一种基于潮霉素筛选的大青杨转基因体系建立的方法与流程
[0001]
本发明属于植物育种技术领域,具体涉及一种基于潮霉素筛选的大青杨转基因体系建立的方法。
背景技术:
[0002]
大青杨(populus ussuriensis kom.)是东北林区主要的乡土树种,耐寒、速生,是山地营造速生用材林的主要树种之一,也是我国东北地区青杨派树种中分布最广、利用价值最大的树种。干形通直,木材洁白,是造纸及胶合板材极好的原料。随着天然林的不断采伐,人们开始营造大青杨的人工林。林地条件往往需要抗逆适应性强的大青杨新品种。但常规的育种周期长、工序复杂,且受外界环境的影响大,不能满足短期内有目的定向培育杨树新品种的需要。而基因工程育种可有目的定向培育杨树新品种,且极大程度上缩短育种周期。因此,大青杨转基因体系的建立尤为重要。
[0003]
目前,大青杨转基因体系已有研究,在构建大青杨转基因体系的过程中,常采用卡那霉素进行筛选,但该筛选方法的筛选效果不佳,大约有一半的概率为假阳性,给大青杨转基因植物的育种带来阻碍。
技术实现要素:
[0004]
有鉴于此,本发明的目的在于提供一种基于潮霉素筛选的大青杨转基因体系建立的方法,降低假阳性筛选结果,为大青杨转基因植物的构建奠定基础。
[0005]
本发明提供了一种基于潮霉素筛选的大青杨转基因体系建立的方法,包括以下步骤:
[0006]
1)将含目标基因的重组载体导入农杆菌中培养,得到农杆菌菌液;
[0007]
2)用所述农杆菌菌液侵染大青杨外植体后共培养,脱菌后,得到侵染后的大青杨外植体;
[0008]
3)将所述侵染后的大青杨外植体接种至含潮霉素的叶片不定芽分化培养基上进行筛选培养,得到不定芽;
[0009]
4)将所述不定芽进行生根培养,获得大青杨转基因再生植株。
[0010]
优选的,步骤3)中所述叶片不定芽分化培养基中潮霉素的浓度为2.5~3.2mg/l。
[0011]
优选的,步骤1)中含目标基因的重组载体的基础骨架载体为pcambia1302。
[0012]
优选的,步骤1)中所述农杆菌为根癌农杆菌;
[0013]
所述根癌农杆菌的菌株包括eha101、eha105或lba4404。
[0014]
优选的,步骤1)中所述农杆菌菌液的od值为0.5~0.8;
[0015]
所述农杆菌菌液的侵染时间为25~50min。
[0016]
优选的,步骤2)中所述农杆菌菌液与大青杨外植体共培养的时间为2~3.5d。
[0017]
优选的,步骤2)中所述农杆菌菌液与大青杨外植体共培养的温度为20~27℃。
[0018]
优选的,步骤2)中所述大青杨外植体包括茎段和叶片。
[0019]
优选的,步骤2)中所述叶片不定芽分化培养基为含0.5~2mg/l 6-ba、0.2mg/lnaa和0~0.1mg/ltdz的ms培养基。
[0020]
优选的,步骤4)中所述生根培养用培养基为1/2ms培养基。
[0021]
本发明提供的基于潮霉素筛选的大青杨转基因体系建立的方法,利用农杆菌介导法将目标基因转到大青杨的基因组中,以此来建立大青杨的转基因受体系统,同时在农杆菌侵染后通过限定以潮霉素为筛选标记,使潮霉素筛选得到的转化子假阳性相对较低,产生抗性的转化子经分子检测后阳性率达95%以上,克服了现有技术中常规采用卡那霉素为筛选标记,筛选效果不好,假阳性率高的问题,从而实现优化大青杨转化体系的目的。实验证明,在转基因系统稳定遗传转化效率已超过10%。
[0022]
进一步的,本发明具体限定了农杆菌介导法中多种能影响转化率的参数,包括潮霉素筛选工作浓度、根癌农杆菌菌株、农杆菌菌液od值、侵染时间、共培养时间以及温度等因素,能大大提高抗性芽诱导率。实验证明,通过严格限定影响转化率的参数的取值范围,使抗性芽诱导率达到30%~37%,同时还能降低污染率。
附图说明
[0023]
图1为基于潮霉素筛选的大青杨转基因体系建立的流程图;
[0024]
图2为转基因植株的gus活性检测;左图为非转基因植株(wt);右图为转基因植株(camv35s::gus);
[0025]
图3为大青杨转基因体系建立的过程;a为在添加2mg/l潮霉素的培养基中进行抗性芽的筛选;b-c为从伤口处长出的抗性芽;d为抗性芽的增殖,e-f为抗性芽的生根及植株再生;g为筛选的潮霉素抗性的转基因再生植株的扩繁结果;h为dna水平上的pcr检测;+表示以带有gus基因的质粒dna为模板进行的pcr作为阳性对照;-表示以非转基因的大青杨dna为模板的阴性对照;i为qrt-pcr检测结果。
具体实施方式
[0026]
本发明提供了一种基于潮霉素筛选的大青杨转基因体系建立的方法,包括以下步骤:
[0027]
1)将含目标基因的重组载体导入农杆菌中培养,得到农杆菌菌液;
[0028]
2)用所述农杆菌菌液侵染大青杨外植体后共培养,脱菌后,得到侵染后的大青杨外植体;
[0029]
3)将所述侵染后的大青杨外植体接种至含潮霉素的叶片不定芽分化培养基上进行筛选培养,得到不定芽;
[0030]
4)将所述不定芽进行生根培养,获得大青杨转基因再生植株。
[0031]
本发明将含目标基因的重组载体导入农杆菌中培养,得到农杆菌菌液。
[0032]
本发明对含目标基因的重组载体的构建方法没有特殊限制,采用本领域所熟知的重组载体的构建方法即可。本发明对所述目标基因的种类没有特殊限制,采用本领域感兴趣的目标基因即可。为了举例说明本发明构建的遗传转化体系的实施过程,本发明以gu报告基因作为目标基因,转gus的植株用x-gluc染色液进行检测,通过显色反应筛选转基因效率。呈阳性的植株会呈现蓝颜色。使用该报告基因很方便,它的阳性转化子很容易被检测。
[0033]
在本发明中,所述含目标基因的重组载体的基础骨架载体优选为pcambia1302载体。所述pcambia1302载体的来源购自addgene官网(https://www.fenghbio.cn/?360)。
[0034]
在本发明对重组载体导入农杆菌的方法没有特殊限制,采用本领域所熟知的转化方法即可。所述农杆菌优选为根癌农杆菌。所述根癌农杆菌的菌株包括eha101、eha105或lba4404。实验表明,不同根癌农杆菌的菌株转染效果存在差异,与gv3101菌株相比,eha101、eha105或lba4404菌株抗性芽诱导率有显著优势,抗性芽诱导率达到30%以上。
[0035]
在本发明中,所述农杆菌菌液的od值优选为0.5~0.8,更优选为0.6~0.8,最优选为0.8。实验表明,并非所述农杆菌菌液的od值越高取得的转基因效率就最高,od
600
达到1.0之后,抗性芽的诱导率是降低的,当od
600
为0.8时转基因效率最高,而且菌液浓度太高,菌生长速度比较快,很难脱菌,造成污染率比较高,因此,本发明选择od
600
0.8最为合适。
[0036]
得到所述农杆菌菌液后,本发明用所述农杆菌菌液侵染大青杨外植体后共培养,脱菌后,得到侵染后的大青杨外植体。
[0037]
在本发明中,所述农杆菌菌液的侵染时间优选为25~50min,更优选为30~45min,最优选为30min。实验表明,浸染时间直接影响转基因效率,侵染时间过短不利于农杆菌将目标基因转入植物细胞中,时间过长容易造成外植体活性降低。浸染30min和45min比15min的抗性芽诱导率要高,而且30min和45min没有差别,因为选择30min比较合适。
[0038]
在本发明中,农杆菌菌液与大青杨外植体共培养的时间优选为2~3.5d,更优选为2.5~3d。实验表明,共培养时间过长也会导致菌生长旺盛,不好脱菌,而共培养超过4天及以后,明显抗性芽的诱导率会下降,因此共培养时间选择3天最佳。所述农杆菌菌液与大青杨外植体共培养的温度优选为20~27℃,更优选为22~25℃,最优选为23℃。
[0039]
在本发明中,所述共培养时,将农杆菌菌液与大青杨置于叶片不定芽分化培养基中进行。所述叶片不定芽分化培养基优选为含0.5~2mg/l 6-ba、0.2mg/lnaa和0~0.1mg/ltdz的ms培养基,更优选为1~2mg/l 6-ba、0.2mg/l naa的ms培养基或1~2mg/l 6-ba、0.2mg/l naa和0.1mg/l tdz的ms培养基。本发明对所述叶片不定芽分化培养基的配制及灭菌方法没有特殊限制,采用本领域所熟知的培养基的制备及灭菌方法即可。
[0040]
在本发明中,所述大青杨外植体优选包括茎段和叶片,更优选为茎段。所述大青杨外植体的来源优选为大青杨的无菌组培苗。所述大青杨的无菌组培苗的制备方法将大青杨组织材料经消毒、水洗后制备外植体,接种至所述叶片不定芽分化培养基,得到的不定芽再接种至生根培养基上生根培养,得到组培苗。所述生根培养基优选为不加任何生长素的1/2ms培养基。所述生根培养的时间优选为四周。
[0041]
在本发明中,所述脱菌的方法采用灭菌水冲洗3~5遍培养的大青杨外植体。
[0042]
得到侵染后的大青杨外植体后,本发明将所述侵染后的大青杨外植体接种至含潮霉素的叶片不定芽分化培养基上进行筛选培养,得到不定芽。
[0043]
在本发明中,所述叶片不定芽分化培养基优选为含0.5~2mg/l 6-ba、0.2mg/lnaa和0~0.1mg/ltdz的ms培养基,更优选为含2~1mg/l 6-ba、0.2mg/lnaa的ms培养基或含2~1mg/l 6-ba、0.2mg/lnaa和0.1mg/l tdz的ms培养基,最优选为含2.0mg/l 6-ba、0.2mg/lnaa的ms培养基。所述叶片不定芽分化培养基中潮霉素的浓度优选为2.5~3.2mg/l,更优选为3.0mg/l。
[0044]
在本发明中,所述筛选培养的温度优选为22~25℃,最优选为23℃。所述筛选培养
的时间优选为12~17d,更优选为15d。选择叶片分化的潮霉素临界浓度的要求是,既能效抑制非转化细胞的生长,使之缓慢死亡,又不影响转化细胞的正常生长。实验表明,随着潮霉素浓度的升高,分化不定芽数下降,潮霉素浓度为1mg/l与2mg/l时仅能产生少量不定芽,而在浓度为3mg/l时不能分化形成芽。因此,进行大青杨遗传转化时,潮霉素浓度为3mg/l时较为合适。
[0045]
在本发明中,所述生根培养用培养基优选为1/2ms培养基。大青杨很容易生根,根系也很发达,不定芽在不含激素的培养基上生长也会自然伸长同时生根,有利于转基因植株的获得。
[0046]
在本发明中,对获得的转基因植株进行检测。所述检测方法包括生理生化检测和分子检测。所述生理生化检测以报告基因的作用进行检查。在本发明实施例中,gus基因能够经化学染色后,植物组织叶片变色,来判断转基因植物为阳性转基因植物。所述分子检测优选采用pcr、qrt-pcr检测方法检测目标基因片段,通过分析目标基因的片段及表达量来判断是否未阳性转基因植物。本发明通过gus组织化学染色、pcr、qrt-pcr检测大青杨转基因系统遗传转化效率,在转基因系统稳定遗传转化效率已超过10%。
[0047]
下面结合实施例对本发明提供的一种基于潮霉素筛选的大青杨转基因体系建立的方法进行详细的说明,但是不能把它们理解为对本发明保护范围的限定。
[0048]
实施例1
[0049]
不定芽的诱导培养基的筛选
[0050]
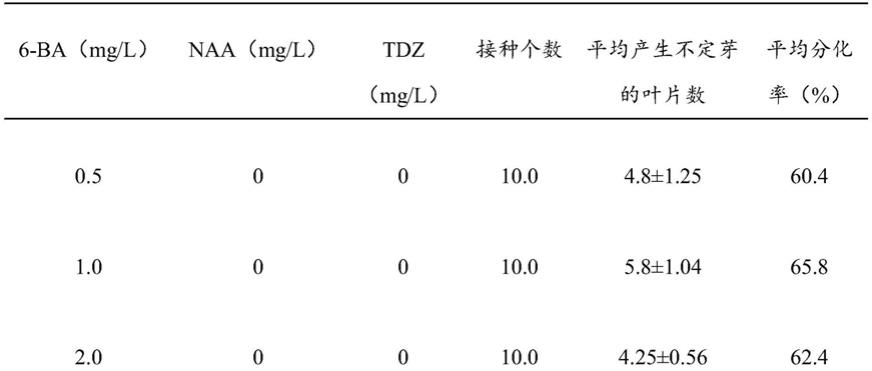
当年生的大青杨实生苗生长到2个月的时候,苗高大概50cm从实生苗选取顶端叶片。放在100ml三角瓶中,纱布包住瓶口,绑紧后放在水龙头下冲洗二至四小时,取出材料放入75%酒精里消毒30s~1min,用高压灭过菌的蒸馏水冲洗3次,然后将材料放入的1%次氯酸钠溶液中消毒20min,用灭菌水冲洗3次。以ms为基本培养基,选定6-ba、naa和tdz三种生长调节剂,设定不同浓度梯度,得到多种不同浓度梯度培养基,再将大青杨组培苗的茎段、叶片分别接种于不同的筛选培养基上,其中每个培养皿培养10个外植体,每个处理重复三次,每四周继代一次,记录不同处理的不定芽诱导率。
[0051]
不同种类筛选培养基的筛选结果见表1。
[0052]
表1不同生长调节物质对不定芽的诱导
[0053]
[0054][0055]
由表1结果表明,不同浓度的6-ba、naa和tdz组合后不定芽的分化情况,其中1.0mg/l 6-ba+0.2mg/lnaa+0.1mg/ltdz条件下的不定芽分化效率最高(见表1),达到了96.3%。
[0056]
实施例2
[0057]
根癌农杆菌介导法不同参数对转化率的影响
[0058]
将报告基因gus插入植物表达载体pcambia1302中,并转化到根癌农杆菌中,在1.0mg/l 6-ba+0.2mg/lnaa+0.1mg/ltdz培养基中培养,经过鉴定,将转化有gus的阳性根癌农杆菌培养至一定浓度后,得到根癌农杆菌菌液。
[0059]
用根癌农杆菌菌液分别侵染大青杨外植体,侵染结束后添加实施例1筛选得到的不定芽诱导培养基进行共培养,脱毒后,得到侵染后的大青杨外植体。将侵染后的大青杨外植体在实施例1筛选得到的含潮霉素的不定芽诱导培养基进行诱导培养,得到不定芽。将不定芽转接至1/2ms培养基上培养,得到转基因大青杨再生植株。
[0060]
1)根癌农杆菌菌株对抗性芽的诱导率的影响
[0061]
使用eha101、eha105、lba4404、gv13014种不同农杆菌菌株对大青杨转基因效率的影响。将构建好的载体转入上述4种菌株中,经检测合格后,培养至od
600
值0.8时收集农杆菌菌液。用农杆菌菌液在常温条件下侵染大青杨的嫩叶30min,每次浸染50个外植体,三次生物学重复,培养一个月之后按式i统计计算抗性芽的诱导率(%)。
[0062]
抗性芽的诱导率(%)=长出抗性芽的叶片个数/转基因外植体的总叶片数(个)式i。
[0063]
表2不同农杆菌菌株对抗性芽诱导率的结果
[0064][0065]
结果表明(表2)不同农杆菌菌株对抗性芽诱导率也有一定的影响,与gv3101相比,eha101、eha105、lba4404对诱导抗性芽具有明显优势,其中eha105对大青杨的抗性芽的诱导率是最高的,高达35.3%,因此,后续用eha105菌株进行转基因。
[0066]
2)根癌农杆菌菌液od值对抗性芽的诱导率的影响
[0067]
农杆菌的浸染浓度也是影响转基因转化效率的一个关键因素,分别用od
600
为0.2、0.5、0.8、1.0、1.2的eha105菌株菌液与外植体混合,常温浸染30min,每次浸染50个外植体,三次生物学重复。侵染后的外植体接种实施例1筛选的不定芽诱导培养基进行共培养,经过灭菌水洗涤脱菌后,接种至含潮霉素不定芽诱导培养基上进行筛选培养,培养一个月之后统计抗性芽的诱导率(%)。
[0068]
表3农杆菌菌液浓度对抗性芽诱导率的影响
[0069][0070][0071]
结果表明(表3),随着农杆菌菌液浓度的提高,抗性芽的诱导率随之提高,但是od
600
达到1.0之后,抗性芽的诱导率开始降低,当od
600
0.8时的转基因效率最高,而且菌液浓度太高,菌生长速度比较快,很难脱菌,造成污染率比较高,因此,选择od
600
为0.8最为合适。
[0072]
3)浸染时间对抗性芽的诱导率的影响
[0073]
为研究浸染时间对转基因效率的影响,用od
600
为0.8的eha105菌株菌液分别对大青杨外植体侵染15min、30min、45min,共培养3天后,在含潮霉素3mg/l的不定芽诱导培养基进行筛选培养,培养一个月之后统计抗性芽的诱导率(%)。
[0074]
表4浸染时间对抗性芽诱导率的影响
[0075][0076]
结果表明(表4)浸染30min和45min比15min的抗性芽诱导率要高,而且30min和45min没有差别,因为选择30min比较合适。
[0077]
4)共培养时间对抗性芽的诱导率的影响
[0078]
为了研究共培养时间影响转基因效率,在上述培养方法基础上,选择了1d、2d、3d、4d四个时间段来判断其对抗性芽诱导率的影响。
[0079]
表5共培养时间对抗性芽诱导率的影响
[0080][0081]
结果表明(表5)共培养3天的效率最高,其次是2天。共培养时间过长也会导致菌生长旺盛,不好脱菌,所以4天后,明显抗性芽的诱导率会下降。因此,选择3天共培养效果最佳。
[0082]
5)共培养温度
[0083]
共培养温度有时对转基因效率也会有影响,分别在21℃、23℃、25℃培养条件下共培养3天,每次浸染50个外植体,三次生物学重复。共培养3天后,在含潮霉素3mg/l的不定芽诱导培养基进行筛选培养培养一个月之后统计抗性芽的诱导率(%)。
[0084]
表6共培养温度对抗性芽诱导率的影响
[0085][0086]
结果表明(表6),共培养温度为23℃时的抗性芽诱导率最高,但是三个温度下的抗性芽诱导率差别不大。
[0087]
6)潮霉素筛选浓度的确定
[0088]
在上述筛选结果的基础上,以野生型大青杨外植体为材料,在固体分化培养基中分别添加浓度为0、1、2、3、4、5、6mg/l 6个潮霉素浓度筛选半个月。由于潮霉素毒性较大,在上述筛选结果的基础上进行培养,统计半个月的存活率,最终以刚好死亡的临界浓度作为筛选转化子的合适浓度潮霉素敏感度测试。
[0089]
表7不同浓度的潮霉素对大青杨不定芽诱导的影响
[0090][0091]
将幼嫩叶片接种在不含潮霉素的叶片不定芽分化培养基对照上,接种后,叶片几乎都能产生大量不定芽,而在附加不同浓度潮霉素的培养基上,叶片的反应不同。由表2可以看出,随着潮霉素浓度的升高,分化不定芽数下降,潮霉素浓度为1mg/l与2mg/l时仅能产生少量不定芽,而在浓度为3mg/l时不能分化形成芽。选择叶片分化的潮霉素临界浓度的要求是既能效抑制非转化细胞的生长,使之缓慢死亡,又不影响转化细胞的正常生长。因此,进行大青杨遗传转化时,潮霉素浓度为3mg/l时较为合适。
[0092]
选择潮霉素为筛选标记,使筛选得到的转化子相对假阳性比较低,产生抗性的转
化子经分子检测后95%以上为阳性,与先前选用的卡那霉素为筛选标记基因筛选效果约有一半概率假阳性结果相比,使用潮霉素作为筛选标记能提高大青杨转化体系的准确性。
[0093]
7)外植体种类:挑选生长良好的组培苗的茎段和叶为材料为外植体,与培养好的农杆菌菌液(od
600
=0.8)共培养30min,每次浸染50个外植体,三次生物学重复,培养一个月之后统计抗性芽的诱导率(%)。
[0094]
表8不同外植体对抗性芽诱导率的影响
[0095][0096]
结果表明(表8),叶片的浸染效率高一些,以叶片为外植体得到的抗性芽比茎段获得的数量要高,而且茎段不太容易脱菌。但抗性芽的分化率均超过20%,而且筛选时间可控制在一个月之内,达到预期技术目标。
[0097]
实施例3
[0098]
一种基于潮霉素筛选的大青杨转基因体系建立的方法,包括以下步骤:
[0099]
1)将含gus基因插入植物表达载体pcambia1302中,得到的重组载体转化根癌农杆菌eha105菌株中,经培养,利用gus基因的特异性引物f:5
′-
aatccatcgcagcgtaatgctct-3
′
(seq id no:1);r:5
′-
gctggcctgcccaacctttcggtat-3
′
(seq id no:2)进行pcr扩增鉴定,得到含gus基因的阳性农杆菌菌液;
[0100]
2)用od
600
为0.8的农杆菌菌液侵染消毒后的大青杨幼嫩叶片30min后,添加叶片不定芽分化培养基(含2mg/l 6-ba、0.2mg/lnaa的ms培养基)在23℃条件下进行共培养3d,用灭菌水清洗3~4次后脱菌,得到侵染后的大青杨外植体;
[0101]
3)将所述侵染后的大青杨外植体接种至含3mg/l潮霉素的叶片不定芽分化培养基上,在23℃条件下筛选培养1个月,得到大量不定芽;统计不定芽数量,计算抗性芽诱导率,抗性芽诱导率高达35%以上;
[0102]
4)将所述不定芽接种至不加任何生长素的1/2ms培养基中继续培养四周,获得大青杨转基因再生植株。
[0103]
5)将得到的大青杨转基因再生植株材料用x-gluc染色液进行检测,同时以野生型大青杨作为对照。通过显色反应筛选转基因效率,筛选的抗性芽绝大多数为阳性(98%以上)。结果见图2。
[0104]
与对照组相比,本发明构建的大青杨转基因再生体系显示蓝色,说明构建的大青杨转基因再生体系成功构建含gus基因的再生体系。
[0105]
同时提取染色结果阳性的转基因植株材料的dna和rna,分别采用普通普通pcr扩增方法和qrt-pcr方法检测gus基因及其表达情况,同时以野生型大青杨(wt)作为对照。
[0106]
①
普通pcr扩增方法
[0107]
设计gus基因的特异性引物f:5
′-
aatccatcgcagcgtaatgctct-3
′
(seq id no:1);r:5
′-
gctggcctgcccaacctttcggtat-3
′
(seq id no:2)。随机选择14个gus阳性转基因植株
株系,提取基因组dna,并且以该基因组dna为模板,以特异性引物f和特异性引物r作为引物对,进行pcr扩增检测,其中pcr程序为94℃预变性3min;94℃变性30s,58℃退火30s,72℃延伸1min,循环30次;72℃总延伸7min。结果如图3中h,7个株系经过pcr检测后呈阳性。
[0108]
②
实时荧光定量pcr(qrt-pcr)分析方法
[0109]
采用试剂盒的方法提取上述7个株系的rna,以puactin基因(引物序列:5
′-
tgttgcccttgactatgagcagga-3
′
,seq id no:5;5
′
acggaatctctcagctccaatggt-3
′
,seq id no:6)为内参基因,以f:5
′-
tacggcaaagtgtgggtcaa-3
′
(seq id no:3)和5
′-
tcttgccgttttcgtcggta-3
′
(seq id no:4)作为引物进行qrt-pcr扩增,其中qrt-pcr总反应体系为20μl:cdna模版1μl、正反向引物各1μl、2
×
easy taq pcr supermix酶10μl、去离子水7μl。反应程序:94℃预变性30s;94℃变性5s,60℃,30s,共45个循环;55℃至99℃每0.5℃读板一次,保持1s。
[0110]
结果如图3中i所示,7个株系中gus基因的表达量水平不同,但7个株系均比wt要高很多倍。
[0111]
实施例4
[0112]
一种基于潮霉素筛选的大青杨转基因体系建立的方法,包括以下步骤:
[0113]
1)将含gus基因插入植物表达载体pcambia1302中,得到的重组载体转化根癌农杆菌eha101菌株中,经培养,经pcr扩增鉴定,得到含gus基因的阳性农杆菌菌液;
[0114]
2)用od
600
为0.7的农杆菌菌液侵染消毒后的大青杨幼嫩茎段30min后,添加叶片不定芽分化培养基(含1mg/l6-ba、0.2mg/lnaa和0.1mg/ltdz的ms培养基)在23℃条件下进行共培养3d,用灭菌水清洗3~4次后脱菌,得到侵染后的大青杨外植体;
[0115]
3)将所述侵染后的大青杨外植体接种至含3mg/l潮霉素的叶片不定芽分化培养基上,在23℃条件下筛选培养1个月,得到大量不定芽;统计不定芽数量,计算抗性芽诱导率,抗性芽诱导率高达35%以上;
[0116]
4)将所述不定芽接种至不加任何生长素的1/2ms培养基中继续培养四周,获得大青杨转基因再生植株,假阳性率低于2%。
[0117]
5)将得到的大青杨转基因再生植株材料用x-gluc染色液进行染色检测,同时以野生型大青杨作为对照。
[0118]
通过gus组织化学染色、pcr、qrt-pcr检测大青杨转基因系统遗传转化效率,在转基因系统稳定遗传转化效率已超过10%。
[0119]
以上所述仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1