一种与花粉发育相关的蛋白及其编码基因的应用
1.本发明属于植物基因工程领域,具体涉及一种与花粉发育相关的蛋白及其编码基因的应用。
背景技术:
2.高等植物的生长发育分为营养生长和生殖生长两个阶段,当到达特定时期,植物由营养生长向生殖生长转换,而正常开花的过程是植物进行生殖生长的重要节点,因此开花被认为是植物个体发育过程中的中心环节。植物的花器官对于人们欣赏、饮食及其果蔬、农作物丰产等方面都具有很重要的作用,因此研究植物开花过程、花发育的调控机理在理论及实践应用中都具有很好的意义。
3.植物的花发育及开花是一个复杂且高度程序化的过程,受外在环境及内在遗传机制的共同作用,其中多种植物激素及其调控基因参与其中。
技术实现要素:
4.本发明的目的是提供一种与花粉发育相关的蛋白及其编码基因的应用。
5.本发明提供了slpp2c5蛋白在调控植物花粉形成中的应用。
6.本发明还提供了slpp2c5蛋白在调控植物花粉发育中的应用;。
7.本发明还提供了slpp2c5蛋白在调控植物花发育中的应用。
8.本发明还提供了编码slpp2c5蛋白的核酸分子在制备花粉畸形率增高的转基因植物中的应用。
9.本发明还提供了编码slpp2c5蛋白的核酸分子在制备雄性不育的转基因植物中的应用。
10.本发明还提供了一种培育转基因植物的方法,包括如下步骤:将编码slpp2c5蛋白的核酸分子导入受体植物中,得到花粉畸形率增高的转基因植物。
11.本发明还提供了一种植物育种方法,包括如下步骤:增加目的植物中slpp2c5蛋白的含量和/或活性,从而使植物花粉畸形率增高。
12.本发明还提供了一种培育转基因植物的方法,包括如下步骤:将编码slpp2c5蛋白的核酸分子导入受体植物中,得到雄性不育的转基因植物。
13.本发明还提供了一种植物育种方法,包括如下步骤:增加目的植物中slpp2c5蛋白的含量和/或活性,从而使植物雄性不育。
14.以上任一所述slpp2c5蛋白,获自番茄(solanum lycopersicum),为是如下(a1)或(a2)或(a3)或(a4):
15.(a1)序列表的序列1所示的蛋白质;
16.(a2)将序列表的序列1所示的蛋白质经过一个或几个氨基酸残基的取代和/或缺失和/或添加且与植物花粉形成和/或花粉发育相关的由其衍生的蛋白质;
17.(a3)在(a1)所述蛋白质的n端或/和c端连接标签得到的融合蛋白;
18.(a4)来源于番茄且与(a1)具有98%以上同一性且与植物花粉形成和/或花粉发育相关的蛋白质。
19.标签具体如表1所示。
20.表1标签的序列
21.标签残基序列poly-arg5-6(通常为5个)rrrrrpoly-his2-10(通常为6个)hhhhhhflag8dykddddkstrep-tag ii8wshpqfekc-myc10eqkliseedlha9ypydvpdya
22.以上任一所述编码slpp2c5蛋白的核酸分子是如下(b1)或(b2)或(b3)或(b4)的dna分子:
23.(b1)编码区如序列表的序列2中第167-1609位核苷酸所示的dna分子;
24.(b2)序列表的序列2所示的dna分子;
25.(b3)在严格条件下与(b1)或(b2)限定的dna分子杂交且编码所述蛋白质的dna分子;
26.(4)来源于番茄且与(b1)或(b2)限定的dna分子至少具有98%同源性且编码所述蛋白质的dna分子。
27.所述严格条件是在2
×
ssc,0.1%sds的溶液中,在68℃下杂交并洗膜2次,每次5min,又于0.5
×
ssc,0.1%sds的溶液中,在68℃下杂交并洗膜2次,每次15min。
28.以上任一所述将编码slpp2c5蛋白的核酸分子导入受体植物具体可为将具有编码slpp2c5蛋白的核酸分子的重组质粒导入受体植物。具体的,所述重组质粒可为将序列表的序列2中第167-1609位核苷酸所示的双链dna分子插入pri101-an载体的多克隆位点(例如xbai和saci酶切位点之间)得到的重组质粒。
29.以上任一所述植物可为双子叶植物。
30.以上任一所述植物可为茄科植物。
31.以上任一所述植物可为番茄属植物。
32.以上任一所述植物可为番茄,例如番茄micro-tom。
33.本发明的发明人发现,在番茄中过表达slpp2c5基因,可使番茄花发育异常、花粉畸形率增高。因此,slpp2c5蛋白与番茄花发育过程相关,与花粉发育相关。slpp2c5蛋白及其编码基因在番茄高品质育种中具有重要作用。
附图说明
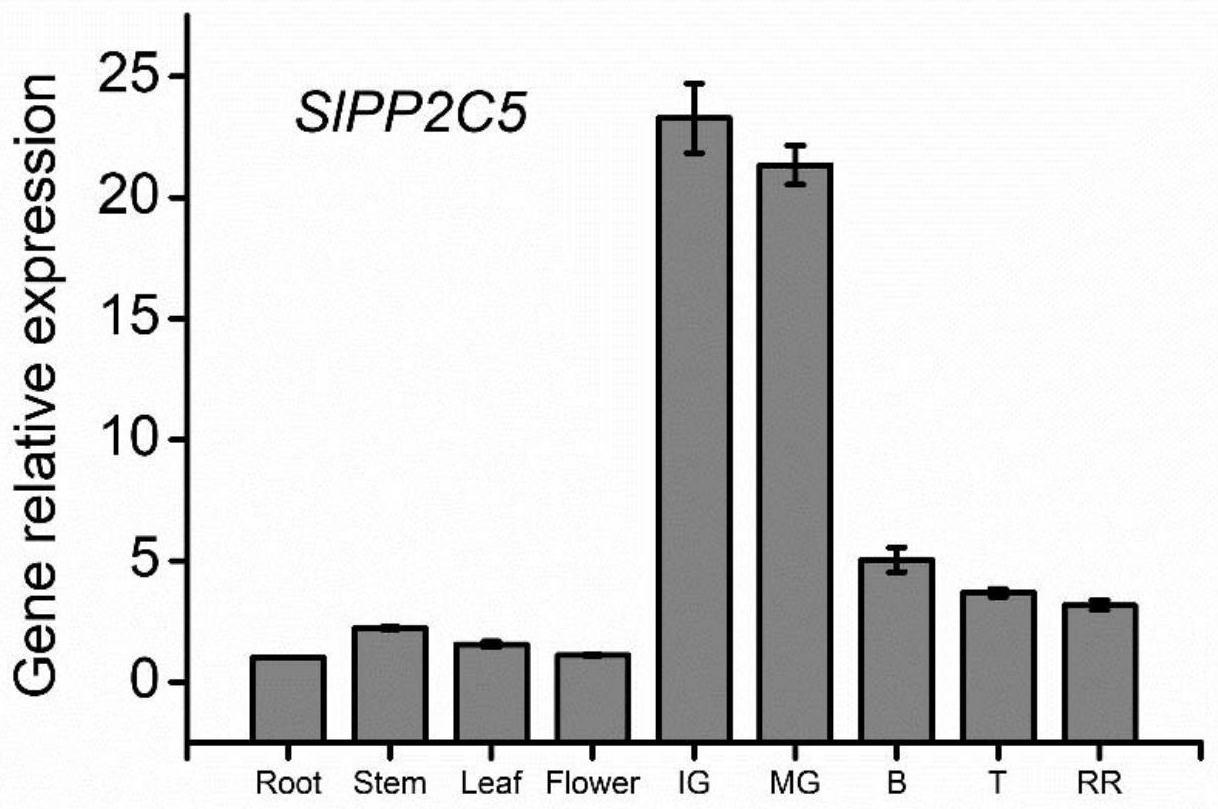
34.图1为slpp2c5基因在番茄果实发育成熟过程中的表达变化情况。
35.图2为花粉电镜下照片。
36.图3为花粉畸形率。
具体实施方式
37.下面结合具体实施方式对本发明进行进一步的详细描述,给出的实施例仅为了阐明本发明,而不是为了限制本发明的范围。以下提供的实施例可作为本技术领域普通技术人员进行进一步改进的指南,并不以任何方式构成对本发明的限制。
38.下述实施例中的实验方法,如无特殊说明,均为常规方法,按照本领域内的文献所描述的技术或条件或者按照产品说明书进行。下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。如无特殊说明,以下实施例中的定量试验,均设置三次重复实验,结果取平均值。番茄micro-tom,番茄常规品种,也称为野生型番茄,用wt表示。番茄的果皮指的是果实除去外果皮、果胶和种子以外的部分。
39.从番茄中发现一个新蛋白,如序列表的序列1所示,命名为slpp2c5蛋白。将编码slpp2c5蛋白的基因命名为slpp2c5基因。番茄cdna中,slpp2c5基因如序列表的序列2所示(开放阅读框如序列2中第167-1609位所示)。
40.实施例1、slpp2c5基因在番茄果实发育成熟过程中的表达变化情况
41.根据番茄植株盛花期后坐果天数及果实大小、颜色变化情况,将果实发育成熟过程分为幼果期(ig)、绿熟期(mg)、破色期(b)、转色期(t)和红熟期(rr)。
42.取番茄micro-tom植株的根、茎、叶、花以及每个时期的果实的果皮进行液氮冻存。取液氮冻存样本,提取总rna,反转录得到cdna。以cdna为模板,通过实时定量qpcr检测slpp2c5基因的表达量(内参基因为slsand基因)。
43.用于检测slpp2c5基因的引物如下:
44.slpp2c5-qf(上游引物):gtgtatttggcgttcttgcaatgtc;
45.slpp2c5-qr(下游引物):caggcagagggttagtcccgttc。
46.反应体系:2
×
sybr premix ex taq 10μl,上游引物(10mm)0.5μl,下游引物(10mm)0.5μl,cdna 1.5μl,ddh2o 7.5μl。
47.结果见图1。
48.实施例2、转slpp2c5基因番茄的获得以及果实相关性状的检测
49.一、重组表达载体的构建
50.将序列表的序列2中第167-1609位核苷酸所示的双链dna分子插入pri101-an载体的xbai和saci酶切位点之间,得到重组质粒。重组质粒已进行测序验证。
51.二、制备转基因番茄植株
52.1、取番茄micro-tom的种子,用75%乙醇水溶液消毒30s,然后用次氯酸钠水溶液消毒15min,然后用无菌水冲洗6-8遍。
53.2、取完成步骤1的种子,置于固体1/2ms培养基,25℃光暗交替培养7-8天(此时子叶充分展开、真叶尚未形成)。
54.3、完成步骤2后,剪取子叶,剪除两端,中部剪成边长0.5cm左右的正方形,即为外植体;将外植体置于预培养培养基,25℃黑暗培养2天。
55.预培养培养基:含2.5mg/l 6-ba和0.2mg/l iaa的固体ms培养基。
56.4、完成步骤3后,将外植体置于侵染液中侵染5min,然后取出外植体并吸干残余菌液,然后置于共培养培养基,25℃黑暗培养2天。
57.侵染液:将步骤一得到的重组质粒导入农杆菌lba4404,得到重组农杆菌;用含
100mg/l as的液体ms培养基重悬重组农杆菌,使od
600nm
=0.5,即为侵染液。
58.共培养培养基:含2.5mg/l 6-ba、0.2mg/l iaa和100mg/l as的固体ms培养基。
59.5、完成步骤4后,将外植体转移至除菌培养基,先25℃黑暗培养2-3天,再25℃光暗交替培养3-4天。
60.除菌培养基:含2.5mg/l 6-ba、0.2mg/l iaa、200mg/l carb、300mg/l cef的固体ms培养基。
61.6、完成步骤5后,将外植体转移至筛选培养基,25℃光暗交替培养30-45天(每隔15天继代一次),得到植株。
62.筛选培养基:含2.5mg/l 6-ba、0.2mg/l iaa、200mg/l carb、300mg/l cef、4mg/l hyg的固体ms培养基。
63.7、完成步骤6后,将植株转移至生根培养基,25℃光暗交替培养,得到生根的植株,即为t0代再生植株。
64.生根培养基:含0.2mg/l iaa和200mg/l carb的1/2ms固体培养基。
65.8、从t0代再生植株中筛选转基因植株。
66.筛选转基因植株的方法:待植株长至根长2-4cm时取植株叶片进行pcr鉴定,如果得到特异性扩增产物,该再生植物为pcr鉴定阳性植株,即转基因植株。
67.用于pcr鉴定的引物如下:
68.35s-f:gcaagacccttcctctatataagg;
69.slpp2c5-r:acttttgcttttgaacttcctgtg。
70.9、t0代转基因植株自交,得到的后代植株即为t1代植株;从t1代植株中筛选转基因植株(方法同步骤8)。
71.10、t1代转基因植株自交,得到的后代植株即为t2代植株;从t2代植株中筛选转基因植株(方法同步骤8)。
72.对于某一t1代转基因植株,如果其自交得到的t2代植株均为转基因植株,该t1代植株为纯合的转基因植株,其自交后代为纯合的转基因株系。取2个转基因株系(即slpp2c5-oe-2株系和slpp2c5-oe-10株系)进行步骤四。
73.三、制备转空载体番茄植株
74.用pri101-an载体代替重组质粒,参照步骤二进行操作,得到转空载体株系。
75.四、果实品质分析
76.供试植株:slpp2c5-oe-2株系的t3植株、slpp2c5-oe-10株系的t3植株、野生型番茄植株、转空载体株系的t3植株。
77.在平行条件下正常培养供试植株,盛花期取花粉进行电镜观察,统计花粉畸形率。
78.花粉电镜下照片见图2。野生型番茄植株的花粉粒呈麦粒型,有三条萌发沟。slpp2c5-oe-2株系植株和slpp2c5-oe-10株系植株的花粉粒空瘪变形,萌发沟浅且中部凸起。花粉管通常从萌发沟进入,因此转基因植株的花粉干瘪及萌发沟畸形会导致花粉活力降低。与野生型番茄植株相比,转空载体株系植株的花粉表型无显著差异。
79.花粉畸形率见图3(每个株系至少检测3株植株)。野生型番茄植株的花粉畸形率为10%。转空载体株系植株的花粉畸形率为11%。slpp2c5-oe-2株系植株和slpp2c5-oe-10株系植株的花粉畸形率分别为66.7%和81.8%,即转基因植株的花粉畸形率显著高于野生型
植株。
80.以上结果表明,表明过表达slpp2c5基因影响了番茄花的正常发育,slpp2c5蛋白在番茄花粉发育过程中发挥重要作用。
81.以上对本发明进行了详述。对于本领域技术人员来说,在不脱离本发明的宗旨和范围,以及无需进行不必要的实验情况下,可在等同参数、浓度和条件下,在较宽范围内实施本发明。虽然本发明给出了特殊的实施例,应该理解为,可以对本发明作进一步的改进。总之,按本发明的原理,本技术欲包括任何变更、用途或对本发明的改进,包括脱离了本技术中已公开范围,而用本领域已知的常规技术进行的改变。按以下附带的权利要求的范围,可以进行一些基本特征的应用。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1