一种抗埃博拉病毒糖蛋白GP2亚基的单克隆抗体2G1及应用的制作方法
本发明公开了一种抗体,属于多肽
技术领域:
。
背景技术:
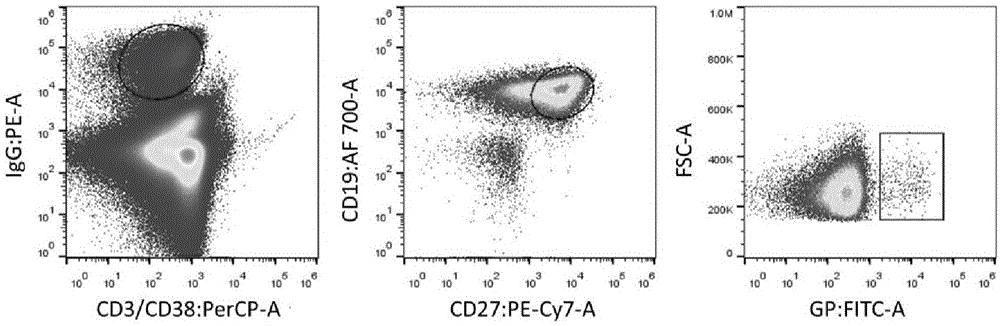
:埃博拉病毒(ebolavirus,ebov)能在人类和非人灵长类体内引发急性严重出血热病,是迄今发现的致死率最高的病毒之一,致死率高达90%,可直接通过接触传播,具有极强的传染性和致死率。埃博拉病毒包膜表面上的糖蛋白(glycoprotein,gp)刺突几乎介导了病毒进入细胞的所有环节,因此埃博拉gp是一个重要的病毒中和抗体作用靶点。埃博拉病毒gp基因被加工成两种蛋白,一种是分泌的非结构型gp(secretedglycoprotein,sgp);另一种是结构型gp。埃博拉gp首先作为一个多肽被合成,接着被furin酶酶切成为gp1(1-501位氨基酸)和gp2(502-676位氨基酸)亚基,两个亚基通过二硫键相连接,形成的三聚体由gp2内部的跨膜区固定在于包膜表面。gp1包含黏蛋白(mucin)、聚糖帽(glycancap)、头部(head)、基部(base)等结构域。furin酶切后形成的gp结构并不能直接激发埃博拉病毒与宿主细胞膜融合过程的发生。埃博拉病毒进入体内后,与细胞膜表面受体结合,但是这些宿主细胞膜表面的粘附因子,并不是真正的埃博拉病毒发生膜融合进入宿主细胞的受体。病毒结合于细胞表面粘附因子后,由网格蛋白介导胞吞和胞饮,发生初级和次级核内体的运输。gp在核内体中被组蛋白酶b和组蛋白酶l酶切,去除gp1上包括黏蛋白和聚糖帽在内的60%的氨基酸,形成激活态gp(primedgp,gpcl),此时激活了决定性的膜融合过程的发生。gpcl与内体膜蛋白niemann-pickc1(npc1)结合发生膜融合,从而使病毒rna进入胞质完成病毒基因组复制和转录,新病毒蛋白合成后组装成子代病毒颗粒,并从宿主细胞表面出芽。内体膜蛋白npc1是ebov感染的必不可少的宿主感染因子。埃博拉病毒被美国nih和cdc列为a类生物剂/生物恐怖剂,将其归为对国家安全和公众健康存在重大隐患和风险的病原体之一,目前仍没有获批的预防或治疗埃博拉病毒的药物。1967年埃博拉病毒被发现后,共经历了12次较大规模的传播。2014年西非爆发了历史上最大规模、最难控制的埃博拉疫情,经鉴定为扎伊尔型埃博拉病毒,此次疫情造成1万多例死亡和2.5万多例感染(根据who2016年2月17日发布的埃博拉疫情报告),疫情首次传播到非洲大陆以外,在世界范围内引起了极大的恐慌。公众和当局的高度关注快速推进了埃博拉疫苗和抗病毒药物的研究。国内外迅速开展了几个有前景的埃博拉疫苗临床试验:大猩猩腺病毒3型载体疫苗、腺病毒5型载体疫苗、水疱性口炎病毒载体疫苗等。不可否认地,发展有效的预防类药物是一个重要的目标,并且预防措施被认为可以挽救大量的生命,然而疫苗在某些情况下难以发挥作用,比如:1)宿主个体可能存在差异(如年长者、年幼者及免疫受损者的免疫能力薄弱);2)疫苗长时间免疫后免疫效果逐渐减弱;3)疫苗激发宿主的免疫无法抵抗高剂量的埃博拉病毒暴露;4)大部分病人仅在出现埃博拉病毒感染信号后才寻求药物帮助,疫苗在短暂的治疗窗口期内难以激发宿主有效的免疫应答。因此,在开发疫苗的进程中埃博拉病毒病治疗药物的研发同样不容忽视。目前埃博拉病毒中和抗体可能通过三种途径发挥保护作用:1)阻断gp被组织蛋白酶酶切水解;2)阻断激活态gp与npc1受体的结合;3)影响与npc1受体结合后到膜融合前gp的变构过程。其中,前两种途径已有文献证实,第三种途径为部分文献的推测,尚无直接证据。尽管目前没有批准的埃博拉病毒病治疗药物,但是有一些实验型抗埃博拉病毒药物正在研究当中,包括小干扰rna、反义寡核苷酸、核苷酸类似物、抗体等。2018年5月,埃博拉疫情在刚果民主共和国再一次爆发,截止到8月12日,造成57人感染,41人死亡。在此次疫情爆发后,who批准了5种处于研究阶段的药物,可以被用于应急治疗埃博拉病毒病,其中包括3种抗体药物(zmapp、regn3470-3471-3479和mab114)和2种小分子药物(remdesivir和favipiravir)。其中,由三株单克隆抗体混合而成的抗体药物“zmapp”及其优化株“mil77”在2014年西非埃博拉疫情爆发期间,成功挽救了多人的生命,其效果和治疗窗口期长度远超于其他种类的抗病毒药物,大大提高了药物安全性和患者治愈率,给了人们极大的鼓舞。应用抗体药物治疗丝状病毒病引起了世界范围内的关注,成为埃博拉病毒病治疗药物研究领域的热点。zmapp是由烟草系统表达的三株人鼠嵌合单抗(c2g4、c4g7、c13c6)组成的鸡尾酒治疗策略,其中c2g4(表位为c511,n550,g553,c556)和c4g7(表位为c511,d552,c556)结合于gp2亚基,表位存在交叠;c13c6(表位为t270,k272)以近似垂直的角度结合于聚糖帽区域。mil77是优化于zmapp的抗体组合,mil77-1/-2/-3分别含有c2g4、c4g7和c13c6的可变区,其骨架区经人源化改造后表达于cho细胞。mil77-1和ml77-3两株抗体的鸡尾酒组合疗法在感染72小时后能够对非人灵长类起到100%的保护活性。鸡尾酒组合疗法所显示的优异效果为埃博拉病毒病的治疗带来了新的启示,研发能够抗独特抗原表位的抗埃博拉病毒单抗成为本领域的一大技术需求,因为如果具有了更多的、不同于现有技术已经出现的抗独特抗原表位的抗埃博拉病毒单克隆抗体,对于埃博拉病毒病的治疗方案就有了更多的选择,尤其是鸡尾酒组合制剂的方案设计。因此本发明的目的就是提供一种能够针对独特抗原表位的抗埃博拉病毒糖蛋白的单克隆抗体,进而提供其在制备埃博拉病毒病治疗药物中的应用。技术实现要素:基于上述目的,本发明首先提供了一种抗埃博拉病毒糖蛋白gp2亚基的单克隆抗体,所述抗体轻链可变区的cdr1、cdr2和cdr3的氨基酸序列分别如seqidno.1的第27-32、50-52和89-96位氨基酸序列所示,所述抗体重链可变区的cdr1、cdr2和cdr3的氨基酸序列分别如seqidno.5的第26-33、51-58和97-113位氨基酸序列所示。在一个优选的实施方案中,所述抗体轻链可变区的氨基酸序列如seqidno.1所示,所述抗体重链可变区的氨基酸序列如seqidno.5所示。在一个更为优选的实施方案中,所述抗体轻链恒定区的氨基酸序列如seqidno.3所示,所述抗体重链恒定的氨基酸序列如seqidno.7所示。其次,本发明还提供了一种上述单克隆抗体轻链和重链的基因编码序列,所述抗体的轻链可变区的基因编码序列由seqidno.2所示,所述抗体的重链可变区的基因编码序列由seqidno.6所示。在一个优选的实施方案中,所述抗体轻链恒定区的基因编码序列由seqidno.4所示,所述抗体重链恒定区的基因编码序列由seqidno.8所示。再次,本发明提供了一种能够表达上述编码单克隆抗体重链和/或轻链的基因编码序列的功能元件。在一个优选的实施方案中,所述功能元件为线性表达框。又次,本发明提供了一种含有上述线性表达框的宿主细胞,所述细胞为293t细胞。最后,本发明提供了上述的单克隆抗体在制备埃博拉病毒病治疗药物中的应用。在一个优选的实施方案中,所述单克隆抗体作为埃博拉病毒病鸡尾酒疗法制剂中的组分使用。本发明提供的抗埃博拉病毒包膜糖蛋白的单克隆抗体具有独特的cdr区,在抗原结合能力上显示了优异的广谱性,与ebovgp、bdbvgp、sudvgp和restvgp具有良好的结合活性,ec50值分别为0.0059μg/ml、0.0075μg/ml、0.0048μg/ml和0.026μg/ml。稳定性试验表明,在四种不同ph的环境中,2g1与gpdm的结合活性基本无变化,证明2g1与gp的结合稳定,不受次级内体低ph环境的影响,为其发挥中和作用提供了基础。与对照抗体相比,2g1在体外可有效地中和ebov、bdbv和sudv假病毒。2g1中和活性随着抗体浓度的升高而增强,在1μg/ml的浓度下即可对三种埃博拉假病毒实现近100%的保护。本发明提供的抗体还具有独特的作用位点,不同于现有技术中已经出现的抗埃博拉病毒包膜糖蛋白单克隆抗体的作用位点,提示2g1具有与其他中和抗体组成鸡尾酒组合疗法的潜力。附图说明图1.流式分选记忆b细胞结果图;图2.巢式pcr扩增抗体基因的电泳鉴定图谱;图3.线性表达框的拼接顺序示意图;图4.抗体结合力筛选示意图;图5.各抗体与抗原结合的光谱性对照分析图;图6.2g1与不同gp抗原的结合活性曲线图;图7.2g1与不同假病毒的中和活性曲线图;图8.截短型gp构建示意图;图9.2g1与各截短型抗原的结合活性曲线图(1);图10.2g1与各截短型抗原的结合活性曲线图(2);图11.抗体之间的竞争结合值示意图;图12.2g1与gp在不同ph下的结合活性曲线图。具体实施方式下面结合具体实施例来进一步描述本发明,本发明的优点和特点将会随着描述而更为清楚。但这些实施例仅是范例性的,并不对本发明的保护范围构成任何限制。实施例1:抗体制备1.血液样品的采集在获得知情同意书后,采集重组埃博拉疫苗临床试验受试者第二次免疫28天后血液样品5ml,用于后续实验。2.fitc标记蛋白gpdm需要通过荧光标记的抗原来分选特异的记忆b细胞,fitc标记截短型抗原蛋白gpdm方法如下:1)fluoresceinisothiocyanate_fitc(sigma,f4274)溶于dmso,浓度为20mg/ml。2)取100μlgpdm(3.3mg/ml),加缓冲液(ph9.6碳酸盐缓冲液)至400μl。3)加8μlfitc至gpdm溶液中,4℃避光孵育3h。4)用50kd的离心浓缩管对溶液加pbs换液,直到滤过液为透明无色。将标记好的蛋白用锡箔纸包好,4℃存放待用。3.流式分选记忆b细胞将采集的血样利用ficoll密度梯度离心法分离pbmc,过程如下:1)取新鲜抗凝全血,edta抗凝。用等体积pbs稀释全血。2)在离心管中加入一定体积的分离液,将稀释后的血样平铺到分离液液面上方,保持两液面界面清晰。分离液、抗凝未经稀释全血、pbs(或生理盐水)体积为1:1:1。3)配平,室温,水平转子800g,加速度3acc,离心30min。4)离心结束后,管底是红细胞,中间层是分离液,最上层是血浆/组织匀浆层,血浆层与分离液层之间是一层薄且较致密的白膜,即:单个核细胞(包括淋巴细胞和单核细胞)层。小心吸取白膜层到另一离心管中。5)用pbs稀释到一定体积,颠倒混匀。室温,水平转子300g,离心10min,弃上清。重复洗涤2次。6)用pbs将淋巴细胞重悬备用。7)将用来分选的细胞计数,取1×106个细胞,体积为100μl,加入下表推荐量的6种荧光染料,4℃避光孵育1h。标记荧光公司/货号体积(每1×106cells)抗原fitcsigma,f42742μliggpebd,55578740μlcd19apc-af700beckman,im247010μlcd3percpbd,55285120μlcd38percpbiolegend,30352010μlcd27pc7beckman,a5482310μl8)使用含2%fbs的pbs重复洗涤2-3次,400μlfpbs重悬,用40μm细胞筛去除细胞团,4℃避光保存供分选。9)使用细胞分选仪(sony,sh800s)分选gp特异的单个记忆b细胞。分选策略为:igg+/cd3-/cd38-/cd19+/cd27+/gpdm+,直接将单个浆细胞分选至96孔板中,96孔板中每孔含有5urna酶抑制剂及19.8μl去rna酶的水中,-80℃保存。图1是细胞分选结果图。图中方框圈出部分的细胞特征为igg+/cd3-/cd38-/cd19+/cd27+/gpdm+,即期望的gp特异性记忆b细胞。4.单细胞pcr扩增全人源单抗1)反转录pcr分选得到80个gpdm特异的记忆b细胞,向每个反应体系中同时加入以下全部的针对重链h、κ轻链、λ轻链各亚型的特异引物(引物序列见表1)。表1反转录pcr引物pcr反应体系中包含:5×缓冲液6μl、dntp1.2μl、反转录酶1.2μl、引物如上、模板为单细胞,水补齐至30μl。pcr反应条件为:50℃反转录30min,95℃预变性15min,接着95℃40s,55℃30s,72℃1min,40个循环,最后72℃延伸10min。2)巢式pcr以反转录产物为模板,分别进行3次巢式pcr反应扩增h、κ、λ(引物序列见表2),具体过程如下:表2巢式pcr引物pcr反应体系中包含:10×缓冲液2.5μl、10mmdntp0.5μl、dna聚合酶0.25μl、引物如上、模板为反转录产物1μl、水补齐至25μl。pcr反应条件为:94℃预变性4min,接着94℃30s,57℃30s,72℃45min,40个循环,最后72℃延伸10min。3)琼脂糖凝胶电泳一个单细胞中重链和轻链基因均扩增成功的克隆,被认为是配对成功的克隆。取5μl巢式pcr扩增产物经1%琼脂糖凝胶电泳后,取配对的阳性克隆进行测序,测序获得的抗体可变区序列用vectornti软件及imgt网站进行分析,并进行抗体蛋白表达和功能验证。图2是对h、κ、λ三种链基因的巢式pcr扩增后琼脂糖凝胶电泳的鉴定图谱。只有当重链和轻链可变区基因从同一个记忆b细胞中扩增获得时,认为是自然配对的抗体基因,并将对应巢式pcr产物测序做进一步研究。5.线性表达框表达抗体通过上述反转录反应,单细胞克隆中获得了25个配对的抗体序列,鉴于传统的克隆表达方法费时费力,构建线性表达框的方法则可以快速表达抗体。该方法的基本原理是直接通过重叠延伸pcr将启动子序列(genbank登记号:x03922.1)、抗体前导肽的编码序列、抗体可变区(从单细胞中扩增获得)、抗体恒定区(生工生物合成)、多聚a尾(genbank登记号:x03896.1)连接起来,将该线性形式的dna转染进入细胞中进行抗体表达。具体过程如下:1)以psectag2(invitrogen)为模板,扩增启动子-前导序列片段、多聚a尾片段。扩增启动子-前导序列片段的pcr反应体系中包含:模板质粒psectag2(invitrogen)1ng,10×缓冲液5μl、10mmdntp1μl、dna聚合酶0.5μl、引物5'cmv-forward(cgatgtacgggccagatatacgcgttg)、引物3'leader-sequence(gtcaccagtggaacctggaaccca)、水补齐至50μl。扩增多聚a尾片段的pcr反应体系中包含:模板质粒psectag2(invitrogen)1ng,10×缓冲液5μl、10mmdntp1μl、dna聚合酶0.5μl、引物5'poly(a)(gcctcgactgtgccttctagttgc)、引物3'poly(a)(tccccagcatgcctgctattgtct)、水补齐至50μl。pcr反应条件为:94℃预变性4min,接着94℃30s,60℃30s,72℃1min,30个循环,最后72℃延伸10min。2)扩增抗体恒定区h链恒定区pcr体系中包含:重链恒定区模板10ng、10×缓冲液5μl、10mmdntp1μl、dna聚合酶0.5μl、引物5'ch(accaagggcccatcggtcttcccc)、引物3'ch(gcaactagaaggcacagtcgaggctttacccggagacaggga)、水补齐至50μl。κ链恒定区pcr体系中包含:κ链恒定区模板10ng、10×缓冲液5μl、10mmdntp1μl、dna聚合酶0.5μl、引物5'cκ(actgtggctgcaccatctgtcttc)、引物3'cκ(gcaactagaaggcacagtcgaggcacactctcccctgttgaagct)、水补齐至50μl。λ链恒定区pcr体系中包含:λ链恒定区模板10ng、10×缓冲液5μl、10mmdntp1μl、dna聚合酶0.5μl、引物5'cλ(gaggagcttcaagccaacaaggccaca)、引物3'cλ(gcaactagaaggcacagtcgaggctgaacattctgtaggggccac)、水补齐至50μl。pcr反应条件为:94℃预变性4min,接着94℃30s,60℃60s,72℃3min,30个循环,最后72℃延伸10min。3)扩增抗体可变区pcr体系中包含:模板为反转录pcr产物10ng,10×缓冲液5μl、10mmdntp1μl、dna聚合酶0.5μl、引物如表3中所示(将重链和轻链引物分别混合后加入体系中)、水补齐至50μl。pcr反应条件为:94℃预变性4min,接着94℃30s,60℃30s,72℃3min,30个循环,最后72℃延伸10min。表3.线性表达框构建pcr引物4)pcr产物回收纯化:将以上pcr产物经1%的琼脂糖凝胶电泳后,切胶并使用omega公司回收试剂盒回收。5)分别扩增重链和轻链的线性表达框线性表达框的拼接顺序示意图如图3所示。图3中,a为h链线性表达框,b为κ链线性表达框,c为λ链线性表达框。pcr反应体系中包括:模板:纯化后的启动子-前导序列片段10ng、重链/轻链可变区片段10ng、重链/轻链恒定区片段10ng、多聚a尾片段10ng,10×缓冲液2.5μl、10mmdntp0.5μl、dna聚合酶0.25μl、引物5'cmv-forward(cgatgtacgggccagatatacgcgttg)和3'poly(a)(tccccagcatgcctgctattgtct)、水补齐至25μl。pcr反应条件为:94℃预变性4min,接着94℃30s,60℃30s,72℃3min,30个循环,最后72℃延伸10min。6)pcr产物回收纯化:pcr产物直接用omega公司回收试剂盒回收。7)dna定量:用nano(gehealthcare)对pcr回收产物进行定量。8)细胞接种:将293t细胞以2×105/ml接种于24孔细胞培养板中,在含有5%co2的细胞温箱中,37℃培养过夜。9)细胞共转染:次日,向200μl无血清的mem培养基中,加入构建成功的重链和轻链线性表达框pcr产物各1μg,混匀后加入4μl转染试剂turbofect(thermoscientific,r0531),共同孵育15-20min后逐滴加至过夜培养的293t细胞培养孔中。在含有5%co2的细胞温箱中,37℃培养48h后收细胞培养上清备用。6.elisa筛选具有结合活性的抗体1)实验前一天96孔酶联板,包被1μg/ml的ebovgp,每孔100μl包被。将包被的酶联板放入湿盒,4℃过夜。2)实验当天用洗板机(bio-tek,405_ls)洗5次。3)每孔加入100μl封闭液,室温下放置1小时。4)洗板机洗5次。5)加入100μl的转染细胞培养上清,室温静置1小时。6)洗板机洗5次。7)将hpr标记的羊抗人igg二抗(abcam,ab97225)以1:10000用稀释液进行稀释,每孔100μl加入到elisa板对应孔中,室温孵育1小时;8)洗板机洗5次。9)每孔加入100μl的tmb单组份显色液,显色6min,室温避光,之后每孔加入50μl终止液终止反应;10)用酶标仪上检测450-630nm处的od值,保存记录原始数据。结果:将25株单抗进行筛选,并对埃博拉gp的结合抗体进行特异性鉴定。有10株与埃博拉gp能够特异结合,包括2a1、2g1、2f4、2e5、2g5、2c6、2a7、2c7、2h7、2a10。图4显示了25株单抗的结合活性对照柱图。7.elisa筛选具有广谱结合活性的抗体为了获得具有广谱结合多种埃博拉病毒的抗体,对结合抗体的广谱性进行分析。实验前一天96孔酶联板,包被1μg/ml的ebovgp、bdbvgp、sudvgp和restvgp。其余方法同6。检测结果如图5所示,10株ebovgp结合抗体中,7株抗体具有广谱结合活性,能与两种或两种以上的埃博拉病毒gp结合。2a1、2g5、2c6能与ebovgp和bdbvgp结合;2g1能与ebovgp、bdbvgp、sudvgp和restvgp结合;2f4能与ebovgp、bdbvgp和sudvgp结合;2e5、2a7能与ebovgp和restvgp结合;2c7、2h7和2a10仅能与ebovgp结合,不具有广谱性。其中,2g1具有最好的广谱性,能与我们有的4种埃博拉病毒gp结合,且结合活性良好。8.2g1抗体序列说明2g1具有良好的广谱结合活性,其序列描述如下:轻链可变区的氨基酸序列如seqidno.1所示,轻链可变区的cdr1、cdr2和cdr3的氨基酸序列分别如seqidno.1的第27-32、50-52和89-96位氨基酸序列所示,轻链可变区的基因编码序列由seqidno:2所示,轻链恒定区的氨基酸序列如seqidno.3所示,轻链恒定区的基因编码序列由seqidno:4所示。重链可变区的氨基酸序列如seqidno.5所示,重链可变区的cdr1、cdr2和cdr3的氨基酸序列分别如seqidno.5的第26-33、51-58和97-113位氨基酸序列所示,重链可变区的基因编码序列由seqidno.6重链恒定的氨基酸序列如seqidno.7所示,所述抗体的重链恒定区的基因编码序列由seqidno.8所示。9.表达质粒构建与抗体制备对2g1构建表达质粒,进行单抗的表达制备。方法如下:1)将2g1h和2g1k线性表达框全长基因用ecori(neb,r3101)和noti(neb,r3189)双酶切,连接至pcdna3.4表达质粒。2)取pcdna3.4-2g1h和pcdna3.4-2g1k各15μg,转染至30mlexpi293体系(life,a14524)中,125rpm,5%co2培养72h。3)3000×g,离心10min收取表达上清,经0.22μm针头滤器抽滤后,采用rproteina亲和纯化。4)用pbs对收集的抗体进行换液,然后用bca蛋白定量试剂盒(thermoscientific,23225)测定抗体浓度。实施例2.elisa检测抗体结合活性1)实验前一天96孔酶联板,包被1μg/ml的ebovgp、bdbvgp、sudvgp和restvgp,每孔100μl包被。将包被的酶联板放入湿盒,4℃过夜。2)实验当天用洗板机洗5次。3)每孔加入100μl封闭液,室温下放置1小时。4)洗板机洗5次。5)首孔加入150μl浓度为10μg/ml的2g1单抗,其余孔加入100μl的稀释液。从首孔吸出50μl加入到次孔,以此类推,按1:3梯度稀释每孔终体积为100μl。室温静置1小时。6)洗板机洗5次。7)将hpr标记的羊抗人igg二抗以1:10000用稀释液进行稀释,每孔100μl加入到elisa板对应孔中,室温孵育1小时。8)洗板机洗5次。9)每孔加入100μl的tmb单组份显色液,显色6min,室温避光,之后每孔加入50μl终止液终止反应;10)用酶标仪上检测450-630nm处的od值,保存记录原始数据。结果:2g1单抗与ebovgp、bdbvgp、sudvgp和restvgp具有良好的结合活性,ec50值分别为0.0059μg/ml、0.0075μg/ml、0.0048μg/ml和0.026μg/ml(检测结果参见图6)。实施例3.假病毒中和实验评价2g1中和活性包装hiv骨架的ebov、bdbv和sudv假病毒,在体外评价2g1的中和活性。评价方法如下:1)用dmem培养基稀释2g1单抗,96孔细胞培养板首孔加入75μl浓度为100μg/ml的抗体稀释液,其余孔加入50μl的dmem培养基。2)从首孔吸取25μl液体加入次孔,混匀,以此类推,按1:3倍比稀释,每孔终体积为50μl。3)将假病毒按1:5用dmem培养基稀释(使对照孔荧光素读值控制在20000~100000之间),加入至各抗体孔,每孔50μl。混匀,37℃孵育1h。4)293t细胞计数,2×105cells/ml,每孔加入100μl。5)将96孔细胞培养板放入37℃恒温箱中培养36~48小时6)取出细胞培养板,小心吸出培养液弃掉。各孔加入100μl细胞裂解液,震荡机上350rpm震荡15min。7)3000rpm室温离心10min。8)将荧光素酶检测系统(promega,e1501)的检测底物冻干剂和检测缓冲液混匀后,充盈glomax96microplateluminometer(promega)检测管路。9)吸取20μl裂解上清读值,读取荧光值,计算抗体对细胞的保护率。结果:与对照抗体相比,2g1在体外可有效地中和ebov、bdbv和sudv假病毒。2g1中和活性随着抗体浓度的升高而增强,在1μg/ml的浓度下即可对三种埃博拉假病毒实现近100%的保护。(检测结果参见图7)。实施例4.截短型抗原分析2g1结合区域2g1呈现出良好的广谱中和活性,希望通过构建截短型gp来分析其大致的结合区域。除了文献中常见的gpdtm、gpdm、gp1、sgp和gpcl外,根据gp结构特点和分析目标,还设计构建了gp33-310;463-558、gp33-227、gp33-158、gp95-295、gp158-295和gp227-295。gp33-310;463-558是在gpdm的基础上去掉了后面的七肽重复区;gp33-227是在sgp的基础上去掉了聚糖帽;gp33-158包含了大部分的base区和head区;gp95-295相比sgp缺失了base区;gp158-295相比sgp基本缺失了base区和head区;gp227-295包含了大部分的聚糖帽区。gpcl是在gpdm的基础上,经嗜热菌蛋白酶(thermolysin)切除聚糖帽得到的,不需构建表达质粒。各截短型gp的结构示意图如图8所示。截短型gp分析2g1结合区域方法如下:1)根据示意图,从gp全长基因中扩增各截短型gp基因片段,羧基端添加his6标签,将基因片段连接至pcdna3.4表达质粒中。2)各截短型gp在expi293系统中表达,ni-nta纯化获得目的蛋白。3)elisa分析2g1对各截短型gp的结合活性,方法同实施例2所述。结果:2g1与gpdm结合ec50值为0.0037μg/ml,与gpdtm基本一致。2g1与gp1的结合ec50值为0.8075μg/ml,相比gpdtm和gpdm而言,结合能力降低了100多倍,提示gp2亚基对2g1的结合起着关键作用。尽管2g1与gpcl的结合最大值降低为其与gpdtm结合最大值的一半左右,但其结合ec50值为0.019μg/ml,结合活性较好。gpcl包含了gp2亚基(502-676位氨基酸)和部分的gp1亚基,而sgp包含了gpcl在gp1亚基上的部分,2g1与sgp基本不结合,进一步说明gp2对2g1结合的重要性。2g1与各截短型抗原的结合活性检测见图9-图10。实施例5.竞争实验采用竞争结合elisa分析2g1的抗原作用表位。用已知结合表位的mil77-1/2/3和其余几株非中和抗体作为对照抗体,通过考察检测抗体与gp的结合是否会被同孵育的竞争抗体所阻断,来反映这些抗体的结合表位是否存在交叠。方法如下:1)称取生物素(thermoscientific,21335)4mg,溶于2ml超纯水中,浓度为2mg/ml。2)取抗体2g1、2e5、5d7、5g11、5b12、1b3、mil77-1、mil77-2、mil77-3各200μg,体积控制在200μl左右。3)以生物素比抗体为20:1的摩尔比标记抗体。将抗体和生物素混合,室温孵育1小时,用50kd的0.5ml离心过滤器换液,每次更换pbs约400μl,换液3次以上。4)最后一次用pbs将过滤器中剩余的液体统一补齐至100μl左右(目的是使各抗体的浓度相近),测定抗体浓度。5)96孔酶联板以1μg/ml的浓度包被gpdm,4℃包被过夜。6)洗板机洗涤elisa板,每孔加入100μl封闭液,37℃孵育1小时。7)实验中检测抗体为生物素标记的抗体,竞争抗体为非生物素标记抗体。稀释液稀释竞争抗体至2.5μg/ml;稀释液稀释检测抗体至25ng/ml。8)洗涤96孔板。各孔加入50μl的检测抗体和50μl的竞争抗体,每孔终体积100μl。9)洗涤,链霉亲和素抗体(thermoscitntific,21126)以1:1000稀释,每孔加入100μl,37℃孵育1小时。10)洗涤,每孔加入100μltmb单组份显色液,避光显色6分钟,加入50μl终止液。11)读取450-630nmod值。结果:竞争结合值小于30认为强竞争;大于30小于60认为弱竞争;大于60认为不竞争。各抗体自身之间很好地竞争结合,根据结果可将这些抗体分成四组。其中,结合于gp1亚基和结合于gp2亚基各两组。2g1与同结合于gp2亚基的非中和抗体5b12、中和抗体mil77-1和mil77-2也基本不存在竞争。竞争结合结果说明2g1的结合表位不同于这些对照抗体,提示2g1具有与其他中和抗体组成鸡尾酒组合疗法的潜力(检测结果参见图11)。实施例6.低ph下2g1与gpdm的结合活性检测抗体与病毒颗粒表面的gp结合后,与病毒颗粒一起进入宿主细胞。在次级内体的酸性(ph约5.5)环境中,gp被酶切水解,进而与受体结合,促进病毒基因组的释放。为进一步考察在2g1与gp结合活性是否受酸性环境的影响,检测了体外在ph7.5、ph6.5、ph5.5和ph4.5的环境中2g1与gp的结合活性,elisa方法同实施例2所述。结果:在四种不同ph的环境中,2g1与gpdm的结合活性基本无变化。2g1与gp的结合稳定,不受次级内体低ph环境的影响,为其发挥中和作用提供了基础(检测结果参见图12)。序列表<110>中国人民解放军军事科学院军事医学研究院<120>一种抗埃博拉病毒糖蛋白gp2亚基的单克隆抗体2g1及应用<160>8<170>siposequencelisting1.0<210>1<211>106<212>prt<213>homosapiens<400>1aspileglnmetthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargthrserglnserileserasntyr202530leuasntrptyrglnglnileproglylysalaprolysleuleuile354045serthralaserasnleuhisserglyvalserserargphesergly505560serglyserglythrhisphethrleuthrileserserleuglnpro65707580gluaspphealathrtyrtyrcysglnglnsertyrserthrproser859095pheglyglnglythrlysvalgluilelys100105<210>2<211>318<212>dna<213>homosapiens<400>2gacatccagatgacccagtctccatcctccctgtctgcatctgtaggagacagagtcacc60atcacttgccggacaagtcagagcattagcaactatttaaattggtatcagcagatacca120gggaaagcccctaaactcctgatctctactgcatccaatttgcacagtggggtctcatca180aggttcagtggcagtggatctgggacacatttcactctcaccatcagcagtctgcaacct240gaagattttgcaacttactactgtcaacagagttacagtaccccctcgttcggccaaggg300accaaggtggagatcaaa318<210>3<211>107<212>prt<213>homosapiens<400>3argthrvalalaalaproservalpheilepheproproseraspglu151015glnleulysserglythralaservalvalcysleuleuasnasnphe202530tyrproargglualalysvalglntrplysvalaspasnalaleugln354045serglyasnserglngluservalthrgluglnaspserlysaspser505560thrtyrserleuserserthrleuthrleuserlysalaasptyrglu65707580lyshislysvaltyralacysgluvalthrhisglnglyleuserser859095provalthrlysserpheasnargglyglucys100105<210>4<211>321<212>dna<213>homosapiens<400>4cgtactgtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatct60ggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacag120tggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggac180agcaaggacagcacctacagcctcagcagtaccctgacgctgagcaaagcagactacgag240aaacacaaagtctacgcctgcgaagtcacccatcagggcctgagttcgcccgtcacaaag300agcttcaacaggggagagtgt321<210>5<211>124<212>prt<213>homosapiens<400>5gluvalglnleuvalgluserglyglyglyleuvallysargglygly151015serleuargleusercysvalvalserglytyrthrpheglyglytyr202530sermethistrpvalargglnalaproglylysglyleuglutrpval354045serglyilesersersersertyrtyrlystyrtyralaaspserval505560lysglyargphethrileserargaspasnalalysasnserleutyr65707580leuglnmetasnserleuargalagluaspthralavaltyrtyrcys859095alaargaspmetglytyrcysserglyglysercysproasnpheasp100105110phetrpglyglnglythrthrvalthrvalserser115120<210>6<211>372<212>dna<213>homosapiens<400>6gaggtgcagctggtggagtctgggggaggcctggtcaagcggggggggtccctgagactc60tcctgtgtagtctccggatacaccttcggtggatatagcatgcactgggtccgccaggct120ccagggaaggggctggagtgggtctcaggcattagtagtagtagttattacaaatactac180gcagactcagtgaagggccgattcaccatctccagagacaacgccaagaactcactgtat240ctgcaaatgaatagcctgagagccgaggacacggctgtttattactgtgcgagagatatg300ggatattgtagtggtggtagctgccctaactttgacttctggggccagggaaccacggtc360accgtctcctca372<210>7<211>330<212>prt<213>homosapiens<400>7alaserthrlysglyproservalpheproleualaproserserlys151015serthrserglyglythralaalaleuglycysleuvallysasptyr202530pheprogluprovalthrvalsertrpasnserglyalaleuthrser354045glyvalhisthrpheproalavalleuglnserserglyleutyrser505560leuserservalvalthrvalproserserserleuglythrglnthr65707580tyrilecysasnvalasnhislysproserasnthrlysvalasplys859095lysvalgluprolyssercysasplysthrhisthrcysproprocys100105110proalaprogluleuleuglyglyproservalpheleuphepropro115120125lysprolysaspthrleumetileserargthrprogluvalthrcys130135140valvalvalaspvalserhisgluaspprogluvallyspheasntrp145150155160tyrvalaspglyvalgluvalhisasnalalysthrlysproargglu165170175gluglntyrasnserthrtyrargvalvalservalleuthrvalpro180185190hisglnasptrpleuasnglylysglutyrlyscyslysvalserasn195200205lysalaleuproalaproileglulysthrileserlysalalysgly210215220glnproarggluproglnvaltyrthrleupropropheargaspglu225230235240leuthrlysasnglnvalserleuthrcysleuvallysglyphetyr245250255proseraspilealavalglutrpgluserasnglyglnprogluasn260265270asntyrlysthrthrproprovalleuaspseraspglyserphephe275280285leutyrserlysleuthrvalasplysserargtrpglnglnglyasn290295300valphesercysservalmethisglualaleuhisasnhistyrthr305310315320glnlysserleuserleuserproglylys325330<210>8<211>990<212>dna<213>homosapiens<400>8gcgtcgaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctggg60ggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcg120tggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctca180ggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacc240tacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagccc300aaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctgggggga360ccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccct420gaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactgg480tacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaac540agcacgtaccgtgtggtcagcgtcctcaccgtcccgcaccaggactggctgaatggcaaa600gagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctcc660aaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccattccgggatgag720ctgaccaagaaccaggtcagcctgacctgcctggtcaagggcttctatcccagcgacatc780gccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtg840ctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtgg900cagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacg960cagaagagcctctccctgtctccgggtaaa990当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1