一种重组质粒、包含其的基因工程菌及其在制备碳酸二甲酯中的应用的制作方法
本发明属于生物化工领域,具体涉及一种重组质粒、由其表达的脂肪酶突变体及其在制备碳酸二甲酯中的应用。
背景技术:
碳酸二甲酯简称dmc,常温时是一种无色透明、略有气味的液体,熔点4℃,沸点90.1℃,密度1.0699/cm3,难溶于水,但可以与醇、醚、酮等几所有的有溶剂混溶。碳酸二甲酯性能优良,用途广泛,除用于甲基化、羰基化和碳基甲氧基化反应外,从dmc出发还可制备多种衍生物,如高性能的树脂、溶剂、染料中间体、药物、香料、润滑油添加剂等。碳酸二甲酯具有无毒、对环境影响小的特性,属于新型的“绿色”化工原料,被称为21世纪有机合成的“新基石”,并在近年来取得了迅猛的发展。
dmc现有几种生产工艺,存在以下缺陷:光气法污染严重,剧毒物的使用不符合绿色化工要求,已经被淘汰;氧化羰基化法反应条件苛刻,设备投资较大,工艺操作费用高,co的引入有潜在爆炸危险;酯交换法单程转化率低,需大回流比提高反应物转化率,能耗高,另外,副产物二元醇受到市场条件限制,影响经济效益。
酶作为一种生物催化剂,近年来已被人们广泛应用于食品生产与检测、环保技术、生物技术、生物医药等领域。目前研究和应用最广泛的脂肪酶(lipase,ec3.1.1.3)是一类能催化油脂和短链醇进行转酯化反应生成脂肪酸甲酯的生物催化剂,脂肪酶所介导的反应具有反应条件温和、醇用量小、产品易收集纯化、无污染物排放等优点。目前国内外用于转酯化研究的脂肪酶主要为诺维信公司生产的novozym435,此酶为树脂固定化酶,载体为大孔丙烯酸树脂,价格高昂,不利于酶法制备化工原料的工业化生产。虽然已有专利报道使用脂肪酶催化甲醇与碳酸乙烯酯或甲醇与碳酸丙烯酯进行转酯反应制备碳酸二甲酯,但反应时间较长,酶活性有待提高。
技术实现要素:
本发明所要解决的技术问题是提供一种可高效催化制备碳酸二甲酯的基因工程菌,本发明提供的基因工程菌可以表达突变脂肪酶,从而有效提高制备碳酸二甲酯的效率,应用前景广阔。
一方面,本发明提供一种重组质粒,其在酶切位点ecori和hindiii之间包含用于表达脂肪酶突变体的核苷酸序列,其中所述序列选自seqidno:3、seqidno:7和seqidno:9。
另一方面,本发明提供一种上述重组质粒的制备方法,所述方法包括:
1)将pel编号为genbank:af284064.1的基因序列通过全基因合成,并连接在质粒pet-28a上,酶切位点为ecori和hindiii,获得质粒pet28a-pel;
2)以质粒pet28a-pel为模板,以下列引物对之一进行高保真pcr扩增:
引物对1:
83正向:5’gctctcatcactcctgtgctctcgggcgtgact3’(seqidno:1)
83反向:5’agtcacgcccgagagcacaggagtgatgagagc3’(seqidno:2);
引物对2:
92正向:5’actttcccctctaatgtgaagatcatg3’(seqidno:5)
92正向:5’catgatcttcacattagaggggaaagt3’(seqidno:6);
3)pcr之后通过琼脂糖电泳验证pcr产物大小,异丙醇沉淀回收pcr产物,dpni酶切去除模板,转化大肠杆菌后提取得到包含e83v或d92n突变酶基因的重组质粒,
或者
所述方法包括:在上述步骤3)之后,以与前次所用引物对不同的引物对重复步骤2)和3),,也即,如果第一次pcr扩增采用引物对1,则在述步骤3)之后,采用引物对2重复步骤2)和3),如果第一次pcr扩增采用引物对2,则在述步骤3)之后,采用引物对1重复步骤2)和3),以获得包含e83v和d92n双突变酶基因的重组质粒。
在具体实施方式中,步骤2)中的pcr条件为:
98℃预变性5min;98℃变性10s;55℃退火15s;72℃延伸6min,25个循环,72℃延伸6min。
另一方面,本发明提供了一种催化制备碳酸二甲酯的基因工程菌,所述基因工程菌包含上述重组质粒。
在具体实施方式中,所述基因工程菌为以大肠杆菌为出发菌株,并包含表达脂肪酶突变体的质粒,
其中,所述脂肪酶突变体相对于扩展青霉pf898的脂肪酶具有选自以下突变中的任一者或两者组合的突变:第83位谷氨酸突变为缬氨酸(e83v),以及第92位天冬氨酸突变为天冬酰胺(d92n)。
在具体实施方式中,所述脂肪酶突变体,是以扩展青霉脂肪酶的氨基酸序列(genbank:aag22769.1)为模板,进行了e83v和d92n的双突变,其具有seqidno:10的序列。
另一方面,本发明提供一种脂肪酶突变体,所述脂肪酶突变体以上述基因工程菌为发酵菌株进行液体发酵而制备,并包含选自seqidno:4、seqidno:8和seqidno:10的氨基酸序列。
再一方面,本发明提供一种制备碳酸二甲酯的方法,所述方法包括利用上述基因工程菌进行液体发酵生产脂肪酶突变体以催化制备碳酸二甲酯的步骤。
在具体实施方式中,所述方法包括以下步骤:
1)上述基因工程菌种子液按照接种量为培养基体积的1.6%接种于培养基,培养温度为37℃,培养基转速为220rpm,ph6.0-8.0,然后加入诱导剂iptg至终浓度0.2mmol/l,30℃培养7h后,8000rpm,4℃离心5min收集菌体;
2)将步骤1)得到的菌体用m9培养基重悬获得菌液;以及
3)将甲醇与碳酸丙烯酯按照摩尔比16:1的比例加入到步骤2)重悬的菌液中,55℃,反应48h。
技术效果
区别于现有的碳酸二甲酯的制备中存在的转化率低、选择性低、工艺复杂、污染环境的情况,本发明使用基因工程菌催化甲醇与碳酸丙烯酯或碳酸乙烯酯进行转酯反应制备碳酸二甲酯,具有以下优点:转化率高、选择性高、普适性高,而且工艺简单、反应条件温和、降低成本、环境友好、应用前景广阔。
附图说明
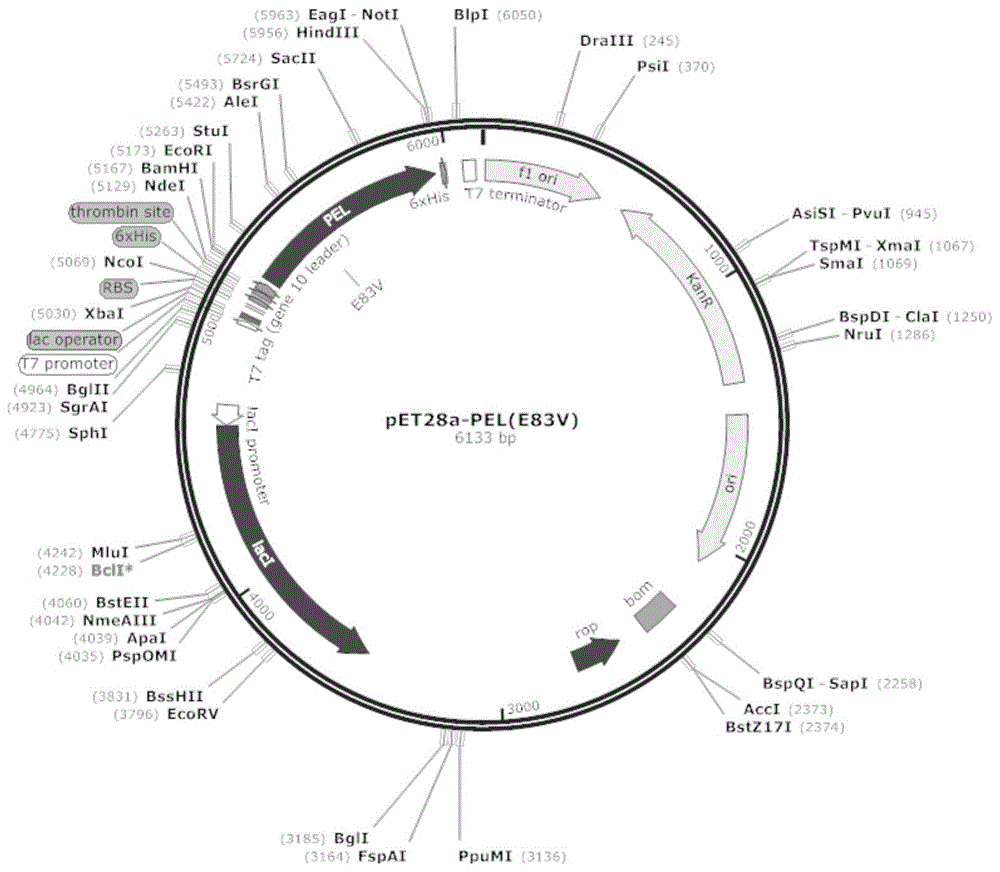
图1为本申请实施例1中的质粒pet28a-pele83v的图谱。
图2为本申请实施例3中的质粒pet28a-peld92n的图谱。
图3为本申请实施例5中的质粒pet28a-pelmut的图谱。
具体实施方式
下面将结合具体实施例对本发明的技术方案进行清楚、完整地描述,显然,所描述的实施例仅仅是本发明的一部分实施例,而不是全部的实施例。基于本发明中的实施例,本领域普通技术人员在没有做出创造性劳动前提下所获得的所有其他实施例,都属于本发明保护的范围。
实施例1:脂肪酶e83v的定点突变
脂肪酶来源于扩展青霉(penicilliumexpansum)pf898,脂肪酶的氨基酸序列编号为genbank:aag22769.1,其基因序列pel编号为genbank:af284064.1。将该基因通过全基因合成,并连接在质粒pet-28a上,酶切位点为ecori和hindiii,获得质粒pet28a-pel。以质粒pet28a-pel为模板,以下列引物进行高保真扩增,将83位氨基酸e突变为v,即将对应的核酸序列329的a突变为t。
83正向:5’gctctcatcactcctgtgctctcgggcgtgact3’(seqidno:1)
83反向:5’agtcacgcccgagagcacaggagtgatgagagc3’(seqidno:2)
98℃预变性5min;98℃变性10s;55℃退火15s;72℃延伸6min,25个循环,72℃延伸6min。琼脂糖电泳验证pcr产物大小。异丙醇沉淀回收pcr产物,dpni酶切(37℃酶切1h)去除模板,转化大肠杆菌后提取质粒送测序,验证突变成功后,保存质粒pet28a-pele83v,质粒图谱如图1,突变酶基因序列及其氨基酸序列分别为:seqidno:3和seqidno:4。
实施例2:基因工程菌bl21(de3)/pet28a-pele83v催化合成碳酸二甲酯
分别取质粒pet28a-pel、pet28a-pele83v2μl,转入感受态bl21(de3)中,所用平板抗性为kan,获得含有野生型脂肪酶的对照菌株bl21(de3)/pet28a-pel,以及含有突变脂肪酶(氨基酸序列:seqidno:4)的基因工程菌株bl21(de3)/pet28a-pele83v。
挑取菌株bl21(de3)/pet28a-pel、bl21(de3)/pet28a-pele83v的单克隆,置于含20ml(含50μg/mlkan)的液体lb培养基的50ml摇瓶中,置于37℃摇床,200rpm,过夜培养活化。分别将活化得到的菌液以2:100的接种量接入摇瓶中,摇瓶规格为500ml,含300ml液体lb培养基(含50μg/mlkan),然后放置37℃摇床,200rpm培养,当细胞od600达到0.6-0.8之间加入终浓度200μmol/l的iptg诱导。iptg诱导后,30℃、180rpm继续培养,7h后收集菌体,4℃,8000rpm,离心5min,然后用5-10mlm9培养基,分别重悬沉淀,并达到相同od600。
按甲醇与碳酸丙烯酯的摩尔比16:1的比例加入底物,然后加入重悬的菌液补齐至5ml,55℃,反应时间48h。结果表明利用bl21(de3)/pet28a-pele83v催化产生碳酸二甲酯的含量是菌株bl21(de3)/pet28a-pel的1.6倍。
实施例3:脂肪酶d92n的突变
以质粒pet28a-pel为模板,以下列引物进行高保真扩增,将92位氨基酸d突变为n,即将对应的核酸序列355的g突变为a。
92正向:5’actttcccctctaatgtgaagatcatg3’(seqidno:5)
92反向:5’catgatcttcacattagaggggaaagt3’(seqidno:6)
98℃预变性5min;98℃变性10s;55℃退火15s;72℃延伸6min,25个循环,72℃延伸6min。琼脂糖电泳验证pcr产物大小。异丙醇沉淀回收pcr产物,dpni酶切(37℃酶切1h)去除模板,转化大肠杆菌后提取质粒送测序,验证突变成功后,保存质粒pet28a-peld92n,其质粒图谱如图2,突变酶基因序列及其氨基酸序列分别为:seqidno:7和seqidno:8。
实施例4:基因工程菌bl21(de3)/pet28a-peld92n催化合成碳酸二甲酯
分别取质粒pet28a-pel、pet28a-peld92n2μl,转入感受态bl21(de3)中,所用平板抗性为kan,获得含有野生型脂肪酶的对照菌株bl21(de3)/pet28a-pel,以及含有突变脂肪酶(氨基酸序列:seqidno:8)的基因工程菌株bl21(de3)/pet28a-peld92n。
挑取菌株bl21(de3)/pet28a-pel、bl21(de3)/pet28a-peld92n的单克隆,置于含20ml(含50μg/mlkan)的液体lb培养基的50ml摇瓶中,置于37℃摇床,200rpm,过夜培养活化。分别将活化得到的菌液以2:100的接种量接入摇瓶中,摇瓶规格为500ml,含300ml液体lb培养基(含50μg/mlkan),然后放置37℃摇床,200rpm培养,当细胞od600达到0.6-0.8之间加入终浓度200μmol/l的iptg诱导。iptg诱导后,30℃、180rpm继续培养,7h后收集菌体,4℃,8000rpm,离心5min,然后用5-10mlm9培养基,分别重悬沉淀,并达到相同od600。
按甲醇与碳酸丙烯酯的摩尔比16:1的比例加入底物,然后加入重悬的菌液补齐至5ml,55℃,反应时间48h。结果表明利用bl21(de3)/pet28a-peld92n催化产生碳酸二甲酯的含量是菌株bl21(de3)/pet28a-pel的1.9倍。
实施例5:脂肪酶e83v和d92n的突变
以质粒pet28a-pele83v为模板,以下列引物进行高保真扩增,将92位氨基酸d突变为n,即将对应的核酸序列355的g突变为a。
92正向:5’actttcccctctaatgtgaagatcatg3’(seqidno:5)
92反向:5’catgatcttcacattagaggggaaagt3’(seqidno:6)
98℃预变性5min;98℃变性10s;55℃退火15s;72℃延伸6min,25个循环,72℃延伸6min。琼脂糖电泳验证pcr产物大小。异丙醇沉淀回收pcr产物,dpni酶切(37℃酶切1h)去除模板,转化大肠杆菌后提取质粒送测序,验证突变成功后,保存质粒pet28a-pelmut其质粒图谱如图3,突变酶基因序列及其氨基酸序列分别为:seqidno:9和seqidno:10。
实施例6:基因工程菌bl21(de3)/pet28a-pelmut催化合成碳酸二甲酯
分别取质粒pet28a-pel、pet28a-pelmut2μl,转入感受态bl21(de3)中,所用平板抗性为kan,获得含有野生型脂肪酶的对照菌株bl21(de3)/pet28a-pel,以及含有突变脂肪酶(氨基酸序列:seqidno:10)的基因工程菌株bl21(de3)/pet28a-pelmut。
挑取菌株bl21(de3)/pet28a-pel、bl21(de3)/pet28a-pelmut的单克隆,置于含20ml(含50μg/mlkan)的液体lb培养基的50ml摇瓶中,置于37℃摇床,200rpm,过夜培养活化。分别将活化得到的菌液以2:100的接种量接入摇瓶中,摇瓶规格为500ml,含300ml液体lb培养基(含50μg/mlkan),然后放置37℃摇床,200rpm培养,当细胞od600达到0.6-0.8之间加入终浓度200μmol/l的iptg诱导。iptg诱导后,30℃、180rpm继续培养,7h后收集菌体,4℃,8000rpm,离心5min,然后用5-10mlm9培养基,分别重悬沉淀,并达到相同od600。
按甲醇与碳酸丙烯酯的摩尔比16:1的比例加入加入重悬的菌液补齐至5ml,55℃,反应时间48h。结果表明利用bl21(de3)/pet28a-pelmut催化产生碳酸二甲酯的含量是菌株bl21(de3)/pet28a-pel的2.9倍。
在脂肪酶催化甲醇与碳酸乙烯酯或甲醇与碳酸丙烯酯反应制备碳酯二甲酯的过程中,碳酸二甲酯含量分析方法为:取50μl反应液离心分层,取上层液样10μl,用290μl环己烷溶解,摇匀,再加入300μl十二烷(2mg/ml)作为内标;取1μl样品进样,由气相色谱测定反应物中的碳酸二甲酯的含量。转化率用山东鲁南化工仪器厂的sp-6890型气相色谱仪测定,色谱柱为se-54型。具体的测试条件为:柱室采用程序升温:100℃维持6分钟,升温速度40℃/分钟,到200℃维持15分钟,气化室320℃,检测室320℃。
从以上实验结果可以看出,本申请获得基因工程菌可以用于高效催化生产碳酸二甲酯。突变酶相较于野生型脂肪酶催化效率提高主要基于以下原因:e83v突变意味着由带负电荷的极性氨基酸突变为不带电荷的非极性氨基酸,由于缬氨酸是脂肪族非极性氨基酸可以帮助蛋白质形成更加紧密的疏水核心,使得其蛋白的包装效率更高,而且也能为蛋白质提供更多的盐桥,这些原因都使得蛋白质的稳定性和活性得到提高。同理,d92n是由带负电荷的极性氨基酸突变为不带电荷的极性氨基酸。
以上所述,仅为本发明的具体实施方式,但本发明的保护范围并不局限于此,任何熟悉本技术领域的技术人员在本发明揭露的技术范围内,可轻易想到变化或替换,都应涵盖在本发明的保护范围之内。因此,本发明的保护范围应所述以权利要求的保护范围为准。
上文中的提及的具体序列分别如下:
e83v突变酶的基因序列(seqidno:3,前后ecori和hindiii酶切位点为斜体标记,加粗为突变氨基酸的碱基)
e83v突变酶的氨基酸序列(seqidno:4,加粗和下划线标记突变氨基酸)
d92n突变酶的基因序列(seqidno:7,前后ecori和hindiii酶切位点为斜体标记,加粗为突变氨基酸的碱基)
d92n突变酶的氨基酸序列(seqidno:8,加粗和下划线标记突变氨基酸)
e83v和d92n双突变酶的基因序列(seqidno:9,加粗为突变氨基酸的碱基)
e83v和d92n双突变酶的氨基酸序列(seqidno:10,加粗和下划线标记突变氨基酸)
序列表
<110>源创核新(北京)新材料科技有限公司
<120>一种重组质粒、包含其的基因工程菌及其在制备碳酸二甲酯中的应用
<130>di20-2121-xc03
<160>10
<170>patentinversion3.5
<210>1
<211>33
<212>dna
<213>人工序列
<220>
<223>正向引物
<400>1
gctctcatcactcctgtgctctcgggcgtgact33
<210>2
<211>33
<212>dna
<213>人工序列
<220>
<223>反向引物
<400>2
agtcacgcccgagagcacaggagtgatgagagc33
<210>3
<211>789
<212>dna
<213>人工序列
<220>
<223>e83v突变酶的核苷酸序列
<400>3
gaattcgcaactgccgacgctgctgccttccctgatctgcaccgtgcagcaaagctctct60
tccgctgcctacacaggttgcatcggaaaggccttcgatgtcactatcgccaagaggatt120
tatgacctcgtgaccgacaccaatggattcgtcggatactccaccgagaagaagaccatc180
gcggtcatcatgaggggctcgactaccatcaccgacttcgtgaacgacattgacattgct240
ctcatcactcctgtgctctcgggcgtgactttcccctctgatgtgaagatcatgagaggt300
gttcacagaccttggtccgctgtacacgacaccatcattactgaagtcaaggctctcatt360
gcgaagtaccctgattacactctggaagcagtcggacattccctcggtggtgccctcaca420
tccattgcccacgttgccctggcccagaacttcccggacaagtcacttgtcagcaatgcc480
cttaacgccttccccatcggcaaccaagcgtgggccgactttggtactgcgcaggccggt540
accttcaaccgcggaaataacgttcttgacggtgtccctaacatgtactcgagcccgctt600
gttaacttcaagcactatggaaccgaatactacagctctggtaccgaggctagcaccgtg660
aagtgcgaaggccagcgtgacaagtcttgctctgccggcaatggcatgtacgctgtcact720
cccggtcacatcgccagctttggcgtcgtgatgcttactgctggttgtggctatctgagc780
tgaaagctt789
<210>4
<211>258
<212>prt
<213>人工序列
<220>
<223>e83v突变酶的氨基酸序列
<400>4
alathralaaspalaalaalapheproaspleuhisargalaalalys
151015
leuserseralaalatyrthrglycysileglylysalapheaspval
202530
thrilealalysargiletyraspleuvalthraspthrasnglyphe
354045
valglytyrserthrglulyslysthrilealavalilemetarggly
505560
serthrthrilethraspphevalasnaspileaspilealaleuile
65707580
thrprovalleuserglyvalthrpheproseraspvallysilemet
859095
argglyvalhisargprotrpseralavalhisaspthrileilethr
100105110
gluvallysalaleuilealalystyrproasptyrthrleugluala
115120125
valglyhisserleuglyglyalaleuthrserilealahisvalala
130135140
leualaglnasnpheproasplysserleuvalserasnalaleuasn
145150155160
alapheproileglyasnglnalatrpalaasppheglythralagln
165170175
alaglythrpheasnargglyasnasnvalleuaspglyvalproasn
180185190
mettyrserserproleuvalasnphelyshistyrglythrglutyr
195200205
tyrserserglythrglualaserthrvallyscysgluglyglnarg
210215220
asplyssercysseralaglyasnglymettyralavalthrprogly
225230235240
hisilealaserpheglyvalvalmetleuthralaglycysglytyr
245250255
leuser
<210>5
<211>27
<212>dna
<213>人工序列
<220>
<223>正向引物
<400>5
actttcccctctaatgtgaagatcatg27
<210>6
<211>27
<212>dna
<213>人工序列
<220>
<223>反向引物
<400>6
catgatcttcacattagaggggaaagt27
<210>7
<211>789
<212>dna
<213>人工序列
<220>
<223>d92n突变酶的核苷酸序列
<400>7
gaattcgcaactgccgacgctgctgccttccctgatctgcaccgtgcagcaaagctctct60
tccgctgcctacacaggttgcatcggaaaggccttcgatgtcactatcgccaagaggatt120
tatgacctcgtgaccgacaccaatggattcgtcggatactccaccgagaagaagaccatc180
gcggtcatcatgaggggctcgactaccatcaccgacttcgtgaacgacattgacattgct240
ctcatcactcctgagctctcgggcgtgactttcccctctaatgtgaagatcatgagaggt300
gttcacagaccttggtccgctgtacacgacaccatcattactgaagtcaaggctctcatt360
gcgaagtaccctgattacactctggaagcagtcggacattccctcggtggtgccctcaca420
tccattgcccacgttgccctggcccagaacttcccggacaagtcacttgtcagcaatgcc480
cttaacgccttccccatcggcaaccaagcgtgggccgactttggtactgcgcaggccggt540
accttcaaccgcggaaataacgttcttgacggtgtccctaacatgtactcgagcccgctt600
gttaacttcaagcactatggaaccgaatactacagctctggtaccgaggctagcaccgtg660
aagtgcgaaggccagcgtgacaagtcttgctctgccggcaatggcatgtacgctgtcact720
cccggtcacatcgccagctttggcgtcgtgatgcttactgctggttgtggctatctgagc780
tgaaagctt789
<210>8
<211>258
<212>prt
<213>人工序列
<220>
<223>d92n突变酶的氨基酸序列
<400>8
alathralaaspalaalaalapheproaspleuhisargalaalalys
151015
leuserseralaalatyrthrglycysileglylysalapheaspval
202530
thrilealalysargiletyraspleuvalthraspthrasnglyphe
354045
valglytyrserthrglulyslysthrilealavalilemetarggly
505560
serthrthrilethraspphevalasnaspileaspilealaleuile
65707580
thrprogluleuserglyvalthrpheproserasnvallysilemet
859095
argglyvalhisargprotrpseralavalhisaspthrileilethr
100105110
gluvallysalaleuilealalystyrproasptyrthrleugluala
115120125
valglyhisserleuglyglyalaleuthrserilealahisvalala
130135140
leualaglnasnpheproasplysserleuvalserasnalaleuasn
145150155160
alapheproileglyasnglnalatrpalaasppheglythralagln
165170175
alaglythrpheasnargglyasnasnvalleuaspglyvalproasn
180185190
mettyrserserproleuvalasnphelyshistyrglythrglutyr
195200205
tyrserserglythrglualaserthrvallyscysgluglyglnarg
210215220
asplyssercysseralaglyasnglymettyralavalthrprogly
225230235240
hisilealaserpheglyvalvalmetleuthralaglycysglytyr
245250255
leuser
<210>9
<211>789
<212>dna
<213>人工序列
<220>
<223>e83v和d92n突变酶的核苷酸序列
<400>9
gaattcgcaactgccgacgctgctgccttccctgatctgcaccgtgcagcaaagctctct60
tccgctgcctacacaggttgcatcggaaaggccttcgatgtcactatcgccaagaggatt120
tatgacctcgtgaccgacaccaatggattcgtcggatactccaccgagaagaagaccatc180
gcggtcatcatgaggggctcgactaccatcaccgacttcgtgaacgacattgacattgct240
ctcatcactcctgtgctctcgggcgtgactttcccctctaatgtgaagatcatgagaggt300
gttcacagaccttggtccgctgtacacgacaccatcattactgaagtcaaggctctcatt360
gcgaagtaccctgattacactctggaagcagtcggacattccctcggtggtgccctcaca420
tccattgcccacgttgccctggcccagaacttcccggacaagtcacttgtcagcaatgcc480
cttaacgccttccccatcggcaaccaagcgtgggccgactttggtactgcgcaggccggt540
accttcaaccgcggaaataacgttcttgacggtgtccctaacatgtactcgagcccgctt600
gttaacttcaagcactatggaaccgaatactacagctctggtaccgaggctagcaccgtg660
aagtgcgaaggccagcgtgacaagtcttgctctgccggcaatggcatgtacgctgtcact720
cccggtcacatcgccagctttggcgtcgtgatgcttactgctggttgtggctatctgagc780
tgaaagctt789
<210>10
<211>258
<212>prt
<213>人工序列
<220>
<223>e83v和d92n突变酶的氨基酸序列
<400>10
alathralaaspalaalaalapheproaspleuhisargalaalalys
151015
leuserseralaalatyrthrglycysileglylysalapheaspval
202530
thrilealalysargiletyraspleuvalthraspthrasnglyphe
354045
valglytyrserthrglulyslysthrilealavalilemetarggly
505560
serthrthrilethraspphevalasnaspileaspilealaleuile
65707580
thrprovalleuserglyvalthrpheproserasnvallysilemet
859095
argglyvalhisargprotrpseralavalhisaspthrileilethr
100105110
gluvallysalaleuilealalystyrproasptyrthrleugluala
115120125
valglyhisserleuglyglyalaleuthrserilealahisvalala
130135140
leualaglnasnpheproasplysserleuvalserasnalaleuasn
145150155160
alapheproileglyasnglnalatrpalaasppheglythralagln
165170175
alaglythrpheasnargglyasnasnvalleuaspglyvalproasn
180185190
mettyrserserproleuvalasnphelyshistyrglythrglutyr
195200205
tyrserserglythrglualaserthrvallyscysgluglyglnarg
210215220
asplyssercysseralaglyasnglymettyralavalthrprogly
225230235240
hisilealaserpheglyvalvalmetleuthralaglycysglytyr
245250255
leuser
- 还没有人留言评论。精彩留言会获得点赞!