腺相关病毒变体衣壳和用于抑制血管生成的应用的制作方法
相关申请的交叉引用
本申请要求2017年11月27日提交的美国临时专利申请序列号62/590,976、2018年4月30日提交的62/664,726的权益,其每一个的全部公开内容通过引用并入本文。
通过引用并入作为文本文件提供的序列表
序列表在本文中作为于2018年11月26日创建并且大小为238kb的文本文件“090400-5010-wo-seq-listing.txt”提供。文本文件的内容通过引用以其全文并入本文。
本文公开的发明总体上涉及包含变体衣壳蛋白的腺相关病毒(aav)病毒粒子领域和使用定向进化技术产生这种变体衣壳。
背景技术:
遗传性视网膜疾病涵盖大量异质性遗传疾病,其影响约3000分之1的人(全世界超过2百万人),并且是严重视力丧失或失明的主要来源。复杂的多因素视网膜疾病如湿性年龄相关性黄斑变性(wamd)和糖尿病性视网膜病变(dr)影响甚至更多的个体,目前有170万美国人患有与wamd相关的严重中央视力丧失,并且患有糖尿病的年龄超过40岁的近三分之一成年人具有视力障碍。这些疾病通常与一种或多种类型的视网膜细胞的功能障碍或死亡相关联,在某些情况下归因于缺乏关键蛋白的表达或功能,例如lca2中的rpe65,在其它情况下归因于产生毒性基因产物的基因突变,例如影响视紫红质蛋白折叠的显性突变,或者在又其它情况下归因于由蛋白质的异位表达诱导的视网膜生理学变化,例如wamd中的vegf
解决这种巨大的未满足医学需求的一种方法是基于基因的腺相关病毒(aav)介导疗法,其中使用重组腺相关病毒(raav)将基因递送到视网膜中的一种或多种类型的细胞以例如替换缺失的基因、纠正显性缺陷基因、或提供连续蛋白质疗法的模板。虽然基于aav的临床基因疗法已经越来越成功,但是在病毒载体性质方面仍然存在缺点,包括例如以高效率靶向期望的视网膜细胞。例如,已经识别并表征了多种同源灵长类动物aav血清型和许多非人灵长类血清型,其中aav2在aav血清型中是最佳表征的并且是第一种适合作为眼中的基因递送媒剂的。然而,尚未报道这些aav(包括aav2)在通过玻璃体内施用递送时有效转导视网膜的较深细胞类型。因此,本领域需要具有优异转导能力的新aav变体,其将为视网膜细胞提供更有效的基于基因的递送以治疗眼病。本领域需要这种aav变体,所述aav变体显示出增强的视网膜转导谱-与本领域已知的野生型aav和aav变体相比,在某些情况下,所述aav变体广泛地用于某些视网膜细胞类型,在其它情况下,所述aav变体优先用于某些视网膜细胞类型。
天然存在的aav是单链dna病毒,其含有三个开放阅读框,rep、cap和aap。第一个基因rep编码对基因组复制而言必需的四种蛋白质(rep78、rep68、rep52和是rep40),第二个基因cap表达组装形成病毒衣壳的三种结构蛋白(vp1-3),并且第三个基因表达对衣壳组装至关重要的组装激活蛋白(aap)。aav依赖于辅助病毒(如腺病毒或疱疹病毒)的存在进行活性复制。在没有辅助病毒的情况下,aav建立潜伏状态,其中其基因组以游离方式维持或整合到aavs1基因座中的宿主染色体中。
体外和体内定向进化技术可以用于选择aav变体,所述aav变体提供对当前基于aav的基因递送载体的改进。这种定向进化技术是本领域已知的并且描述于例如pct公开wo2014/194132和kotterman&schaffer(naturereviewgenetics,aop,2014年5月20日在线出版;doi:10.1038/nrg3742),这两者均通过引用以其全部并入本文。定向进化是一种衣壳工程化方法,其通过迭代轮次的遗传多样化和选择过程模拟自然进化,从而能够积累逐步改善生物分子的功能如基于aav的病毒粒子的有益的突变。在此方法中,使野生型aavcap基因多样化以产生大的遗传文库,其被包装以产生病毒颗粒文库,并且应用选择性压力以分离具有可以克服基因递送障碍的优良表型的独特变体。
以下中已经公开了aav变体:例如美国专利号9,193,956;9;186;419;8,632,764;8,663,624;8,927,514;8,628,966;8,263,396;8,734,809;8,889,641;8,632,764;8,691,948;8,299,295;8,802,440;8,445,267;8,906,307;8,574,583;8,067,015;7,588,772;7,867,484;8,163,543;8,283,151;8,999,678;7,892,809;7,906,111;7,259,151;7,629,322;7,220,577;8,802,080;7,198,951;8,318,480;8,962,332;7,790,449;7,282,199;8,906,675;8,524,446;7,712,893;6,491,907;8,637,255;7,186,522;7,105,345;6,759,237;6,984,517;6,962,815;7,749,492;7,259,151;和6,156,303;美国公开号2013/0295614;2015/0065562;2014/0364338;2013/0323226;2014/0359799;2013/0059732;2014/0037585;2014/0056854;2013/0296409;2014/03350542013/0195801;2012/0070899;2011/0275529;2011/0171262;2009/0215879;2010/0297177;2010/0203083;2009/0317417;2009/0202490;2012/0220492;2006/0292117;和2004/0002159;欧洲公开号2692731a1;2383346b1;2359865b1;2359866b1;2359867b1;和2357010b1;1791858b1;1668143b1;1660678b1;1664314b1;1496944b1;1456383b1;2341068b1;2338900b1;1456419b1;1310571b1;1456383b1;1633772b1;和1135468b1;和国际(pct)公开号wo2014/124282;wo2013/170078;wo2014/160092;wo2014/103957;wo2014/052789;wo2013/174760;wo2013/123503;wo2011/038187;和wo2008/124015;wo2003/054197;然而,这些参考文献没有公开本文所描述的aav变体的实施方式和/或特征和/或物质结构的组合物。
本文引用的和引用的专利文献中引用的所有文献和参考文献均通过引用结合到本文中。
技术实现要素:
本文提供了变体腺相关病毒(aav)衣壳蛋白,其相对于亲本aav衣壳蛋白具有在氨基酸序列上的一个或多个修饰,当存在于aav病毒粒子中时,与包含未修饰的亲本aav衣壳蛋白的aav病毒粒子对视网膜细胞的感染性相比,所述变体aav衣壳蛋白赋予对一种或多种类型的视网膜细胞增加的感染性。还提供了重组aav病毒粒子和其包含本文所描述的变体aav衣壳蛋白的药物组合物,制造raav衣壳蛋白和病毒粒子的方法,以及在研究中和在临床实践中,例如在例如将核酸序列递送至视网膜的一个或多个细胞以治疗视网膜病症和疾病中使用这些raav衣壳蛋白和病毒粒子的方法。
在本公开的一些方面,提供了变体腺相关病毒(aav)衣壳蛋白,这些变体aav衣壳蛋白相对于亲本aav衣壳具有在氨基酸序列上的一个或多个修饰,当存在于aav病毒粒子中时,所述变体aav衣壳蛋白赋予对一种或多种类型的视网膜细胞(例如,感光细胞(例如视杆;视锥)、视网膜神经节细胞(rgc)、神经胶质细胞(例如穆勒胶质细胞、小胶质细胞)、双极细胞、无长突细胞、水平细胞和/或视网膜色素上皮(rpe)细胞)与包括亲本aav衣壳蛋白的aav病毒粒子对所述视网膜细胞的感染性相比增加的感染性,所述亲本aav衣壳蛋白不包括氨基酸序列修饰。
在本公开的一些方面,提供了重组aav(raav)病毒粒子,这些raav病毒粒子包括如本文所述的变体衣壳蛋白,其中所述raav病毒粒子表现出对一种或多种类型的视网膜细胞(例如感光细胞)(例如,视杆;视锥)、视网膜神经节细胞(rgc)、神经胶质细胞(例如穆勒胶质细胞、小胶质细胞)、双极细胞、无长突细胞、水平细胞和/或视网膜色素上皮(rpe)细胞)相对于包括相应未修饰的亲本aav衣壳蛋白的aav病毒粒子对所述视网膜细胞的感染性的增加的感染性。在一些实施方式中,所述raav病毒粒子相对于包含亲本aav衣壳蛋白的aav病毒粒子,表现出对所有视网膜细胞的增加的感染性。在其它实施方式中,所述raav病毒粒子相对于包含亲本aav衣壳蛋白的aav病毒粒子,表现出对视网膜的某些细胞类型而不是的其它细胞类型的增加的感染性。换句话说,所述raav病毒粒子表现出对视网膜的某些细胞类型而不是对其它细胞类型优选增加的感染性,例如raav表现出优选地对选自感光细胞、视网膜神经节细胞、神经胶质细胞、双极细胞、无长突细胞、水平细胞和/或视网膜色素上皮(rpe)细胞中的一种或多种细胞类型的增加的感染性,但未表现出对所有细胞类型的增加的感染性。
在一些实施方式中,raav病毒粒子包括异源核酸。在一些此类实施方式中,异源核酸对编码多肽的rna进行编码。在其它此类实施方式中,异源核酸序列对不编码多肽的rna进行编码,例如异源核酸序列编码针对核酸酶的rna干扰剂、指导rna等。
本文还提供了包括主题感染性raav病毒粒子和药学上可接受的载剂的药物组合物。
还提供了包含本文所述的变体衣壳蛋白的raav病毒粒子在通过使靶细胞与raav病毒粒子接触而将异源核酸递送到靶细胞(例如视网膜细胞)的方法中的用途。在一些实施方式中,靶细胞在体内,例如在需要治疗眼部疾病的个体的眼睛中。在其它实施方式中,靶细胞在体外。
还提供了通过向需要这种治疗的受试者施用有效量的raav病毒粒子来治疗眼病的方法,所述病毒粒子包含如本文所述的变体衣壳蛋白或包含有效量的raav病毒粒子的药物组合物。
还提供了一种分离的核酸和包含所述分离的核酸的宿主细胞,所述分离的核酸包含对如本文所述的变体aav衣壳蛋白进行编码的序列。在又其它实施方式中,分离的核酸和/或分离的宿主细胞包含raav。
在一些方面,变体aav衣壳蛋白包含在相对于相应的亲本aav衣壳蛋白的所述衣壳蛋白的gh-环中约5个氨基酸至约20个氨基酸的插入(“异源肽”或“肽插入”),其中当存在于aav病毒粒子中时,与包括相应亲本aav衣壳蛋白的aav病毒粒子对视网膜细胞的感染性相比,所述变体衣壳蛋白赋予对视网膜细胞增加的感染性。在一些实施方式中,所述肽包括选自由以下组成的组的序列:qadttkn(seqidno:13)、isdqtkh(seqidno:14)、asdstka(seqidno:15)、nqdytkt(seqidno:16)、hditkni(seqidno:17)、hpdttkn(seqidno:18)、hqdttkn(seqidno:19)、nkttnkd(seqidno:20)、isneneh(seqidno:21)、qananen(seqidno:22)、gkskvid(seqidno:23)、tnrtspd(seqidno:24)、pnsthgs(seqidno:25)、kdrapst(seqidno:26)、laqadttkna(seqidno:27)、laisdqtkha(seqidno:28)、lgisdqtkha(seqidno:29)、laasdstkaa(seqidno:30)、lanqdytkta(seqidno:31)、lahditknia(seqidno:32)、lahpdttkna(seqidno:33)、lahqdttkna(seqidno:34)、lankttnkda(seqidno:35)、lpisneneha(seqidno:36)、lpqananena(seqidno:37)、lagkskvida(seqidno:38)、latnrtspda(seqidno:39)、lapnsthgsa(seqidno:40)和lakdrapsta(seqidno:41)。在一些实施方式中,所述肽基本上由选自由以下组成的组的序列组成:qadttkn(seqidno:13)、isdqtkh(seqidno:14)、asdstka(seqidno:15)、nqdytkt(seqidno:16)、hditkni(seqidno:17)、hpdttkn(seqidno:18)、hqdttkn(seqidno:19)、nkttnkd(seqidno:20)、isneneh(seqidno:21)、qananen(seqidno:22)、gkskvid(seqidno:23)、tnrtspd(seqidno:24)、pnsthgs(seqidno:25)、kdrapst(seqidno:26)、laqadttkna(seqidno:27)、laisdqtkha(seqidno:28)、lgisdqtkha(seqidno:29)、laasdstkaa(seqidno:30)、lanqdytkta(seqidno:31)、lahditknia(seqidno:32)、lahpdttkna(seqidno:33)、lahqdttkna(seqidno:34)、lankttnkda(seqidno:35)、lpisneneha(seqidno:36)、lpqananena(seqidno:37)、lagkskvida(seqidno:38)、latnrtspda(seqidno:39)、lapnsthgsa(seqidno:40)和lakdrapsta(seqidno:41)。在一些方面,变体aav衣壳蛋白包含相对于相应的亲本aav衣壳蛋白的一个或多个氨基酸取代,其中当存在于aav病毒粒子中时,与包含相应亲本aav衣壳蛋白的aav病毒粒子对视网膜细胞的感染性相比,所述变体衣壳蛋白赋予对视网膜细胞增加的感染性。
在相关方面,变体aav衣壳蛋白包含相对于相应的亲本aav衣壳蛋白的肽插入和一个或多个氨基酸取代,其中当存在于aav病毒粒子中时,与包含相应亲本aav衣壳蛋白的aav病毒粒子对视网膜细胞的感染性相比,所述变体衣壳蛋白赋予对视网膜细胞增加的感染性。
本文还公开了变体aav衣壳蛋白,其包含异源肽laisdqtkha(seqidno:28)和相对于aav2的p34a取代。在相关实施方式中,本文公开了感染性重组aav,其包含与seqidno:42所示序列具有至少90%同一性的变体aav衣壳蛋白和包含编码vegf抑制剂(优选vegfa抑制剂)的序列的异源核酸,以及公开了包含所述感染性重组aav的药物组合物。在一些实施方式中,编码vegf抑制剂的异源核酸序列选自编码阿柏西普、雷珠单抗、单链形式的雷珠单抗(sc-雷珠单抗lh或hl)、溴珠单抗与iggfc结构域融合的sc-雷珠单抗(sc-雷珠单抗-fc)或与iggfc结构域融合的溴珠单抗(溴珠单抗-fc)的序列。在相关实施方式中,重组aav包含含有两个以上序列的异源核酸,所述序列中的每一个编码vegfa抑制剂(例如编码阿柏西普的第一序列和编码溴珠单抗的第二序列)。在优选的实施方式中,异源核酸序列具有seqidno:65、67、69、70、72、74、76中的任一个序列或与其具有至少90%同一性的序列。在其它相关实施方式中,提供了治疗患有与眼内vegfa水平升高相关的眼病的患者的方法,其包括优选通过玻璃体内注射向患者施用有效量的感染性重组aav和异源核酸,所述感染性重组aav包含与seqidno:42所示序列具有至少90%同一性的变体aav衣壳蛋白,所述异源核酸包含编码vegf抑制剂的序列。
本文还公开了变体aav衣壳蛋白,其包含异源肽laisdqtkha(seqidno:28)和相对于aav2的氨基酸取代n312k、n449d、n551s、i698v和l735q。
本文还公开了用于制造和/或递送包含如本文中公开的变体aav衣壳的raav的方法。另外,本文提供了包含raav的试剂盒和用于本文所述的方法的用途,所述raav包含如本文公开的变体aav衣壳。
在其它实施方式中,包含前述段落中的变体衣壳蛋白的aav病毒粒子可以并入任何前述或随后公开的实施方式。实际上,应理解的是,为清楚起见在单独的实施方式的上下文中描述的本发明的某些特征也可以在单个实施方式中组合提供。相反,为简洁起见而在单个实施方式的背景下描述的本发明的不同特征也可以单独地或以任何适合的子组合形式提供。本发明具体涵盖与本发明有关的实施方式的所有组合,并且在本文中公开,就好像每个和每个组合被单独和明确地公开一样。此外,本发明还具体涵盖了各种实施方式和其元件的所有子组合,并且在本文中公开,就好像每个子组合和每个这种子组合在本文中单独和明确地公开一样。
发明内容并非旨在限定权利要求,也不旨在以任何方式限制本发明的范围。
根据以下附图、具体实施方式和权利要求,本文公开的本发明的其它特征和优点将显而易见。
在描述本发明的方法和组合物之前,应当理解,本发明不限于所述的特定方法或组合物,因此可以变化。还应理解,本文所用的术语仅用于描述特定实施方式的目的,而不是旨在限制,因为本发明的范围仅由所附权利要求书限制。
在附图和说明书中说明了本文公开的发明。然而,尽管在附图中示出了特定实施方式,但是不旨在将本发明限制于所示出和/或公开的一个或多个特定实施方式。相反,本文公开的发明旨在覆盖落入本发明的精神和范围内的所有修改、替代构造和等同物。因此,附图旨在是说明性的而非限制性的。
在提供数值范围的情况下,应当理解,除非上下文另外明确指出,否则也具体公开了该范围的上限和下限之间的每个中间值,至下限单位的十分之一。在所述范围内的任何所述值或中间值与该所述范围内的任何其它所述值或中间值之间的每个较小范围都包括在本发明内。这些较小范围的上限和下限可以独立地包括在该范围内或排除在该范围外,并且其中任一限值、两个限值都不包括在较小范围内的每个范围也包括在本发明内,服从在所述范围内的任何具体排除的限值。当所述范围包括一个或两个极限时,排除那些包括的极限中的一个或两个的范围也包括在本发明中。
除非另有定义,否则本文所用的所有科学技术术语具有与本发明所属领域的普通技术人员通常理解的相同的含义。尽管与本文所述的那些类似或等同的任何方法和材料可用于本发明的实践或测试,但现在描述一些可能的和优选的方法和材料。本文提及的所有出版物均通过引用并入本文以公开和描述与所引用的出版物相关的方法和/或材料。应当理解,本公开在存在矛盾的程度上取代了所并入的出版物的任何公开内容。
本领域技术人员在阅读本公开后将明白,本文描述和说明的单独实施方式具有离散的组件和特征,其可容易地与其它几个实施方式中的任一个的特征分开或组合,而不脱离本发明的范围或精神。任何列举的方法可以以列举的事件的顺序或以逻辑上可能的任何其它顺序来执行。
应注意,如本文和所附权利要求中所用,单数形式“一”、“一个”和“该”包括复数指代物,除非上下文另外明确指出。因此,例如,提及“重组aav病毒粒子”包括多个这样的病毒粒子,提及“感光细胞”包括提及一个或多个感光细胞及其本领域技术人员已知的等同物,等等。还应注意,权利要求书可被撰写为排除任何可选元素。因此,该陈述旨在用作使用与权利要求要素的叙述有关的诸如“单独地”、“仅”等的排他性术语或使用“否定型”限定的先行基础。
本文所讨论的出版物仅提供其在本申请的申请日之前的公开内容。本文不应被解释为承认本发明无权凭借在先发明先于这些出版物。此外,所提供的出版日期可能与实际出版日期不同,实际出版日期可能需要独立确认。
附图说明
当结合附图阅读时,从以下详细描述中可以最好地理解本发明。本专利或申请文件含有至少一幅彩色附图。在请求并支付必要的费用后,官方将会提供带有一幅或多幅彩色附图的本专利或专利申请公开物的副本。需要强调的是,根据惯例,附图的各种特征不是按比例的。相反,为了清楚起见不同特征的尺寸被任意地扩大或减少。附图中包含以下图:
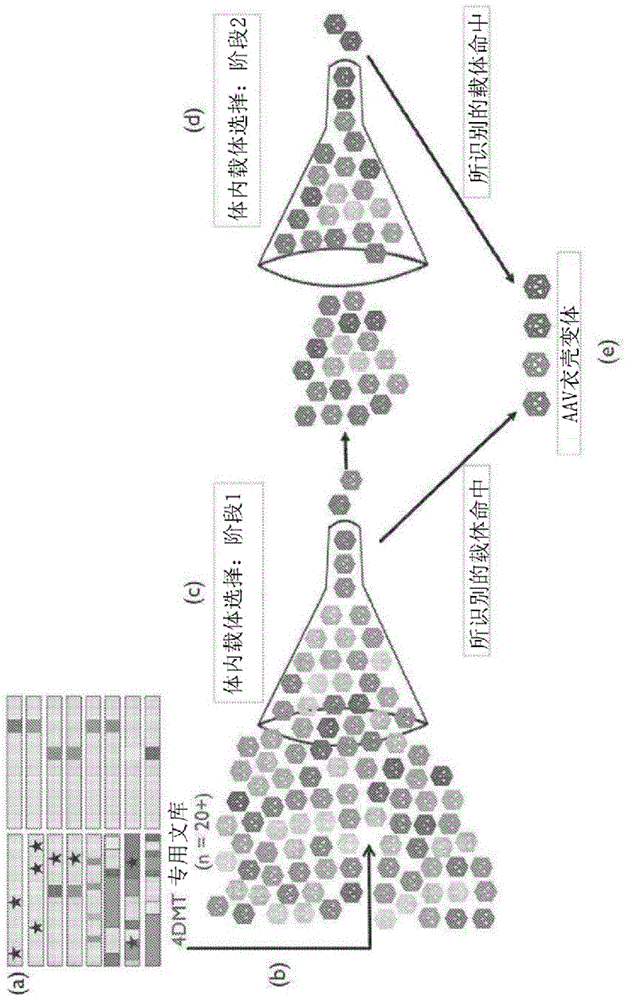
图1描绘了定向进化方法论的实施方式。步骤(a)描绘了包括dna突变技术和cap基因的组合的病毒衣壳文库的产生。步骤(b)描绘了病毒的包装,使得每个病毒颗粒由围绕编码该衣壳和纯化的cap基因的突变衣壳组成。然后将衣壳文库置于体外或体内选择压力下。在定向进化技术的这个方面,收获感兴趣的组织或细胞材料以分离已成功感染该靶标的aav变体,并回收成功的病毒。步骤(c)描绘了通过重复选择的成功克隆的阶段1富集。步骤(d)描绘了选择的cap基因的阶段2富集,所选择的cap基因经历再多样化(re-diversification)和进一步选择步骤以迭代地增加病毒拟合性。步骤(e)描绘了在载体选择阶段1和2期间识别为命中的变体,其将作为重组aav载体制造并表征各种细胞类型和组织靶标的转导水平。根据aav定向进化过程的性质,本文公开的变体已经证明在选择过程期间转导视网膜细胞并递送基因组(对变体cap基因进行编码的基因组)的能力。
图2提供了视网膜平面安装示意图,其显示了在视网膜的广泛区域从何处收集由其扩增病毒基因组的样品。
图3示出了来自代表性选择轮的神经节细胞层(gcl)、内核层(inl)、光感受器/外核层(onl)和视网膜色素上皮(rpe)层视网膜组织的病毒基因组的pcr扩增。右眼(顶部图像)和左眼(底部图像)都注入了文库。对内视网膜(内)、中间视网膜(中)和外/周边视网膜(外)进行取样。红框内的条带代表病毒基因组的成功扩增。
图4a-4d示出了测序分析内的基序的频率。图4a提供了第3轮测序分析。图4b提供了第4轮测序分析。图4c提供了第5轮测序分析。图4d提供了第6轮测序分析。
图5提供了在氨基酸587后含有随机七聚体的aav2的代表性三维模型。
图6a-6w提供了野生型aavseqidnos:1-11的比对,其示出了野生型(天然存在的)血清型aav1、aav2、aav3a、aav3b与aav4-10之间和跨其的氨基酸位置。
图7提供了用在玻璃体内施用2x1011个aav2的载体基因组(vg)后的非洲绿猴视网膜的heidelbergspectralistm拍摄的眼底荧光图像,所述aav2在cmv启动子的控制下递送gfp转基因(aav2.cmv.gfp)。图像是在注射后的基线(a)和第14天(b)、第28天(c)和第42天(d)拍摄的。
图8提供了在用玻璃体内施用2x1011个新型aav变体laisdqtkha+p34a的载体基因组(vg)后的非洲绿猴视网膜的heidelbergspectralistm拍摄的眼底荧光图像,所述aav变体在cmv启动子的控制下递送gfp转基因(laisdqtkha+p34a.cmv.gfp)。图像是在注射后的基线(a)和第14天(b)、第28天(c)和第42天(d)拍摄的。
图9提供了在用玻璃体内施用新型aav变体laisdqtkha+p34a后的非洲绿猴视网膜的heidelbergspectralistm拍摄的眼底荧光图像,所述aav变体在cag启动子的控制下递送gfp转基因(laisdqtkha+p34a.cag.egfp)。(a)用2x1011vg载体玻璃体内注射猴子的视网膜,注射后第14天(a1)、第21天(a2)和第28天(a3)成像。(b)用1x1012vg载体玻璃体内注射猴子的视网膜,注射后第14天(b1)和第21天(b2)成像。
图10a至图10e提供了在注射后三周分析的玻璃体内注射1x1012vg新型aav变体laisdqtkha+p34a的猴视网膜的免疫组织化学分析的结果,所述新型aav变体在cag启动子控制下递送gfp转基因。所有免疫组织化学与相应的眼底荧光图像一起提供,红色框表示大约在视网膜中进行分析的位置。图10a:使用gfp特异性抗体(红色)观察到稳健的视网膜色素上皮(rpe)和光感受器转导。使用m/l视蛋白抗体的视锥光感受器免疫染色示出为白色。图10b和图10c:通过直接egfp荧光(绿色)和通过使用gfp特异性抗体(红色)的免疫组织化学观察到稳健的视杆和视锥光感受器(图10b)和rpe(图10c)转导。rpe中的黑素体在图像中显示为黑色。图10d:通过直接egfp荧光(绿色)和通过使用gfp特异性抗体(红色)的免疫组织化学观察到视锥光感受器(由m/l视蛋白识别,白色)和视网膜神经节细胞(rgc)在中央凹中和周围的转导。中间图中的图像是由左图中白色框表示的区域的更高放大(63倍)。图10e:通过直接egfp荧光观察视网膜神经节细胞(rgc)和视网膜神经节细胞层的转导(右图,绿色;右下图是右上图的63倍放大);左上方示出了明场照明下的区域。
图11a至图11f提供了体外通过重组aav病毒转导人视网膜色素上皮(rpe)细胞的数据,所述重组aav病毒包含在cag启动子控制下的新型aav变体laisdqtkha+p34a衣壳和gfp转基因。用新型aav变体laisdqtkha+p34a.cag.gfp或野生型对照aav2.cag.gfp感染从人胚胎干细胞系(图11a和图11c)或源自人成纤维细胞的诱导多能干细胞(fb-ipsc)分化成rpe细胞的细胞(图11b和11d)。图11a和11b:在moi为500的感染后7天对细胞培养物进行免疫荧光成像表明新型aav变体衣壳(左图)比野生型aav2衣壳更好地转导rpe细胞(右图)。图11c和图11d:通过流式细胞术定量每种培养物中gfp阳性rpe细胞的百分比揭示,与野生型aav2衣壳相比,新型aav变体衣壳提供了转导细胞的数量的显著剂量依赖性改善,而与细胞来源无关。图11e和图11f:通过蛋白质印迹定量每种培养物中gfp的量揭示,与野生型aav2衣壳相比,新型aav变体提供了转基因表达方面的显著改善,而与细胞来源无关。
图12a-12f描述了抗vegf蛋白的实施方式。图12a描述了阿柏西普设计,其由与人igg1的fc区融合的人fltl信号肽、vegfr1结构域2和vegfr2结构域3组成。图12b-c描述了雷珠单抗的设计,其包括柔性的蛋白质接头以将双链抗原结合片段(fab)转化为单链fab(scfab)。图12b描述了轻-重(lh)形式,其由通过柔性肽连接的人igκ轻链信号肽、雷珠单抗的可变轻链、恒定轻链、可变重链和恒定重链1结构域组成。图12c描绘了重-轻(hl)形式,与lh形式相比,其是相似的,除了信号肽源自人igg重链并且重链和轻链结构域在接头的相反侧上。图12d描述了溴珠单抗设计,其包括通过柔性肽连接的可变轻链和可变重链结构域。图12e描述sc-雷珠单抗lh-fc设计,其由与人iggl的fc区融合的scfablh形式组成。图12f描述了溴珠单抗-fc设计,其由与人igg1的fc区融合的溴珠单抗组成。红色条带表示互补决定区(cdr)。基因经密码子优化以改善从人细胞的表达,并通过geneart或genscript合成,并插入到paav-cag-sv40pa载体中cag启动子和sv40polya信号之间。
图13a-b显示了检测与vegf结合的蛋白质的elisa结果。图13a显示在来自转染有阿柏西普(seqidno:65)或单链(sc)雷珠单抗(seqidno:67、69和70)表达质粒的hek293t细胞的培养基中检测到vegf结合活性,但没有在模拟转染或转染有gfp表达载体的细胞中检测到。图13b显示在来自转染有阿柏西普(seqidno:65)、sc-雷珠单抗lh1(seqidno:69)或溴珠单抗(seqidno:74)表达质粒的hek293t细胞的培养基中检测到vegf结合活性,但没有在来自转染有gfp表达载体的细胞中检测到。使用溴珠单抗的信号非常低,最可能是由于检测抗体的识别差。误差棒代表一式四份转染孔的标准偏差。
图14a-b提供了来自用抗vegf构建体转染的hek293t细胞的培养基的代表性蛋白质印迹。图14a显示阿柏西普(seqidno:65)或sc-雷珠单抗(seqidno:67、69和70)表达质粒。将临床eylea和阿柏西普样品都还原成表观分子量为58kd的二聚体的一半(包括糖基化)。临床lucentis被还原成24kd的单独轻链和重链,而sc-雷珠单抗蛋白没有被分离,以约48kd迁移。与hl形式的蛋白质相比,存在更高量的lh形式的蛋白质,这与通过elisa获得的蛋白质定量一致。图14b显示阿柏西普(seqidno:65)、sc-雷珠单抗lh1(seqidno:69)或溴珠单抗(seqidno:74)表达质粒。溴珠单抗的信号低,最可能是由于检测抗体的识别差。蛋白质以26kd的正确分子量迁移。在任何模拟转染或gfp阴性对照样品中都没有信号。
图15a-b显示在与来自用抗vegf构建体转染的hek293t细胞的培养基温育后检测游离vegf的竞争elisa结果。图15a显示阿柏西普(seqidno:65)或sc-雷珠单抗(seqidno:67、69和70)表达质粒。来自转染样品的四种抗vegf蛋白的抑制曲线与临床比较蛋白eylea和lucentis非常相似。阿柏西普和eylea与sc-雷珠单抗变体和lucents相比竞争vegf更强。sc-雷珠单抗的所有三种形式几乎相同。图15b显示阿柏西普(seqidno:65)、sc-雷珠单抗lh1(seqidno:69)或溴珠单抗(seqidno:74)表达质粒。gfp阴性对照样品没有竞争活性。误差棒表示一式两份转染孔的标准偏差。
图16显示细胞vegf中和测定的结果。该测定使用表达vegf受体/β-半乳糖苷酶融合蛋白的hek293细胞,所述融合蛋白在vegf结合时产生活性β-半乳糖苷酶。将细胞与vegf和各种稀释的培养基的混合物一起温育,所述培养基来自转染有阿柏西普(seqidno:65)或sc-雷珠单抗(seqidno:69)表达质粒的hek293t细胞。来自转染样品的抗vegf蛋白的抑制曲线证明,抗vegf蛋白以与临床比较蛋白相等的水平中和vegf活性。误差棒表示一式两份测定孔的标准偏差。
图17a-b显示了用来自转染有抗vegf质粒的hek293t细胞的等体积培养基进行的细胞vegf中和测定的结果。图17a显示gfp、阿柏西普(seqidno:65)、sc-雷珠单抗hl(seqidno:67)或sc-雷珠单抗lh1(seqidno:69)表达质粒。所有抗vegf构建体评估中和的vegf。图17b显示gfp、阿柏西普(seqidno:65)、sc-雷珠单抗lh1(seqidno:69)和溴珠单抗(seqidno:74)表达质粒。所有抗vegf构建体评估中和的vegf。在测定的稀释液中,gfp对照样品有轻微的基质效应。误差棒表示一式两份转染孔的标准偏差。
图18显示检测来自转染有阿柏西普(seqidno:65)或sc-雷珠单抗(seqidnos:67、69和70)表达质粒的rpe细胞的培养基中的vegf的elisa结果。观察到阿柏西普表达和sc-雷珠单抗表达都降低了vegf水平。所有三种sc-雷珠单抗形式对vegf水平的作用是相似的。误差棒表示一式四份转染孔的标准偏差。
图19显示在用表达阿柏西普(seqidno:65)、sc-雷珠单抗(seqidno:67和69)或溴珠单抗(seqidno:74)转基因的r100衣壳(具有seqidno:42所示的氨基酸序列)转导后六或十天,检测从rpe细胞收集的培养基中vegf的elisa结果。用gfp对照载体转导的细胞所显示的vegf内源水平为4,500到8,300pg/ml。用所有抗vegf载体转导在培养基中产生不可检测的vegf水平。误差棒表示一式四份转导孔的标准偏差。
图20显示在用表达抗vegf转基因的r100衣壳(具有seqidno:42所示的氨基酸序列)转导后六或十天,检测从rpe细胞收集的培养基中与vegf结合的蛋白质的elisa结果。在来自用阿柏西普(seqidno:65)、sc-雷珠单抗(seqidno:67和69)或溴珠单抗(seqidno:74)表达载体转导的细胞的培养基中检测到vegf结合活性,但在从用gfp表达载体转导的细胞中未检测到。溴珠单抗的信号非常低,最可能是由于检测抗体的识别差。误差棒表示一式四份转导孔的标准偏差。
图21提供了从用表达抗vegf转基因的r100衣壳(具有seqidno:42所示的氨基酸序列)转导六或十天后,从rpe细胞收集的培养基的代表性蛋白质印迹。将临床eylea和阿柏西普(seqidno:65)样品都还原成表观分子量为60kd的二聚体的一半(包括糖基化),如黑色箭头所示。在gfp阴性对照样品中没有正确迁移的条带。临床lucentis被还原成24kd的单独轻链和重链,而sc-雷珠单抗hl和lh(seqidno:67和69)没有分离,并且以58kd的表观分子量迁移,如灰色箭头所示。溴珠单抗(seqidno:74)的信号低,最可能是由于检测抗体的识别差。蛋白质以26kd的正确分子量迁移,如点画箭头所示。
图22显示在用表达抗vegf转基因的r100衣壳(具有seqidno:42所示的氨基酸序列)转导六或十天后,与从rpe细胞收集的培养基温育后,检测游离vegf的竞争elisa的结果。所有抗vegf构建体竞争vegf。gfp阴性对照样品没有竞争活性。由于rpe细胞产生的内源性vegf,游离vegf水平在最低稀释度时较高。误差棒表示一式两份测定孔的标准偏差。
图23显示了在用表达抗vegf转基因的r100衣壳(具有seqidno:42所示的氨基酸序列)转导六天后,用从rpe细胞收集的培养基进行的细胞vegf中和测定的结果。所有抗vegf构建体评估中和的vegf。用来自gfp对照转导的培养基没有观察到vegf中和。误差棒表示一式两份测定孔的标准偏差。
定义
除非另有定义,否则在此所使用的所有科学和技术术语均具有与本技术所属领域技术人员所通常理解的相同含义。
腺相关病毒是一种非致病性细小病毒,其由无包膜二十面体衣壳内的4.7kb单链dna基因组组成。基因组包含三个侧接有作为病毒复制起点和包装信号的反向末端重复序列(itr)的开放阅读框(orf)。reporf编码四种非结构蛋白,这四种非结构蛋白在病毒复制、转录调节、位点特异性整合和病毒粒子组装中发挥作用。caporf编码三种结构蛋白(vp1-3),这三种结构蛋白组装形成60-mer病毒衣壳。最后,作为cap基因内的替代阅读框存在的orf产生组装激活蛋白(aap),aap是一种病毒蛋白,其将aav衣壳蛋白定位于细胞核并在衣壳组装过程中起作用。
存在若干种天然存在的(“野生型”)血清型和超过100种已知的aav变体,所述变体中的每种变体的氨基酸序列不同,特别是在衣壳蛋白的高变区内以及因此在其基因递送性质方面。没有aav与任何人类疾病相关,使得重组aav对临床应用具有吸引力。
出于本文公开的目的,术语“aav”是腺相关病毒的缩写,包括但不限于病毒本身和其衍生株。除非另有说明,否则术语是指所有亚型或血清型以及复制能力型和重组型。术语“aav”包括但不限于aav1型(aav-1或aav1)、aav2型(aav-2或aav2)、aav3a型(aav-3a或aav3a)、aav3b型(aav-3b或aav3b)、aav4型(aav-4或aav4)、aav5型(aav-5或aav5)、aav6型(aav-6或aav6)、aav7型(aav-7或aav7)、aav型8(aav-8或aav8)、aav9型(aav-9或aav9)、aav10型(aav-10或aav10或aavrh10)、禽aav、牛aav、犬aav、山羊aav、马aav、灵长类动物aav、非灵长类动物aav和绵羊aav。“灵长类动物aav”是指感染灵长类动物的aav,“非灵长类动物aav”是指感染非灵长类哺乳动物的aav,“牛aav”是指感染牛哺乳动物的aav等。
各种血清型aav的基因组序列以及天然末端重复(tr)、rep蛋白和衣壳亚基的序列是本领域已知的。这种序列可以在文献中或在如genbank等公共数据库中找到。参见,例如,genbank登录号nc_002077.1(aav1)、af063497.1(aav1)、nc_001401.2(aav2)、af043303.1(aav2)、j01901.1(aav2)、u48704.1(aav3a)、nc_001729.1(aav3a)、af028705.1(aav3b)、nc_001829.1(aav4)、u89790.1(aav4)、nc_006152.1(aa5)、af085716.1(aav-5)、af028704.1(aav6)、nc_006260.1(aav7)、af513851.1(aav7)、af513852.1(aav8)nc_006261.1(aav-8)、ay530579.1(aav9)、aat46337(aav10)和aao88208(aavrh10);其公开内容通过引用并入本文以教导aav核酸和氨基酸序列。参见,例如,srivistava等人,(1983)j.virology45:555;chiorini等人,(1998)j.virology71:6823;chiorini等人,(1999)j.virology73:1309;bantel-schaal等人,(1999)j.virology73:939;xiao等人,(1999)j.virology73:3994;muramatsu等人,(1996)virology221:208;shadeet.al.(1986)j.virol.58:921;gao等人,(2002)proc.nat.acad.sci.usa99:11854;moris等人,(2004)virology33:375-383;国际专利公开wo00/28061,wo99/61601,wo98/11244;和美国专利号6,156,303。
与aav血清型相关的天然存在的cap(衣壳)蛋白的序列是本领域已知的并且包含本文公开的那些序列号:如aav1(seqidno:1)、aav2(seqidno:2)、aav3a(seqidno:3)、aav3b(seqidno:4)、aav4(seqidno:5)、aav5(seqidno:6)、aav6(seqidno:7)、aav7(seqidno:8)、aav8(seqidno:9)、aav9(seqidno:10)、aav10(seqidno:11)和aavrh10(seqidno:12)。术语“变体aav衣壳蛋白”或“aav变体”是指包含氨基酸序列的aav衣壳蛋白,所述氨基酸序列包含相对于天然存在的或“野生型”aav衣壳蛋白序列(例如如本文seqidno:1-12所阐述的)的至少一个修饰或取代(包含缺失、插入、点突变等)。变体aav衣壳蛋白可以与野生型衣壳蛋白的氨基酸序列具有约80%以上的同一性,例如与野生型衣壳蛋白的氨基酸序列具有85%以上的同一性、90%以上的同一性、或95%以上的同一性,例如与野生型衣壳蛋白具有98%或99%的同一性。变体aav衣壳蛋白可能不是野生型衣壳蛋白。
出于本文公开的目的,“aav病毒粒子”或“aav病毒颗粒”是指由至少一种aav衣壳蛋白和衣壳化的aav多核苷酸构成的病毒颗粒。
出于本文公开的目的,术语“raav”是指重组腺相关病毒的缩写。应用于多核苷酸的“重组”是指多核苷酸是克隆、限制或连接步骤、以及导致不同于在自然中找到的的多核苷酸的构建体的其它过程的各种组合的产物。重组病毒是包含重组多核苷酸的病毒颗粒。所述术语分别包含原始多核苷酸构建体的复制和原始病毒构建体的后代。
术语“raav载体”涵盖raav病毒粒子(即raav病毒粒子)(例如,感染性raav病毒粒子),其定义包括raav多核苷酸;并且还涵盖编码raav的多核苷酸(例如,编码raav的单链多核苷酸(ss-raav);编码raav的双链多核苷酸(ds-raav),例如编码raav的质粒;等等)。
如果aav病毒粒子包含异源多核苷酸(即除野生型aav基因组以外的多核苷酸,例如待递送到靶细胞的转基因、待递送到靶细胞的rnai剂或crispr剂等),它通常被称为“重组aav(raav)病毒粒子”或“raav病毒颗粒”。通常,异源多核苷酸侧接至少一个,并且通常是两个aav反向末端重复序列(itr)。
术语“包装”是指导致aav颗粒的组装和衣壳化的一系列细胞内事件。aav“rep”和“cap”基因是指编码腺相关病毒的复制和衣壳化蛋白的多核苷酸序列。aavrep和cap在本文中称为aav“包装基因”。
术语aav的“辅助病毒”是指允许aav(例如野生型aav)被哺乳动物细胞复制和包装的病毒。用于aav的各种各样的辅助病毒是本领域已知的,包括腺病毒、疱疹病毒和痘病毒如牛痘病毒。腺病毒涵盖许多不同的亚组,尽管最常用的是亚组c的腺病毒5型。人、非人哺乳动物和禽类的许多腺病毒是已知的并且可从如atcc等保藏机构获得。疱疹病毒家族的病毒包括例如单纯疱疹病毒(hsv)和爱泼斯坦-巴尔病毒(ebv)、以及巨细胞病毒(cmv)和伪狂犬病病毒(prv);这些也可以从如atcc等保藏机构获得。
术语“一个或多个辅助病毒功能”是指在辅助病毒基因组中编码的一个或多个功能,其允许aav复制和包装(结合本文所述的复制和包装的其它要求)。如本文所述,“辅助病毒功能”可以多种方式提供,包括通过提供辅助病毒或向反式生产细胞提供对一个或多个必需功能进行编码的多核苷酸序列。例如,将包含编码一种或多种腺病毒蛋白的核苷酸序列的质粒或其它表达载体与raav载体一起转染到生产细胞中。
术语“感染性”病毒或病毒颗粒是包含有效组装的病毒衣壳并且能够将多核苷酸组分递送到病毒物种为热带的细胞中的术语。所述术语不一定意味着病毒的任何复制能力。在本公开内容和本领域中描述了用于计数感染性病毒颗粒的测定。病毒感染性可以表示为感染性病毒颗粒与总病毒颗粒的比例。确定感染性病毒颗粒与总病毒颗粒的比例的方法是本领域已知的。参见例如,grainger等人,(2005)mol.ther.11:s337(描述tcid50感染性滴度测定);和zolotukhin等人(1999)genether.6:973。还参见实施例。
如本文所用的术语“趋向性”是指病毒(例如,aav)优先靶向特定宿主物种的细胞或宿主物种内的特定细胞类型。例如,相对于仅可能感染肺和肌肉细胞的病毒,可以感染心脏、肺、肝和肌肉细胞的病毒具有更宽(即增加)的趋向性。趋向性还可以包含病毒对宿主的特定类型的细胞表面分子的依赖性。例如,一些病毒能够只感染具有表面糖胺聚糖的细胞,而其它病毒能够只感染具有唾液酸的细胞(这种依赖性可以使用缺乏特定类别分子的各种细胞系作为病毒感染的潜在宿主细胞进行测试)。在某些情况下,病毒的趋向性描述了病毒的相对偏好。例如,第一种病毒可能能够感染所有细胞类型,但在感染具有表面糖胺聚糖的这些细胞方面成功得多。如果第二种病毒也更喜欢相同的特征(例如,第二种病毒在感染具有表面糖胺聚糖的这些细胞方面也更成功),则可以认为第二种病毒与第一种病毒具有相似(或相同)的趋向性,即使绝对转导效率不相似。例如,第二种病毒在感染每种测试的给定细胞类型时可能比第一种病毒更有效,但如果相对偏好相似(或相同),则第二种病毒仍可被认为具有与第一种病毒相似(或相同)的趋向性。在一些实施方式中,相对于天然存在的病毒粒子,包含主题变体aav衣壳蛋白的病毒粒子的趋向性没有改变。在一些实施方式中,相对于天然存在的病毒粒子,包含主题变体aav衣壳蛋白的病毒粒子的趋向性被扩展(即,变宽)。在一些实施方式中,相对于天然存在的病毒粒子,包含主题变体aav衣壳蛋白的病毒粒子的趋向性降低。
术语“具有复制能力的”病毒(例如,具有复制能力的aav)是指具有感染性并且还能够在受感染的细胞中复制(即在存在辅助病毒或辅助病毒功能的情况下)的表型野生型病毒。在aav的情况下,复制能力通常需要存在功能性aav包装基因。通常,由于缺乏一种或多种aav包装基因,如本文所述的raav载体在哺乳动物细胞(特别是在人细胞中)中是不能复制的。通常,此类raav载体缺乏任何aav包装基因序列,以最小化通过aav包装基因与进入的raav载体之间的重组产生复制能力aav的可能性。在许多实施方式中,如本文所述的raav载体制剂是那些含有很少(如果有的话)具有复制能力的aav(rcaav,也称为rca)的那些载体制剂(例如,每102个raav颗粒小于约1rcaav、每104个raav颗粒小于约1rcaav、每10个raav颗粒小于约1rcaav、每1012个raav颗粒小于约1rcaav、或没有rcaav)。
术语“多核苷酸”是指任何长度的聚合形式的核苷酸,包含脱氧核糖核苷酸或核糖核苷酸或其类似物。多核苷酸可以包含修饰的核苷酸,如甲基化的核苷酸和核苷酸类似物,并且可以被非核苷酸组分中断。如果存在对核苷酸结构的修饰,那么在组装聚合物之前或之后进行。如本文所用,术语多核苷酸可互换地指双链和单链分子。除非另有说明或要求,否则本文包含多核苷酸的任何实施方式涵盖双链形式和已知或预测构成双链形式的两种互补单链形式中的每一种。
多核苷酸或多肽与另一多核苷酸或多肽具有一定百分比的“序列同一性”,这意味着当比对时,碱基或氨基酸的百分比在与两个序列比较时是相同的。序列相似性可以以多种不同方式确定。为了确定序列同一性,可以使用包含可通过万维网ncbi.nlm.nih.gov/blast/获得的blast的方法和计算机程序对序列进行比对。另一种比对算法是fasta,其可从是oxfordmoleculargroup,inc的全资子公司美国威斯康星州麦迪逊市的遗传计算组(geneticscomputinggroup,gcg)包中获得。其它用于比对的技术描述于methodsinenzymology,第266卷:computermethodsformacromolecularsequenceanalysis(1996),ed.doolittle,academicpress,inc.,adivisionofharcourtbrace&co.,sandiego,california,usa.。特别感兴趣的是允许序列中存在空位的比对程序。smith-waterman是一种允许序列比对空位的算法。参见meth.mol.biol.70:173-187(1997)。此外,使用needleman和wunsch比对方法的gap程序可以用于比对序列。参见j.mol.biol.48:443-453(1970)。
术语“基因”是指在细胞中发挥某种功能的多核苷酸。例如,基因可以含有能够编码基因产物的开放阅读框。基因产物的一个实例是从基因转录和翻译的蛋白质。基因产物的另一个实例是被转录但不翻译的rna,例如功能性rna产物,例如核酸酶的适体、干扰rna、核糖体rna(rrna)、转移rna(trna)、非编码rna(ncrna)、指导rna等。
术语“基因表达产物”或“基因产物”是由如上文限定的特定基因的表达产生的分子。基因表达产物包括例如多肽、适体、干扰rna、信使rna(mrna)、rrna、trna、非编码rna(ncrna)等。
术语“sirna剂”(“小干扰”或“短干扰rna”(或sirna))是靶向感兴趣基因(“靶基因”)的核苷酸的rna双链体。“rna双链体”是指通过rna分子的形成双链rna区域(dsrna)的两个区域之间的互补配对而形成的结构。sirna“靶向”基因,因为sirna的双链体部分的核苷酸序列与靶基因的核苷酸序列互补。在一些实施方式中,sirna的双链体的长度小于30个核苷酸。在一些实施方式中,双链体的长度可以是29、28、27、26、25、24、23、22、21、20、19、18、17、16、15、14、13、12、11或10个核苷酸。在一些实施方式中,双链体的长度为19-25个核苷酸。在一些实施方式中,sirna介导的基因靶向是通过使用dna指导的rna干扰(ddrnai)实现的,所述ddrnai是利用dna构建体激活动物细胞的内源性rna干扰(rnai)途径的基因沉默技术。这种dna构建体被设计成表达一旦被加工就引起一个靶基因或多个靶基因沉默的自身互补的双链rna,通常是短发夹rna(shrna)。包括内源性mrna或病毒rna的任何rna可以通过将构建体设计成表达与期望mrna靶标互补的双链rna而被沉默。因此,sirna剂的rna双链体部分可以是被称为shrna的短发夹结构的一部分。除了双链体部分之外,发夹结构可以含有位于形成双链体的两个序列之间的环部分。所述环的长度可以变化。在一些实施方式中,环的长度为5、6、7、8、9、10、11、12或13个核苷酸。发夹结构还可以包含3'或5'悬垂部分。在一些实施方式中,悬垂是长度为0、1、2、3、4或5个核苷酸的3'或5'悬垂。通常,靶基因的表达产物(例如,mrna、多肽等)的水平被含有特异性双链核苷酸序列的sirna剂(例如,sirna、shrna等)降低,所述特异性双链核苷酸序列与长度至少为19-25个核苷酸区段(例如,20-21个核苷酸序列)的靶基因转录物互补,包括5'非翻译(ut)区、orf或3'ut区。在一些实施方式中,短干扰rna的长度为约19-25nt。参见,例如,pct申请wo00/44895、wo99/32619、wo01/75164、wo01/92513、wo01/29058、wo01/89304、wo02/16620和wo02/29858;以及描述sirna技术的美国专利公开号2004/0023390。sirna和/或shrna可以由核酸序列编码,并且核酸序列还可以包括启动子。核酸序列还可以包括多腺苷酸化信号。在一些实施方式中,多腺苷酸化信号是合成的最小多腺苷酸化信号。
术语“反义rna”涵盖与基因表达产物互补的rna。例如,靶向特异性mrna的反义rna是基于rna的试剂(或可以是修饰的rna),所述基于rna的试剂与mrna互补,其中反义rna与mrna的杂交改变mrna的表达(例如,通过改变rna的稳定性、改变rna的翻译等)。“反义rna”中还包括编码反义rna的核酸。
关于“crispr/cas9剂”,术语“crispr”涵盖聚集的规则间隔短回文重复/crispr相关(cas)系统,其通过使用crisprrna(crrna)指导入侵核酸沉默而进化以提供针对病毒和质粒的细菌和古细菌的适应性免疫。cas9蛋白(或其功能等同物和/或其变体,即cas9样蛋白)天然含有dna核酸内切酶活性,其依赖于蛋白质与被称为crrna和tracrrna(也称为指导rna)的两种天然存在的或合成的rna分子缔合。在一些情况下,两个分子共价连接以形成单个分子(也称为单个指导rna(“sgrna”))。因此,cas9或cas9样蛋白与dna靶向rna(所述术语涵盖双分子指导rna构造和单分子指导rna构造)相关联,所述dna靶向rna激活cas9或cas9样蛋白并将蛋白质指导到靶核酸序列。
如果cas9或cas9样蛋白保留其天然酶功能,则它将切割靶dna以产生双链断裂,这可导致基因组改变(即编辑:缺失、插入(当存在供体多核苷酸时)、替换等),从而改变基因表达。cas9的一些变体(其变体被术语cas9样涵盖)已被改变,使得它们具有降低的dna切割活性(在一些情况下,它们切割单链而不是靶dna的两条链,而在其它情况下,它们严重减少到没有dna切割活性)。具有降低的dna切割活性(甚至没有dna切割活性)的cas9样蛋白仍然可以被引导到靶dna以阻断rna聚合酶活性。可替代地,cas9或cas9样蛋白可以通过将vp64转录激活结构域与cas9蛋白融合并用ms2-p65-hsf1辅助蛋白和在四环上包含ms2rna适体的单一指导rna编码协同递送来修饰以在激活转录的细胞中形成协同激活介质(cas9-sam)复合物。因此,酶促失活的cas9样蛋白可以通过靶向dna的rna来靶向靶dna中的特定位置,以阻断或激活靶dna的转录。如本文所用的术语“crispr/cas9剂”涵盖如上所述或本领域已知的所有形式的crispr/cas9。
关于crispr剂的详细信息可以在以下文献中找到:例如(a)jinek等人,science.2012aug17;337(6096):816-21:"aprogrammabledual-rna-guideddnaendonucleaseinadaptivebacterialimmunity";(b)qi等人,cell.2013feb28;152(5):1173-83:"repurposingcrisprasanrna-guidedplatformforsequence-specificcontrolofgeneexpression",和(c)美国专利申请号13/842,859和pct申请号pct/us13/32589;其通过引用以其全部内容并入本文。因此,本文使用的术语“crispr剂”涵盖可以在基于cas9的系统(例如,cas9或cas9样蛋白;靶向dna的rna的任何组分,例如crrna样rna、tracrrna样rna、单一指导rna等;供体多核苷酸;等等)中使用的包含天然存在和/或合成序列的任何试剂。
“锌指核酸酶”(zfn)是指通过将锌指dna结合结构域与dna切割结构域融合而产生的人工dna内切核酸酶。zfn可以被工程化成靶向期望的dna序列,并且这使得锌指核酸酶能够切割独特的靶序列。当引入细胞中时,zfn可以用于通过诱导双链断裂来编辑细胞中的靶dna(例如,细胞的基因组)。有关使用zfn的更多信息,请参见例如:asuri等人,molther.2012feb;20(2):329-38;bibikova等人,science.2003may2;300(5620):764;wood等人,science.2011jul15;333(6040):307;ochiai等人,genescells.2010aug;15(8):875-85;takasu等人,insectbiochemmolbiol.2010oct;40(10):759-65;ekker等人,zebrafish2008summer;5(2):121-3;young等人,procnatlacadsciusa.2011apr26;108(17):7052-7;goldberg等人,cell.2010mar5;140(5):678-91;geurts等人,science.2009jul24;325(5939):433;flisikowska等人,plosone.2011;6(6):e21045.doi:10.1371/journal.pone.0021045.epub2011jun13;hauschild等人,procnatlacadsciusa.2011jul19;108(29):12013-7;andyu等人,cellres.2011nov;21(l1):1638-40;所有这些文献通过引用其与zfn相关的教导并入本文。术语“zfn剂”涵盖锌指核酸酶和/或包含编码锌指核酸酶的核苷酸序列的多核苷酸。
术语“转录激活子样效应子核酸酶”或“talen”剂是指转录激活子样效应子核酸酶(talen)是通过将tal(转录激活子样)效应子dna结合结构域与dna切割结构域融合而产生的人工dna核酸内切酶。talens可以被快速工程化成实际上结合任何期望dna序列,并且当引入细胞时,talen可以通过诱导双链断裂来编辑细胞中的靶dna(例如,细胞的基因组)。有关使用talen的更多信息,请参见例如:hockemeyer等人,natbiotechnol.2011jul7;29(8):731-4;wood等人,science.2011jul15;333(6040):307;tesson等人,natbiotechnol.2011aug5;29(8):695-6;andhuang等人,natbiotechnol.2011aug5;29(8):699-700;所有这些文献通过引用其与talen相关的教导并入本文。术语“talen剂”涵盖talen和/或包含编码talen的核苷酸序列的多核苷酸。
术语“控制元件”或“控制序列”是指参与分子相互作用的核苷酸序列,所述核苷酸序列有助于多核苷酸的功能调节,包括多核苷酸的复制、重复、转录、剪接、翻译或简并。所述调节可以影响过程的频率、速度或特异性,并且可能本质上是增强或抑制的。本领域已知的控制元件包括例如转录调节序列,如启动子和增强子。启动子是能够在某些条件下与rna聚合酶结合并且启动通常位于启动子下游(3'方向)的编码区的转录的dna区域。启动子可以是普遍起作用型的,即在许多细胞类型中有活性,例如cag或cmv启动子;或是组织或细胞特异性型的,例如在视杆中有活性的rho启动子,或在视锥中有活性的视蛋白启动子。
术语“可操作地连接(operativelylinked或operablylinked)”是指遗传元件的并置,其中所述元件处于允许其以预期方式操作的关系中。例如,如果启动子有助于启动编码序列的转录,则启动子可操作地连接到编码区。在启动子和编码区之间可以存在间插残基,只要保持这种功能关系即可。
术语“表达载体”涵盖包含对感兴趣多肽进行编码的多核苷酸区域的载体,并且用于实现蛋白质在指定靶细胞中的表达。表达载体还可以包含与编码区可操作地连接的控制元件,以促进蛋白质在靶标中的表达。控制元件以及与其可操作地连接以进行表达的一个基因或多个基因的组合有时被称为“表达盒”,其中许多是本领域已知的并可获得的,或者可以容易地由在本领域可获得的组分构造。
术语“异源”是指源自与其进行比较的实体的其余部分的基因型不同的实体。例如,通过基因工程技术引入到衍生自不同物种的质粒或载体中的多核苷酸是异源多核苷酸。从其天然编码序列中除去并与天然未发现连接的编码序列可操作地连接的启动子是异源启动子。因此,例如,包含对异源基因产物进行编码的异源核酸序列的raav是包括通常不包括在天然存在的野生型aav中的多核苷酸的raav,并且编码的异源基因产物是通常不由天然存在的野生型aav编码的基因产物。
术语“遗传改变”和“遗传修饰”(和其语法变体)在本文中可互换使用以指代其中遗传元件(例如,多核苷酸)被引入除有丝分裂或减数分裂之外的细胞中的过程。所述元件对于细胞可以是异源的,或者它可以是已经存在于细胞中的元件的额外拷贝或改进版本。遗传改变可以例如通过用重组质粒或其它多核苷酸通过本领域已知的任何方法转染细胞来实现,如电穿孔、磷酸钙沉淀、或与多核苷酸-脂质体复合物接触。遗传改变还可以例如通过用dna或rna病毒或病毒载体转导或感染来实现。通常,遗传元件被引入细胞中的染色体或微型染色体中;但是任何改变细胞和其后代的表型和/或基因型的改变都包含在此术语中。
关于细胞修饰,术语(例如通过重组病毒)由外源性dna进行的“遗传修饰”或“转化”或“转染”或“转导”的术语是指当这种dna已引入细胞内时。外源dna的存在导致永久或瞬时的遗传变化。转化dna可以整合或可以不整合(共价连接)到细胞的基因组中。“克隆”是通过有丝分裂从单个细胞或共同祖先衍生的细胞群。“细胞系”是能够在体外稳定生长许多代的原代细胞的克隆。
如本文所用,如果序列可用于在体外延长的细胞培养期间和/或在体内延长的时间段执行其功能,则称细胞被“稳定地”改变、转导、遗传修饰或用基因序列转化。通常,这种细胞被“可遗传地”改变(遗传修饰),因为引入了还可通过改变的细胞的后代遗传的遗传改变。
在此可互换使用的术语“多肽”、“肽”和“蛋白质”是指具有任何长度的氨基酸的聚合物。这些术语还涵盖已修饰的氨基酸聚合物;例如,二硫键形成、糖基化、脂化、磷酸化或与标记组分的缀合。当在向哺乳动物受试者递送基因产物的背景下讨论时,多肽如抗血管生成多肽、神经保护多肽等和其组合物是指保留了完整蛋白质所需的生化功能的相应的完整多肽、或其任何片段或基因工程衍生物。类似地,提及编码抗血管生成多肽的核酸、编码神经保护多肽的核酸和其它用于将基因产物递送到哺乳动物受试者(其可以被称为待递送到受体细胞的“转基因”)的此类核酸,包括编码完整多肽的多核苷酸或具有期望生化功能的任何片段或基因工程化衍生物。
如本文所用,“分离的”质粒、核酸、载体、病毒、病毒粒子、宿主细胞、蛋白质或其它物质是指缺乏至少一些其它组分的物质的制剂,所述其它组分还可以存在于天然存在或最初由其制备的物质或类似物质中。因此,例如,可以通过使用纯化技术从源混合物中富集分离的物质来制备所述分离的物质。富集可以在绝对基础上测量,如每体积溶液的重量,或者可以相对于源混合物中存在的第二种潜在干扰物质进行测量。本公开的实施方式的增加的富集是越来越分离的。在一些实施方式中,分离的质粒、核酸、载体、病毒、宿主细胞或其它物质是纯化的,例如纯度为约80%至约90%、纯度至少为约90%、纯度至少为约95%、纯度至少为约98%、或纯度至少为约99%或更多。
如本文所用,术语“治疗(treatment、treating)”等是指获得期望药理学和/或生理学效果。就完全或部分预防疾病或其症状而言,所述效果可以是预防性的,和/或就部分或完全治愈疾病和/或可归因于疾病的副作用而言可以是治疗性的。如本文所用,“治疗”涵盖哺乳动物,特别是人类的疾病的任何治疗,并且包括:(a)预防可能在受试者中发生的疾病(和/或由疾病引起的症状),所述受试者可能易患所述疾病或有患病风险但尚未被诊断为患有所述疾病;(b)抑制疾病(和/或由疾病引起的症状),即阻止其发展;以及(c)缓解疾病(和/或由疾病引起的症状),即引起疾病(和/或由疾病引起的症状)消退,即改善疾病和/或疾病的一种或多种症状。例如,主题组合物和方法可以针对视网膜疾病的治疗。用于评估视网膜疾病和其治疗的非限制性方法包括测量视网膜功能和其变化,例如视力的变化(例如最佳矫正视力[bcva]、移动、导航、物体检测和辨别)、视野的变化(例如,静态和动态视野的视野检查)、临床检查(例如,眼睛前后段的裂隙灯检查)、对所有光明和黑暗波长的电生理反应(例如所有形式的视网膜电图(erg)[全视野,多视野和模式]、所有形式的视觉诱发电位(vep)、眼电图(eog)、色觉、暗适应和/或对比敏感度;使用解剖学和/或摄影措施来测量解剖学或健康的变化,例如光学相干断层扫描(oct)、眼底照相、自适应光学扫描激光检眼镜、荧光和/或自发荧光;测量眼球动性和眼球运动(例如,眼球震颤、固定偏好和稳定性)、测量报告的结果(患者报告的视觉和非视觉引导行为和活动的变化、患者报告的结果[pro]、基于问卷的生活质量评估、日常活动和神经功能的测量(例如功能性磁共振成像(mri))。
术语“个体”,“宿主”,“受试者”和“患者”在本文中可互换使用,并且指哺乳动物,包括但不限于:人;非人类灵长类动物,包含猿猴;哺乳动物运动动物(如马);哺乳动物农场动物(如绵羊,山羊等);哺乳动物宠物(狗,猫等);和啮齿动物(例如,小鼠,大鼠等)。
在一些实施方式中,个体是先前已经天然暴露于aav并且因此携带抗aav抗体(即,aav中和抗体)的人。在一些实施方式中,个体是先前已经施用aav载体的人(并且因此可能携带抗aav抗体)并且需要再次施用载体以治疗不同病症或进一步治疗相同病症。基于将aav基因递送到受针对该载剂的中和抗体影响的所有组织例如肝脏、肌肉和视网膜的临床试验中的阳性结果,存在许多此类治疗应用/疾病靶。
如本文所用的,术语“有效量”是足以实现有益或期望的临床结果的量。有效量可以在一次或多次施用中施用。出于本公开的目的,有效量的化合物(例如,感染性raav病毒粒子)是足以减轻、改善、稳定、逆转、预防、减缓或延迟(和/或与其相关的症状)特定疾病状态(例如,视网膜疾病)的进展的量。因此,有效量的感染性raav病毒粒子是能够将异源核酸有效递送到个体的靶细胞(或多个靶细胞)的感染性raav病毒粒子的量。有效量可以通过例如使用本领域熟知的技术例如rt-pcr、蛋白质印迹、elisa、荧光或其它报道因子读数等在细胞或组织中检测由异源核酸序列编码的基因产物(rna,蛋白质)来临床前确定。如本文所述和如本领域已知的,有效量可以通过例如使用本领域已知的方法例如眼底自发荧光、荧光血管造影、oct、微观测量、自适应光学等检测疾病的发作或进展的变化来临床确定。
术语“视网膜细胞”在本文中是指包含视网膜的任何细胞类型,如但不限于视网膜神经节(rg)细胞、无长突细胞、水平细胞、双极细胞、感光细胞、穆勒神经胶质细胞、小神经胶质细胞和视网膜色素上皮(rpe)。术语“感光细胞”在本文中是指但不限于视杆细胞或“视杆”和视锥细胞或“视锥”。术语“穆勒细胞”或“穆勒神经胶质”是指支持脊椎动物视网膜中神经元的神经胶质细胞。
术语“定向进化”是指体外和/或体内衣壳工程方法论,其通过迭代轮次的遗传多样化和选择过程模拟自然进化,从而积累逐渐改善生物分子的功能的有益突变。定向进化通常涉及被称为“生物淘选”的体内方法,其用于从文库中选择aav变体,所述变体具有感兴趣的细胞或组织类型的更有效水平的感染性。
具体实施方式
腺相关病毒(aav)是一种细小病毒家族,其具有包含在无包膜衣壳内的4.7kb的单链dna基因组。天然存在的aav的病毒基因组具有2个反向末端重复序列(itr)-其作为复制和包装信号的病毒源起作用-侧接2个主要开放阅读框(orf):rep(编码在病毒复制中起作用的蛋白质、转录调节、位点特异性整合和病毒粒子装配)和cap。caporf编码3种结构蛋白,这3种结构蛋白组装形成60-mer病毒衣壳。已经分离出许多天然存在的aav变体和血清型,并且没有一种与人类疾病相关。
aav的重组版本可以用作基因递送载体,其中感兴趣的标记或治疗基因插入取代rep和cap的itr之间。已经示出这些载体在体外和体内转导分裂和非分裂细胞,并且可以在有丝分裂后组织中导致稳定的转基因表达多年。参见例如,knipedm,howleypm.fields’virology.lippincottwilliams&wilkins,philadelphia,pa,usa,2007;gaog-p,alviramr,wangl,calcedor,johnstonj,wilsonjm.noveladeno-associatedvirusesfromrhesusmonkeysasvectorsforhumangenetherapy.procnatlacadsciusa2002;99:11854–9;atchisonrw,castobc,hammonwm.adenovirus-associateddefectivevirusparticles.science1965;149:754–6;hogganmd,blacklownr,rowewp.studiesofsmalldnavirusesfoundinvariousadenoviruspreparations:physical,biological,andimmunologicalcharacteristics.procnatlacadsciusa1966;55:1467–74;blacklownr,hogganmd,rowewp.isolationofadenovirus-associatedvirusesfromman.procnatlacadsciusa1967;58:1410–5;bantel-schaalu,zurhausenh.characterizationofthednaofadefectivehumanparvovirusisolatedfromagenitalsite.virology1984;134:52–63;mayorhd,melnickjl.smalldeoxyribonucleicacid-containingviruses(picodnavirusgroup).nature1966;210:331–2;moris,wangl,takeuchit,kandat.twonoveladeno-associatedvirusesfromcynomolgusmonkey:pseudotypingcharacterizationofcapsidprotein.virology2004;330:375–83;flottetr.genetherapyprogressandprospects:recombinantadeno-associatedvirus(raav)vectors.genether2004;11:805–10。
重组aav(在本文中简称为“aav”)在越来越多的临床试验中具有有希望的结果。然而,基因递送的障碍可能限制aav的效用,如抗衣壳免疫应答、某些组织的低转导、不能靶向递送到特定细胞类型和相对低的携带能力。在许多情况下,没有足够的机械知识来有效地赋予具有改善aav的能力的合理设计。作为替代方案,定向进化已成为创建满足特定生物医学需求的新型aav变体的策略。定向进化策略利用遗传多样化和选择过程来能够积累逐步改善生物分子的功能的有益突变。在此过程中,野生型aavcap基因通过若干种方法多样化以产生大型遗传文库,所述大型遗传文库被包装以产生病毒颗粒文库,并且然后应用选择性压力以分离可以克服基因递送障碍的新型变体。重要的是,基因递送问题的机制基础不需要为功能的定向进化所知,这可以因此加速增强的载体的开发。
通常,本文公开的变体是通过使用一个aav文库和/或多个aav文库产生的。通过病毒基因组工程化领域的技术人员已知且可容易获得的一系列定向进化技术,使cap基因突变而产生此类一种aav文库或多种aav文库,所述cap基因是一种编码aav衣壳的结构蛋白的基因。参见例如,bartel等人,am.soc.genecellther.15thannu.meet.20,s140(2012);bowles,d.等人,j.virol.77,423-432(2003);gray等人,mol.ther.18,570-578(2010);grimm,d.等人,j.virol.82,5887-5911;koerber,j.t.等人,mol.ther.16,1703-1709(2008);liw.等人,mol.ther.16,1252-1260(2008);koerber,j.t.等人,methodsmol.biol.434,161-170(2008);koerber,j.t.等人,hum.genether.18,367-378(2007);和koerber,j.t.等人,mol.ther.17,2088-2095(2009)。这种技术非限制性地如下:i)用于以预定的可修改的速率将随机点突变引入aavcap开放阅读框(orf)的易错pcr;ii)体外或体内病毒重组或“dna改组”,其用于产生aavcap基因的随机嵌合体以产生具有多种aav血清型的基因文库;iii)通过在caporf中连接简并寡核苷酸而在衣壳的限定位点处的随机肽插入;iv)使用转座子诱变将肽编码序列限定插入aavcaporf的随机位置;v)用基于天然aav血清型和变体中每个氨基酸位置的保守水平而生物信息设计的肽序列文库替换aav衣壳的表面环,以产生“环交换”文库;vi)在aav血清型之间的简并性位置处的随机氨基酸取代以产生祖先变体的文库(santiago-ortiz等人,2015);和其此类技术的组合。
dna改组产生嵌合体,所述嵌合体以独特且通常有益的方式结合其亲本特性;然而,有些可能无法进行包装,这实际上会降低文库的多样性。通过肽插入技术实现文库的多样性浓度,例如但不限于上述iii-iv)。文库的多样性也集中在诸如上述v)的技术中,并且这种浓度指向位于aav衣壳的表面暴露环上的多个高变区。虽然许多技术产生变异衣壳,其中只有衣壳的小部分突变,但这些技术可以与额外诱变策略配对以修饰完整的衣壳。
一旦产生一个aav文库或多个aav文库,病毒然后就被包装,使得每个aav颗粒由围绕对突变衣壳进行编码的cap基因的突变衣壳组成并纯化。然后使文库的变体经受aav领域的技术人员已知的并且容易获得的体外和/或体内选择性压力技术。参见例如maheshri,n.等人,naturebiotech.24,198-204(2006);dalkara,d.等人,sci.transl.med.5,189ra76(2013);lisowski,l.等人,nature.506,382-286(2013);yang,l.等人,pnas.106,3946-3951(2009);gao,g.等人,mol.ther.13,77-87(2006);和bell,p.等人,hum.gene.ther.22,985-997(2011)。例如但非限制性地,可以使用以下选择aav变体:i)亲和柱,其中不同级分的洗脱产生具有改变的结合特性的变体;ii)原代细胞-其从模拟人体细胞行为的组织样品或永生细胞系中分离-其产生具有提高的效率和/或组织特异性的aav变体;iii)动物模型-其模拟临床基因疗法环境-其产生已成功感染靶组织的aav变体;iv)人异种移植模型,其产生已感染移植的人细胞的aav变体;和/或其选择技术的组合。
一旦选择了病毒,就可以通过已知技术回收它们,例如但不限于腺病毒介导复制、pcr扩增、新一代测序和克隆等。然后通过重复轮次的选择技术富集病毒克隆,并分离aavdna以回收所选择的感兴趣的变体cap基因。这种选择的变体可以经受进一步的修饰或突变,并且因此作为进一步选择步骤的新起点以迭代地增加aav病毒适合度。然而,在某些情况下,已经生成了成功的衣壳而没有额外的突变。
本文公开的aav变体至少部分地通过使用体内定向进化方法(例如上述技术)产生,所述方法涉及在玻璃体内施用后使用灵长类视网膜筛选。因此,本文公开的aav变体衣壳包含氨基酸序列中的一个或多个修饰,与相应的亲本aav衣壳蛋白相比,其赋予对灵长类动物视网膜细胞更有效的转导。如本文所用,“相应的亲本aav衣壳蛋白”是指与主题变体aav衣壳蛋白相同的野生型或变体aav血清型的aav衣壳蛋白,但不包含主题变体aav衣壳蛋白的一个或多个氨基酸序列修饰。
在一些实施方式中,主题变体aav衣壳蛋白包含相对于相应的亲本aav衣壳蛋白通过共价连接插入aav衣壳蛋白gh环或环iv中的约5个氨基酸至约20个氨基酸的异源肽。aav衣壳蛋白的“gh环”或环iv,是指本领域中称为aav衣壳蛋白的gh环或环iv的溶剂可及部分。对于aav衣壳的gh环/环iv,参见例如vanvliet等人,(2006)mol.ther.14:809;padron等人,(2005)j.virol.79:5047;和shen等人,(2007)mol.ther.15:1955。因此,例如,插入位点可以在aavvp1衣壳蛋白的氨基酸约411-650内。例如,插入位点可以在aav1vp1的氨基酸571-612、aav2vp1的氨基酸570-611内、aav3avp1的氨基酸571-612内、在aav3bvp1的氨基酸571-612内、在aav4vp1的氨基酸569-610内、在aav5vp1的氨基酸560-601内、在aav6vp1的氨基酸571-612内、在aav7vp1的氨基酸572-613内、在aav8vp1的氨基酸573-614内、在aav9vp1的氨基酸571-612内、或aav10vp1的氨基酸573-614内、或其任何变体的相应氨基酸。基于各种aav血清型的衣壳蛋白的氨基酸序列的比较,本领域技术人员将知道“对应于aav2的氨基酸”的插入位点将在任何给定aav血清型的衣壳蛋白中。还参见图6以了解野生型aavseqidno:1-11的比对,其提供野生型(天然存在的)血清型aav1、aav2、aav3a、aav3b和aav4-10之间和跨其的氨基酸位置。
在某些实施方式中,插入位点是位于任何野生型aav血清型或aav变体的vp1的氨基酸570-614之间的两个相邻氨基酸之间的单个插入位点,例如,插入位点在位于任何aav血清型或变体的vp1的氨基酸570-610、氨基酸580-600、氨基酸570-575、氨基酸575-580、氨基酸580-585、氨基酸585-590、氨基酸590-600、或氨基酸600-614的两个相邻氨基酸之间。例如,插入位点可以在氨基酸580与581之间、氨基酸581与582之间、氨基酸583与584之间、氨基酸584与585之间、氨基酸585与586之间、氨基酸586与587之间、氨基酸587与588之间、氨基酸588与589之间、或氨基酸589与590之间。插入位点可以在氨基酸575与576之间、氨基酸576与577之间、氨基酸577与578之间、氨基酸578与579之间、或氨基酸579与580之间。插入位点可以在氨基酸590与591之间、氨基酸591与592之间、氨基酸592与593之间、氨基酸593与594之间、氨基酸594与595之间、氨基酸595与596之间、氨基酸596与597之间、氨基酸597与598之间、氨基酸598与599之间、或氨基酸599与600之间。例如,插入位点可以在aav2的氨基酸587与588之间、aav1的氨基酸590与591之间、aav3a的氨基酸588与589之间、aav3b的氨基酸588与589之间、aav4的氨基酸584与585之间、aav5的氨基酸575与576之间、aav6的氨基酸590与591之间、aav7的氨基酸589与590之间、aav8的氨基酸590与591之间、aav9的氨基酸588与589之间、或aav10的氨基酸588与589之间。
在一些实施方式中,本文公开的肽插入的长度为5个氨基酸、6个氨基酸、7个氨基酸、8个氨基酸、9个氨基酸、10个氨基酸、11个氨基酸、12个氨基酸、13个氨基酸、14氨基酸、15个氨基酸、16个氨基酸、17个氨基酸、18个氨基酸、19个氨基酸或20个氨基酸。在另一个实施方式中,本文公开的肽插入在本文公开的肽插入中的任何一种的氨基末端(n末端)和/或羧基末端(c末端)包含1至4个间隔氨基酸。示例性间隔氨基酸包括但不限于亮氨酸(l)、丙氨酸(a)、甘氨酸(g)、丝氨酸(s)、苏氨酸(t)和脯氨酸(p)。在某些实施方式中,肽插入包含在n末端处的2个间隔氨基酸和在c末端处的2个间隔氨基酸。在其它实施方式中,肽插入包含在n末端处的2个间隔氨基酸和在c末端处的1个间隔氨基酸。
本文公开的肽插入先前未描述和/或插入aav衣壳中。不希望受理论束缚,任何公开的肽插入的存在可以起到降低变体衣壳对硫酸肝素的亲和力的作用,这可能降低与灵长类动物视网膜前面的细胞外基质的结合。此外,本文公开的肽插入基序可通过添加细胞表面受体结合结构域而赋予对灵长类动物视网膜细胞增强的转导。
在一些优选的实施方式中,插入肽包含下式中任一个的氨基酸序列。
在一些方面,插入肽可以是式1a的长度为7至10个氨基酸的肽:
y1y2x1x2x3x4x5x6x7y3
其中y1-y3中的每一个(如果存在的话)独立地选自ala、leu、gly、ser、thr、pro
x1选自gln、asn、his、ile和ala
x2选自ala、gln、asp、ser、lys和pro
x3选自asp、ile、thr和asn
x4选自thr、ser、tyr、gln、glu和ala
x5选自thr、lys和asn
x6选自lys、asn和glu
x7选自asn、thr、ile、his、asp和ala。
在某些实施方式中,式1a的插入肽包含选自以下的氨基酸序列:qadttkn(seqidno:13)、isdqtkh(seqidno:14)、asdstka(seqidno:15)、nqdytkt(seqno:16)、hditkni(seqidno:17)、hpdttkn(seqidno:18)、hqdttkn(seqidno:19)、nkttnkd(seqidno:20)、isneneh(seqidno:21)和qananen(seqidno:22)。
在其它方面,插入肽可以是式1b的长度为7至10个氨基酸的肽:
y1y2x1x2x3x4x5x6x7y3
其中y1-y3中的每一个(如果存在的话)独立地选自ala、leu、gly、ser、thr、pro
x1选自gln、asn、his和ile
x2选自ala、gln、asp和ser
x3选自asp和ile
x4选自thr、tyr和gln
x5选自thr和lys
x6选自lys和asn
x7选自asn、thr、ile和his
在某些实施方式中,式1b的插入肽包含选自以下的氨基酸序列:qadttkn(seqidno:13)、isdqtkh(seqidno:14)、nqdytkt(seqno:16)、hditkni(seqidno:17)和hqdttkn(seqidno:19)。
在其它方面,插入肽可以是式1c的长度为7至10个氨基酸的肽:
y1y2x1x2aspx3thrlysx4y3
其中y1-y3中的每一个(如果存在的话)独立地选自ala、leu、gly、ser、thr、pro
x1选自gln、asn、his和ile
x2选自ala、gln和ser
x3选自thr、tyr和gln
x4选自asn、thr和his
在某些实施方式中,式1c的插入肽包含选自以下的氨基酸序列:qadttkn(seqidno:13)、isdqtkh(seqidno:14)、nqdytkt(seqno:16)和hqdttkn(seqidno:19)。
在其它方面,插入肽可以是式1d的长度为7至10个氨基酸的肽:
y1y2x1x2aspx3thrthrx4y3
其中y1-y3中的每一个(如果存在的话)独立地选自ala、leu、gly、ser、thr、pro
x1选自gln和ile
x2选自ala和ser
x3选自thr和gln
x4选自asn和his
在某些实施方式中,式1d的插入肽包含选自以下的氨基酸序列:qadttkn(seqidno:13)和isdqtkh(seqidno:14)。
在其它方面,插入肽可以是式1e的长度为7至11个氨基酸的肽:
y1y2x1x2asnx3asnglux4y3
其中y1-y3中的每一个(如果存在的话)独立地选自ala、leu、gly、ser、thr、pro
x1选自gln和ile
x2选自ala和ser
x3选自glu和ala
x4选自asn和his
在其它实施方式中,式1e的插入肽包含选自以下的氨基酸序列:isneneh(seqidno:21)和qananen(seqidno:22)。
在又另一个实施方式中,插入肽可以是式iia的长度为7至11个氨基酸的肽:
y1y2x1x2dx3tkx4y3.
其中y1-y3中的每一个(如果存在的话)独立地选自ala、leu、gly、ser、thr、pro
x1选自q、n、a、h和i;
x2选自q、a、p和s
x3选自t、y、s和q;并且
x4选自t、n、a和h。
在式x1x2dx3tkx4的氨基酸序列的肽插入的另一个实施方式中,肽插入选自由以下组成的组:qadttkn(seqidno:13)、isdqtkh(seqidno:14)、asdstka、nqdytkt(seqno:16)、hqdttkn(seqidno:19)和hpdttkn(seqidno:18)。
在一些这种实施方式中,插入肽可以是式iib的长度为7至11个氨基酸的肽:
y1y2x1x2dx3tkx4y3
其中y1-y3中的每一个(如果存在的话)独立地选自ala、leu、gly、ser、thr、pro
x1选自n、a和h;
x2选自q、p和s;
x3选自t、y和s;并且
x4选自t、n和a。
在式x1x2dx3tkx4的氨基酸序列的肽插入的另一个实施方式中,肽插入选自由以下组成的组:asdstka、nqdytkt(seqno:16)、hqdttkn(seqidno:19)和hpdttkn(seqidno:18)。
在其它实施方式中,插入肽包含选自以下的氨基酸序列:kdrapst(seqidno:26)、tnrtspd(seqidno:24)、pnsthgs(seqidno:25)和gkskvid(seqidno:23)。
在一些实施方式中,插入肽包含选自以下的氨基酸序列:asdstka(seqidno:15)、qananen(seqidno:22)、qadttkn(seqidno:13)、isdqtkh(seqidno:14)、nqdytkt(seqidno:16)、hditkni(seqidno:17)、hpdttkn(seqidno:18)、hqdttkn(seqidno:19)、nkttnkd(seqidno:20)、isneneh(seqidno:21)、gkskvid(seqidno:23)、tnrtspd(seqidno:24)、pnsthgs(seqidno:25)和kdrapst(seqidno:26)。
在其它优选的实施方式中,插入肽在选自以下的氨基酸序列的氨基和/或羧基末端具有1至3个间隔氨基酸(y1-y3):qadttkn(seqidno:13)、isdqtkh(seqidno:14)、asdstka(seqidno:15)、nqdytkt(seqidno:16)、hditkni(seqidno:17)、hpdttkn(seqidno:18)、hqdttkn(seqidno:19)、nkttnkd(seqidno:20)、isneneh(seqidno:21)、qananen(seqidno:22)、gkskvid(seqidno:23)、tnrtspd(seqidno:24)、pnsthgs(seqidno:25)和kdrapst(seqidno:26)。在某些这种实施方式中,插入肽选自由以下组成的组:laqadttkna(seqidno:27)、laisdqtkha(seqidno:28)、lgisdqtkha(seqidno:29)、laasdstkaa(seqidno:30)、lanqdytkta(seqidno:31)、lahditknia(seqidno:32)、lahpdttkna(seqidno:33)、lahqdttkna(seqidno:34)、lankttnkda(seqidno:35)、lpisneneha(seqidno:36)、lpqananena(seqidno:37)、lagkskvida(seqidno:38)、latnrtspda(seqidno:39)、lapnsthgsa(seqidno:40)和lakdrapsta(seqidno:41)。
在一些实施方式中,主题变体aav衣壳蛋白不包含除gh环或环iv中约5个氨基酸至约20个氨基酸的肽插入之外的任何其它氨基酸序列修饰。例如,在一些实施方式中,主题变体aav衣壳蛋白包含肽插入,所述肽插入包含选自由以下组成的组的氨基酸序列:qadttkn(seqidno:13)、isdqtkh(seqidno:14)、asdstka(seqidno:15)、nqdytkt(seqidno:16)、hditkni(seqidno:17)、hpdttkn(seqidno:18)、hqdttkn(seqidno:19)、nkttnkd(seqidno:20)、isneneh(seqidno:21)、qananen(seqidno:22)、gkskvid(seqidno:23)、tnrtspd(seqidno:24)、pnsthgs(seqidno:25)、kdrapst(seqidno:26)、laqadttkna(seqidno:27)、laisdqtkha(seqidno:28)、lgisdqtkha(seqidno:29)、laasdstkaa(seqidno:30)、lanqdytkta(seqidno:31)、lahditknia(seqidno:32)、lahpdttkna(seqidno:33)、lahqdttkna(seqidno:34)、lankttnkda(seqidno:35)、lpisneneha(seqidno:36)、lpqananena(seqidno:37)、lagkskvida(seqidno:38)、latnrtspda(seqidno:39)、lapnsthgsa(seqidno:40)和lakdrapsta(seqidno:41),并且变体aav衣壳不包含任何其它氨基酸取代、插入或缺失(即,变体aav衣壳蛋白包含所述插入,并且在其它方面与相应的aav衣壳蛋白一致)。换句话说,包含所述插入的变体aav衣壳蛋白在其它方面与肽已插入的亲本aav衣壳蛋白一致。作为另一个实例,主题变体aav衣壳蛋白包含具有选自以下的氨基酸序列的肽插入:qadttkn(seqidno:13)、isdqtkh(seqidno:14)、asdstka(seqidno:15)、nqdytkt(seqidno:16)、hditkni(seqidno:17)、hpdttkn(seqidno:18)、hqdttkn(seqidno:19)、nkttnkd(seqidno:20)、isneneh(seqidno:21)、qananen(seqidno:22)、gkskvid(seqidno:23)、tnrtspd(seqidno:24)、pnsthgs(seqidno:25)、kdrapst(seqidno:26)、laqadttkna(seqidno:27)、laisdqtkha(seqidno:28)、lgisdqtkha(seqidno:29)、laasdstkaa(seqidno:30)、lanqdytkta(seqidno:31)、lahditknia(seqidno:32)、lahpdttkna(seqidno:33)、lahqdttkna(seqidno:34)、lankttnkda(seqidno:35)、lpisneneha(seqidno:36)、lpqananena(seqidno:37)、lagkskvida(seqidno:38)、latnrtspda(seqidno:39)、lapnsthgsa(seqidno:40)和lakdrapsta(seqidno:41),其中肽插入位于aav2衣壳的vp1的氨基酸587与588或另一种亲本aav的vp1的相应氨基酸之间,例如在aav1、aav3a、aav3b、aav6或aav9的vp1的氨基酸588与589之间、在aav4的vp1的氨基酸586与587之间、在aav5的vp1的氨基酸577与578之间、在aav7的vp1的氨基酸589与590之间、在aav8或aav10的vp1的氨基酸590到591之间等,其中变体aav衣壳蛋白序列在其它方面与相应的亲本aav衣壳蛋白序列一致,例如seqidno:1-12中的任何一个。
在其它实施方式中,除了包含gh环中例如如本文公开的或如本领域已知的肽插入外,主题变体aav衣壳蛋白包含约1至约100个氨基酸取代或缺失,例如,与亲本aav衣壳蛋白相比1个至约5个、约2个至约4个、约2个至约5个、约5个至约10个、约10个至约15个、约15个至约20个、约20个至约25个、约25个至约50个、约50个至约100个氨基酸取代或缺失。因此,在一些实施方式中,主题变体衣壳蛋白包含与相应的亲本aav衣壳(例如seqidno:1-12中所示的野生型衣壳蛋白)具有85%或更高、90%或更高、95%或更高、或98%或更高例如或99%的序列同一性。
在进一步的实施方式中,所述一个或多个氨基酸取代位于如肽插入前编号的aav2vp1衣壳蛋白的一个或多个氨基酸残基1、15、34、57、66、81、101、109、144、164、176、188、196、226、236、240、250、312、363、368、449、456、463、472、484、524、535、551、593、698、708、719、721和/或735或另一种aav衣壳蛋白的相应的一个或多个氨基酸残基处。在一些这种实施方式中,所述一个或多个氨基酸取代选自由以下组成的组:如肽插入前编号的aav2vp1衣壳蛋白的m1l、l15p、p34a、n57d、n66k、r81q、q101r、s109t、r144k、r144m、q164k、t176p、l188i、s196y、g226e、g236v、i240t、p250s、n312k、p363l、d368h、n449d、t456k、s463y、d472n、r484c、a524t、p535s、n551s、a593e、i698v、v708i、v719m、s721l和l735q或另一种aav衣壳蛋白的相应的一个或多个氨基酸残基。
在优选的实施方式中,提供了一种变体aav衣壳蛋白,其包含a)在衣壳蛋白的gh-环中的插入肽,其中肽插入包含选自以下的氨基酸序列:isdqtkh(seqidno:14)、lgisdqtkha(seqidno:29)和laisdqtkha(seqidno:28),和b)与aav2的氨基酸序列(seqidno:2)相比以下氨基酸取代中的一个或多个或另一个aav亲本血清型(即除了aav2之外)的相应取代,其中取代的一个或多个氨基酸不天然存在于以下的相应位置:m1l、l15p、p34a、n57d、n66k、r81q、q101r、s109t、r144k、r144m、q164k、t176p、l188i、s196y、g226e、g236v、i240t、p250s、n312k、p363l、d368h、n449d、t456k、s463y、d472n、r484c、a524t、p535s、n551s、a593e、i698v、v708i、v719m、s721l、l735q和其组合。在一些实施方式中,所述一个或多个氨基酸取代选自由以下组成的组:m1l+l15p+p535s、p34a、p34a+s721l、n57d、n66k、r81q、q101r、s109t、r144k、r144m、q164k、q164k+v708i、t176p、l188i、s196y、g226e、g236v、i240t、n312k、n312k+n449d+d472n+n551s+i698v+l735q、p363l、r484c+v708i、t456k和v708i。优选地,肽插入位点位于aav2衣壳的氨基酸587与588之间或另一种aav血清型的衣壳蛋白中的相应位置。
在特别优选的实施方式中,变体aav衣壳包含肽插入,所述肽插入包含氨基酸序列isdqtkh(seqidno:14)或包含、基本上由以下组成、或由以下组成:在aav2的vp1的氨基酸587与588之间或另一个aav衣壳的相应氨基酸之间的氨基酸序列laisdqtkha(seqidno:28)或lgisdqtkha(seqidno:29),并且进一步包含在相对于aav2衣壳的氨基酸序列(seqidno:2)的残基34或另一种aav衣壳的相应残基处的p34a氨基酸取代。变体aav衣壳可以与seqidno:2所示的氨基酸序列或相应的亲本aav衣壳的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%、或更高的氨基酸序列同一性。在特别优选的实施方式中,变体aav衣壳具有氨基酸序列,所述氨基酸序列与以下氨基酸序列具有至少约85%、至少约90%、至少约95%、至少约98%序列同一性或与以下氨基酸序列具有100%同一性:
maadgylpdwledtlsegirqwwklkpgppppkaaerhkddsrglvlpgykylgpfngldkgepvneadaaalehdkaydrqldsgdnpylkynhadaefqerlkedtsfggnlgravfqakkrvleplglveepvktapgkkrpvehspvepdsssgtgkagqqparkrlnfgqtgdadsvpdpqplgqppaapsglgtntmatgsgapmadnnegadgvgnssgnwhcdstwmgdrvittstrtwalptynnhlykqissqsgasndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevtqndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmvpqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylsrtntpsgtttqsrlqfsqagasdirdqsrnwlpgpcyrqqrvsktsadnnnseyswtgatkyhlngrdslvnpgpamashkddeekffpqsgvlifgkqgsektnvdiekvmitdeeeirttnpvateqygsvstnlqrgnlaisdqtkharqaatadvntqgvlpgmvwqdrdvylqgpiwakiphtdghfhpsplmggfglkhpppqilikntpvpanpsttfsaakfasfitqystgqvsveiewelqkenskrwnpeiqytsnynksvnvdftvdtngvyseprpigtryltrnl(seqidno:42)
在另一个特别优选的实施方式中,变体aav衣壳包含肽插入,所述肽插入包含氨基酸序列isdqtkh(seqidno:14)或包含、基本上由以下组成、或由以下组成:在aav2衣壳蛋白的氨基酸587与588之间或另一个aav血清型的衣壳蛋白中的相应位置的氨基酸序列laisdqtkha(seqidno:28)或lgisdqtkha(seqidno:29),并且包含与aav2衣壳的氨基酸序列(seqidno:2)相比的n312k氨基酸取代或在另一种aav亲本血清型中的相应取代,并且任选地进一步包含与aav2衣壳的氨基酸序列或另一种aav亲本血清型的相应取代相比的n449d、d472n、n551s、i698v和/或l735q氨基酸取代。在另一个特别优选的实施方式中,变体aav衣壳包含肽插入,所述肽插入包含氨基酸序列isdqtkh(seqidno:14)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间或另一个aav血清型的衣壳蛋白中的相应位置的氨基酸序列laisdqtkha(seqidno:28)或lgisdqtkha(seqidno:29),并且包含与aav2衣壳的氨基酸序列(seqidno:2)相比的n312k、n449d、d472n、n551s、i698v和l735q氨基酸取代或在另一种aav亲本血清型中的相应残基的取代。变体aav衣壳可以与seqidno:2所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或更高的氨基酸序列同一性。在特别优选的实施方式中,变体aav衣壳具有氨基酸序列,所述氨基酸序列与以下氨基酸序列具有至少约85%、至少约90%、至少约95%、至少约98%序列同一性或与以下氨基酸序列具有100%同一性:
maadgylpdwledtlsegirqwwklkpgppppkpaerhkddsrglvlpgykylgpfngldkgepvneadaaalehdkaydrqldsgdnpylkynhadaefqerlkedtsfggnlgravfqakkrvleplglveepvktapgkkrpvehspvepdsssgtgkagqqparkrlnfgqtgdadsvpdpqplgqppaapsglgtntmatgsgapmadnnegadgvgnssgnwhcdstwmgdrvittstrtwalptynnhlykqissqsgasndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlkfklfniqvkevtqndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmvpqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylsrtdtpsgtttqsrlqfsqagasdirnqsrnwlpgpcyrqqrvsktsadnnnseyswtgatkyhlngrdslvnpgpamashkddeekffpqsgvlifgkqgsektsvdiekvmitdeeeirttnpvateqygsvstnlqrgnlaisdqtkharqaatadvntqgvlpgmvwqdrdvylqgpiwakiphtdghfhpsplmggfglkhpppqilikntpvpanpsttfsaakfasfitqystgqvsveiewelqkenskrwnpevqytsnynksvnvdftvdtngvyseprpigtryltrnq(seqidno:43)
在另一个实施方式中,提供了一种变体aav衣壳蛋白,其包含a)位于aav1、aav3a、aav3b、aav6或aav9的vp1的氨基酸588与589之间、aav4的氨基酸586与587之间、aav5的氨基酸577与578之间、aav7的氨基酸589与590之间、或aav8或aav10的氨基酸590至591之间的肽插入,肽插入包含选自以下的氨基酸序列:isdqtkh(seqidno:14)、lgisdqtkha(seqidno:29)和laisdqtkha(seqidno:28),和b)aav3a或aav3b的氨基酸709处的缬氨酸到异亮氨酸取代、aav1或aav6的位置709处的丙氨酸到异亮氨酸取代、aav4的氨基酸707处或aav9的氨基酸709处的天冬酰胺到异亮氨酸取代、或aav7的氨基酸710处或aav8或aav10的氨基酸711处的苏氨酸到异亮氨酸取代、或aav5的氨基酸697处的谷氨酰胺到异亮氨酸取代,并且任选地在其它方面与seqidno:1和3-12中的任一个一致。在优选的实施方式中,变体衣壳蛋白包含:a)肽插入,其包含氨基酸序列isdqtkh(seqidno:14)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列laisdqtkha(seqidno:28)或lgisdqtkha(seqidno:29)和b)与aav2的氨基酸序列相比在氨基酸708处的缬氨酸到异亮氨酸氨基酸取代,其中变体衣壳蛋白包含2个至5个、5个至10个、或10个至15个氨基酸取代。
在又另一个实施方式中,变体衣壳蛋白包含:a)肽插入,其包含氨基酸序列isdqtkh(seqidno:14)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列laisdqtkha(seqidno:28)或lgisdqtkha(seqidno:29)和b)与aav2的氨基酸序列相比氨基酸708处的缬氨酸到异亮氨酸氨基酸取代,并且在其它方面与氨基酸序列seqidno:2一致。
在又另一个实施方式中,变体衣壳蛋白包含:a)肽插入,其包含氨基酸序列isdqtkh(seqidno:14)或包含、基本上由以下组成、或由以下组成:aav2衣壳的氨基酸587与588之间的氨基酸序列laisdqtkha(seqidno:28)或lgisdqtkha(seqidno:29),并且在其它方面与氨基酸序列seqidno:2一致。在一些实施方式中,变体aav衣壳具有氨基酸序列,所述氨基酸序列与以下氨基酸序列具有至少约85%、至少约90%、至少约95%、至少约98%序列同一性或与以下氨基酸序列具有100%同一性:
maadgylpdwledtlsegirqwwklkpgppppkpaerhkddsrglvlpgykylgpfngldkgepvneadaaalehdkaydrqldsgdnpylkynhadaefqerlkedtsfggnlgravfqakkrvleplglveepvktapgkkrpvehspvepdsssgtgkagqqparkrlnfgqtgdadsvpdpqplgqppaapsglgtntmatgsgapmadnnegadgvgnssgnwhcdstwmgdrvittstrtwalptynnhlykqissqsgasndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevtqndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmvpqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylsrtntpsgtttqsrlqfsqagasdirdqsrnwlpgpcyrqqrvsktsadnnnseyswtgatkyhlngrdslvnpgpamashkddeekffpqsgvlifgkqgsektnvdiekvmitdeeeirttnpvateqygsvstnlqrgnlgisdqtkharqaatadvntqgvlpgmvwqdrdvylqgpiwakiphtdghfhpsplmggfglkhpppqilikntpvpanpsttfsaakfasfitqystgqvsveiewelqkenskrwnpeiqytsnynksvnvdftvdtngvyseprpigtryltrnl(seqidno:44)
在优选的实施方式中,提供了一种变体aav衣壳蛋白,其包含a)在衣壳蛋白的gh-环中的插入肽,其中肽插入包含选自以下的肽插入:qadttkn(seqidno:13)和laqadttkna(seqidno:27),和b)与aav2的氨基酸序列(seqidno:2)相比以下氨基酸取代中的一个或多个或在另一个aav亲本血清型(即除了aav2之外)中的相应取代,其中取代的一个或多个氨基酸不天然存在于相应的位置:m1l、l15p、p34a、n57d、n66k、r81q、q101r、s109t、r144k、r144m、q164k、t176p、l188i、s196y、g226e、g236v、i240t、p250s、n312k、p363l、d368h、n449d、t456k、s463y、d472n、r484c、a524t、p535s、n551s、a593e、i698v、v719m、s721l、l735q和其组合,优选地选自s109t、p250s、a524t、a593e、i698v、v708i和/或v719m。肽插入位点优选地位于aav2衣壳的氨基酸587与588之间或在另一种aav血清型的衣壳蛋白中的相应位置。在特别优选的实施方式中,变体aav衣壳包含肽插入,所述肽插入包含氨基酸序列qadttkn(seqidno:13)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间或在另一种aav血清型的衣壳蛋白中的相应位置的氨基酸序列laqadttkna(seqidno:27),并且包含与aav2的氨基酸序列相比的i698v氨基酸取代或在另一种aav亲本血清型中的相应取代,其中取代的一种或多种氨基酸不会自然地出现在相应的位置。变体aav衣壳可以与seqidno:2所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或更高的氨基酸序列同一性。在一些实施方式中,相应的氨基酸取代是与aav3a、aav3b或aav9衣壳的氨基酸序列相比的i699v氨基酸取代、与aav5衣壳的氨基酸序列相比的i687v取代、与aav7的氨基酸序列相比的i700v取代、与aav8或aav10的氨基酸序列相比的i701v取代。在特别优选的实施方式中,变体aav衣壳具有氨基酸序列,所述氨基酸序列与以下氨基酸序列具有至少约85%、至少约90%、至少约95%、至少约98%序列同一性或与以下氨基酸序列具有100%同一性:
maadgylpdwledtlsegirqwwklkpgppppkpaerhkddsrglvlpgykylgpfngldkgepvneadaaalehdkaydrqldsgdnpylkynhadaefqerlkedtsfggnlgravfqakkrvleplglveepvktapgkkrpvehspvepdsssgtgkagqqparkrlnfgqtgdadsvpdpqplgqppaapsglgtntmatgsgapmadnnegadgvgnssgnwhcdstwmgdrvittstrtwalptynnhlykqissqsgasndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevtqndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmvpqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylsrtntpsgtttqsrlqfsqagasdirdqsrnwlpgpcyrqqrvsktsadnnnseyswtgatkyhlngrdslvnpgpamashkddeekffpqsgvlifgkqgsektnvdiekvmitdeeeirttnpvateqygsvstnlqrgnlaqadttknarqaatadvntqgvlpgmvwqdrdvylqgpiwakiphtdghfhpsplmggfglkhpppqilikntpvpanpsttfsaakfasfitqystgqvsveiewelqkenskrwnpevqytsnynksvnvdftvdtngvyseprpigtryltrnl(seqidno:45)
在其它优选的实施方式中,变体aav衣壳包含肽插入,所述肽插入包含氨基酸序列qadttkn(seqidno:13)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间或在另一种aav血清型的衣壳蛋白中的相应位置的氨基酸序列laqadttkna(seqidno:27),并且包含与aav2的氨基酸序列相比的v719m氨基酸取代以及任选地v708i取代或在另一种aav亲本血清型中的相应取代,其中取代的一种或多种氨基酸不会自然地出现在相应的位置。
在另一个实施方式中,提供了一种变体aav衣壳蛋白,其包含a)位于aav1、aav3a、aav3b、aav6或aav9的vp1的氨基酸588与589之间、aav4的氨基酸586与587之间、aav5的氨基酸577与578之间、aav7的氨基酸589与590之间、或aav8或aav10的氨基酸590到591之间的肽插入,肽插入包含选自以下的氨基酸序列:qadttkn(seqidno:13)和laqadttkna(seqidno:27),和b)aav3a或aav3b的氨基酸709处的缬氨酸到异亮氨酸取代、aav1或aav6的位置709处的丙氨酸到异亮氨酸取代、aav4的氨基酸707处或aav9的氨基酸709处的天冬酰胺到异亮氨酸取代、或aav7的氨基酸710处或aav8或aav10的氨基酸711处的苏氨酸到异亮氨酸取代、或aav5的氨基酸697处的谷氨酰胺到异亮氨酸取代。在另一个实施方式中,提供了一种变体aav衣壳蛋白,其包含a)位于aav1、aav3a、aav3b、aav6或aav9的vp1的氨基酸588与589之间、aav4的氨基酸586与587之间、aav5的氨基酸577与578之间、aav7的氨基酸589与590之间、或aav8或aav10的氨基酸590到591之间的肽插入,肽插入包含选自以下的氨基酸序列:qadttkn(seqidno:13)和laqadttkna(seqidno:27),和b)与aav1、aav3a、aav3b、aav4、aav7、aav8、aav9或aav10的氨基酸序列相比位置109处或与aav5或aav6的氨基酸序列相比位置108处的丝氨酸到苏氨酸氨基酸取代。在优选的实施方式中,变体aav衣壳包含肽插入,其包含氨基酸序列qadttkn(seqidno:13)或包含、基本上由以下组成、或由以下组成:aav2衣壳的氨基酸587与588之间的氨基酸序列laqadttkna(seqidno:27),并且包含与aav2氨基酸序列相比氨基酸109处的丝氨酸到苏氨酸取代(s109t)或氨基酸708处的缬氨酸到异亮氨酸氨基酸取代(v708i),其中变体衣壳蛋白包含1个至5个、5个至10个、或10个至15个氨基酸取代,并且与seqidno:2所示的氨基酸序列的整个长度优选地具有至少约85%、至少约90%、至少约95%、至少约98%或更高的氨基酸序列同一性。在其它优选的实施方式中,变体aav衣壳包含肽插入,所述肽插入包含氨基酸序列qadttkn(seqidno:13)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间或在另一种aav血清型的衣壳蛋白中的相应位置的氨基酸序列laqadttkna(seqidno:27),并且包含与aav2的氨基酸序列相比氨基酸109处的丝氨酸到苏氨酸取代和氨基酸708处的缬氨酸到异亮氨酸氨基酸取代。
在又另一个实施方式中,变体衣壳蛋白包含:a)肽插入,其包含氨基酸序列qadttkn(seqidno:13)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列laqadttkna(seqidno:27)和b)至少一个氨基酸取代,其中变体衣壳的氨基酸序列不包含与aav2的氨基酸序列相比氨基酸708处的缬氨酸到异亮氨酸氨基酸取代,并且不包含与aav2的氨基酸序列相比氨基酸109处的丝氨酸到苏氨酸取代。
在又另一个实施方式中,变体衣壳蛋白包含:a)肽插入,其包含氨基酸序列qadttkn(seqidno:13)或包含、基本上由以下组成、或由以下组成:aav2衣壳的氨基酸587与588之间的氨基酸序列laqadttkna(seqidno:27),并且在其它方面与氨基酸序列seqidno:2一致。
在另一个优选的实施方式中,提供了一种变体aav衣壳蛋白,其包含a)在衣壳蛋白的gh-环中的插入肽,其中肽插入包含选自以下的肽插入:hditkni(seqidno:17)、iahditknia(seqidno:60)和lahditknia(seqidno:32),和b)与aav2的氨基酸序列(seqidno:2)相比以下氨基酸取代中的一个或多个或另一个aav亲本血清型(即除了aav2之外)的相应取代,其中取代的一个或多个氨基酸不天然存在于相应的位置:m1l、l15p、p34a、n57d、n66k、r81q、q101r、s109t、r144k、r144m、q164k、t176p、l188i、s196y、g226e、g236v、i240t、p250s、n312k、p363l、d368h、r389s、n449d、t456k、s463y、d472n、r484c、a524t、p535s、n551s、a593e、i698v、v708i、v719m、s721l、l735q和其组合。在一些实施方式中,aav衣壳蛋白包含选自s109t、r389s、a593e和/或v708i的一个或多个氨基酸取代。优选地,肽插入位点位于aav2衣壳的氨基酸587与588之间或另一种aav血清型的衣壳蛋白中的相应位置。在一个优选的实施方式中,变体aav衣壳包含肽插入,所述肽插入包含氨基酸序列hditkni(seqidno:17)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列iahditknia(seqidno:60)或lahditknia(seqidno:32),并且包含与aav2衣壳的氨基酸序列相比的s109t氨基酸取代或另一种aav亲本血清型中的相应取代。变体aav衣壳可以与seqidno:2所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或更高的氨基酸序列同一性。
在又另一个实施方式中,变体衣壳包含:a)肽插入,其包含氨基酸序列hditkni(seqidno:17)或包含、基本上由以下组成、或由以下组成:aav2衣壳的氨基酸587与588之间的氨基酸序列iahditknia(seqidno:60)或lahditknia(seqidno:32),并且在其它方面与seqidno:2中所述的氨基酸序列一致。在一些实施方式中,变体aav衣壳具有氨基酸序列,所述氨基酸序列与以下氨基酸序列具有至少约85%、至少约90%、至少约95%、至少约98%序列同一性或与以下氨基酸序列具有100%同一性:
maadgylpdwledtlsegirqwwklkpgppppkpaerhkddsrglvlpgykylgpfngldkgepvneadaaalehdkaydrqldsgdnpylkynhadaefqerlkedtsfggnlgravfqakkrvleplglveepvktapgkkrpvehspvepdsssgtgkagqqparkrlnfgqtgdadsvpdpqplgqppaapsglgtntmatgsgapmadnnegadgvgnssgnwhcdstwmgdrvittstrtwalptynnhlykqissqsgasndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevtqndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmvpqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylsrtntpsgtttqsrlqfsqagasdirdqsrnwlpgpcyrqqrvsktsadnnnseyswtgatkyhlngrdslvnpgpamashkddeekffpqsgvlifgkqgsektnvdiekvmitdeeeirttnpvateqygsvstnlqrgnlahditkniarqaatadvntqgvlpgmvwqdrdvylqgpiwakiphtdghfhpsplmggfglkhpppqilikntpvpanpsttfsaakfasfitqystgqvsveiewelqkenskrwnpeiqytsnynksvnvdftvdtngvyseprpigtryltrnl(seqidno:46)
在其它实施方式中,变体衣壳包含:a)肽插入,其包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列lahditknia,和b)至少一个氨基酸取代,其中变体衣壳的氨基酸序列不包含与aav2的氨基酸序列相比氨基酸708处的缬氨酸到异亮氨酸氨基酸取代。在又其它实施方式中,变体衣壳包含:a)肽插入,其包含氨基酸序列ditknia(seqidno:61)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列iahditknia(seqidno:60)或lahditknia(seqidno:32),和b)与aav2的氨基酸序列相比的v708i取代。在其它实施方式中,变体衣壳包含:a)肽插入,其包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列lahditknia(seqidno:32),和b)两个或更多个氨基酸取代,其中变体衣壳的氨基酸序列包含与aav2的氨基酸序列相比氨基酸708处的缬氨酸到异亮氨酸氨基酸取代。
在另一个优选的实施方式中,提供了一种变体aav衣壳蛋白,其包含a)在衣壳蛋白的gh-环中的插入肽,其中肽插入包含选自以下的肽插入:nqdytkt(seqidno:16)和lanqdytkta(seqidno:31),和b)与aav2的氨基酸序列(seqidno:2)相比以下氨基酸取代中的一个或多个或另一个aav亲本血清型(即除了aav2之外)中的相应取代,其中取代的一个或多个氨基酸不天然存在于相应的位置:m1l、l15p、p34a、n57d、n66k、r81q、q101r、s109t、r144k、r144m、q164k、t176p、l188i、s196y、g226e、g236v、i240t、p250s、p363l、d368h、n449d、t456k、s463y、d472n、r484c、a524t、p535s、n551s、a593e、i698v、v708i、v719m、s721l、l735q和其组合。在一些实施方式中,aav衣壳蛋白包含选自s109t、s109t+s463y、d368h和v708i的一个或多个氨基酸取代。优选地,肽插入位点位于aav2衣壳的氨基酸587与588之间或在另一种aav血清型的衣壳蛋白中的相应位置。在一个优选的实施方式中,变体aav衣壳包含肽插入,所述肽插入包含氨基酸序列nqdytkt(seqidno:16)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列lanqdytkta(seqidno:31),并且包含与aav2衣壳的氨基酸序列相比的v708i氨基酸取代或在另一种aav亲本血清型中的相应取代。变体aav衣壳可以与seqidno:2所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或更高的氨基酸序列同一性。在又另一个实施方式中,变体衣壳包含:a)肽插入,其包含氨基酸序列nqdytkt(seqidno:16)或包含、基本上由以下组成、或由以下组成:aav2衣壳的氨基酸587与588之间的氨基酸序列lanqdytkta(seqidno:31),并且在其它方面与seqidno:2中所述的氨基酸序列一致。在一些实施方式中,变体aav衣壳具有氨基酸序列,所述氨基酸序列与以下氨基酸序列具有至少约85%、至少约90%、至少约95%、至少约98%序列同一性或与以下氨基酸序列具有100%同一性:
maadgylpdwledtlsegirqwwklkpgppppkpaerhkddsrglvlpgykylgpfngldkgepvneadaaalehdkaydrqldsgdnpylkynhadaefqerlkedtsfggnlgravfqakkrvleplglveepvktapgkkrpvehspvepdsssgtgkagqqparkrlnfgqtgdadsvpdpqplgqppaapsglgtntmatgsgapmadnnegadgvgnssgnwhcdstwmgdrvittstrtwalptynnhlykqissqsgasndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevtqndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmvpqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylsrtntpsgtttqsrlqfsqagasdirdqsrnwlpgpcyrqqrvsktsadnnnseyswtgatkyhlngrdslvnpgpamashkddeekffpqsgvlifgkqgsektnvdiekvmitdeeeirttnpvateqygsvstnlqrgnlanqdytktarqaatadvntqgvlpgmvwqdrdvylqgpiwakiphtdghfhpsplmggfglkhpppqilikntpvpanpsttfsaakfasfitqystgqvsveiewelqkenskrwnpeiqytsnynksvnvdftvdtngvyseprpigtryltrnl(seqidno:47)
在其它实施方式中,变体衣壳包含:a)肽插入,其包含氨基酸序列nqdytkt(seqidno:16)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列lanqdytkta(seqidno:31)和b)与seqidno:2的序列相比的s109t氨基酸取代以及和任选地s463y氨基酸取代,其中变体衣壳与seqidno:2所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%同一性。在相关实施方式中,变体衣壳包含:a)肽插入,其包含氨基酸序列nqdytkt(seqidno:16)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列lanqdytkta(seqidno:31)和b)与氨基酸序列seqidno:2相比的s109t氨基酸取代,并且在其它方面与氨基酸序列seqidno:2一致。
在另一个实施方式中,提供了一种变体aav衣壳蛋白,其包含a)位于aav1、aav3a、aav3b、aav6或aav9的vp1的氨基酸588与589之间、aav4的氨基酸586与587之间、aav5的氨基酸577与578之间、aav7的氨基酸589与590之间、或aav8或aav10的氨基酸590到591之间的肽插入,肽插入包含选自以下的氨基酸序列:nqdytkt(seqidno:16)和lanqdytkta(seqidno:31)和b)与aav1或aav6的氨基酸序列相比位置313处或与aav9的氨基酸序列相比位置314处的天冬酰胺到赖氨酸氨基酸取代、或aav3a或aav3b的位置312处或aav8或aav10的位置315处的丝氨酸到赖氨酸取代、或aav4或aav5的位置303处或aav7的位置314处的精氨酸到赖氨酸取代。在另一个实施方式中,变体衣壳包含:a)肽插入,其包含氨基酸序列nqdytkt(seqidno:16)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列lanqdytkta(seqidno:31)和b)n312k氨基酸取代,其中变体衣壳蛋白包含1个至5个、5个至10个、或10个至15个氨基酸取代。
在另一个实施方式中,提供了一种变体aav衣壳蛋白,其包含a)在衣壳蛋白的gh-环中的插入肽,其中肽插入包含选自以下的肽插入:pnsthgs(seqidno:25)和lapnsthgsa(seqidno:40),和b)与aav2的氨基酸序列(seqidno:2)相比以下氨基酸取代中的一个或多个或在另一个aav亲本血清型(即除了aav2之外)中的相应取代,其中取代的一个或多个氨基酸不天然存在于相应的位置:m1l、l15p、p34a、n57d、n66k、r81q、q101r、s109t、r144k、r144m、q164k、t176p、l188i、s196y、g226e、g236v、i240t、p250s、n312k、p363l、d368h、n449d、t456k、s463y、d472n、r484c、a524t、p535s、n551s、a593e、i698v、v708i、v719m、s721l、l735q和其组合。优选地,肽插入位点位于aav2衣壳的氨基酸587与588之间或在另一种aav血清型的衣壳蛋白中的相应位置。在一个优选的实施方式中,变体aav衣壳包含肽插入,所述肽插入包含氨基酸序列pnsthgs(seqidno:25)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列lapnsthgsa(seqidno:40),并且包含与aav2衣壳的氨基酸序列相比的v708i氨基酸取代或在另一种aav亲本血清型中的相应取代。变体aav衣壳可以与seqidno:2所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或更高的氨基酸序列同一性。在又另一个实施方式中,变体衣壳包含:a)肽插入,其包含氨基酸序列pnsthgs(seqidno:25)或包含、基本上由以下组成、或由以下组成:aav2衣壳的氨基酸587与588之间的氨基酸序列lapnsthgsa(seqidno:40),并且在其它方面与seqidno:2中所述的氨基酸序列一致。在一些实施方式中,变体aav衣壳具有氨基酸序列,所述氨基酸序列与以下氨基酸序列具有至少约85%、至少约90%、至少约95%、至少约98%序列同一性或与以下氨基酸序列具有100%同一性:
maadgylpdwledtlsegirqwwklkpgppppkpaerhkddsrglvlpgykylgpfngldkgepvneadaaalehdkaydrqldsgdnpylkynhadaefqerlkedtsfggnlgravfqakkrvleplglveepvktapgkkrpvehspvepdsssgtgkagqqparkrlnfgqtgdadsvpdpqplgqppaapsglgtntmatgsgapmadnnegadgvgnssgnwhcdstwmgdrvittstrtwalptynnhlykqissqsgasndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevtqndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmvpqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylsrtntpsgtttqsrlqfsqagasdirdqsrnwlpgpcyrqqrvsktsadnnnseyswtgatkyhlngrdslvnpgpamashkddeekffpqsgvlifgkqgsektnvdiekvmitdeeeirttnpvateqygsvstnlqrgnlapnsthgsarqaatadvntqgvlpgmvwqdrdvylqgpiwakiphtdghfhpsplmggfglkhpppqilikntpvpanpsttfsaakfasfitqystgqvsveiewelqkenskrwnpeiqytsnynksvnvdftvdtngvyseprpigtryltrnl(seqidno:48)
在另一个实施方式中,提供了一种变体aav衣壳蛋白,其包含a)在衣壳蛋白的gh-环中的插入肽,其中肽插入包含选自以下的肽插入:nkttnkda(seqidno:62)和lankttnkda(seqidno:35),和b)与aav2的氨基酸序列(seqidno:2)相比以下氨基酸取代中的一个或多个或另一个aav亲本血清型(即除了aav2之外)的相应取代,其中取代的一个或多个氨基酸不天然存在于相应的位置:m1l、l15p、p34a、n57d、n66k、r81q、q101r、s109t、r144k、r144m、q164k、t176p、l188i、s196y、g226e、g236v、i240t、p250s、n312k、p363l、d368h、n449d、t456k、s463y、d472n、r484c、a524t、p535s、n551s、a593e、i698v、v708i、v719m、s721l、l735q和其组合。优选地,肽插入位点位于aav2衣壳的氨基酸587与588之间或另一种aav血清型的衣壳蛋白中的相应位置。在一个优选的实施方式中,变体aav衣壳包含肽插入,所述肽插入包含氨基酸序列nkttnkda(seqidno:62)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列lankttnkda(seqidno:35),并且包含与aav2衣壳的氨基酸序列相比的n449d氨基酸取代或在另一种aav亲本血清型中的相应取代。变体aav衣壳可以与seqidno:2所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或更高的氨基酸序列同一性。在又另一个实施方式中,变体衣壳包含:a)肽插入,其包含氨基酸序列nkttnkda(seqidno:62)或包含、基本上由以下组成、或由以下组成:aav2衣壳的氨基酸587与588之间的氨基酸序列lankttnkda(seqidno:35),并且在其它方面与seqidno:2中所述的氨基酸序列一致。在一些实施方式中,变体aav衣壳具有氨基酸序列,所述氨基酸序列与以下氨基酸序列具有至少约85%、至少约90%、至少约95%、至少约98%序列同一性或与以下氨基酸序列具有100%同一性:
maadgylpdwledtlsegirqwwklkpgppppkpaerhkddsrglvlpgykylgpfngldkgepvneadaaalehdkaydrqldsgdnpylkynhadaefqerlkedtsfggnlgravfqakkrvleplglveepvktapgkkrpvehspvepdsssgtgkagqqparkrlnfgqtgdadsvpdpqplgqppaapsglgtntmatgsgapmadnnegadgvgnssgnwhcdstwmgdrvittstrtwalptynnhlykqissqsgasndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevtqndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmvpqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylsrtntpsgtttqsrlqfsqagasdirdqsrnwlpgpcyrqqrvsktsadnnnseyswtgatkyhlngrdslvnpgpamashkddeekffpqsgvlifgkqgsektnvdiekvmitdeeeirttnpvateqygsvstnlqrgnlankttnkdarqaatadvntqgvlpgmvwqdrdvylqgpiwakiphtdghfhpsplmggfglkhpppqilikntpvpanpsttfsaakfasfitqystgqvsveiewelqkenskrwnpeiqytsnynksvnvdftvdtngvyseprpigtryltrnl(seqidno:49)
在另一个实施方式中,提供了一种变体aav衣壳蛋白,其包含a)在衣壳蛋白的gh-环中的插入肽,其中肽插入包含选自以下的肽插入:tnrtspd(seqidno:24)和latnrtspda(seqidno:39),和b)与aav2的氨基酸序列(seqidno:2)相比以下氨基酸取代中的一个或多个或在另一个aav亲本血清型(即除了aav2之外)中的相应取代,其中取代的一个或多个氨基酸不天然存在于相应的位置:m1l、l15p、p34a、n57d、n66k、r81q、q101r、s109t、r144k、r144m、q164k、t176p、l188i、s196y、g226e、g236v、i240t、p250s、n312k、p363l、d368h、n449d、t456k、s463y、d472n、r484c、a524t、p535s、n551s、a593e、i698v、v719m、s721l、l735q和其组合。优选地,肽插入位点位于aav2衣壳的氨基酸587与588之间或在另一种aav血清型的衣壳蛋白中的相应位置。在相关实施方式中,提供了一种变体aav衣壳蛋白,其包含a)位于aav1、aav3a、aav3b、aav6或aav9的vp1的氨基酸588与589之间、aav4的氨基酸586与587之间、aav5的氨基酸577与578之间、aav7的氨基酸589与590之间、或aav8或aav10的氨基酸590到591之间的肽插入,肽插入包含选自以下的氨基酸序列:tnrtspd(seqidno:24)和latnrtspda(seqidno:39),和b)aav3a或aav3b的氨基酸709处的缬氨酸到异亮氨酸取代、aav1或aav6的位置709处的丙氨酸到异亮氨酸取代、aav4的氨基酸707处或aav9的氨基酸709处的天冬酰胺到异亮氨酸取代、或aav7的氨基酸710处或aav8或aav10的氨基酸711处的苏氨酸到异亮氨酸取代、或aav5的氨基酸697处的谷氨酰胺到异亮氨酸取代。在其它实施方式中,变体衣壳蛋白包含:a)肽插入,其包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列latnrtspda(seqidno:39)和b)与aav2的氨基酸序列相比在氨基酸708处的缬氨酸到异亮氨酸氨基酸取代,其中变体衣壳蛋白包含1个至5个、5个至10个、或10个至15个氨基酸取代。在又另一个实施方式中,变体衣壳蛋白包含:a)肽插入,其包含在aav2衣壳的氨基酸587与588之间的氨基酸序列tnrtspd(seqidno:24)和b)与aav2的氨基酸序列相比在氨基酸708处的缬氨酸到异亮氨酸氨基酸取代。变体aav衣壳可以与seqidno:2所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或更高的氨基酸序列同一性。
在又另一个实施方式中,变体衣壳蛋白包含:a)肽插入,其包含氨基酸序列tnrtspd(seqidno:24)或包含、基本上由以下组成、或由以下组成:aav2衣壳的氨基酸587与588之间的氨基酸序列latnrtspda(seqidno:39),并且在其它方面与氨基酸序列seqidno:2一致。在一些实施方式中,变体aav衣壳具有氨基酸序列,所述氨基酸序列与以下氨基酸序列具有至少约85%、至少约90%、至少约95%、至少约98%序列同一性或与以下氨基酸序列具有100%同一性:
maadgylpdwledtlsegirqwwklkpgppppkpaerhkddsrglvlpgykylgpfngldkgepvneadaaalehdkaydrqldsgdnpylkynhadaefqerlkedtsfggnlgravfqakkrvleplglveepvktapgkkrpvehspvepdsssgtgkagqqparkrlnfgqtgdadsvpdpqplgqppaapsglgtntmatgsgapmadnnegadgvgnssgnwhcdstwmgdrvittstrtwalptynnhlykqissqsgasndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevtqndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmvpqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylsrtntpsgtttqsrlqfsqagasdirdqsrnwlpgpcyrqqrvsktsadnnnseyswtgatkyhlngrdslvnpgpamashkddeekffpqsgvlifgkqgsektnvdiekvmitdeeeirttnpvateqygsvstnlqrgnlatnrtspdarqaatadvntqgvlpgmvwqdrdvylqgpiwakiphtdghfhpsplmggfglkhpppqilikntpvpanpsttfsaakfasfitqystgqvsveiewelqkenskrwnpeiqytsnynksvnvdftvdtngvyseprpigtryltrnl(seqidno:50)
在另一个实施方式中,提供了一种变体aav衣壳蛋白,其包含a)在衣壳蛋白的gh-环中的插入肽,其中肽插入包含选自以下的肽插入:gkskvid(seqidno:23)和lagkskvida(seqidno:38),和b)与aav2的氨基酸序列(seqidno:2)相比以下氨基酸取代中的一个或多个或在另一个aav亲本血清型(即除了aav2之外)中的相应取代,其中取代的一个或多个氨基酸不天然存在于相应的位置:m1l、l15p、p34a、n57d、n66k、r81q、q101r、s109t、r144k、r144m、q164k、t176p、l188i、s196y、g226e、g236v、i240t、p250s、n312k、p363l、d368h、n449d、t456k、s463y、d472n、r484c、a524t、p535s、n551s、a593e、i698v、v708i、v719m、s721l、l735q和其组合。优选地,肽插入位点位于aav2衣壳的氨基酸587与588之间或在另一种aav血清型的衣壳蛋白中的相应位置。变体aav衣壳可以与seqidno:2所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或更高的氨基酸序列同一性。在一些实施方式中,变体aav衣壳包含位于aav2衣壳的氨基酸587与588之间的肽插入,所述肽插入包含氨基酸序列gkskvid(seqidno:23)或包含、基本上由以下组成、或由以下组成:氨基酸序列lagkskvida(seqidno:38),并且在其它方面与氨基酸序列seqidno:2一致。在其它实施方式中,变体aav衣壳包含:a)肽插入,其包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列lagkskvida(seqidno:38),并且包含至少一个氨基酸取代。
在一些实施方式中,变体aav衣壳具有氨基酸序列,所述氨基酸序列与以下氨基酸序列具有至少约85%、至少约90%、至少约95%、至少约98%序列同一性或与以下氨基酸序列具有100%同一性:
maadgylpdwledtlsegirqwwklkpgppppkpaerhkddsrglvlpgykylgpfngldkgepvneadaaalehdkaydrqldsgdnpylkynhadaefqerlkedtsfggnlgravfqakkrvleplglveepvktapgkkrpvehspvepdsssgtgkagqqparkrlnfgqtgdadsvpdpqplgqppaapsglgtntmatgsgapmadnnegadgvgnssgnwhcdstwmgdrvittstrtwalptynnhlykqissqsgasndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevtqndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmvpqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylsrtntpsgtttqsrlqfsqagasdirdqsrnwlpgpcyrqqrvsktsadnnnseyswtgatkyhlngrdslvnpgpamashkddeekffpqsgvlifgkqgsektnvdiekvmitdeeeirttnpvateqygsvstnlqrgnlagkskvidarqaatadvntqgvlpgmvwqdrdvylqgpiwakiphtdghfhpsplmggfglkhpppqilikntpvpanpsttfsaakfasfitqystgqvsveiewelqkenskrwnpeiqytsnynksvnvdftvdtngvyseprpigtryltrnl(seqidno:51)
在另一个实施方式中,提供了一种变体aav衣壳蛋白,其包含a)在衣壳蛋白的gh-环中的插入肽,其中肽插入包含选自以下的肽插入:asdstka(seqidno:15)和laasdstkaa(seqidno:30),和b)与aav2的氨基酸序列(seqidno:2)相比以下氨基酸取代中的一个或多个或在另一个aav亲本血清型(即除了aav2之外)中的相应取代,其中取代的一个或多个氨基酸不天然存在于相应的位置:m1l、l15p、p34a、n57d、n66k、r81q、q101r、s109t、r144k、r144m、q164k、t176p、l188i、s196y、g226e、g236v、i240t、p250s、n312k、p363l、d368h、n449d、t456k、s463y、d472n、r484c、a524t、p535s、n551s、a593e、i698v、v708i、v719m、s721l、l735q和其组合。优选地,肽插入位点位于aav2衣壳的氨基酸587与588之间或在另一种aav血清型的衣壳蛋白中的相应位置。变体aav衣壳可以与seqidno:2所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或更高的氨基酸序列同一性。在又另一个实施方式中,变体衣壳包含肽插入,其包含氨基酸序列asdstka(seqidno:15)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列laasdstkaa(seqidno:30),并且在其它方面与seqidno:2中所述的氨基酸序列一致。在一些实施方式中,变体aav衣壳具有氨基酸序列,所述氨基酸序列与以下氨基酸序列具有至少约85%、至少约90%、至少约95%、至少约98%序列同一性或与以下氨基酸序列具有100%同一性:
maadgylpdwledtlsegirqwwklkpgppppkpaerhkddsrglvlpgykylgpfngldkgepvneadaaalehdkaydrqldsgdnpylkynhadaefqerlkedtsfggnlgravfqakkrvleplglveepvktapgkkrpvehspvepdsssgtgkagqqparkrlnfgqtgdadsvpdpqplgqppaapsglgtntmatgsgapmadnnegadgvgnssgnwhcdstwmgdrvittstrtwalptynnhlykqissqsgasndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevtqndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmvpqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylsrtntpsgtttqsrlqfsqagasdirdqsrnwlpgpcyrqqrvsktsadnnnseyswtgatkyhlngrdslvnpgpamashkddeekffpqsgvlifgkqgsektnvdiekvmitdeeeirttnpvateqygsvstnlqrgnlaasdstkaarqaatadvntqgvlpgmvwqdrdvylqgpiwakiphtdghfhpsplmggfglkhpppqilikntpvpanpsttfsaakfasfitqystgqvsveiewelqkenskrwnpeiqytsnynksvnvdftvdtngvyseprpigtryltrnl(seqidno:52)
在另一个实施方式中,提供了一种变体aav衣壳蛋白,其包含a)在衣壳蛋白的gh-环中的插入肽,其中肽插入包含选自以下的肽插入:kdrapst(seqidno:26)和lakdraptsa(seqidno:41),和b)与aav2的氨基酸序列(seqidno:2)相比以下氨基酸取代中的一个或多个或在另一个aav亲本血清型(即除了aav2之外)中的相应取代,其中取代的一个或多个氨基酸不天然存在于相应的位置:m1l、l15p、p34a、n57d、n66k、r81q、q101r、s109t、r144k、r144m、q164k、t176p、l188i、s196y、g226e、g236v、i240t、p250s、n312k、p363l、d368h、n449d、t456k、s463y、d472n、r484c、a524t、p535s、n551s、a593e、i698v、v708i、v719m、s721l、l735q和其组合。优选地,肽插入位点位于aav2衣壳的氨基酸587与588之间或在另一种aav血清型的衣壳蛋白中的相应位置。变体aav衣壳可以与seqidno:2所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或更高的氨基酸序列同一性。在又另一个实施方式中,变体衣壳包含肽插入,其包含氨基酸序列kdrapst(seqidno:26)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列lakdraptsa(seqidno:41),并且在其它方面与seqidno:2中所述的氨基酸序列一致。在一些实施方式中,变体aav衣壳具有氨基酸序列,所述氨基酸序列与以下氨基酸序列具有至少约85%、至少约90%、至少约95%、至少约98%序列同一性或与以下氨基酸序列具有100%同一性:
maadgylpdwledtlsegirqwwklkpgppppkpaerhkddsrglvlpgykylgpfngldkgepvneadaaalehdkaydrqldsgdnpylkynhadaefqerlkedtsfggnlgravfqakkrvleplglveepvktapgkkrpvehspvepdsssgtgkagqqparkrlnfgqtgdadsvpdpqplgqppaapsglgtntmatgsgapmadnnegadgvgnssgnwhcdstwmgdrvittstrtwalptynnhlykqissqsgasndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevtqndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmvpqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylsrtntpsgtttqsrlqfsqagasdirdqsrnwlpgpcyrqqrvsktsadnnnseyswtgatkyhlngrdslvnpgpamashkddeekffpqsgvlifgkqgsektnvdiekvmitdeeeirttnpvateqygsvstnlqrgnlakdrapstarqaatadvntqgvlpgmvwqdrdvylqgpiwakiphtdghfhpsplmggfglkhpppqilikntpvpanpsttfsaakfasfitqystgqvsveiewelqkenskrwnpeiqytsnynksvnvdftvdtngvyseprpigtryltrnl(seqidno:53)
在另一个实施方式中,提供了一种变体aav衣壳蛋白,其包含a)在衣壳蛋白的gh-环中的插入肽,其中肽插入包含选自以下的肽插入:hqdttkn(seqidno:19)和lahqdttkna(seqidno:34),和b)与aav2的氨基酸序列(seqidno:2)相比以下氨基酸取代中的一个或多个或在另一个aav亲本血清型(即除了aav2之外)中的相应取代,其中取代的一个或多个氨基酸不天然存在于相应的位置中的一个或多个位置:m1l、l15p、p34a、n57d、n66k、r81q、q101r、s109t、r144k、r144m、q164k、t176p、l188i、s196y、g226e、g236v、i240t、p250s、n312k、p363l、d368h、n449d、t456k、s463y、d472n、r484c、a524t、p535s、n551s、a593e、i698v、v708i、v719m、s721l、l735q和其组合。优选地,肽插入位点位于aav2衣壳的氨基酸587与588之间或另一种aav血清型的衣壳蛋白中的相应位置。变体aav衣壳可以与seqidno:2所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或更高的氨基酸序列同一性。在又另一个实施方式中,变体衣壳包含肽插入,其包含氨基酸序列hqdttkn(seqidno:19)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列lahqdttkna(seqidno:34),并且在其它方面与seqidno:2中所述的氨基酸序列一致。在一些实施方式中,变体aav衣壳具有氨基酸序列,所述氨基酸序列与以下氨基酸序列具有至少约85%、至少约90%、至少约95%、至少约98%序列同一性或与以下氨基酸序列具有100%同一性:
maadgylpdwledtlsegirqwwklkpgppppkpaerhkddsrglvlpgykylgpfngldkgepvneadaaalehdkaydrqldsgdnpylkynhadaefqerlkedtsfggnlgravfqakkrvleplglveepvktapgkkrpvehspvepdsssgtgkagqqparkrlnfgqtgdadsvpdpqplgqppaapsglgtntmatgsgapmadnnegadgvgnssgnwhcdstwmgdrvittstrtwalptynnhlykqissqsgasndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevtqndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmvpqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylsrtntpsgtttqsrlqfsqagasdirdqsrnwlpgpcyrqqrvsktsadnnnseyswtgatkyhlngrdslvnpgpamashkddeekffpqsgvlifgkqgsektnvdiekvmitdeeeirttnpvateqygsvstnlqrgnlahqdttknarqaatadvntqgvlpgmvwqdrdvylqgpiwakiphtdghfhpsplmggfglkhpppqilikntpvpanpsttfsaakfasfitqystgqvsveiewelqkenskrwnpeiqytsnynksvnvdftvdtngvyseprpigtryltrnl(seqidno:54)
在另一个实施方式中,提供了一种变体aav衣壳蛋白,其包含a)在衣壳蛋白的gh-环中的插入肽,其中肽插入包含选自以下的肽插入:isneneh(seqidno:21)和lpisneneha(seqidno:36),和b)与aav2的氨基酸序列(seqidno:2)相比以下氨基酸取代中的一个或多个或在另一个aav亲本血清型(即除了aav2之外)中的相应取代,其中取代的一个或多个氨基酸不天然存在于相应的位置中的一个或多个位置:m1l、l15p、p34a、n57d、n66k、r81q、q101r、s109t、r144k、r144m、q164k、t176p、l188i、s196y、g226e、g236v、i240t、p250s、n312k、p363l、d368h、n449d、t456k、s463y、d472n、r484c、a524t、p535s、n551s、a593e、i698v、v708i、v719m、s721l、l735q和其组合。优选地,肽插入位点位于aav2衣壳的氨基酸587与588之间或在另一种aav血清型的衣壳蛋白中的相应位置。变体aav衣壳可以与seqidno:2所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或更高的氨基酸序列同一性。在又另一个实施方式中,变体衣壳包含肽插入,其包含氨基酸序列isneneh(seqidno:21)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列lpisneneha(seqidno:36),并且在其它方面与seqidno:2中所述的氨基酸序列一致。在一些实施方式中,变体aav衣壳具有氨基酸序列,所述氨基酸序列与以下氨基酸序列具有至少约85%、至少约90%、至少约95%、至少约98%序列同一性或与以下氨基酸序列具有100%同一性:
maadgylpdwledtlsegirqwwklkpgppppkpaerhkddsrglvlpgykylgpfngldkgepvneadaaalehdkaydrqldsgdnpylkynhadaefqerlkedtsfggnlgravfqakkrvleplglveepvktapgkkrpvehspvepdsssgtgkagqqparkrlnfgqtgdadsvpdpqplgqppaapsglgtntmatgsgapmadnnegadgvgnssgnwhcdstwmgdrvittstrtwalptynnhlykqissqsgasndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevtqndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmvpqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylsrtntpsgtttqsrlqfsqagasdirdqsrnwlpgpcyrqqrvsktsadnnnseyswtgatkyhlngrdslvnpgpamashkddeekffpqsgvlifgkqgsektnvdiekvmitdeeeirttnpvateqygsvstnlqrgnlpisneneharqaatadvntqgvlpgmvwqdrdvylqgpiwakiphtdghfhpsplmggfglkhpppqilikntpvpanpsttfsaakfasfitqystgqvsveiewelqkenskrwnpeiqytsnynksvnvdftvdtngvyseprpigtryltrnl(seqidno:55)
在另一个实施方式中,提供了一种变体aav衣壳蛋白,其包含a)在衣壳蛋白的gh-环中的肽插入,其中肽插入包含选自以下的肽插入:qananen(seqidno:22)和lpqananena(seqidno:37),和b)与aav2的氨基酸序列(seqidno:2)相比以下氨基酸取代中的一个或多个或在另一个aav亲本血清型(即除了aav2之外)中的相应取代,其中取代的一个或多个氨基酸不天然存在于相应的位置:m1l、l15p、p34a、n57d、n66k、r81q、q101r、s109t、r144k、r144m、q164k、t176p、l188i、s196y、g226e、g236v、i240t、p250s、n312k、p363l、d368h、n449d、t456k、s463y、d472n、r484c、a524t、p535s、n551s、a593e、i698v、v708i、v719m、s721l、l735q和其组合。优选地,肽插入位点位于aav2衣壳的氨基酸587与588之间或在另一种aav血清型的衣壳蛋白中的相应位置。变体aav衣壳可以与seqidno:2所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或更高的氨基酸序列同一性。在又另一个实施方式中,变体衣壳包含肽插入,其包含氨基酸序列qananen(seqidno:22)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列lpqananena(seqidno:37),并且在其它方面与seqidno:2中所述的氨基酸序列一致。在一些实施方式中,变体aav衣壳具有氨基酸序列,所述氨基酸序列与以下氨基酸序列具有至少约85%、至少约90%、至少约95%、至少约98%序列同一性或与以下氨基酸序列具有100%同一性:
maadgylpdwledtlsegirqwwklkpgppppkpaerhkddsrglvlpgykylgpfngldkgepvneadaaalehdkaydrqldsgdnpylkynhadaefqerlkedtsfggnlgravfqakkrvleplglveepvktapgkkrpvehspvepdsssgtgkagqqparkrlnfgqtgdadsvpdpqplgqppaapsglgtntmatgsgapmadnnegadgvgnssgnwhcdstwmgdrvittstrtwalptynnhlykqissqsgasndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevtqndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmvpqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylsrtntpsgtttqsrlqfsqagasdirdqsrnwlpgpcyrqqrvsktsadnnnseyswtgatkyhlngrdslvnpgpamashkddeekffpqsgvlifgkqgsektnvdiekvmitdeeeirttnpvateqygsvstnlqrgnlpqananenarqaatadvntqgvlpgmvwqdrdvylqgpiwakiphtdghfhpsplmggfglkhpppqilikntpvpanpsttfsaakfasfitqystgqvsveiewelqkenskrwnpeiqytsnynksvnvdftvdtngvyseprpigtryltrnl(seqidno:56)
在另一个实施方式中,提供了一种变体aav衣壳蛋白,其包含a)在衣壳蛋白的gh-环中的插入肽,其中肽插入包含选自以下的肽插入:hpdttkn(seqidno:18)和lahpdttkna(seqidno:33),和b)与aav2的氨基酸序列(seqidno:2)相比以下氨基酸取代中的一个或多个或在另一个aav亲本血清型(即除了aav2之外)中的相应取代,其中取代的一个或多个氨基酸不天然存在于相应的位置:m1l、l15p、p34a、n57d、n66k、r81q、q101r、s109t、r144k、r144m、q164k、t176p、l188i、s196y、g226e、g236v、i240t、p250s、n312k、p363l、d368h、n449d、t456k、s463y、d472n、r484c、a524t、p535s、n551s、a593e、i698v、v708i、v719m、s721l、l735q和其组合。优选地,肽插入位点位于aav2衣壳的氨基酸587与588之间或在另一种aav血清型的衣壳蛋白中的相应位置。变体aav衣壳可以与seqidno:2所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或更高的氨基酸序列同一性。在又另一个实施方式中,变体衣壳包含肽插入,其包含氨基酸序列hpdttkn(seqidno:18)或包含、基本上由以下组成、或由以下组成:在aav2衣壳的氨基酸587与588之间的氨基酸序列lahpdttkna(seqidno:33),并且在其它方面与seqidno:2中所述的氨基酸序列一致。在一些实施方式中,变体aav衣壳具有氨基酸序列,所述氨基酸序列与以下氨基酸序列具有至少约85%、至少约90%、至少约95%、至少约98%序列同一性或与以下氨基酸序列具有100%同一性:
maadgylpdwledtlsegirqwwklkpgppppkpaerhkddsrglvlpgykylgpfngldkgepvneadaaalehdkaydrqldsgdnpylkynhadaefqerlkedtsfggnlgravfqakkrvleplglveepvktapgkkrpvehspvepdsssgtgkagqqparkrlnfgqtgdadsvpdpqplgqppaapsglgtntmatgsgapmadnnegadgvgnssgnwhcdstwmgdrvittstrtwalptynnhlykqissqsgasndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevtqndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmvpqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylsrtntpsgtttqsrlqfsqagasdirdqsrnwlpgpcyrqqrvsktsadnnnseyswtgatkyhlngrdslvnpgpamashkddeekffpqsgvlifgkqgsektnvdiekvmitdeeeirttnpvateqygsvstnlqrgnlahpdttknarqaatadvntqgvlpgmvwqdrdvylqgpiwakiphtdghfhpsplmggfglkhpppqilikntpvpanpsttfsaakfasfitqystgqvsveiewelqkenskrwnpeiqytsnynksvnvdftvdtngvyseprpigtryltrnl(seqidno:57).
在若干方面,提供了一种变体aav衣壳蛋白,其包含相对于相应的亲本aav衣壳蛋白的一个或多个氨基酸取代,其中当存在于aav病毒粒子中时,与包含相应亲本aav衣壳蛋白的aav病毒粒子对视网膜细胞的感染性相比,所述变体aav衣壳蛋白赋予对视网膜细胞增加的感染性。
在一些实施方式中,变体aav衣壳蛋白包含与aav2衣壳的氨基酸序列(seqidno:2)相比的p34a氨基酸取代或与aav5衣壳的氨基酸序列(seqidno:6)相比的p33a氨基酸取代。在一些优选的实施方式中,变体衣壳蛋白包含氨基酸序列,所述氨基酸序列与seqidno:2或seqidno:6中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更多氨基酸序列同一性,并且包含分别与aav2或aav5衣壳的氨基酸序列相比p34a或p33a氨基酸取代。在一些优选的实施方式中,变体衣壳蛋白包含氨基酸序列,所述氨基酸序列包含与seqidno2中所示的氨基酸序列相比的p34a氨基酸取代,并且在其它方面与seqidno:2中所示的氨基酸序列一致。在相关实施方式中,变体衣壳蛋白包含与氨基酸序列seqidno:2相比的p34a氨基酸取代,其中变体衣壳蛋白包含与seqidno:2中所示的aav2衣壳蛋白的氨基酸序列相比的1个至5个、5个至10个、或10个至15个氨基酸取代。
在其它实施方式中,变体aav衣壳蛋白包含与aav2衣壳的氨基酸序列(seqidno:2)相比氨基酸164或在另一种aav亲本血清型(即除了aav2之外)中的相应位置处的氨基酸取代,其中取代的氨基酸不会自然地出现在相应的位置。在一些优选的实施方式中,变体衣壳蛋白包含氨基酸序列,所述氨基酸序列与seqidno:2中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更多氨基酸序列同一性,并且包含与aav2衣壳的氨基酸序列(seqidno:2)相比氨基酸164处的氨基酸取代。在一些实施方式中,raav病毒粒子包含与aav1、aav2或aav6的氨基酸序列相比氨基酸164处或与aav7、aav8或aav10的氨基酸序列相比氨基酸165处的谷氨酰胺到赖氨酸氨基酸取代;或包含在aav5的氨基酸160处的丝氨酸到赖氨酸取代或在aav9的氨基酸164处的丙氨酸到赖氨酸取代。在相关实施方式中,变体衣壳蛋白包含与aav2衣壳的氨基酸序列(seqidno:2)相比的氨基酸164(例如,q164k)处的氨基酸取代,其中变体衣壳蛋白包含与seqidno:2中所示的aav2衣壳蛋白的氨基酸序列相比的1个至5个、5个至10个、或10个至15个氨基酸取代。在一些优选的实施方式中,变体衣壳蛋白包含氨基酸序列,所述氨基酸序列包含与seqidno:2中所示的氨基酸序列相比的q164k氨基酸取代,并且在其它方面与seqidno:2中所示的氨基酸序列一致。在其它实施方式中,变体衣壳蛋白包含与aav2衣壳的氨基酸序列(seqidno:2)相比的q164k和v708i氨基酸取代或在另一种aav亲本血清型(即除aav2之外)中的相应取代,并且与seqidno:2中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更高的氨基酸序列同一性。
在其它实施方式中,变体aav衣壳蛋白包含与aav2衣壳的氨基酸序列(seqidno:2)相比氨基酸698或在另一种aav亲本血清型(即除了aav2之外)中的相应位置处的氨基酸取代,其中取代的氨基酸不会自然地出现在相应的位置。在一些优选的实施方式中,变体衣壳蛋白包含氨基酸序列,所述氨基酸序列与seqidno:2中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更多氨基酸序列同一性,并且包含与aav2衣壳的氨基酸序列(seqidno:2)相比氨基酸698处的氨基酸取代。在一些实施方式中,raav病毒粒子包含与aav2的氨基酸序列相比氨基酸698处、或与aav3a、aav3b或aav9的氨基酸序列相比氨基酸699处、或aav5的氨基酸687处、或aav7的氨基酸700处、或aav8或aav10的氨基酸701处的异亮氨酸到缬氨酸氨基酸取代。在相关实施方式中,变体衣壳蛋白包含与aav2衣壳的氨基酸序列(seqidno:2)相比的氨基酸699(例如,i698v)处的氨基酸取代,其中变体衣壳蛋白包含与seqidno:2中所示的aav2衣壳蛋白的氨基酸序列相比的1个至5个、5个至10个、或10个至15个氨基酸取代。在一些优选的实施方式中,变体衣壳蛋白包含氨基酸序列,所述氨基酸序列包含与seqidno:2中所示的氨基酸序列相比的i698v氨基酸取代,并且在其它方面与seqidno:2中所示的氨基酸序列一致。
在其它实施方式中,变体aav衣壳蛋白包含与aav2衣壳的氨基酸序列(seqidno:2)相比氨基酸109或在另一种aav亲本血清型(即除了aav2之外)中的相应位置处的氨基酸取代。在一些优选的实施方式中,变体衣壳蛋白包含氨基酸序列,所述氨基酸序列与seqidno:2中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更多氨基酸序列同一性,并且包含与aav2衣壳的氨基酸序列(seqidno:2)相比氨基酸109处的氨基酸取代。在一些实施方式中,变体衣壳蛋白包含与aav1、aav3a、aav3b、aav4、aav7、aav8、aav9或aav10的氨基酸序列相比位置109处或与aav5或aav6的氨基酸序列相比位置108处的丝氨酸到苏氨酸氨基酸取代。在相关实施方式中,变体衣壳蛋白包含与氨基酸序列aav2相比的s109t氨基酸取代,其中变体衣壳蛋白包含1个至5个、5个至10个、或10个至15个氨基酸取代。在其它相关实施方式中,变体衣壳蛋白包含与aav2的氨基酸序列相比的s109t氨基酸取代和a593e氨基酸取代。在一些实施方式中,变体衣壳蛋白包含与aav2衣壳的氨基酸序列(seqidno:2)相比的s109t和a493v以及任选地a593e和/或v708i氨基酸取代或在另一种aav亲本血清型(即除aav2之外)中的相应取代,并且与seqidno:2中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更高的氨基酸序列同一性。在一些优选的实施方式中,变体衣壳蛋白包含与aav2衣壳的氨基酸序列(seqidno:2)相比的s109t、a493v、a593e和v708i氨基酸取代或另一种aav亲本血清型(即除aav2之外)的相应取代,并且与seqidno:2中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更高的氨基酸序列同一性。在其它优选的实施方式中,变体衣壳蛋白包含与aav2的氨基酸序列相比的s109t和v708i氨基酸取代,并且与seqidno:2中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更多氨基酸序列同一性,并且在其它方面与氨基酸序列seqidno:2一致。
在其它实施方式中,变体aav衣壳蛋白包含与aav2衣壳的氨基酸序列(seqidno:2)相比氨基酸593或在另一种aav亲本血清型(即除了aav2之外)中的相应位置处的氨基酸取代。在一些优选的实施方式中,变体衣壳蛋白包含氨基酸序列,所述氨基酸序列与seqidno:2中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更多氨基酸序列同一性,并且包含与aav2衣壳的氨基酸序列(seqidno:2)相比氨基酸593处的氨基酸取代。在一些实施方式中,变体衣壳蛋白包含与aav1、aav3a、aav6或aav9的氨基酸序列相比氨基酸594处、aav5的氨基酸583处或aav8或aav10的氨基酸596处的甘氨酸到谷氨酸氨基酸取代、或aav3b的氨基酸594处的精氨酸到谷氨酸氨基酸取代、或aav4的氨基酸592处的天冬氨酸到谷氨酸氨基酸取代或aav7的位置595处的谷氨酰胺到谷氨酸氨基酸取代。在其它实施方式中,变体衣壳蛋白包含与aav2的氨基酸序列相比的a593e氨基酸取代,并且不包含与aav2的氨基酸序列相比的以下氨基酸取代中的一个或多个:i19v、v369a、k26r、n215d、g355s、v46a和s196p。在相关实施方式中,变体衣壳蛋白包含与aav2的氨基酸序列相比的a593e和n596d氨基酸取代,并且与seqidno:2中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%的同一性。在其它实施方式中,变体衣壳包含与aav2的氨基酸序列相比的a593e和n596d氨基酸取代,并且在其它方面与aav2的氨基酸序列一致。在其它实施方式中,变体衣壳蛋白包含与aav2的氨基酸序列相比的a593e和v708i氨基酸取代,并且与seqidno:2中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%的同一性。在其它实施方式中,变体衣壳包含与aav2的氨基酸序列相比的a593e和v708i氨基酸取代,并且在其它方面与aav2的氨基酸序列一致。
在其它实施方式中,变体aav衣壳蛋白包含与aav2衣壳的氨基酸序列(seqidno:2)相比氨基酸708或在另一种aav亲本血清型(即除了aav2之外)中的相应位置处的氨基酸取代,其中取代的氨基酸不会自然地出现在相应的位置。优选地,raav病毒粒子不包含与aav2相比的氨基酸250或在另一种aav亲本血清型中的相应氨基酸处的脯氨酸到丝氨酸取代。在一些实施方式中,变体衣壳蛋白包含氨基酸序列,所述氨基酸序列与seqidno:2中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更多氨基酸序列同一性,并且包含与aav2衣壳的氨基酸序列(seqidno:2)相比氨基酸708处的氨基酸取代。在优选的实施方式中,变体衣壳蛋白包含与aav2衣壳的氨基酸序列相比的氨基酸708处的缬氨酸到异亮氨酸(v708i)取代,并且与seqidno:2中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更多氨基酸序列同一性,并且在其它方面与氨基酸序列seqidno:2一致,其中变体衣壳蛋白不包含p250s氨基酸取代。在一些实施方式中,变体衣壳蛋白包含aav3a或aav3b的氨基酸709处的缬氨酸到异亮氨酸取代、aav1或aav6的位置709处的丙氨酸到异亮氨酸取代、aav4的氨基酸707处或aav9的氨基酸709处的天冬酰胺到异亮氨酸取代、或aav7的氨基酸710处或aav8或aav10的氨基酸711处的苏氨酸到异亮氨酸取代、或aav5的氨基酸697处的谷氨酰胺到异亮氨酸取代。在相关实施方式中,变体衣壳蛋白包含与aav2的氨基酸序列相比的v708i氨基酸取代,其中变体衣壳蛋白包含2个至5个、5个至10个、或10个至15个氨基酸取代,并且其中变体衣壳蛋白不包含p250s氨基酸取代。在其它实施方式中,变体衣壳蛋白包含v708i氨基酸取代,并且还包含与aav2的氨基酸序列相比的a593e和/或s109t氨基酸取代。在其它相关实施方式中,变体衣壳包含与aav2的氨基酸序列相比的v708i和a593e氨基酸取代,其中变体衣壳蛋白在其它方面与aav2的氨基酸序列一致。在其它相关实施方式中,变体衣壳包含与aav2的氨基酸序列相比的v708i和s109t氨基酸取代,其中变体衣壳蛋白在其它方面与aav2的氨基酸序列一致。在其它实施方式中,变体衣壳蛋白包含与aav2的氨基酸序列相比的v708i和v719m氨基酸取代,并且与seqidno:2中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更多氨基酸序列同一性,并且在其它方面与氨基酸序列seqidno:2一致。在其它实施方式中,变体衣壳蛋白包含与aav2的氨基酸序列相比的v708i和r733c氨基酸取代,并且与seqidno:2中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更多氨基酸序列同一性,并且在其它方面与氨基酸序列seqidno:2一致。在其它实施方式中,变体衣壳蛋白包含与aav2的氨基酸序列相比的v708i和g727d氨基酸取代,并且与seqidno:2中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更多氨基酸序列同一性,并且在其它方面与氨基酸序列seqidno:2一致。
在其它实施方式中,变体aav衣壳蛋白包含与aav2衣壳的氨基酸序列(seqidno:2)相比氨基酸196或在另一种aav亲本血清型(即除了aav2之外)中的相应位置处的氨基酸取代,其中取代的氨基酸不会自然地出现在相应的位置并且任选地除了脯氨酸之外。在一些优选的实施方式中,变体衣壳蛋白包含氨基酸序列,所述氨基酸序列与seqidno:2中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更多氨基酸序列同一性,并且包含与aav2衣壳的氨基酸序列(seqidno:2)相比氨基酸196处的氨基酸取代并且任选地除了s196p取代之外。在优选的实施方式中,变体衣壳蛋白包含aav2或aav9的氨基酸196处或aav7、aav8或aav10的氨基酸197处或aav5的氨基酸186处的丝氨酸到酪氨酸氨基酸取代;或aav1或aav6的氨基酸196处的丙氨酸到酪氨酸取代;或aav4的氨基酸191处的甲硫氨酸到酪氨酸取代;或aav3a或aav3b的氨基酸196处的苏氨酸到酪氨酸取代。在相关实施方式中,变体衣壳蛋白包含氨基酸序列,所述氨基酸序列包含与seqidno:2中所示的氨基酸序列相比的s196y氨基酸取代,并且在其它方面与seqidno:2中所示的氨基酸序列一致。在相关实施方式中,变体衣壳蛋白包含与aav2衣壳的氨基酸序列(seqidno:2)相比的氨基酸196处的除了s196p取代(例如,包含s196y取代)之外的氨基酸取代,其中变体衣壳蛋白包含与seqidno:2中所示的aav2衣壳蛋白的氨基酸序列相比的1个至5个、5个至10个、或10个至15个氨基酸取代。
在其它实施方式中,变体aav衣壳蛋白包含与aav2衣壳的氨基酸序列(seqidno:2)相比氨基酸175或在另一种aav亲本血清型(即除了aav2之外)中的相应位置处的氨基酸取代,其中取代的氨基酸不会自然地出现在相应的位置。在一些优选的实施方式中,变体衣壳蛋白包含氨基酸序列,所述氨基酸序列与seqidno:2中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更多氨基酸序列同一性,并且包含与aav2衣壳的氨基酸序列(seqidno:2)相比氨基酸175处的氨基酸取代。在一些实施方式中,变体衣壳包含与seqidno:2中所示的aav2的氨基酸序列相比的q175h氨基酸取代或在另一种aav亲本血清型中的相应位置处的谷氨酰胺到组氨酸取代。在相关实施方式中,变体衣壳蛋白包含与aav2衣壳的氨基酸序列(seqidno:2)相比的氨基酸175(例如,q175h)处的氨基酸取代,其中变体衣壳蛋白包含与seqidno:2中所示的aav2衣壳蛋白的氨基酸序列相比的1个至5个、5个至10个、或10个至15个氨基酸取代。
在其它实施方式中,变体aav衣壳蛋白包含与aav2衣壳的氨基酸序列(seqidno:2)相比氨基酸64或在另一种aav亲本血清型(即除了aav2之外)中的相应位置处的氨基酸取代,其中取代的氨基酸不会自然地出现在相应的位置。在一些优选的实施方式中,变体衣壳蛋白包含氨基酸序列,所述氨基酸序列与seqidno:2中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更多氨基酸序列同一性,并且包含与aav2衣壳的氨基酸序列(seqidno:2)相比氨基酸64处的氨基酸取代。在一些实施方式中,raav病毒粒子包含与seqidno:2中所示的aav2的氨基酸序列相比的p64s氨基酸取代或另一种aav亲本血清型中的相应位置处的脯氨酸到丝氨酸取代。在相关实施方式中,变体衣壳蛋白包含与aav2衣壳的氨基酸序列(seqidno:2)相比的氨基酸64(例如,p64s)处的氨基酸取代,其中变体衣壳蛋白包含与seqidno:2中所示的aav2衣壳蛋白的氨基酸序列相比的1个至5个、5个至10个、或10个至15个氨基酸取代。
在其它实施方式中,变体aav衣壳蛋白包含与选自由以下组成的组的野生型aav衣壳序列具有至少85%、至少90%、至少95%或至少98%同一性的氨基酸序列:seqidnos:1、2、3、4、5、6、7、8、10、11和12,并且还包含i)一个或多个选自由以下组成的组的氨基酸取代:p34a、s109t+v708i、a593e+n596d、v708i+v719m、v708i+g727d、s109t+a493v+a593e+v708i、v708i+r733c、q164k和i698v和/或(ii)选自由以下组成的组的肽插入:qadttkn(seqidno:13)、isdqtkh(seqidno:14)、asdstka(seqidno:15)、nqdytkt(seqidno:16)、hditkni(seqidno:17)、pqananen(seqidno:63)、tnrtspd(seqidno:24)、pnsthgs(seqidno:25)、kdrapst(seqidno:26)、hqdttkn(seqidno:19)、hpdttkn(seqidno:18)、nkttnkd(seqidno:20)、gkskvid(seqidno:23)、pisneneh(seqidno:64)、laqadttkna(seqidno:27)、laisdqtkha(seqidno:28)、lgisdqtkha(seqidno:29)、laasdstkaa(seqidno:30)、lahditknia(seqidno:32)、lpqananena(seqidno:37)、lanqdytkta(seqidno:31)、latnrtspda(seqidno:39)、lapnsthgsa(seqidno:40)、lakdrapsta(seqidno:41)、lahqdttkna(seqidno:34)、lahpdttkna(seqidno:33)、lankttnkda(seqidno:35)、lagkskvida(seqidno:38)和lpisneneha(seqidno:36)。在一些实施方式中,变体aav衣壳包含指定的一个或多个氨基酸取代和/或肽插入,并且在其它方面与选自由以下组成的组的序列一致:seqidno:1-12。
在一些实施方式中,变体aav衣壳蛋白是祖先衣壳蛋白。祖先衣壳蛋白是指今天在自然界中发现的衣壳蛋白的进化祖先,例如aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aavrh10、aav11、aav12、aav13,其是通过在今天自然界中发现的aav衣壳蛋白之间的简并性位置处的随机氨基酸取代来计算机模拟产生的。下面提供了祖先衣壳的一个非限制性实例,其中简并性的位置(残基264、266、268、448、459、460、467、470、471、474、495、516、533、547、551、555、557、561、563、577、583、593、596、661、662、664、665、710、717、718、719、723))被标记为“x”:
maadgylpdwlednlsegirewwdlkpgapkpkanqqkqddgrglvlpgykylgpfngldkgepvnaadaaalehdkaydqqlkagdnpylrynhadaefqerlqedtsfggnlgravfqakkrvleplglveegaktapgkkrpvepspqrspdsstgigkkgqqpakkrlnfgqtgdsesvpdpqplgeppagpsglgsgtmaagggapmadnnegadgvgnasgnwhcdstwlgdrvittstrtwalptynnhlykqissxsxgxtndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevttndgvttiannltstvqvfsdseyqlpyvlgsahqgclppfpadvfmipqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylxrtqstggtagxxellfsqxgpxxmsxqaknwlpgpcyrqqrvsktlxqnnnsnfawtgatkyhlngrxslvnpgvamathkddexrffpssgvlifgkxgagxnntxlxnvmxtxeeeikttnpvatexygvvaxnlqssntapxtgxvnsqgalpgmvwqnrdvylqgpiwakiphtdgnfhpsplmggfglkhpppqilikntpvpanppxxfxxakfasfitqystgqvsveiewelqkenskrwnpeiqytsnyaksxnvdfavxxxgvyxeprpigtryltrnl(seqidno:58)
在一些实施方式中,祖先衣壳蛋白包含氨基酸序列,所述氨基酸序列与seqidno:58中所示的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更多氨基酸序列同一性。在一些实施方式中,祖先衣壳蛋白包含氨基酸序列,所述氨基酸序列与例如seqidno:2中所示的aav2的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更多氨基酸序列同一性。在一些实施方式中,祖先衣壳蛋白包含氨基酸序列,所述氨基酸序列与seqidno:58或seqidno:2中公开的祖先序列的氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%或更高的氨基酸序列同一性,并且包含选自由以下组成的组的一个或多个氨基酸残基:264处的丙氨酸(a)、266处的丙氨酸(a)、268处的丝氨酸(s)、448处的丙氨酸(a)、459处的苏氨酸(t)、460处的精氨酸(r)、467处的丙氨酸(a)、470处的丝氨酸(s)、471处的天冬酰胺(n)、474处的丙氨酸(a)、495处的丝氨酸(s)、516处的天冬酰胺(d)、533处的天冬酰胺(d)、547处的谷氨酰胺(q)、551处的丙氨酸(a)、555处的丙氨酸(a)、557处的谷氨酸(e)、561处的甲硫氨酸(m)、563处的丝氨酸(s)、577处的谷氨酰胺(q)、583处的丝氨酸(s)、593处的缬氨酸(v)、596处的苏氨酸(t)、661处的丙氨酸(a)、662处的缬氨酸(v)、664处的苏氨酸(t)、665处的脯氨酸(p)、710处的苏氨酸(t)、717处的天冬氨酸(d)、718处的天冬酰胺(n)、719处的谷氨酸(e)、以及723处的丝氨酸(s)。在一些优选的实施方式中,变体衣壳蛋白包含氨基酸序列,所述氨基酸序列与以下氨基酸序列的整个长度具有至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%、在一些情况下100%的氨基酸序列同一性,并且包含选自由以下组成的组的一个或多个氨基酸残基:264处的丙氨酸(a)、266处的丙氨酸(a)、268处的丝氨酸(s)、448处的丙氨酸(a)、459处的苏氨酸(t)、460处的精氨酸(r)、467处的丙氨酸(a)、470处的丝氨酸(s)、471处的天冬酰胺(n)、474处的丙氨酸(a)、495处的丝氨酸(s)、516处的天冬酰胺(d)、533处的天冬酰胺(d)、547处的谷氨酰胺(q)、551处的丙氨酸(a)、555处的丙氨酸(a)、557处的谷氨酸(e)、561处的甲硫氨酸(m)、563处的丝氨酸(s)、577处的谷氨酰胺(q)、583处的丝氨酸(s)、593处的缬氨酸(v)、596处的苏氨酸(t)、661处的丙氨酸(a)、662处的缬氨酸(v)、664处的苏氨酸(t)、665处的脯氨酸(p)、710处的苏氨酸(t)、717处的天冬氨酸(d)、718处的天冬酰胺(n)、719处的谷氨酸(e)和723处的丝氨酸(s):
maadgylpdwlednlsegirewwdlkpgapkpkanqqkqddgrglvlpgykylgpfngldkgepvnaadaaalehdkaydqqlkagdnpylrynhadaefqerlqedtsfggnlgravfqakkrvleplglveegaktapgkkrpvepspqrspdsstgigkkgqqpakkrlnfgqtgdsesvpdpqplgeppagpsglgsgtmaagggapmadnnegadgvgnasgnwhcdstwlgdrvittstrtwalptynnhlykqissasagstndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevttndgvttiannltstvqvfsdseyqlpyvlgsahqgclppfpadvfmipqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylartqstggtagtrellfsqagpsnmsaqaknwlpgpcyrqqrvsktlsqnnnsnfawtgatkyhlngrdslvnpgvamathkddedrffpssgvlifgkqgaganntalenvmmtseeeikttnpvateqygvvasnlqssntapvtgtvnsqgalpgmvwqnrdvylqgpiwakiphtdgnfhpsplmggfglkhpppqilikntpvpanppavftpakfasfitqystgqvsveiewelqkenskrwnpeiqytsnyakstnvdfavdnegvyseprpigtryltrnl(seqidno:59).
在其它实施方式中,变体aav衣壳蛋白包含与选自由以下组成的组的野生型aav衣壳序列至少85%、至少90%、至少95%或至少98%一致的氨基酸序列:如本文seqidno:58中公开的祖先变体,包含选自由以下组成的组的一个或多个氨基酸残基:264处的丙氨酸(a)、266处的丙氨酸(a)、268处的丝氨酸(s)、448处的丙氨酸(a)、459处的苏氨酸(t)、460处的精氨酸(r)、467处的丙氨酸(a)、470处的丝氨酸(s)、471处的天冬酰胺(n)、474处的丙氨酸(a)、495处的丝氨酸(s)、516处的天冬酰胺(d)、533处的天冬酰胺(d)、547处的谷氨酰胺(q)、551处的丙氨酸(a)、555处的丙氨酸(a)、557处的谷氨酸(e)、561处的蛋氨酸(m)、563处的丝氨酸(s)、577处的谷氨酰胺(q)、583处的丝氨酸(s)、593处的缬氨酸(v)、596处的苏氨酸(t)、661处的丙氨酸(a)、662处的缬氨酸(v)、664处的苏氨酸(t)、665处的脯氨酸(p)、710处的苏氨酸(t)、717处的天冬氨酸(d)、718处的天冬酰胺(n)、719处的谷氨酸(e)和723处的丝氨酸(s);并且还包含i)一个或多个选自由以下组成的组的氨基酸取代:p34a、s109t+v708i、a593e+n596d、v708i+v719m、v708i+g727d、s109t+a493v+a593e+v708i、v708i+r733c、q164k和i698v和/或(ii)选自由以下组成的组的肽插入:qadttkn(seqidno:13)、isdqtkh(seqidno:14)、asdstka(seqidno:15)、nqdytkt(seqidno:16)、hditkni(seqidno:17)、pqananen(seqidno:63)、tnrtspd(seqidno:24)、pnsthgs(seqidno:25)、kdrapst(seqidno:26)、hqdttkn(seqidno:19)、hpdttkn(seqidno:18)、nkttnkd(seqidno:20)、gkskvid(seqidno:23)、pisneneh(seqidno:64)、laqadttkna(seqidno:27)、laisdqtkha(seqidno:28)、lgisdqtkha(seqidno:29)、laasdstkaa(seqidno:30)、lahditknia(seqidno:32)、lpqananena(seqidno:37)、lanqdytkta(seqidno:31)、latnrtspda(seqidno:39)、lapnsthgsa(seqidno:40)、lakdrapsta(seqidno:41)、lahqdttkna(seqidno:34)、lahpdttkna(seqidno:33)、lankttnkda(seqidno:35)、lagkskvida(seqidno:38)和lpisneneha(seqidno:36)。在一些实施方式中,变体aav衣壳包含指定的一个或多个氨基酸取代和/或肽插入,并且在其它方面与seqidno:59一致。
本文公开的aav变体通过使用体内定向进化产生,其包括在玻璃体内施用后使用灵长类视网膜筛选。在一些实施方式中,与通过包含相应亲本aav衣壳蛋白或野生型aav的aav病毒粒子转导视网膜细胞相比,当存在于aav病毒粒子中时,本文公开的变体衣壳蛋白赋予对视网膜细胞增加的转导。例如,在一些实施方式中,当存在于aav病毒粒子中时,与包含相应亲本aav衣壳蛋白或野生型aav衣壳蛋白的aav病毒粒子相比,本文公开的变体衣壳蛋白赋予对灵长类视网膜细胞更有效的转导,例如视网膜细胞摄取比包含亲本aav衣壳蛋白或野生型aav的aav病毒粒子更多的包含受试者变体aav衣壳蛋白的aav病毒粒子。在一些这种实施方式中,与通过野生型aav病毒粒子或包含相应亲本aav衣壳蛋白的raav转导视网膜细胞相比,aav变体病毒粒子或变体raav对视网膜细胞表现出至少2倍、至少5倍、至少10倍、至少15倍、至少20倍、至少25倍、至少50倍或超过50倍的增加的转导。在某些此类实施方式中,当存在于aav病毒粒子中时,与包含相应亲本aav衣壳蛋白或野生型aav衣壳蛋白的aav病毒粒子相比,本文公开的变体衣壳蛋白赋予对灵长类视网膜细胞更广泛的转导。换句话说,变体aav病毒粒子转导未被包含相应亲本aav衣壳蛋白的病毒粒子转导的细胞类型,因此在视网膜中转导比相应亲本aav病毒粒子更多类型的细胞。在一些实施方式中,aav变体病毒粒子优先转导视网膜细胞,例如,与另一种视网膜细胞或非视网膜细胞,例如眼睛外的细胞相比,主题raav病毒粒子以2倍、5倍、10倍、15倍、20倍、25倍、50倍或多于50倍特异性感染视网膜细胞。在一些实施方式中,转导的视网膜细胞是感光细胞(例如,视杆;视锥)。在一些实施方式中,视网膜细胞是视网膜神经节细胞(rgc)。在一些实施方式中,视网膜细胞是视网膜上皮细胞(rpe细胞)。在一些实施方式中,视网膜细胞是穆勒神经胶质细胞。在一些实施方式中,视网膜细胞是小神经胶质细胞。在一些实施方式中,视网膜细胞是无长突细胞。在一些实施方式中,视网膜细胞是双极细胞。在一些实施方式中,视网膜细胞是水平细胞。通过任何数量的本领域中用于测量基因表达的方法,可以容易地在体外或体内评估视网膜细胞转导的增加,例如增加的转导效率、更广泛的转导、更优先的转导等。例如,aav可以与包含表达盒的基因组一起包装,所述表达盒包含在普遍存在的或组织特异性启动子的控制下的报告基因,例如荧光蛋白,并且通过由例如荧光显微镜检测荧光蛋白来评估转导程度。作为另一个实例,aav可以与包含条形码核酸序列的基因组一起包装,并且通过由例如pcr检测核酸序列来评估转导程度。作为另一个实例,aav可以与包含表达盒的基因组一起包装,所述表达盒包含用于治疗视网膜疾病的治疗基因,并且通过检测施用aav的受困扰的患者中视网膜疾病的治疗来评估转导程度。
可使用本文公开的变体raav载体或病毒粒子和/或方法治疗的眼部疾病包括但不限于单基因疾病、复杂遗传疾病、获得性疾病和创伤性损伤。单基因疾病的实例包括但不限于巴德-毕德(bardet-biedl)综合征;巴藤病(batten'sdisease);bietti氏结晶营养不良;无脉络膜症;脉络膜视网膜萎缩;脉络膜视网膜变性;视锥或视锥-视杆状营养不良(常染色体显性遗传、常染色体隐性遗传和x连锁);先天性静止性夜盲症(常染色体显性遗传、常染色体隐性遗传和x连锁);色觉障碍,包含全色盲(包含achm2、achm3、achm4和achm5)、红色盲、绿色盲和蓝色盲;弗里德赖希共济失调;莱伯氏先天性黑朦(常染色体显性遗传和常染色体隐性遗传),包括但不限于lca1、lca2、lca3、lca4、lca6、lca7、lca8、lca12和lca15;莱伯氏遗传性视神经病变;黄斑营养不良(常染色体显性遗传和常染色体隐性遗传),包括但不限于急性黄斑变性、最佳卵黄样黄斑营养不良、图形样营养不良、北卡罗莱纳黄斑营养不良(northcarolinamaculardystrophy)、遗传性玻璃疣、索斯比眼底营养不良(sorsby'sfundusdystrophy)、malattialevantanese和遗传决定的早产儿视网膜病;眼视网膜发育疾病;眼白化病;视神经萎缩(常染色体显性遗传、常染色体隐性遗传和x连锁);色素性视网膜炎(常染色体显性遗传、常染色体隐性遗传、x连锁和线粒体遗传性状),其实例包含rp1、rp2、rp3、rp10、rp20、rp38、rp40和rp43;x连锁视网膜劈裂症;斯特格氏病(stargardtdisease);以及尤塞氏综合症(ushersyndrome),包括但不限于ush1b、ush1c、ush1d、ush1f、ush1g、ush2a、ush2c、ush2d和ush3。复杂遗传疾病的实例包括但不限于青光眼(开角型、闭角型、低眼压性、正常眼压性、先天性、新生血管性、色素性、假性剥脱);年龄相关和其它形式的黄斑变性,渗出性和非渗出性形式(常染色体显性遗传和常染色体隐性遗传),如急性黄斑变性,卵黄样黄斑变性;早产儿视网膜病;和vogtkoyanagi-harada(vkh)综合症。获得性疾病的实例包括但不限于急性黄斑神经视网膜病;前部缺血性视神经病和后部缺血性视神经病;白塞氏病(behcet'sdisease);分支视网膜静脉阻塞;脉络膜新生血管;糖尿病视网膜病变,包含增殖性糖尿病视网膜病变和相关并发症;糖尿病性葡萄膜炎;水肿,如黄斑水肿、囊样黄斑水肿和糖尿病性黄斑水肿;视网膜前膜病症;黄斑毛细血管扩张症;多灶性脉络膜炎;非视网膜病变糖尿病性视网膜功能障碍;眼部肿瘤;视神经萎缩;视网膜脱离;视网膜病症,如中央视网膜静脉阻塞、增殖性玻璃体视网膜病变(pvr)、视网膜动脉和静脉闭塞性疾病、血管闭塞、葡萄膜炎性视网膜疾病;葡萄膜渗漏;视网膜感染和浸润性疾病;视神经疾病,如获得性视神经萎缩。创伤性损伤的实例包括但不限于组织胞浆菌病;视神经损伤;影响后眼部位或位置的眼外伤;视网膜损伤;眼睛的病毒感染;视神经的病毒感染;由眼部激光治疗引起或受其影响的后眼部病症;由光动力疗法引起或受其影响的后眼部病症;光凝,放射性视网膜病变;以及交感性眼炎。
在另一个实施方式中,本文公开的变体衣壳包含异源核酸,其包含编码基因产物的核苷酸序列,例如但不限于干扰rna、长非编码rna、短非编码rna、反义rna、适体、多肽、分泌的抗体、单链抗体、vhh结构域、可溶性受体、亲和体(affibody)、打结素(knottin)、darpin、centurin、伴侣蛋白、提供基因功能的位点特异性敲低的位点特异性核酸酶或提供转录的基因特异性活化的修饰的位点特异性核酸酶。
本文公开的raav变体病毒粒子包含异源核酸,其包含编码基因产物的核苷酸序列。在一些实施方式中,基因产物是干扰rna。在一些实施方式中,基因产物是长的非编码rna。在一些实施方式中,基因产物是短的非编码rna。在一些实施方式中,基因产物是反义rna。在一些实施方式中,基因产物是适体。在一些实施方式中,基因产物是多肽。在一些实施方式中,基因产物是分泌的抗体。在一些实施方式中,基因产物是单链抗体。在一些实施方式中,基因产物是vhh结构域。在一些实施方式中,基因产物是可溶性受体。在一些实施方式中,基因产物是亲和体。在一些实施方式中,基因产物是打结素。在一些实施方式中,基因产物是darpin。在一些实施方式中,基因产物是centurin。在一些实施方式中,基因产物是伴侣蛋白。在一些实施方式中,基因产物是提供基因功能的位点特异性敲低的位点特异性核酸酶。
基因产物的用途包括但不限于:提高细胞中因子的水平;通过分泌因子来提高相邻细胞中因子的水平;降低细胞中因子的水平;或者通过分泌因子来降低相邻细胞中因子的水平。基因产物可以设计为:补充缺失基因产物缺陷的水平;降低缺失基因产物缺陷的水平;引入新的支持基因产物;补充支持基因产物的水平;降低阻碍基因产物的水平;或者既降低阻碍基因产物的水平又引入或补充支持基因产物的水平。
由主题aav变体递送的基因产物可以用于改变直接或间接与视网膜疾病和创伤相关的基因产物或基因产物活性的水平。基因产物与遗传疾病直接或间接相关的基因包括例如adp-核糖基化因子样6(arl6);bbsome相互作用蛋白1(bbip1);bbsome蛋白1(bbs1);bbsome蛋白2(bbs2);bbsome蛋白4(bbs4);bbsome蛋白5(bbs5);bbsome蛋白7(bbs7);bbsome蛋白9(bbs9);bbsome蛋白10(bbs10);bbsome蛋白12(bbs12);中心体蛋白290kda(cep290);细胞纤毛内运输蛋白172(ift172);细胞纤毛内运输蛋白27(ift27);肌醇多磷酸-5-磷酸酶e(inpp5e);内向整流钾通道亚家族j成员13(kcnj13);亮氨酸拉链转录因子样-1(lztfl1);mckusick-kaufman综合征蛋白(mkks);梅克尔(meckel)综合征1型蛋白(mks1);肾结核3蛋白(nphp1);血清学定义的结肠癌抗原8(sdccag8);含有三重基序的蛋白32(trim32);三角形四肽(tetratricopeptide)重复结构域8(ttc8);巴特病蛋白(cln3);拉布(rab)护送蛋白1(chm);(prdm13);(rgr;(tead1);芳香烃相互作用受体蛋白样1(aipl1);视锥-视杆otx样光感受器同源框转录因子(crx);鸟苷酸环化酶活化蛋白1a(guca1a);视网膜特异性鸟苷酸环化酶(gucy2d);磷脂酰肌醇转移膜相关家族成员3(pitpnm3);prominin1(prom1);外周蛋白(prph);外周蛋白2(prph2);调节突触膜胞吐蛋白1(rims1);臂板蛋白4a(sema4a);秀丽隐杆线虫(c.elegans)unc119蛋白的人类同源物(unc119);atp结合盒转运蛋白-视黄醛(abca4);adam金属肽酶结构域9(adam9);活化转录因子6(atf6);21号染色体开放阅读框2(c21orf2);8号染色体开放阅读框37(c8orf37);钙通道;电压依赖性;α2/δ亚基4(cacna2d4);钙粘蛋白相关的家庭成员1(原钙粘附蛋白21)(cdhr1);神经酰胺激酶样蛋白(cerkl);锥形光感受器cgmp门控阳离子通道α亚基(cnga3);锥环核苷酸门控阳离子通道β3亚基(cngb3);细胞周期蛋白m4(cnnm4);鸟嘌呤核苷酸结合蛋白(g蛋白);α转导活性多肽2(gnat2);钾通道亚家族v成员2(kcnv2);磷酸二酯酶6c(pde6c);磷酸二酯酶6h(pde6h);中心粒1中心蛋白b蛋白质组(poc1b);ras癌基因家族的rab28成员(rab28);视网膜和前神经折叠同源框2转录因子(rax2);11-顺式视黄醇脱氢酶5(rdh5);rpgtp酶调节子相互作用蛋白1(rpgrip1);微管蛋白酪氨酸连接酶样家族成员5(ttll5);l型电压门控钙通道α-1亚基(cacna1f);视网膜色素变性gtp酶调节子(rpgr);(gnat1);(pde6b);(rho);cabp4);gpr179);(grk1);grm6);lrit3);slc24a1);trpm1);nyx);opn1lw);opn1mw);蓝视锥视蛋白(opn1sw);共济蛋白(fxn);(impdh1);(otx2);crb1);dthd1gdf6);ift140);iqcb1);lca5);lrat);nmnat1);rd3);rdh12);rpe65);spata7);tulp1);线粒体基因(kss、lhon、mt-atp6、mt-th、mt-tl1、mt-tp、mt-ts2,线粒体编码的nadh脱氢酶[mt-nd]);(best1);c1qtnf5efemp1);elovl4);fscn2);guca1b);hmcn1);impg1);rp1l1);timp3);dram2);mfn2);nr2f1);视神经萎缩1(opa1);tmem126a);timm8a);ca4);hk1);klhl7);nr2e3);nrl);or2w3);prpf3);prpf4);prpf6);prpf8);prpf31);rom1);视网膜色素变性蛋白(rp1);rp9);snrnp200);spp2);topors);arl2bp);c2orf71);clrn1);cnga1);cngb1);cyp4v2);dhdds);dhx38);emc1);eys);fam161a);gpr125);hgsnat);idh3b);impg2);kiaa1549);kiz);mak);mertk);mvk);nek2);neurod1);pde6a);pde6g);prcd);rbp3);rlbp1);slc7a14);ush2a);znf408);znf513);ofd1);rp2);视网膜劈裂蛋白(rs1);abhd12);cdh23);cep250);cib2);dfnb31);gpr98);hars);myo7a);pcdh15);ush1c);ush1g);ndp);pgk1);capn5);fzd4);itm2b);lrp5);mir204);rb1);tspan12);c12orf65);cdh3);mfrp);oat);pla2g5);rbp4);rgs9);rgs9bp);arms2;ercc6);fbln5);htra1);tlr3);和tlr4).
基因产物诱导或促进凋亡的基因在本文中称为“促凋亡基因”,这些基因(mrna;蛋白质)的产物被称为“促凋亡基因产物”。促凋亡靶标包含例如bax基因产物;bid基因产物;bak基因产物;bad基因产物;bcl-2;bcl-x1。抗凋亡基因产物包含x连锁的凋亡抑制剂。
基因产物诱导或促进血管生成的基因在本文中称为“促血管生成基因”,这些基因(mrna;蛋白质)的产物被称为“促血管生成基因产物”。促血管生成靶标包含例如血管内皮生长因子(vegfa、vegfb、vegfc、vegfd);血管内皮生长因子受体1(vegfr1);血管内皮生长因子受体2(vegfr2);fms相关酪氨酸激酶1(flt1);胎盘生长因子(pgf);血小板源性生长因子(pdgf);血管生成素;声波刺猬。基因产物抑制血管生成的基因在本文中称为“抗血管生成基因”,并且这些基因(mrna;蛋白质)的产物被称为“抗血管生成基因产物”。抗血管生成基因产物包含内皮抑素;肿瘤抑素;血管抑素;色素上皮源性因子(pedf),以及特异于促血管生成靶标和/或其受体的融合蛋白或抗体,例如抗vegf融合蛋白sflt1或eylea,vegf特异性抗体lucentistm和avastintm等。
在一些实施方式中,由主题aav变体递送的基因产物用于抑制血管生成。在某些优选的实施方式中,由主题aav变体递送的基因产物用于抑制一种或多种哺乳动物vegf蛋白的活性,所述vegf蛋白选自由vegfa、vegfb、vegfc、vegfd和pgf组成的组。在特别优选的实施方式中,由主题aav变体递送的基因产物抑制vegfa的活性。vegfa具有通过可变剪接产生的9种同种型,其中生理学上最相关的是vegf165。已经发现患有湿性年龄相关性黄斑变性、糖尿病性黄斑水肿和视网膜静脉阻塞的患者的玻璃体中vegfa水平升高。抑制眼中vegfa活性并因此有效治疗玻璃体vegfa升高的患者的基因产物包括但不限于阿柏西普、雷珠单抗、溴珠单抗、贝伐单抗和可溶性fms-样酪氨酸激酶1(sflt1)(genbank登录号u01134)。在一些实施方式中,提供了感染性重组aav(raav)病毒粒子,其包含(i)如本文所述的变体aav衣壳蛋白和(ii)包含多个序列的异源核酸,所述多个序列中的每一个编码不同的vegfa抑制剂。
在一个优选实施方式中,基因产物是阿柏西普。阿柏西普
atggtttcttactgggacaccggcgtgctgctgtgtgccctgctttcttgtctgctgctgaccggctctagcagcggctctgataccggcagacccttcgtggaaatgtacagcgagatccccgagatcatccacatgaccgagggcagagagctggtcatcccttgcagagtgacaagccccaacatcaccgtgactctgaagaagttccctctggacacactgatccccgacggcaagagaatcatctgggacagccggaagggcttcatcatcagcaacgccacctacaaagagatcggcctgctgacctgtgaagccaccgtgaatggccacctgtacaagaccaactacctgacacacagacagaccaacaccatcatcgacgtggtgctgagccctagccacggcattgaactgtctgtgggcgagaagctggtgctgaactgtaccgccagaaccgagctgaacgtgggcatcgacttcaactgggagtaccccagcagcaagcaccagcacaagaaactggtcaaccgggacctgaaaacccagagcggcagcgagatgaagaaattcctgagcaccctgaccatcgacggcgtgaccagaagtgaccagggcctgtacacatgtgccgccagctctggcctgatgaccaagaaaaacagcaccttcgtgcgggtgcacgagaaggacaagacccacacctgtcctccatgtcctgctccagaactgctcggcggaccttccgtgttcctgtttcctccaaagcctaaggacaccctgatgatcagcagaacccctgaagtgacctgcgtggtggtggatgtgtcccacgaggatcccgaagtgaagttcaattggtacgtggacggcgtggaagtgcacaacgccaagaccaagcctagagaggaacagtacaatagcacctacagagtggtgtccgtgctgaccgtgctgcaccaggattggctgaacggcaaagagtacaagtgcaaggtgtccaacaaggccctgcctgctcctatcgagaaaaccatctccaaggccaagggccagcctagggaaccccaggtttacacactgcctccaagcagggacgagctgacaaagaaccaggtgtccctgacctgcctggtcaagggcttctacccttccgatatcgccgtggaatgggagagcaatggccagcctgagaacaactacaagacaacccctcctgtgctggacagcgacggctcattcttcctgtacagcaagctgacagtggacaagagcagatggcagcagggcaacgtgttcagctgctccgtgatgcacgaggccctgcacaaccactacacccagaagtccctgagcctgtctcctggcaaatga(seqidno:65)。
在一些实施方式中,提供了编码阿柏西普的核酸序列,其包含与seqidno:65中所示核酸序列的全长或与seqidno:65的核苷酸79-1377(不具有编码flt1信号序列的下划线核苷酸)具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%,或更高的核酸同一性的核酸序列。在相关实施方式中,由主题aav变体递送的基因产物由核酸序列编码,所述核酸序列由seqidno:65的核酸序列或与seqidno:65的核酸序列具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%同一性的核酸组成,或包含其。在其它相关实施方式中,由主题aav变体递送的基因产物包含与下列氨基酸序列具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%同一性或100%同一性的氨基酸序列:
mvsywdtgvllcallscllltgsssgsdtgrpfvemyseipeiihmtegrelvipcrvtspnitvtlkkfpldtlipdgkriiwdsrkgfiisnatykeiglltceatvnghlyktnylthrqtntiidvvlspshgielsvgeklvlnctartelnvgidfnweypsskhqhkklvnrdlktqsgsemkkflstltidgvtrsdqglytcaassglmtkknstfvrvhekdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsrdeltknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk(seqidno:66)
在其它相关实施方式中,aav变体包含异源核酸,该异源核酸包含编码与seqidno:66的氨基酸27-458(对应于不具有下划线信号肽序列的阿柏西普的氨基酸序列)具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%同一性或100%同一性的氨基酸序列的核苷酸序列。
在另一个优选的实施方式中,基因产物是单链形式的雷珠单抗(sc-雷珠单抗)。雷珠单抗
atggactggacctggtccatcctgtttctggtggctgccgccacaggcacatactctgaagtgcagctggtggaatctggcggcggacttgttcaacctggcggctctctgagactgagctgtgccgcctctggctacgactttacccactacggcatgaactgggtccgacaggcccctggcaaaggccttgaatgggtcggatggatcaacacctacaccggcgagccaacatacgccgccgacttcaagcggagattcaccttcagcctggacaccagcaagagcaccgcctacctccagatgaacagcctgagagccgaggacaccgccgtgtactactgcgccaagtatccctactactacggcaccagccactggtacttcgacgtgtggggacagggcacactggtcacagtgtctagcgcctctacaaagggccccagcgttttcccactggctcctagcagcaagtctacctccggtggaacagccgctctgggctgtctggtcaaggactactttcccgagcctgtgaccgtgtcctggaatagcggagcactgacaagcggcgtgcacacctttccagccgtgctgcaaagcagcggcctgtactctctgagcagcgtcgtgacagtgccaagcagctctctgggcacccagacctacatctgcaatgtgaaccacaagcctagcaacaccaaggtggacaagaaggtggaacccaagagctgcgacaagacacacctcggcggaagctctggaagcggctctggatctaccggcacaagctctagcggaacaggcacaagcgctggcacaaccggaacaagcgcttctacatctggctctggttctggcggaggcggaggatcaggtggtggtggatctgctggcggaacagctacagctggcgcctcttctggcagcgacattcagctgacacagagcccttctagcctgagcgcctctgtgggcgacagagtgaccatcacatgtagcgccagccaggacatctccaactacctgaactggtatcagcagaagcccggcaaggcccctaaggtgctgatctactttaccagcagcctgcactccggcgtgcccagcagattttctggatctggctccggcaccgacttcaccctgacaatatctagcctccagcctgaggacttcgccacctactactgccagcagtacagcaccgtgccttggacatttggccagggcacaaaggtggaaatcaagcggacagtggccgctcctagcgtgttcatctttccacctagcgacgagcagctgaagtctggcacagcctctgtcgtgtgcctgctgaacaacttctaccccagagaagccaaggtgcagtggaaagtggacaacgccctccagtccggcaacagccaagagtctgtgaccgagcaggacagcaaggactccacctacagcctgtccagcacactgacactgagcaaggccgactacgagaagcacaaagtgtacgcctgcgaagtgacccaccagggcctttctagccctgtgaccaagagcttcaaccggggcgagtgttga(seqidno:67)
在一些实施方式中,提供了编码sc-雷珠单抗hl的核酸序列,其包含与seqidno:67中所示核酸序列的全长或与seqidno:67的核苷酸58-1575(不具有编码人ighv7-8信号序列的下划线核苷酸)具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%或更高核酸同一性的核酸序列。在相关实施方式中,由主题aav变体递送的基因产物由核酸序列编码,所述核酸序列由seqidno:67的核酸序列或与seqidno:67的核酸序列具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%同一性的核酸组成,或包含其。在其它相关实施方式中,由主题aav变体递送的基因产物包含与下列氨基酸序列具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%同一性或100%同一性的氨基酸序列:
mdwtwsilflvaaatgtysevqlvesggglvqpggslrlscaasgydfthygmnwvrqapgkglewvgwintytgeptyaadfkrrftfsldtskstaylqmnslraedtavyycakypyyygtshwyfdvwgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthlggssgsgsgstgtsssgtgtsagttgtsastsgsgsgggggsggggsaggtatagassgsdiqltqspsslsasvgdrvtitcsasqdisnylnwyqqkpgkapkvliyftsslhsgvpsrfsgsgsgtdftltisslqpedfatyycqqystvpwtfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec(seqidno:68)。
在其它相关实施方式中,aav变体包含异源核酸,所述异源核酸包含编码与seqidno:68的氨基酸20-524(对应于不具有下划线信号肽序列的sc-雷珠单抗hl的氨基酸序列)具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%同一性或100%同一性的氨基酸序列的核苷酸序列。
在另一个特别优选的实施方式中,提供了编码对应于图12b的单链轻-重(lh)形式的雷珠单抗(sc-雷珠单抗lh)的新型密码子经优化的核酸序列,所述核酸序列包含以下或由以下组成:
atggtcctccagacacaggtgttcatcagcctgctgctgtggatctctggcgcctacggcgatatccagctgacacagagccctagcagcctgtctgcctctgtgggcgacagagtgaccatcacatgtagcgccagccaggacatcagcaactacctgaactggtatcagcagaagcccggcaaggcccctaaggtgctgatctactttaccagcagcctgcacagcggcgtgcccagcagattttctggctctggcagcggcaccgacttcaccctgacaatatctagcctccagcctgaggacttcgccacctactactgccagcagtacagcaccgtgccttggacatttggccagggcaccaaggtggaaatcaagcggacagtggccgctcctagcgtgttcatctttccacctagcgacgagcagctgaagtctggcacagcctctgtcgtgtgcctgctgaacaacttctaccccagagaagccaaggtgcagtggaaggtggacaacgccctccagagcggcaatagccaagagagcgtgaccgagcaggacagcaaggactctacctactctctgagcagcacactgaccctgagcaaggccgactacgagaagcacaaagtgtacgcctgcgaagtgacccaccagggcctttctagccctgtgaccaagagcttcaacagaggcgagtgtggcggcagctctggaagcggatctggaagcacaggcacaagctctagcggaaccggaacaagcgctggcacaacaggcacatctgccagcacaagcggttctggttctggcggaggcggaggatctggtggtggcggatctgctggcggaacagctacagctggtgcctcttctggatccgaggtgcagctggttgaatctggcggtggactggttcagccaggcggatctctgagactgtcttgtgccgccagcggctacgatttcacccactacggcatgaactgggtccgacaggcccctggcaaaggccttgaatgggtcggatggatcaacacctacaccggcgagccaacatacgccgccgacttcaagcggagattcaccttcagcctggacacctccaagagcaccgcctacctccagatgaacagcctgagagccgaggacaccgccgtgtactactgcgccaagtatccctactactacggcaccagccactggtacttcgacgtgtggggacagggcacactggtcacagtgtctagcgcctctacaaagggccccagcgttttcccactggctcctagcagcaagagcacatcaggcggaactgctgccctgggctgtctggtcaaggactactttcctgagcctgtgaccgtgtcctggaacagcggagcactgacatctggcgtgcacacctttccagccgtgctccaaagcagcggcctgtattctctgtccagcgtcgtgacagtgcctagcagctctctgggcacccagacctacatctgcaatgtgaaccacaagcctagcaacaccaaggtcgacaagaaggtggaacccaagagctgcgacaagacccacctctga(seqidno:69)
在另一个特别优选的实施方式中,提供了编码对应于图12b的单链轻-重(lh)形式的雷珠单抗(sc-雷珠单抗lh)的新型密码子经优化的核酸序列,所述核酸序列包含以下或由以下组成:
atggtgctccagacccaggtgtttatttcactgctgctgtggattagcggggcttacggagacattcagctgacccagagtccttcatctctgagcgcctccgtgggcgacagggtgaccatcacatgctctgccagccaggatatctccaactatctgaattggtaccagcagaagcccggcaaggcccctaaggtgctgatctatttcaccagctccctgcacagcggagtgccatcccgcttctccggctctggcagcggcaccgactttaccctgacaatctctagcctccagccagaggatttcgccacatactattgccagcagtacagcaccgtgccctggacatttggccagggcaccaaggtggagatcaagcggacagtggccgccccaagcgtgttcatctttccccctagcgacgagcagctgaagtctggcaccgccagcgtggtgtgcctgctgaacaatttctatcccagagaggccaaggtgcagtggaaggtggataacgccctccagtccggcaattctcaggagagcgtgaccgagcaggactccaaggattctacatacagcctgtcctctaccctgacactgtccaaggccgactatgagaagcacaaggtgtacgcatgcgaggtgacccaccagggcctgagctccccagtgacaaagagctttaacaggggagagtgtggaggatctagcggatccggatctggaagcaccggcacatcctctagcggaaccggcacaagcgccggcaccacaggcacctccgcctctacaagcggcagcggatctggcggaggaggaggatccggaggaggaggatctgccggcggcaccgccacagccggcgcctcctctggctccgaggtgcagctggtggagtctggaggaggcctggtgcagcctggaggctccctgaggctgtcttgcgcagcaagcggctatgatttcacccactacggaatgaactgggtgcgccaggcacctggcaagggcctggagtgggtgggctggatcaatacctatacaggcgagccaacctacgccgccgactttaagcggagattcacattttccctggataccagcaagtccacagcctacctccagatgaacagcctgagggcagaggacaccgccgtgtactattgcgccaagtatccttactattacggcacaagccactggtacttcgacgtgtggggacagggcaccctggtgacagtgagctccgccagcaccaagggcccatccgtgtttcctctggccccatctagcaagtctaccagcggaggaacagccgccctgggatgtctggtgaaggactacttcccagagcccgtgaccgtgtcctggaattctggcgccctgacctccggcgtgcacacatttcccgccgtgctccagtcctctggcctgtatagcctgagctccgtggtgaccgtgccttctagctccctgggcacccagacatacatctgtaacgtgaatcacaagccttcaaataccaaagtcgataaaaaagtggaaccaaaatcctgtgataaaacccatctgtga(seqidno:70)。
在一些实施方式中,提供了编码sc-雷珠单抗lh的核酸序列,其包含与seqidno:69或seqidno:70中所示核酸序列的全长或与seqidno:69或seqidno:70的核苷酸61-1578(不具有编码igκ信号序列的下划线核苷酸)具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%或更高核酸同一性的核酸序列。在相关实施方式中,由主题aav变体递送的基因产物由核酸序列编码,所述核酸序列由seqidno:69或seqidno:70的核酸序列或者与seqidno:69或seqidno:70的核酸序列具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%同一性的核酸组成,或包含其。在其它相关实施方式中,由主题aav变体递送的基因产物包含与下列氨基酸序列具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%同一或100%同一性的氨基酸序列:
mvlqtqvfislllwisgaygdiqltqspsslsasvgdrvtitcsasqdisnylnwyqqkpgkapkvliyftsslhsgvpsrfsgsgsgtdftltisslqpedfatyycqqystvpwtfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgecggssgsgsgstgtsssgtgtsagttgtsastsgsgsgggggsggggsaggtatagassgsevqlvesggglvqpggslrlscaasgydfthygmnwvrqapgkglewvgwintytgeptyaadfkrrftfsldtskstaylqmnslraedtavyycakypyyygtshwyfdvwgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthl(seqidno:71)
在其它相关实施方式中,aav变体包含异源核酸,该异源核酸包含编码与seqidno:71的氨基酸21-525(对应于不具有下划线信号肽序列的sc-雷珠单抗lh的氨基酸序列)具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%同一性或100%同一性的氨基酸序列的核苷酸序列。
在另一个优选的实施方式中,提供了编码对应于图12e的、与人igg1的fc区融合的单链轻-重(lh)形式的雷珠单抗(sc-雷珠单抗-fc)的新型核酸序列,所述核酸序列包含以下或由以下组成:
atggtcctccagacacaggtgttcatcagcctgctgctgtggatctctggcgcctacggcgatatccagctgacacagagccctagcagcctgtctgcctctgtgggcgacagagtgaccatcacatgtagcgccagccaggacatcagcaactacctgaactggtatcagcagaagcccggcaaggcccctaaggtgctgatctactttaccagcagcctgcacagcggcgtgcccagcagattttctggctctggcagcggcaccgacttcaccctgacaatatctagcctccagcctgaggacttcgccacctactactgccagcagtacagcaccgtgccttggacatttggccagggcaccaaggtggaaatcaagcggacagtggccgctcctagcgtgttcatctttccacctagcgacgagcagctgaagtctggcacagcctctgtcgtgtgcctgctgaacaacttctaccccagagaagccaaggtgcagtggaaggtggacaacgccctccagagcggcaatagccaagagagcgtgaccgagcaggacagcaaggactctacctactctctgagcagcacactgaccctgagcaaggccgactacgagaagcacaaagtgtacgcctgcgaagtgacccaccagggcctttctagccctgtgaccaagagcttcaacagaggcgagtgtggcggcagctctggaagcggatctggaagcacaggcacaagctctagcggaaccggaacaagcgctggcacaacaggcacatctgccagcacaagcggttctggttctggcggaggcggaggatctggtggtggcggatctgctggcggaacagctacagctggtgcctcttctggatccgaggtgcagctggttgaatctggcggtggactggttcagccaggcggatctctgagactgtcttgtgccgccagcggctacgatttcacccactacggcatgaactgggtccgacaggcccctggcaaaggccttgaatgggtcggatggatcaacacctacaccggcgagccaacatacgccgccgacttcaagcggagattcaccttcagcctggacacctccaagagcaccgcctacctccagatgaacagcctgagagccgaggacaccgccgtgtactactgcgccaagtatccctactactacggcaccagccactggtacttcgacgtgtggggacagggcacactggtcacagtgtctagcgcctctacaaagggccccagcgttttcccactggctcctagcagcaagagcacatcaggcggaactgctgccctgggctgtctggtcaaggactactttcctgagcctgtgaccgtgtcctggaacagcggagcactgacatctggcgtgcacacctttccagccgtgctccaaagcagcggcctgtattctctgtccagcgtcgtgacagtgcctagcagctctctgggcacccagacctacatctgcaatgtgaaccacaagcctagcaacaccaaggtcgacaagaaggtggaacccaagagctgcgacaagacccacacctgtcctccatgtcctgctccagaactgctcggcggaccttccgtgttcctgtttcctccaaagcctaaggacaccctgatgatcagcagaacccctgaagtgacctgcgtggtggtggatgtgtcccacgaggatcccgaagtgaagttcaattggtacgtggacggcgtggaagtgcacaacgccaagaccaagcctagagaggaacagtacaatagcacctacagagtggtgtccgtgctgaccgtgctgcaccaggattggctgaacggcaaagagtacaagtgcaaggtgtccaacaaggccctgcctgctcctatcgagaaaaccatctccaaggccaagggccagcctagggaaccccaggtttacacactgcctccaagcagggacgagctgacaaagaaccaggtgtccctgacctgcctggtcaagggcttctacccttccgatatcgccgtggaatgggagagcaatggccagcctgagaacaactacaagacaacccctcctgtgctggacagcgacggctcattcttcctgtacagcaagctgacagtggacaagagcagatggcagcagggcaacgtgttcagctgctccgtgatgcacgaggccctgcacaaccactacacccagaagtccctgagcctgtctcctggcaaatgagccacgcgtaacacgtgcatgcgagagatctga(seqidno:72)。
在一些实施方式中,提供了编码sc-雷珠单抗-fc的核酸序列,其包含与seqidno:72中所示核酸序列的全长或与seqidno:72的核苷酸61-2277(不具有编码igκ信号序列的下划线核苷酸)具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%或更高的核酸同一性的核酸序列。在相关实施方式中,由主题aav变体递送的基因产物由核酸序列编码,所述核酸序列由seqidno:72的核酸序列或与seqidno:72的核酸序列具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%同一性的核酸组成,或包含其。在其它相关实施方式中,由主题aav变体递送的基因产物包含与下列氨基酸序列具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%同一性或100%同一性的氨基酸序列:
mvlqtqvfislllwisgaygdiqltqspsslsasvgdrvtitcsasqdisnylnwyqqkpgkapkvliyftsslhsgvpsrfsgsgsgtdftltisslqpedfatyycqqystvpwtfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgecggssgsgsgstgtsssgtgtsagttgtsastsgsgsgggggsggggsaggtatagassgsevqlvesggglvqpggslrlscaasgydfthygmnwvrqapgkglewvgwintytgeptyaadfkrrftfsldtskstaylqmnslraedtavyycakypyyygtshwyfdvwgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthldkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsrdeltknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk(seqidno:73)
在其它相关实施方式中,aav变体包含异源核酸,该异源核酸包含编码与seqidno:73的氨基酸21-752(对应于不具有下划线信号肽序列的sc-雷珠单抗-fc的氨基酸序列)具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%同一性或100%同一性的氨基酸序列的核苷酸序列。
在另一个优选的实施方式中,基因产物是溴珠单抗。溴珠单抗(rth258)是一种结合并阻断vegfa的所有同种型的单链可变片段(scfv)。溴珠单抗目前正处于iii期临床研究中,评价用于治疗湿性年龄相关性黄斑变性的3mg和6mg剂量。在特别优选的实施方式中,提供了编码溴珠单抗(对应于图12d)的新型密码子经优化的核酸序列,所述核酸序列包含以下或由以下组成:
atggtcctccagacacaggtgttcatcagcctgctgctgtggatctctggcgcctatggcgagatcgtgatgacacagagccccagcacactgtctgccagcgtgggagacagagtgatcatcacatgccaggccagcgagatcatccacagctggctggcttggtatcagcagaagcctggcaaggcccctaagctgctgatctacctggcctctacactggccagcggagtgcctagcagattttctggctctggatctggcgccgagttcaccctgacaatctctagcctccagcctgacgacttcgccacctactactgccagaacgtgtacctggccagcaccaacggcgccaattttggccagggcaccaagctgacagtgcttggcggaggcggaggttctggtggcggaggaagtggcggcggaggatcaggcggtggtggatctgaagtgcagctggtggaatcaggcggaggactggttcaacctggcggctctctgagactgagctgtaccgcctctggcttctccctgaccgactactactacatgacctgggtccgacaggcccctggcaaaggacttgagtgggtcggattcatcgaccccgacgacgatccttactacgccacatgggccaagggcagattcaccatcagccgggacaacagcaagaacaccctgtacctccagatgaacagcctgagagccgaggacaccgccgtgtactattgtgccggcggagatcacaatagcggctggggactcgatatctggggccagggaacactggtcaccgtgtctagttga(seqidno:74)。
在一些实施方式中,提供了编码溴珠单抗的核酸序列,所述核酸序列包含与seqidno:74中所示核酸序列的全长或与seqidno:74的核苷酸61-816(不具有编码igκ信号序列的下划线核苷酸)具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%或更高核酸同一性的核酸序列。在相关实施方式中,由主题aav变体递送的基因产物由核酸序列编码,所述核酸序列由seqidno:74的核酸序列或与seqidno:74的核酸序列具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%同一性的核酸组成,或包含其。在其它相关实施方式中,由主题aav变体递送的基因产物包含与下列氨基酸序列具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%同一性或100%同一性的氨基酸序列:
mvlqtqvfislllwisgaygeivmtqspstlsasvgdrviitcqaseiihswlawyqqkpgkapklliylastlasgvpsrfsgsgsgaeftltisslqpddfatyycqnvylastnganfgqgtkltvlgggggsggggsggggsggggsevqlvesggglvqpggslrlsctasgfsltdyyymtwvrqapgkglewvgfidpdddpyyatwakgrftisrdnskntlylqmnslraedtavyycaggdhnsgwgldiwgqgtlvtvss(seqidno:75)
在其它相关实施方式中,aav变体包含异源核酸,该异源核酸包含编码与seqidno:75的氨基酸21-271(对应于不具有下划线信号肽序列的溴珠单抗的氨基酸序列)具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%同一性或100%同一性的氨基酸序列的核苷酸序列。
在另一个优选的实施方式中,提供了编码对应于图12f的、与人igg1的fc区融合的溴珠单抗(溴珠单抗-fc)的新型核酸序列,所述核酸序列包含以下或由以下组成:
atggtcctccagacacaggtgttcatcagcctgctgctgtggatctctggcgcctatggcgagatcgtgatgacacagagccccagcacactgtctgccagcgtgggagacagagtgatcatcacatgccaggccagcgagatcatccacagctggctggcttggtatcagcagaagcctggcaaggcccctaagctgctgatctacctggcctctacactggccagcggagtgcctagcagattttctggctctggatctggcgccgagttcaccctgacaatctctagcctccagcctgacgacttcgccacctactactgccagaacgtgtacctggccagcaccaacggcgccaattttggccagggcaccaagctgacagtgcttggcggaggcggaggttctggtggcggaggaagtggcggcggaggatcaggcggtggtggatctgaagtgcagctggtggaatcaggcggaggactggttcaacctggcggctctctgagactgagctgtaccgcctctggcttctccctgaccgactactactacatgacctgggtccgacaggcccctggcaaaggacttgagtgggtcggattcatcgaccccgacgacgatccttactacgccacatgggccaagggcagattcaccatcagccgggacaacagcaagaacaccctgtacctccagatgaacagcctgagagccgaggacaccgccgtgtactattgtgccggcggagatcacaatagcggctggggactcgatatctggggccagggaacactggtcaccgtgtctagtgacaagacccacacctgtcctccatgtcctgctccagaactgctcggcggaccttccgtgttcctgtttcctccaaagcctaaggacaccctgatgatcagcagaacccctgaagtgacctgcgtggtggtggatgtgtcccacgaggatcccgaagtgaagttcaattggtacgtggacggcgtggaagtgcacaacgccaagaccaagcctagagaggaacagtacaatagcacctacagagtggtgtccgtgctgaccgtgctgcaccaggattggctgaacggcaaagagtacaagtgcaaggtgtccaacaaggccctgcctgctcctatcgagaaaaccatctccaaggccaagggccagcctagggaaccccaggtttacacactgcctccaagcagggacgagctgacaaagaaccaggtgtccctgacctgcctggtcaagggcttctacccttccgatatcgccgtggaatgggagagcaatggccagcctgagaacaactacaagacaacccctcctgtgctggacagcgacggctcattcttcctgtacagcaagctgacagtggacaagagcagatggcagcagggcaacgtgttcagctgctccgtgatgcacgaggccctgcacaaccactacacccagaagtccctgagcctgtctcctggcaaatgagccacgcgtaacacgtgcatgcgagagatctga(seqidno:76)。
在一些实施方式中,提供了编码溴珠单抗-fc的核酸序列,所述核酸序列包含与seqidno:76所示核酸序列的全长或与seqidno:76的核苷酸61-1530(不具有编码igκ信号序列的下划线核苷酸)具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%或更高核酸同一性的核酸序列。在相关实施方式中,由主题aav变体递送的基因产物由核酸序列编码,所述核酸序列由seqidno:76的核酸序列或与seqidno:76的核酸序列具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%同一性的核酸组成,或包含其。在其它相关实施方式中,由主题aav变体递送的基因产物包含与下列氨基酸序列具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%同一性或100%同一性的氨基酸序列:
mvlqtqvfislllwisgaygeivmtqspstlsasvgdrviitcqaseiihswlawyqqkpgkapklliylastlasgvpsrfsgsgsgaeftltisslqpddfatyycqnvylastnganfgqgtkltvlgggggsggggsggggsggggsevqlvesggglvqpggslrlsctasgfsltdyyymtwvrqapgkglewvgfidpdddpyyatwakgrftisrdnskntlylqmnslraedtavyycaggdhnsgwgldiwgqgtlvtvssdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsrdeltknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk(seqidno:77)。
在其它相关实施方式中,aav变体包含异源核酸,该异源核酸包含编码与seqidno:77的氨基酸21-498(对应于不具有下划线信号肽序列的溴珠单抗的氨基酸序列)具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%同一性或100%同一性的氨基酸序列的核苷酸序列。
在特别优选的实施方式中,提供了感染性重组aav(raav)病毒粒子,其包含变体aav衣壳蛋白和编码抑制vegf(优选vegfa)活性的多肽的异源核酸,所述变体aav衣壳蛋白相对于相应的亲本aav衣壳蛋白包含在衣壳蛋白的gh环中的肽插入,其中所述肽插入包含氨基酸序列isdqtkh(seqidno:14)或laisdqtkha(seqidno:28),其中与相应的亲本aav衣壳蛋白对视网膜细胞的感染性相比,所述变体衣壳蛋白赋予对视网膜细胞增加的感染性。在一些实施方式中,插入位点在对应于aav2的vp1(seqidno:2)的氨基酸587和588之间或在另一aav血清型的衣壳蛋白中的相应位置处的氨基酸。优选地,变体aav衣壳蛋白还包含相对于aav2的vp1衣壳(seqidno:2)的p34a氨基酸取代,或在另一种aav血清型中的相应取代。
在一些实施方式中,提供了包含变体aav衣壳蛋白和异源核酸的raav病毒粒子,所述变体aav衣壳蛋白包含(i)肽插入,其位于对应于aav2的vp1(seqidno:2)的氨基酸587和588之间,或在所述衣壳蛋白的另一aav血清型的衣壳蛋白中相对于相应亲本aav衣壳蛋白的相应位置,和(ii)相对于aav2的vp1衣壳(seqidno:2)的p34a氨基酸取代或在另一aav血清型中的相应取代,所述异源核酸包含编码阿柏西普的序列。在优选的实施方式中,编码阿柏西普的核酸序列由seqidno:65的核酸序列或与seqidno:65的核酸序列具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%同一性的核酸序列组成,或包含其。在特别优选的实施方式中,提供raav病毒粒子,其包含变体aav衣壳蛋白和包含seqidno:65的核酸序列的异源核酸,所述变体aav衣壳蛋白具有与seqidno:42所示的序列具有至少90%同一性、至少95%同一性或至少99%同一性的氨基酸序列,其中与相应亲本aav衣壳蛋白对视网膜细胞的感染性相比,所述变体衣壳蛋白赋予对视网膜细胞增加的感染性。在其它相关实施方式中,异源核酸还包含一个或多个序列,每个序列编码另外的vegfa抑制剂,所述抑制剂优选选自雷珠单抗、sc-雷珠单抗hl、sc-雷珠单抗lh、sc-雷珠单抗-fc、溴珠单抗和溴珠单抗-fc。在相关实施方式中,提供了包含这种raav的药物组合物。在其它相关实施方式中,提供了治疗vegfa相关眼病的方法,其包括向有需要的受试者施用有效量的raav病毒粒子,所述raav病毒粒子包含具有与seqidno:42所示序列具有至少90%同一性的氨基酸序列的变体aav衣壳蛋白和包含seqidno:65的核酸序列的异源核酸以及任选地一种或多种另外的核酸序列,所述另外的核酸序列各自编码不同的vegfa抑制剂。raav可通过视网膜下、脉络膜上、局部、前房内或玻璃体内注射来施用,但优选通过玻璃体内注射来施用。在一些实施方式中,vegfa相关眼病选自:湿性(新生血管性、渗出性)年龄相关性黄斑变性;视网膜静脉阻塞后的黄斑水肿;由视网膜静脉阻塞引起的视网膜新生血管形成;糖尿病性黄斑水肿、糖尿病性视网膜病变(包括所有阶段的非增殖性糖尿病性视网膜病变和增殖性糖尿病性视网膜病变)、近视性黄斑变性、分支视网膜静脉阻塞、半侧性视网膜静脉阻塞和中央视网膜静脉阻塞;早产儿视网膜病变;特发性脉络膜新生血管形成;近视性黄斑变性和继发性视网膜和脉络膜新生血管形成;视网膜毛细血管扩张;新生血管性青光眼;玻璃体出血;继发于视网膜疾病的视网膜和脉络膜新生血管形成,包括但不限于葡萄膜炎、创伤、视网膜变性病症、遗传性视网膜和/或脉络膜疾病、眼睛肿瘤、角膜和虹膜新生血管形成。在一些优选的实施方式中,vegfa相关眼病选自湿性(新生血管性、渗出性)年龄相关性黄斑变性;糖尿病性黄斑水肿;视网膜静脉阻塞后的黄斑水肿;糖尿病性视网膜病变;和近视性脉络膜新生血管形成。
在一些实施方式中,提供了包含变体aav衣壳蛋白和异源核酸的raav病毒粒子,所述变体aav衣壳蛋白包含(i)肽插入,其位于对应于aav2的vp1(seqidno:2)的氨基酸587和588的氨基酸之间,或在所述衣壳蛋白的另一aav血清型的衣壳蛋白中相对于相应亲本aav衣壳蛋白的相应位置,和(ii)相对于aav2的vp1衣壳(seqidno:2)的p34a氨基酸取代或在另一aav血清型中的相应取代,所述异源核酸包含编码雷珠单抗、sc-雷珠单抗hl、sc-雷珠单抗lh或sc-雷珠单抗-fc的核苷酸序列。在优选的实施方式中,编码sc-雷珠单抗的核酸序列由seqidno:67、69、70和72中任一个所示的核酸序列组成或包含其,或者与其具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%同一性。在特别优选的实施方式中,提供了raav病毒粒子,其包含变体aav衣壳蛋白和异源核酸,所述变体aav衣壳蛋白具有与seqidno:42所示序列具有至少90%同一性、至少95%同一性或至少99%同一性的氨基酸序列,所述异源核酸包含seqidno:67、69、70和72中任一个的核酸序列,其中与相应的亲本aav衣壳蛋白对视网膜细胞的感染性相比,所述变体衣壳蛋白赋予对视网膜细胞增加的感染性。在其它相关实施方式中,异源核酸还包含一种或多种另外的核酸序列,所述另外的核酸序列各自编码不同的vegfa抑制剂,所述vegfa抑制剂优选选自阿柏西普、溴珠单抗和溴珠单抗-fc。在相关实施方式中,提供了包含这种raav的药物组合物。在其它相关实施方式中,提供了治疗与眼内vegfa升高相关的眼病的方法,所述方法包括向有需要的受试者施用有效量的raav病毒粒子,所述raav病毒粒子包含变体aav衣壳蛋白和异源核酸序列,所述变体aav衣壳蛋白具有与seqidno:42所示序列具有至少90%同一性的氨基酸序列,所述异源核酸序列包含seqidno:67、69,70和72中任一个的核酸序列。优选地,raav通过玻璃体内注射施用。在一些实施方式中,vegfa相关眼病选自:湿性(新生血管性、渗出性)年龄相关性黄斑变性;视网膜静脉阻塞后的黄斑水肿;由视网膜静脉阻塞引起的视网膜新生血管形成;糖尿病性黄斑水肿、糖尿病性视网膜病变(包括所有阶段的非增殖性糖尿病性视网膜病变和增殖性糖尿病性视网膜病变)、近视性黄斑变性、分支视网膜静脉阻塞、半侧性视网膜静脉阻塞和中央视网膜静脉阻塞;早产儿视网膜病变;特发性脉络膜新生血管形成;近视性黄斑变性和继发性视网膜和脉络膜新生血管形成;视网膜毛细血管扩张;新生血管性青光眼;玻璃体出血;继发于视网膜疾病的视网膜和脉络膜新生血管形成,包括但不限于葡萄膜炎、创伤、视网膜变性病症、遗传性视网膜和/或脉络膜疾病、眼睛肿瘤、角膜和虹膜新生血管形成。在一些优选的实施方式中,vegfa相关眼病选自湿性(新生血管性、渗出性)年龄相关性黄斑变性;糖尿病性黄斑水肿;视网膜静脉阻塞后的黄斑水肿;糖尿病性视网膜病变;和近视性脉络膜新生血管形成。
在一些实施方案中,提供了包含变体aav衣壳蛋白和异源核酸的raav病毒粒子,所述变体aav衣壳蛋白包含(i)肽插入,其位于对应于aav2的vp1(seqidno:2)的氨基酸587和588的氨基酸之间,或在所述衣壳蛋白的另一aav血清型的衣壳蛋白中相对于相应亲本aav衣壳蛋白的相应位置,和(ii)相对于aav2的vp1衣壳(seqidno:2)的p34a氨基酸取代或在另一aav血清型中的相应取代,所述异源核酸包含编码溴珠单抗或溴珠单抗-fc的核苷酸序列。在优选的实施方式中,编码珠单抗或溴珠单抗-fc的核酸序列由seqidno:74或seqidno:76所示的核酸序列组成或包含其,或者与其具有至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%同一性。在特别优选的实施方式中,提供了raav病毒粒子,其包含变体aav衣壳蛋白和异源核酸,所述变体aav衣壳蛋白具有与seqidno:42所示序列具有至少90%同一性、至少95%同一性或至少99%同一性的氨基酸序列,所述异源核酸包含seqidno:74或seqidno:76的核酸序列,其中与相应的亲本aav衣壳蛋白对视网膜细胞的感染性相比,所述变体衣壳蛋白赋予对视网膜细胞增加的感染性。在其它相关实施方式中,异源核酸还包含一种或多种核苷酸序列,所述核苷酸序列各自编码不同的vegfa抑制剂,所述vegfa抑制剂优选选自阿柏西普、雷珠单抗、sc-雷珠单抗hl、sc-雷珠单抗lh和sc-雷珠单抗-fc。在相关实施方式中,提供了包含这种raav的药物组合物。在其它相关实施方式中,提供了治疗与眼内vegfa升高相关的眼病的方法,所述方法包括向有需要的受试者施用有效量的raav病毒粒子,所述raav病毒粒子包含变体aav衣壳蛋白和异源核酸序列,所述变体aav衣壳蛋白具有与seqidno:42所示序列具有至少90%同一性的氨基酸序列,所述异源核酸序列包含seqidno:74或seqidno:76的核酸序列。优选地,raav通过玻璃体内注射施用。在一些实施方式中,vegfa相关眼病选自:湿性(新生血管性、渗出性)年龄相关性黄斑变性;视网膜静脉阻塞后的黄斑水肿;由视网膜静脉阻塞引起的视网膜新生血管形成;糖尿病性黄斑水肿、糖尿病性视网膜病变(包括所有阶段的非增殖性糖尿病性视网膜病变和增殖性糖尿病性视网膜病变)、近视性黄斑变性、分支视网膜静脉阻塞、半侧性视网膜静脉阻塞和中央视网膜静脉阻塞;早产儿视网膜病变;特发性脉络膜新生血管形成;近视性黄斑变性和继发性视网膜和脉络膜新生血管形成;视网膜毛细血管扩张;新生血管性青光眼;玻璃体出血;继发于视网膜疾病的视网膜和脉络膜新生血管形成,包括但不限于葡萄膜炎、创伤、视网膜变性病症、遗传性视网膜和/或脉络膜疾病、眼睛肿瘤、角膜和虹膜新生血管形成。在一些优选的实施方式中,vegfa相关眼病选自湿性(新生血管性、渗出性)年龄相关性黄斑变性;糖尿病性黄斑水肿;视网膜静脉阻塞后的黄斑水肿;糖尿病性视网膜病变;和近视性脉络膜新生血管形成。
基因产物充当免疫调节剂的基因例如补体因子、toll样受体,被称为“免疫调节基因”。示例性的免疫调节基因包含细胞因子、趋化因子和对它们和/或其受体具有特异性的融合蛋白或抗体,例如抗il-6融合蛋白rilonacepttm、补体因子h特异性抗体拉帕单抗(lampamizumab)等。基因产物充当神经保护因子的基因例如:血小板源性生长因子受体(pdgfr);神经胶质源性神经营养因子(gdnf);视杆源性视锥生存力因子(rod-derivedconviabilityfactor,rdcvf);成纤维细胞生长因子(fgf);神经秩蛋白(ntn);睫状神经营养因子(cntf);神经生长因子(ngf);神经营养因子-4(nt4);脑源性神经营养因子(bdnf);表皮生长因子。基因产物充当光响应视蛋白的基因,例如视蛋白;视紫红质;通道视紫红质;晕光视紫红质。
在一些情况下,关注的基因产物是位点特异性内切核酸酶,其提供基因功能的位点特异性敲低,例如,其中内切核酸酶敲除与视网膜疾病相关的等位基因。例如,当显性等位基因编码基因的缺陷拷贝时,所述缺陷拷贝当为野生型时是视网膜结构蛋白和/或提供正常的视网膜功能,可以将位点特异性内切核酸酶靶向缺陷等位基因并敲除缺陷等位基因。
除了敲除缺陷性等位基因外,位点特异性核酸酶还可以用于刺激与供体dna的同源重组,所述供体dna编码由缺陷等位基因编码的蛋白质的功能性拷贝。因此,例如,主题raav病毒粒子可以用于递送敲除缺陷等位基因的位点特异性内切核酸酶,并且可以用于递送缺陷等位基因的功能性拷贝,从而导致缺陷等位基因的修复,由此提供功能性视网膜蛋白(例如,功能性视网膜劈裂蛋白、功能性rpe65、功能性外周蛋白等)的产生。参见li等人,(2011)nature475:217。在一些实施方式中,本文公开的raav病毒粒子包含编码位点特异性内切核酸酶的异源核苷酸序列;和编码缺陷等位基因的功能性拷贝的异源核苷酸序列,其中功能性拷贝编码功能性视网膜蛋白。功能性视网膜蛋白包含例如视网膜劈裂蛋白、rpe65、视网膜色素变性gtp酶调节剂(rgpr)-相互作用蛋白-1、外周蛋白、外周蛋白-2等。
适合使用的位点特异性内切核酸酶包含例如大范围核酸酶;锌指核酸酶(zfn);转录活化剂样效应核酸酶(talen);以及定期聚集的间隙短回文重复/crispr相关(cas),其中这些位点特异性核酸内切酶是非天然存在的,并被修饰以靶向特定基因。可以工程化这种位点特异性核酸酶以切割基因组内的特定位置,并且然后非同源末端连接可以在插入或缺失几个核苷酸时修复断裂。这样的位点特异性内切核酸酶(也称为“indel”)然后将蛋白质抛出框架并有效地敲除基因。参见,例如美国专利公开号2011/0301073。
在本文公开的变体raav载体的一些实施方式中,编码关注的基因产物的核苷酸序列与组成型启动子可操作地连接。合适的组成型启动子包含例如巨细胞病毒启动子(cmv)(stinski等人,(1985)journalofvirology55(2):431-441),cmv早期增强子/鸡β-肌动蛋白(cba)启动子/兔β-珠蛋白内含子(cag)(miyazaki等人,(1989)gene79(2):269-277,cbsb(jacobson等人,(2006)moleculartherapy13(6):1074-1084),人类延伸因子1α启动子(ef1α)(kim等人,(1990)gene91(2):217-223),人类磷酸甘油酸激酶启动子(pgk)(singer-sam等人,(1984)gene32(3):409-417,线粒体重链启动子(loderio等人,(2012)pnas109(17):6513-6518),泛素启动子(wulff等人,(1990)febsletters261:101-105)。在其它实施方式中,编码关注的基因产物的核苷酸序列与诱导型启动子可操作地连接。在一些情况下,编码关注的基因产物的核苷酸序列与组织特异性或细胞类型特异性调节元件可操作地连接。例如,在一些情况下,编码关注的基因产物的核苷酸序列与光感受器特异性调节元件(例如,光感受器特异性启动子)可操作地连接,所述光感受器特异性调节元件例如在感光细胞中赋予可操作连接基因的选择性表达的调节元件。合适的光感受器特异性调节元件包含例如视紫红质启动子;视紫红质激酶启动子(young等人,(2003)ophthalmol.vis.sci.44:4076);β磷酸二酯酶基因启动子(nicoud等人,(2007)j.genemed.9:1015);视网膜色素变性基因启动子(nicoud等人,(2007)同上);光感受器间维甲酸结合蛋白(irbp)基因增强子(nicoud等人,(2007)同上);irbp基因启动子(yokoyama等人,(1992)expeyeres.55:225),视蛋白基因启动子(tucker等人,(1994)pnas91:2611-2615),视网膜劈裂蛋白基因启动子(park等人,(2009)genetherapy16(7):916-926),crx同源域蛋白基因启动子(furukawa等人,(2002)thejournalofneuroscience22(5):1640-1647),鸟嘌呤核苷酸结合蛋白α转导活性多肽1(gnat1)基因启动子(lee等人,(2010)genetherapy17:1390-1399,神经视网膜特异性亮氨酸拉链蛋白(nrl)基因启动子(akimoto等人,(2006)pnas103(10):3890-3895),人类视锥抑制蛋白(hcar)启动子(li等人,(2002)biochemistryandmolecularbiology43:1375-1383),以及pr2.1、pr1.7、pr1.5和pr1.1启动子(ye等人,(2016)humangenetherapy27(1):72-82))。在一些情况下,编码关注的基因产物的核苷酸序列与视网膜色素上皮细胞(rpe)细胞特异性调节元件(例如,rpe特异性启动子)可操作地连接,所述细胞特异性调节元件例如在rpe细胞中赋予可操作连接的基因选择性表达的调节元件。合适的rpe特异性调节元件包含例如rpe65基因启动子(meur等人,(2007)genetherapy14:292-303),细胞视黄醛结合蛋白(cralbp)基因启动子(kennedy等人,(1998)journalofbiologicalchemistry273:5591-5598),色素上皮源性因子(pedfakaserpinf1)基因启动子(kojima等人,(2006)molecularandcellularbiochemistry293(1-2):63-69),和卵黄样黄斑营养不良(vmd2)启动子(esumi等人,(2004)thejournalofbiologicalchemistry279(18):19064-19073)。在一些情况下,编码关注的基因产物的核苷酸序列与穆勒神经胶质细胞特异性调节元件(例如,神经胶质特异性启动子)可操作地连接,所述细胞特异性调节元件例如在视网膜神经胶质细胞中赋予可操作连接基因的选择性表达的调节元件。合适的神经胶质特异性调节元件包含例如神经胶质原纤维酸性蛋白(gfap)启动子(besnard等人,(1991)journalofbiologicalchemistry266(28):18877-18883)。在一些情况下,编码关注的基因产物的核苷酸序列与双极细胞特异性调节元件(例如,双极特异性启动子)可操作地连接,所述细胞特异性调节元件例如在双极细胞中赋予可操作连接基因的选择性表达的调节元件。合适的双极特异性调节元件包含例如grm6启动子(cronin等人,(2014)embomolecularmedicine6(9):1175-1190)。
出于本发明的目的,本文的公开内容提供了分离的核酸,其包含编码如上所述的变体aav衣壳蛋白的核苷酸序列。分离的核酸可以是aav载体,例如重组aav载体。
本文的公开内容还提供了一种治疗视网膜疾病的方法,所述方法包括向有此需要的个体施用有效量的raav变体病毒粒子,其包含如上所述和本文公开的关注的转基因。本领域普通技术人员将能够容易地确定主题raav病毒粒子的有效量,并且通过测试一个或多个功能或解剖学参数的变化来治疗疾病,所述参数例如视敏度、视野、对亮和暗的电生理学反应、色觉、对比敏感度、解剖学、视网膜健康和脉管系统、眼能动性、固定偏好和稳定性。
用于评估视网膜功能和其变化的非限制性方法包括:评估视敏度(例如最佳矫正视力[bcva]、移动、导航、物体检测和辨别);评估视野(例如静态和动态视野测量);进行临床检查(例如,对眼睛的前段和后段进行裂隙灯检查);评估对所有亮和暗波长的电生理反应(例如所有形式的视网膜电图(erg)[全视角、多焦点和模式]、所有形式的视觉诱发电位(vep)、眼电图(eog)、色觉、暗适应和/或对比敏感度)。用于评估解剖学和视网膜健康和其变化的非限制性方法包含光学相干断层扫描(oct)、眼底摄影、自适应光学扫描激光检眼镜检查(ao-slo)、荧光和/或自发荧光;测量眼球动性和眼球运动(例如,眼球震颤、固定偏好和稳定性)、测量报告的结果(患者报告的视觉和非视觉引导行为和活动的变化、患者报告的结果[pro]、基于问卷的生活质量评估、日常活动和神经功能的测量(例如功能性磁共振成像(mri))。
在一些实施方式中,有效量的主题raav病毒粒子导致视网膜功能、解剖学完整性或视网膜健康的丧失速率降低,例如丧失速率以及因此疾病的进展降低2倍、3倍、4倍或5倍或者更多,例如丧失速率以及因此疾病的进展降低10倍或更多。在一些实施方式中,有效量的主题raav病毒粒子导致视觉功能、视网膜功能、视网膜解剖学或健康的改善、和/或眼能动性的改善和/或神经功能的改善增加,例如视网膜功能、视网膜解剖学或健康和/或眼能动性的改善提高2倍、3倍、4倍或5倍或者更高,例如视网膜功能、视网膜解剖学或健康的改善和/或眼能动性的改善提高10倍或更多。如普通技术人员将容易理解的,实现期望的治疗效果所需的剂量通常在1x108到约1x1015个重组病毒粒子的范围内,通常被普通技术人员称为1x108到约1x1015个“载体基因组”。
主题raav病毒粒子可以通过眼内注射施用,例如通过玻璃体内注射、通过视网膜下注射、通过脉络膜上注射或通过将导致raav病毒粒子递送至眼睛的任何其它方便的施用模式或途径。其它方便的施用模式或途径包括但不限于静脉内、动脉内、眼周、前房内、结膜下和眼球囊下(sub-tenons)注射和局部施用和鼻内施用。当通过玻璃体内注射施用时,主题raav病毒粒子能够穿过玻璃体并穿过内界膜(在本文中也称为内界膜,或“ilm”;视网膜的表面上的形成视网膜与玻璃体之间的边界的薄的、透明的无细胞膜,其由星形胶质细胞和穆勒细胞的终足形成),和/或与包含相应亲本aav衣壳蛋白的aav病毒粒子的能力相比,更有效地移动通过视网膜层。
本文公开的变体衣壳蛋白被分离,例如纯化。在一些实施方式中,本文公开的变体衣壳蛋白包括在aav载体或重组aav(raav)病毒粒子中。在其它实施方式中,这种aav变体载体和/或aav变体病毒粒子用于治疗灵长类动物视网膜中的眼病的体内或离体方法。
本文的公开内容还提供了宿主细胞,例如但不限于包含主题核酸的分离的(遗传修饰的)宿主细胞。本文公开的根据本发明的宿主细胞可以是分离的细胞,如来自体外细胞培养物的细胞。如本文所述,这种宿主细胞用于产生主题raav变体病毒粒子。在一个实施方式中,用核酸对这种宿主细胞进行稳定地遗传修饰。在其它实施方式中,用核酸对宿主细胞进行瞬时遗传修饰。使用已建立的技术将这种核酸稳定地或瞬时地引入宿主细胞中,所述技术包括但不限于电穿孔、磷酸钙沉淀、脂质体介导的转染等。为了稳定转化,核酸通常还包含可选择标记,例如,几种众所周知的可选择标记中的任何一种,如新霉素抗性等。通过将核酸引入多种细胞中的任何细胞中来产生这种宿主细胞,所述多种细胞例如哺乳动物细胞,包括例如鼠类细胞和灵长类动物细胞(例如人类细胞)。示例性的哺乳动物细胞包括但不限于原代细胞和细胞系,其中示例性的细胞系包括但不限于293细胞、cos细胞、hela细胞、vero细胞、3t3小鼠成纤维细胞、c3h10t1/2成纤维细胞、cho细胞等。示例性的宿主细胞包括但不限于hela细胞(例如,美国典型培养物保藏中心(atcc)编号ccl-2)、cho细胞(例如,atcc编号crl9618、ccl61、crl9096)、293细胞(例如,atcc编号crl-1573)、vero细胞、nih3t3细胞(例如,atcc编号crl-1658)、huh-7细胞、bhk细胞(例如,atcc编号ccl10)、pc12细胞(atcc编号crl1721)、cos细胞、cos-7细胞(atcc编号crl1651)、rat1细胞、小鼠l细胞(atcc编号ccli.3)、人胚肾(hek)细胞(atcc编号crl1573)、hlhepg2细胞等等。还可以使用杆状病毒来感染宿主细胞以感染昆虫细胞,如sf9细胞,其产生aav(参见,例如,美国专利号7,271,002;美国专利申请序列号12/297,958)。在一些实施方式中,如上所述,除了包含编码变体aav衣壳蛋白的核苷酸序列的核酸之外,遗传修饰的宿主细胞包括包含编码一种或多种aavrep蛋白的核苷酸序列的核酸。在其它实施方式中,宿主细胞进一步包含raav变体载体。可以使用此类宿主细胞产生raav变体病毒粒子。例如美国专利公开号2005/0053922和美国专利公开号2009/0202490中描述了产生raav病毒粒子的方法。
本公开在本文中另外提供了一种药物组合物,其包含:a)如上所述和本文公开的raav变体病毒粒子;b)药学上可接受的载剂、稀释剂、赋形剂或缓冲剂。在一些实施方式中,药学上可接受的载剂、稀释剂、赋形剂或缓冲剂适用于人或非人患者。此类赋形剂、载剂、稀释剂和缓冲剂包括可以在没有过度毒性的情况下施用的任何试剂。药学上可接受的赋形剂包含但不局限于液体,例如水、盐水、甘油、以及乙醇。其中可以包括药学上可接受的盐,例如无机酸盐,如盐酸盐、氢溴酸盐、磷酸盐、硫酸盐等;和有机酸的盐,如乙酸盐、丙酸盐、丙二酸盐、苯甲酸盐等。此外,在这种载剂中可以存在辅助物质,如润湿剂或乳化剂、ph缓冲物质等。多种药学上可接受的赋形剂是本领域已知的,无需在本文详细讨论。已在各种出版物中充分描述了药学上可接受的赋形剂,包含例如a.gennaro(2000)“remington:thescienceandpracticeofpharmacy”,第20版,lippincott,williams,&wilkins;pharmaceuticaldosageformsanddrugdeliverysystems(1999)h.c.ansel等人,eds.,7thed.,lippincott,williams,&wilkins;和handbookofpharmaceuticalexcipients(2000)a.h.kibbe等人,eds.,3rded.amer.pharmaceuticalassoc。在本发明的一些方面,本发明提供了一种药物组合物,其包含约1×108到约1×1015个重组病毒或1×108到约1×1015个载体基因组,其中每个所述重组病毒包含编码一种或多种基因产物的基因组。
本发明的一些实施方式在以下项目1至36中举例说明:
1.一种变体腺相关病毒(aav)衣壳蛋白,所述变体腺相关病毒(aav)衣壳蛋白包含相对于相应的亲本aav衣壳蛋白在所述衣壳蛋白的gh环中的肽插入,其中所述肽插入包含氨基酸序列isdqtkh(seqidno:14),并且其中与所述相应的亲本aav衣壳蛋白对视网膜细胞的感染性相比,所述变体衣壳蛋白赋予对视网膜细胞增加的感染性。
2.根据项目1所述的变体aav衣壳蛋白,其中所述肽插入包含氨基酸序列y1y2isdqtkhy3,其中y1-y3中的每一个独立地选自ala、leu、gly、ser、thr和pro。
3.根据项目2所述的变体aav衣壳蛋白,其中所述肽插入包含氨基酸序列laisdqtkha(seqidno:28)。
4.根据项目1-3中任一项所述的变体aav衣壳蛋白,其中插入位点位于对应于aav2的vp1(seqidno:2)的氨基酸587和588的氨基酸之间或在另一aav血清型的衣壳蛋白中的相应位置。
5.根据项目1-4中任一项所述的变体aav衣壳蛋白,其中所述衣壳蛋白包含相对于aav2的vp1衣壳(seqidno:2)的一个或多个氨基酸取代或在另一aav血清型中的一个或多个相应取代。
6.根据项目1-5中任一项所述的变体aav衣壳蛋白,其中所述衣壳蛋白包含相对于aav2的vp1衣壳(seqidno:2)的p34a氨基酸取代或在另一aav血清型中的相应取代。
7.根据项目1-6中任一项所述的变体aav衣壳蛋白,其中所述衣壳蛋白包含(i)氨基酸序列isdqtkh(seqidno:14)和(ii)p34a氨基酸取代,并且与seqidno:42所示的序列具有至少90%同一性、至少95%同一性或至少99%同一性。
8.根据项目7所述的变体aav衣壳蛋白,其中所述衣壳蛋白基本上由seqidno:42所示的氨基酸序列组成。
9.一种重组aav(raav)病毒粒子,其包含根据项目1至8中任一项所述的变体aav衣壳蛋白和异源核酸,所述异源核酸包含编码基因产物的核苷酸序列。
10.根据项目9所述的raav,其中所述基因产物是多肽。
11.根据项目10所述的raav,其中所述异源核酸包含编码抑制血管内皮生长因子(vegf)的活性的多肽的核苷酸序列。
12.根据项目11所述的raav,其中所述核苷酸序列编码融合蛋白。
13.根据项目12所述的raav,其中所述核苷酸序列编码阿柏西普(eylea)。
14.根据项目13所述的raav,其中所述核苷酸序列与seqidno:65所示的核酸序列或与seqidno:65的核苷酸79-1377具有至少90%同一性、至少95%同一性或至少99%同一性,且优选编码seqidno:66所示的氨基酸序列或seqidno:66的氨基酸27-458。
15.根据项目11所述的raav,其中所述核苷酸序列编码单克隆抗体或其抗原结合片段。
16.根据项目15所述的raav,其中所述核苷酸序列编码雷珠单抗(lucentis)。
17.根据项目15所述的raav,其中所述核苷酸序列(i)包含与seqidno:67(sc-雷珠单抗hl)的序列或与seqidno:67的核苷酸58-1575具有至少90%同一性、至少95%同一性或至少99%同一性的序列,并且编码seqidno:68所示的氨基酸序列,或(ii)编码seqidno:68的氨基酸20-524的氨基酸序列。
18.根据项目15所述的raav,其中所述核苷酸序列(i)具有与seqidno:69(sc-雷珠单抗lh1)或seqidno:70(sc-雷珠单抗lh2)的序列或者与seqidno:69或seqidno:70的核苷酸61-1578具有至少90%同一性、至少95%同一性或至少99%同一性的序列,且编码seqidno:71所示的氨基酸序列,或(ii)编码seqidno:71的氨基酸21-525的氨基酸序列。
19.根据项目15所述的raav,其中所述核苷酸序列(i)具有与seqidno:72(sc-雷珠单抗-fc)的序列或与seqidno:72的核苷酸61-2277具有至少90%同一性、至少95%同一性或至少99%同一性的序列,并且编码seqidno:73所示的氨基酸序列,或(ii)编码seqidno:73的氨基酸21-752的氨基酸序列。
20.根据项目15所述的raav,其中所述核苷酸序列编码溴珠单抗。
21.根据项目20所述的raav,其中所述核苷酸序列具有与seqidno:74所示的核酸序列或与seqidno:74的核苷酸61-816具有至少90%同一性、至少95%同一性或至少99%同一性的序列,且优选编码seqidno:75所示的氨基酸序列或seqidno:75的氨基酸21-271的氨基酸序列。
22.根据项目15所述的raav,其中所述核苷酸序列(i)具有与seqidno:76(溴珠单抗-fc)的序列或与seqidno:76的核苷酸61-1530具有至少90%同一性、至少95%同一性或至少99%同一性的序列,并且编码seqidno:77所示的氨基酸序列,或(ii)编码seqidno:77的氨基酸21-498的氨基酸序列。
23.根据项目11所述的raav,其中所述异源核酸包含(i)编码阿柏西普的序列,优选地其中编码阿柏西普的序列与seqidno:65所示的核酸序列具有至少90%同一性、至少95%同一性或至少99%同一性,且编码seqidno:66所示的氨基酸序列,和(ii)编码溴珠单抗的序列,优选地其中编码溴珠单抗的序列与seqidno:74或seqidno:76所示的核酸序列具有至少90%同一性、至少95%同一性或至少99%同一性,且优选地编码seqidno:75或seqidno:77所示的氨基酸序列。
24.根据项目9-23中任一项所述的raav,其中编码基因产物的核苷酸序列可操作地连接至表达控制序列。
25.一种药物组合物,其包含根据项目11-24中任一项的raav和药学上可接受的载体。
26.一种用于将vegf抑制剂递送至受试者的视网膜细胞、脉络膜细胞、晶状体细胞、睫状细胞、虹膜细胞、视神经细胞和/或角膜细胞的方法,其包括向所述受试者施用根据项目11-24中任一项的raav病毒粒子或根据权利要求24所述的药物组合物。
27.根据项目26所述的方法,其中向所述受试者玻璃体内施用所述raav病毒粒子或药物组合物。
28.一种治疗需要治疗vegfa相关眼病的受试者中的vegfa相关眼病的方法,所述vegfa相关眼病选自湿性(新生血管性、渗出性)年龄相关性黄斑变性;视网膜静脉阻塞后的黄斑水肿;由视网膜静脉阻塞引起的视网膜新生血管形成;糖尿病性黄斑水肿,糖尿病性视网膜病变(包括所有阶段的非增殖性糖尿病性视网膜病变和增殖性糖尿病性视网膜病变);近视性黄斑变性;分支视网膜静脉阻塞、半侧性视网膜静脉阻塞和中央视网膜静脉阻塞;早产儿视网膜病变;特发性脉络膜新生血管形成;近视性黄斑变性和继发性视网膜和脉络膜新生血管形成;视网膜毛细血管扩张;新生血管性青光眼;玻璃体出血;继发于视网膜疾病的视网膜和脉络膜新生血管形成,包括但不限于葡萄膜炎、创伤、视网膜变性病症、遗传性视网膜和/或脉络膜疾病、眼睛肿瘤、角膜和虹膜新生血管形成,所述方法通过向所述受试者施用有效量的根据项目11-24中任一项所述的raav或根据权利要求25所述的药物组合物来进行
29.根据项目28所述的方法,其中所述vegfa相关眼病选自湿性(新生血管性、渗出性)年龄相关性黄斑变性;糖尿病性黄斑水肿;视网膜静脉阻塞后的黄斑水肿;糖尿病性视网膜病变;和近视性脉络膜新生血管形成。
30.根据项目26至29中任一项所述的方法,其中所述raav包含基本上由seqidno:42所示的氨基酸序列组成的衣壳蛋白和包含编码阿柏西普的序列的异源核酸,优选地其中编码阿柏西普的序列具有seqidno:65所示的序列。
31.根据项目26至29中任一项所述的方法,其中所述raav包含基本上由seqidno:42所示的氨基酸序列组成的衣壳蛋白和包含编码雷珠单抗、sc-雷珠单抗hl、sc-雷珠单抗lh或sc-雷珠单抗-fc的序列的异源核酸,优选地其中编码sc-雷珠单抗hl、sc-雷珠单抗lh或sc-雷珠单抗-fc的序列具有seqidno:67、69、70和72中任一个所示的序列。
32.根据项目26至29中任一项所述的方法,其中所述raav包含基本上由如seqidno:42所示的氨基酸序列组成的衣壳蛋白和包含编码溴珠单抗或溴珠单抗-fc的序列的异源核酸,优选地其中所述编码溴珠单抗或溴珠单抗-fc的序列具有seqidno:74或seqidno:76所示的序列。
33.根据项目28-32中任一项所述的方法,其中向所述受试者玻璃体内施用所述raav或药物组合物。
34.根据项目26-33中任一项所述的方法,其中所述受试者是人。
35.一种分离的核酸,其包含编码根据项目1-8中任一项的变体aav衣壳蛋白的核苷酸序列。
36.一种分离的、遗传修饰的宿主细胞,其包含根据项目35所述的核酸。
实施例
提出以下实施例以便向本领域普通技术人员提供如何制备和使用本发明的完整披露和描述,并且不旨在限制诸位发明人看待其发明的范围,它们也不旨在表示以下实验是进行的全部或仅有的实验。虽然已尽力确保所用数字(例如量、温度等)的准确性,但仍应考虑一些实验误差和偏差。除非另外指明,份数是重量份,分子量是重量平均分子量,温度是摄氏度,并且压力是大气压或接近大气压。
可以在这种标准教科书中找到分子和细胞生物化学的一般方法,例如,molecularcloning:alaboratorymanual,第3版,(sambrook等人,harborlaboratorypress2001);shortprotocolsinmolecularbiology第4版,(ausubel等人,eds.,johnwiley&sons1999);proteinmethods(bollag等人,johnwiley&sons1996);nonviralvectorsforgenetherapy(wagner等人,eds.,academicpress1999);viralvectors(kaplift&loewyeds.,academicpress1995);immunologymethodsmanual(i.lefkovitsed.,academicpress1997);以及cellandtissueculture:laboratoryproceduresinbiotechnology(doyle&griffiths,johnwiley&sons1998)。其公开内容通过引用并入本文。本公开中提及的用于遗传操纵的试剂、克隆载体和试剂盒可从商业供应商获得,如biorad、stratagene、invitrogen、sigma-aldrich和clontech。
实施例1
玻璃体内注射和组织收获。通过玻璃体内注射通过巩膜(使用适于人类使用的程序和递送装置在角膜缘后约3mm)为年龄为4到10岁并且体重至少为4kg的单一雄性食蟹猴(macacafascicularis)给药。将动物麻醉,并给予局部麻醉剂。将100μl文库施用到每一只眼睛。
在第14±3天,由经过培训的兽医人员使用100mg/kg戊巴比妥钠静脉注射进行安乐死。将眼睛成核并储存在4℃下直至解剖。
组织解剖。用手术刀沿锯齿锯缘切开眼睛,并移除前段。在中央凹周围的视网膜上进行减压切割,以使视网膜能够平坦安装,并移除玻璃体。收集来自每个象限(上、下、鼻和颞)的六个视网膜样品,如图2所示,分离出对应于rpe细胞的细胞材料、光感受器、生物极细胞、无长突细胞、水平细胞和/或神经节细胞。
定向进化。图1a-图1e中示出了定向进化过程。简而言之,创建了包含20多种dna突变技术和cap基因的专有组合的病毒衣壳文库(图1a)。然后包装病毒(图1b)—使得每个颗粒由围绕编码该衣壳的cap基因的突变衣壳组成—并且被纯化。将衣壳文库置于体内选择压力下。收获感兴趣的组织或细胞材料以分离已成功感染该靶标的aav变体,并回收成功的病毒。通过重复选择来富集成功的克隆(阶段i到图1d)。选择的cap基因然后经历专有的再分散,并通过进一步的选择步骤富集以迭代地增加病毒适应性(阶段2–图1d)。在载体选择阶段1和2期间识别的变体证明了转导灵长类动物视网膜细胞的能力(图1e)。
aav衣壳基因组成功回收。第1到6轮。从每轮选择中回收的衣壳用于包装注射的文库以开始随后的一轮选择。从组织中回收衣壳基因表示文库载体成功地内化到感兴趣的组织中。在第4轮之后,在文库包装和第5轮注射之前,将文库的额外重新多样化纳入其中。图3中示出了来自代表性选择轮次的rpe、pr、内核层(inl)和神经节细胞层(gcl)视网膜组织的病毒基因组的回收。盒内的条带代表病毒基因组的成功回收。
测序分析:第3到6轮。在第3到6轮期间,对文库内的单个克隆进行测序以确定群体内变体的频率。评估变体在测序数据中是否存在基序。基于在多个序列中发生的统一变异(例如,衣壳内一致位置中的特异性点突变或特异性肽插入序列)的存在,将变体分组为基序。图4a(第3轮测序分析)、图4b(第4轮测序分析)、图4c(第5轮测序分析)和4d(第6轮测序分析)中表示了代表两轮或更多轮选择中至少5%的测序群体或一轮或更多轮选择中至少10%的测序群体的基序。
下表1中列出了赋予对视网膜细胞增加的感染性的识别的若干个代表性克隆(每个克隆含有识别的一个或多个取代和/或肽插入,并且在其它方面与seqidno:2一致;列出了针对每个克隆的选择轮次、序列编号和频率(在括号中):
表1.赋予对视网膜的一种或多种细胞增加的感染性的aavvp1衣壳蛋白的氨基酸序列修饰。第2列中列出的取代基于野生型aav2的氨基酸序列,即不存在插入的肽。
还识别为对一种或多种视网膜细胞具有增加的感染性的衣壳是具有以下祖先vp1衣壳序列的克隆:
maadgylpdwlednlsegirewwdlkpgapkpkanqqkqddgrglvlpgykylgpfngldkgepvnaadaaalehdkaydqqlkagdnpylrynhadaefqerlqedtsfggnlgravfqakkrvleplglveegaktapgkkrpvepspqrspdsstgigkkgqqpakkrlnfgqtgdsesvpdpqplgeppagpsglgsgtmaagggapmadnnegadgvgnasgnwhcdstwlgdrvittstrtwalptynnhlykqissasagstndnhyfgystpwgyfdfnrfhchfsprdwqrlinnnwgfrpkrlnfklfniqvkevttndgvttiannltstvqvfsdseyqlpyvlgsahqgclppfpadvfmipqygyltlnngsqavgrssfycleyfpsqmlrtgnnftfsytfedvpfhssyahsqsldrlmnplidqylyylartqstggtagtrellfsqagpsnmsaqaknwlpgpcyrqqrvsktlsqnnnsnfawtgatkyhlngrdslvnpgvamathkddedrffpssgvlifgkqgaganntalenvmmtseeeikttnpvateqygvvasnlqssntapvtgtvnsqgalpgmvwqnrdvylqgpiwakiphtdgnfhpsplmggfglkhpppqilikntpvpanppavftpakfasfitqystgqvsveiewelqkenskrwnpeiqytsnyakstnvdfavdnegvyseprpigtryltrnl.(seqidno:59)
这种祖先衣壳变体是从祖先衣壳seqidno:58进化而来的,其中简并性位置(残基264、266、268、448、459、460、467、470、471、474、495、516、533、547、551、555、557、561、563、577、583、593、596、661、662、664、665、710、717、718、719、723)进化成包含:264处的丙氨酸(a)、266处的丙氨酸(a)、268处的丝氨酸(s)、448处的丙氨酸(a)、459处的苏氨酸(t)、460处的精氨酸(r)、467处的丙氨酸(a)、470处的丝氨酸(s)、471处的天冬酰胺(n)、474处的丙氨酸(a)、495处的丝氨酸(s)、516处的天冬酰胺(d)、533处的天冬酰胺(d)、547处的谷氨酰胺(q)、551处的丙氨酸(a)、555处的丙氨酸(a)、557处的谷氨酸(e)、561处的甲硫氨酸(m)、563处的丝氨酸(s)、577处的谷氨酰胺(q)、583处的丝氨酸(s)、593处的缬氨酸(v)、596处的苏氨酸(t)、661处的丙氨酸(a)、662处的缬氨酸(v)、664处的苏氨酸(t)、665处的脯氨酸(p)、710处的苏氨酸(t)、717处的天冬氨酸(d)、718处的天冬酰胺(n)、719处的谷氨酸(e)、以及723处的丝氨酸(s)。
本文公开的aav变体病毒粒子可以并入合理的合理设计参数、特征、修饰、优点和变化,这些是本领域技术人员在工程化aav病毒载体领域中显而易见的。
实施例2
采用定向进化来发现在玻璃体内(ivt)施用后对视网膜细胞具有优异基因递送的新型腺相关病毒(aav)变体,玻璃体内(ivt)施用是显著优于将基因递送到人眼的其它方法的施用途径(实施例1)。玻璃体内施用包含p34a取代和插入氨基酸588处的肽laisdqtkha(seqidno:28)的新型aav变体(laisdqtkha+p34a;seqidno:42)后的细胞趋向性,在非人灵长类动物(nhp)中体内评估为含有isdqtkh(seqidno:14)的aav变体转导视网膜细胞的能力的代表性实例。
使用标准方法制造重组aav病毒粒子,其包含aav2衣壳或新型变体衣壳laisdqtkha+p34a(seqidno:42)以及包含与cmv启动子(分别为aav2.cmv.gfp和laisdqtkha+p34a.cmv.gfp)或cag启动子(分别为aav2.cag.egfp和laisdqtkha+p34a.cag.egfp)可操作地连接的绿色荧光蛋白(gfp)转基因的基因组。非洲绿猴(图7、图8)或食蟹猴(图9)玻璃体内注射有各种剂量的载体,每只眼睛注射范围为4×1010vg到1×1012vg(详见图例),并且用heidelbergspectralistm通过眼底荧光成像评估视网膜细胞转导寿命。
包含新型变体laisdqtkha+p34a(seqidno:42)的aav的玻璃体内递送导致跨nhp视网膜的转基因表达比aav2更广泛且更稳健(图7-图9)。图像显示新型aav变体衣壳在中央凹(富含视锥的区域)的中心内提供稳健的表达;早在注射后2周,在旁中心凹环(富含视网膜神经节细胞的区域)和在周边(富含许多类型的细胞的区域,包含视杆、穆勒胶质细胞、无长突细胞,双极细胞)中。相比之下并且与由其它人报道的结果一致,野生型aav2提供较弱的表达,其主要在旁中心凹环中并且仅可以在稍后的时间点检测到。注射后3周对视网膜的各个区域进行的免疫组织化学分析证实,已经在整个视网膜中成功转导了许多类型的视网膜细胞,包含视网膜色素上皮细胞、视杆和视锥光感受器、以及视网膜神经节细胞(图10a-图10e)。
此研究表明了与临床相关的aav2相比在临床上优选的施用途径后包含isdqtkh的变体的优异基因递送。用包含此肽插入基序的其它变体可实现类似的功效。同样地,使用本文中公开的使用相同的定向进化方法识别的其它变体可以实现类似的功效。
实施例3
用于视网膜色素上皮(rpe)细胞和光感受器(pr)细胞的新型aav变体laisdqtkha+p34a(seqidno:42)的细胞趋向性,在体外使用rpe细胞和由成纤维细胞源性人诱导多能干细胞(fb-ipsc)或人胚胎干细胞(esc)产生的pr细胞进行评估。
使用标准方法来制造aav病毒粒子,其包括aav2衣壳或新型变体衣壳laisdqtkha+p34a(seqidno:42)和包含与cag启动子(分别为aav2.cag.egfp和laisdqtkha+p34a.cag.egfp)可操作地连接的绿色荧光蛋白(egfp)转基因的基因组。使用45天的分化方案,由人胚胎干细胞系esi-017或人成纤维细胞源性诱导多能干细胞(“fb-ipsc”)产生人rpe细胞培养物。通过检测以下来确认成熟为rpe细胞:包含rpe65和best1的成熟rpe标记物的表达;vegf和pedf的合成;和吞噬视杆外段的能力。通过多步眼杯形成范例产生pr培养物,并通过在培养179天后检测恢复蛋白和s视蛋白的表达而确认包含pr。
相对于aav2,如通过免疫荧光(图11a至图11b)、流式细胞术(增加2.7倍;图11c至图11d)和蛋白质印迹分析(图11e至图11f)所确定的,laisdqtkha+p34a(seqidno:42)在感染后7天在人rpe培养物中提供显著更高的转导效率和转基因表达。在感染后32天,使用laisdqtkha+p34a.cag.egfp在人pr培养物中同样实现了稳健的转导和表达。此研究说明了包含isdqtkh(seqidno:14)的变体将基因递送至视网膜细胞的优异能力。
实施例4
抗vegf表达载体的设计和构建。阿柏西普的氨基酸序列由与人igg1的fc区融合的人flt1信号肽、vegfr1结构域2和vegfr2结构域3组成(图12a;seqidno:66)。通过柔性蛋白质接头连接雷珠单抗的轻链和重链的氨基酸序列,以将双链抗原结合片段(fab)转化为单链fab(scfab)。轻-重(lh)形式由通过柔性肽连接的人igκ轻链信号肽、雷珠单抗的可变轻链、恒定轻链、可变重链和恒定重链1结构域组成(图12b;seqidno:71)。与lh形式相比,重-轻(hl)形式是相似的,除了信号肽来源于人igg重链并且重链和轻链结构域在接头的相反侧(图12c;seqidno:68)。溴珠单抗设计包括通过柔性肽连接的可变轻链和可变重链结构域,以及用于从哺乳动物细胞分泌的人igκ轻链信号肽(图12d;seqidno:75)。为了改善从人细胞的表达,对开放阅读框进行密码子优化,通过geneart或genscript进行合成,并提供在标准质粒克隆载体中。用限制酶psti和bglii从该质粒切下感兴趣的dna,并将其插入paav-cag-sv40pa质粒中的cag启动子和sv40polya信号之间。sc-雷珠单抗-fc设计由与来自阿柏西普设计的人igg1的fc区融合的lh形式sc-雷珠单抗组成(图12e;seqidno:73)。溴珠单抗-fc设计由与来自阿柏西普设计的人igg1的fc区融合的溴珠单抗组成(图12f;seqidno:77)。通过pcr从起始克隆载体扩增sc-雷珠单抗lh、溴珠单抗和fc区,所述起始克隆载体具有在3'末端(sc-雷珠单抗lh和溴珠单抗)或5'末端(fc)含有iis型限制酶bsmbi位点的延伸。将得到的pcr产物用限制酶psti和bsmbi(sc-雷珠单抗lh和溴珠单抗)或bsmbi和bglii(fc)消化,并插入paav-cag-sv40pa质粒中的cag启动子和sv40polya信号之间。使用连接反应物来转化大肠杆菌,并通过限制性消化来鉴定阳性克隆。然后使克隆以更大的规模生长,并用qiagen无内切maxiprep试剂盒纯化质粒dna。通过限制酶切消化和测序验证质粒的身份。paav-cag-sv40polya质粒含有aav2反向末端重复(itr),其允许抗vegf序列被包装在aav衣壳内并通过玻璃体内施用来递送至视网膜中的感兴趣的细胞。
在两个独立的实验中,使用fugene6转染试剂,用含有在cag启动子控制下的gfp或抗vegf转基因(sc-rnbzhl(seqidno:67)、sc-rnbzlh1(seqidno:69)和sc-rnbz-lh2(seqidno:70)、aflb(seqidno:65)、bro(seqidno:74))的质粒模拟转染或转染hek293t细胞。转染后48小时收集培养基,用阿柏西普elisa试剂盒(eaglebiosciences,immunoguideig-aa115)进行测定。通过在试剂盒中作为试剂提供的hrp-抗-iggfc(模拟、gfp和aflb)或hrp-抗-iggh+l(sc-rnbzhl、sc-rnbzlh1和sc-rnbzlh2)检测抗-vegf蛋白。相对于临床eylea(模拟、gfp和aflb)或lucentis(sc-rnbzhl、sc-rnbzlh1、sc-rnbzlh2和bro)计算浓度。在两个实验的第一个中,阿柏西普表达约为15μg/ml,sc-雷珠单抗表达范围为1.6-2.8μg/ml,lh形式表达相似的浓度,hl形式表达低约两倍浓度(图13a)。在第二个实验中,阿柏西普的表达为约50μg/ml,sc-雷珠单抗lh1的表达为约4.0μg/ml(图13b)。溴珠单抗的信号非常低,最可能是由于检测抗体的识别差。在模拟或gfp转染的培养基中没有检测到vegf结合活性。
实施例5
为了使抗vegf蛋白直接可视化并确认蛋白的大小,对来自用gfp、阿柏西普(seqidno:65)、sc-雷珠单抗(seqidno:67、69和70)或溴珠单抗(seqidno:74)表达质粒转染的细胞的培养基进行蛋白质印迹分析(图14)。在bolt4-12%bis-trisplus凝胶(invitrogencat#nw04122box)上运行来自转染的hek293t细胞的培养基,并用iblot2装置将分离的蛋白质转移至硝酸纤维素。使用ibindflex装置,用hrp缀合的羊抗人iggfc(thermocat#31413,左图)或羊抗人iggfab(thermocat#31482,右图)探测印迹,并用supersignalwestdura化学发光底物(thermocat#34076)使其可视化。用ibrightfl1000成像仪捕获图像。
阿柏西普样品似乎与临床比较蛋白eylea相似。在阴性对照样品中没有信号。临床比较蛋白lucentis被还原成24kd的单独轻链和重链,而sc-雷珠单抗蛋白运行在约48kd,这归因于存在将轻链和重链结合成单一蛋白的多肽接头。与hl形式的蛋白质相比,存在更高量的lh形式的蛋白质,这与通过elisa获得的蛋白质定量一致(图13a)。溴珠单抗信号低,最可能是由于检测抗体的识别差。蛋白质以24kd的正确分子量迁移。
实施例6
作为确定vegf结合活性的另一种方法,进行vegf竞争elisa(图15)。临床比较蛋白eylea或lucentis和来自转染的hek293t细胞的培养基与13pmvegf在室温下温育过夜。使用quantikinevegfelisa试剂盒(r&dsystemscat#dve00)测定样品的游离vegf。在两个实验的第一个中,来自转染样品的四种抗vegf蛋白的抑制曲线与临床eylea和lucentis非常相似。阿柏西普和eylea与sc-雷珠单抗变体和lucents相比竞争vegf更强(图15a)。sc-雷珠单抗的所有三种形式(seqidno:67、69和70)几乎相同。在第二个实验中,所有的抗vegf构建体竞争vegf(图15b)。gfp阴性对照样品没有竞争活性。
实施例7
为了确认从转染的细胞表达的抗vegf蛋白阻断vegf与其受体的结合并由此阻断其功能,将相等浓度的临床比较蛋白和来自转染的hek293t细胞的培养基与20ng/mlvegf混合,并置于pathhunterkdr细胞(discoverx93-0996y1)上。这些细胞表达vegf受体/β-半乳糖苷酶融合蛋白,其在vegf结合时产生活性β-半乳糖苷酶。22小时后裂解所述细胞并测定β-半乳糖苷酶活性。来自转染样品的抗vegf蛋白的抑制曲线与临床比较蛋白相同,证明表达的蛋白在基于细胞的测定中阻断vegf功能(图16)。在随后的两个实验中,细胞vegf中和测定也用等体积的培养基进行,所述培养基来自用抗vegf构建体转染的hek293t细胞。在这些实验的第一个中,评价gfp、阿柏西普(seqidno:65)、sc-雷珠单抗hl(seqidno:67)或sc-雷珠单抗lh1(seqidno:69)表达质粒。所有抗vegf构建体评估中和的vegf(图17a)。sc-雷珠单抗lh形式比sc-雷珠单抗hl形式更强地中和vegf。在第二个实验中,将gfp、阿柏西普(seqidno:65)、sc-雷珠单抗lh1(seqidno:69)或溴珠单抗(seqidno:74)表达质粒与8ng/mlvegf混合,并置于pathhunterkdr细胞(discoverx93-0996y1)上。18小时后裂解所述细胞并测定β-半乳糖苷酶活性。再一次,所有抗vegf构建体评估中和的vegf(图17b)。在测定的稀释液中,gfp对照样品有轻微的基质效应。
实施例8
使用fugenehd转染试剂,用含有gfp或抗vegf转基因的质粒模拟转染或转染视网膜色素上皮细胞(rpe)细胞,所述转基因在cag启动子的控制下。在转染后48小时收集培养基。如gfp表达细胞的数目所证明的(数据未显示),rpe细胞的转染效率极低,因此,在来自这些转染的培养基中不能检测到vegf结合活性。评估培养基中vegf的总浓度以确定低浓度的抗vegf剂是否能减少vegf的rpe分泌。转染后48小时收集培养基,用quantikinevegfelisa试剂盒(r&dsystemscat#dve00)进行测定。观察到在阿柏西普(seqidno:65)表达后vegf水平有很大的降低,在sc-雷珠单抗(seqidno:67、69和70)表达后vegf水平有轻微的降低(图18)。所有三种sc-雷珠单抗形式对vegf水平的作用是相似的。一旦通过更有效的aav转导进行递送,抗vegf转基因就导致产生更高浓度的抗vegf蛋白,其通过阿柏西普elisa试剂盒定量。
实施例9
使用携带阿柏西普(seqidno:65)、sc-雷珠单抗(seqidno:67和69)或溴珠单抗(seqidno:74)的基因的aav表达质粒将病毒dna包装在r100变体衣壳(具有seqidno:42所示的氨基酸序列)中。在1.0mlx-vivo10培养基中用这些载体以moi5,000接种12孔板后30天,转导成熟rpe细胞。转导后三天改变培养基,转导后六天收集并更换,转导后十天再次收集并更换。
用quantikinevegfelisa试剂盒(r&dsystemscat#dve00)测定培养基中vegf的总浓度,以确定用抗vegf载体转导对由rpe细胞分泌到培养基中的游离可检测内源vegf水平的影响(图19)。用gfp对照载体转导的细胞所显示的vegf内源水平在第6天样品中为4,500pg/ml,在第10天样品中为8,300pg/ml。用所有抗vegf载体转导在培养基中产生很少至没有可检测的vegf。很可能rpe细胞分泌的vegf的量没有改变,而是培养基中的vegf被抗vegf蛋白结合,使得通过elisa不能检测到。可进行包括蛋白质印迹在内的其它实验以确定结合和未结合vegf的总量。
来自rpe转导的培养基也用阿柏西普elisa试剂盒(eaglebiosciences,immunoguideig-aa115)进行测定。用hrp-抗-iggh+l抗体(thermofishercat#31410)检测抗-vegf蛋白。相对于临床eylea(aflb和gfp)或lucentis(sc-rnbzhl、sc-rnbzlh1和bro)计算浓度。阿柏西普(seqidno:65)的表达在第6天的样品中约为1,800ng/ml,在第10天的样品中为2,800ng/ml。sc-雷珠单抗表达范围为hl(seqidno:67)第6天样品的700ng/ml至lh(seqidno:69)第10天样品的1,700ng/ml(图20)。利用溴珠单抗(seqidno:74)的信号非常低,最可能是由于检测抗体的识别差。在来自gfp转导的培养基中没有检测到vegf结合活性。
实施例10
为了使抗vegf蛋白直接可视化并确认蛋白的大小,对来自转导的rpe细胞的培养基进行蛋白质印迹分析(图21)。在bolt4-12%bis-trisplus凝胶(invitrogencat#nw04122box)上运行等体积的第6天和第10天培养基,并用iblot2装置将分离的蛋白质转移至硝酸纤维素。使用ibindflex装置,用hrp缀合的羊抗人iggfc(thermocat#31413,左图)或羊抗人iggfab(thermocat#31482,右图)探测印迹,并用supersignalwestdura化学发光底物(thermocat#34076)观察。用chemidocmp成像仪捕获图像。
阿柏西普(seqidno:65)样品似乎与临床比较蛋白eylea(黑色箭头)相似。在gfp阴性对照样品中没有正确迁移率的条带。临床lucentis被还原成24kd的单独轻链和重链,而sc-雷珠单抗hl(seqidno:67)和lh(seqidno:69)没有分离,并且以58kd的表观分子量迁移,如灰色箭头所示。溴珠单抗(seqidno:74)的信号低,最可能是由于检测抗体的识别差。蛋白质以26kd的正确分子量迁移,如点箭头所示。在第6天和第10天样品中蛋白质水平相当相似。
实施例11
作为确定vegf结合活性的另一种方法,进行vegf竞争elisa(图22)。来自转导的rpe细胞的等体积培养基与13pmvegf在室温下温育过夜。使用quantikinevegfelisa试剂盒(r&dsystemscat#dve00)测定样品的游离vegf。来自用所有抗vegf构建体转导的细胞的培养基竞争vegf。第6天和第10天样品的结果相似。gfp阴性对照样品没有竞争活性。由于rpe细胞产生的内源性vegf,所以在最低稀释度时游离vegf水平较高。
实施例12
为了比较在来自病毒载体的rpe细胞中表达的抗vegf蛋白的vegf中和活性,将来自转导的rpe细胞的等体积培养基与8ng/mlvegf混合,并置于pathhunterkdr细胞(discoverx93-0996y1)上(图23)。18小时后裂解所述细胞并测定β-半乳糖苷酶活性。来自用所有抗vegf构建体转导的细胞的培养基中和vegf。用来自gfp对照转导的培养基没有观察到vegf中和。
前述内容仅说明了本发明的原理。将理解的是,本领域技术人员将能够设计不同的安排,所述不同的安排虽然没有在此明确地描述或显示,但体现本发明的原理并且包括在其精神和范围之内。另外,在此叙述的所有实例和条件性语言主要旨在帮助读者理解诸位发明人所贡献的本发明的原理和概念以推动本领域发展,并且将被解释为而不限于这些特别叙述的实例和条件。
此外,引用本发明的原理、方面、以及实施方式的所有在此的陈述连同其具体实例旨在涵盖其结构和功能等效物两者。另外,预期此类等效物包括当前已知的等效物以及未来开发的等效物两者,即不论结构而执行相同功能的开发的任何要素。因此,本发明的范围不是旨在受限于在此显示和描述的示例性实施方式。相反,本发明的范围和精神通过所附权利要求书来体现。
- 还没有人留言评论。精彩留言会获得点赞!