水稻叶绿体蛋白OsFBN7及其应用的制作方法
本发明属于分子生物学领域,具体涉及水稻叶绿体蛋白osfbn7及其应用。
背景技术:
脂质不仅是生物体内高效的能量贮存物质,也是生物膜的主要成分和重要的信号分子,广泛参与各种生命活动过程,是植物生长发育所必需的基本成分。脂肪酸是糖脂、磷脂及储存性脂质合成过程中必不可少的前体物,其合成受阻将对植物生长发育造成严重影响。水稻是世界上重要的粮食作物之一,当水稻细胞的脂质含量不够或其合成受阻时,将对水稻的生长发育产生直接影响,可能导致减产和品质下降。
种子中的脂肪酸稻谷除去稻壳后即为糙米。糙米由米糠层、胚及胚乳组成。米糠层约占糙米总重的10%。米糠层由果皮、种皮、外胚乳及糊粉层组成。除富含淀粉和蛋白质外,糙米中还含有2-5%的脂类。糙米中油脂主要分布在胚和米糠层的糊粉层中。脂类的主要成分为:甘油三脂(tag)占76.4-80.5%,游离脂肪酸(ffas)占7.2-9.8%,磷脂(pls)占3.5-3.6%。游离脂肪酸的主要成分为:棕榈酸(c16:0)占15-20%、油酸(c18:1)占36-48%,亚油酸(c18:2)占30-38%。它们可以降低胆固醇含量、治疗脑震荡、高血脂和心脑血管疾病。米糠油是国内外公认的营养健康油(birlaetal,2017;yinetal,2014;qiuetal,2008)。在我国长江中下游和东北稻区等水稻主产区拥有资源优势,是我国米糠油加工的主要区域。但是,我国米糠油年产量不足12万吨,利用率不到10%,开发米糠油产品发展前景和潜力很大。因此,开发高油酸水稻品种和生产技术,发展营养健康型稻米对建设“健康中国2030规划纲要”具有重要的社会效益和经济效益。
叶绿体蛋白fbn叶绿体是植物细胞特有的细胞器。它是由叶绿体两层被膜、类囊体和基质组成的。类囊体膜和叶绿体内被膜主要含有单半乳糖-二酰甘油(mgdg)、双半乳糖-二酰甘油(dgdg)和硫代异鼠李糖二酰甘油(sqdg)以及少量磷脂。mgdg和dgdg分别占叶绿体脂类的52%和26%(kalischetal,2016)。
1994年,首次从甜椒果皮纤维状有色体原纤丝中分离出与类胡萝卜素结合的32kda纤丝状蛋白,被称为纤维蛋白(叶绿体,fbn)(deruere,1994)。后来,在马铃薯叶片、豌豆叶片、拟南芥叶片、黄瓜花冠和油菜花药的质体小球中分离了fbn同源蛋白。最终,把这些来源于质体小球的蛋白总称为纤维蛋白(fbn)。在拟南芥的14个atfbns中,fbn1a/1b,fbn2,fbn4,fbn7a/7b和fbn8是质体小球的核心蛋白。fbn1a与fbn1b互作,fbn1a/1b均能与淀粉合酶ss4互作,影响叶绿体中淀粉的生物合成(gámez-arjonaetal,2014a;gámez-arjonaetal,2014b)。fbn4影响叶绿体中质体小球的亲脂性、叶绿体发育和耐光氧化能力(singhetal,2010)。在水稻的11个osfbns中,fbn5与水稻的耐光氧化抑制有关(otsuboetal,2018;kimetal,2015);fbn1基因超表达增加叶绿体中质体小球的数量,负调控水稻开花期的耐热性(lietal,2019)。但是,到目前为止,还没有关于osfbn7蛋白功能的报道。
脂肪酸的从头生物合成目前,水稻叶绿体中脂肪酸的从头合成途径还未深入研究(kimetal,2015;quetal,2008)。一般认为,植物叶绿体中脂肪酸的从头合成、延长和去饱和过程主要是在叶绿体中进行的。其基本过程大致如下:丙酮酸在位于叶绿体被膜上的丙酮酸脱氢酶系作用下产生乙酰coa。在叶绿体基质中,乙酰coa羧化酶(accase)催化产生丙二酸单酰coa,在acp-丙二酸单酰基转移酶(mcmt)催化下产生丙二酸单酰-acp。然后,乙酰-coa和丙二酸单酰-acp通过缩合-还原-脱水-再还原循环,催化该循环的4个酶类依次为kasiii(β-酮脂酰-acp合酶iii)、kar(β-酮脂酰-acp还原酶)、had(β-羟脂酰-acp脱水酶)和ear(β-烯脂酰-acp还原酶),合成丁酰-acp。类似地,通过6次循环,在kasi(β-酮脂酰-acp合酶i)、kar、had和ear的作用下,合成c16:0-acp。最后,在kasii(β-酮脂酰-acp合酶ii)、kar、had和ear作用下,合成c18:0-acp。其中,kas(β-ketoacyl-acpsynthase)是决定脂肪酸碳链延长的最主要关键酶。特别是,atkasi和oskasi影响叶绿体的发育、种子的成熟和脂肪酸积累过程(wu&xue,2010;dingetal,2015)。其次,在酰基acp硫酯酶(acyl-acpthioesterase,fat)作用下合成游离的c16和c18脂肪酸,终止脂肪酸碳链的延长反应。水稻含有25个fat基因,fata和fatb最重要。其中,fata特异催化油脂酰acp产生油酸(c18:1),fatb催化软脂酰acp和硬脂酸acp分别产生软脂酸和硬脂酸(salasetal,2002)。脂肪酸去饱和酶有两大类。①类:脂酰acp去饱和酶。水稻有7个硬脂酰acp去饱和酶(sad)基因,均编码叶绿体蛋白。sad2和sad5最为重要。②类:游离脂肪酸去饱和酶(fad)。水稻含有20个基因:4个osfad2亚族基因,2个osfad3亚族基因,1个osfad6,1个osfad7,1个osfad8,9个osfab2亚族基因,1个osdes1和1个ossld1。其中,osfad2、osfad3存在于内质网中,osfad6、osfad7、osfad8存在于叶绿体中(eetal,2019)。利用rnai干涉和crispr-cas9基因编辑技术获得osfad2-1基因敲除表达突变体,其成熟种子中油滴变大,油酸(c18:1)含量显著增加,亚油酸(c18:2)含量降低(abeetal,2018;tiwarietal,2016;zaplinetal,2013)。
mgdg和dgdg的生物合成在磷酸-3-甘油-酰基转移酶(gpat)催化作用下,c16:0-acp或c18:1-acp与甘油-3-磷酸(g-3-p)反应合成溶血磷脂酸(lpa)。在lpa酰基转移酶(lpaat)作用下进一步合成磷脂酸(pa)。然后,在磷脂酸磷酸酶(pap)作用下合成二酰甘油(dag)。在叶绿体中,mgdg合酶(mgd)和dgdg合酶(dgd)均定位于叶绿体的双层被膜中。mgd催化dag和鸟苷二磷酸-半乳糖(udp-gal)反应合成mgdg,dgd催化mgdg和udp-gal反应合成dgdg。然后,这些糖脂(mgdg、dgdg)主要用于类囊体膜、叶绿体内被膜和质体小球的合成与组装。
内质网中油体(tag)的生物合成一般认为,在内质网(er)中存在油体(tag)的合成-装配-积累途径(
在水稻种子成熟过程中,水稻叶绿体中从头合成的fas转变为脂酰coa转运至er中。在er中,与甘油-3-磷酸(g-3-p)反应生成tag。tag是胚和糊粉层油体中的主要脂类,占总脂类含量的76.4-80.5%。在胚特异基因(reg-2)启动子驱动下超表达2个大豆oleosin基因,分别增加水稻种子中油体含量36.93%和46.06%,但不改变tag中脂肪酸的组成(liuetal,2012)。前人研究表明,oskasi基因的ems(甲基磺酸乙酯)化学诱变水稻突变体生长受抑制,结实率下降,种子中脂肪酸总含量下降,油酸(c18:1)含量明显下降(dingetal,2015)。但是,到目前为止,还有没有关于osfbn7通过正调控oskasib活性提高水稻叶绿体和成熟种子中脂肪酸含量的报道。
技术实现要素:
本发明所要解决的技术问题是提供一种调控水稻叶绿体中脂肪酸从头合成途径的水稻叶绿体蛋白osfbn7的编码基因,将其转入目的植物中,能够提高其叶绿体与成熟种子中脂类和脂肪酸含量,达到培育种子中高油脂含量目的植物的目的。
为解决上述技术问题,本发明采用的技术方案如下:
一种水稻叶绿体蛋白osfbn7,其氨基酸序列如seqidno.1所示,或为如seqidno.1所示的氨基酸序列经替换、缺失或添加一个或几个氨基酸形成的具有同等功能的氨基酸序列,或为与如seqidno.1所示的氨基酸序列具有至少90%同源性且具有相同功能蛋白的氨基酸序列。
一种水稻叶绿体蛋白osfbn7的编码基因的转基因细胞系。
一种水稻叶绿体蛋白osfbn7的编码基因的重组菌。
一种水稻叶绿体蛋白osfbn7的编码基因的重组载体。
一种水稻叶绿体蛋白osfbn7或其编码基因在提高植物叶绿体脂肪酸含量中的应用。
一种水稻叶绿体蛋白osfbn7或其编码基因在培育高油脂植物品种中的应用。
一种提高目的植物油脂质含量的方法,将所述的水稻叶绿体蛋白osfbn7的编码基因转入目的植物中超表达,得到油脂含量高于所述目的植物的转基因植物。
一种水稻叶绿体蛋白osfbn7或其编码基因在调控水稻叶绿体脂肪酸从头合成途径中的应用。
与现有技术相比,本发明的优点在于:
本发明发现了osfbn7基因具有促进叶绿体中脂质合成的功能。通过实验证明,将osfbn7基因在水稻中超表达,所获得的转基因植株的种子相较于野生型表现出显著的高油脂表型。osfbn7蛋白及其编码基因可在实践中用于高油脂植物品种的培育和鉴定,为从源头上调控植物叶绿体中脂肪酸的从头合成、提高植物叶绿体脂肪酸及培育高油脂植物新品种提供了新基因资源和新方法。
附图说明
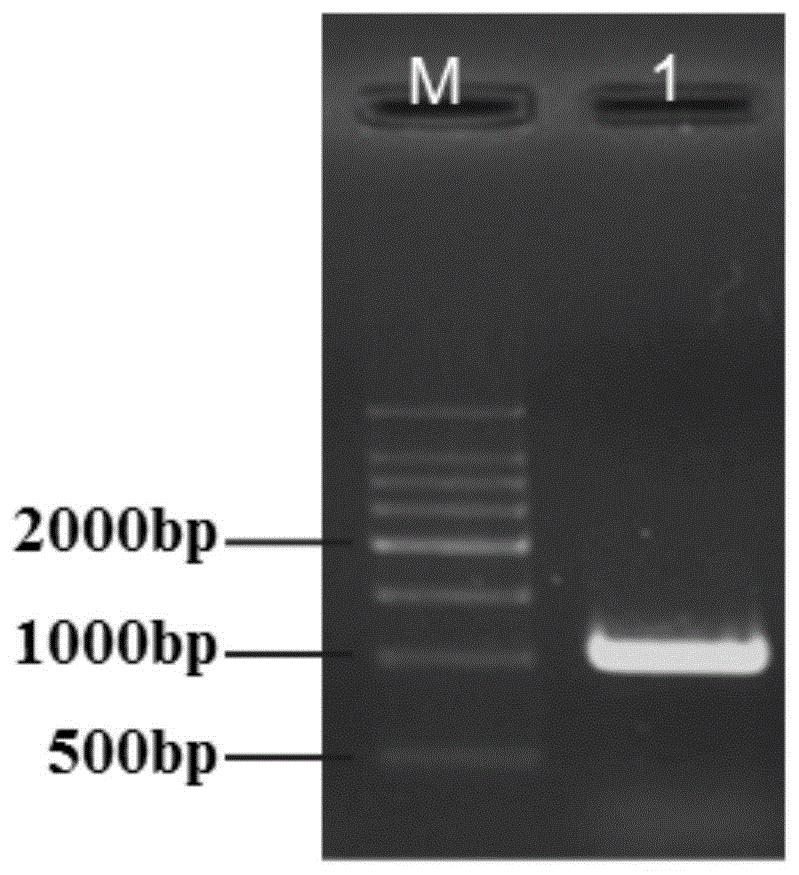
图1为本发明实施例提供的osfbn7基因pcr扩增产物;泳道1为产物条带(1023bp)。
图2为本发明实施例提供的osfbn7-pmd18-t载体双酶切鉴定图;泳道分别为marker条带,1,2为2个阳性克隆质粒双酶切后的载体和目标片段(1023bp)。
图3为本发明实施例提供的osfbn7-ptck303菌落pcr鉴定图;泳道1-5为5个阳性单菌落的pcr检测目标片段的克隆成功,因为均含有1023bp的目标片段。
图4为本发明实施例提供的osfbn7-ptck303载体酶切鉴定图;泳道m为marker;泳道1为酶切条带,含有目标片段(1023bp)。
图5为本发明实施例提供的超表达株系osfbn7中的osfbn7基因表达检测结果图;其中,wt为中花11野生型植株,oe-osfbn7为osfbn7基因超表达转基因株系(#1、#12和#17)。
图6为本发明实施例提供的osfbn7超表达转基因株系(图中oe-osfbn7,#17)和野生型植株(图中为wt-zh11)叶绿体中脂类含量的检测结果;脂类物质包括pc、pe、pg、pi、ps、pa、mgdg、dgdg、dag和tag。
图7为本发明实施例提供的osfbn7超表达转基因株系(图中oe-osfbn7,#17)和野生型植株(图中为wt-zh11)成熟种子中甘油三脂(tag)脂肪酸含量的检测结果;甘油三脂的脂肪酸链长包括c50:0、c50:2、c52:3、c52:4、c54:3,c54:4,和c54:5。
图8为本发明实施例提供的osfbn7超表达转基因株系(图中oe-osfbn7,#17)和野生型植株(图中为wt-zh11)成熟种子中游离脂肪酸含量的检测结果;游离脂肪酸包括c16:0、c16:1、c16:2、c18:0、c18:1、c18:2、c18:3、c20:0、c22:0和c24:0。
图9为本发明实施例提供的osfbn7在叶绿体中与脂肪酸从头合成途径关键酶kasib互作的酵母双杂交(y2h)鉴定的结果图。
图10为本发明实施例提供的osfbn7超表达转基因株系(图中oe-osfbn7,#1,#12,#17)和野生型植株(图中为wt-zh11)开花期剑叶中kasi酶活性elisa检测结果图。
具体实施方式
为了使本发明的目的、技术方案及优点更加清楚明白,以下结合附图及实施例,对本发明进行进一步详细说明。应当理解,此处所描述的具体实施例仅仅用以解释本发明,并不用于限定本发明。
下述实施例中所使用的实验方法如无特殊说明,均为常规方法。
下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
以下实施例中ptck303质粒可市购;pmd18-t为常用的克隆载体,可市购;水稻品种为中花11(野生型);水稻遗传转化的农杆菌菌株eha105。
以下实施例中的主要试剂如下:各种限制性内切酶、taq酶、t4连接酶、kod等购自于takara(大连)、neb、toyobo等生物公司;dntps购自于genestar公司;质粒小提试剂盒和琼脂糖凝胶回收试剂盒购自于上海捷瑞生物工程公司;ms培养基,琼脂粉,琼脂糖,氨苄青霉素(amp)、卡那霉素(kan)、利福平(rif)等抗生素及lbmedium等均为进口或国产分析纯试剂。
以下实施例中所使用的引物由天一辉远公司合成,并进行相关测序。
水稻基因osfbn7转基因株系的构建与筛选
osfbn7基因超表达株系的构建与筛选的具体方法如下:
1、从水稻中花11品种中克隆编码植物叶绿体叶绿体蛋白的基因osfbn7,连接到具有ubi启动子的超表达载体ptck303上。
根据编码区序列分析(osfbn7基因的序列如seqidno.2所示),设计引物扩增osfbn7基因的编码区。提取水稻的总rna,经反转录获得cdna,以水稻cdna为模板,利用上游引物f1和下游引物r1扩增osfbn7基因,osfbn7基因的扩增引物如下:
f1:5’-ggatccgcaactcatctcggcacca-3’(如seqidno.3所示,下划线为bamhi酶切位点);
r1:5’-ggtaccaagcgtgaaaagtggctgaac-3’(如seqidno.4所示,下划线为kpni酶切位点)。
将扩增得到的osfbn7基因(pcr产物总长1023bp,如图1所示)连接到具有pubi启动子的载体ptck303上,具体方法如下:
(1)用t4连接酶将pcr产物与pmd18-t载体相连,连接产物命名为osfbn7-pmd18-t(如图2所示);
(2)利用bamhi和kpni酶切osfbn7-pmd18-t载体得到osfbn7,连入ptck303载体,连接产物命名为osfbn7-ptck303。
(3)将步骤(2)所得的osfbn7-ptck303质粒用bamhi和kpni酶切,利用1%的琼脂糖凝胶,120v、50ma电泳,紫外凝胶分析系统扫描成像。经酶切鉴定成功后,将质粒测序,待测序正确后进行下一步试验。
任何含有上述增强叶绿体脂质含量的相关蛋白编码基因osfbn7的重组载体、转基因细胞系和重组菌均属于本发明的保护范围。
可用现有的植物表达载体构建含有osfbn7基因的重组表达载体。所述植物表达载体包括双元农杆菌载体和可用于植物微弹轰击的载体等,如ptck303、pcambia3301、pcambia1300、pbi121、pbin19、pcambia2301、pcambia1301-ubin或其它衍生植物表达载体。
使用osfbn7基因构建重组表达载体时,可在其转录起始核苷酸前加上任何一种增强型、组成型、组织特异型或诱导型启动子,如花椰菜花叶病毒(camv)35s启动子、泛素(ubiquitin)基因启动子(pubi)等,它们可单独使用或与其它的植物启动子结合使用;此外,使用本发明的基因构建植物表达载体时,还可使用增强子,包括翻译增强子或转录增强子,这些增强子区域可以是atg起始密码子或邻接区域起始密码子等,但必需与编码序列的阅读框相同,以保证整个序列的正确翻译。所述翻译控制信号和起始密码子的来源是广泛的,可以是天然的,也可以是合成的。翻译起始区域可以来自转录起始区域或结构基因。
2、为了便于对转基因植物细胞或植物进行鉴定及筛选,可对所用植物表达载体进行加工,如加入在植物中表达可产生颜色变化的酶或发光化合物的基因(gus基因、gfp基因、萤光素酶基因等)、具有抗性的抗生素标记物(潮霉素、庆大霉素、卡那霉素等)或是抗化学试剂标记基因(如抗除莠剂基因)等。从转基因植物的安全性考虑,可不加任何选择性标记基因,直接以逆境筛选转化植株。
携带有本发明的与植物叶绿体发育相关蛋白编码基因osfbn7的植物表达载体可通过ti质粒、ri质粒、植物病毒载体、直接dna转化、显微注射、电导、农杆菌介导等常规生物学方法转化到目的植物细胞或组织中。上述基因osfbn7是通过上述重组表达载体导入上述目的植物中的。
将上述实施例中正确构建的osfbn7-ptck303载体利用电击转化法转化到农杆菌eha105菌株中,经过鉴定得到含有osfbn7-ptck303载体的阳性单克隆菌(如图3和图4)再将含上述载体的阳性单克隆菌株转化到野生型水稻植株中,得到水稻转基因幼苗,具体的方法如下:
(1)将含有osfbn7-ptck303载体的农杆菌接种于100mlyep(加入rif+kan)液体培养基中,28℃、200rpm振荡培养至菌液的od600为0.6~1.0时,5000rpm离心5min,收集菌体;用含有10mg/las(乙酰丁香酮)的aam液体培养基重悬菌体,调整菌液浓度od600至0.1~0.3;
(2)将od600为0.1~0.3的aam农杆菌悬液加入干燥且颜色亮黄的愈伤组织中,浸泡3~4min,倒掉多余的菌液并吹干愈伤组织;将愈伤组织置于n6-as共培养培养基中,22℃黑暗条件下培养2~3d;
(3)将共培养后的愈伤组织转移至50ml三角瓶中,灭菌水清洗,用无菌吸水纸吸干残留的水;将愈伤组织以几块愈伤堆在一起的形式接种至含有250mg/l羧苄青霉素和50mg/l潮霉素的选择培养基上,28℃,暗培养2~3周,直到看到有新的抗性愈伤长出;
(4)经过两次筛选后,将长出的抗性愈伤转至盛有再生ms-nk的培养基中,正常光周期14h/10h(光/暗)、28℃培养直至分化出幼苗。
上述ptck303载体所携带筛选抗性基因为潮霉素,用潮霉素抗性对水稻转基因幼苗进行筛选获得纯合转基因植株。具体方法如下:将上述获得的t1代具有潮霉素抗性的阳性苗进行单株收种得到t2代;对t2代幼苗进行潮霉素抗性的测试,选择3/4具有抗性而1/4没有抗性的株系,说明在该株系中含有目的基因的超表达载体以单拷贝形式插入;将上述株系中具有潮霉素抗性的植株移出,再进行单株收种得到t3代;对t3代幼苗进行潮霉素抗性筛选,选择没有分离的纯合转基因株系。该纯合株系可以用于繁种、叶绿体脂质含量检测、酶活性测定和种子中脂类和脂肪酸含量测定等实验。
3、采用qrt-pcr的方法检测所得野生型和osfbn7超表达转基因株系oe-osfbn7中osfbn7基因表达情况。
具体方法如下:提取野生型(wt)、osfbn7超表达转基因纯合植株的总rna,进行反转录得到cdna;利用下面引物进行qrt-pcr检测:
f2:5’-agcccgtcttctgtttgaga-3’(如seqidno.5所示);
r2:5’-gaccctccacagacaactga-3’(如seqidno.6所示);
actin1作为内参基因进行检测,所以引物如下,
f3:5’-tgctatgtacgtcgccatccag-3’(如seqidno.7所示);
r3:5’-aatgagtaaccacgctccgtca-3’(如seqidno.8所示);
qrt-pcr检测结果如图5所示,结果显示,超表达株系中osfbn7基因表达量较野生型显著提高。在其它植物中超表达osfbn7基因的方法可以参考本实施例进行。
超表达osfbn7转基因水稻株系叶绿体中脂质含量的检测
上述实施例中得到的osfbn7超表达转基因水稻纯合株系中叶绿体脂质含量检测具体方法如下:
选取生长良好的osfbn7超表达转基因纯合株系和野生型(zh11)水稻叶片5~10g,通过percoll梯度制备的方法,提取完整叶绿体。
取0.5~1mg叶绿体于50ml离心管中,加入10ml甲醇、10ml氯仿和5mlddh2o涡旋使溶液浑浊,1500g离心5min,取下层氯仿相(脂质),低压旋转蒸发器蒸发掉溶剂,取250μl氯仿溶解后,进行脂质含量测定(gc-ms)。
脂质含量测定结果如图6所示。结果显示,osfbn7基因超表达转基因纯合株系的叶绿体中甘油二脂(dag)和糖脂(mgdg和dgdg)含量较野生型显著增加。
超表达osfbn7转基因水稻种子中脂质和脂肪酸含量检测
成熟种子中油脂的提取与测定方法如下:
成熟水稻种子在磨粉机中磨碎后,取水稻粉末(100g)与300ml氯仿/甲醇(2:1,v/v)混匀后,轻轻震动提取2h,重复提取三次。然后,将20mlkcl(0.75%)添加到提取物中后,提取氯仿层,在旋转蒸发器中进行真空浓缩。然后,将脂类转移至25ml棕色玻璃瓶中,加入氯仿/甲醇(2:1,v/v)。利用氮气吹干后,在-25℃中保存,用于后续分析测定。提取的脂类经过甲酯化反应制备游离脂肪酸甲酯组分,利用esi-ms/ms测定水稻成熟种子中tag(甘油三脂)和脂肪酸的含量。
成熟水稻种子中tag含量和游离脂肪酸含量的测定结果如图7和图8所示。
图7结果显示,osfbn7基因超表达转基因纯合株系成熟种子中tag含量较野生型明显增加。osfbn7超表达转基因株系中tag含量平均为17.5μmol.mg-1dw(干重种子),较野生型中花11号(14.1μmol.mg-1dw)增加了24.1%。这表明osfbn7促进了成熟种子中油体(tag)的合成与积累。其中,osfbn7超表达明显提高c52:2、c52:3、c52:4和c54:3油体脂肪酸的组成和含量。
图8测定结果表明,成熟水稻种子中,c16:0、c18:0、c18:1和c18:2游离脂肪酸含量较野生型中花11的含量明显提高。而且,tag中脂肪酸种类和游离脂肪酸组成是一致的。因此,osfbn7促进了成熟水稻种子中游离脂肪酸的合成与积累。
osfbn7蛋白与kasib互作鉴定
osfbn7与oskasib互作的点对点酵母双杂交(y2h)鉴定方法具体如下:
水稻中存在2个kasi(β-酮脂酰-acp合酶i)基因:oskasia和oskasib,这两个基因编码蛋白的同源性高达84.5%,可能具有相似的功能。因此,在酵母双杂交文库筛选结果的基础上,为了验证osfbn7与oskasib蛋白之间的互作,本发明构建了oskasib-pgadt7质粒与osfbn7-pgbkt7质粒,进行了点对点酵母双杂交验证。
1、将候选基因的酵母质粒oskasib-pgadt7转入大肠杆菌jm109,提取含候选基因的大肠杆菌质粒。
2、将含候选基因的大肠杆菌质粒oskasib-pgadt7与osfbn7-pgbkt7重组质粒通过共转法转化酵母菌ah109。
3、将含有目的基因的重组质粒oskasib-pgadt7或osfbn7-pgbkt7,单独转化酵母菌ah109。
4、将转化好的酵母菌ah109分别在二缺(sd-trp-his)、三缺(sd-trp-ade-his)和含20mg/lα-x-gal的四缺(sd-trp-leu-his-ade+α-x-gal)的营养缺陷型选择培养基上进行涂板筛选。如果共转化酵母菌在四缺(sd-trp-leu-his-ade+α-x-gal)培养基上生长正常生且菌落变蓝,则可认为酵母双杂交验证的候选互作蛋白基因。图9结果显示,oskasib与osfbn7蛋白之间在水稻叶绿体中确实存在互作。
osfbn7超表达株系叶片中kasi酶活性测定
在鉴定osfbn7与oskasib互作的基础上,本发明测定了osfbn7超表达转基因纯合株系叶片中kasi酶活性,具体方法如下:
选取上述实施例中生长良好的osfbn7超表达转基因纯合株系和野生型zh11水稻开花期剑叶1g,并进行酶液提取。
利用武汉力博瑞公司的β-酮脂酰-acp合酶i酶联免疫分析试剂盒,检测kasi酶活性。每个样品进行4个生物学重复测定。
以标准物的浓度为横坐标,od值为纵坐标,利用excel绘出标准曲线,计算出标准曲线的回归方程。将样品的od值代入方程,即可计算出样品浓度,再乘以稀释倍数,即为样品的实际浓度。
水稻叶片中kasi酶活性的检测结果如图10所示,结果表明,超表达osfbn7显著提高叶片中kasi的酶活性。因此,本实验充分地证明了osfbn7与oskasib之间存在互作。而且,在osfbn7超表达转基因纯合株系中oskasi的酶促活性较野生型明显增加。表明osfbn7通过促进oskasi的酶促活性来增强水稻叶绿体中脂肪酸的从头生物合成途径。最终,提高了成熟水稻种子中脂肪酸的含量。
综上所述,植物叶绿体中脂肪酸从头合成途径是植物主要的脂肪酸合成途径。脂肪酸从头合成途径的最初步骤由乙酰辅酶a羧化酶(accase)催化。然后,由三种类型的β-酮脂酰-acp合酶(kasiii、i、ii)缩合,导致脂肪酸碳链的分阶段延长。这些kas蛋白在植物叶绿体中脂肪酸碳链从c2到c18缩合和延伸中的作用至关重要。目前,水稻叶绿体中存在5种kas蛋白:包括oskasia/ib、oskasiia/iib和oskasiii。oskasiii主要负责碳链c2-c4的延伸,oskasi主要负责脂肪酸的c4-c16的延伸,oskasii主要负责脂肪酸的c16-c18的延伸。因此,在脂肪酸从头生物合成过程中起非常关键的作用。本发明证明osfbn7蛋白可以通过与oskasib蛋白互作来促进oskasi的酶活性,进而提高植物叶绿体中脂质的含量。最终,在成熟水稻种子中油脂和脂肪酸的含量明显提高,这些结果为培育高油脂植物种子提供了一种新思路和新途径。因此,在生产上,利用调控水稻脂肪酸从头生物合成的基因osfbn7,将为植物高油脂含量品种培育提供有特色和应用潜力的基因资源。该基因将在运用基因工程手段提高植物油脂含量和调控脂肪酸合成途径研究中发挥重要作用,具有重要的实用价值和经济效益。
以上所述本发明的具体实施方式,并不构成对本发明保护范围的限定。任何根据本发明的技术构思所做出的各种其他相应的改变与变形,均应包含在本发明权利要求的保护范围内。
序列表
<110>华中农业大学
<120>水稻叶绿体蛋白osfbn7及其应用
<160>8
<170>siposequencelisting1.0
<210>1
<211>301
<212>prt
<213>水稻(oryzasativa)
<400>1
metalaalaalaalaalaalaalaleuilethralaalaserthrpro
151015
pheproleuvalserpheargserargargaspglyhisleuserleu
202530
serproproargargproglyalaglyargcysargalaseralapro
354045
thrpheglnglyglyproalaalasertyralaargglumetgluarg
505560
leuseralalysgluserleuleuleualapheargaspalaglygly
65707580
phegluserleuvalserglylysthrthrglymetglnlysileasp
859095
valasngluargilevalglyleugluargleuasnprothrproarg
100105110
prothrthrserpropheleugluglyargtrpasnpheglutrpphe
115120125
glyaspserserproglyalaleualaalaargleuleuphegluarg
130135140
serprothrthrvalalahisphethrglyleuaspvalleuilelys
145150155160
aspglytyrserlysileserserasnvallyspheleuasnthrval
165170175
glnserlyspheleuleuthrthrglnleuservalgluglyproile
180185190
argmetlysgluglutyrvalgluglyleuilegluileproargile
195200205
argglugluthrleuproaspglnleulysglyphepheglyglnthr
210215220
alaglyalaleuglnglnleuproalaproileargaspalavalser
225230235240
gluglyilelysleuproleuasnglymetpheglnargleuphemet
245250255
ilesertyrleuaspglugluileleuileileargaspalasergly
260265270
alaproaspvalleuthrargleugluglyproglnproasnserile
275280285
aspglythrseraspalavalleuserglutyrgluser
290295300
<210>2
<211>906
<212>dna
<213>水稻(oryzasativa)
<400>2
atggccgccgccgccgccgcggcgctcatcaccgccgcctccactcctttcccgctggtc60
tccttccgcagccggcgggatggccacctgagcctttcgcctcctcgccgccccggcgcc120
gggcgctgcagggcctcggcgccgacgttccaggggggacccgccgccagctacgcccgc180
gagatggagcgcctctccgccaaggagtccctcctcctcgccttcagagatgctggaggg240
tttgaatccttggttagtggaaagactacagggatgcagaagattgatgtcaatgagcgg300
attgttgggctcgagcgtctaaaccccactccacggcccacaacatctccctttctggaa360
ggtcgatggaactttgaatggtttggtgacagcagtcctggagcacttgcagcccgtctt420
ctgtttgagaggtcacctacaactgttgcacactttacaggacttgatgtcctgatcaag480
gatggatactccaaaatttcttccaacgtgaagttcttaaatacggtgcaaagcaagttc540
ctgctgactactcagttgtctgtggagggtcctattaggatgaaagaggaatatgtcgag600
ggcctaatagagatccctaggatcagagaagaaacactgcctgatcaactaaagggtttc660
tttggacaaactgcaggtgctctgcaacaactccctgcccccataagggacgctgtttca720
gaggggattaaattgccacttaatgggatgttccagcgcctattcatgatttcttacttg780
gatgaggaaatactgattatcagagatgcatctggagcacctgatgtcctaacaagattg840
gaagggccacagccaaattcaattgatggcacatcagacgcagtgctgtcagaatatgaa900
agctag906
<210>3
<211>25
<212>dna
<213>f1(artificialsequence)
<400>3
ggatccgcaactcatctcggcacca25
<210>4
<211>27
<212>dna
<213>r1(artificialsequence)
<400>4
ggtaccaagcgtgaaaagtggctgaac27
<210>5
<211>20
<212>dna
<213>f2(artificialsequence)
<400>5
agcccgtcttctgtttgaga20
<210>6
<211>20
<212>dna
<213>r2(artificialsequence)
<400>6
gaccctccacagacaactga20
<210>7
<211>21
<212>dna
<213>f3(artificialsequence)
<400>7
gctatgtacgtcgccatccag21
<210>8
<211>22
<212>dna
<213>r3(artificialsequence)
<400>8
aatgagtaaccacgctccgtca22
- 还没有人留言评论。精彩留言会获得点赞!