水稻丝氨酸羟甲基转移酶编码基因OsSHM4突变体及其应用的制作方法
本发明属于植物基因工程技术领域,涉及水稻丝氨酸羟甲基转移酶基因osshm4突变体及其应用。
背景技术:
硒(se),是人类和动物所必须的营养元素,在保持健康的免疫系统和降低患癌症的风险中起到重要作用(sorsetal.,2005)。在中国的许多地区平均硒摄入量是低于世界卫生组织(who)建议的40μgd-1,甚至在我国一些硒元素贫瘠地区,硒摄入量低于10μgd-1(combs,2001;chenetal.,2002)。为了减少由缺硒引起的癌症、心血管疾病、病毒性疾病等健康问题,提高食物中硒含量是满足人类硒摄入的一种良好途径。在我国水稻是日常重要主食之一,然而大米中硒含量往往较低甚至不足以满足膳食所需(williamsetal.,2009),因此施加硒肥料对农作物而言可作为一种有效的生物强化方法(hawkesford&zhao,2007)。已有研究表明稻田土壤中se含量在0.5-47.7mgkg-1范围内,水稻中se含量与土壤中se含量呈一定线性关系,水稻中se积累量从高到低分别是:秸秆>麸皮>全谷物>精米>稻壳(sunetal.,2010)。当然,进一步探索研究植物从土壤中吸收se,从根部向地上部的转移和地上部se的再分配这一系列过程,将会更加高效和经济地提高农作物se积累量(zhuetal.,2009)。
在碱性和良好氧化的土壤中,硒酸盐是主要形态,而亚硒酸盐普遍存在于酸性到中性的土壤中或还原性土壤中,如水稻土(elramadyetal.,2015)。由于硫酸盐和硒酸盐的化学相似性,硒酸盐可以通过硫酸盐转运蛋白吸收进入根部而亚硒酸盐的吸收机制尚不明确。在土壤中,亚硒酸盐易被铁氧化物和氢氧化物强烈吸附(barrow&whelan,2010),所以硒酸盐是植物硒元素吸收的主要来源。
目前,水稻中直接影响到水稻地上部和籽粒中硒积累的基因报道较少。osshm4基因在genbank中的登录号为ak103554.1,注释为丝氨酸羟甲基转移酶(serinehydroxymethyltransferase4,shm4)。
技术实现要素:
本发明的目的是提供水稻丝氨酸羟甲基转移酶编码基因osshm4的突变体。
本发明的目的是提供水稻丝氨酸羟甲基转移酶编码基因突变体的应用。
水稻丝氨酸羟甲基转移酶编码基因突变体,为野生型水稻osshm4基因cds序列区域第461位的t突变为c,导致其编码的氨基酸由亮氨酸突变为脯氨酸;所述的野生型水稻osshm4基因cds序列如seqidno.1所示;所述的水稻丝氨酸羟甲基转移酶编码基因突变体cds序列如seqidno.3所示。
所述的水稻丝氨酸羟甲基转移酶编码基因突变体编码的蛋白,其氨基酸序列如seqidno.4所示。
进一步地,含有上述水稻osshm4基因突变体的表达载体属于本发明的保护范围。
进一步地,含有上述水稻osshm4基因突变体的宿主细胞属于本发明的保护范围。
本发明所述的水稻丝氨酸羟甲基转移酶编码基因突变体在增强水稻根部对硫酸盐的吸收能力并且显著提高了水稻地上部总硫积累中的应用。
本发明所述的水稻丝氨酸羟甲基转移酶编码基因突变体在增强根部对硒酸盐的吸收能力并且显著提高了水稻地上部总硒积累中的应用。
本发明所述的水稻丝氨酸羟甲基转移酶编码基因突变体在显著提高水稻籽粒中总s和se的积累且不显著影响农艺性状中的应用。
本发明所述的水稻丝氨酸羟甲基转移酶编码基因突变体编码的蛋白在增强水稻根部对硫酸盐的吸收能力并且显著提高了水稻地上部总硫积累中的应用。
本发明所述的水稻丝氨酸羟甲基转移酶编码基因突变体编码的蛋白在增强根部对硒酸盐的吸收能力并且显著提高了水稻地上部总硒积累中的应用。
本发明所述的水稻丝氨酸羟甲基转移酶编码基因突变体编码的蛋白在显著提高水稻籽粒中总s和se的积累且不显著影响农艺性状中的应用。
本发明的有益效果
1,本发明通过系统研究,首次提供了一种水稻丝氨酸羟甲基转移酶编码基因osshm4突变体及其生物学功能。
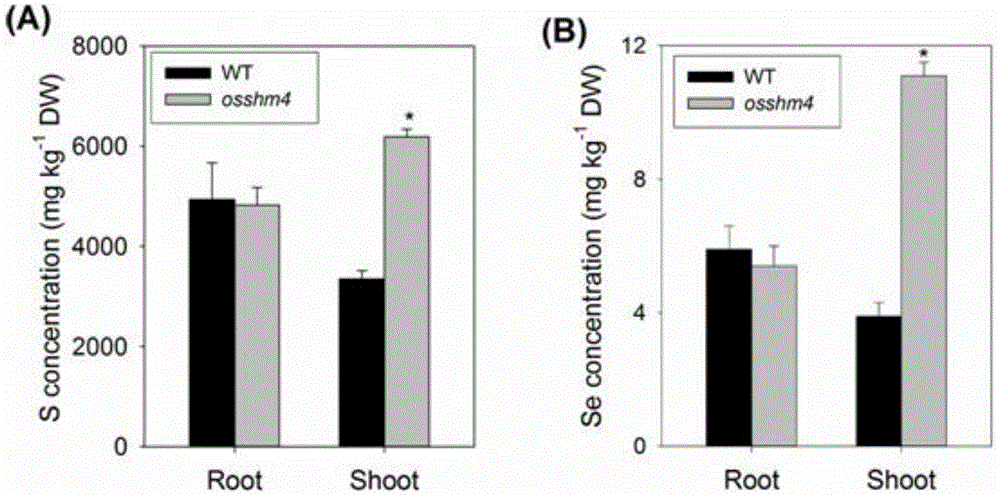
2,该丝氨酸羟甲基转移酶编码基因osshm4突变后,在水培条件下显著提高了水稻地上部硫和硒元素的积累量,而不影响根部硫和硒元素的积累量(图1)。
3,在硒元素吸收动力学实验中,丝氨酸羟甲基转移酶编码基因osshm4突变后根部se吸收能力显著提高(图2);
4,该丝氨酸羟甲基转移酶编码基因osshm4突变后,在田间实验条件下显著提高了水稻地上部硫和硒含量,而不影响水稻的生物量(图3;表1;表2)。
5,野生型丝氨酸羟甲基转移酶编码基因osshm4转基因回补突变体后(图4),回补株系的地上部硫和硒含量恢复至野生型水培(图5)。
附图说明
图1为osshm4基因突变后显著提高了地上部硫元素和硒元素的积累。
a:在1/2kimurab营养液中水培三周,osshm4突变体材料与野生型相比,地上部积累了一倍多的总s而根部s积累量无差异;
b:在1/2kimurab营养液中水培三周后用1μm硒酸钠处理72小时,地上部积累了约三倍的总se而根部总se含量无差异;
图2为osshm4基因突变体材料与野生型相比,根部对硒酸盐的吸收能力显著提高。
图3为田间试验中,osshm4突变体材料与野生型相比,水稻籽粒中积累更多的s和se元素。a:栽种在不同稻田下,野生型和osshm4突变体材料中籽粒s积累量;
b:栽种在不同稻田下,野生型和osshm4突变体材料中籽粒se积累量;
图4为回补载体pzh2b的质粒图谱。
图5为野生型osshm4基因回补突变体后可以将地上部总硫和硒含量恢复至野生型水平。
a:在1/2kimurab营养液中水培三周,野生型基因osshm4回补转基因株系地上部的总硫含量恢复到野生型的水培,回补成功。
b:在1/2kimurab营养液中水培三周,5μm硒酸钠处理72小时,野生型基因osshm4回补转基因株系地上部的总硒含量恢复到野生型的水培,回补成功。
图6载体pos-sgrna和pos-cas9图谱
具体实施方式
以下实施例用于说明本发明,但不用来限制本发明的范围。在不背离本发明精神和本质的情况下,对本发明方法、步骤或条件所作的修改或替换,均属于本发明的范围。
实施例1丝氨酸羟甲基转移酶基因osshm4水稻突变体的获得
1)水稻甲基磺酸乙酯(ems)化学诱变库的构建:约1kg水稻种子(中嘉早17)37℃浸泡24小时后,用1%(v/v)终浓度甲基磺酸乙酯(ems)进行化学诱变8小时,然后用自来水冲洗干净去除残留的ems。在37℃条件下催芽48小时,然后将这些诱变后的水稻种子播种到稻田,水稻成熟后每个单株收获种子m2代并干燥保存备有,完成水稻突变体库的构建。
2)水稻突变体的筛选及基因克隆:在筛选水稻突变体库时,以2μm氯化镉筛选得到一株耐镉水稻突变体并对其进行水稻全基因组测序,定位到突变基因为水稻丝氨酸羟甲基转移酶编码基因osshm4,突变位点为osshm4基因cds序列区域的第一个外显子第461位的t突变为c,导致单个氨基酸替换(亮氨酸—脯氨酸)。
本实施例中得到了一株中嘉早17水稻背景的丝氨酸羟甲基转移酶编码基因osshm4突变体,该突变体中除本发明所述的水稻丝氨酸羟甲基转移酶编码基因osshm4在cds序列区域的第一个外显子第461位存在t-c突变外,基因组其与位置均没有发生突变。
实施例2
osshm4突变体材料与野生型水稻地上部和根部总s和se积累量比较,具体实施过程如下;
1)将水稻野生型与osshm4突变体种子在37℃下浸泡48小时,自来水冲洗干净后播种在悬浮的塑料小黑网上,用0.5mmcacl2溶液培养1周,每三天换次溶液。
2)将水稻野生型与osshm4突变体幼苗转移至4l的塑料黑桶进行培养,用1/2kimurab(ph4.5-5.0)培养两周。4l黒桶分别种4株野生型和4株osshm4突变体,共三桶作为三个生物学重复,然后用终浓度为1mm硒酸钠的营养液处理72小时。
3)收集地上部与根部样品,并用去离子水清洗3次去除表面吸附的离子及杂质,放入65℃烘箱干燥3天后进行消煮。
4)称取全部地上部或根部样品置于石墨消煮管中,加入5ml混酸溶液(85%:15%,v/v,硝酸:高氯酸,优级纯)后放置在石墨消煮炉中进行样品消煮,温控条件为:60℃1小时,90℃1小时,120℃1小时,150℃1小时。然后在190℃条件下完全将管内剩余酸液赶尽,用2%硝酸溶液定容到25ml,混匀后转移至50ml塑料离心管中。
5)用电感耦合等离子体质谱(icp-ms)测定样品溶液中s和se含量,按照样品量和稀释倍数最终计算出水稻野生型和突变体地上部和根部总s和se的含量。
本实施例结果表明osshm4突变体材料与野生型相比,地上部积累更多的总s而非在根部(图1a),并且地上部总se含量也得到了大幅度提高而不影响根部se含量(图1b)。
实施例3
野生型水稻和osshm4突变体材料在硒酸盐吸收能力上的比较,具体实施过程如下:
1)将水稻野生型与osshm4突变体种子在37℃下浸泡48小时,自来水冲洗干净后播种在悬浮的塑料小黑网上,用0.5mmcacl2溶液培养1周,每三天换次溶液。
2)将水稻野生型与osshm4突变体幼苗转移至4l的塑料黑桶进行培养,用1/2kimurab(ph4.5-5.0)培养两周,待用。
3)处理时,将生长三周左右的水稻野生型与osshm4突变体幼苗转移至1l容器中,分别用1l硒酸钠溶液处理20分钟,浓度分别为0,1,2,5,10,20和60μm,水温28℃,室温28℃。每个浓度设置三个生物学重复,一个生物学重复即为1l容器中的三株水稻野生型与三株osshm4突变体材料。
4)处理20分钟后,仅取根部然后用去离子水清洗三次以去吸附的离子及杂质,放在65℃烘箱干燥3天后进行消煮。
5)称取根部样品置于石墨消煮管中,加入5ml混酸溶液(85%:15%,v/v,硝酸:高氯酸,优级纯)后放置在石墨消煮炉中进行样品的的消煮,温控条件为:60℃1小时,90℃1小时,120℃1小时,150℃1小时。然后在190℃条件下完全将管内剩余酸液赶尽,用2%硝酸溶液定容到25ml,混匀后转移至50ml塑料离心管中。
6)用电感耦合等离子体质谱(icp-ms)测定样品溶液中se的含量后,按照样品量和稀释倍数最终计算出水稻野生型和突变体根部吸收的se含量。
本实施例表明osshm4突变体材料与野生型相比,根部吸收硒酸钠[se(v)]的能力大大提高(图2)。线性拟合后,km值(wt,9.4±4.6μmvsosshm4,9.0±0.8μm)没有变化而osshm4突变体材料的vmax值(wt,70.6±12.1mgkg-1h-1vscdt1,164.3±5.6mgkg-1h-1)显著提高,表明了osshm4突变体根部se(v)的吸收能力显著提高。
实施例4
野生型水稻和osshm4突变体材料在大田实验中籽粒总s和se积累量的比较,具体实施过程如下:
1)将水稻野生型与osshm4突变体种子在育秧田播种育秧,长至三四片叶时移苗至水稻田中,每穴一株,每株间距10cm,行间距15cm。期间,正常水分,养分管理控制及病虫害防治。
2)种植在不同地方的水稻田中(南京,湘潭和长兴),并对土壤取样测定ph值及se含量。
3)待水稻成熟后,取四个单株的水稻籽粒混合为一个样品,共设置这样的三个生物学重复,测定水稻野生型与osshm4突变体籽粒中的总s和se的含量。
4)待水稻成熟后,选取每行中间水稻单株共10株以测定水稻野生型与osshm4突变体的农艺性状,取样时从地上部分与土壤接触处剪断。
5)水稻籽粒中总s和se含量测定:称取0.25g干燥籽粒于微波消煮管中,加入5ml浓硝酸(优级纯)进行微波消煮,完成后置于160℃温度下去除残留酸液,赶酸完成后用5%硝酸定容至10ml转移至15ml离心管中保存。利用icp-ms测定籽粒样品中的s含量,用液相色谱-原子荧光光谱仪(lc-afs8500)测定籽粒样品中的se含量。
6)土壤样品ph值测定:称取10g研磨好的干燥稻田土壤,加入25ml去离子水,混合均匀后静置待土壤完全沉降后用ph计测定上层液体的ph值。
7)土壤总se测定:称取0.2g研磨好的干燥稻田土壤于石墨消煮管中,先加入4ml浓盐酸(优级纯)再加入1ml浓硝酸(优级纯),土壤样品消煮温度控制如下:
8)消煮结束后,用2%硝酸定容至25ml,涡旋后静置,再将上清液倒进50ml离心管保存。离心取上清溶液待测,方法同上述水稻籽粒中se的测定。
本实施例结果表明种植在不同地区的稻田时,osshm4突变体材料与野生型相比,其籽粒中富集更多的s(图3a)和se(图3b),并且农艺性状没有太大变化(表1)。总体而言,各地稻田土壤的ph值偏酸且土壤se含量较低都在1mgkg-1以下(表2)。
表1栽种在不同地点的情况下,野生型和osshm4突变体材料的农艺性状指标
表2栽种在不同地点的情况下,稻田土壤样品的ph值和总se含量
实施例5
将野生型丝氨酸羟甲基转移酶编码基因osshm4回补至突变体osshm4中,验证是否能够成功回补地上部高s和se积累的突变表型,具体实施过程如下:
以水稻野生型的基因组dna为模板,回补引物序列如下所示,并通过
回补引物为:
osshm4-com-f:agaggatccccgggtaccttgtttgctaggattgggtt(seqidno.5)
osshm4-com-r:attgttaattaagaattcgatgctagacagaggccgagt(seqidno.6)
将构建好的质粒送武汉伯远生物公司完成突变体水稻的转基因工作,以水稻突变体osshm4为转基因背景。
提取阳性转基因株系单株叶片dna用上述引物扩增片段以进行测序验证,最终挑选出阳性转基因回补株系,连续自交繁种得到t3代纯合材料以用作后续回补表型验证实验。
将水稻野生型,osshm4突变体和三个独立转基因回补株系的种子在37℃下浸泡48小时,自来水冲洗干净后播种在悬浮的塑料网上,用0.5mmcacl2溶液培养1周,每三天换次溶液。然后转移至4l的塑料黑桶用1/2kimurab(ph4.5-5.0)培养两周。
4l黒桶中分别种植水稻野生型,osshm4突变体和三个独立回补株系各四株,共三桶作为三个生物学重复,然后用终浓度为5mm硒酸钠[se(vi)]营养液处理72小时。
收集地上部与根部样品,并用去离子水清洗3次去除表面吸附的离子及杂质,放入65℃烘箱干燥3天后进行消煮。称取全部地上部或根部样品置于石墨消煮管中,加入5ml混酸溶液(85%:15%,v/v,硝酸:高氯酸,优级纯)后放置在石墨消煮炉中进行样品的的消煮,温控条件为:60℃1小时,90℃1小时,120℃1小时,150℃1小时。然后在190℃条件下完全将管内剩余酸液赶尽,用2%硝酸溶液定容到25ml,混匀后转移至50ml塑料离心管中。
用电感耦合等离子体质谱(icp-ms)测定样品溶液中s和se的含量后,按照样品量和稀释倍数最终计算出地上部和根部总s和se的含量。
本实施例结果(图5)显示野生型基因osshm4转基因回补株系地上部的总s和总se含量已恢复至野生型水稻的正常水平,证明丝氨酸羟甲基转移酶编码基因osshm4影响了水稻突变体地上部高s和se积累表型。
实施例6
通过crisper基因敲除技术构建水稻osshm4基因点突变材料并进行验证,具体实施过程如下:
1)根据水稻osshm4基因组序列并利用broadinstitute(http://www.broadinstitute.org/rnai/public/analysis-tools/sgrna-design)设计osshm4基因上的敲除靶点sgrnas,并通过引物合成:
sgrna-f(5’-3’):ggcaaggctcattgctgataagtg(seqidno.7)
sgrna-r(5’-3’):aaaccacttatcagcaatgagcct(seqidno.8)
2)通过t4连接酶将sgrna-f和sgrna-r片段插入到线性化载体pos-sgrna的bsai酶切位点处,再用gateway方法中的lr反应将中间载体与终载pos-cas9相融合,载体图谱如图6所示。测序验证后送至武汉伯远生物科技公司进行水稻转基因,在粳稻中花11背景上通过crispr技术敲除osshm4基因获得点突变材料。
3)用潮霉素标记筛选出阳性转基因苗,以t0代单株叶片dna为模板,正向引物f(5’-3’):gcccaaggaccgtatcat(seqidno.9),反向引物r(5’-3’):gcattgtgatgttgaatggt(seqidno.10),pcr扩增出敲除靶位点sgrna上下游约250bp序列并测序验证检测osshm4基因的敲除结果,crisper敲除所针对敲除位点为osshm4基因第一外显子999-1019bp的敲除靶点序列(5’-aggctcattgctgataagtg-3’(seqidno.11))。
4)严格自交至t2代,以纯合osshm4敲除突变体作表型分析。
5)本实施例结果显示osshm4点突转基因株系地上部的总s和总se含量显著高于野生型水稻,证明丝氨酸羟甲基转移酶编码基因osshm4功能缺失导致了水稻地上部高s和se积累。
参考文献
barrownj,whelanbr.2010.testingamechanisticmodel.viii.theeffectsoftimeandtemperatureofincubationonthesorptionandsubsequentdesorptionofseleniteandselenatebyasoil.europeanjournalofsoilscience40:29-37.
chenl,yangf,xuj,huy,huq,zhangy,pang.2002.determinationofseleniumconcentrationofriceinchinaandeffectoffertilizationofseleniteandselenateonseleniumcontentofrice.journalofagriculturalandfoodchemistry50:5128-5130.
combsgf.2001.seleniuminglobalfoodsystems.britishjournalofnutrition85:517-547.
elramadyh,abdallan,alshaalt,domokosszabolcsye,elhawatn,prokischj,sztrika,farim,elmarsafawys,shamsms.2015.seleniuminsoilsunderclimatechange,implicationforhumanhealth.environmentalchemistryletters13:1-19.
hawkesfordmj,zhaof.2007.strategiesforincreasingtheseleniumcontentofwheat.journalofcerealscience46:282-292.
sorstg,ellisdr,saltde.2005.seleniumuptake,translocation,assimilationandmetabolicfateinplants.photosynthesisresearch86:373-389.
sung,liux,williamspn,zhuy.2010.distributionandtranslocationofseleniumfromsoiltograinanditsspeciationinpaddyrice(oryzasatival.).environmentalscience&technology44:6706-6711.
williamspn,lombie,sung,scheckelkg,zhuy,fengx,zhuj,careya,adomakoee,lawgaliy.2009.seleniumcharacterizationintheglobalricesupplychain.environmentalscience&technology43:6024-6030.
zhuy-g,pilon-smitseah,zhaof-j,williamspn,mehargaa.2009.seleniuminhigherplants:understandingmechanismsforbiofortificationandphytoremediation.trendsinplantscience14:436-442.
序列表
<110>南京农业大学
<120>水稻丝氨酸羟甲基转移酶编码基因osshm4突变体及其应用
<160>11
<170>siposequencelisting1.0
<210>1
<211>1803
<212>dna
<213>水稻(oryzasativa)
<400>1
atggatctctcgcggccggaatcgtcggacctgtcgctgggtttccactcgctcggccac60
gcccgcgggcacgcggtcacggggccgctgcggctgttcgatgacatggaggacgccaag120
ccggagaagagcgtcggtggcggcggaggaggagggggaggaggaggcgaggaggaggat180
ggggaggaagggggcgaccagcacttctccctcctcggccacgcgctctgcgtcaagcgt240
ccacgccgcgcgctctacggaggcggaggcggaggcggggcaggaggtggaggaggagga300
ggaggggaggcgtcctcctgctcgtcttcctcctcgtcgctgcacccggcgaagcggcag360
gcgacggcggagcgcggcgcggatctggaggcccgccgcggcgccgtccgcgcgtggggc420
aaccaggcgctggcggaggccgacccggacgtgcacgcgctgatggagctcgagcgcgac480
cgccaggtgcgcgggatcgagctgatcgcgtcggagaacttcgtctgccgcgcggtgctc540
gaggcgctcggcagccacctcaccaacaagtactctgagggccatcccggggctcggtac600
tatggcgggaaccagcacatcgacggcatcgagcgcctctgccatgagcgcgccctcgcc660
gccttcggtctcgaccccgcctgctggggggtcaacgtccagccatactcctgcacctct720
gcaaacctagctgtttacacaggtctcctcctgcccaaggaccgtatcatggggctcgag780
ccgccttccggtggccatgtcagccacgggtactacacaccaagtggtaagaaggtgtct840
ggagcatccatattcttcgagagcttgtcgtacaaggtgaatccgcagactgggtatatt900
gattacgacaagcttgaggagcgggcaatggatttccaccctaagattctcatatgtggt960
gggagctcataccccagggagtgggattttgcacggatgaggctcattgctgataagtgc1020
ggtgcggtgctcatgtgcgatatggcacatatcagcgggcttgttgcggccaaggaatgc1080
cgtagtccctttgattactgtgatgtggttacatcaactactcacaagaatctgagaggt1140
cctagaggtggtataatatttttcaggagagggaaaaacttaaggaggcgtactggatct1200
ttttctcaagcggatgagaatgattatgacttcgaggacaggataaattttgcagttttc1260
ccttcaatgcaaggaggaccccataataatcatattgcagccttggcgataactctgaag1320
caagtagcaacacctgaatacaaagcatacatcatacaagttaagaaaaatgcccaagct1380
ttggcatcagccttactcagaagaaaatgcagattggttactggtggcaccgataaccac1440
ttggtactttgggacctcagaaatttaggcttgactggaaagaactttgagaaggtctgt1500
gaggcatgccacatctccataaataagatgccgatatatggggataatggctcaatatct1560
cctgggggtgttcggattggaacacctgcaatgactactagaggttgcttggaagatgac1620
tttgaggtgatcgcagatttcctcatcagagccacccaaatagctagtaatcttatgaaa1680
gaacatggaaaaatgcagaaggaattcctgagaggcctacagaacaataaagatatcata1740
gaactccgtaaccaggttgagaattttgcatcgcagtttgcaatgccaggttttgatgta1800
tga1803
<210>2
<211>600
<212>prt
<213>水稻(oryzasativa)
<400>2
metaspleuserargprogluserseraspleuserleuglyphehis
151015
serleuglyhisalaargglyhisalavalthrglyproleuargleu
202530
pheaspaspmetgluaspalalysproglulysservalglyglygly
354045
glyglyglyglyglyglyglyglygluglugluaspglygluglugly
505560
glyaspglnhispheserleuleuglyhisalaleucysvallysarg
65707580
proargargalaleutyrglyglyglyglyglyglyglyalaglygly
859095
glyglyglyglyglyglyglualasersercysserserserserser
100105110
serleuhisproalalysargglnalathralagluargglyalaasp
115120125
leuglualaargargglyalavalargalatrpglyasnglnalaleu
130135140
alaglualaaspproaspvalhisalaleumetgluleugluargasp
145150155160
argglnvalargglyilegluleuilealasergluasnphevalcys
165170175
argalavalleuglualaleuglyserhisleuthrasnlystyrser
180185190
gluglyhisproglyalaargtyrtyrglyglyasnglnhisileasp
195200205
glyilegluargleucyshisgluargalaleualaalapheglyleu
210215220
aspproalacystrpglyvalasnvalglnprotyrsercysthrser
225230235240
alaasnleualavaltyrthrglyleuleuleuprolysaspargile
245250255
metglyleugluproproserglyglyhisvalserhisglytyrtyr
260265270
thrproserglylyslysvalserglyalaserilephephegluser
275280285
leusertyrlysvalasnproglnthrglytyrileasptyrasplys
290295300
leuglugluargalametaspphehisprolysileleuilecysgly
305310315320
glysersertyrproargglutrpaspphealaargmetargleuile
325330335
alaasplyscysglyalavalleumetcysaspmetalahisileser
340345350
glyleuvalalaalalysglucysargserpropheasptyrcysasp
355360365
valvalthrserthrthrhislysasnleuargglyproargglygly
370375380
ileilephepheargargglylysasnleuargargargthrglyser
385390395400
pheserglnalaaspgluasnasptyraspphegluaspargileasn
405410415
phealavalpheprosermetglnglyglyprohisasnasnhisile
420425430
alaalaleualailethrleulysglnvalalathrproglutyrlys
435440445
alatyrileileglnvallyslysasnalaglnalaleualaserala
450455460
leuleuargarglyscysargleuvalthrglyglythraspasnhis
465470475480
leuvalleutrpaspleuargasnleuglyleuthrglylysasnphe
485490495
glulysvalcysglualacyshisileserileasnlysmetproile
500505510
tyrglyaspasnglyserileserproglyglyvalargileglythr
515520525
proalametthrthrargglycysleugluaspaspphegluvalile
530535540
alaasppheleuileargalathrglnilealaserasnleumetlys
545550555560
gluhisglylysmetglnlysglupheleuargglyleuglnasnasn
565570575
lysaspileilegluleuargasnglnvalgluasnphealasergln
580585590
phealametproglypheaspval
595600
<210>3
<211>1803
<212>dna
<213>水稻(oryzasativa)
<400>3
atggatctctcgcggccggaatcgtcggacctgtcgctgggtttccactcgctcggccac60
gcccgcgggcacgcggtcacggggccgctgcggctgttcgatgacatggaggacgccaag120
ccggagaagagcgtcggtggcggcggaggaggagggggaggaggaggcgaggaggaggat180
ggggaggaagggggcgaccagcacttctccctcctcggccacgcgctctgcgtcaagcgt240
ccacgccgcgcgctctacggaggcggaggcggaggcggggcaggaggtggaggaggagga300
ggaggggaggcgtcctcctgctcgtcttcctcctcgtcgctgcacccggcgaagcggcag360
gcgacggcggagcgcggcgcggatctggaggcccgccgcggcgccgtccgcgcgtggggc420
aaccaggcgctggcggaggccgacccggacgtgcacgcgccgatggagctcgagcgcgac480
cgccaggtgcgcgggatcgagctgatcgcgtcggagaacttcgtctgccgcgcggtgctc540
gaggcgctcggcagccacctcaccaacaagtactctgagggccatcccggggctcggtac600
tatggcgggaaccagcacatcgacggcatcgagcgcctctgccatgagcgcgccctcgcc660
gccttcggtctcgaccccgcctgctggggggtcaacgtccagccatactcctgcacctct720
gcaaacctagctgtttacacaggtctcctcctgcccaaggaccgtatcatggggctcgag780
ccgccttccggtggccatgtcagccacgggtactacacaccaagtggtaagaaggtgtct840
ggagcatccatattcttcgagagcttgtcgtacaaggtgaatccgcagactgggtatatt900
gattacgacaagcttgaggagcgggcaatggatttccaccctaagattctcatatgtggt960
gggagctcataccccagggagtgggattttgcacggatgaggctcattgctgataagtgc1020
ggtgcggtgctcatgtgcgatatggcacatatcagcgggcttgttgcggccaaggaatgc1080
cgtagtccctttgattactgtgatgtggttacatcaactactcacaagaatctgagaggt1140
cctagaggtggtataatatttttcaggagagggaaaaacttaaggaggcgtactggatct1200
ttttctcaagcggatgagaatgattatgacttcgaggacaggataaattttgcagttttc1260
ccttcaatgcaaggaggaccccataataatcatattgcagccttggcgataactctgaag1320
caagtagcaacacctgaatacaaagcatacatcatacaagttaagaaaaatgcccaagct1380
ttggcatcagccttactcagaagaaaatgcagattggttactggtggcaccgataaccac1440
ttggtactttgggacctcagaaatttaggcttgactggaaagaactttgagaaggtctgt1500
gaggcatgccacatctccataaataagatgccgatatatggggataatggctcaatatct1560
cctgggggtgttcggattggaacacctgcaatgactactagaggttgcttggaagatgac1620
tttgaggtgatcgcagatttcctcatcagagccacccaaatagctagtaatcttatgaaa1680
gaacatggaaaaatgcagaaggaattcctgagaggcctacagaacaataaagatatcata1740
gaactccgtaaccaggttgagaattttgcatcgcagtttgcaatgccaggttttgatgta1800
tga1803
<210>4
<211>600
<212>prt
<213>水稻(oryzasativa)
<400>4
metaspleuserargprogluserseraspleuserleuglyphehis
151015
serleuglyhisalaargglyhisalavalthrglyproleuargleu
202530
pheaspaspmetgluaspalalysproglulysservalglyglygly
354045
glyglyglyglyglyglyglyglygluglugluaspglygluglugly
505560
glyaspglnhispheserleuleuglyhisalaleucysvallysarg
65707580
proargargalaleutyrglyglyglyglyglyglyglyalaglygly
859095
glyglyglyglyglyglyglualasersercysserserserserser
100105110
serleuhisproalalysargglnalathralagluargglyalaasp
115120125
leuglualaargargglyalavalargalatrpglyasnglnalaleu
130135140
alaglualaaspproaspvalhisalaprometgluleugluargasp
145150155160
argglnvalargglyilegluleuilealasergluasnphevalcys
165170175
argalavalleuglualaleuglyserhisleuthrasnlystyrser
180185190
gluglyhisproglyalaargtyrtyrglyglyasnglnhisileasp
195200205
glyilegluargleucyshisgluargalaleualaalapheglyleu
210215220
aspproalacystrpglyvalasnvalglnprotyrsercysthrser
225230235240
alaasnleualavaltyrthrglyleuleuleuprolysaspargile
245250255
metglyleugluproproserglyglyhisvalserhisglytyrtyr
260265270
thrproserglylyslysvalserglyalaserilephephegluser
275280285
leusertyrlysvalasnproglnthrglytyrileasptyrasplys
290295300
leuglugluargalametaspphehisprolysileleuilecysgly
305310315320
glysersertyrproargglutrpaspphealaargmetargleuile
325330335
alaasplyscysglyalavalleumetcysaspmetalahisileser
340345350
glyleuvalalaalalysglucysargserpropheasptyrcysasp
355360365
valvalthrserthrthrhislysasnleuargglyproargglygly
370375380
ileilephepheargargglylysasnleuargargargthrglyser
385390395400
pheserglnalaaspgluasnasptyraspphegluaspargileasn
405410415
phealavalpheprosermetglnglyglyprohisasnasnhisile
420425430
alaalaleualailethrleulysglnvalalathrproglutyrlys
435440445
alatyrileileglnvallyslysasnalaglnalaleualaserala
450455460
leuleuargarglyscysargleuvalthrglyglythraspasnhis
465470475480
leuvalleutrpaspleuargasnleuglyleuthrglylysasnphe
485490495
glulysvalcysglualacyshisileserileasnlysmetproile
500505510
tyrglyaspasnglyserileserproglyglyvalargileglythr
515520525
proalametthrthrargglycysleugluaspaspphegluvalile
530535540
alaasppheleuileargalathrglnilealaserasnleumetlys
545550555560
gluhisglylysmetglnlysglupheleuargglyleuglnasnasn
565570575
lysaspileilegluleuargasnglnvalgluasnphealasergln
580585590
phealametproglypheaspval
595600
<210>5
<211>38
<212>dna
<213>人工序列(artificialsequence)
<400>5
agaggatccccgggtaccttgtttgctaggattgggtt38
<210>6
<211>39
<212>dna
<213>人工序列(artificialsequence)
<400>6
attgttaattaagaattcgatgctagacagaggccgagt39
<210>7
<211>24
<212>dna
<213>人工序列(artificialsequence)
<400>7
ggcaaggctcattgctgataagtg24
<210>8
<211>24
<212>dna
<213>人工序列(artificialsequence)
<400>8
aaaccacttatcagcaatgagcct24
<210>9
<211>18
<212>dna
<213>人工序列(artificialsequence)
<400>9
gcccaaggaccgtatcat18
<210>10
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>10
gcattgtgatgttgaatggt20
<210>11
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>11
aggctcattgctgataagtg20
- 还没有人留言评论。精彩留言会获得点赞!