一种靶向人程序性死亡配体1(PD-L1)的单可变域抗体及其衍生物的制作方法
一种靶向人程序性死亡配体1(pd
‑
l1)的单可变域抗体及其衍生物
1.本专利申请要求于2020年4月22日提交的申请号为cn 202010324761.8的中国发明专利申请的优先权权益,在此将其全部内容引入作为参考。
技术领域
2.本发明涉及抗体药物领域,具体而言,本发明涉及靶向人程序性死亡配体1(pd
‑
l1)的单可变域抗体、其衍生蛋白及其用于制备药物的用途,特别是在治疗和/或预防、或诊断pd
‑
l1相关疾病例如肿瘤中的用途。
背景技术:
3.pd
‑
1及其配体pd
‑
l1是肿瘤免疫的重要靶点。pd
‑
1、pd
‑
l1是一对免疫抑制分子,是免疫系统防止自身免疫过激的重要组成部分,其通路的激活具有抑制肿瘤免疫应答、诱导肿瘤特异性t细胞凋亡的作用,与肿瘤发展关系密切。pd
‑
1(cd279)是一种i型跨膜蛋白,属于免疫球蛋白超家族成员,主要表达于活化的cd4+t细胞、cd8+t细胞及b细胞等免疫细胞上。其配体pd
‑
l1(又名b7
‑
h1,cd274)属于b7家族成员,在肿瘤浸润免疫细胞(tic),以及多种恶性肿瘤细胞中高表达,如恶性黑色素瘤、非小细胞肺癌、头颈鳞癌等。利用单克隆抗体阻断pd
‑
1、pd
‑
l1通路治疗肿瘤,目前在临床上显示出良好的疗效和安全性,已有多种抗体类药物获批上市,适应症包括黑色素瘤、非小细胞肺癌、晚期肾细胞癌等多种恶性肿瘤,同时很多正在进行的临床试验尝试开发更多新的适应症。
4.虽然pd
‑
1/pd
‑
l1单抗在各种恶性肿瘤临床治疗中显示出良好治疗效果,但是存在用药量大、总体反应率低等问题。其原因主要包括pd
‑
l1表达量低、肿瘤微环境中t细胞耗竭等。因此仍有必要对pd
‑
1/pd
‑
l1靶点进一步深度开发,研制具有更好临床效果的治疗药物。在该靶点的药物研发方面,通过以下途径可能会增加临床患者的获益。(1)进一步开发亲和力更高,活性更好的抗体分子;(2)基于该靶点的双特异抗体或类似物;(3)初步研究表明pd
‑
l1阳性肿瘤患者对pd
‑
l1抑制剂的应答率远高于pd
‑
l1阴性肿瘤患者,因此需要有效的生物标志物来预测pd
‑
l1阳性肿瘤患者或提前筛选患者,以降低治疗费用和可能的严重不良反应;(4)为了提高应答率,联合用药成为肿瘤免疫治疗的发展趋势,如与其他肿瘤免疫药物的联用,与靶向药物、化疗或放疗的联用。
5.单可变域抗体是目前最小的抗体分子,其分子量是普通抗体的1/10,最初由比利时科学家hamers,r在骆驼血液中发现,它是工程化抗体产品中备受关注的一类。单可变域抗体除具备单克隆抗体的抗原反应性外,还拥有一些独特的功能特性,如分子质量小,稳定性强、可溶性好、易表达、靶向性强、人源化简单等,尤其是适合进行双/多特异治疗性抗体的开发和car
‑
t/m/nk等疗法的开发。目前单可变域抗体和/或基于单可变域抗体的双/多特异性抗体开发已成为研发热点。国际上ablynx公司在单可变域抗体领域做了广泛布局。由该公司研发的caplacizumab已于2019年2月被fda批准用于罕见病
‑‑
获得性血栓性血小板减少性紫癜(attp)的治疗。获得性血栓性血小板减少性紫癜特征是小血管中血液凝固过
多,caplacizumab是首个被批准用于该疾病的药物,也是第一种靶向血管性血友病因子(von willebrand factor,vwf)的药物,vwf是血液凝固级联中的关键蛋白。同时,caplacizumab也是fda批准的第一个单可变域抗体,caplacizumab的获批是单可变域抗体这一药物领域正式走上人类疾病治疗舞台的一件标志性事件。中国也有企业在积极开发单可变域抗体,其中包括深圳国创单可变域抗体技术公司、深圳普瑞金生物药业有限公司、苏州博生吉(安科)公司以及苏州康宁杰瑞公司等。其中国内康宁杰瑞最先步入单可变域抗体领域,其开发的pd
‑
l1单可变域抗体(kn035,皮下给药)于2016年获cfda和fda批准进入临床试验,并于2017年6月底获得日本药品监督管理部门(pmda)临床批件。kn035是全球首个pd
‑
l1单可变域抗体。
6.目前pd
‑
1/pd
‑
l1单抗在各种恶性肿瘤临床治疗中显示出良好治疗效果,但是存在用药量大、总体反应率低等问题。其原因主要包括pd
‑
l1表达量低、肿瘤微环境中t细胞耗竭等。因此,需要发现新的抗pd
‑
l1抗体类药物。在候选分子开发方面,通过以下途径可能会获得治疗效果更优的抗体分子:(1)进一步开发亲和力更高,活性更好的抗体分子;(2)基于该靶点的双特异抗体或类似物;(3)开发更有效的诊断抗体,通过检测pd
‑
l1表达预测pd
‑
l1阳性肿瘤患者或提前筛选患者,以降低治疗费用和可能的严重不良反应。骆驼来源的单可变域抗体以其独特的性质,预期可以用来很好的解决以上问题。
技术实现要素:
7.为解决上述问题,本公开提供一种靶向人程序性死亡配体1(pd
‑
l1)的单可变域抗体及其衍生物。由人pd
‑
l1免疫骆驼提取pbmc构建噬菌体表面展示vhh抗体库,筛选鉴定获得抗人pd
‑
l1特异性单可变域抗体2
‑
2f2,在此基础上制备嵌合抗体chf2和人源化改造的抗体hzf2突变体。所述hzf2突变体的亲和力能够达到甚至超过初始单可变域抗体2
‑
2f2,在体外能够阻断pd
‑
1和pd
‑
l1的结合,在荷瘤小鼠体内试验中能够抑制肿瘤的生长。具体而言:
8.第一方面,本发明提供一种抗pd
‑
l1单可变域抗体,其特征在于所述单可变域抗体可变区中的cdr1
‑
cdr3分别如seq id no:43
‑
45所示。
9.进一步,本发明所述的抗pd
‑
l1单可变域抗体,其特征在于所述单可变域抗体不具有恒定区或具有1
‑
3个重链恒定区。
10.进一步,本发明所述抗pd
‑
l1单可变域抗体,其特征在于所述单可变域抗体可变区的氨基酸序列如seq id no:1所示。
11.第二方面,本发明提供一种抗pd
‑
l1单可变域抗体,其特征在于所述单可变域抗体为人
‑
骆驼嵌合单可变域抗体,包括本发明第一方面所述单可变域抗体的可变区和人重链恒定区。
12.进一步,本发明所述的抗pd
‑
l1单可变域抗体,其特征在于所述嵌合单可变域抗体具有seq id no:3所示的氨基酸序列。
13.第三方面,本发明提供一种抗pd
‑
l1单可变域抗体,其特征在于所述单可变域抗体为人源化单可变域抗体,其可变区是在本发明第一方面所述单可变域抗体可变区的基础上进行了人源化改造获得。
14.进一步,本发明所述的抗pd
‑
l1单可变域抗体,其特征在于所述单可变域抗体的可变区具有seq id no:7所示的氨基酸序列。
15.进一步,本发明所述的抗pd
‑
l1单可变域抗体,其特征在于所述单可变域抗体具有seq id no:9所示的氨基酸序列。
16.第四方面,本发明提供一种抗pd
‑
l1单可变域抗体,其特征在于所述单可变域抗体为突变的抗pd
‑
l1人源化单可变域抗体,其是在本发明第三方面所述抗pd
‑
l1单可变域抗体的基础上对可变区cdrs进行1个、2个、3个或4个氨基酸残基的突变产生的;所述突变的抗pd
‑
l1人源化单可变域抗体至少部分的保留了对pd
‑
l1的特异性结合能力。
17.进一步,本发明所述的抗pd
‑
l1单可变域抗体,其特征在于其可变区选自由seq id no:11
‑
26组成的组。
18.第五方面,本发明提供一种组合物,其包含一种或多种抗pd
‑
l1,所述一种或多种抗pd
‑
l1选自由本发明第一方面至第四方面任一所述抗pd
‑
l1单可变域抗体组成的组。
19.进一步,本发明所述组合物,其特征在于还包含药学上可接受的载体,并作为药物组合物使用,优选所述药物组合物为水剂、针剂、粉针剂。
20.第六方面,本发明提供抗体或其片段在制备治疗异常增殖性疾病药物中的应用,其特征在于:所述抗体选自由本发明第一方面至第四方面任一所述抗pd
‑
l1单可变域抗体组成的组。
21.进一步,本发明所述的应用,其特征在于所述异常增殖性疾病包括肿瘤,优选黑色素瘤、非小细胞肺癌、头颈鳞癌、肾癌、结肠癌等。
22.第七方面,本发明提供抗体或其片段在制备多特异性抗体或抗体靶向药物中的应用,其特征在于:所述抗体选自由本发明第一方面至第四方面任一所述抗pd
‑
l1单可变域抗体组成的组。
23.第八方面,本发明提供一种多核苷酸,其编码本发明第一方面至第四方面任一所述抗pd
‑
l1单可变域抗体。
24.第九方面,本发明提供一种载体,其包含本发明第八方面所述的多核苷酸。
25.第十方面,本发明提供一种宿主细胞,其包含本发明第七方面所述的多核苷酸或本发明第八方面所述的载体。
26.第十一方面,本发明提供一种制备抗pd
‑
l1单可变域抗体的方法,其包括以下步骤:
27.(1)在适合表达重组抗pd
‑
l1单可变域抗体的条件下培养本发明第十方面所述宿主细胞;
28.(2)从细胞培养物中分离纯化抗pd
‑
l1单可变域抗体。
29.除非另外指出,无论在本文中用于指代重链抗体或指代常规4
‑
链抗体,术语“免疫球蛋白序列”均用作通用术语,包括全尺寸抗体,其单独的链,以及其所有部分、结构域或片段(分别包括但不限于抗原结合结构域或片段如vhh结构域或vh/vl结构域)。此外,如在本文中所使用的术语“序列”(例如在如“免疫球蛋白序列”、“抗体序列”、“可变结构域序列”、“vhh序列”或“蛋白质序列”的术语中)通常应被理解为包括相关氨基酸序列以及编码其的核酸或核苷酸序列二者,除非上下文需要更有限制性的解释。
30.可以使用免疫球蛋白单可变结构域作为用于制备含有可以充当结合单元(即,针对相同靶标的相同或不同表位和/或针对一个或多个不同靶标)的一个或多个另外的免疫球蛋白单可变结构域的多肽的“结合单元”、“结合结构域”或“构建单元”(这些术语可互换
使用)。
31.可与“单可变结构域”(“svd”)互换使用的术语“免疫球蛋白单可变结构域”(“isvd”)定义了其中抗原结合位点存在于单一免疫球蛋白结构域上并且由单一免疫球蛋白结构域形成的分子。这使得免疫球蛋白单可变结构域与“常规”免疫球蛋白或它们的片段不同,其中两个免疫球蛋白结构域可变结构域,尤其是两个可变结构域,相互作用以形成抗原结合位点。通常,在常规免疫球蛋白中,重链可变结构域(vh)和轻链可变结构域(vl)相互作用以形成抗原结合位点。在这种情况中,vh和vl二者的互补决定区(cdr)将会有利于抗原结合位点,即在抗原结合位点形成中将会涉及总计6个cdr。
32.相比之下,免疫球蛋白单可变结构域的结合位点由单一vh或vl结构域形成。因此,免疫球蛋白单可变结构域的抗原结合位点由不多于三个cdr形成。
33.术语“免疫球蛋白单可变结构域”和“单可变结构域”因此不包括需要至少两个可变结构域相互作用以形成抗原结合位点的常规免疫球蛋白或它们的片段。然而,这些术语包括其中通过单可变结构域形成抗原结合位点的常规免疫球蛋白的片段。
34.通常,单可变结构域将会是基本上由4个构架区(分别为fr1至fr4)和3个互补决定区(分别为cdr1至cdr3)组成的氨基酸序列。这样的单可变结构域和片段是最优选的,从而它们包含免疫球蛋白折叠或者能够在适合条件下形成免疫球蛋白折叠。因此,单可变结构域可以例如包含轻链可变结构域序列(例如,vl序列)或其适合的片段;或重链可变结构域序列(例如,vh序列或vhh序列)或其适合的片段;只要其能够形成单一抗原结合单元(即,基本上由单可变结构域组成的功能性抗原结合单元,以使得单一抗原结合单元不需要与另一个可变结构域相互作用以形成功能性抗原结合单元,例如对于在例如常规抗体和scfv片段中存在的、需要例如通过vh/vl相互作用而与另一个可变结构域相互作用以形成功能性抗原结合结构域的可变结构域来说是这样的情况)。
35.在本发明的一个实施方案中,免疫球蛋白单可变结构域是轻链可变结构域序列(例如,vl序列)或重链可变结构域序列(例如,vh序列);更具体地,免疫球蛋白单可变结构域可以是来源于常规四链抗体的重链可变结构域序列或来源于重链抗体的重链可变结构域序列。
36.例如,单可变结构域或免疫球蛋白单可变结构域(或适合用作免疫球蛋白单可变结构域的氨基酸)可以是(单)结构域抗体(或适合用作(单)结构域抗体的氨基酸)、“dab”或dab(或适合用作dab的氨基酸)或纳米抗体(如在本文中定义的,并且包括但不限于vhh);其他单可变结构域,或它们中任一种的任何适合的片段。
37.对于(单)结构域抗体的概括描述,还参考在本文中引用的现有技术以及参考ep0368684。用于术语“dab”,例如参考ward等人,1989(nature341:544
‑
546),参考holt等人,2003(trends biotechnol.21:484
‑
490);以及参考例如wo04/068820、wo 06/030220、wo06/003388、wo 06/059108、wo 07/049017、wo 07/085815和domantis ltd.的其他公布的专利申请。还应当指出,尽管因为它们不是哺乳动物来源的而在本发明上下文中较不优选,单可变结构域可以来源于鲨鱼的某些种(例如,所谓的“ignar结构域”,参见例如wo 05/18629)。
38.尤其是,免疫球蛋白单可变结构域可以是(如在本文中定义的)或其适合的片段。[注释:是埃博灵克斯股
份有限公司(ablynx n.v.)的注册商标]对于纳米抗体的概括描述参考以下的进一步描述,以及参考在本文中引用的现有技术,如,例如,在wo 08/020079(第16页)中描述的。
[0039]
对于vhh和纳米抗体的进一步描述,参考muyldermans 2001(分子生物技术中的综述(reviews in molecular biotechnology)74:277
‑
302)的综述文章,以及参考作为一般背景技术提及的以下专利申请:vrijeuniversiteit brussel的wo94/04678、wo 95/04079和wo 96/34103;unilever的wo 94/25591、wo99/37681、wo 00/40968、wo 00/43507、wo00/65057、wo 01/40310、wo01/44301、ep 1134231和wo 02/48193;vlaams instituutvoor biotechnologie(vib)的wo 97/49805、wo 01/21817、wo 03/035694、wo 03/054016和wo03/055527;algonomics n.v.和埃博灵克斯股份有限公司的wo 03/050531;加拿大国立研究委员会(national research council of canada)的wo 01/90190;抗体研究所(institute of antibodies)的wo 03/025020;以及埃博灵克斯股份有限公司的wo 04/041867、wo 04/041862、wo 04/041865、wo 04/041863、wo04/062551、wo 05/044858、wo06/40153、wo 06/079372、wo 06/122786、wo 06/122787和wo 06/122825和埃博灵克斯股份有限公司的另外的公布的专利申请。还参考在这些申请中提及的另外的现有技术,并且尤其参考在国际申请wo 06/040153的第41
‑
43页提及的参考文献清单,该清单和参考文献通过引用结合在本文中。如在这些参考文献中描述的,纳米抗体(尤其是vhh序列和部分人源化的纳米抗体)的特征可以尤其在于在一个或多个构架序列中存在一个或多个“标志残基”。可以例如在wo 08/101985和wo 08/142164中找到纳米抗体的进一步描述,包括纳米抗体的人源化和/或骆驼源化,以及其他修饰、部分或片段、衍生物或“纳米抗体融合体”、多价构建体(包括接头序列的一些非限制性实例)和增加纳米抗体半衰期的不同修饰和它们的制备。
[0040]
因此,在本发明的意义中,术语“免疫球蛋白单可变结构域”或“单可变结构域”包括来源于非人来源、优选骆驼科动物、优选骆驼科动物重链抗体的多肽。如之前描述的,它们可以是人源化的。此外,该术语包括来源于非骆驼科动物来源例如小鼠或人的已经“骆驼源化”的多肽,例如在davies和riechmann 1994(febs 339:285
‑
290)、1995(biotechnol.13:475
‑
479)、1996(prot.eng.9:531
‑
537)和riechmann和muyldermans 1999(j.immunol.methods231:25
‑
38)中描述的。
[0041]
术语“免疫球蛋白单可变结构域”包括不同来源的免疫球蛋白序列,包括小鼠、大鼠、兔、驴、人和骆驼科动物免疫球蛋白序列。其还包括完全人的、人源化的或嵌合的免疫球蛋白序列。例如,它包括骆驼科动物免疫球蛋白序列和人源化骆驼科动物免疫球蛋白序列,或骆驼源化免疫球蛋白单可变结构域,例如由ward等人,1989描述的骆驼源化dab(参见例如wo 94/04678以及davies和riechmann 1994、1995和1996)和骆驼源化vh。
[0042]
同样地,这样的免疫球蛋白单可变结构域可以以任何适合的方式来源于任何适合的来源,并且例如可以是自然存在的vhh序列(即,来自适合的骆驼科动物物种)或合成或半合成氨基酸序列,包括但不限于部分或完全“人源化”的vhh、“骆驼源化”的免疫球蛋白序列(并且尤其是骆驼源化vh),以及通过以下技术获得的纳米抗体和/或vhh:如亲和力成熟(例如,从合成的、随机的或自然存在的免疫球蛋白序列如vhh序列开始)、cdr移植、镶盖术(veneering)、组合来源于不同免疫球蛋白序列的片段、使用重叠引物的pcr装配、和技术人员公知的用于改造免疫球蛋白序列的类似技术;或前述中任一项的任何适合的组合。
[0043]
免疫球蛋白单可变结构域的氨基酸序列和结构可以被认为——然而不限于——
由四个构架区或“fr”组成,其在本领域中和在本文中分别被称为“构架区1”或“fr1”;“构架区2”或“fr2”;“构架区3”或“fr3”;以及“构架区4”或“fr4”;所述构架区被三个互补决定区或“cdr”隔开,其在本领域中分别被称为“互补决定区1”或“cdr1”;“互补决定区2”或“cdr2”;以及“互补决定区3”或“cdr3”。
[0044]
在免疫球蛋白单可变结构域中的氨基酸残基的总数量可以在110
‑
120的范围内,优选为112
‑
115,并且最优选113。
[0045]
如在wo 08/020079(通过引用结合在本文中)的第58和59页的q)段中进一步描述的,根据由kabat等人给出的针对vh结构域的通用编号(“kabat编号”)(“免疫学目标蛋白序列(sequence of proteins ofimmunological interest)”,美国公众健康服务中心(uspublic healthservices),nih bethesda,md,公布号91)对免疫球蛋白单可变结构域的氨基酸残基进行编号,如在riechmann和muyldermans 2000(j.immunol.methods 240:185
‑
195;参见例如该出版物的图2)的文章中应用于骆驼科动物的vhh结构域的,并且相应地,免疫球蛋白单可变结构域的fr1包含在1
‑
30位的氨基酸残基,免疫球蛋白单可变结构域的cdr1包含在31
‑
35位的氨基酸残基,免疫球蛋白单可变结构域的fr2包含在36
‑
49位的氨基酸,免疫球蛋白单可变结构域的cdr2包含在50
‑
65位的氨基酸残基,免疫球蛋白单可变结构域的fr3包含在66
‑
94位的氨基酸残基,免疫球蛋白单可变结构域的cdr3包含在95
‑
102位的氨基酸残基,并且免疫球蛋白单可变结构域的fr4包含在103
‑
113位的氨基酸残基。
[0046]
基于在本文中以及在wo 08/020079中、在wo 06/040153中和在另外的关于免疫球蛋白单可变结构域的在其中引用的参考文献中给出的免疫球蛋白单可变结构域序列的实例,将会清楚的是,氨基酸残基的精确数量还将会取决于在免疫球蛋白单可变结构域中存在的特定cdr的长度。就cdr而言,如在本领域中公知的,存在多种定义并且描述vh或vhh片段的cdr的习惯,如kabat定义(其基于序列可变性并且是最常用的)和chothia定义(其基于结构环区域的位置)。例如,参考网站http://www.bioinf.org.uk/abs/。对于本说明书和权利要求的目的来说,即使也可以提及根据kabat的cdr,最优选基于abm定义(其基于oxfordmolecular的abm抗体建模软件)来定义cdr,因为认为这是在kabat和chothia定义之间的最佳折衷。再次参考网站http://www.bioinf.org.uk/abs/)。
[0047]
在一个实施方案中,fr4包含c端氨基酸序列vtvss,即109、110、111、112和113位中的每一个。本发明还包括在109、110、111或112位终止的isvd。在本发明的一个方面中,fr4以c端氨基酸序列vtvs(109
‑
112位)终止,fr4以c端氨基酸序列vtv(109
‑
111位)终止,fr4以c端氨基酸序列vt(109
‑
110位)终止,或者fr4以c端氨基酸v(109位)终止。c端延伸可以存在于最后一个(位于最c端)的isvd的fr4的最后一个氨基酸残基例如v109、t110、v111、s112或s113的c端,其中本发明的半胱氨酸部分优选存在于或者位于c端延伸的c末端。在一个实施方案中,fr4包含c端氨基酸序列vtvss并且c端延伸是半胱氨酸(例如,本发明的多肽以vtvssc终止)。在一个实施方案中,fr4包含c端氨基酸序列vtvs并且c端延伸是半胱氨酸(例如,本发明的多肽以vtvsc终止)。在一个实施方案中,fr4包含c端氨基酸序列vtv并且c端延伸是半胱氨酸(例如,本发明的多肽以vtvc终止)。在一个实施方案中,fr4包含c端氨基酸序列vt并且c端延伸是半胱氨酸(例如,本发明的多肽以vtc终止)。在一个实施方案中,fr4包含c端氨基酸v并且c端延伸是半胱氨酸(例如,本发明的多肽以vc终止)。
[0048]
在一个实施方案中,本发明涉及如在本文中所描述的二聚体,其中isvd是轻链可
变结构域序列(vl),是重链可变结构域序列(vh),来源于常规四链抗体或来源于重链抗体。
[0049]
在一个实施方案中,本发明涉及如在本文中所描述的二聚体,其中所述isvd选自由下列各项组成的组:单一结构域抗体、结构域抗体、适合用作单一结构域抗体的氨基酸序列、适合用作结构域抗体的氨基酸序列、dab、适合用作dab的氨基酸序列、纳米抗体、vhh、人源化vhh、和骆驼源化vh。优选地,isvd包含100至140个之间的氨基酸,如110
‑
130个之间的氨基酸。
[0050]
与现有技术相比,本发明的技术方案具有以下优点:
[0051]
第一,本发明抗体为具有高亲和力的抗pd
‑
l1人源化单可变域抗体。人源化抗pd
‑
l1单可变域抗体hzf2高亲和力特异结合人pd
‑
l1蛋白,其亲和力(kd)为1.1nm,与对照抗体kn035相当。亲和力高特异性好的基本特性为hzf2对pd
‑
1/pd
‑
l1信号通路的抑制作用提供了理论依据;而单可变域抗体与传统的单克隆抗体相比,应用形式更加灵活,更适合进行双/多特异治疗性抗体的开发。
[0052]
第二,本发明抗体具有良好的生物学活性。hzf2可以有效的结合细胞表面重组表达的人pd
‑
l1,与重组表达了人pd
‑
l1的cho细胞(cho
‑
pd
‑
l1)结合的ec50为1.01nm;可以有效阻断重组人pd
‑
l1与其受体pd
‑
1的结合作用,ic50为4.3nm;通过使用jurkat
‑
pd1
‑
nfat细胞和cho
‑
pd
‑
l1
‑
cd3l细胞报告基因方法检测hzf2对pd
‑
1/pd
‑
l1信号通路的阻断,结果活性ec50为5.45nm;hzf2具有良好的体内稳定性,并且在免疫系统人源化异种移植皮下黑色素瘤a375模型中可以有效抑制肿瘤生长。
[0053]
第三,本发明在hzf2的基础上提供了多个突变体。部分突变体在特异性、亲和力等性能参数上表现出优于初始抗体hzf2的效果,这些不同的单可变域抗体突变体在基于pd
‑
l1的肿瘤检测、靶向治疗、药物递送等方面提供了更多的选择,进一步丰富和拓展了hzf2的应用潜力。
附图说明
[0054]
通过阅读下文优选实施方式的详细描述,各种其他的优点和益处对于本领域普通技术人员将变得清楚明了。附图仅用于示出优选实施方式的目的,而并不认为是对本发明的限制。而且在整个附图中,用相同的参考符号表示相同的部件。在附图中:
[0055]
图1:elisa检测单可变域抗体对人pd
‑
l1与其受体pd
‑
1结合的抑制作用。
[0056]
图2:facs分析chf2与细胞表面抗原的结合活性。
[0057]
图3:elisa分析chf2与重组pd
‑
l1结合特异性。
[0058]
图4:chf2对细胞表面pd
‑
l1结合特异性分析。
[0059]
图5:hzf2对人pd
‑
l1与其受体pd
‑
1结合的抑制作用分析结果图。
[0060]
图6:报告基因系统评估抗pd
‑
l1vhh抗体fc融合蛋白的体外阻断活性。
[0061]
图7:hzf2在balb/c小鼠体内单次给药药
‑
时曲线。
[0062]
图8:hzf2对人pd
‑
l1转基因小鼠皮下同种移植mc38
‑
hpdl1鼠结肠癌肿瘤模型的抗肿瘤药效实验结果图(肿瘤体积)。
[0063]
图9:hzf2对人pd
‑
l1转基因小鼠皮下同种移植mc38
‑
hpdl1鼠结肠癌肿瘤模型的抗肿瘤药效实验结果图(肿瘤重量)。
具体实施方式
[0064]
下面将参照附图更详细地描述本公开的示例性实施方式。虽然附图中显示了本公开的示例性实施方式,然而应当理解,可以以各种形式实现本公开而不应被这里阐述的实施方式所限制。相反,提供这些实施方式是为了能够更透彻地理解本公开,并且能够将本公开的范围完整的传达给本领域的技术人员。
[0065]
实施例1:噬菌体展示骆驼单可变域抗体免疫库构建
[0066]
利用抗原免疫骆驼,分离外周血单核细胞(pbmc)并提取总rna进行反转录,以反转录产物为模板扩增单可变域抗体重链可变区(variable domain of the heavy
‑
chain of heavychain antibody,vhh)并连入噬菌体展示载体,电转入大肠杆菌tg1感受态细胞,构建骆驼免疫库。骆驼免疫两周一次,共4次。每次注射0.8mg pd
‑
l1胞外区重组蛋白(自主表达纯化,基因序列号:np_054862.1,19aa
‑
238aa),佐以弗氏不完全佐剂(sigma,cat:f5506
‑
10ml),采取皮下多点注射的方式。每次免疫后2周采集1ml血分离血清,用免疫原作为测定抗原,通过elisa法分别测定血清中全抗体(igg)和单可变域抗体(heavy chain antibody,hcab)的滴度。当血清滴度达到建库要求后,采集100ml骆驼外周血并用分离试剂盒(天津灏洋,cat:tbd2011cm)分离pbmc,提取pbmc的总rna反转获得cdna,作为后续扩增vhh片段的模板。根据相关文献和数据库中检索到驼源vhh抗体的基因,设计并合成vhh抗体库构建引物,pcr扩增出抗体可变区基因序列。随后利用核酸内切酶对载体和扩增的抗体片段进行酶切。采用t4连接酶的连接方式构建连接产物,利用电转技术将连接产物转入到tg1菌种之中。最终构建了一个1.8
×
108的骆驼抗人pd
‑
l1 vhh抗体免疫库,用于特异性抗人pd
‑
l1单可变域抗体的筛选。为检测文库的正确率,随机选取50个克隆进行菌落pcr,结果显示插入率已达到100%。
[0067]
实施例2:抗人pd
‑
l1特异性单可变域抗体筛选
[0068]
通过固相筛选的方法对所构建的骆驼免疫库进行筛选,获得特异性噬菌体展示单可变域抗体。
[0069]
(1)原始库呈现。骆驼免疫库转接至含有氨苄青霉素和四环素的2yt培养基,培养至对数生长期,加入m13辅助噬菌体,然后加入卡那霉素,在较低温度条件下呈现过夜。第二天收取培养上清,peg沉淀对噬菌体进行浓缩,获得高滴度的抗体库呈现产物用于后续筛选。
[0070]
(2)筛选。采用固相法进行特异性抗体的筛选。将特异性抗原包被在免疫管表面,用封闭剂分别封闭免疫管和抗体库后,将抗体库加至免疫管中孵育,然后进行反复洗涤,最后用ph2.2的酸进行洗脱,洗脱物中和至中性后与对数生长期的xl
‑
blue混合孵育进行感染,进一步进行噬菌体展示,回收特异性噬菌体颗粒。筛选2
‑
3轮后进行单克隆鉴定。
[0071]
(3)鉴定。感染回收特异性噬菌体颗粒的xl
‑
blue涂布平板,长出克隆后进行单克隆鉴定。挑取单克隆培养至对数增长期,加入m13辅助噬菌体感染,然后加入卡那霉素,在30℃培养过夜。第二天取上清,加入包被了pd
‑
l1的酶联板中进行elisa反应。反应呈阳性的克隆提取噬粒(含有抗体基因的噬菌体展示载体)进行测序,确定vhh抗体基因序列。通过筛选获得了5株可结合人pd
‑
l1重组蛋白的噬菌体展示的单可变域抗体(vhh):1
‑
4g1,1
‑
6c4,2
‑
3d6,2
‑
5b7,2
‑
2f2。
[0072]
实施例3:抗人pd
‑
l1特异性单可变域抗体初步鉴定
[0073]
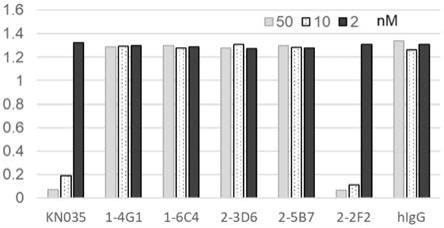
将获得的5株单域vhh抗体在大肠杆菌tg1中诱导表达,诱导条件为1mm iptg、30℃、150rpm过夜培养。诱导表达的细菌样品通过超声破碎处理并过滤,然后使用镍柱进行亲和纯化,并进行超滤处理,获得单域vhh抗体。随后通过elisa检测单域对人pd
‑
l1与其受体pd
‑
1结合的抑制作用。具体的,将人pd
‑
1胞外区和人fc的融合蛋白(pd
‑1‑
hfc,pd
‑
1序列号:np_005009.2,21aa
‑
167aa),浓度0.5μg/ml,4℃包被过夜,用5%bsa于37℃恒温培养箱封闭60min。将单域vhh抗体(浓度为50、10、2nm)和1μg/ml的pd
‑
l1
‑
mfc共孵育,37℃恒温培养箱反应60min。pbst洗板4次;然后加入1:5000稀释的hrp
‑
anti
‑
mouse fc(jackson immuno research,cat:115
‑
035
‑
071),反应45min,加入tmb(北京泰天河生物,cat:me142)底物显色15min,2m hcl终止后读板。以630nm为参比波长,读取并记录波长450nm下孔板的吸光度值a450nm
‑
630nm。结果显示2
‑
2f2可以有效阻断重组人pd
‑
l1与其受体pd
‑
1的结合作用。结果显示,2
‑
2f2具有良好的阻断活性(图1)。选取该分子作为后续开发的初始分子,简写为vhh
‑
f2。该单可变域抗体可变区氨基酸序列见seq id no.1,可变区核苷酸序列见seq id no.2。
[0074]
设计特异性引物,以阳性克隆噬粒为模板,pcr获得驼源抗体vhh
‑
f2的可变区基因,通过酶切连接,将可变区基因克隆入含有人fc(igg1,hfc)编码基因的真核表达载体上。获得序列正确的表达质粒后,转染293f细胞进行瞬时表达,通过protein a纯化,最终获得人
‑
骆驼嵌合单可变域抗体融合蛋白(vhh
‑
f2
‑
human
‑
fc chimeric antibody,简称chf2)。chf2抗体分子全长氨基酸序列见seq id no.3,核苷酸序列见:seq id no.4。
[0075]
参照who公布的envafolimab抗体序列(who drug information,vol.33,no.3,2019,page634
‑
635,envafolimab),全合成了kn035的可变区基因,kn035可变区氨基酸序列见seq id no.5,核苷酸序列见seq id no.6。并利用上述相同的策略构建chf2相同形式的kn035的fc融合蛋白,简称kn035。
[0076]
实施例4:抗人pd
‑
l1嵌合单可变域抗体结合活性分析
[0077]
方法1.bli测定结合活性
[0078]
利用fortebio公司的octet qke system仪器,采用抗人抗体fc段的捕获抗体(ahc)生物探针捕获抗体fc段的方法测定chf2结合重组抗原的能力。测定时将chf2用pbs缓冲液稀释至4μg/ml,流经ahc探针(cat:18
‑
0015,pall)表面,时间为120s。人pd
‑
l1重组蛋白作为流动相,与捕获在芯片表面的抗体作用。pd
‑
l1重组蛋白浓度为60nm。每个抗原的结合时间为300s,最后的解离时间为300s。结果显示(表1),当前实验条件下,chf2和pd
‑
l1重组蛋白具有较高亲和力,与对照抗体kn035亲和力相当。
[0079]
表1.chf2和kn035与人pd
‑
l1重组蛋白亲和力测定
[0080][0081]
方法2.facs分析结合活性
[0082]
离心收集细胞(cho
‑
pd
‑
l1
‑
cd3l细胞),将细胞分为5
×
105cells/样品/100μl,加入梯度稀释的单可变域抗体,加入的抗体终浓度为:最高浓度66nm,3倍连续稀释10个梯度。冰上孵育2h,冰冷pbs(含0.05%吐温)洗涤细胞2遍;加入fitc标记的抗人fc二抗(cat.:f9512,sigma),冰上孵育1h,冰冷pbs(含0.05%吐温)洗涤细胞2遍,重悬于200μl流式缓冲
液中,流式细胞仪(型号b49007ad,snaw31211,beckman coulter)检测细胞的平均荧光强度(mfi)。检测结果显示,chf2和kn035与表达在细胞表面的pd
‑
l1结合活性相当。其半数有效结合浓度(ec50)值分别为1.04nm、1.27nm nm(图2)。
[0083]
实施例5:抗人pd
‑
l1嵌合单可变域抗体特异性分析
[0084]
方法1.elisa鉴定嵌合抗体与重组抗原的特异性
[0085]
将人重组蛋白(pd
‑
l1、pd
‑
1、b7h3、b7h4、ctla4、cd28、icos等)用pbs稀释至1μg/ml,100μl/孔包被酶联板,4℃包被过夜;5%bsa封闭液37℃恒温培养箱封闭60min,pbst洗板3次;加入稀释至1μg/ml的chf2,37℃反应60min,pbst洗板4次;加入1:5000稀释的hrp
‑
anti
‑
human igg反应45min,pbst洗板4次;最后加入tmb底物显色,37℃恒温培养箱反应15min,2m hcl终止反应,以630nm为参比波长,读取并记录波长450nm下孔板的吸光度a450nm
‑
630nm。结果显示(图3),chf2和pd
‑
l1特异性结合,与其他重组蛋白不结合。
[0086]
方法2.bli鉴定种属特异性
[0087]
利用fortebio公司的octet qke system仪器,采用抗人抗体fc段的捕获抗体(ahc)生物探针捕获抗体fc段的方法测定chf2是否可以结合重组猴、鼠pd
‑
l1。测定时将chf2用pbs缓冲液稀释至4μg/ml,流经ahc探针(pall,cat:18
‑
0015)表面,时间为120s。猴pd
‑
l1重组蛋白、鼠pd
‑
l1重组蛋白作为流动相,与捕获在芯片表面的抗体作用。pd
‑
l1重组蛋白浓度为60nm。每个抗原的结合时间为300s,最后的解离时间为300s。结果显示,chf2和kn035均与猴pd
‑
l1重组蛋白结合,且亲和力相当;而不和鼠pd
‑
l1重组蛋白结合。(表2)
[0088]
表2.chf2和kn035与猴、鼠pd
‑
l1重组蛋白结合活性分析
[0089][0090]
方法3.facs鉴定嵌合抗体特异性
[0091]
离心收集细胞,分为3
×
105cells/样品/100μl,加入20μg/ml的单可变域抗体。冰上孵育2h,冰冷pbs(含0.05%吐温)洗涤细胞2遍;加入fitc标记的抗人fc二抗(sigma,f9512),冰上孵育1h,冰冷pbs(含0.05%吐温)洗涤细胞2遍,重悬于200μl流式缓冲液中,流式细胞仪检测。检测结果显示(图4),chf2对各肿瘤细胞的反应性与对照抗体kn035完全相同,均特异性结合表达人pd
‑
l1的细胞株,而不结合非表达细胞株。
[0092]
实施例6:chf2人源化改造
[0093]
选择可变区与骆驼抗体vhh
‑
f2可变区同源性最高的人重链可变区框架,通过cdr移植,并保留部分支撑结构氨基酸,对vhh
‑
f2的可变区进行人源化改造,设计人源化单可变域抗体融合蛋白(vhh
‑
f2
‑
human
‑
fc humanized antibody,简写为hzf2)可变区氨基酸序列如seq id no.7所示,全长氨基酸序列如seq id no.9所示。合成hzf2可变区核苷酸序列如seq id no.8所示,并构建hzf2的重组表达载体,构建的全长hzf2可变区核苷酸序列如seq id no.10所示。随后进行真核表达后用bli法测定亲和力,并进行相关分析。
[0094]
对抗体株hzf2进行进一步突变,获得从大量突变体抗体,突变序列cdr区氨基酸序
列如下表3所示,突变体可变区氨基酸序列如表4所示(seq id no.11~seq id no.26),核苷酸序列见seq id no.27~seq id no.42,部分突变体结合和解离常数变化如表5所示。
[0095]
表3.hzf2突变体cdr区氨基酸序列及亲和力变化情况
[0096][0097][0098]
表4.hzf2突变体可变区氨基酸序列
[0099]
[0100][0101]
表5.hzf2突变体与人pd
‑
l1重组蛋白亲和力测定结果
[0102][0103]
实施例7:elisa检测hzf2对人pd
‑
l1与其受体pd
‑
1结合的抑制作用
[0104]
将人pd
‑1‑
hfc(pd
‑
1序列号:np_005009.2,21aa
‑
167aa)用pbs稀释至0.5μg/ml,4
℃包被过夜,用5%bsa于37℃恒温培养箱封闭60min。梯度稀释hzf2和对照抗体kn035,以及同型对照nc
‑
higg1(起始工作浓度为50nm,1.5倍稀释10个浓度梯度),加入工作浓度为0.5μg/ml的pd
‑
l1
‑
mfc(pd
‑
l1序列号:np_054862.1,19aa
‑
238aa)共孵育,37℃恒温培养箱反应60min。pbst洗板4次;然后加入1:5000稀释的hrp
‑
anti
‑
mouse fc(cat:115
‑
035
‑
071,jackson immuno research),反应45min,加入tmb(cat:me142,北京泰天河生物)底物显色15min,2m hcl终止后读板。以630nm为参比波长,读取并记录波长450nm下孔板的吸光度值a450nm
‑
630nm。结果显示(图5),hzf2可以有效阻断重组人pd
‑
l1与其受体pd
‑
1的结合作用。其半数有效抑制浓度(ic50)为4.3nm。
[0105]
实施例8 pd
‑
1、pd
‑
l1报告基因法评价细胞阻断活性
[0106]
使用jurkat
‑
pd1
‑
nfat细胞和cho
‑
pd
‑
l1
‑
cd3l细胞报告基因方法(reporter gene assay,rga)检测hzf2对pd
‑
1和pd
‑
l1通路的阻断作用。具体如下:对数生长期的cho
‑
pd
‑
l1
‑
cd3l细胞将细胞密度调整至5
×
105个/ml,以100μl/孔铺板培养过夜。抗体样品用培养基品分步预稀释至20μg/ml,再进行2倍梯度稀释,共10个点,将稀释后样品加入到过夜培养的细胞中,50μl/孔。同时加入2
×
106个/ml浓度的jurkat
‑
pd1
‑
nfat细胞,50μl/孔。置于细胞培养箱孵育6h。提前1~2h取出适量分装bio
‑
glo
tm luciferase底物并融化,室温避光放置。从培养箱取出细胞板,平衡至室温后(约10~15min),向细胞板100μl/孔加入bio
‑
glo
tm luciferase底物,将细胞板置于微孔板恒温振荡器中,800rpm避光孵育20min。设置多功能酶标仪为luminescence模式,intergration选择500(仪器默认值),读取rlu。利用softmax软件分析数据,以样品浓度为x轴,rlu平均检测值为y轴,选择四参数方程绘制标准曲线。根据参比品和供试品曲线拟合结果的ec
50
值,计算得到供试品的相对生物学活性。结果如图6所示。hzf2对pd
‑
l1、pd
‑
1的阻断活性与kn035基本相当。ec50分别为hzf2=5.45nm;kn035=4.90nm。
[0107]
实施例9:hzf2在小鼠体内的半衰期测定
[0108]
将健康雌性5周龄裸小鼠,3只一组,经尾静脉将抗体注射进小鼠体内,给药剂量为单次单剂量15mg/kg,分别于给药后2h、4h、8h、24h、48h、96h、144h、196h尾静脉采血,收集血样并离心分离血清,
‑
20℃保存。观察其药代动力学特性。采血全部完成后,包被pd
‑
l1
‑
his(序列号:np_054862.1,19aa
‑
238aa)于96孔酶联板,0.5ug/ml,100ul/孔,4℃过夜,pbs洗板3次后,加入5%bsa pbs,37℃封闭60min,pbst洗板3次;加入待检测血清样本(10000,20000倍稀释),设置hzf2标准曲线孔(起始浓度为0.05μg/ml,2倍连续稀释,12个梯度),37℃孵育60min,pbst洗板4次;加入1:5000稀释的hrp
‑
羊抗人igg(fcr)(cat:109
‑
035
‑
098,jackson immuno research),37℃孵育40min,pbst洗板4次;加入tmb底物(cat:me142,北京泰天河生物技术有限公司)显色,37℃孵育10min后,加入2m hcl终止反应;以630nm为参比波长,读取并记录波长450nm下孔板的吸光度a450nm
‑
630nm。以标准抗体的浓度为y轴,od值为x轴,绘制时间——抗体浓度曲线图,进行线性拟合,并根据公式t
1/2
=|0.693/k|,计算药物半衰期t
1/2
。
[0109]
最终结果显示(图7),当前条件下,hzf2在小鼠体内半衰期分别为83.1小时,提示hzf2具有良好的体内半衰期和稳定性。
[0110]
实施例10:hzf2对人pd
‑
l1转基因小鼠皮下同种移植mc38
‑
hpdl1鼠结肠癌肿瘤模型的抗肿瘤药效检测
[0111]
将高表达人pdl1的鼠结肠癌肿瘤细胞mc38
‑
hpdl1接种于雌性b6
‑
hpdl1小鼠(c57来源的人pd
‑
l1转基因小鼠)右侧前胁肋部皮下,在肿瘤生长至100mm3左右时分组给予hzf2、kn035或同型对照igg,给药剂量为10mg/kg,每周2次,给药6次。每次给药的同时测量肿瘤体积及体重,记录荷瘤鼠体重和肿瘤体积的变化与给药时间的关系。实验结束时,荷瘤鼠安乐死后剥离肿瘤称重、拍照。计算治疗组与对照组相对肿瘤体积比值(t/c)和肿瘤生长抑制率(1
‑
t/c)并进行统计学分析。结果显示,受试药物hzf2有效地抑制了肿瘤的生长(图8,图9)。
[0112]
本技术相关的氨基酸和核苷酸序列如下:
[0113]
seq id no.1:骆驼来源单可变域抗体vhh
‑
f2可变区氨基酸序列
[0114]
qvqlqesgggsvqtggslrlacavsrdsddgascmgwfrqapgkgregvaiifnagertdygdsvkgrftisqdnakntlflqmnslkpedsamyycatvwcgswvarsfgqgtqvtvss
[0115]
其中:
[0116]
重链cdr1氨基酸(seq id no.43):rdsddgascmg
[0117]
重链cdr2氨基酸(seq id no.44):iifnagertdygdsvkg
[0118]
重链cdr3氨基酸(seq id no.45):vwcgswvars
[0119]
seq id no.2:骆驼来源单可变域抗体vhh
‑
f2可变区核苷酸序列
[0120]
caggtgcagctgcaggagtctggaggaggctcggtgcagactggagggtctctgagactcgcctgtgcagtctctagagacagtgacgacggtgccagctgtatggggtggttccgccaggctccagggaaggggcgcgagggggtcgcaatcatttttaatgctggtgaacgtaccgactatggcgactccgtgaagggccgattcaccatctcccaagacaacgccaagaacacgctgtttctacaaatgaacagcctgaaacctgaggacagtgccatgtactattgtgcgacagtttggtgtggttcttgggtcgcgcgttctttcggccaggggacccaggtcaccgtctcctca
[0121]
其中:
[0122]
重链cdr1核苷酸:
[0123]
agagacagtgacgacggtgccagctgtatgggg
[0124]
重链cdr2核苷酸:
[0125]
atcatttttaatgctggtgaacgtaccgactatggcgactccgtgaagggc重链cdr3核苷酸:
[0126]
gtttggtgtggttcttgggtcgcgcgttct
[0127]
seq id no.3:嵌合单可变域抗体chf2全长氨基酸序列
[0128]
qvqlqesgggsvqtggslrlacavsrdsddgascmgwfrqapgkgregvaiifnagertdygdsvkgrftisqdnakntlflqmnslkpedsamyycatvwcgswvarsfgqgtqvtvssasepkssdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsrdeltknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspg
[0129]
其中:
[0130]
重链cdr1氨基酸:rdsddgascmg
[0131]
重链cdr2氨基酸:iifnagertdygdsvkg
[0132]
重链cdr3氨基酸:vwcgswvars
[0133]
seq id no.4:嵌合单可变域抗体chf2全长核苷酸序列
[0134]
caggtgcagctgcaggagtctggaggaggctcggtgcagactggagggtctctgagactcgcctgtgca
gtctctagagacagtgacgacggtgccagctgtatggggtggttccgccaggctccagggaaggggcgcgagggggtcgcaatcatttttaatgctggtgaacgtaccgactatggcgactccgtgaagggccgattcaccatctcccaagacaacgccaagaacacgctgtttctacaaatgaacagcctgaaacctgaggacagtgccatgtactattgtgcgacagtttggtgtggttcttgggtcgcgcgttctttcggccaggggacccaggtcaccgtctcctcagctagcgagcccaaatctagcgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcctccatctcgggatgagctgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctatagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtccccgggt
[0135]
其中:
[0136]
重链cdr1核苷酸:
[0137]
agagacagtgacgacggtgccagctgtatgggg
[0138]
重链cdr2核苷酸:
[0139]
atcatttttaatgctggtgaacgtaccgactatggcgactccgtgaagggc
[0140]
重链cdr3核苷酸:
[0141]
gtttggtgtggttcttgggtcgcgcgttct
[0142]
seq id no.5:kn035可变区氨基酸序列
[0143]
qvqlvesggglvqpggslrlscaasgkmssrrcmawfrqapgkerervak
[0144]
llttsgstyladsvkgrftisrdnskntvylqmnslraedtavyycaadsfe
[0145]
dptctlvtssgafqywgqgtlvtvss
[0146]
其中:
[0147]
重链cdr1氨基酸:rrcma
[0148]
重链cdr2氨基酸:kllttsgstyladsvkg
[0149]
重链cdr3氨基酸:dsfedptctlvtssgafqy
[0150]
seq id no.6:kn035可变区核苷酸序列
[0151]
caggtgcagctggtggagtctggaggaggcctggtgcagcctggaggctccctgaggctgtcctgcgctgcctctggcaagatgtcctccagacggtgcatggcctggttccgacaggctcctggcaaggagcgagagcgggtggccaagctgctgaccacctccggctccacctacctggccgactccgtgaagggacggttcaccatctccagggacaactccaagaacaccgtgtacctgcagatgaactccctgcgagctgaggacaccgccgtgtactactgcgctgcagactccttcgaggaccccacctgcaccctggtgacctcctctggagccttccagtactggggacagggcaccctggtgaccgtgtcctcc
[0152]
其中:
[0153]
重链cdr1核苷酸:
[0154]
agacggtgcatggcc
[0155]
重链cdr2核苷酸:
[0156]
aagctgctgaccacctccggctccacctacctggccgactccgtgaaggga
[0157]
重链cdr3核苷酸:
[0158]
gactccttcgaggaccccacctgcaccctggtgacctcctctggagccttccagtac
[0159]
seq id no.7:人源化单可变域抗体hzf2可变区氨基酸序列
[0160]
evqlvesggglvqpggslrlscaasrdsddgascmgwfrqapgkglegvaiifnagertdygdsvkgrftisrdnakntlylqmnslraedtavyycatvwcgswvarswgqgtlvtvss
[0161]
其中:
[0162]
重链cdr1氨基酸:rdsddgascmg
[0163]
重链cdr2氨基酸:iifnagertdygdsvkg
[0164]
重链cdr3氨基酸:vwcgswvars
[0165]
seq id no.8:人源化单可变域抗体hzf2可变区核苷酸序列
[0166]
gaggtgcagctggtggagtctggaggtggcctggtgcagcctggaggctccctgaggctgtcctgcgctgcctctcgggactccgacgacggagcctcctgcatgggctggttcaggcaggctcctggcaagggcctggagggagtggccatcatcttcaacgctggcgagcggaccgactacggcgactccgtgaagggacggttcaccatctccagggacaacgccaagaacaccctgtacctgcagatgaactccctgagagccgaggacacagccgtgtactactgcgctaccgtgtggtgtggctcctgggtggctcggtcctggggacagggcaccctggtgaccgtgtcctcc
[0167]
其中:
[0168]
重链cdr1核苷酸:
[0169]
cgggactccgacgacggagcctcctgcatgggc
[0170]
重链cdr2核苷酸:
[0171]
atcatcttcaacgctggcgagcggaccgactacggcgactccgtgaaggga
[0172]
重链cdr3核苷酸:
[0173]
gtgtggtgtggctcctgggtggctcggtcc
[0174]
seq id no.9:人源化单可变域抗体hzf2全长氨基酸序列
[0175]
evqlvesggglvqpggslrlscaasrdsddgascmgwfrqapgkglegvaiifnagertdygdsvkgrftisrdnakntlylqmnslraedtavyycatvwcgswvarswgqgtlvtvssasepkssdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsrdeltknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspg
[0176]
其中:
[0177]
重链cdr1氨基酸:rdsddgascmg
[0178]
重链cdr2氨基酸:iifnagertdygdsvkg
[0179]
重链cdr3氨基酸:vwcgswvars
[0180]
seq id no.10:人源化单可变域抗体hzf2全长核苷酸酸序列
[0181]
gaggtgcagctggtggagtctggaggtggcctggtgcagcctggaggctccctgaggctgtcctgcgctgcctctcgggactccgacgacggagcctcctgcatgggctggttcaggcaggctcctggcaagggcctggagggagtggccatcatcttcaacgctggcgagcggaccgactacggcgactccgtgaagggacggttcaccatctccagggacaacgccaagaacaccctgtacctgcagatgaactccctgagagccgaggacacagccgtgtactactgcgctaccgtgtggtgtggctcctgggtggctcggtcctggggacagggcaccctggtgaccgtgtcctccgctagcgagcccaaatc
tagcgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcctccatctcgggatgagctgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctatagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtccccgggt
[0182]
其中:
[0183]
重链cdr1核苷酸:
[0184]
cgggactccgacgacggagcctcctgcatgggc
[0185]
重链cdr2核苷酸:
[0186]
atcatcttcaacgctggcgagcggaccgactacggcgactccgtgaaggga
[0187]
重链cdr3核苷酸:
[0188]
gtgtggtgtggctcctgggtggctcggtcc
[0189]
seq id no.11:人源化单可变域抗体hzf2
‑
m1可变区氨基酸序列
[0190]
evqlvesggglvqpggslrlscaasrdsddgassmgwfrqapgkglegvaiifnagertdygdsvkgrftisrdnakntlylqmnslraedtavyycatvwsgswvarswgqgtlvtvss
[0191]
seq id no.12:人源化单可变域抗体hzf2
‑
m2可变区氨基酸序列
[0192]
evqlvesggglvqpggslrlscaasgdsddgascmgwfrqapgkglegvaiifnagertdygdsvkgrftisrdnakntlylqmnslraedtavyycatvwcgswvarswgqgtlvtvss
[0193]
seq id no.13:人源化单可变域抗体hzf2
‑
m3可变区氨基酸序列
[0194]
evqlvesggglvqpggslrlscaasrdsssgascmgwfrqapgkglegvaiifnagertdygdsvkgrftisrdnakntlylqmnslraedtavyycatvwcgswvarswgqgtlvtvss
[0195]
seq id no.14:人源化单可变域抗体hzf2
‑
m4可变区氨基酸序列
[0196]
evqlvesggglvqpggslrlscaasrdsndgascmgwfrqapgkglegvaiifnagertdygdsvkgrftisrdnakntlylqmnslraedtavyycatvwcgswvarswgqgtlvtvss
[0197]
seq id no.15:人源化单可变域抗体hzf2
‑
m5可变区氨基酸序列
[0198]
evqlvesggglvqpggslrlscaasrdsddaascmgwfrqapgkglegvaiifnagertdygdsvkgrftisrdnakntlylqmnslraedtavyycatvwcgswvarswgqgtlvtvss
[0199]
seq id no.16:人源化单可变域抗体hzf2
‑
m6可变区氨基酸序列
[0200]
evqlvesggglvqpggslrlscaasrdsdegascmgwfrqapgkglegvaiifnagertdygdsvkgrftisrdnakntlylqmnslraedtavyycatvwcgswvarswgqgtlvtvss
[0201]
seq id no.17:人源化单可变域抗体hzf2
‑
m7可变区氨基酸序列
[0202]
evqlvesggglvqpggslrlscaasrdsddsascmgwfrqapgkglegvaiifnagertdygdsvkgrftisrdnakntlylqmnslraedtavyycatvwcgswvarswgqgtlvtvss
[0203]
seq id no.18:人源化单可变域抗体hzf2
‑
m8可变区氨基酸序列
[0204]
evqlvesggglvqpggslrlscaasrdsddgascmgwfrqapgkeregvaiifnvgertdygdsvkgrf
tisrdnakntlylqmnslraedtavyycatvwcgswvarswgqgtlvtvss
[0205]
seq id no.19:人源化单可变域抗体hzf2
‑
m9可变区氨基酸序列
[0206]
evqlvesggglvqpggslrlscaasrdsdegascmgwfrqapgkeregvaiifnagertdygdsvkgrftisrdnakntlylqmnslraedtavyycatvwcgswvarswgqgtlvtvss
[0207]
seq id no.20:人源化单可变域抗体hzf2
‑
m10可变区氨基酸序列
[0208]
evqlvesggglvqpggslrlscaasrdsddgascmgwfrqapgkeregvaiifnagertdygdsvkgrftisrdnakntlylqmnslraedtavyycatvycgswvarswgqgtlvtvss
[0209]
seq id no.21:人源化单可变域抗体hzf2
‑
m11可变区氨基酸序列
[0210]
evqlvesggglvqpggslrlscaasrdsddgascmgwfrqapgkeregvaiifnagertdygdsvkgrftisrdnakntlylqmnslraedtavyycatvycgsyvarswgqgtlvtvss
[0211]
seq id no.22:人源化单可变域抗体hzf2
‑
m12可变区氨基酸序列
[0212]
evqlvesggglvqpggslrlscaasrdsdegascmgwfrqapgkeregvaiifnvgertdygdsvkgrftisrdnakntlylqmnslraedtavyycatvwcgswvarswgqgtlvtvss
[0213]
seq id no.23:人源化单可变域抗体hzf2
‑
m13可变区氨基酸序列
[0214]
evqlvesggglvqpggslrlscaasrdsddgascmgwfrqapgkeregvaiifnagertdygdsvkgrftisrdnakntlylqmnslraedtavyycatvfcgsfvarswgqgtlvtvss
[0215]
seq id no.24:人源化单可变域抗体hzf2
‑
m14可变区氨基酸序列
[0216]
evqlvesggglvqpggslrlscaasrdsdegascmgwfrqapgkeregvaiifnvgertdygdsvkgrftisrdnakntlylqmnslraedtavyycatvycgsyvarswgqgtlvtvss
[0217]
seq id no.25:人源化单可变域抗体hzf2
‑
m15可变区氨基酸序列
[0218]
evqlvesggglvqpggslrlscaasrdsdegascmgwfrqapgkeregvaiifnvgertdygdsvkgrftisrdnakntlylqmnslraedtavyycatvfcgsyvarswgqgtlvtvss
[0219]
seq id no.26:人源化单可变域抗体hzf2
‑
m16可变区氨基酸序列
[0220]
evqlvesggglvqpggslrlscaasrdsdegascmgwfrqapgkeregvaiifnvgertdygdsvkgrftisrdnakntlylqmnslraedtavyycatvfcgsfvarswgqgtlvtvss
[0221]
seq id no.27:人源化单可变域抗体hzf2
‑
m1可变区核苷酸序列
[0222]
gaggtgcagctggtggagtctggaggtggcctggtgcagcctggaggctccctgaggctgtcctgcgctgcctctcgggactccgacgacggagcctccagcatgggctggttcaggcaggctcctggcaagggcctggagggagtggccatcatcttcaacgctggcgagcggaccgactacggcgactccgtgaagggacggttcaccatctccagggacaacgccaagaacaccctgtacctgcagatgaactccctgagagccgaggacacagccgtgtactactgcgctaccgtgtggtccggctcctgggtggctcggtcctggggacagggcaccctggtgaccgtgtcctcc
[0223]
seq id no.28:人源化单可变域抗体hzf2
‑
m2可变区核苷酸序列
[0224]
gaggtgcagctggtggagtctggaggtggcctggtgcagcctggaggctccctgaggctgtcctgcgctgcctctggagactccgacgacggagcctcctgcatgggctggttcaggcaggctcctggcaagggcctggagggagtggccatcatcttcaacgctggcgagcggaccgactacggcgactccgtgaagggacggttcaccatctccagggacaacgccaagaacaccctgtacctgcagatgaactccctgagagccgaggacacagccgtgtactactgcgctaccgtgtggtgtggctcctgggtggctcggtcctggggacagggcaccctggtgaccgtgtcctcc
[0225]
seq id no.29:人源化单可变域抗体hzf2
‑
m3可变区核苷酸序列
[0226]
gaggtgcagctggtggagtctggaggtggcctggtgcagcctggaggctccctgaggctgtcctgcgct
gcctctcgggactccagcagcggagcctcctgcatgggctggttcaggcaggctcctggcaagggcctggagggagtggccatcatcttcaacgctggcgagcggaccgactacggcgactccgtgaagggacggttcaccatctccagggacaacgccaagaacaccctgtacctgcagatgaactccctgagagccgaggacacagccgtgtactactgcgctaccgtgtggtgtggctcctgggtggctcggtcctggggacagggcaccctggtgaccgtgtcctcc
[0227]
seq id no.30:人源化单可变域抗体hzf2
‑
m4可变区核苷酸序列
[0228]
gaggtgcagctggtggagtctggaggtggcctggtgcagcctggaggctccctgaggctgtcctgcgctgcctctcgggactccaacgacggagcctcctgcatgggctggttcaggcaggctcctggcaagggcctggagggagtggccatcatcttcaacgctggcgagcggaccgactacggcgactccgtgaagggacggttcaccatctccagggacaacgccaagaacaccctgtacctgcagatgaactccctgagagccgaggacacagccgtgtactactgcgctaccgtgtggtgtggctcctgggtggctcggtcctggggacagggcaccctggtgaccgtgtcctcc
[0229]
seq id no.31:人源化单可变域抗体hzf2
‑
m5可变区核苷酸序列
[0230]
gaggtgcagctggtggagtctggaggtggcctggtgcagcctggaggctccctgaggctgtcctgcgctgcctctcgggactccgacgacgctgcctcctgcatgggctggttcaggcaggctcctggcaagggcctggagggagtggccatcatcttcaacgctggcgagcggaccgactacggcgactccgtgaagggacggttcaccatctccagggacaacgccaagaacaccctgtacctgcagatgaactccctgagagccgaggacacagccgtgtactactgcgctaccgtgtggtgtggctcctgggtggctcggtcctggggacagggcaccctggtgaccgtgtcctcc
[0231]
seq id no.32:人源化单可变域抗体hzf2
‑
m6可变区核苷酸序列
[0232]
gaggtgcagctggtggagtctggaggtggcctggtgcagcctggaggctccctgaggctgtcctgcgctgcctctcgggactccgacgagggagcctcctgcatgggctggttcaggcaggctcctggcaagggcctggagggagtggccatcatcttcaacgctggcgagcggaccgactacggcgactccgtgaagggacggttcaccatctccagggacaacgccaagaacaccctgtacctgcagatgaactccctgagagccgaggacacagccgtgtactactgcgctaccgtgtggtgtggctcctgggtggctcggtcctggggacagggcaccctggtgaccgtgtcctcc
[0233]
seq id no.33:人源化单可变域抗体hzf2
‑
m7可变区核苷酸序列
[0234]
gaggtgcagctggtggagtctggaggtggcctggtgcagcctggaggctccctgaggctgtcctgcgctgcctctcgggactccgacgactctgcctcctgcatgggctggttcaggcaggctcctggcaagggcctggagggagtggccatcatcttcaacgctggcgagcggaccgactacggcgactccgtgaagggacggttcaccatctccagggacaacgccaagaacaccctgtacctgcagatgaactccctgagagccgaggacacagccgtgtactactgcgctaccgtgtggtgtggctcctgggtggctcggtcctggggacagggcaccctggtgaccgtgtcctcc
[0235]
seq id no.34:人源化单可变域抗体hzf2
‑
m8可变区核苷酸序列
[0236]
gaggtgcagctggtggagtctggaggtggcctggtgcagcctggaggctccctgaggctgtcctgcgctgcctctcgggactccgacgacggagcctcctgcatgggctggttcaggcaggctcctggcaaggagagagagggagtggccatcatcttcaacgtgggcgagcggaccgactacggcgactccgtgaagggacggttcaccatctccagggacaacgccaagaacaccctgtacctgcagatgaactccctgagagccgaggacacagccgtgtactactgcgctaccgtgtggtgtggctcctgggtggctcggtcctggggacagggcaccctggtgaccgtgtcctcc
[0237]
seq id no.35:人源化单可变域抗体hzf2
‑
m9可变区核苷酸序列
[0238]
gaggtgcagctggtggagtctggaggtggcctggtgcagcctggaggctccctgaggctgtcctgcgctgcctctcgggactccgacgagggagcctcctgcatgggctggttcaggcaggctcctggcaaggagagagagggagtggccatcatcttcaacgctggcgagcggaccgactacggcgactccgtgaagggacggttcaccatctccagggacaacgccaagaacaccctgtacctgcagatgaactccctgagagccgaggacacagccgtgtactactgcgctaccgtg
tggtgtggctcctgggtggctcggtcctggggacagggcaccctggtgaccgtgtcctcc
[0239]
seq id no.36:人源化单可变域抗体hzf2
‑
m10可变区核苷酸序列
[0240]
gaggtgcagctggtggagtctggaggtggcctggtgcagcctggaggctccctgaggctgtcctgcgctgcctctcgggactccgacgacggagcctcctgcatgggctggttcaggcaggctcctggcaaggagagagagggagtggccatcatcttcaacgctggcgagcggaccgactacggcgactccgtgaagggacggttcaccatctccagggacaacgccaagaacaccctgtacctgcagatgaactccctgagagccgaggacacagccgtgtactactgcgctaccgtgtactgtggctcctgggtggctcggtcctggggacagggcaccctggtgaccgtgtcctcc
[0241]
seq id no.37:人源化单可变域抗体hzf2
‑
m11可变区核苷酸序列
[0242]
gaggtgcagctggtggagtctggaggtggcctggtgcagcctggaggctccctgaggctgtcctgcgctgcctctcgggactccgacgacggagcctcctgcatgggctggttcaggcaggctcctggcaaggagagagagggagtggccatcatcttcaacgctggcgagcggaccgactacggcgactccgtgaagggacggttcaccatctccagggacaacgccaagaacaccctgtacctgcagatgaactccctgagagccgaggacacagccgtgtactactgcgctaccgtgtactgtggctcctacgtggctcggtcctggggacagggcaccctggtgaccgtgtcctcc
[0243]
seq id no.38:人源化单可变域抗体hzf2
‑
m12可变区核苷酸序列
[0244]
gaggtgcagctggtggagtctggaggtggcctggtgcagcctggaggctccctgaggctgtcctgcgctgcctctcgggactccgacgagggagcctcctgcatgggctggttcaggcaggctcctggcaaggagagagagggagtggccatcatcttcaacgtgggcgagcggaccgactacggcgactccgtgaagggacggttcaccatctccagggacaacgccaagaacaccctgtacctgcagatgaactccctgagagccgaggacacagccgtgtactactgcgctaccgtgtggtgtggctcctgggtggctcggtcctggggacagggcaccctggtgaccgtgtcctcc
[0245]
seq id no.39:人源化单可变域抗体hzf2
‑
m13可变区核苷酸序列
[0246]
gaggtgcagctggtggagtctggaggtggcctggtgcagcctggaggctccctgaggctgtcctgcgctgcctctcgggactccgacgacggagcctcctgcatgggctggttcaggcaggctcctggcaaggagagagagggagtggccatcatcttcaacgctggcgagcggaccgactacggcgactccgtgaagggacggttcaccatctccagggacaacgccaagaacaccctgtacctgcagatgaactccctgagagccgaggacacagccgtgtactactgcgctaccgtgttctgtggctccttcgtggctcggtcctggggacagggcaccctggtgaccgtgtcctcc
[0247]
seq id no.40:人源化单可变域抗体hzf2
‑
m14可变区核苷酸序列
[0248]
gaggtgcagctggtggagtctggaggtggcctggtgcagcctggaggctccctgaggctgtcctgcgctgcctctcgggactccgacgagggagcctcctgcatgggctggttcaggcaggctcctggcaaggagagagagggagtggccatcatcttcaacgtgggcgagcggaccgactacggcgactccgtgaagggacggttcaccatctccagggacaacgccaagaacaccctgtacctgcagatgaactccctgagagccgaggacacagccgtgtactactgcgctaccgtgtactgtggctcctacgtggctcggtcctggggacagggcaccctggtgaccgtgtcctcc
[0249]
seq id no.41:人源化单可变域抗体hzf2
‑
m15可变区核苷酸序列
[0250]
gaggtgcagctggtggagtctggaggtggcctggtgcagcctggaggctccctgaggctgtcctgcgctgcctctcgggactccgacgagggagcctcctgcatgggctggttcaggcaggctcctggcaaggagagagagggagtggccatcatcttcaacgtgggcgagcggaccgactacggcgactccgtgaagggacggttcaccatctccagggacaacgccaagaacaccctgtacctgcagatgaactccctgagagccgaggacacagccgtgtactactgcgctaccgtgttctgtggctcctacgtggctcggtcctggggacagggcaccctggtgaccgtgtcctcc
[0251]
seq id no.42:人源化单可变域抗体hzf2
‑
m16可变区核苷酸序列
[0252]
gaggtgcagctggtggagtctggaggtggcctggtgcagcctggaggctccctgaggctgtcctgcgct
gcctctcgggactccgacgagggagcctcctgcatgggctggttcaggcaggctcctggcaaggagagagagggagtggccatcatcttcaacgtgggcgagcggaccgactacggcgactccgtgaagggacggttcaccatctccagggacaacgccaagaacaccctgtacctgcagatgaactccctgagagccgaggacacagccgtgtactactgcgctaccgtgttctgtggctccttcgtggctcggtcctggggacagggcaccctggtgaccgtgtcctcc
[0253]
以上所述,仅为本发明较佳的具体实施方式,但本发明的保护范围并不局限于此,任何熟悉本技术领域的技术人员在本发明揭露的技术范围内,可轻易想到的变化或替换,都应涵盖在本发明的保护范围之内。因此,本发明的保护范围应以所述权利要求的保护范围为准。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1