非洲猪流感病毒DNA连接酶的晶体及其制备方法和应用与流程
本发明属于生物技术领域,具体涉及来源于非洲猪流感病毒asfivirus属中dna连接酶asfvlig表达纯化、晶体生长方法、晶体结构及其应用。
背景技术:
非洲猪流感病毒(africanswinefevervirus,asfv)是一种双链dna病毒,归属于asfivirus属,并且是目前仅知的asfarviridae家族病毒。asfv病毒的传染性非常强,既能感染野生猪,也能感染家猪。asfv病毒的潜伏期非常短,有些情况下两天左右便会出现病症。非洲猪流感的早期症状包括发热、厌食、四肢泛力等,这些同正常的猪流感的症状很相似,因此早期非洲猪流感常被误诊为正常的猪流感。后期的时候,感染asfv的猪会出现体温下降、皮肤发红等与正常猪流感不同的症状,但此时猪的淋巴结、肾脏、胃肠粘膜等部位已明显出血,并会迅速死亡。
中国、美国以及联合国卫生部们在内的多个国家和组织开展了很多合作研究。但是,目前还没有任何有效的asfv病毒检测手段和疫苗被研发出来,大规模屠宰和焚烧受感染的猪仍是控制asfv病毒漫延的最主要手段。asfv病毒是已报道的最复杂的dna病毒之一,其基因组全长在170-190kb之间,编码了150-170种不同的蛋白。这些蛋白参与了asfv病毒生命循环的多个过程,包括基因表达、dna复制、病毒颗粒组装、宿主细胞侵染以及宿主免疫应答的抑制等。同正常的猪流感病毒相比,asfv病毒基因组的突变频率要高很多。仅通过对其中一个基因b6464l的3’-端序列测定,就有20多个不同的asfv基因型被鉴定出来。这些不同基因型的形成使得asfv疾病的症状、诊断和预防都变得更加困难。
asfv病毒可以在猪的多种不同细胞中复制,其中单核巨噬细胞是asfv病毒最主要的宿主细胞。虽然asfv病毒的dna合成起始于细胞核,但其主要的dna合成以及病毒颗粒组装的过程则是在细胞质中完成的。巨噬细胞的细胞质为富氧环境,其中氧自由基的含量很高。这些高浓度氧自由基的存在对asfv病毒的基因组造成了持续不断的损伤,导致了asfvdna链的断裂以及碱基脱嘌呤环等。为了有效地对抗这类dna损伤,asfv基因组编码了三个dna修复蛋白(包括dna剪切酶asfvap,dna合成酶asfvpolx以及dna连接酶asfvlig),构成了一条完整的baseexcisionrepair(ber)修复通路。其中asfvap负责将脱嘌呤的碱基(df)从dna链上剪切掉,形成缺口;asfvpolx则负责将缺失的dna补充完整;在atp的参与下,asfvlig将dna骨架上的缺口连接起来,完成修复过程。
根据发明人课题组先前的研究结果发现asfvpolx在缺少thumbsubdomain的情况下,进化出了一个与其他dna聚合酶完全不同的底物5’-磷酸根结合位点,为针对asfvpolx的小分子抑制剂的设计提供了结构基础。虽然asfvpolx具有如上的特异性,但是由于asfvpolx与其同家族成员相比,其余结构域的特异性很低。为此发明人需要寻找一个具有更高特异性的,用来作为药物靶点与抗体作用位点的蛋白。
技术实现要素:
本发明的目的是提供一种来源于非洲猪流感病毒asfivirus属中dna连接酶asfvlig的晶体结构、制备方法及其应用。
本发明通过一系列的结晶工艺程序,得到非洲猪流感病毒asfivirus属中dna连接酶asfvlig的晶体,其参数为:
晶体空间群:p21
晶胞参数:a=72.257å,b=55.159å,c=79.875å,α=γ=90°,β=103.15°。
具体而言,非洲猪瘟病毒dna连接酶的晶体三维结构中,一个晶体学不对称单位中有一个蛋白分子,由9个α螺旋以及17个β折叠组成。具体组成如下:α螺旋1,即含有asn3-gln4,β折叠1,即含有gln8-ser10,β折叠2,即含有ile13-cys15,β折叠3,即含有ile19-thr23,β折叠4,即含有gly26-val38,:α螺旋2,即含有asp48-ser52,β折叠5,即含有val59-val68,β折叠6,即含有thr79-val81,α螺旋3,即含有val95-ala111,β折叠6,即含有ile147-lys151,β折叠6,即含有ile155-lys170,α螺旋4,即含有asp181-phe191,β折叠7,即含有tyr199-tyr205,α螺旋5,即含有leu211-asn219,β折叠8,即含有his228-phe235,α螺旋6,即含有ser244-gln257,β折叠9,即含有ile266-gln269,α螺旋7,即含有val275-glu288,β折叠10,即含有glu291-arg296,β折叠11,即含有leu314-leu317,β折叠12,即含有leu321-gln332,β折叠13,即含有ile342-leu348,β折叠14,即含有lys351-lys359,α螺旋8,即含有tyr363-lys372,α螺旋9,即含有pro375-leu382,β折叠15,即含有gly384-ser395,β折叠16,即含有gly399-leu402,β折叠17,即含有arg405-phe409。
本发明提供的非洲猪流感病毒dna连接酶asfvlig的晶体的制备方法,具体步骤如下:
(1)将预先制备好的高纯度asfvlig蛋白浓缩至0.4-0.6mm,并加入等体积的对应的浓度为0.4-0.6mm的dna底物,所述dna底物含有三条dna链,其序列分别为:ccagtccgacccgcatcccgga(seq.id.no.2),tccgggatgcgt(seq.id.no.3),以及pho-gtcggactgg(seq.id.no.4);
(2)使用商业化结晶试剂盒作为晶体生长的初筛条件,采用座滴气象扩散法进行结晶,通过优化调整,最终在0.05-0.5m的醋酸铵(ammoniunacetate),ph为4-6、浓度0.1-0.3m的bis-tris,5-18%的分子量为100-10000的peg分子条件下,得到衍射分辨率高的晶体。
本发明步骤(1)中,所述非洲猪流感病毒dna连接酶asfvlig的蛋白纯度在90%以上,优选为95%以上,更优选98%以上。
本发明步骤(1)中,所述asfvlig蛋白浓缩至0.45-0.55mm,更优选浓缩至0.48mm。
本发明步骤(1)中,所述dna底物的浓度优选为0.5-0.6mm,更优选为0.5mm。
本发明步骤(1)的条件为:将预先制备好的高纯度asfvlig蛋白浓缩至0.45-0.55mm,并加入等体积的对应的浓度为0.5-0.6mm的dna底物。
本发明步骤(2)中,使用的结晶试剂盒为商业化的结晶试剂盒,优选来自hamptonresearch等公司的crystalscreenkiti/ii,index,salt,peg/ion等。
本发明步骤(2)中,结晶溶液优选为:0.1-0.2m的ammoniunacetate,0.1-0.2m的bis-trisph5-5.5,10-18%分子量为1000-10000的peg。更优选结晶溶液为0.1m的ammoniunacetate,0.01m的bis-trisph5.5,10%peg1000条件下,可以得到衍射分辨率高的晶体。
本发明步骤(2)的条件为:作为晶体生长的初筛条件,采用座滴气象扩散法进行结晶,通过优化调整,最终在0.1-0.2mammoniunacetate,0.01mbis-trisph5.5,10%peg1000条件下可以得到衍射分辨率高的晶体。
本发明还公开了非洲猪流感病毒dna连接酶asfvlig的晶体结构在药物设计、制造或药物筛选中的用途。
asfvlig的序列与结构同源性非常低,虽然asfvlig与其余ligase一样具有三个结构域,即负责dna结合的dna结合结构域(dnabindingdomain,dbd,),负责atp活化的addomain(ad结构域),以及负责辅助dna形成从a-form向b-form转变的obdomain(ob结构域)。
根据现有研究结果与已有数据对比发现,asfvlig的n-terminaldomain(ntd)是一个在结构与序列上完全特异性的dna结合结构域。通过与人源dnaligase1-4(hslig1-4)进行序列与结构比对,发现asfvlig的ntd与hslig1-4的dbd在序列与结构上完全不同。由于猪源dnaligase1-4(suslig1-4)与人源dnaligase1-4(hslig1-4)序列同源性极高(同源性高于90%,而且在主要功能区域序列完全一致),发明人认为suslig1-4与hslig1-4存在结构上的一致性。那么asfvligntd与hslig1-4dbd的高度差异性也将存在于asfvligntd与suslig1-4dbd之间。
同时由于asfvligntd会通过与ab与ob互作促使asfvlig形成可以行使功能的活性构象。发明人的研究结果也表明,如果将asfvligntd截除,那么asfvlig的活性将彻底丧失。在不截除asfvligntd的情况下将asfvligntd行使重要结合功能的位点灭活,asfvlig的dna结合能力与活性催化能力都会有极大幅度的下降。
除了ntd结构域外,asfvlig的obdomain也具有很高的特异性。尽管asfvlig的ob结构域与hslig1-4的ob在序列与结构上有一定相似性,但是在反应活性的关键位点上asfvlig与hslig1-4存在显著的区别。
发明人通过解析asfvlig的复合物结构发现asfvlig的活性反应中心与hslig1的活性反应中心存在着明显的不同。从图上可以发现asfvlig在活性反应中心周围更倾向于形成一个较为松散的口袋,而不是像hslig1一样形成一个更为紧密以及刚性的结构。
通过活性验证,发现松散的活性中心构象是asfvlig特异性的。无论是将asfvlig的活性口袋附近的氨基酸残基按照hslig1进行单点突变(l402r、q403f突变体)、l402r\q403f双点突变体(doublemutation)、l402r\q403f\n153d三点突变体(triplemutation)还是l402r\q403f\n153d\l211f四点突变体(quadruplemutation)均会导致asfvlig催化活性的显著降低。
研究结果显示,asfvlig的ntd结构域以及催化位点(n153、l211、l402、q403)附近的口袋都是asfvlig特异性的,均可以作为理想的药物设计靶点,用于设计针对非洲猪瘟病毒的小分子药物与筛选大分子抗体。这些小分子和抗体将有助于针对非洲猪瘟的诊断和防治,具有极高的经济价值。
附图说明
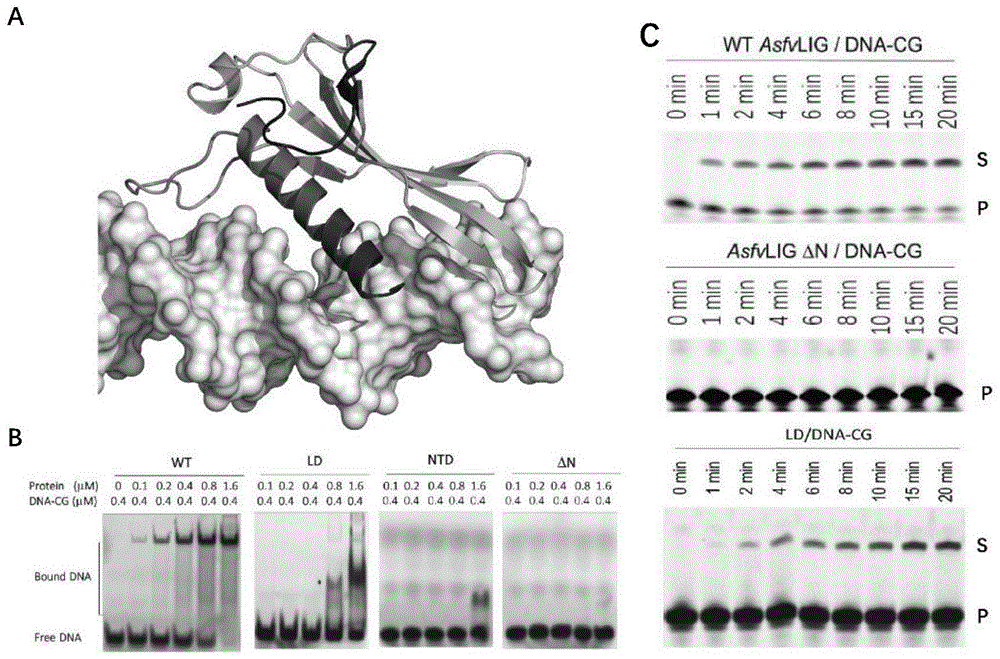
图1:asfvligntd结构及其对asfvlig结合和催化底物dna连接的影响。其中,(a)asfvligntd与dna的结合模式。(b)野生型asfvlig(wt)与asfvligntd(ntd)、asfvligntd截除(δn)及asfvligntd的dna结合loop删除(ld)蛋白的dna结合能力比对。(c)野生型asfvlig(wt)与asfvligntd截除(δn)及asfvligntd的dna结合loop删除(ld)蛋白的催化能力比对。
图2:asfvlig与同源蛋白ob结构域的序列比对。
图3:asfvlig具有独特的活性中心构象。(a)asfvlig的催化位点附近的氨基酸n153、l211、l402、q403构象。(b)hslig1的催化位点附近的氨基酸d570、f635、r871、f872构象。(c)asfvlig与hslig1的催化位点附近的氨基酸构象比较。
具体实施方式
下面通过具体实施例进一步说明本发明的内容。
不能认定本发明的具体实施只局限于这些说明。对于本发明所属技术领域的普通技术人员来说,在不脱离本发明构思的前提下,还可以做出若干简单推演或替换,都应当视为属于本发明的保护范围。
实施例1、来源于非洲猪瘟病毒的dna连接酶的制备
来源于非洲猪瘟病毒的dna连接酶序列如seqidno:1所示,其来自于文献报道(hammondj.et.al.,nucleicacidsresearch,1992,20(11):2667-2671)。其单字母氨基酸序列如下所示:
mlnqfpgqysnnifcfppiesetksgkkaswiicvqvvqhntiipitdemfstdvkdava
eiftkffveegavriskmtrvtegknlgkknattvvhqafkdalskynrharqkrgahtn
rgmippmlvkyfniipktffeeetdpivqrkrngvravacqqgdgcillysrtekeflgl
dnikkelkqlylfidvrvyldgelylhrkplqwiagqanaktdsselhfyvfdcfwsdql
qmpsnkrqqlltnifkqkedltfihqvenfsvknvdealrlkaqfikegyegaivrnang
pyepgynnyhsahlaklkplldaefilvdytqgkkgkdlgailwvcelpnkkrfvvtpkh
ltyadryalfqkltpalfkkhlygkeltveyaelspktgiplqaravgfrepinvleii
使用分子克隆技术将非洲猪瘟病毒dna连接酶经过密码子优化的全长基因导入pet28b-sumo载体上,其可表达一个在氨基末端带有6个连续组氨酸标签的融合蛋白。将该重组质粒通过热激活方法转入到大肠杆菌bl21(de3)中,通过37摄氏度振荡培养复苏该转染菌株10小时,而后将20毫升复苏菌液转入1升lb培养基内,继续37摄氏度振荡扩增至od值0.6~0.8时加入50μl1miptg诱导表达蛋白。表达条件16摄氏度,20-22小时。以上所有流程培养基内皆含有50μg/ml的卡那霉素作为选择压力。
离心收集菌体,以适量buffer(20mmtrisph8.0,500mmnacl,25mm咪唑)重悬菌体。利用低温高压裂解菌体,将裂解产物以17000rpm进行40分钟高速离心以去除沉淀和其它颗粒状杂质。以2ml/min速度使上清液与ni亲和介质结合,先后使用20mmtrisph8.0,1mnacl,25mm咪唑buffer和20mmtrisph8.0,500mmnacl,100mm咪唑的缓冲体系冲洗。最后使用20mmtrisph8.0,500mmnacl,500mm咪唑缓冲体系集中收集目的蛋白。使用ulp1蛋白酶切除his-sumo标签,同时使用20mmtrisph8.0,500mmnacl缓冲体系以透析方式降低蛋白溶液中的咪唑浓度。3小时后将切除标签的蛋白样品再次与ni亲和介质结合,收集流穿样品。将流穿样品通过稀释降至150mmnacl,将稀释后样品以3ml/min流速与阳离子交换柱结合,使用20mmtrisph8.0,200mmnacl,2mmdtt缓冲体系冲洗介质,20mmtrisph8.0,350mmnacl,2mmdtt缓冲体系收集目的蛋白。将收集的目的蛋白浓缩至25mg/ml,液氮速冻后备用。
实施例2、非洲猪瘟病毒dna连接酶活性验证
使用反应体系100mmnacl;20mmtrisph8.0,10mmmgcl2,1mmatp,0.8μmfam荧光标记dna,0.05μm蛋白。在37摄氏度先分别孵育蛋白和底物5min,而后将蛋白加入底物中,按照时间梯度取样,每个样品按照1:1比例加入终止缓冲液(90%甲酰胺,20mmedta,0.05%溴酚蓝,0.05%二甲苯蓝)95摄氏度加热终止反应。将反应终止后的样品使用16%尿素胶跑胶分离,使用typhoonfla9000扫描,而后使用imagequanttl与graphpadprism软件定量分析结果。
实施例3、非洲猪瘟病毒dna连接酶的结晶方法
将预先制备好的高纯度蛋白浓缩至0.48mm,并加入等体积的对应的浓度为0.5mm的长度为22bp的双链dna,使用结晶试剂盒(来自hamptonresearch等公司的crystalscreenkiti/ii,index,salt,peg/ion等)作为晶体生长的初筛条件,采用座滴气象扩散法进行结晶,通过优化调整,最终在0.1mammoniunacetate,0.01mbis-trisph5.5,10%peg1000条件下可以得到衍射分辨率高的晶体。
实施例4、非洲猪瘟病毒dna连接酶的数据收集及结构解析
使用25%甘油作为防冻液冻存晶体于液氮中,在ssrf19u1线站收集数据,使用hkl3000对数据进行处理,该晶体结构通过硒代蛋白提供反常信息使用hkl2map软件进行解析,利用phenix软件进行修正。非洲猪流感病毒(africanswinefevervirus,asfv)asfivirus属中dna连接酶asfvlig的晶体,其参数为:
晶体空间群:p21
晶胞参数:a=72.257å,b=55.159å,c=79.875å,α=γ=90°,β=103.15°。
具体而言,非洲猪瘟病毒dna连接酶的晶体三维结构中,一个晶体学不对称单位中有一个蛋白分子,由9个α螺旋以及17个β折叠组成。具体组成如下:α螺旋1,即含有asn3-gln4,β折叠1,即含有gln8-ser10,β折叠2,即含有ile13-cys15,β折叠3,即含有ile19-thr23,β折叠4,即含有gly26-val38,:α螺旋2,即含有asp48-ser52,β折叠5,即含有val59-val68,β折叠6,即含有thr79-val81,α螺旋3,即含有val95-ala111,β折叠6,即含有ile147-lys151,β折叠6,即含有ile155-lys170,α螺旋4,即含有asp181-phe191,β折叠7,即含有tyr199-tyr205,α螺旋5,即含有leu211-asn219,β折叠8,即含有his228-phe235,α螺旋6,即含有ser244-gln257,β折叠9,即含有ile266-gln269,α螺旋7,即含有val275-glu288,β折叠10,即含有glu291-arg296,β折叠11,即含有leu314-leu317,β折叠12,即含有leu321-gln332,β折叠13,即含有ile342-leu348,β折叠14,即含有lys351-lys359,α螺旋8,即含有tyr363-lys372,α螺旋9,即含有pro375-leu382,β折叠15,即含有gly384-ser395,β折叠16,即含有gly399-leu402,β折叠17,即含有arg405-phe409。
asfvlig蛋白晶体原子坐标群见附表。
实施例5:非洲猪瘟病毒dna连接酶结构和活性分析
晶体结构显示,asfvlig具有一个独特的n-端结构域(图1(a)),该结构域对于底物的结合十分关键(图1(b))。将n-端结构域截除或是将其中结合dna的loop去除均会严重地影响asfvlig的催化活性(图1(c))。序列比对显示asfvlig与同源蛋白的同源性比较低(图2)。在asfvlig底物催化位点附近含有几个独特的氨基酸,包括n153、l211、l402和q403(图3)。对于这些氨基酸残基的突变均会严重地降低asfvlig的催化活性(表1)。蛋白突变、体外dna结合以及活性研究显示,催化位点附近的氨基酸对催化形式asfvdnal-dna复合物的形成及活性至关重要。
表1、野生型和突变体asfvlig催化活性的比较
实施例6非洲猪瘟病毒dna连接酶结晶结构的用途
asfv病毒是一种独特的双链dna病毒,既能感染野生猪,也能感染家猪。一旦爆发,将会导致近100%受感染猪的死亡。自1921年被发现以来,asfv病毒受到了科研人员的广泛关注,但目前仍没有任何有效的疫苗被报道。asfv病毒对全球多个国家和地区的猪饲养业都是一种巨大的威胁,并于2018年在中国首度爆发,并导致了严重的经济损失。asfv病毒编码了多个蛋白,形成了一套完整的dna修复通路。与通常的dna修复通路不同,asfvdna修复通路中的dna聚合酶(asfvpolx)和连接酶asfvdnal的保真性都非常低,在asfv病毒基因组的修复和突变过程中均发挥了关键作用。甘建华课题组和麻锦彪课题组的早期研究发现,asfvpolx拥有一个独特的磷酸根识别位点,对其底物结合和催化活性十分关键(plosbiology,2017)。
本发明的结构显示asfvlig具有一个独特的n-端结构域,该结构域可以作为抗体筛选的靶点。n-端结构域与催化结构域及c-端dna结合结构域之间的协同作用对催化复合物的形成非常关键,这些结构域之间的接触界面可以作为小分子药物的靶点。另外,asfvlig活性位点附近的独特的氨基酸构型(n153、l211、l402和q403)也可以作为小分子药物的靶点。
序列表
<110>复旦大学
<120>非洲猪流感病毒dna连接酶的晶体及其制备方法和应用
<160>4
<170>siposequencelisting1.0
<210>1
<211>419
<212>prt
<213>asfivirus
<400>1
metleuasnglnpheproglyglntyrserasnasnilephecysphe
151015
proproileglusergluthrlysserglylyslysalasertrpile
202530
ilecysvalglnvalvalglnhisasnthrileileproilethrasp
354045
glumetpheserthraspvallysaspalavalalagluilephethr
505560
lysphephevalglugluglyalavalargileserlysmetthrarg
65707580
valthrgluglylysasnleuglylyslysasnalathrthrvalval
859095
hisglnalaphelysaspalaleuserlystyrasnarghisalaarg
100105110
glnlysargglyalahisthrasnargglymetileproprometleu
115120125
vallystyrpheasnileileprolysthrphephegluglugluthr
130135140
aspproilevalglnarglysargasnglyvalargalavalalacys
145150155160
glnglnglyaspglycysileleuleutyrserargthrglulysglu
165170175
pheleuglyleuaspasnilelyslysgluleulysglnleutyrleu
180185190
pheileaspvalargvaltyrleuaspglygluleutyrleuhisarg
195200205
lysproleuglntrpilealaglyglnalaasnalalysthraspser
210215220
sergluleuhisphetyrvalpheaspcysphetrpseraspglnleu
225230235240
glnmetproserasnlysargglnglnleuleuthrasnilephelys
245250255
glnlysgluaspleuthrpheilehisglnvalgluasnpheserval
260265270
lysasnvalaspglualaleuargleulysalaglnpheilelysglu
275280285
glytyrgluglyalailevalargasnalaasnglyprotyrglupro
290295300
glytyrasnasntyrhisseralahisleualalysleulysproleu
305310315320
leuaspalaglupheileleuvalasptyrthrglnglylyslysgly
325330335
lysaspleuglyalaileleutrpvalcysgluleuproasnlyslys
340345350
argphevalvalthrprolyshisleuthrtyralaaspargtyrala
355360365
leupheglnlysleuthrproalaleuphelyslyshisleutyrgly
370375380
lysgluleuthrvalglutyralagluleuserprolysthrglyile
385390395400
proleuglnalaargalavalglyphearggluproileasnvalleu
405410415
gluileile
<210>2
<211>22
<212>dna
<213>人工序列(artificialsequence)
<400>2
ccagtccgacccgcatcccgga22
<210>3
<211>12
<212>dna
<213>人工序列(artificialsequence)
<400>3
tccgggatgcgt12
<210>4
<211>10
<212>dna
<213>人工序列(artificialsequence)
<400>4
gtcggactgg10
- 还没有人留言评论。精彩留言会获得点赞!