axsymtm分析(abbottlaboratories,abbottpark,il,usa),以1∶20稀释度测定hbsag、hbeag、抗-hbs和抗-hbc的血清水平。如先前所述(untergasser等,hepatology2006;43:539-47)通过实时聚合酶链式反应进行对血清hbv效价的定量。
中和分析
在来自接种疫苗的小鼠的血清(1∶33、1∶100、1∶333和1∶1000)的系列稀释液存在下,一式两份地用200个包含dna的包膜的hbv颗粒/细胞(ayw亚型)感染如所述(lucifora等,jhepatol2011;55:996-1003)分化和培养的heparg细胞。作为阳性对照,采用0.8个国际单位的hepatecttmcp抗体(biotestpharmagmbh,dreieich,germany)。感染后24小时,将细胞用pbs洗涤3次并加入1ml分化培养基。在感染后第4、7和10天收集上清液,并以1∶20稀释度通过免疫分析检测hbsag。
统计分析
采用prism5软件(graphpad,sandiego,usa)进行统计分析。结果表示为平均值±平均值的标准误差。使用双尾student′st-检验分析组间差异的统计显著性。
mva疫苗的产生
如先前所述通过同源重组和宿主范围选择产生重组mva(staib等,biotechniques2003;34:694-6,698,700)。将整个hbcag(d基因型,ayw亚型)和hbsag开放阅读框(d基因型,ayw或adw亚型)克隆到mva转移质粒piiiδhr-ph5或piiiδhr-p7.5中,由此将hbv蛋白置于早期/晚期牛痘病毒特异性启动子ph5(hbcagayw/hbsagayw/hbsagadw)或p7.5(hbsagayw)的控制下。在构建每种病毒后,验证基因表达、插入dna的序列和病毒纯度。为了产生疫苗制剂,常规地使mva在cef中繁殖、通过蔗糖超速离心纯化、在1mmtris-hclph9.0中重构并按照标准方法(staib等,methodsmolbiol2004;269:77-100)滴定。
免疫印迹
在补充有10%fcs、100u/ml青霉素和100μg/ml链霉素的rpmi1640培养基中培养nih-3t3小鼠成纤维细胞(crl-1658)。感染后16小时,将细胞收获于裂解缓冲液(50mmtris-hcl[ph8.0],150mmnacl,1%nonidetp-40,0.02%nan3和100μg/ml苯甲磺酰氟)中,在12%十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sds-page)上分散,并且印迹至硝酸纤维素膜(0.45μm;bio-rad,munich,germany)上。将膜用抗-hbc(antiserumh800;由h.schaller友情提供)、抗-hbs(murexhbsagversion3;abbott,abbottpark,il,usa)或抗-肌动蛋白(sigma,munich,germany)抗体分别以1∶10000、1∶50和1∶10000的稀释度于4℃下温育。在21℃下,以1∶5000的稀释度使用辣根过氧化物酶标记的二级小鼠和兔抗体(dianova,hamburg,germany)达1小时。将抗体稀释于含有5%脱脂牛奶的磷酸盐缓冲盐水中。根据指导使用增强的化学发光(roche,mannheim,germany)。
采用abbottaxsymhbsag分析(abbottlaboratories,abbottpark,il,usa)测定所培养细胞的上清中分泌的hbsag。
实施例1:蛋白质-初次/mva-加强疫苗接种诱导强抗-hbv免疫
从hbvd基因型、ayw亚型产生两种表达hbs(mva-s)或hbc(mva-核心)的mva疫苗(图1)。蛋白质印迹和hbsagelisa证实了来自任一mva的正确蛋白质表达(图7a、b)。
为了检查免疫原性,用mva-s、mva-核心或mvawt(108感染单位)腹膜内(i.p.)免疫c57bl/6(wt)小鼠,并在免疫后第8天通过细胞内细胞因子染色(ics)测定产生ifnγ的cd8+t细胞的频率。基于全身性mva分布的目的,采用所述i.p.途径。mva-s、mva-核心和mvawt免疫诱导了与来自mva的免疫显性b8r肽(第二次免疫后其被加强)相当的cd8+t细胞应答(图1b)。相对地,没有检测到hbv-特异性cd8+t细胞应答(图1b)。
为了诱导hbv-特异性免疫,进行了异源初次-加强疫苗接种。使小鼠接受12μgcpg作为佐剂的重组颗粒状hbsag或hbcag(ayw亚型)。hbsag疫苗接种后8天,响应于s190和s208肽刺激而分泌ifnγ的脾cd8+t细胞的频率在0.6%左右,而在hbcag免疫后几乎没有检测到针对c93的cd8+t细胞应答(图1c)。第21天,采用mva-s或mva-核心的加强免疫接种能够分别诱导高频率的hbsag-或hbcag-特异性cd8+t细胞以及高抗-hbs或抗-hbc效价(图1d)。总之,这些数据表明,需要异源初次-加强免疫接种来诱导hbv-特异性t细胞。
实施例2:高抗原血症防止了抗-hbv免疫的诱导
为了研究hbv抗原负载对诱导hbv-特异性免疫应答的影响,在接种疫苗前,根据小鼠的血清hbeag水平,将hbv1.3.32转基因(hbvtg)小鼠分为低、中和高抗原血症组(图10)。在加强免疫后6天,与来自中-抗原血症组的小鼠相比,低-抗原血症组中的接受hbsag/mva-s的小鼠的抗-hbs效价更高,并且在高-抗原血症小鼠中仍未检测到抗-hbs效价(图2a)。即使在mva-s加强后监测35天,高-抗原血症小鼠也没有发展可检测到的抗-hbs效价,并且hbsag保持在低水平(图11)。为了研究抗-hbs是否可能与过量的hbsag复合并因此未能检测到,用尿素溶解沉淀的131种蛋白复合物并重复免疫分析。由此,我们在接种的高和中抗原血症hbvtg小鼠中发现了hbsag-抗-hbs免疫复合物,但是未在低抗原血症hbvtg小鼠中发现(图14)。然而,在所有组的hbcag/mva-核心接种的小鼠的血清中检测到了hbcag-特异性抗体,但效价再次显示与抗原血症呈负相关的趋势(图2b)。为了分析疫苗接种诱导的cd4+t细胞应答,用hbsag刺激脾细胞和肝相关淋巴细胞(lal)。尽管在wt小鼠中发现了高频率的hbsag-特异性cd4+t细胞,但未能在任何组的hbvtg小鼠中检测到hbv-特异性cd4+t细胞。
与抗体效价类似,我们观察到抗原血症与hbv-特异性cd8+t细胞应答之间呈负相关。在高-抗原血症小鼠中,hbsag/mva-s以及hbcag/mva-核心免疫既不在外周(脾)也不在肝(hbv-复制位点)中诱导可检测的hbsag-或hbcag-特异性cd8+t细胞(图3a、b)。具有中或低hbeag负荷的小鼠对hbsag/mva-s疫苗接种产生s190-和s208-特异性cd8+t细胞应答,和对hbcag/mva-c疫苗接种产生c93-特异性t细胞应答。在低-抗原血症小鼠中检测到的hbv-特异性cd8+t细胞频率比在中-抗原血症小鼠中发现的该细胞的频率更高。重要的是,mvab8r特异性cd8+t细胞频率独立于抗原血症,并且在所有组之间具有可比性,这表明了相同的疫苗接种效率(图3a、b)。
在慢性感染和抗原持续存在期间,cd8+t细胞可以发展为耗竭的、功能失调的表型。在这种条件下,通过比如ifnγ产生的功能测试大大低估了抗原-特异性t细胞的数量。因此,进行了s190-、c93-和b8r-特异性多聚体染色,其未在免疫的高-抗原血症小鼠的脾或肝中检测到hbv-特异性cd8+t细胞,而b8r-多聚体阳性cd8+t细胞是易于检测到的。这表明该疫苗确实未能诱导该组中的hbv-特异性t细胞。在低-和中-抗原血症组中,hbv-s190-特异性应答和mva-b8r-特异性应答显示多聚体阳性和ifnγ阳性cd8+t细胞的相似比率(图13)。总之,hbv抗原水平影响通过异源初次-加强疫苗接种能够诱导hbv-特异性抗体以及t细胞应答的效率。
实施例3:抗原血症影响疫苗接种诱导的应答的质量
除了产生ifnγ外,cd8+t细胞的重要的效应功能还包括产生il-2和tnfα,以及响应于肽刺激而脱粒的能力,所述脱粒的能力可通过cd107a的表面表达进行分析。在s190肽刺激下,通过hbsag/mva-s免疫在wt小鼠和低-抗原血症hbvtg小鼠中诱导的ifnγ+s190-特异性脾cd8+t细胞脱粒至相似比率(分别为72.3%和73.4%)(图4a),并且在来源于中-抗原血症小鼠的脾和肝cd8+t细胞中显示表达相当的il-2和tnfα,尽管程度较低(62%),所述细胞在肽刺激后也能脱粒,但缺乏il-2表达(图4b)。
为了确定多聚体结合细胞的增殖的分化状态,我们对cd127和klrg-1进行染色。wt和低-抗原血症hbvtg小鼠的疫苗接种在肝和脾中诱导了高百分比的s190-特异性cd127+klrg-1-多聚体结合细胞,该种细胞被认为是具有增殖潜力并产生新的效应细胞后代的长寿细胞的短暂前体(图4c)。然而,在中-抗原血症小鼠中,这些细胞很难被检测到(图4c)。这些数据表明hbv抗原表达减少了cd8+t细胞的多功能性,特别是效应细胞的il-2分泌和增殖能力。
实施例4:用于蛋白质初次疫苗接种的佐剂的比较
为了研究聚磷腈佐剂pcep将增强cpg的免疫刺激作用,使用pcep代替cpg或者将其加入至cpg以用于蛋白质疫苗制剂以及组合的hbsag和hbcag。
第0天,给野生型小鼠接种与相应的佐剂一起的16μghbsag(ayw亚型)和16μghbcag(ayw亚型)。第28天,将小鼠用mva-ph5-s(5×107i.u.;ayw亚型)和mva-核心(5×107i.u.)加强。加强后的第6天,分析血清中抗-hbs(图9a)和抗-hbc(图9b)的存在。加强后的第6天,采用hbsag(s190和s208adw)-或hbcag(c93)-特异性肽刺激后,通过ics分析脾细胞(图9c)。柱示出了背景扣除后对ifnγ染色阳性的cd8+细胞的百分比(平均值±sem)。i.u.:感染单位。
在wt小鼠中蛋白质初次/mva-加强疫苗接种后,pcep(单独或与cpg联合)的使用在诱导抗-hbs和抗hbc-抗体应答方面优于单独的cpg,而cd8+t细胞应答是相当的(图9a-c)。
实施例5:佐剂组合的比较
接下来的目的是确定不同佐剂对体液和细胞免疫应答的影响。因此,用在不同佐剂制剂中复合的颗粒状hbcag和hbsag初次免疫野生型ch57bi/6小鼠。在第1至3组(n=3)中,hbsag与氢氧化铝(alumnhydroxide)复合并与hbcag、cpg或聚磷腈佐剂或两者组合。第4至6组(n=3)使用采用cpg或聚磷腈或两者但没有采用任何铝(alumn)辅佐的颗粒状hbcag和hbsag接种。在第28天,将所有动物用表达核心和sadw亚型的mva加强。
28天(仅蛋白质初次免疫)后和34天(蛋白质初次免疫加mva加强)后测定针对hbs和hbc的抗体应答。图15a和15b表明,当未使用明矾(alum)时,抗体应答,特别是针对hbs的抗体应答出乎意料地更明显。图15c-d示出了在初次免疫后(图15c)和所述mva-加强后(图15d),针对s和核心表位的cd8+t细胞应答。
当疫苗制剂不含铝(alumn)时,在初次免疫后已可检测到t细胞应答。有趣的是,在所有组中以相同的效率进行mva加强(由b8r特异性应答表示)后,所有小鼠均产生核心特异性t细胞应答,而再次,在没有铝(alumn)的情况下接种的动物出现更明显的s-特异性cd8+t细胞应答。
实施例6:在高-抗原血症hbvtg小鼠中,用异源hbsag亚型疫苗接种破坏t细胞耐受并诱导强抗体产生
在接种疫苗后分析小鼠血清的中和能力。即使在高达1∶1000的高度稀释下,接种了hbsagadw亚型的小鼠也能够交叉中和hbvayw亚型(图15e)。
接下来,旨在增强异源蛋白质初次/mva-加强疫苗接种的免疫原性,以破坏在更高的hbv抗原负荷存在下的耐受性。
为了测试更强的抗原触发是否会提高疫苗接种效率,表达hbsag的新mva也工程化为在更强的启动子ph5的控制下(图5a)并且在第14天进行第二次蛋白质疫苗接种(图12)。另外,比较了ayw(与hbvtg小鼠相同)和adw亚型的hbsag。此外,在初次免疫和加强期间,在cpg中添加pcep用于蛋白疫苗制剂以及组合的hbsag和hbcag,以实现对多种hbv抗原的免疫应答。当应用该种修饰的疫苗接种方案(在第0天和第14天用cpg和pcep辅佐的组合的hbsag/hbcag初次免疫,然后在第28天使用在较强的ph5启动子下表达抗原的mva-s/mva-核心加强)时,能够破坏高抗原血症hbvtg小鼠和诱导的hbsag-和hbcag-特异性cd8+和cd4+t细胞中的耐受性(图5b、c)。
之后,研究疫苗和靶抗原之间的部分错配是否会改善疫苗效力(schirmbeck等,eurjimmunol2003;33:3342-52)。如用亚型特异性肽s208(图11)所测定的,无论是疫苗抗原、sayw还是sadw均诱导针对两种亚型的cd8+t细胞(图5b)。然而,含异种sadw的疫苗诱导更强的cd8+和可检测的cd4+t细胞应答(图5b、c)。与我们用来自中-抗原血症小鼠的s-特异性cd8+t细胞所观察到的(图4b)类似,发现脾s208-特异性cd8+t细胞分泌ifnγ并且在一定程度上分泌tnfα,但仅显示il-2的最低限度(marginal)表达(图5d)。
值得注意的是,只有含有sadw,而不是含有sayw的疫苗制剂能够在高抗原血症小鼠中诱导可检测的抗-hbs抗体应答(图6a),而两种制剂均能诱导抗-hbc抗体(图6b)。重要的是,由sadw产生的抗-hbs抗体能够中和ayw亚型的hbv颗粒(图6c)。伴随在sadw疫苗接种组中诱导中和性抗-hbs,hbsag水平显著下降至低水平(图6d)。总之,这表明修改的疫苗接种方案确实允许破坏hbvtg小鼠中的b-和t-细胞耐受性。
实施例7:由多抗原性mva诱导的广泛免疫应答
接下来,旨在比较使用多抗原性mva针对一种、两种和若干种hbv抗原的免疫应答的诱导。出乎意料的是,当所述mva疫苗载体共表达核心时,针对s或s和pol的体液以及细胞免疫应答得到改善。
实施例8:rnai和治疗性疫苗接种的组合
使用靶向所有hbvrna的3′区的hbv-特异性sirna处理表达高效价hbv抗原的hbv转基因小鼠品系hbvxfs。sirna处理使hbeag和hbsag水平降低了90%。8周后,给动物接种蛋白质初次-mva-hbv加强疫苗以诱导抗-hbs抗体和hbv特异性t细胞(图27)。作为蛋白质疫苗,颗粒状hbsag和hbcag以cpg和pcep辅佐。mva疫苗载体表达hbvs和核心蛋白的完整的开放阅读框。对照为无sirna、无疫苗接种及其组合。
加强疫苗接种后6天,处死小鼠,测定hbeag和hbsag水平以及抗-hbs效价(图28a、b)。将t细胞从肝和脾分离,用hbv特异性肽离体刺激,通过细胞内细胞因子染色对干扰素γ表达进行染色并通过流式细胞术分析(图28c)。
实施例9:最佳mva剂量的评估
在第一组实验中,我们旨在评价异源蛋白质-初次/mva-加强疫苗接种的最低mva剂量,该最低mva剂量将显示令人满意的免疫原性并且能够破坏hbvtg小鼠中的免疫耐受性。因此,以两周的间隔,用15μg二-(3′,5′)-环二聚单磷酸腺苷酸(c-di-amp)辅佐的颗粒状hbcag免疫低和中抗原血症hbvtg小鼠组两次。第28天,将小鼠用4种不同剂量的mva-核心(分别为3×106、1×107、3×107、1×108pfu)加强(图29a)。加强免疫后7天(第35天),评估由采用不同mva剂量的免疫方案引发的体液和细胞免疫应答。
分析第0天和第35天(加强后第7天)小鼠的血清中hbsag、hbeag、抗-hbs和抗-hbc抗体(图29b)。第35天,在hbvtg小鼠的血清中,全部免疫方案均引发了相似的抗-hbc抗体水平。此外,所有小鼠组均显示血液中hbsag的显著减少。这种效应不是由抗hbs抗体介导的,因为免疫方案未包括hbsag,这对于hbvtg模型中的hbsag转化是关键的。有趣的是,来自接受较高剂量mva-核心(3×107和1×108pfu)作为加强的组的hbvtg小鼠显示血清hbeag水平显著降低(图29b)。此外,在接受较高剂量的mva-核心(3×107和1×108pfu)的小鼠组中也观察到肝丙氨酸转移酶(alt)的轻微升高(图29c)。这些数据表明,这两组小鼠的疫苗接种方案导致肝中hbv复制受抑制,这可能是由于hbcag-特异性t细胞的活性增强。实际上,肝相关淋巴细胞(lal)和脾细胞的细胞内ifnγ染色显示,用较高剂量的mva-核心免疫的hbvtg小鼠可以(特别是在肝中)增加更有效的hbv-特异性cd8+t细胞应答(图29d)。同时,mva-特异性cd8+t细胞应答不随着用于免疫的更高mva剂量而显著增加。
从这些结果可以得出,用于异源蛋白质-初次/mva-加强疫苗接种的3×107pfu的mva-核心剂量足以破坏低和中抗原血症hbvtg小鼠的免疫耐受性。
实施例10:c-di-amp作为用于蛋白质初次免疫(priming)的佐剂的评估.
诱导平衡的th1/th2cd4+t细胞应答的安全有效的佐剂的缺乏可能是启动临床试验的障碍。此外,触发新识别的胞质模式识别受体sting是治疗性疫苗接种的有趣的替代方案。因此,我们旨在研究c-di-amp作为治疗性乙型肝炎疫苗的潜在新佐剂的功效。为了这一目的,中和高抗原血症hbvtg小鼠组(n=7)小鼠接受两次蛋白质初次免疫和mva加强免疫。将用于蛋白质初次免疫的颗粒状hbsag和hbcag组合并用c-di-amp或上文已建立的cpg和聚磷腈(pcep)的组合辅佐。第28天,将小鼠用mva-s/核心的混合物加强(图30a)。将接受c-di-amp注射和“空”mva(mvawt)的hbvtg小鼠(n=4)用作对照。第34天(加强后6天)比较疫苗制剂诱导体液和细胞免疫应答的功效。
在高抗原血症hbvtg小鼠中,c-di-amp和cpg/pcep免疫方案均未对血清hbeag水平产生影响(图30c)。两种测试的疫苗制剂诱导了显著的抗-hbc应答。然而,与cpg/pcep方案相比,采用c-di-amp的免疫诱导了显著更高的抗-hbc抗体效价(p<0.05)(图30c)。有趣的是,在全部检查的hbvtg小鼠中,两种免疫方案均导致hbsag向抗-hbs的血清转化(图30d)。由c-di-amp-或cpg/pcep辅佐的疫苗引发的高水平的抗hbc抗体与循环hbsag复合并将其从小鼠的血清中移除。相对地,接受仅c-di-amp并随后被mvawt加强的hbvtg小鼠没有产生任何抗-hbs,且这些小鼠血清中的hbsag水平保持不变。重要地,两种疫苗制剂均诱导在hbvtg小鼠脾中(p<0.05)、并且特别地在肝相关淋巴细胞中(p<0.05)可检测的显著的hbsag-特异性(s208)和hbcag-特异性(c93)cd8+t-细胞应答(图30e),伴随有由于alt增加导致的轻度t-细胞-诱导的肝损伤(图30b)。在由c-di-amp或cpg/pcep方案引发的hbv-特异性cd8+t细胞应答的量级上没有统计学显著差异。
鉴于这些数据,即使在高抗原血症hbvtg小鼠中,认为c-di-amp是用于治疗性蛋白质初次-mva加强疫苗接种的强力的佐剂。
实施例11:对于各种佐剂:c-di-amp、聚-lcic和rig-i配体,最佳递送途径的评估.
对于用于治疗性异源蛋白质-初次/mva-加强疫苗接种的蛋白质初次免疫的适当佐剂,扩大了筛选。我们的目标是比较c-di-amp与两种潜在的新佐剂(聚-lcic和rig-i配体)用于治疗性乙型肝炎疫苗的功效。此外,我们检查了各种免疫方案以找到最有效的应用途径。为此,使低和中抗原血症hbvtg小鼠组(n=5)接受两次蛋白质初次和mva加强免疫(图30a)。将用于蛋白质初次免疫的颗粒状hbsag和hbcag组合并用c-di-amp、聚-lcic或rig-i配体辅佐。第28天,将小鼠用mva-sag和mva-核心的混合物加强。仅通过肌肉内(i.m.)途径进行免疫,或者皮下(s.c.)施用蛋白质初次免疫,然后腹膜内(i.p.)加强。就第34天(加强后6天)诱导体液和细胞免疫应答,比较不同疫苗制剂和应用途径的功效。
所有的疫苗接种方案都有效地降低了hbvtg小鼠血清中的hbsag水平。这是由于以下事实:所有检查的佐剂和递送途径都可以引起高效价的与小鼠血液中的hbsag复合的抗-hbs抗体。类似地,这些小鼠组之间诱导的抗-hbc抗体的水平是相当的,其中轻微趋势是肌肉内免疫途径在诱导抗-hbc中更有效。然而,仅在通过i.m.或者s.c./i.p途径接受c-di-amp或者通过i.m.途径接受聚-lcic的hbvtg小鼠组中观察到,通过hbeag水平间接检测到的较低hbv复制(图31a)。不幸的是,用肌肉内方式进行聚-lcic免疫是唯一被检查的方案,其导致hbvtg小鼠体重明显下降(图31b)。可认为c-di-amp在诱导免疫的hbvtg小鼠的脾、特别是肝中hbcag-特异性(c93)和hbsag-特异性(s208)cd8+t细胞应答方面也是优越的,与采用哪种给药途径无关(图31c)。有趣的是,聚-lcic在肌肉内递送时导致剧烈的肝内hbcag-特异性cd8+t细胞应答,而当以s.c./i.p.途径递送时主要引发hbsag-特异性(s208)cd8+t细胞应答。rig-i配体能够诱导hbv-特异性体液应答,但未能诱导初次hbv特异性cd8+t细胞应答。作为对照,mva-特异性cd8+t细胞应答在所有免疫组的脾和肝中均相当,这表明了相同的疫苗接种效率。
这些数据表明c-di-amp是非常有效的佐剂,聚-lcic显示出中等效力,而rig-i配体对治疗性蛋白质初次-mva加强疫苗接种不足够有效。在i.m.和s.c./i.p两种应用途径中,c-di-amp同样有效。
实施例12:c57bl/6小鼠中新mva构建体(mva-hbvvac)的评估.
此外,评估了新构建的多顺反子mva的体内免疫原性,该mva表达hbsag、hbcag(覆盖主要hbva、b、c、d基因型的序列)和hbv聚合酶的rt结构域(mva-hbvvac)。产生两种多顺反子(polycystronic)疫苗接种构建体的示意图:涵盖所有主要hbv基因型的hbv核心和s以及hbv聚合酶的rt结构域的hbvvac,以及表达hbv核心和s的c/s(c/s)(图32a)。通过蛋白质印迹来验证蛋白质表达。图32b示出了表达hbvvac或s/c的不同重组mva-克隆的s-表达。
用c-di-amp辅佐的颗粒状hbsag和hbcag的混合物初次免疫c57bl/6小鼠组(n=5)一次。两周后,将小鼠用mva-s和mva-核心的混合物或等量的新mva-hbvvac加强。第21天将小鼠处死以评估hbv-特异性体液和细胞免疫应答(图32c)。
新的多顺反子mva引起显著的抗-hbs和抗-hbc抗体应答,与mva-s和mva-核心构建体的组合的混合物相当(图32d)。此外,用mva-hbvvac进行免疫引起了强烈的hbsag-特异性(s190、s208和spool)和hbcag-特异性(c93,cpool)cd8+t细胞应答(通过分析脾细胞测定),这些应答在量级上类似于由mva-s和mva-核心的混合物诱导的应答(图32e)。此外,即使没有将rt蛋白用于初次免疫,用mva-hbvvac免疫导致检测到对于肽rt61、rt333和rt肽库2(用箭头标记)的低水平的rt特异性cd8+t细胞应答。
这些数据表明,该多顺反子mva(mva-hbvvac)表达所有预期的蛋白质,并在c57bl/6小鼠中显示出优异的体内免疫原性。
实施例13:通过共表达cd70增加mva构建体的免疫原性.
为了改善基于mva的疫苗载体的体内免疫原性,构建了以双顺反子方式表达cd70的mva载体(图34a)。
用cpg和pcep辅佐的颗粒状hbcag初次免疫c57bl/6小鼠组(n=6-7)一次。两周后,用mva-核心或用另外表达cd70的等量mva核心-cd70或作为对照的野生型mva加强小鼠(图34b)。第35天,将小鼠处死以评估hbv特异性体液和细胞免疫应答。尽管在mva核心和mva核心-cd70加强后体液免疫应答相同,但在用mva核心-cd70加强的小鼠中,针对mva(b8r)特异性细胞因子的cd8+t细胞应答轻微增加,并且针对hbv(c93)特异性细胞因子的cd8+t细胞应答显著增加(图34c)。重复实验得出了相同的结果。
在第三个实验中,使在c57bl/6背景下喂养的hbv转基因小鼠组(n=5-6)接种。在这些动物中,在治疗性疫苗接种后可以破坏免疫耐受性。在用cpg和pcep辅佐的颗粒状hbcag初次免疫一次后,将小鼠用mva-核心或用等量的表达cd70的mva核心-cd70加强(图35a和b)。接种野生型mva的小鼠作为对照。第35天,将小鼠处死以评估mva-和hbv-特异性t细胞应答。与接种mva核心的小鼠相比,当小鼠接种mva核心-cd70时,门控到肝相关淋巴细胞上的cd8+t细胞在分别用mva-和hbv核心-特异性肽再刺激时显示出更显著的ifng和il2分泌。在用mvawt加强的小鼠中,检测到mva-特异性t细胞应答,但是未检测到hbv-特异性t细胞应答(图35c)。
关于上述结果,可以得出结论:cd70与hbv特异性抗原的共表达显著提高了体内mva-和hbv-特异性t细胞应答。
在本文中说明性阐述的本发明,适当地可在本文中未具体公开的任何一个或多个要素、限制不存在的情况下实施。因此,例如,术语“包含”、“包括”和“含有”等应当扩展性且无限制地理解。此外,本文已采用的术语和表达是用作描述而非限制的术语,并且不意图在使用这类术语和表达时将所示和所述特征的任何等价物或其部分排除在外,而是认识到,在要求保护的本发明的范围之内的多种修改是可能的。因此,应理解的是,尽管已通过示例性实施方案和可选特征具体公开本发明,但是本领域技术人员可采用本文公开的其中实施的本发明的修改和变化,并且此类修改和变化被认为在本发明的范围内。
本文中已经广泛地和一般性地描述了本发明。落入一般公开内容中的每个所述较窄的种类和亚属群组也构成本发明的一部分。这包括从属中移除任何主题的附带条件或否定限制的本发明的一般描述,不管去除的材料是否在本文具体列举。
其它实施方案在随后的权利要求范围内。此外,当以马库什组的方式阐述本发明的特征或方面时,本领域技术人员将认识到,本发明亦由此以该马库什组的任何单个成员或成员亚组的形式来阐述。
本文中所引用的所有文献和专利文件的内容均整体以引用的方式并入本文。
- 还没有人留言评论。精彩留言会获得点赞!