制备D-木糖酸盐的方法和棒状杆菌型细菌与流程
d-木糖酸(c5h10o6)是一种有机酸,其如toivari等人在applmicrobiolbiotechnol,2012,96(1):1-8中阐述的那样可以用作聚酰胺、聚酯和1,2,4-丁三醇的前体并因此在制药工业、食品工业和化学工业中具有广泛的应用潜力。以下的说明涉及d-木糖酸盐,即d-木糖酸的盐。d-木糖酸盐属于潜在高品质、基于第二代可再生原材料(例如含戊糖的半纤维素)的前体化学品的前30名。d-木糖酸盐与全球市场为80kt/年的d-葡糖酸盐(c6h11o7)相似。
d-木糖酸盐通过两步反应在一些细菌中天然形成。在第一反应中,d-木糖被氧化成d-木糖酸内酯,其中在此取决于生物体,特异性脱氢酶具有催化活性。然后可以通过特异性内酯酶或在没有酶催化剂的情况下自发地将d-木糖酸内酯转化为d-木糖酸盐。例如对于菌种氧化葡糖杆菌(gluconobacteroxydans)和莓实假单胞菌(pseudomonasfragi),报道描述了高产品滴度(produkttiter)的d-木糖酸盐(buchert等人,appliedmicrobiologyandbiotechnology,27(4):333-336,toivari等人,applmicrobiolbiotechnol,2012,96(1):1-8)。
此外,替代的d-木糖酸盐生产菌株(例如菌种酿酒酵母(saccharomycescerevisiae)的酵母、菌种大肠杆菌(escherichiacoli)的细菌和菌种黑曲霉(aspergillusniger)的真菌)通过例如来自新月柄杆菌(caulobactercrescentus)的d-木糖脱氢酶的异源表达生成,如(liu等人,bioresourtechnol,2012,115:244-248,richard等人,2012,us20120005788a1(us13/256,559),toivari等人,metabeng,2012,14(4):427-436)阐述。
除了微生物制备外,d-木糖酸盐也可以通过电化学(jokic等人,journalofappliedelectrochemistry,21(4):321-326)、通过酶(pezzotti等人,carbohydrres,2006,341(13):2290-2292)或通过化学氧化(isbell等人,bureauofstandardsjournalofresearch,1932,8(3):327-338)来制备。
已知的是,有机酸或它们的盐可以通过棒状杆菌型细菌菌株,特别是谷氨酸棒杆菌的发酵来制备。
文献us2013/0295621a1公开了通过基因技术修饰的细菌制备d-木糖酸的方法。
现有技术中已知的具有内源d-木糖酸盐合成能力的生产菌株,例如氧化葡糖杆菌不需要特定的培养基用于其生长,由此使得d-木糖酸盐的生产变得显然更复杂、更昂贵和因此更不经济。
到目前为止描述的所有“非天然”d-木糖酸盐生产者是在用于规范基因技术的法规(gentg)§3范围内的基因技术修饰的生物体,尤其是各种酵母、真菌和细菌;当生物体能够异源表达或者包含对于该生物体而言外源的基因,即经重组修饰时,该生物体根据所述法规是基因技术修饰的。由于复杂的许可程序,对于在特定的工业领域,例如食品工业和制药工业中的应用产生缺点。
需要提供一种方法和微生物,通过它们可以以高对映体纯度和高产率制备d-木糖酸盐和因此d-木糖酸。尤其应当提供一种微生物和方法,其可以在工业应用中,特别是在食品工业、制药工业和化学工业中应用并容易地满足用于食品工业和制药工业以及化学工业的许可条件。优选地应当避免与在用于规范基因技术的法规§3范围内的基因修饰的生物体相关的缺点,尤其是与重组修饰的生物体相关的缺点。尤其应当可以使用简单且成本有利的培养基用于培养,这种培养基便宜且易于使用。所使用的微生物应当在特定的培养基上具有高生长速率、实现高生物量产量并简化后续产品后处理。
从权利要求1和并列权利要求的上位概念出发,根据本发明通过在权利要求的特征部分中给出的特征来实现所述目的。
通过本发明的微生物、其基因组和本发明的制备方法,现在可以以高产率(例如>0.5克d-木糖酸盐/克d-木糖)以100%的对映体纯度制备d-木糖酸盐。本发明的微生物非常适合于工业使用,并且在一个优选实施方案中克服了在食品工业和制药工业以及化学工业中使用的许可标准,特别是它们根据基因技术法规被认为是非重组修饰的。本发明的微生物在特定的培养基上实现高生长速率和高生物量产量,并且实现简单的产品后处理。克服了现有技术的缺点。所述特定的培养基在使用时是成本更有利且较不复杂的。与在非特定的培养基的情况下相比,生产生物体的生长更具可再现性。培养基的复杂性低,因此减少了产品后处理所需的方法步骤的数量。
在从属权利要求中给出了本发明的有利扩展方案。
在下文中以其一般形式描述本发明,而不是限制性地解释。
已经令人惊讶地发现,使用其中编码肌醇代谢的调节因子的基因iolr(在下文中称为iolr基因)的活性降低或消除或者其中iolr基因至少部分缺失的棒状杆菌型细菌,可以由作为底物的d-木糖酸盐出发获得非常高产率的对映体纯d-木糖酸盐。
因此,本发明的主题是棒状杆菌型细菌,其中iolr基因的活性相对于野生型或野生型的突变体,优选含有该基因的非重组突变体降低或完全消除,或者其中所述iolr基因至少部分缺失;以及使用棒状杆菌型细菌制备d-木糖酸盐的方法,其中所述棒状杆菌型细菌的iolr基因在其活性方面降低或完全消除,或者所述iolr基因至少部分缺失。
根据本发明修饰的棒状杆菌型细菌可以源自于野生型,或者相对于野生型经基因修饰、优选非重组修饰的形式。
来自于野生型的菌株对于制药工业和食品工业而言特别优选,因为它们特别容易被许可生产。作为本发明方法的起始菌株,所述野生型包含天然的iolr基因。
基因修饰的菌株在其起始形式中同样包含天然的iolr基因。在本发明中的基因修饰的菌株是指例如如此基因修饰的菌株,从而使得更多地表达调节因子iolr的靶向或目标基因。由此可以过表达在肌醇代谢中重要的并且编码肌醇代谢的酶的所有或一部分基因。为此,例如可以将基因整合到编码这些酶并且允许这些基因的表达增加的基因组中。所述基因也可以以染色体方式引入。还可以设想用于肌醇代谢的酶的表达增加的其它活化机制,例如引入更强的启动子和调整转译效率。此外,还可以增加肌醇代谢的酶的稳定性。
特别地,可以使用如下类型的棒状杆菌型细菌:棒状杆菌(corynebacterium)、短杆菌(brevibacterium),特别是谷氨酸棒杆菌(corynebacteriumglutamicum)、醋麸酸棒状杆菌(corynebacteriumacetoglutamicum)、嗜热产氨棒杆菌(corynebacteriumthermoaminogenes)、黄色短杆菌(brevibacteriumflavum)、乳酸发酵短杆菌(brevibacteriumlactofermentum)或叉开短杆菌(brevibacteriumdivaricatum)。
优选为由野生型出发根据本发明修饰的棒状杆菌型细菌,例如使用如下野生型:
谷氨酸棒杆菌atcc13032
醋麸酸棒状杆菌atcc15806
嗜乙酰乙酸棒杆菌(corynebacteriumacetoacidophilum)atcc13870
嗜热产氨棒杆菌fermbp-1539
黄色短杆菌atcc14067
乳酸发酵短杆菌atcc13869和
叉开短杆菌atcc14020。所述列举不是限制性的,而是示例性的。
谷氨酸棒杆菌用作生产生物体提供了特别的优点,因为
i)其是可以在所有的工业领域中使用的“一般认为安全”(gras)-生物体;
ⅱ)在特定的培养基上因此实现高生长速率和生物量产量(unthan等人,biotechnolbioeng,2014,111(2):359-371),和
ⅲ)在工业应用中存在丰富的经验(becker等人,curropinbiotechnol,2012,23(4):631-640)。
在棒状杆菌型细菌,优选谷氨酸棒杆菌atcc13032中的肌醇代谢调节因子iolr,特别是iolr(cg0196)的部分失活、失活或部分缺失或完全缺失使得由d-木糖形成d-木糖酸盐。该优选实施方案的优点在于,其作为根据本发明修饰的野生型不遭受化学、制药或食品生产中的许可限制。
在所述优选的实施方案中,所述iolr基因是完全消除的。
这可以优选地通过iolr基因的完全缺失来实现。如果iolr基因的失活是完全的,这也可以通过部分缺失来实现。
所述iolr基因的完全失活也可以通过本领域技术人员已知的其它措施来实现。例如可以提及适当的培养(kulturführung),这例如通过使用肌醇作为iolr的底物/诱导物,将另一dna序列插入iolr基因,特别是iolr基因cg0196的基因区域中。在本发明中的基因区域是指其中的修饰仍对iolr基因的活性产生影响的区域。该区域可以位于iolr基因的开放阅读框之前、之后或之中。
完全失活还可以通过基因表达的信号结构的基因修饰(突变)通过iolr的基因表达的完全消除而实现。基因表达的信号结构是例如阻遏基因、激活基因、操纵基因、启动子、弱化子、核糖体结合位点、起始密码子和终止子。本领域技术人员例如在如下文献中找到对此的说明:专利申请wo96/15246,boyd和murphy(boyd等人,jbacteriol,1988,170(12):5949-5952),voskuil和chambliss(voskuil等人,nucleicacidsresearch,1998,26(15):3584-3590),jensen和hammer(jensen等人,biotechnolbioeng,1998,58(2-3):191-195),patek等人(patek等人,microbiology,1996,142(pt5):1297-1309)以及已知的遗传学和分子生物教科书,例如knippers的教科书("molekularegenetik",第8版,georgthiemeverlag,stuttgart,deutschland,2001)或winnacker的教科书("geneundklone",vchverlagsgesellschaft,weinheim,deutschland,1990)。
导致iolr的功能特性改变,特别是导致改变的底物特异性的突变同样可以导致iolr基因活性的完全消除。突变包括转换、颠换、插入和缺失,以及定向进化的方法。产生这种突变和蛋白质的指导属于现有技术中,并且可以从已知的教科书(r.knippers"molekularegenetik",2001年第8版,georgthiemeverlag,stuttgart,deutschland)或综述文章(pokala等人,jstructbiol,2001,134(2-3):269-281,jaenicke等人,angewchemintedengl,2003,42(2):140-142,lilie,emborep,2003,4(4):346-351,pei等人,procnatlacadsciusa,2003,100(20):11361-11366,dokholyan,proteins,2004,54(4):622-628,tramontano,angewchemintedengl,2004,43(25):3222-3223)获知。
以这种方式产生的棒状杆菌型细菌绝对没有iolr基因活性,并且导致对映体纯d-木糖酸盐的高产率生产。这是本发明的最佳实施方案,特别优选为完全缺失。
在谷氨酸棒杆菌atcc13032δiolr的情况下,所述iolr基因cg0196完全缺失。
在另一个实施方案中,iolr基因的活性降低。
这也可以通过部分缺失来实现,只要活性由此没有完全消除。
所述iolr基因的部分失活也可以通过本领域技术人员已知的其它措施来实现。例如可以提及适当的培养,这例如通过使用肌醇作为iolr的底物/诱导物,将另一dna序列插入iolr基因,特别是iolr基因cg0196的基因区域中。在本发明中的基因区域是指其中的修饰仍对iolr基因的活性产生影响的区域。该区域可以位于iolr基因的开放阅读框之前、之后或之中。
部分失活还可以通过基因表达的信号结构的基因修饰(突变)通过iolr的基因表达的部分消除而实现。基因表达的信号结构是例如阻遏基因、激活基因、操纵基因、启动子、弱化子、核糖体结合位点、起始密码子和终止子。原则上,对此可以使用如在完全缺失的情况下相同的方法。
导致iolr的功能特性改变,特别是导致改变的底物特异性的突变同样可以导致iolr基因活性的部分消除。突变包括转换、颠换、插入和缺失,以及定向进化的方法。同样,可以使用上述方法。
所给出的本发明的基因修饰产生本发明的棒状杆菌型细菌,以及在分离的情况下产生本发明的基因组。
iolr基因cg0196显示在序列表编号1中。
具有开放阅读框的棒状杆菌型细菌atcc13032的iolr基因显示在序列编号2中。
令人惊讶地,在具有用于生长的碳源和能量源例如d-葡萄糖和作为其它碳源和能量源的d-木糖的特定培养基上的iolr基因失活导致d-木糖酸盐的形成。d-木糖酸盐自然不会被棒状杆菌型细菌代谢,因此在培养基中积累,随后可以从培养基中纯化。
根据本发明修饰的棒状杆菌型细菌生物体以高产率产生100%对映体纯的d-木糖酸盐或d-木糖酸。以这种方式产生的本发明基因组使得棒状杆菌型细菌能够以高产率和100%的对映体纯度制备d-木糖酸盐或d-木糖酸。
通过本发明的棒状杆菌型细菌生产d-木糖酸盐可以用实验室或生产中常用的已知发酵方法进行。
根据本发明制备的微生物可以连续或不连续地以分批方法(批次培养)或补料分批(进料法)或重复补料分批(重复进料法)培养以进行d-木糖酸盐生产。已知培养方法的概述描述在chmiel的教科书(bioprozesstechnik1.einführungindiebioverfahrenstechnik(gustavfischerverlag,stuttgart,1991))或者storhas的教科书(bioreaktorenundperiphereeinrichtungen(viewegverlag,braunschweig/wiesbaden,1994))中。
使用的培养基必须适当地满足各微生物的要求。各种微生物的培养基的说明包含在美国细菌学学会的的手册“manualofmethodsforgeneralbacteriology”(washingtond.c.,usa,1981)中。
除了d-木糖作为形成d-木糖酸盐的起始底物外,还可以使用糖和碳水化合物例如葡萄糖、蔗糖、乳糖、果糖、麦芽糖、糖蜜、淀粉和纤维素,油和脂肪例如大豆油、葵花籽油、花生油和椰子脂肪,脂肪酸例如棕榈酸、硬脂酸和亚油酸,醇例如甘油和乙醇,以及有机酸例如乙酸作为用于生长的碳源。这些物质可以单独使用或作为混合物使用。特别优选的是具有d-木糖和d-葡萄糖作为碳源和能量源的cgxii培养基(unthan等人,biotechnolbioeng,2014,111(2):359-371)。由此可以实现高达0.61h-1的谷氨酸棒杆菌atcc13032的高比生长速率。此外,cgxii培养基的使用确保了目标产物d-木糖酸盐与培养上清液容易分离,这例如通过乙醇沉淀与然后真空干燥(liu等,bioresourcetechnology,2012,115:244-248)。简单的培养基是要实现的目的的一部分。
作为氮源,可以使用含氮的有机化合物例如蛋白胨、酵母提取物、肉提取物、麦芽提取物、玉米浆、大豆粉和尿素,或者无机化合物例如硫酸铵、氯化铵、磷酸铵、碳酸铵和硝酸铵。氮源可以单独使用或作为混合物使用。作为磷源,可以使用磷酸二氢钾或磷酸氢二钾,或相应的含钠盐。
此外,所述培养基应当含有金属盐,例如生长所需的硫酸镁或硫酸铁。
最后,除了上述物质之外,还可以使用必要的生长素如氨基酸和维生素。
所述原料可以以一次批次的形式加入培养物中或在培养期间以合适的方式加料。
为了控制培养物的ph,可以适当地使用碱性化合物例如氢氧化钠、氢氧化钾、氨,或者酸性化合物例如盐酸、磷酸或硫酸。细胞生长的优选ph范围为6至8。
为了控制起泡,可以使用消泡剂,例如脂肪酸聚二醇酯。
为了维持需氧条件,优选向培养物中引入氧气或含氧气体混合物,例如空气。
培养物的温度通常为20℃至45℃,优选为25℃至40℃。
优选继续培养,直至形成最大量的d-木糖酸盐。这个目的通常在10小时至160小时内达到。
附图显示了实施例的实验结果,其证明了本发明的优点。
其显示了:
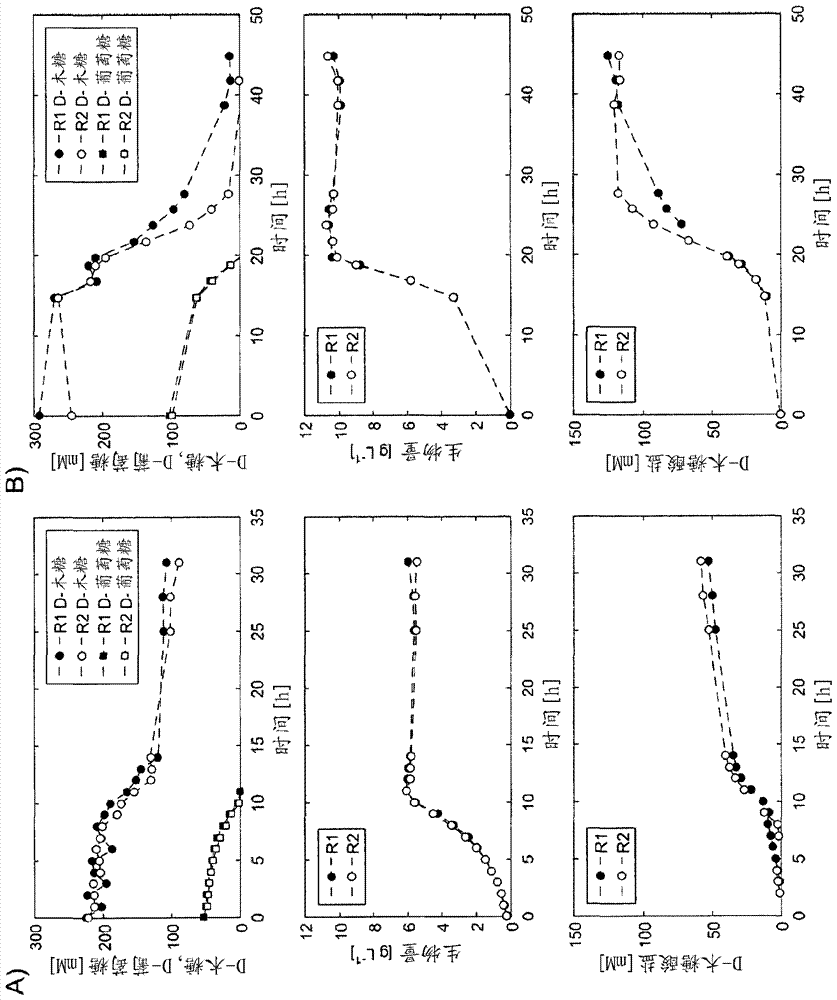
图1:菌株谷氨酸棒杆菌δiolr的培养曲线;
图2:纯化沉淀物的gc-tof-ms谱图。
在图1中,在两个子图a)和b)中显示了菌株谷氨酸棒杆菌δiolr的培养曲线。所有子图中图表的横坐标是以小时h为单位的时间。第一行的纵坐标表示以毫摩尔[mm]为单位的d-木糖和d-葡萄糖的量。第二行的纵坐标表示以克/升[g/l]为单位的生物量,且第三行的纵坐标表示以毫摩尔[mm]为单位的d-木糖酸盐的量。
子图1a显示了在分批方法中菌株谷氨酸棒杆菌δiolr的培养曲线。达到d-木糖酸盐的最终滴度为9.2±0.6g/l。比产率和生产速率为yp/s=0.49±0.01gd-木糖酸盐/gd-木糖和0.30±0.02g/(lh)。子图1b显示了在补料分批方法中菌株谷氨酸棒杆菌δiolr的培养曲线。达到d-木糖酸盐的最终滴度为20.0±1.0g/l。比产率和生产速率为yp/s=0.51±0.02gd-木糖酸盐/gd-木糖和0.45±0.02g/(lh)。
图2显示了通过gc-tof-ms分析产生的纯化沉淀物的谱图(paczia等,microbialcellfactories,2012,11)。其中绘制了纵坐标中的相对峰强度与作为横坐标的色谱运行时间。
实施例:
下面作为实施例给出实验证据,这不解释为限制性的。
为了产生菌株谷氨酸棒杆菌δiolr,使阅读框中(“框内”)的基因iolr缺失,从而尽可能不影响位于下游的基因的表达。为此,首先进行交叉pcr。在两个第一pcr中,将待缺失的基因的两个分别400bp长的位于下游和上游的侧翼dna区域分开扩增。将为此使用的寡核苷酸(iolr_df1_forcccaagcttgaggtacttgccgaaagattg(hindiii),iolr_df1_revctcgattacttggccggagggctacttggaagtagagg,iolr_df2_fortccggccaagtaatcgag,iolr_df2_revcgggatccatcgcgttggcattcttc(bamhi))如此修饰,以使得产生的pcr片段在5'-端分别与18个核苷酸长的同源序列连接,该同源序列在所述片段的情况下彼此互补。对于交叉pcr,第一pcr的产物用作dna模板,其中通过互补区域在反应过程中发生积聚(anlagerung)。通过使用外部引物,由此产生缺失构建体,其在载体pk19mobsacb中克隆。在将所述不可自由复制的载体转化到谷氨酸棒杆菌中和随后对由pk19mobsacb-δiolr2介导的卡那霉素抗性选择之后,分离其中质粒通过同源重组整合到染色体中的克隆。通过在没有卡那霉素的情况下在完全培养基中培养这些细胞,可以通过现在在染色体中双重存在的dna区域发生第二重组事件。将细菌培养物以不同的稀释在经由10%蔗糖的lb培养基上铺板。由于通过sacb在pk19mobsacb-δiolr2上编码的果聚糖-蔗糖酶,蔗糖聚合成果聚糖。该聚合物进而导致致死性。因此,只有那些通过第二重组已失去质粒的克隆可以生长。在此时蔗糖抗性和卡那霉素敏感的克隆的情况下,又建立出野生型基因状况,或者所希望的基因缺失。在谷氨酸棒杆菌的基因组中的缺失通过具有引物的pcr确认,该引物与所述基因外的区域互补(k_diolr_forgcacgttatgacctgcaaactc,k_diolr_revtacggtctggctatctacatcc)。
以这种方式产生的菌株谷氨酸棒杆菌δiolr以分批和补料分批方法在1.5l生物反应器(dasgipag,jülich,德国)中在30℃和1vvm的恒定通气速率下培养。连续的需氧条件(>30%的溶解氧)通过具有匹配的搅拌器转数(400-1200rpm)的氧气引入而实现。培养基中的ph值用5mh3po4和5mnh4oh恒定调节在ph7.1。分批方法在两个独立的反应器中由具有40g/ld-葡萄糖的cgxii培养基上的od为1的过夜预培养物(unthan等人,biotechnolbioeng,2014,111(2):359-371)接种在不含尿素且具有10g/ld-葡萄糖以及30g/ld-木糖的cgxii培养基中。补料分批方法在两个独立的反应器中由具有40g/ld-葡萄糖的od为1的cgxii培养基上的过夜预培养物接种在不含尿素且具有20g/ld-葡萄糖以及40g/ld-木糖的cgxii培养基中。在使用葡萄糖限制的20h处理时间之后,开始以2.5ml/h(反应器1)和5ml/h的速度(反应器2)连续地进料100g/ld-葡萄糖。所有培养物每隔一定时间采样,将样品离心,并且使用所得到的无细胞的上清液用于底物和产物分析。对于底物分析(d-葡萄糖和d-木糖),采用两种酶法测定(unthan等,microbcellfact,2015,14:32),产物(d-木糖酸盐)借助hplc(300×8mm有机酸柱,cs色谱法,langerwehe,德国)在80℃和具有0.1mh2so4的等度洗脱和0.6ml/min的流动速率下通过dad检测器(agilent,santaclara,ca,usa)进行定量。
在补料分批培养结束时,完全收获清空(abgeerntet)反应器用于产物纯化和分析。对于产物纯化,如下使用现有方案(liu等,bioresourcetechnology,2012,115:244-248):1.通过离心(4500rpm,在4℃下10分钟)分离细胞;2.将所得到的上清液在活性炭(ac)中脱色;3.将经ac处理的上清液过滤(0.22μm)并通过旋转蒸发仪(100mbar,60℃水浴)浓缩;4.将浓缩物过滤(0.22μm)并通过加入etoh(3:1,v/v)沉淀d-木糖酸盐;在-10℃下将产物真空干燥至少12小时。
具有开放阅读框的iolr基因序列(粗体)以及上游和下游250bp。
- 还没有人留言评论。精彩留言会获得点赞!