选择性地操纵靶细胞中对两个或更多个共有受体多肽识别的配体的细胞反应的方法与流程
本申请是申请日为2013年8月8日,国际申请号为pct/us2013/054164,国家申请号为201380047839.4,发明名称为“超级因子和合成因子:具有新的和增强的信号传导活性的经改造细胞因子”的发明专利申请的分案申请。
相关申请的交叉引用
本申请要求在2013年5月21日提交的美国临时专利申请第61/825,980号、在2012年11月13日提交的美国临时专利申请第61/725,791号,以及在2012年8月9日提交的美国临时专利申请第61/681,490号的优先权,上述每一申请的公开内容出于所有目的以引用方式整体并入本文。
本发明涉及免疫学领域,具体涉及一种超级因子和合成因子,即具有新的和增强的信号传导活性的经改造的细胞因子。
背景技术:
细胞因子是由许多细胞分泌的小的细胞信号传导分子并且是广泛用于细胞间通信的一类信号传导分子。细胞因子调节关键的细胞功能,包括分化、增殖和凋亡/抗凋亡。
许多细胞因子通过以下方式介导刺激作用:首先与相对高亲和力的细胞因子受体链(通常标记为“α”)相互作用,之后与不同细胞因子共有共有的受体链(共有受体链)相对低亲和力的相互作用。细胞因子与第一高亲和力受体的结合产生复合表面,所述共有受体链可随后与该复合表面结合。
白细胞介素-4(il-4)为这种细胞因子之典型。il-4的主结合链是il-4受体α(il-4rα)。il-4/il-4rα复合物充当il-4受体的第二组分γc的配体。另外,il-4/il-4rα复合物充当白细胞介素-13(il-13)受体α1(il-13rα1)的配体。与il-4不同,il-13不结合至il-4rα,但是il-13/il-13rα1复合物结合至il-4rα。
因为il-4和il-13可以通过不同的受体进行信号传导,所以可以假设il-4和il-13能够激活不同的信号转导通道。实际上,γc激活酪氨酸激酶janus激酶3(jak3),而il-13rα1激活tyk2和jak2。被激活的jak介导il-4r对保守的酪氨酸残基的胞质尾区磷酸化,所述保守的酪氨酸残基充当包含src同源2(sh2)域的蛋白的停靠位点。三个紧密成簇的酪氨酸残基充当信号转导和转录激活蛋白6(stat6)的停靠位点,所述stat6是选择性地偶联至il-4rα链的转录因子。il-13与il-13rα1的结合还通过il-13/il-13rα1复合物与il-4rα的结合而激活stat6。
除了stat6,il-4募集和激活irs-2。结构-功能分析已经显示il-4rα的跨膜结构域上的酪氨酸残基[tyr497,胰岛素/il-4r基序(i4r)的部分]是在il-4rα已经被il-4激活之后irs-2停靠至il-4rα所必需的。jak1和jak3随后磷酸化结合il-4rα的irs-2。irs-2的激活导致磷脂酰肌醇3-激酶(pi3k)和下游蛋白丝氨酸/苏氨酸激酶akt的激活,这种途径被认为介导许多细胞类型中的生长信号和存活信号。实际上,这种途径对表达i型il-4r的细胞(nk细胞、t细胞,以及b细胞)中的il-4-介导的生长是重要的。
虽然il-4rα是普遍存在的,但是γc而非il-13rα1存在于t细胞、自然杀伤(nk)细胞、嗜碱性粒细胞、肥大细胞以及大部分小鼠b细胞上(大部分人b细胞表达γc和il-13rα1两者)。因此,il-4而不是il-13,促进天然t细胞分化成th2细胞,并且il-4显得比il-13对小鼠ige反应的诱导更加重要。
一些骨髓源细胞,包括巨噬细胞和树突状细胞,表达γc和il-13rα1两者并且因此对il-4和il-13两者应答。这两种受体亚基在这些细胞的不同亚群上的相对丰度差异可以部分地负责其对il-4相对于对il-13的相对反应性。在大部分非骨髓源细胞(包括平滑肌细胞和上皮细胞)上发现的是il-13rα1,但很少或没有γc亚基;因此,与il-13相比,il-4在刺激这些细胞方面并没有固有的优势。
在20世纪90年代早期,进行了使用il-4治疗癌症的临床试验。已经观察到il-4诱导体外白血病淋巴母细胞的生长抑制和凋亡。这些观测已在使用移植到免疫缺陷型小鼠中的人白血病细胞的实验中得到证实。不幸地是,il-4的临床有效性受到细胞因子的多效活性的限制,所述多效活性包括肾毒性、肝毒性、神经毒性和胃肠道毒性以及血管渗漏综合征,这与il-4与非造血细胞的结合相关。因此,“野生型”il-4作为治疗剂的用途受到其结合造成不良反应的细胞类型的能力的限制。
因此,本领域中存在对具有对一种受体相对于另一种受体的增加的选择性的分子的需求;使用il-4作为实例,其对γc相对于il-13rα1增加的选择性可以是有利的,或者反之亦然。
此外,与野生型细胞因子的使用相关的一些毒性可以是高剂量施用的结果。因此,可用较低剂量实现所需共有受体的激活的分子也是有利的。
因此,本发明解决了本领域中的这些和其他需要。
技术实现要素:
许多细胞因子在制药领域正在被开发用于潜在的临床应用。在这些情况的大多数中,正在尝试通常伴随剂量限制性毒性或不足疗效的内源性野生型细胞因子。
一种改良细胞因子的可能性是使其偏向对某些希望细胞类型的优选活性。通过异二聚体受体复合物起作用的细胞因子尤其适合这种方法。蛋白工程可用于提供新的细胞因子突变蛋白,所述突变蛋白改变了对共有受体的相对亲和力。
因此,在一些实施方案中提供了一种用于选择性地操纵靶细胞中对两个或更多个共有受体多肽识别的配体的细胞反应的方法。所述方法包括提供突变蛋白配体,其中所述突变蛋白配体以比所述配体更高的亲和力结合所述两个或更多个共有受体多肽中的一个。在其他实施方案中,所述突变蛋白配体以比所述配体更高的亲和力结合所述两个或更多个共有受体多肽中的两个,从而选择性地操纵所述细胞反应。而在其他实施方案中,所述突变蛋白配体以比所述配体更高的亲和力结合所述两个或更多个共有受体多肽中的一个,并且以比所述配体更低的亲和力结合所述两个或更多个共有受体多肽中的另一个,从而选择性地操纵所述细胞反应。在其他实施方案中,所述两个或更多个共有受体多肽不均等地存在于所述靶细胞表面。在其他实施方案中,被所述突变蛋白配体结合的共有受体以比未被所述突变蛋白配体结合的共有受体更低的水平存在于所述靶细胞表面上。在其他实施方案中,被所述突变蛋白配体结合的共有受体以比未被所述突变蛋白配体结合的共有受体更高的水平存在于所述靶细胞表面上。在本申请所述的方法中,其中未被所述突变蛋白配体结合的共有受体具有减少的对所述突变蛋白配体的可及性。在其他实施方案中,通过提供干扰未被所述突变蛋白配体结合的共有受体可及性的试剂来减少对未被所述突变蛋白配体结合的共有受体的可及性。在其他实施方案中,所述试剂是识别未被所述突变蛋白配体结合的共有受体的抗体。在其他实施方案中,所述突变蛋白是il-4突变蛋白。在其他实施方案中,所述共有受体是常见的γ链(γc)或者白细胞介素-13受体α1(il-13rα1)。在其他实施方案中,所述更高的亲和力是至少10倍。在其他实施方案中,所述更低的亲和力是至少5倍。在其他实施方案中,所述配体是生长因子或者细胞因子。
在一些实施方案中,提供了包含细胞因子突变蛋白的组合物,其中相对于野生型细胞因子细胞因子突变蛋白导致与共有细胞因子受体更高亲和力的结合。在一些实施方案中,相对于野生型细胞因子所述细胞因子突变蛋白导致与第一共有细胞因子受体结合亲和力的相对升高,并且与第二共有细胞因子受体降低的结合亲和力。在其他实施方案中,相对于野生型细胞因子细胞因子突变蛋白导致与两个共有细胞因子受体结合亲和力的相对升高。在一些实施方案中,第一共有细胞因子受体以比第二共有细胞因子受体更低的水平表达,并且在一些实施方案中,第一共有细胞因子受体以比第二共有细胞因子受体更高的水平表达。在一些实施方案中,相对于野生型细胞因子细胞因子突变蛋白导致与第一和第二共有细胞因子受体更高亲和力的结合。在一些实施方案中,所述亲和力升高是至少10倍。在一些实施方案中,所述亲和力降低是至少5倍。
在一些实施方案中,共有细胞因子受体是常见的γ链(γc)。在其他实施方案中,共有细胞因子受体是il-13受体α1。在一些实施方案中,第一共有细胞因子受体是γc,并且第二共有细胞因子受体是il-13rα1。在一些实施方案中,第一共有细胞因子受体是il-13rα1,并且第二共有细胞因子受体是γc。在一些实施方案中,第一和第二共有细胞因子受体是γc和il-13rα1。
在一些实施方案中,所述细胞因子突变蛋白是il-4突变蛋白。
在一些实施方案中,提供具有改变的信号传导性质的il-4突变蛋白,其中与il-4突变蛋白接触的ramos细胞中的stat磷酸化相对于与相同浓度的il-4接触的ramos细胞更大,因此证明il-4突变蛋白具有改变的信号传导性质。在一些实施方案中,提供具有改变的信号传导性质的il-4突变蛋白,其中与il-4突变蛋白接触的a549细胞中的stat磷酸化相对于与相同浓度的il-4接触的a549细胞更大,因此证明il-4突变蛋白具有改变的信号传导性质。
在一些实施方案中,提供包含在位置117、118、121、122、124、125、128和129处的两个、三个、四个、五个、六个、七个或者八个氨基酸取代的il-4突变蛋白,其中氨基酸编号根据野生型人il-4(seqidno:1)。在一些实施方案中,取代在il-4的位置121处进行。在一些实施方案中,取代在il-4的位置121和124处进行。在其他实施方案中,取代在位置121、124和125处进行。
在一些实施方案中,取代在il-4的位置117、118、121、122、124、125、128和129处进行。在其他实施方案中,所公开的il-4突变蛋白具有序列117r、118v、121q、122s、124w、125f、128g和129a。
在一些实施方案中,提供包含细胞因子突变蛋白的药物组合物,其中突变蛋白的施用导致相对于野生型突变蛋白减少的副作用。在一些实施方案中,副作用是血管渗漏综合征。在其他实施方案中,提供包含细胞因子突变蛋白的药物组合物,其中用相对于野生型突变蛋白较少的细胞因子突变蛋白实现相同的治疗效力。
在一些实施方案中,提供包含所描述的细胞因子突变蛋白中的任何一个或多个的药物组合物。在其他实施方案中,提供一种治疗受试者的方法,所述方法包括所描述的细胞因子突变蛋白中任何一个或多个的组合物。在一些实施方案中,所述受试者患有炎性疾病。
现在,将在以下详述中更具体地描述进一步的特征和优点。
附图简述
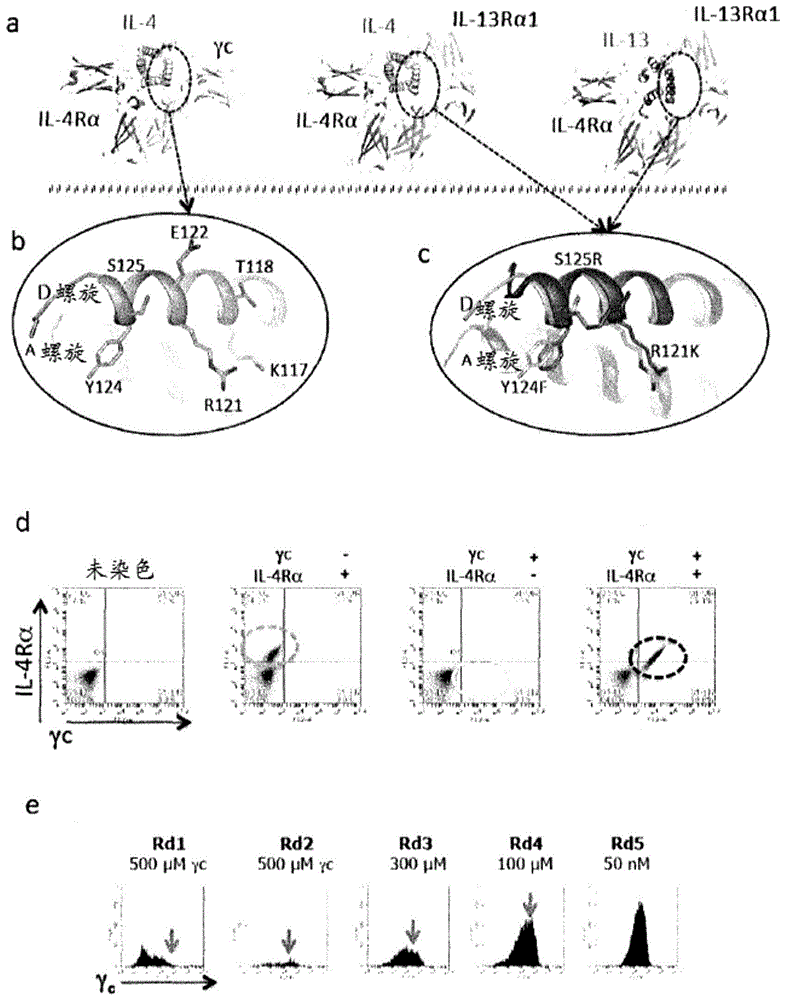
图1描绘il-4超级因子(superkine)的基于结构的工程改造。(a)il-4和il-13i型和ii型三元胞外域复合物6的晶体结构。(b)和(c)分别在il-4和il-13的d螺旋上的主要的γc和il-13rα1结合位点。(b)il-4界面库残基,在(b)中显示了在il-4位点2库中随机化的位置,以及(c)il-4与il-13重叠,在(c)中在所述受体复合物中il-4和il-13的结构重合显示il-4的位置121、124和125紧密重叠在il-13的类似位置上,导致我们要用il-13的那些位置来取代这些位置。在(c)中il-13显示为紫色,并且il-4显示为浅绿色,经取代的残基显示为红色。(d)用受体ecd对il-4-酵母facs染色。在展示il-4的酵母上il-4/il-4rα/γc三元胞外域复合物的协作组装。(e)γc胞外域四聚体上il-4位点2库的顺序富集。在图(b)中显示的il-4位点2库通过分选γc四聚体而逐步富集。在第一分选步骤中用il-4rα复合所述库,后续的循环不包括il-4rα。最终的第六分选循环是对1μm的γc单体的分选(未显示)。
图2描绘了il-4和合成因子与通过c末端生物素化固定在表面上的γc结合的表面等离子体共振分析。
图3描绘il-4对γc的亲和力增强的结构基础。(a)super-4与γc复合。super-4/γc二元复合物在
图4描绘super-4螺旋d的电子密度。
图5描绘super-4/γc界面中重塑的氢键键合相互作用。
图6描绘细胞系上il-4受体组分的表达。(a)il-4rα、γc和il-13rα1的mrna的表达是通过对已经饥饿过夜的细胞以材料和方法中描述的定量pcr来测量的。结果显示三次独立实验的平均值和sem。(b)通过流细胞计数法测定指定细胞系的细胞表面上il-4rα、γc和il-13rα1的表达。用特异性抗体测量的mfi标准化为相同同种型的对照抗体。
图7描绘il-4超级因子对细胞内信号传导的影响。(a)经过夜饥饿的ramos细胞未经刺激,或者用100pg/ml的il-4、super-4或者kfr刺激所示时间。细胞随后被固定、透化,以及用抗磷酸化stat6的抗体染色。进行三次独立实验,具有所示的平均值和sem。(b)用递增量的il-4或super-4刺激ramos细胞15分钟,随后进行如图7(a)中所示的分析。重复该实验三次;指示平均值和sem。(c)ramos细胞经饥饿过夜,接着用递增量的il-4或所指示的超级因子刺激15分钟。指示了三次独立实验的平均值和±sem。(d)a549细胞经饥饿过夜,用指示浓度的il-4或超级因子刺激15分钟。细胞如7(a)中所示地固定和准备;进行三次独立实验,并指示平均值和sem。(e)u937细胞经饥饿过夜,接着用il-4或者超级因子刺激15分钟;pstat6如在7(a)所示测量。重复实验三次;指示了平均值和sem。(f)如所示用il-4或者super-4刺激ramos细胞8小时,接着用cd23进行表面染色。重复实验三次;指示了cd23上调的平均值和sem。
图8描绘单次实验中响应于il-4和超级因子的stat6磷酸化染色。
图9描绘响应于第二链募集的增强和第二链的数量变化的受体组装的模型化。使用matlab脚本来计算仅表达i型il-4受体的细胞表面上il-4受体的组装。(a)在上方的三个图中,il-4rα数目设置为1500。第二链的数目从500升至4500,并且il-4rα复合物对于第二链的二维平衡常数在从0.01μm2到1μm2的范围内,如所指示的。计算针对100pg/ml和1000pg/ml配体,γc分子为每细胞500个、1500个和4500个时最高第二链k值(1.0μm2)对最低第二链k值(0.01μm2)的组装链比率。(b)在中间三个图中,il-4rα的数量设置为1500。二维平衡常数从1μm2变化至0.01μm2,以及第二链数量从每细胞167个到每细胞4500个。显示用每细胞167条γc链组装的复合物,il-4或者突变蛋白浓度为100pg/ml和1000pg/ml,二维平衡常数是1.0μm2、0.1μm2或者0.01μm2。在底部的图中,绘制组装复合物的数量与il-4或者super-4浓度的关系,并且绘制在il-4浓度范围为10,000pg/ml到30,000pg/ml时逼近组装受体的最大数量的线条。
图10描绘改变γc可用性对il-4诱导的stat6激活或者超级因子诱导的stat6激活的影响。(a)用指示浓度的il-4或突变蛋白刺激hh细胞,进行如图7所示的细胞内染色和分析。显示了具有相同结果的两次独立实验的代表性实验。(b)经饥饿的ramos细胞保持未处理,或者用5μg/ml或者50μg/ml的抗γc封闭抗体孵育1小时。随后用100pg/ml的il-4或者所示超级因子刺激所述细胞。随后如图7a所示地进行分析。进行三次独立实验,指示了平均值和sem。(c)经饥饿的u937细胞如10b中的ramos细胞一样进行处理和分析。
图11描绘超级因子对人原代细胞的作用。(a)用指示浓度的il-4、super-4或kfr刺激pbl15分钟,或者pbl保持为未经刺激。如图7所示地计算pstat6的诱导;指示了来自5个供体的数据的平均值和sem(l=淋巴细胞,m=单核细胞,ne=中性粒细胞)。(b)如图6b所示地测量来源于单一供体的人pbl上il-4受体链的表达。对于il-4rα、γc和il-13rα1表达的测量,b细胞和t细胞通过细胞表面标志物(cd19、cd3)门控,而单核细胞和中性粒细胞则基于其区别性的前向和侧向散射而被识别为cd19和cd3阴性细胞。合适的同种型对照充当阴性对照。(c)如在图11b中所示地测量来源于六个供体的pbl上il-4r受体链的表达;提供了平均值和sem(t=t细胞,b=b细胞,m=单核细胞,ne=中性粒细胞)。(d)人pbl上il-4i型和ii型受体链的表达。对于il-4rα、γc和il-13rα1表达的测量,b细胞和t细胞是通过细胞表面标志物(cd19、cd4、cd8)门控的,而单核细胞则被识别为cd14+细胞。合适的同种型对照充当阴性对照,提供了平均值和sd。(h)基于il-4和超级因子的s形剂量响应曲线(图19)获得的标准化pstat6ec50值。来源于il-4wt的pstat6ec50被标准化为1,并且相应地计算super-4和kfr的ec50值。配对t检验被于确定显著的变化。
图12描绘在cd4和cd8t细胞中,相较于il-4和kfr,super-4诱导更强的stat6磷酸化。基于il-4和超级因子的s形剂量响应曲线获取的标准化p-stat6ec50值。将来源于il-4野生型的p-stat6ec50值标准化至1,并且相应地计算super-4和kfr的ec50值(l=淋巴细胞,m=单核细胞,ne=中性粒细胞)。
图13描绘由il-4和超级因子表现出的功能活性。(a)在tgf和指示浓度的il-4、super-4或者kfr的存在下,用抗cd3/抗cd28包被珠培养人天然cd4+cd45ra+cd45ro-cd25-t细胞。4天后,在布雷菲德菌素a存在下用pma和离子霉素再刺激细胞另外4小时。随后分析细胞的il-9和foxp3的细胞内表达。数据(平均值和sem)来源于>4个供体的3次独立实验。(b)从健康的血液供体获取的外周血单核细胞分离cd14+单核细胞(>97%纯度),并且仅用50ng/ml的gm-csf培养所述细胞,或者用指示浓度的il-4、kfr或者super-4培养所述细胞。
图14描绘超级因子图解树突状细胞的成熟依赖于ii型il-4受体复合物的激活。cd14、cd86和cd209表面标志物的表达用于指示超级因子对dc成熟的影响。为了这个实验,分离cd14+单核细胞(>97%纯度),并在所示抗体的存在下用50ng/ml的gm-csf和2μg/ml的il-4、kfr或super-4培养。在第6天处理所述细胞,随后用dapi、荧光标记的同种型对照mab、或者抗cd14、cd86和cd209的mab染色。数据(平均值和sem)来源于3个供体。
图15描绘单核细胞中的il-4和两种超级因子的信号传导和内化动力学。从外周血单核细胞分离cd14+单核细胞(>97%纯度),并用30pm的il-4、super-4或者kfr刺激所示时间。随后收获细胞,并保持在4℃并且用特异于il-13rα1和γc受体链的抗体染色以用于受体下调实验,或者用4%的pfa固定,用100%的甲醇透化,并用抗stat6和irs1的磷酸特异性抗体染色以用于信号传导动力学实验。在上述两种情况中,细胞都是用流式细胞计数法分析的。数据(平均值和sem)来源于4个健康的供体。
图16描绘由单核细胞中的il-4和两种超级因子诱导的基因表达谱。用单独或者与il-4或所述两种超级因子组合的gm-csf刺激来源于5名健康供体的高度纯化的单核细胞六小时,提取所述细胞的rna,并且用双色agilent微阵列(60,000特性)分析。(a)显示了比较三个细胞因子的基因诱导的散布图相关性。(b)显示了维恩图(venndiagram),其中当与gm-csf对照条件相比时,基因被该三个细胞因子显著地下调/上调。(c)热图(heatmap)显示被il-4/super-4对kfr、il-4/kfr对super-4,或者il-4对super-4/kfr显著调节的所选基因列表(表4)(p<0.05,配对t检验)。蓝色代表较高的表达,并且红色代表较低的表达。
图17描绘由不成熟的以及lps-成熟的dc中的il-4和两种超级因子诱导的不同模式的细胞因子分泌。用单独或者与il-4、kfr或者super-4(20ng/ml)组合的gm-csf(50ng/ml)培养来源于3名健康供体的经纯化的单核细胞7天,随后用lps(2μg/ml)刺激(或不刺激)另外24小时。用luminex估量培养上清液中51种细胞因子、趋化因子和生长因子的相对量(用热图列出)。热图的右侧图是产物的代表性实例,其分泌未改变(n=19),仅lps刺激增强(n=20),或者在存在或者不存在lps的情况中被超级因子调节(n=12)。数据代表来源于3名健康供体的平均值和sd(标准化至单独gm-csf的组)。配对t检验用于测定显著的变化,*p<0.05)。
图18显示在抗γc(0μg/ml、5μg/ml,或者50μg/ml)存在下,响应于il-4、super-4和kfr,ramos细胞和u937细胞中stat6的磷酸化。不存在抗γc下的响应被标准化至100%,并且存在抗γc下的响应被表述为与标准值有关。数据(平均值±s.e.m)来源于三次独立实验。
图19显示由il-4和两种超级因子诱导的stat6磷酸化。用指定剂量的il-4和两种超级因子刺激细胞,并且使用荧光标记的磷酸stat6特异性抗体和流式细胞计数法来检测(a)cd4t细胞、(b)cd8t细胞、(c)单核细胞、(d)b细胞、和(e)hh细胞中stat6磷酸化的水平。平均值和sem是从三个不同的实验获得的。
图20示例说明由单核细胞中的il-4和两种超级因子诱导的受体下调与stat6和irs1磷酸化动力学。用50nm的il-4或者两种超级因子刺激单核细胞所示时间,并且细胞(a-b)经固定并且用100%的甲醇透化30分钟并且用磷酸-stat6抗体和磷酸-irs1抗体染色;或者(c-d)用抗il-13rα1或γc特异性抗体染色并且保持在4℃下30分钟以防止受体内化,用4%的pfa固定并且通过流式细胞计数法分析。数据(平均值和sem)来源于四个健康供体。
图21示例说明il-4、krf和super-4对细胞因子分泌的差异性诱导。用50ng/ml的gm-csf,使用或者不使用il-4、kfr或者super-4(20ng/ml),培养单核细胞7天,随后用lps(2mg/ml)刺激24小时,数据代表来源于3名健康供体的平均值±sd(标准化至未经刺激的单独gm-csf的组)。*p<0.05(配对t检验)。
图22示例说明用il-4超级因子呈现的树突状细胞的分化潜能。分离cd14+单核细胞(>97%纯度),并随后用50ng/ml的单独gm-csf或者用指示浓度的il-4、kfr或者super-4培养。在第6-7天处理细胞,并用针对cd86、cd209的荧光标记抗体染色。通过使用bdlsrii流式细胞仪的流式细胞计数法来评估树突状细胞的分化,并在flowjo软件(treestar)上测定平均荧光强度(mfi)。数据代表来源于3个健康供体的平均值±sem。
图23示例说明在il-4、kfr或者super-4的存在下,表面标志物的差异表达。用50ng/ml的gm-csf,使用或者不使用il-4、kfr或者super-4(20ng/ml),培养单核细胞7天。数据代表来源于3个健康供体的平均值±sd(标准化至单独gm-csf的组)。*p<0.05(配对t检验)。未检测到高于背景水平的cd1c(bdca-1)和cd141(bdca-3)。
具体实施方式
介绍
细胞因子被广泛地定义为由一个细胞产生并且作用于另一细胞的分子。这些分子还称为白细胞介素、干扰素、生长因子和tnf,以及其他名称,参与基本上每一个重要的生物过程,从细胞增殖到炎症、免疫性、迁移、纤维化、修复和血管形成。由于这些性质,已经探索细胞因子作为治疗剂的用途。不幸地是,在大多数情况下,使用野生型细胞因子作为治疗剂往往与剂量限制性毒性或疗效不足相关。
细胞因子可以基于三维结构划分为两类,i型和ii型。
i型细胞因子具有四个α螺旋形管束结构,具有“上-上-下-下”的构造。这些细胞因子可以基于α螺旋的长度以及一些其他结构/拓扑因素而进一步划分为短链和长链的四α螺旋形管束细胞因子。在短链细胞因子中,螺旋长度通常是约15个氨基酸,而长链细胞因子的螺旋长度通常是25个氨基酸。
在短链细胞因子中,ab环在cd环“下面”,而在长链细胞因子中该ab环在cd环“上面”。另一不同性质是仅短链细胞因子具有在ab环和cd环内的β折叠结构。
短链细胞因子的实例是il-2、il-3、il-4、il-5、il-7、il-9、il-13、il-15和il-21,而示例性的长链细胞因子是il-6、il-11和il-12。
ii型细胞因子具有不同的结构。例如,干扰素(ifn)-β具有额外的螺旋代替cd链,ifn-γ结合为六个螺旋/二聚物的α二聚物,以及il-10结合至其受体作为两组二聚物,每个il-10域由六个α螺旋组成,所述α螺旋由两个缠绕在一起的肽链组装而成,螺旋a-d来源于一个链,并且螺旋e和f来源于双股相关链。
其他的ii型细胞因子是il-20和il-22。
细胞因子通过与在靶细胞上表达的特异性受体相互作用来发挥其功能。像细胞因子一样,细胞因子受体可以基于结构相似性而分组成不同的相关蛋白家族。这些受体落入不同的相关蛋白家族,并且有时被分类成四大组。i型细胞因子受体属于造血因子受体家族。
i型细胞因子受体是多聚的,因为它们具有一个α受体和一个或两个可共有的次要组分。i型细胞因子受体可以基于常见的受体组分的共有而被分成三个亚家族。i型细胞因子受体可以共有常见的β链(βc)、gp130或者常见的γ链(γc)。
共有βc的细胞因子的实例是il-3、il-5和gm-csf;共有gp130的细胞因子是例如il-6和il-11;以及共有γc的细胞因子是例如il-2、il-3、il-7、il-9、il-13、il-15和il-21。
改良细胞因子的一种可能性是使其偏向对某些希望细胞类型的优选活性。α受体负责基于不同类型细胞表面上选择性表达的细胞因子特异性和细胞特异性。共有受体也是差异表达的。因此,细胞因子对某些细胞类型优选的活性可以通过操纵配体来工程改造。例如,操纵共有受体募集的结合参数可以使细胞因子的活性向某些细胞类型偏移。这可以应用于通过共有和/或异源二聚体受体进行信号传导的宽范围细胞因子。
为了使本发明更易于理解,将在下文以及贯穿说明书定义某些术语和短语。
定义
除非另外定义,否则在本文中使用的所有技术和科学术语具有本发明所属领域的普通技术人员所通常理解的相同含义。通常,本文所使用的命名法则和在细胞培养、分子遗传学、有机化学中所使用的实验方法是本领域中众所周知并且通常使用的那些。标准技术用于核酸和肽的合成。通常根据本领域和各种一般参考文献中的常规方法(一般参见,sambrook等人,molecularcloning:alaboratorymanual,第2版(1989)coldspringharborlaboratorypress,coldspringharbor,n.y.,该参考文献以引用方式并入本文)来进行所述技术和方法,其在本文件通篇提供。本文所使用的命名法则和如下所述的分析与合成有机化学的实验方法是本领域熟知且常用的那些。标准技术或其改良用于化学合成和化学分析。
本文中的“野生型”或者“wt”或“天然的”表示在自然界中发现的氨基酸序列或者核苷酸序列,包括等位基因变化形式。“野生型il-4”表示il-4,无论是天然的还是重组的,具有天然人il-4的129个正常存在的氨基酸序列,seqidno:1,不包含24个氨基酸il-4信号肽。
如本文所使用的,“突变蛋白”表示相对于野生型多肽在一个或多个位点处有氨基酸插入、缺失、取代和修饰的多肽。示例性的突变蛋白可以包括对1个、2个、3个、4个、5个、6个、7个、8个、9个、10个或更多个氨基酸的取代。
突变蛋白还包括对整个野生型多肽或多聚核苷酸序列的保守修饰和取代(例如,对突变蛋白的二级结构或三级结构具有最小影响的那些)。这种保守性取代包括在dayhoff的theatlasofproteinsequenceandstructure5(1978)和argos的emboj.,8:779-785(1989)中描述的那些。例如,属于下列组之一的氨基酸代表保守性变化:组i:ala、pro、gly、gln、asn、ser、thr;组ii:cys、ser、tyr、thr;组iii:val、ile、leu、met、ala、phe;组iv:lys、arg、his;组v:phe、tyr、trp、his;以及组vi:asp、glu。
如本文所使用的,“共有受体多肽”指多于一个配体之间共有的多肽受体。示例性共有受体是被il-4结合的那些,包括但不限于γc、gp130、βc或者il-13rα1。
如本文所使用的,“配体”指能够特异性地结合至另一分子例如受体的任何分子。术语“配体”包括激动剂和拮抗剂两者,并且可以是例如小分子、抗体片段、sirna、反义核酸、诸如突变蛋白的多肽、dna和/或rna。
如本文所使用的,“靶细胞”指表达共有受体多肽的细胞,或者表达能够结合至本发明的突变蛋白的受体多肽的细胞。靶细胞的实例可以包括但不限于b细胞、cd4t细胞、巨噬细胞、单核细胞以及上皮细胞。
“根据野生型编号”表示提及所选氨基酸在野生型多肽的成熟序列中正常存在的位置来鉴别该氨基酸,例如118应指在seqidno:1中存在的第一百一十八个氨基酸。
如本文提及多肽或dna序列使用的术语“同一性”指两个分子之间亚基序列的同一性。当两个分子中的亚基位置被相同的单体亚基(例如,相同的氨基酸残基或核苷酸)占据时,则所述分子在该位置处是同一的。两个氨基酸或者两个核苷酸序列之间的相似性是同一性位置数量的直接函数。一般说来,对所述序列进行比对以便获得更高级别的匹配。如有必要,可以使用公开的技术和广泛可用的计算机程序来计算同一性,诸如gcs程序包(devereux等人,nucleicacidsres.12:387,1984)、blastp、blastn、fasta(atschul等人,j.molecularbiol.215:403,1990)。可以使用序列分析软件来测量序列同一性,所述序列分析软件诸如universityofwisconsinbiotechnologycenter(1710universityavenue,madison,wis.53705)的geneticscomputergroup的sequenceanalysissoftwarepackage,使用软件的默认参数。
术语“多肽”、“蛋白”或者“肽”指氨基酸残基的任何链,而不论其长度或翻译后修饰(例如,糖基化或磷酸化)。
在本公开的突变蛋白多肽是“基本上纯的”的情况下,突变蛋白多肽可以占目标多肽的至少约60重量%(干重)。例如,所述多肽可以占目标多肽的至少约75重量%、约80重量%、约85重量%、约90重量%、约95重量%或者约99重量%。可以通过任何合适的标准方法来测量纯度,例如柱色谱法、聚丙烯酰胺凝胶电泳或者hplc分析。
如本文所使用的术语“有效量”指引起所需的生物反应所必需的量。药物的有效量可根据诸如以下因素而变化:所需的生物终点、待递送的药物、任何其他活性或失活成分的组成,等等。
本文使用的术语“表达”表示从dna产生多肽的过程。所述过程包括基因转录成mrna,以及此mrna翻译成多肽。根据其使用的上下文,“表达”可指rna、蛋白、或rna和蛋白两者的产生。
如本文所使用的术语“基因产物”表示由该基因编码的rna(例如信使rna(mrna)或微rna(mirna))或者蛋白。
如本文所使用的,术语“经分离的”指分子是基本上纯的。经分离的蛋白可以是基本上纯的,例如60%、65%、70%、75%、80%、85%、90%、95%或者99%不含其他不同的蛋白分子。
如本文所使用的,术语“调节(modulate/modulation)”或者“操纵(manipulate/manipulation)”泛指特异性靶向基因(包括其rna和/或蛋白产物)、信号传导途径、细胞和/或靶向表型的下调(例如,抑制或遏制),或者靶向基因的上调(例如,诱导或增加)。
“患者”或“受试者”表示哺乳动物,例如人,其患有或有风险发展诸如炎性疾病的疾病或病症,或者患有或被诊断为患有炎性疾病,或者可以其他方式受益于本文所描述的组合物和方法。
如本文所使用的,术语“减少”以及其语法上的等效形式指对表达或基因产物活性的任何抑制、减少、降低、遏制、下调或阻止。例如,表达或者活性的水平可以是未受抑制的表达或活性的例如100%或者小于100%,例如小于95%、小于90%、小于85%、小于80%、小于75%、小于70%、小于65%、小于60%、小于55%、小于50%、小于45%、小于40%、小于35%、小于30%、小于25%、小于20%、小于15%、小于10%,或者小于5%。
术语“治疗(treating/treatment)”或者“减轻”或者“改善”指治疗性治疗和预防或防治性措施,其中目的是预防或者延迟(减轻)靶向的病理病症或疾患。
il-4
il-4是经典的四α螺旋管束细胞因子。其主结合链是il-4rα,il-4rα是ii型细胞因子受体链,由胞外域、跨膜域和长胞内尾部组成,含有box1域、ptb域蛋白的结合位点以及三个stat6停靠和激活位点。
野生型人il-4包括24个氨基酸的信号肽和129个氨基酸的氨基酸链肽。seqidno:1是成熟的129个氨基酸的人il-4序列。
mgltsqllpplffllacagnfvhghkcditlqeiiktlnslteqktlcteltvtdifaaskntteketfcraatvlrqfyshhekdtrclgataqqfhrhkqlirflkrldrnlwglaglnscpvkeanqstlenflerlktimrekyskcss(seqidno:1)。
il-4/il-4rα复合物充当il-4受体的第二组分的配体,其对于i型受体是γc并且对于ii型受体是il-13rα1。细胞表面上il-4/il-4rα/γc或者il-4/il-4rα/il-13rα1复合物的形成激活细胞内的信号传导途径,所述信号传导途径包括jak-stat途径和pi3k/akt途径,后一途径通过ptb域蛋白(特别是irs2)的募集来动员。与il-4结合的i型和ii型il-4受体的胞外域的晶体结构的最近研究(图1a)显示il-4位于由il-4rα和第二受体链形成的“y形叉”中,并且通过细胞因子的d螺旋上的结合表面与第二受体链直接接触。
il-4与il-4rα的结合是非常高亲和力的,具有约10-10m的kd。il-4/il-4rα复合物与γc或者il-13rα1的后续结合是相对低亲和力的。由于il-4与il-4rα的结合的非常高的亲和力,在大部分情况下信号传导复合物的形成是由第二链的相对表达水平来决定的。此外,替代性的第二链具有不同的细胞表达模式,γc主要在造血细胞上表达,并且il-13rα1主要在非造血细胞上表达,尽管一些造血细胞也表达可变量的il-13rα1。il-4的大部分调节活性是由主要表达i型受体的b细胞和t细胞介导的,而其大部分效应子功能(其模拟il-13)是由唯一表达ii型受体并且也对il-13响应的细胞介导的。
功能上,il-4通过分别作用于天然cd4t细胞、b细胞和巨噬细胞来调节cd4t细胞的th2分化、免疫球蛋白类型转换成ige,以及替代性的巨噬细胞激活。因为il-4结合至主要在非造血细胞上表达的ii型受体,与其结合i型受体一样有效,其可以诱导效应子功能以及调节功能。效应子功能是气道超敏性和杯状细胞化生。然而,后面的这些活性在生理上主要是利用il-13由ii型受体诱导的,因为il-13的量远大于il-4。此外,因为il-13无法结合至主要在造血细胞上表达的i型受体,所以il-13很少或没有“调节”活性。总的说来,通过使用i型和ii型il-4受体,il-4在th2型炎症中起重要作用,不论该炎症是由过敏原引发的还是由侵入的细胞外寄生虫引发的。
il-4当前并未用作治疗剂,但是其已经在过去被考虑用于此类用途,并且如果无毒性,可以被考虑用于以下目的:诸如在疫苗接种期间指导cd4t细胞分化,或者鉴于对已分化的cd4t细胞的可塑性的最近识别来改变分化的产生模式。在20世纪90年代早期,进行临床试验,在所述临床试验中将il-4施用给癌症患者,以期促进t细胞反应或者参与先天免疫系统。
然而,大剂量(600μg/m2/天)il-4的静脉内施用导致研究组中三分之二的患者的血管渗漏综合征。在这些研究和临床前分析中还遭遇了其他毒性。虽然这些毒性中有许多毒性涉及il-4作用于非造血细胞,但是il-4在小鼠体内具有明显的巨噬细胞介导的毒性。将il-4施用至小鼠导致由替代地激活的巨噬细胞介导的噬血细胞综合征。
因为il-4通过结合至ii型受体和后续的效应子功能而对非造血细胞的活性,使用il-4药理学地调节淋巴细胞分化是复杂的。因此产生无法激活ii型受体或者其中i型受体的激活以比ii型受体的激活明显降低的浓度实现的il-4突变蛋白将是有利的。
因此,在一些实施方式中提供了经工程改造的il-4突变蛋白,与野生型il-4相比,该il-4突变蛋白具有对i型和ii型受体的第二链改变的相对结合活性。在一些实施方案中,il-4突变蛋白调节或增强信号传导,而不是阻断信号传导。
本文描述了人il-4突变蛋白的产生,人il-4突变蛋白被称为“超级因子”,具有对γc非常高的亲和力以及对il-13rα1减弱的亲和力,并且相反地,那些il-4突变蛋白以比il-4高得多的亲和力结合至il-13rα1,其对γc的亲和力很少或者没有变化。
因此,在一些实施方案中,il-4突变蛋白具有对γc相对高的亲和力和对il-13rα1减弱的亲和力。在其他实施方案中,il-4突变蛋白具有比il-4相对高得多的对il-13rα1的亲和力,其对γc的亲和力很少或没有变化。
il-4超级因子(突变蛋白)
在各种实施方案中,本公开提供了il-4突变多肽,所述突变多肽可以但不一定是基本上纯化的,并且可以作用为野生型il-4的激动剂;实现il-4的一种或多种生物学活性(例如,刺激细胞增殖)。
示例性的突变il-4多肽包括具有与seqidno:1至少约80%的同一性的氨基酸序列,该氨基酸序列以比seqidno:1代表的多肽结合至γc的亲和力更大的亲和力结合γc,并且具有对il-13rα1减弱的亲和力。例如,相对于野生型il-4,突变il-4多肽可以具有至少一个突变(例如,1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、11个、12个、13个、14个、15个、16个、17个、18个、19个、20个或更多个氨基酸残基的缺失、添加或者取代),并且以比seqidno:1的il-4更大的亲和力结合γc,并且与seqidno:1对il-13rα1的亲和力相比,具有对il-13rα1减弱的亲和力。
示例性的突变il-4多肽包括具有与seqidno:1至少约80%的同一性的氨基酸序列,该氨基酸序列以比il-4(seqidno:1)更高的亲和力与il-13rα1结合,相对于seqidno:1的多肽,该突变il-4多肽对γc的相对亲和力很少或没有变化。例如,相对于野生型il-4,突变il-4多肽可以具有至少一个突变(例如,1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、11个、12个、13个、14个、15个、16个、17个、18个、19个、20个或更多个氨基酸残基的缺失、添加或者取代),并且该突变il-4多肽以比il-4(seqidno:1)更大的亲和力与il-13rα1结合,相对于seqidno:1的多肽,该突变il-4多肽对γc的相对亲和力很少或没有变化。
示例性的突变il-4多肽可以是与野生型il-4至少约50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%同一的。所述突变可以由氨基酸残基的数量或含量变化组成。例如,突变il-4可以具有比野生型il-4更多或更少数量的氨基酸残基。替代地或者附加地,示例性的突变多肽可以含有对存在于野生型il-4中的一个或多个氨基酸残基的取代。在各种实施方案中,所述突变il-4多肽可以通过单一氨基酸残基的添加、缺失或者取代而不同于野生型il-4,例如对位置121处的残基的取代。类似地,示例性的突变il-4多肽可以通过对两个、三个、四个、五个、六个、七个、八个或者更多个氨基酸残基的取代而不同于野生型,所述残基例如seqidno:1的位置117、118、21、122、124、125、128和129处的残基。
举例而言,包括与seqidno:1的参考氨基酸序列至少90%同一的氨基酸序列的多肽是包含与参考序列同一的序列的多肽,除了包含对seqidno:1的参考氨基酸的至多13个改变。例如,参考序列中至多10%的氨基酸残基可以缺失或者被另一种氨基酸取代,或者占参考序列中总氨基酸残基至多10%的大量氨基酸被插入到参考序列中。参考序列的这些改变可以发生在参考氨基酸序列的氨基(n--)或羧基(c--)末端位置,或者发生在那些末端位置之间的任何地方,被个体地引入参考序列的残基中或者以一个或多个连续组引入参考序列中。
经取代的氨基酸残基可以但不一定是保守性的取代,所述取代通常包括在以下组内的取代:甘氨酸、丙氨酸;缬氨酸、异亮氨酸、亮氨酸;天冬氨酸、谷氨酸;天冬酰胺、谷氨酰胺;丝氨酸、苏氨酸;赖氨酸、精氨酸;以及苯丙氨酸、酪氨酸。
例如,取代r121q指变体多肽,在这种情况下其中位置121处的精氨酸被谷氨酰胺替代。而且,取代可以仅在亲本多肽中的位置74处进行,例如121q将指用谷氨酰胺取代位置121处的氨基酸,而不管亲本多肽的位置121处存在何种氨基酸。
本文的“蛋白变体”或者“变体蛋白”或者“变体多肽”表示借助于至少一个氨基酸修饰而与野生型蛋白不同的蛋白。亲本多肽可以是天然存在的或者野生型(wt)多肽,或者可以是wt多肽的改良形式。变体多肽可以指多肽本身、包括该多肽的组合物,或者编码该多肽的氨基酸序列。优选地,该变体多肽与亲本多肽相比具有至少一个氨基酸修饰,例如与亲本相比,从约一个到约十个氨基酸修饰,以及优选地从约一个到约五个氨基酸修饰。
本文使用的“亲本多肽”、“亲本蛋白”、“前体多肽”或者“前体蛋白”表示未经修饰的多肽,该多肽随后将经修饰以产生变体。亲本多肽可以是野生型多肽,或者是野生型多肽的变体或者工程改造形式。亲本多肽可以指多肽本身、包括该亲本多肽的组合物,或者编码该亲本多肽的氨基酸序列。
关于亲和力,本文公开了示例性的突变il-4多肽,该突变il-4多肽以比野生型il-4多肽更高的亲和力结合γc,该亲和力比野生型il-4多肽高至少约2%、至少约5%、至少约10%、至少约20%、至少约30%,或者至少约40%或更高。该野生型il-4多肽以约3300nm的kd结合γc。所公开的示例性突变il-4多肽的结合亲和力还可以表示为比野生型il-4高1.2、1.4、1.5、2、5、10、15、20、25、50、100、200、250、500、1000、1500、2000、2500、3000、3500或更多倍的对γc的亲和力。
示例性的突变il-4多肽可以具有表现出与γc受体亚基升高的缔合速率的能力。
本公开还提供了以比野生型il-4多肽低的亲和力结合γc的突变il-4多肽,该突变il-4多肽的γc亲和力比该野生型il-4多肽低至少约2%、至少约5%、至少约10%、至少约20%、至少约30%,或者至少约40%或者更多。所公开的示例性突变il-4多肽的结合亲和力还可以表示为比野生型il-4低1.2、1.4、1.5、2、3、4、5、10、15、20或更多倍的对γc的亲和力。
关于亲和力,本文公开了示例性的突变il-4多肽,该突变il-4多肽以比野生型il-4多肽更高的亲和力结合il-13rα1,该亲和力比野生型il-4多肽高至少约2%、至少约5%、至少约10%、至少约20%、至少约30%,或者至少约40%或更高。该野生型il-4多肽以约4200nm的kd结合il-13rα1。所公开的示例性突变il-4多肽对il-13rα1的结合亲和力还可以表示为比野生型il-4高1.2、1.4、1.5、2、5、10、15、20、25、50、100、200、250、400、450、500、1000、1500、2000或更多倍的对il-13rα1的亲和力。
示例性的突变il-4多肽可以具有表现出与il-13rα1受体亚基升高的缔合速率的能力。
本公开还提供了以比野生型il-4多肽低的亲和力结合il-13rα1的突变il-4多肽,该突变il-4多肽的il-13rα1亲和力比该野生型il-4多肽低至少约2%、至少约5%、至少约10%、至少约20%、至少约30%,或者至少约40%或者更多。所公开的示例性突变il-4多肽的结合亲和力还可以表示为比野生型il-4低1.2、1.4、1.5、2、3、4、5、10、15、20或更多倍的对il-13rα1的亲和力。
还公开了与野生型il4相比,具有升高的γc亲和力和减小的il-13rα1亲和力这两种性质的示例性突变il-4多肽。
还公开了与野生型il4相比,具有升高的γc亲和力和升高的il-13rα1亲和力这两种性质的示例性突变il-4多肽。
还公开了与野生型il4相比,具有减小的γc亲和力和减小的il-13rα1亲和力这两种性质的示例性突变il-4多肽。
与细胞因子相关的疾病
增殖和分化疾患
在一些实施方案中,突变细胞因子多肽和/或表达突变细胞因子多肽的核酸可以施用至受试者以治疗与异常的凋亡或者分化过程相关的疾患(例如,细胞增殖疾患或者细胞分化疾患,诸如癌症)。
如本文所使用的,术语“癌症”(或者“癌性的”)、“过度增殖”和“肿瘤的”是指细胞具有自主性生长能力(例如,以快速增殖的细胞生长为特征的异常状态或状况)。过度增殖和肿瘤疾病状态可以被分类为病理性的(例如,表征或构成疾病状态),或者过度增殖和肿瘤疾病状态可以被分类为非病理性的(例如,偏离正常但与疾病状态无关)。这些术语意欲包括各种类型的癌性生长或致癌过程、转移性组织,或者恶性转化的细胞、组织或器官,而不论其组织病理类型或侵袭阶段。
癌症的实例包括但不限于癌瘤、肉瘤,和造血系统肿瘤性疾患,例如,白血病)。
突变细胞因子多肽可用于治疗患有、怀疑患有或可能处于高风险发展任何类型癌症或任何病毒性疾病的患者,所述癌症包括肾癌或者黑素瘤。示例性的癌瘤包括从宫颈、肺、前列腺、乳腺、头颈、结肠和卵巢的组织形成的那些。该术语还包括癌肉瘤,癌肉瘤包括由癌组织和肉瘤组织组成的恶性肿瘤。
如本文所使用的,术语“造血系统肿瘤性疾病”包括涉及造血系统起源的增生性/肿瘤性细胞的疾病,例如起源于骨髓细胞谱系、淋巴细胞谱系或红细胞谱系,或者其前体细胞。优选地,该疾病起源于不良分化的急性白血病(例如,成红血球细胞白血病和急性成巨核细胞白血病)。其他示例性的骨髓疾患包括但不限于急性早幼粒细胞白血病(apml)、急性髓性白血病(aml)和慢性粒性白血病(cml)(参见vaickus,l.(1991)critrev.inoncol./hemotol.11:267-97);淋巴系统恶性肿瘤包括但不限于:包括b系all和t系all的急性淋巴母细胞性白血病(all),慢性淋巴细胞性白血病(cll),幼淋巴细胞性白血病(pll)、多毛细胞白血病(hll)和瓦尔登斯特伦氏巨球蛋白血症(wm)。
恶性淋巴瘤的其他形式包括但不限于非霍奇金淋巴瘤及其变体、外周t细胞淋巴瘤、成年t细胞性白血病/淋巴瘤(atl)、皮肤t细胞淋巴瘤(ctcl)、大颗粒淋巴细胞白血病(lgf)、霍奇金氏病和瑞德-史登伯氏病。
增殖和/或分化疾患的其他实例包括皮肤疾患。皮肤疾患可以涉及真皮层、表皮层或者皮下层中的一种细胞或者一组细胞或层的异常活性,或者真皮-表皮连接的异常。
例如,皮肤疾患可以涉及角化细胞的异常活性(例如,过度增殖的基底和上基部角化细胞)、黑色素细胞、郎格罕氏细胞、默克尔细胞(merkelcell)、免疫细胞以及在表皮层的一个或多个层中发现的其他细胞的异常活性,所述一个或多个层例如基底层(生发层)、棘层、颗粒层、透明层或角质层。
在其他实施方案中,该疾患可以涉及真皮细胞的异常活性,所述细胞例如在真皮层中发现的真皮内皮细胞、成纤维细胞、免疫细胞(例如,肥大细胞或者巨噬细胞),所述真皮层例如乳头层或者网状层。
皮肤疾患的实例包括银屑病、银屑病关节炎、皮炎(湿疹),例如,剥脱性皮炎或者特异反应性皮炎、毛发红糠疹、玫瑰糠疹、类银屑病、苔癣样糠疹、扁平苔癣、光泽苔癣、鱼鳞病样皮肤病、角皮病、皮肤病、斑秃、坏疽性脓皮病、白斑病、类天疱疮(例如,眼瘢痕性类天疱疮或者大疱性类天疱疮)、荨麻疹、汗孔角化症(prokeratosis)、类风湿性关节炎,所述类风湿性关节炎涉及衬垫关节囊的上皮相关细胞的增生和发炎;皮炎,诸如脂谥性皮炎和日晒性皮炎;角化病,诸如脂溢性角化病、老年角化病、光化性角化病、光诱导的角化症(photo-inducedkeratosis)和毛囊角化病;播散性痤疮;瘢痕瘤和抗瘢痕瘤形成的预防法;痣;疣,包括瘊、湿疣或者尖端湿疣,以及人乳头瘤病毒(hpv)感染,诸如性病疣;黏膜白斑病;扁平苔癣;以及角膜炎。该皮肤疾患可以是皮炎,例如特异反应性皮炎或者过敏性皮炎,或者是银屑病。
作为直接施用给患者的方法的替代或附加,在一些实施方案中,突变细胞因子多肽可以体外方法使用。例如,细胞(例如,外周血单核细胞,或者从患者分离并且放置或者维持在培养物中的经纯化淋巴细胞群)可以在培养基中体外培养,并且接触步骤可以通过添加细胞因子突变体到培养基而实现。培养步骤可以包括其他步骤,其中用其他药剂刺激或处理细胞,例如以刺激增殖,或者以扩增对相关抗原(例如,癌抗原或者病毒抗原)反应性的细胞群。该细胞则在处理之后施用至患者。
肺部疾病
适合用所公开的细胞因子突变蛋白组合物及其使用方法改善的病理性病症包括例如肺部疾病,诸如哮喘和过敏性炎症反应。
例如,哮喘的特征包括呼吸道症状的周期性发作;自发或治疗经常可逆的可变气道阻塞;气道高反应性的存在;以及慢性气道炎症,其中涉及许多细胞和细胞要素,包括例如肥大细胞、嗜酸性细胞、t淋巴细胞、巨噬细胞、中性粒细胞、和上皮细胞(参见,nationalheart,lung,andbloodinstitute:nationalasthmaeducationandpreventionprogram.expertpanelreportguidelinesforthediagnosisandmanagementofasthma.j.allergyclin.immunol.88:425-534,1991;nationalheart,lung,andbloodinstitutenationalasthmaeducationprogramexpertpanelreportii:guidelinesforthediagnosisandmanagementofasthma;1997.nihpublication第97-4051a号)。虽然任何给定的哮喘患者不一定表现出所有这些特征,并且尚未或广泛认可建立哮喘诊断的绝对“最低标准”,但是在具有当前症状和活性哮喘的患者中普遍发现气道高反应性的存在。
气道高反应性是哮喘的特征,并且由以下组成:气道对吸气收缩肌激动剂的敏感性升高,剂量响应曲线的斜率较陡,以及对激动剂更大的最大反应(byrne和inman,chest,2003)。气道反应性的测量可用于哮喘诊断,特别是对具有与哮喘一致的症状但是没有气道阻塞迹象的患者。包括例如环境过敏原在内的某些吸入刺激物增强了气道炎症并且增强了气道高反应性。与具有持续气道高反应性的哮喘患者的气道高反应性变化相比,健康受试者的气道高反应性变化的数量级要小的多。哮喘的发病机理,以及由于根据本方法治疗而获得的改善可通过支气管镜检、支气管肺泡灌洗、气道活组织检查、气道气体测量以及熟练临床医师已知的其他此类方法跟踪。
制剂
可以用药学上可接受的载体配制单独或与一种或多种化疗剂组合的一种或多种治疗剂,以施用至受试者。活性组分可以单独(个体地)配制以顺序施用,或者可以一起配制以同时施用。
如本文所使用的术语“药学上可接受的载体”表示适合施用至受试者的一种或多种相容的固体或液体填充剂、稀释剂或者包封物质。药物组合物的组分还能够以使得没有明显损害希望药效的相互作用的方式互相混合。这种制剂可以常规含有药学上可接受浓度的盐、缓冲剂、防腐剂、相容的载体、助剂以及任选其他治疗成分。
本文描述的组合物可以作为游离碱或作为药学上可接受的盐施用。这种药理学上以及药学上可接受的盐包括但不限于用下列酸制备的那些:盐酸、氢溴酸、硫酸、硝酸、磷酸、顺丁烯二酸、醋酸、水杨酸、对甲苯磺酸、酒石酸、柠檬酸、甲基磺酸、蚁酸、丙二酸、琥珀酸、萘磺酸,以及苯磺酸。而且,药学上可接受的盐可以制备为碱金属盐或碱土盐,诸如羧酸基团的钠盐、钾盐或钙盐。
药物组合物还可包含合适的固相或凝胶相载体或者赋形剂。这种载体或者赋形剂的实例包括但不限于碳酸钙、磷酸钙、各种糖、淀粉、纤维素衍生物、明胶,以及诸如聚乙二醇的聚合物。
合适的缓冲剂包括:乙酸和盐(1-2%w/v);柠檬酸和盐(1-3%w/v);硼酸和盐(0.5-2.5%w/v);以及磷酸和盐(0.8-2%w/v)。合适的防腐剂包括苯扎氯铵(0.003-0.03%w/v);氯丁醇(0.3-0.9%w/v);对羟苯甲酸(0.01-0.25%w/v)和硫柳汞(0.004-0.02%w/v)。
合适的液体或者固体药物制剂形式是例如用于吸入的水或盐水溶液、微囊化、蜗形化(encochleated)、包被在显微金颗粒上、包含在脂质体中(包括ph依赖性控释制剂)、类脂(lipidoid)、喷雾的、气雾剂、用于植入入皮肤的丸剂,或者干燥于尖锐的物体上以刮入皮肤。所述药物组合物还包括颗粒剂、粉剂、片剂、糖衣片剂、(微)胶囊剂、栓剂、糖浆、乳剂、悬浮液、乳霜、滴剂,或者组合物缓释的制剂,在这些制剂中,如上所述地照例使用赋形剂和添加剂和/或辅助剂,诸如崩解剂、粘合剂、包衣剂、膨胀剂、润滑剂、调味剂、增甜剂或者增溶剂。所述药物组合物适合用于多种药物递送系统。关于药物递送方法的简要综述,参见langer,science249:1527-1533,1990以及langer和tirrell,nature,2004年4月1日;428(6982):487-92。
所述组合物可以方便地以单位剂量形式提供,并且可以用药学领域熟知的任何方法制备。在某些实施方案中,组合物以粉末或颗粒形式而非溶液施用。设想为本发明部分的颗粒形式的实例在u.s.2002/0128225中提供。在一些实施方案中,组合物以气雾剂形式施用。在其他实施方案中,组合物可以是粉末形式的,以在使用前用合适的媒介物、例如无菌无热原的水配制。
此外,本文描述的组合物可以配制作为贮库制剂、定时释放、延迟释放或者持续释放的递送系统。此类系统可以避免本文描述的组合物的重复施用,增加对受试者和医师的便利性。此类长效制剂可以用合适的聚合或者疏水性材料(例如作为可接受的油中的乳液)或者离子交换树脂配制,或者配制作为微溶的衍生物,例如作为微溶的盐。
许多类型的释放递送系统是本领域普通技术人员已知并且可获得的。它们包括:基于聚合物的系统,诸如聚乳酸和聚乙醇酸、β-葡聚糖颗粒、聚酐和聚己内酯;为脂类的非聚合物系统,包括固醇(诸如胆固醇、胆固醇酯和脂肪酸)、中性脂(诸如,甘油单酯、甘油二酯和甘油三酯),或者类脂;水凝胶释放系统;硅胶系统;基于肽的系统;蜡质涂层、使用常规粘合剂和赋形剂的压制片剂,部分融合植入剂等等。此外,可以使用基于泵的硬件递送系统,其中一些适用于植入。
还可以用适当的生物相容且可生物降解的赋形剂材料来实现控释。这些实现缓释的聚合物材料可以是用于产生颗粒的任何合适的聚合物材料,包括但不限于不可生物降解的/不可生物降解和生物可降解的/生物可降解的聚合物。这种聚合物已经在现有技术中非常详细的描述,并且包括但不限于:β-葡聚糖颗粒、聚酰胺类、聚碳酸酯类、聚亚烷基类、聚亚烷基二醇、聚氧烷撑、聚亚烷基对苯二酸酯、聚乙烯醇、聚乙烯醚、聚乙烯酯、聚乙烯基卤化物、聚乙烯吡咯烷酮、聚乙醇酸交酯、聚硅氧烷、聚氨酯及其共聚物、烷基纤维素、羟烷基纤维素、纤维素醚、纤维素酯、硝化纤维素、丙烯酸酯和甲基丙烯酸酯的聚合物、甲基纤维素、乙基纤维素、羟丙基纤维素、羟丙基甲基纤维素、羟丁基甲基纤维素、乙酸纤维素、丙酸纤维素、乙酸丁酸纤维素、醋酞纤维素、羧乙基纤维素、三乙酸纤维素、纤维素硫酸钠、聚(甲基丙烯酸甲酯)、聚(甲基丙烯酸乙酯)、聚(甲基丙烯酸丁酯)、聚(甲基丙烯酸异丁酯)、聚(甲基丙烯酸己酯)、聚(甲基丙烯酸异癸酯)、聚(甲基丙烯酸月桂酯)、聚(甲基丙烯酸苯酯)、聚(丙烯酸甲酯)、聚(丙烯酸异丙酯)、聚(丙烯酸异丁酯)、聚(丙烯酸十八酯)、聚乙烯、聚丙烯聚(乙二醇)、聚(环氧乙烷)、聚(对苯二甲酸乙二酯)、聚(乙烯醇)、聚(乙酸乙烯酯)、聚氯乙烯聚苯乙烯、聚乙烯吡咯啶酮、透明质酸,以及硫酸软骨素。在一个实施方案中,所述缓释聚合物是嵌段共聚物,诸如聚(乙二醇)(peg)/聚(乳酸-羟基乙酸共聚物)(plga)嵌段共聚物。
不可生物降解的聚合物的实例包括乙烯乙酸乙烯酯、聚(甲基)丙烯酸、聚酰胺,及其共聚物和混合物。
生物可降解的聚合物的实例包括合成聚合物,例如β-葡聚糖颗粒、乳酸和乙醇酸的聚合物、聚酐、聚(邻)酯类、聚氨酯、聚(丁酸)、聚(戊酸)、聚(己内酯)、聚(羟基丁酸酯)、聚(丙交酯-乙交酯)共聚物和聚(丙交酯-己内酯)共聚物,以及天然聚合物,诸如海藻酸盐和其他的多聚糖,包括葡聚糖和纤维素、胶原,其化学衍生物(化学基团的取代、添加,例如,烷基化、烯化、羟化、氧化,以及由本领域内技术熟练人员按惯例做出的其他修改)、白蛋白和其他亲水性蛋白、玉米醇溶蛋白和其他醇溶蛋白以及疏水性蛋白,及其共聚物和混合物。通常,这些材料通过酶促水解或者在体内暴露于水,通过表面或者整体腐蚀而降解。前述材料可以单独使用,作为物理混合物(掺混物)使用,或者作为共聚物使用。优选的聚合物是聚酯、聚酐、聚苯乙烯及其掺混物。
将有效量的本文描述的组合物施用至需要这种治疗的受试者。有效量是导致病症、疾病或疾患,或者所述病症、疾病或疾患的症状得到所需改善的那些量。
有效剂量范围从1ng/kg体重至100mg/kg体重,或者从100ng/kg体重至50mg/kg体重,或者从1μg/kg体重至10mg/kg体重,取决于施用方式。或者,有效剂量可以范围从每4平方厘米面积的细胞3微克至14毫克。绝对量将取决于多种因素(包括施用是否和其他治疗方法一起进行,剂量数以及个体患者参数,包括年龄、身体状况、大小和体重),并且可以用常规实验确定。一种可能的有用剂量是符合合理医学判断的最高安全剂量。
各种活性剂的递送之间的时间可以用动力学、递送、释放、药剂药效学、药剂药代学或其任何组合的基本原理合理地确定。或者,各种药剂的递送之间的时间可以通过实验凭经验确定,以确定何时可以实现最大作用。
施用方式
施用方式可以是任何医学上可接受的方式,包括:口服施用、舌下施用、鼻内施用、气管内施用、吸入、眼部施用、局部施用、经皮施用、皮内施用、直肠施用、阴道施用、皮下施用、静脉内施用、肌内施用、腹膜内施用、胸骨内施用,或者经粘膜施用。此外,施用方式可以通过体外装置和/或组织穿透电磁装置。
所选择的具体方式将取决于所选择的具体活性剂、希望的结果、被治疗的具体病症,以及实现疗效所需的剂量。一般说来,本文所描述的方法可以使用任何医学上可接受的施用方式来进行,例如使用产生有效水平的炎性反应改变但不引起临床上不可接受的副作用的任何方式。
根据疾患和施用方式,组合物可以不同的容器、媒介物或制剂提供。例如,对于口服应用,组合物可以作为舌下片剂、口香糖、漱口剂、牙膏、糖果、凝胶剂、膜剂等等施用;对于眼部应用,组合物可作为滴眼剂以滴管施用,作为眼膏、眼凝胶剂、眼膜,作为隐形眼镜或者眼内透镜的被膜,以隐形眼镜贮存液或洗液的方式等等施用;对于局部应用,作为洗液、软膏、凝胶剂、乳霜、喷雾剂、棉纸、拭子、搽剂等等;对于阴道或者直肠应用,作为软膏、棉塞、栓剂、黏膜粘着制剂等等。
组合物可以通过注射施用,例如通过快速推注注射或者连续输注,通过静脉内、皮下、肌内、腹膜内、胸骨内途径。用于注射的制剂可以单位剂量形式提供,例如在安瓿或者多剂容器中,含有添加的防腐剂。组合物可以采取诸如油性或水性媒介物中的悬浮液、溶液或者乳液的形式,并且可以含有配制剂,诸如悬浮剂、稳定剂和/或分散剂。对于口服施用,组合物可以通过将组合物与本领域熟知的药学上可接受的载体组合而容易地配制,例如作为舌下片剂、液体制剂或者口服凝胶剂。
对于通过吸入施用,组合物可以气雾剂喷雾形式从加压包或喷雾器方便地递送,借助合适的推进剂,例如二氯二氟甲烷、三氯氟甲烷、二氯四氟乙烷、二氧化碳或者其他合适的气体。在加压气雾剂的情况下,可以通过提供阀门以递送计量的量来确定剂量单位。用于吸入器或者吹入器的例如明胶的胶囊剂和药筒可以配制为含有组合物的粉末混合物和合适的粉末基底诸如乳糖或者淀粉。用于吸入治疗剂的医疗设备是本领域已知的。在一些实施方案中,所述医疗设备是吸入器。在其他实施方案中,所述医疗设备是计量吸入器、碟型吸入器、涡轮吸入器(turbuhaler)、干粉吸入器(diskus)或者雾化器(spacer)。在这些实施方案中的某些中,所述吸入器是spinhaler(rhone-poulencrorer,westmalling,kent)。其他医疗设备是本领域已知的,并且包括inhale/pfizer、mannkind/glaxo和advancedinhalationresearch/alkermes。
组合物还可以配制成直肠或者阴道组合物,诸如栓剂或者保留灌肠剂,例如含有常规的栓剂基底,诸如可可脂或者其他甘油酯。
其他用途
本文描述的突变蛋白可以直接或者间接地用是标记或者产生信号的部分标记,所述部分例如酶、放射性同位素标记、表位、或者荧光蛋白(诸如,绿色荧光蛋白)。本文描述的突变蛋白可以与样品或者细胞接触,以例如使用标准免疫印迹法、免疫荧光法、酶免疫分析法(eia)、放射免疫测定法(ria)、荧光能量传递、蛋白质印迹以及其他诊断和检测技术来确定该样品中或者细胞上是否存在受体。
本文描述的突变蛋白还可以经标记以用于体内检测和施用给受试者。受试者可以例如通过nmr或者其他层析成像手段来成像。例如,粘合剂可以用放射性同位素标记(诸如,131i、111in、123i、99mtc、32p、125i、3h、14c和188rh)、荧光标记(诸如荧光素和若丹明)、核磁共振活性标记、正电子发射层析成象(“pet”)扫描仪可检测的正电子发射同位素、化学发光剂(诸如荧光素)和酶标志物(诸如,过氧化物酶或者磷酸酶)来标记。本文描述的突变蛋白可以用造影剂标记,所述造影剂例如顺磁性剂和铁磁性剂或超顺磁性剂。
本文描述的突变蛋白可以偶联至放射性同位素,诸如α辐射源、β辐射源,或者γ辐射源。放射性同位素的实例包括碘(诸如,131i、111in、123i、99mtc、32p、125i、3h、14c和188rh)、荧光标记(诸如荧光素和若丹明)、核磁共振活性标记、正电子发射层析成象(“pet”)扫描仪可检测的正电子发射同位素、化学发光剂(诸如荧光素)和酶标志物(诸如,过氧化物酶或者磷酸酶)来标记。本文描述的突变蛋白可以用造影剂标记,所述造影剂例如顺磁性剂和铁磁性剂或超顺磁性剂。本文描述的突变蛋白可以偶联至放射性同位素,诸如α辐射源、β辐射源,或者γ辐射源。放射性同位素的实例包括碘(131i或者125i)、钇(90y)、镥(177lu)、锕(225ac)、镨,或者铋(212bi或者213bi)。结合蛋白可以偶联至生物蛋白、植物源或者细菌源的分子(或其衍生物),例如,美登素类(例如,美登木醇、其类似物或者dm1),以及紫杉烷(例如,紫杉酚或者多西紫杉醇),或者卡奇霉素。美登木醇类似物的实例包括具有经修饰的芳环(例如,c-19-脱氯、c-20-脱甲氧基、c-20-酰氧基)的那些和具有在其他位置的修饰(例如,c-9-ch、c-14-烷氧甲基、c-14-羟甲基或者c-14-酰氧基甲基、c-15-羟基/酰氧基、c-15-甲氧基、c-18-n-脱甲基、4,5-脱氧基)的那些。例如在u.s.6,333,410中描述了美登木醇和美登木醇类似物。美登木醇可以使用例如n-琥珀酰亚胺基3-(2-吡啶二硫代)丙酸酯(又名n-琥珀酰亚胺基4-(2-吡啶二硫代)戊酸酯或者spp)、4-琥珀酰亚胺基-羰氧基-a-(2-吡啶二硫代)-甲苯(smpt)、n-琥珀酰亚胺基3-(2-吡啶二硫代)丁酸酯(sdpb)、2-亚氨基硫烷盐酸盐或s-乙酰琥珀酸酐偶联。
以下实施例仅作为示例说明而提供,并且无意限制本发明的范围。本领域技术人员将容易地认识到可以改变或者调整许多非关键参数来得到基本上相似的结果。
实施例
实施例1:开发具有改变的结合亲和力的il-4突变蛋白
使用il-4/il-13系统测试以下构思:共有细胞因子受体的相对亲和力的操纵可以产生具有特征性反应性模式的细胞因子。采取两种方法来工程改造il-4以更高亲和力结合至γc(图1b)或者il-13rα1(图1c)。
最近已经解析完整的与配体结合的i型和ii型受体三元复合物的三维结构。il-13具有与il-4相比相对高的对il-13rα1亲和力(kd30nm对kd>1μm)。因此,从il-4和il-13在两种ii型三元复合物(il-4/il-4rα/il-13rα1和il-13/il-4rα/il-13rα1)中的结构来比对il-4和il-13的结构。
il-4和il-13的d螺旋在各个复合物中的比对非常接近,d螺旋形成与il-13rα1的大量接触。三个il-4d螺旋残基,r121、y124和s125,在il-4ii型受体三元复合物中形成与γc的接触,不同于il-13中的那些。因此交换这些残基以获得其il-13位置等效物(图1c),并且形成两种il-4变体,称为kf的保守双重突变体r121k、y124f,和三重突变体kfr,其中所有三个残基r121k、y124f和s125r都被交换。
为了提高il-4对γc的亲和力,应用使用亲和力成熟的酵母表面展示的组合库方法。il-4与酵母表面蛋白aga2p融合,其在那里被il-4受体胞外域(ecd)识别(图1d)。为了实现这种融合,将人il-4dna克隆到酵母展示载体pct302中,形成pct302_il-4载体。酿酒酵母(s.cerevisiae)菌株eby100用pct302_il-4载体转化并在sdcaa板上于30℃生长3天。il-4酵母的个体菌落在sdcaa液体培养基(ph4.5)中于30℃生长过夜,接着在sgcaa液体培养基(ph7.0)中于20℃诱导2天。
产生il-4rα、γc和il-13rα1的c末端的生物素化的胞外域,用作通过偶联至链霉亲和素来选择四聚物的染色和分选试剂。用生物素化的il-4rα(b-il-4rα)、四聚体化的生物素化γc(b-γc),或者在b-il-4rα存在下的b-γc对酵母进行染色。通过在冰上使2μm的b-γc与470nm的sav-pe(链霉亲和素-藻红素缀合物,invitrogen)孵育15分钟来形成最高浓度的γc四聚物。分析是在
展示在酵母上的il-4以高亲和力结合il-4rα(图1d,第二小图),但是在il-4rα不存在的情况下不结合至γc(图1d,第三小图)。在il-4rα的存在下,酵母上的il-4结合至γc胞外域四聚物,表明异二聚体受体复合物的协同组装(图1d,最右边的小图)。这些实验证明酵母上野生型il-4的位点1和位点2分别被il-4rα和γc顺序接合。
为了建立il-4的d螺旋库,分析i型受体三元复合物的晶体结构中的il-4/γc界面,d螺旋是细胞因子的主要的与γc相互作用的螺旋(图1b)。基于原子间的接触,建立关注的库,其中螺旋d面上的八个残基被随机化至简并密码子nnk(n=a、c、g或者t;k=g或者t),这使得能够利用20种氨基酸中的任一种(图1b)。使用组装pcr,建立了具有2×108个变体的酵母库,代表完全理论多样性(2.6×1010)的约1%。
使用11个重叠引物进行组装pcr,所述引物中的一个含有随机序列。pcr产物进一步使用引物5'ctagtggtggtggtggttctggtggtggtggttctggtggtggtggttctgctagccacaagtgcgatatcaccttac3'(seqidno:2)和5'cagatctcgagcaagtcttcttcggagataagcttttgttcgccaccagaggatcc3'(seqidno:3)扩增,所述引物还包含必要的载体序列同源性以在酵母中同源重组。如先前所描述的,插入dna与线性化的载体骨架和电感受态eby100组合,电穿孔,并拯救。该电穿孔产生2×108转化子的库。
使用磁激活细胞分选法(macs,miltenyi)进行选择。第一轮选择用酵母库的2×109个细胞进行,是转化子数量的约10倍范围。后续轮次的选择使用1×107个酵母细胞。
通过用il-4rα修饰酵母库以在酵母上建立il-4/il4rα位点2来进行选择,以及随后使用磁激活细胞分选法(macs)、通过五轮体外选择来富集γc-结合酵母(图1e)。最初轮次选择的酵母在il-4rα的存在下结合至四聚体γc。相继选择轮次(二到五轮)通过减少四聚体γc的浓度来进行,其中最终选择,即第六轮,使用1μm浓度的单体γc。高亲和力的γc四聚物是检测相对弱的γc结合和早期轮次选择必要的。后续轮次选择使用1×107个酵母细胞。在sdcaa液体培养基(ph4.5)中于30℃新鲜生长il-4库过夜,接着在sgcaa液体培养基(ph7.0)中于20℃诱导2天。用生物素化的il-4rα(b-il-4rα)、四聚体化的生物素化的γc(b-γc)或者在b-il-4rγ的存在下的b-γc,以及1xpbs和0.5%bsa的缓冲溶液在4c对酵母染色2小时。γc的四聚物是通过使2μm的b-γc与470nm的sav-pe(链霉亲和素-藻红素缀合物,invitrogen)在冰上孵育15分钟而形成的。使用γc的单体选择是通过顺序结合γc、2μg/ml的sa-pe和50μl的miltenyi抗pe微珠/1×108个酵母细胞来进行的。分析是在accuric6流式细胞仪上进行的。
il-4选择的变体的测序显示二个独特序列,‘rq’和‘rga’变体,其中之一rga是高度富集的(表1)。rq仅含有与野生型il-4的单一残基偏差r121q。有趣地是,此位置处的q在大部分γc-结合细胞因子中存在。另一种所选择的细胞因子rga含有r121q突变以及在螺旋d中的7个其他变化(rq和rga的序列请参见表1)。
实施例2:il-4突变蛋白的第二受体链结合特性
表征得到的il-4突变蛋白的结合特性。重组il-4和使用杆状病毒的变体kf、kfr、rq和rga与il-4rα形成复合物,以通过表面等离子体共振(spr)来测量其位点2对il-13rα1和γc的结合亲和力和动力学(见表1和图2)。
将人il-4变体(氨基酸1-129)、il-4rα胞外域(氨基酸1-202)、il-13rα1(氨基酸1-310)和γc(氨基酸34-232)与n末端gp67信号序列和c末端六组氨酸标签框内克隆到pacgp67a载体(bdbiosciences),并使用杆状病毒表达系统产生。杆状病毒储备是通过在于sf900ii培养基(invitrogen)中生长的斜纹夜蛾(spodopterafrugiperda)(sf9)细胞中转染和扩增而制备的,并且在于insect-xpresstm培养基(lonza)中生长的悬浮粉纹夜蛾(trichoplusiani(hifive))细胞中进行蛋白表达。蛋白经表达并且在48-60小时之后用镍琼脂糖(qiagen)从hifive上清液捕获,经浓缩并且通过superdextm200柱(gehealthcare)上尺寸排阻色谱纯化,在10mm的hepes(ph7.2)和150mm的nacl中平衡。在基于细胞的测定中使用的il-4变体表达为充分糖基化的。在spr测量中使用的高亲亲和力二元复合物(il-4rα与il-4变体)是共表达的并且充分糖基化的。为了生物素化受体的表达,将il-4rα、il-13rα1和γc克隆到pacgp67-a载体中,该载体具有c末端生物素受体肽(bap)lndifeaqkiewhe和六组氨酸标签。受体蛋白与bira连接酶共表达,具有过量的生物素(100μm)。对于下文讨论的结晶,il-4rga与γc共表达,其中n连接的糖基化位点asn53突变成gln。在镍纯化之后,结晶蛋白用羧肽酶-a处理过夜,接着进行尺寸排阻。蛋白浓缩到8-20mg/ml以结晶。
在biacoret100仪器上进行表面等离子共振实验。使用nanodrop2000分光光度计(thermoscientific)以280nm的uv光谱定量蛋白浓度。所有数据使用biacoret100评价软件2.0版本按1:1的langmuir结合模型分析。实验使用biacoresa传感器芯片(gehealthcare)。以低密度(100-200ru)捕获生物素化的受体并且以40μl/分钟进行动力学电泳。不相关的生物素化的蛋白被固定作为sa传感器芯片的参考表面,具有与实验表面匹配的ru。所有的测量使用在电泳缓冲液(1xhbs-p(gehealthcare),0.01%的bsa)中3倍连续稀释的il-4变体进行。用7mm甘氨酸(ph3.0)和250mm的nacl再生细胞因子il-4rq和il-4rga的γc表面。使用120s至190s的il-4变体缔合和20s至900s的解离来测定动力学数据。
表1.超级因子对γc和il-13rα1的结合亲和力
*u=无法测量地弱
野生型il-4/il-4rα对il-13rα1和γc的kd分别是4200nm和3300nm。kf/il-4rα以更大的亲和力结合至il-13rα1(kd=250nm)和γc(kd=330nm)。添加s125r突变体到kfr中导致对il-13rα1的亲和力440倍的改善(kd=9.6nm),但是对γc的亲和力减小(kd=6400nm)。因此,kfr赋予il-4在il-13rα1结合亲和力方面2.6对数改善,以及相应的对γc亲和力的2倍减小。所述亲和力改善主要是由于解离速率的3对数减小。在这方面,接枝是非常成功的,并导致对il-13rα1相对于γc的3对数选择性。附加地,还选择针对il-13rα1的il-4d螺旋库,并且这种选择令人惊讶地导致共有序列,其中il-4残基122、124和125分别被lys、phe和arg取代。这与基于结构重叠,通过接枝il-13残基到野生型il-4上而工程改造的kfr几乎相同(图1b)。
当用酵母展示选择的rq变体和rga变体与il4rα复合时,表现出明显更高的结合至γc的亲和力(表1,图2)。与il-4/il-4rα相比,rq/il-4rα显示出对γc的36倍更高的亲和力(kd=91nm),并且rga/il-4rα表现出3700倍更高的亲和力(kd=0.89nm)。rq和rga的亲和力增加主要是由于解离速率降低,而且由于7倍和6倍更快的结合速率。rq和rga超级因子两者都表现出明显减小的对il-13rα1的结合(分别为kd=29,000nm,以及kd=21,000nm)。在生理浓度时,这些变体将表现出可忽略的ii型受体结合。
因此,rga已获得约4对数结合亲和力,并且对i型受体复合物的选择性比对ii型受体的选择性高约6对数。总起来说,基于结构的和体外进化方法已经产生了具有更高亲和力和受体选择性的il-4变体以供功能测试。这些il-4突变蛋白被称为il-4‘超级因子’,具体称rga变体为“super-4”。
实施例3:il-4对γc的亲和力改善的结构基础
理想地,il-4突变蛋白(诸如,super-4)的亲和力升高是通过对野生型il-4/γc结合定向的最小干扰来实现的。为了确定super-4与第二链γc的停靠方式相对于野生型il-4是否受到干扰,进行结晶学分析。假设自由(例如,不结合至il-4rα)的super4对γc的位点2亲和力更高(kd为约300nm),有可能在il-4rα不存在的情况下使二元super-4/γc复合物结晶至
表2.数据收集和细化统计(分子置换)。
为了实现之,通过混合0.1μl的蛋白[在10mm的hepes-naoh(ph7.2)和150mm的nacl中20mg/ml]与等体积的50mm的hepes(ph7.2)、200mm的nacl和30%的peg-4000,使il-4rga/γc晶体在25℃以坐滴法生长。晶体在5-10天中生长到200×40×40μm的最大尺寸。使用含有25%乙二醇作为防冻剂的母液,在液氮中速冻晶体。在advancedlightsource以光束8-2收集
il-4rga二元晶体结构通过用程序phaser,分别使用il-4和γc的坐标(pdb代码3bpl),通过分子置换解析。在所有域定位之后,将野生型序列转换成il-4rga,并使用经phenix迭代轮次精制和经coot模型调整来细化结构。使用molprobity(超文本传输协议://molprobity.biochem.duke.edu)进行ramachandran分析。使用proteininterfaces、surfaces和assemblies(pisa)服务器(超文本传输协议://www.ebi.ac.uk/msd-srv/prot_int/pistart.html)来计算被遮盖的表面积值。il-4rga由2个二元复合物组成。所有的结构图和覆盖使用pymol作出。
二元super-4/γc复合物与三元i型信号传导复合物的重合表明在细胞因子-受体定向中没有大的干扰。
在super-4/γc界面中,super-4螺旋d残基117127的侧链密度是清晰的(图4);这些侧链以与il-4拓扑学类似的方式接合γc结合位点,其中γc热点残基y124占据中心位置(图3c)。看起来,super-4的增强的亲和力的潜在可能机制是用f替代s125(图3d,左小图),其现在插入γc的先前未被占据的大的疏水口袋中,促成附加
表3.配体-受体接触表
表3中的粗体代表从γc到super-4的可能的h-键(
实施例4:il-4突变蛋白能够激活细胞
测试所公开的il-4突变蛋白对各种细胞类型的影响。因为超级因子的第二链募集的平衡常数显著不同于il-4的平衡常数,测试超级因子在主要表达i型或ii型il-4受体或者大量的i型和ii型il-4受体两者的细胞类型中的作用。
在最初,测定il-4rα、γc和il-13rα1在若干不同的人细胞系中的rna和蛋白表达,所述细胞系包括ramosb细胞、hht细胞、u937单核细胞和a549上皮细胞。ramos细胞、u937细胞、a549细胞和hh细胞在含有10%fbs、青霉素/链霉素和l-谷氨酰胺(2mm)的rpmi中生长,并在37℃、5%co2维持。细胞经血清饥饿过夜并收集。用
(qiagen)从经饥饿的细胞分离rna。使用用于rt-pcr的
因此,基于mrna水平,ramos细胞具有相对少的ii型il-4受体。虽然ramos细胞具有大量的il-4rα,但是它们的i型受体量受到相对低表达的γc的限制。a549细胞具有大量的ii型受体和很少或没有i型受体。最后,u937细胞具有大量的i型和ii型受体链rna。
通过facs染色,ramos显示出高的il-4rα表达。hh和u937具有最高的γc表达,而ramos显示出该链相对低的表达。il-13rα1表达在a549细胞中是最高的(图6b)。il-4受体链的细胞表面表达是在封闭fc受体i、ii和iii之后进行的。cd23抗体、il-4rα抗体和γc抗体购自bd,并且针对il-13rα1的抗体购自r&d。
stat6y641磷酸化的测定提供测量il-4信号传导中的非常早期事件的定量工具,所述早期事件直接取决于负责转导细胞内信号的受体复合物的组装。stat6y641磷酸化需要激活jak激酶以及il-4rα的细胞内酪氨酸、尤其是y575、y603和y631的磷酸化。这些经磷酸化的酪氨酸充当stat6的sh2域的停靠位点,所述sh2域随后在y641被磷酸化,导致stat6二聚化和随后进入细胞核。
为了研究对il-4和其超级因子的反应,使用如上所述的细胞类型。测试il-4、super-4和kfr超级因子的刺激活性。当复合至il-4rα时,super-4具有结合至γc的最高kd,并且kfr具有结合至il-13rα1的最高kd。
使用ramos细胞研究由i型受体复合物主导的il-4反应(图7和图8)。用100pg/ml(约7pm)的il-4或super-4或kfr超级因子刺激ramos细胞各种时间,发现在刺激20分钟时有最大的诱导,并在30分钟时持续,随后在60分钟时略有减少(图7a)。用il-4、super-4和kfr刺激stat6磷酸化的时程是类似的,但是在测量的所有时间点super-4比il-4或者kfr诱导明显更多的磷酸化;在20分钟的刺激之后,用super-4诱导的stat6磷酸化的平均mfi是19.6,用il-4诱导的是7.7,并且用kfr诱导的是5.4。
在刺激之前,细胞在含有2%的fbs的生长培养基中培养过夜(“经饥饿”)。在用冰冷的甲醇(90%)透化之后进行细胞内pstat6染色。抗pstat6抗体购自bdbiosciences。通过从未经刺激的样品的平均荧光强度(mfi)中减去经刺激的样品的mfi来计算stat6磷酸化的诱导。
也测定了il-4、super-4、kfr以及rq和kf的浓度/反应曲线。用从30pg/ml(2pm)至100,000pg/ml(7.4nm)浓度范围的il-4或超级因子刺激经饥饿的ramos细胞15分钟,尽管在大部分实验中该范围限于30至1,000pg/ml。il-4和super-4两者引起类似量的stat6磷酸化,但是super-4需要1,000pg/ml来达到高峰水平,而il-4需要在10,000和30,000pg/ml之间来达到高峰水平(图7b)。
进行ramos细胞对在30至1000pg/ml的浓度范围中的il-4和四种超级因子的反应的分析(图7c和图8)。在所有测试浓度下,super-4优于il-4;为了实现与用super-4实现的相同程度的stat6磷酸化,需要约3倍更多的il-4。kf、rq和il4通常彼此类似;kfr引起的刺激比il-4少,是稍微小于3倍的活性。因此,super-4比kfr活性大约10倍。因此,定性地,形成具有最高的γc亲和力的il-4/il-4rα复合物的超级因子il-4是ramos细胞中最有活性的,并且具有最低的亲和力的超级因子是活性最差的。然而,与复合至il-4rα时super-4和il-4对γc的溶液平衡常数的约3700倍差异比较,super-4对比il4的相对优势是相对适中的。
对a549细胞进行类似的分析,如上所述a549细胞主要利用il-13rα1作为其第二链。对于这些细胞,kfr和kf的刺激性比il-4大3-10倍;super-4和rq与il-4难以区别(图7d)。虽然复合至il-4rα的kfr在溶液中具有比il-4大约400倍的il-13rα1亲和力,但kf相对于il-4仅具有约10倍优势。依然是定性协议,因为越高亲和力的超级因子引起越高的反应,但是其优势程度不与其溶液亲和力差异直接相关。
实施例5:受体组装的模型化
super-4是stat6磷酸化的约3-10倍更有效激活剂,而对于γc的三维平衡比il-4高约3700的观念推动第二链的表达如何指挥信号诱导受体形成的分析。已经计算出在巨噬细胞中,约40个受体将给出几乎最大的stat6激活。因此,super-4在细胞水平的令人难惊讶的适中优势可能不得不伴随相对低数量的形成的受体复合物,和/或仅当第二链的数量较低时(例如配体将竞争可用的第二链)可以观察到。
为了解决这个问题,使用一种计算方法。应用
因为ramos细胞上il-4rα链的数量是约1500条,所以受体的组装固定为这个数量。当γc数量设置成4500时,计算出将第二链k值从0.01升高到1.0的相对适中的影响,但是当γc链数量设置成500时,第二链的k值的升高对所组装的受体链的数量有强烈影响(图9a,比较左小图和右小图)。
因此,用100pg/ml的细胞因子浓度,当第二链的数量是4500时k=1μm2相较于k=0.01μm2组装的复合物的比率是6.7,而当γc数量设置成500时,这个比率是34.5。当细胞因子浓度十倍更高,即1000pg/ml时,4500γc分子的k=1μm2/k=0.01μm2比率是6.8,而当γc数量是500时,该比率是25.6。因此当第二链的数量相对低时,升高第二链的k更有用。这有力说明表达低水平的γc或者il-13rα1的细胞将最大地受益于对第二链增强的亲和力,但表达大量γc或il-13rα1的细胞应在高亲和力超级因子和il-4之间差异较小。
另一个问题将是升高超级因子/il-4rα复合物对每个细胞具有非常少的γc分子的细胞上的复合物组装的影响。计算当二维k保持不变并且γc分子的数量从4500变化至167时受体复合物的组装。在用于大部分体外刺激测定的细胞因子浓度下,具有0.01μm2的对γc的二维k(这是il-4的推荐二维k)的il4/il-4rα复合物在具有仅167个γc分子的细胞上组装非常少的受体(在100pg/ml时<1,以及在1000pg/ml时为3.6)(图9b,右小图)。因此,这种细胞应该是对这些细胞因子浓度的il-4基本上无反应的。相反,具有1μm的对γc的二维k的超级因子将在100pg/ml时组装33个复合物,并且在1000pg/ml时组装109个复合物(图9b,左小图)。
为了评估多少组装的复合物对ramos细胞中的最大刺激来说是理论上足够的,确定il-4和rga是否对于stat6磷酸化刺激类似的坪值。实际上,il-4和rga确实对于stat6磷酸化刺激类似的坪值;在ramos细胞中获得10,000pg/ml至30,000pg/ml的il-4的坪值(图7b)。使用中间数量的γc链(1500),计算10,000pg/ml和30,000pg/ml时对于第二链具有低亲和力(0.01μm)的配体的组装复合物的数量。组装链的值在65和71之间(图9c),表示组装更多的链可能不产生任何更大的信号传导。如果选择低γc数量(500),那么用于最大刺激的组装链数量已经相应地减少。然而,带来最大信号的约70个组装受体的值是保守估计。
因此,通过在100pg/ml时使用高亲和力的超级因子获得的33个组装复合物对于表达167个γc分子的细胞的明显反应应该是足够的,而在1000pg/ml时组装的109个复合物应产生最大反应。这表示着在这些低浓度时不会对il-4响应的细胞将会对高亲和力的超级因子产生响应,提高了通过对第二链具有极高亲和力的工程化il-4超级因子可能实现意料外反应(和毒性)的可能性。
实施例6:改变第二链的表达
因为可以预料到γc表达的升高会减少super-4相对于il-4的优势,所以在具有比ramos细胞高得多的γc表达的hh细胞系中研究对il-4和super-4的敏感性(图6a和图6b)。在10pg/ml到10,000pg/ml的浓度范围,super-4在hh细胞中诱导stat6磷酸化方面并不优于il-4(图10a)。
第二链数量对针对il-4和超级因子的相对反应的贡献的替代试验将是减少γc的可及性。为了这个目的,将改变浓度的抗γc抗体添加到刺激培养中。用100pg/mlsuper-4刺激ramos细胞导致相对于100pg/ml的il-4相当大的优势,如通过stat6激活判断的(图7c)。在存在或者不存在抗γc的情况下,用100pg/ml的il-4或者super-4/rq/kf/kfr超级因子刺激这些细胞,用流式细胞计数法测量stat6磷酸化,并计算由抗γc引起的stat6磷酸化的百分比减少。用il-4诱导的stat6磷酸化被50μg/ml的抗γc减少58%,而对于super-4,减少仅12%(图10b;图18)。对于rq和kf超级因子,抑制是约40%,并且对于kfr,抑制类似于il-4的抑制。这些结果与il-4和超级因子结合到γc的相对溶液kd一致(super-4>rq、kf>il-4=kfr,表1),并且支持以下概念:对第二链的亲和力升高导致当第二链表达减少时更大的刺激区别。
对u937单核细胞系进行类似的γc封闭实验。对于γc封闭实验来说,在37℃用封闭抗体孵育经过夜饥饿的ramos或者u937细胞1小时。
u937细胞系表达i型和ii型il-4受体两者,并且可以预计γc的封闭会减少il-4反应,而对kfr超级因子活性应该很少影响,因为kfr超级因子主要利用ii型受体。实际上,封闭u937细胞中的γc导致响应于il-4的stat6磷酸化减少44%,但是响应于kfr超级因子的stat6磷酸化减少仅7%(图10c)。
实施例7:人原代细胞对il-4和超级因子的反应
为了证实对各种超级因子的反应取决于这些超级因子主要使用哪种第二链,测试人外周血液白细胞(pbl)的stat6磷酸化反应(图11)。pbl未受刺激,或者使用il-4或者各种超级因子刺激15分钟。用流式细胞计数法测量stat6y641磷酸化以测试各种外周血液细胞类型的反应性。淋巴细胞对super-4的反应在较低配体浓度时显著更好。
100pg/ml时super-4相对于il-4对stat6磷酸化的优势比较的p值是0.0003,在1ng/ml时p值是0.0104,以及在10ng/ml时p值不是统计学上显著的(0.1493)。在比较中,用于对比il-4相对于kfr超级因子在淋巴细胞stat6磷酸化中的优势的相应p值是0.0005、0.0004和0.5364。这些发现与主要使用γc作为人淋巴细胞中的第二受体链一致(图11a)。单核细胞和中性粒细胞在其对il-4、super-4和kfr的反应方面显示很少差异。
为了确定原代细胞上i型和ii型il-4受体的表达程度,用流式细胞计数法测量来自6个健康供体的b细胞、t细胞、单核细胞和中性粒细胞中的il-4rα、γc和il-13rα1。il-4rα表达在b细胞上是最高的,而单核细胞和中性粒细胞在il-4rα表达方面显示很少差异,并且t细胞具有最小的il-4rα表达(图11b、图11c和图11d)。对于γc,单核细胞和cd4t细胞之间存在相对很少的表达差异。b细胞具有略少的γc,并且cd8t细胞具有最低水平。如所预计的,il-13rα1表达在单核细胞上是最高的,而b细胞和t细胞显示非常低的该链的表达。pbl不经刺激,或者使用il-4或者各种超级因子刺激15分钟;用流式细胞计数法测量stat6y641磷酸化。在cd4和cd8t细胞中,super-4比il-4诱导更强的stat6磷酸化,以及比kfr强很多的磷酸化(图11e)。单核细胞在其对il-4、super-4和kfr的反应方面显示很少差异,跟随其γc和il-13rα1两者的表达。
在cd4和cd8t细胞中,super-4比il-4诱导更强的stat6磷酸化,以及比kfr明显更强的磷酸化(图12)。单核细胞在其对il-4、super-4和kfr的反应方面显示很少差异。一定程度上令人惊讶地,b细胞对kfr的响应较好,尽管其相对低水平的可检测的il-13rα1。
th9分化测定
为了研究il-4和超级因子的功能特异性和免疫调节能力,进行涉及cd4t细胞和单核细胞的一系列实验。tgf-β和il-4的组合促进天然人cd4t细胞分化成th9细胞。为了测试super-4是否比野生型il-4更有力地诱导th9分化,分离出天然的cd4+cd45ra+cd45ro-cd25-t细胞。
在用ficoll-paquetmplus(gehealthcare)进行密度梯度离心之前,使用rosetteseptm人cd4+t细胞富集(stemcelltechnologies)从自健康供体(stanfordbloodcenter)获得的白细胞层制备富集的cd4+t细胞。用天然cd4+t细胞分离试剂盒ii(miltenyibiotec)磁性分选天然cd4+cd45ra+cd45ro-cd25-t细胞。
随后在37℃,在48孔平底板(falcon)中补充有10%ab型人血清(lonza)、100单位/ml青霉素/链霉素、l-谷氨酰胺(invitrogen)和50μm的β–巯基乙醇(sigma-aldrich)的x-vivotm15培养基(lonza)中培养细胞。在存在5ng/ml的tgf-β(ebioscience)和指示浓度的il-4、super-4或kfr的情况下,按照1:1的珠与细胞比率,用抗cd3/cd28包被珠(invitrogen)以2.5x105个细胞/ml的浓度培养细胞。
在培养4天后,将珠磁性地去除,并且在存在布雷菲德菌素a(ebioscience)的情况下,用25ng/ml的pma和750ng/ml的离子霉素(invitrogen)再刺激细胞4小时。随后用
用10μg/ml或者100μg/ml的super-4引发导致与用相同浓度的il-4或kfr进行引发相比,在用pma和离子霉素连续刺激后细胞以显著更高的百分比产生il-9(图13a)。
树突状细胞分化测定
il-4与gm-csf组合诱导从人单核细胞体外分化成树突状细胞(dc)。这个过程中i型或ii型受体的相对作用尚还有待阐明。假设super-4优先结合γc,并且kfr倾向于il-13rα1连接,阐明了在人dc分化期间对γc和il-13rα1途径的需要。
通过使用
在第6天用5mm的edta处理细胞,并且随后用dapi(invitrogen)、荧光标记的同位型对照mab、或者抗cd14、cd86、cd209和hla-dr的mab(bdbiosciences)对细胞染色。用bdlsrii流式细胞仪,通过流细胞计数法评估树突状细胞的分化,并且用flowjo(treestar)检测和分析细胞表面标志物表达的平均荧光强度。
虽然il-4和kfr引发单核细胞分化成dc,如通过cd209、cd86和hla-dr的上调所评估的(图13b和图22),但是super-4未能如此,表明dc分化主要通过ii型il-4受体复合物的信号传导驱动,而ii型il-4受体复合物很难被super-4接合。单核细胞成熟化为dc还与cd14的下调有关;在cd14的下调方面,super-4的效果比kfr或者il-4差(图13b)。另外,用于区分不同树突状细胞子集的其他标志物的分析显示,在有或没有super-4下用gm-csf诱导的细胞是表型相同的(图23),意味着super-4诱导的细胞未完全分化,并非分化成独特的树突状细胞子集。
为了确认i型和ii型il-4受体复合物在dc分化中的相对作用,本文显示抗il-4rα抗体(i型和ii型受体特异性的)响应于il-4和kfr而减少cd86和cd209的表达,而抗γc(i型受体特异性的)未能如此(图14)。如前所述,super-4引起这些标志物的非常适中的诱导。super-4诱导的cd14下调被抗il-4rα而非抗γc部分抑制。有趣的是,当使用抗γc的中和抗体时,il-4和kfr仍然诱导与对照条件相同水平的dc成熟(图14),证实dc分化通过ii型il-4受体复合物发生。
实施例8:在单核细胞中通过il-4和超级因子诱导的信号激活特征
确定了super-4诱导dc分化的能力,因为il-4和两个超级因子在单核细胞中将stat6激活到相同程度(图11(e)、图12和图19)。用两种剂量的细胞因子处理经纯化的单核细胞,一种剂量对应于p-stat6ec50值(30pm)(图15),而另一剂量对应于饱和(50nm)(图20)。在指定时间分析stat6和irs1磷酸化的水平,以及γc和il-13rα1受体的下调。在低剂量时,当与il-4比较时,super-4和kfr表现出对stat6和irs1的延迟激活。γc或者il-13rα1均未观察到显著的内化(图15)。在高剂量时,三种细胞因子诱导相同动力学特征的stat6和irs1激活。在刺激的晚期,kfr表现出更强的il-13rα1内化(图20)。总的来说,这些结果显示表面受体内化和信号传导激活之间缺乏相关性。此外,仅信号传导激活的延迟动力学无法解释super-4不能诱导dc分化,表明dc分化需要ii型受体特异性的信号传导。
实施例9:在单核细胞中通过il-4和超级因子诱导的基因表达谱
为了获得在分化dc中il-4和超级因子诱导的遗传程序冗余程度的定性理解,进行对用gm-csf同时处理的单核细胞中响应于wtil-4和两种超级因子的基因表达的全基因组分析。cd14+单核细胞是从5个健康的供体分离的,并且仅用50ng/ml的gm-csf,或者用与20ng/ml的il-4、super-4或kfr一起的50ng/ml的gm-csf刺激6小时。细胞在pbs中洗涤并且在1ml的trizol试剂(sigma)中溶解。使用组合的trizol/rneasymicro(qiagen)方案分离总rna。用2100bioanalyzer(agilent)确认rna品质。使用two-colorlowinputquickamplabeling试剂盒(agilent),用cy3标记的单一处理样品(仅用gm-csf)和用cy5标记的双处理样品,产生和标记cdna。对于每一供体,将来自每一cy5标记的双处理(含有il-4、super-4或者kfr的gm-csf)样品与相应的cy3标记的用gm-csf单处理样品在8x60ksureprintg3基因表达微阵列(agilent)上共杂交并按照制造商的说明书处理。使用featureextraction软件(agilent)标准化原始数据。使用配对t检验分析来统计分析双处理和单处理之间的变化,以及变体之间的变化。在genespringgx11.5和excel上进行基因表达分析。仅以小于6个阵列检测实体,排除质量控制探针和用于长基因间非编码rna的探针。保留与单独gm-csf相比至少一个双处理组(cy5对cy3)中有显著变化(配对t检验)的实体。用配对t检验确定两种给定的细胞因子之间的倍数变化统计分析。微阵列数据已经提交到ncbi的geneexpressionomnibus(geo)数据库(geo系列登录号:gse40200)。
如散点图的相关性所示,三种细胞因子诱导大多数基因至相同的程度(图16a)。然而,有趣的是,在il-4和两种超级因子之间还可以观测到基因表达特异性的小口袋。大量基因仅被所使用的细胞因子中的一种或两种显著地诱导,如图16b中的维恩图所显示的。图16c中的热图显示细胞因子选择性基因的代表性设置,其中观测到il-4和两种超级因子诱导的表达模式之间的明显差异。表4中提供了在单核细胞中由超级因子和il-4差异调节的基因清单。与super-4相比,il-4和kfr明显诱导dc特异性基因(诸如,tpa1、hla-dpa和cish)至更高水平,与来自ii型il-4受体的特异性信号一致,ii型il-4受体可以偏移il-4诱导的树突状细胞分化过程。
表4.超级因子选择性基因的诱导倍数(平均值,数据的前三列)和p值(配对t检验,数据的后三列)。
实施例10:通过il-4和两种超级因子诱导的细胞因子分泌特征
为了进一步评估工程改造的细胞因子诱导的dc的功能性,通过对在过去的24小时有或没有lps刺激下培养8天的细胞上清液进行luminex测定来比较细胞因子、趋化因子和生长因子的分泌模式。
为了表征luminex细胞因子,在第8天收集培养上清液(有或者没有用2μg/ml的lps刺激24小时)。人51plex试剂盒购自affymetrix并且根据制造商的推荐按如下所述调整而使用。简要地说,将样品在96孔过滤底板上与抗体连接的聚苯乙烯珠混合,并且在室温下孵育2小时,随后在4℃过夜孵育。对板进行真空过滤,并且用洗涤缓冲液洗涤两次,然后与生物素化的检测抗体在室温下孵育2小时。然后过滤样品并如上所述地洗涤两次,并重悬于链霉亲和素-pe。在室温下孵育40分钟后,进行两次额外的真空洗涤,并将样品重悬于读数缓冲液。使用luminex200仪器以每细胞因子每样品100珠的下限读板。针对从仅gm-csf培养的未经刺激的细胞获得的值标准化mfi。
在51个分析物中,20个显示处理间(超级因子和lps)无表达差异,并且19个被lps上调(最显著的是il-6、ccl3、ccl5和cxcl1),在il-4和超级因子之间没有差异(图17)。其余12个产物的表达区分了仅gm-csf或者gm-csf+super-4诱导的细胞与gm-csf+il-4或kfr诱导的dc(图17和图21)。前两个子集是非常相似的,并且相较于后两个子集(也是非常相似的)产生更多的g-csf、hgf、il-1α、il-1β、il-10、il-12p40、lif、tnfα和更少的mcp3、mip1β、pdgf、tgfα。大部分差异是在lps刺激之后观察到,但是也有一些存在于未激活的细胞中。总起来说,这些数据说明super-4对单核细胞的作用没有超过单独的gm-csf,而il-4或者kfr的添加导致表型和功能上不同的dc。因此,经工程改造的细胞因子似乎具有新且不同的功能活性。
- 还没有人留言评论。精彩留言会获得点赞!