新型细菌蛋白质纤维
本发明涉及用于作为生物纳米材料应用的芽孢杆菌内生孢子附属物(ena)和新型蛋白质多聚体和纤维组装体的领域。具体地,本发明涉及由含有细菌duf3992结构域的蛋白质亚基组成的自组装蛋白质,其含有保守的n-末端含半胱氨酸区域,以及涉及工程化改造的蛋白质和它们的多聚体和纤维。此外,所述自组装蛋白质亚基的重组表达提供了新型蛋白质纳米纤维和修饰的展示表面(例如,芽孢杆菌孢子)的生产方法。最后,本文描述了所述多聚体、纤维和表面在生物医学和生物技术应用中的用途。
背景技术:
1、自组装分子为控制化学功能和形态以及生物活性提供了具有挑战性的机会。蛋白质的独特特性,包括其模块化性质、生物相容性和生物降解性,为设计智能纳米材料提供了令人兴奋的机会(herrera estrada&champion,2015年;jain等人,2018年)。受自然界的启发,一些蛋白质/肽被设计成自组装成各种复杂的结构,包括纳米颗粒、囊泡、网箱和纤维组装体;这些可以被赋予新的功能,在生物工程的不同领域提供大量应用(matsuurua 2014;katyal等人,2019)。改变自组装肽和蛋白质的氨基酸序列并操纵环境参数,可以调节特性并控制自组装以根据需要获得多样化的超分子纳米结构(lombardi等人,2019)。氨基酸侧链的各种特性为它们的化学修饰提供了具有无限序列组合的可能性,以及修饰蛋白质的氨基和/或羧基末端可以将蛋白质聚合物的自组装调整为特定的纳米构造(aluri等人,2012;yu等人,1996)。因此,天然的自组装蛋白质或肽可以被工程化改造以诱导自组装以外的各种特性,包括自我修复、剪切稀化、形状记忆等(chen和zou,2019)。
2、当面临不利的生长条件时,属于厚壁菌门的细菌可以分化为代谢休眠和无生产性内生生孢子状态。由于其脱水状态和独特的多层细胞结构,这些内生孢子对环境压力源表现出极强的恢复力,甚至在其形成数百年后仍能萌发进入代谢活跃和复制的营养生长状态(setlow,2014)。通过这种方式,属于厚壁菌门的芽孢杆菌和梭状芽孢杆菌能够承受长时间的干旱、饥饿、高氧或抗生素胁迫。内生孢子通常由包含细菌dna的最内层脱水核心组成。核心被一层内膜包围,内膜被薄的肽聚糖层环绕,所述肽聚糖将作为孢子萌发过程中出现的营养细胞的细胞壁发挥功能。然后是厚的修饰肽聚糖的皮层,其对休眠至关重要(atrihand foster,1999)。皮层又被一些蛋白质包被层环绕。在一些梭状芽胞杆菌和大多数蜡样芽孢杆菌(bacillus cereus)菌群的菌种中,孢子被最外层松散配合的由(糖)蛋白和脂质组成的次晶外生孢子层包围(stewart,2015)。芽孢杆菌和梭状芽胞杆菌内生孢子的表面也可以装饰有数微米长和几纳米宽的丝状附属物,所述附属物在菌株和菌种之间显示非常大的结构多样性(hachisuka and kuno,1976;rode et al.,1971;walker et al.,2007)。蜡样芽孢杆菌广义上是一组革兰氏阳性形成内生孢子的细菌,尽管它们具有系统发育关系,但仍显示出高度的生态多样性。由于其脱水状态和独特的多层细胞结构,其内生孢子对环境压力源表现出极强的恢复力,甚至在其形成数百年后仍能萌发进入代谢活跃和复制的营养生长状态(setlow,2014)。蜡样芽孢杆菌内生孢子装饰有标识和功能未知的微米长附属物。蜡样芽孢杆菌群菌株和菌种之间内生孢子附属物(以下称为ena)的数量和形态各不相同,一些菌株甚至同时表达不同形态的ena(smirnova et al.,2013)。在营养细胞表面没有观察到类似ena的结构,表明它们代表孢子特异性的纤维。ena似乎是蜡样芽孢杆菌群菌株的孢子中普遍的性质。ankolekar等人表明,47种蜡样芽孢杆菌的食品分离株全部产生带有附属物的内生孢子(ankolekar&labbe,2010)。在12种食品源性苏云金芽孢杆菌(bacillusthuringiensis)(与蜡样芽孢杆菌密切相关,并以其杀虫活性而闻名)的产肠毒素分离株的10种的孢子上也发现了附属物(ankolekar&labbe,2010)。总而言之,这使得这些ena结构成为新的可持续生物材料工程化改造的有趣起点。值得注意的是,早在60年代就已报道了属于蜡样芽孢杆菌群的菌种中存在孢子附属物,但由于溶解和酶促消化纤维中的困难,表征其组成和遗传标识的努力失败了(gerhardt&ribi,1964;desrosier&lara,1981)。因此,对此类内生孢子附属物的结构表征是有趣的,并且是需要的,以允许设计、开发和生产新型智能生物材料,这些材料具有改进的特性,例如在恶劣环境条件下的可持续性。
技术实现思路
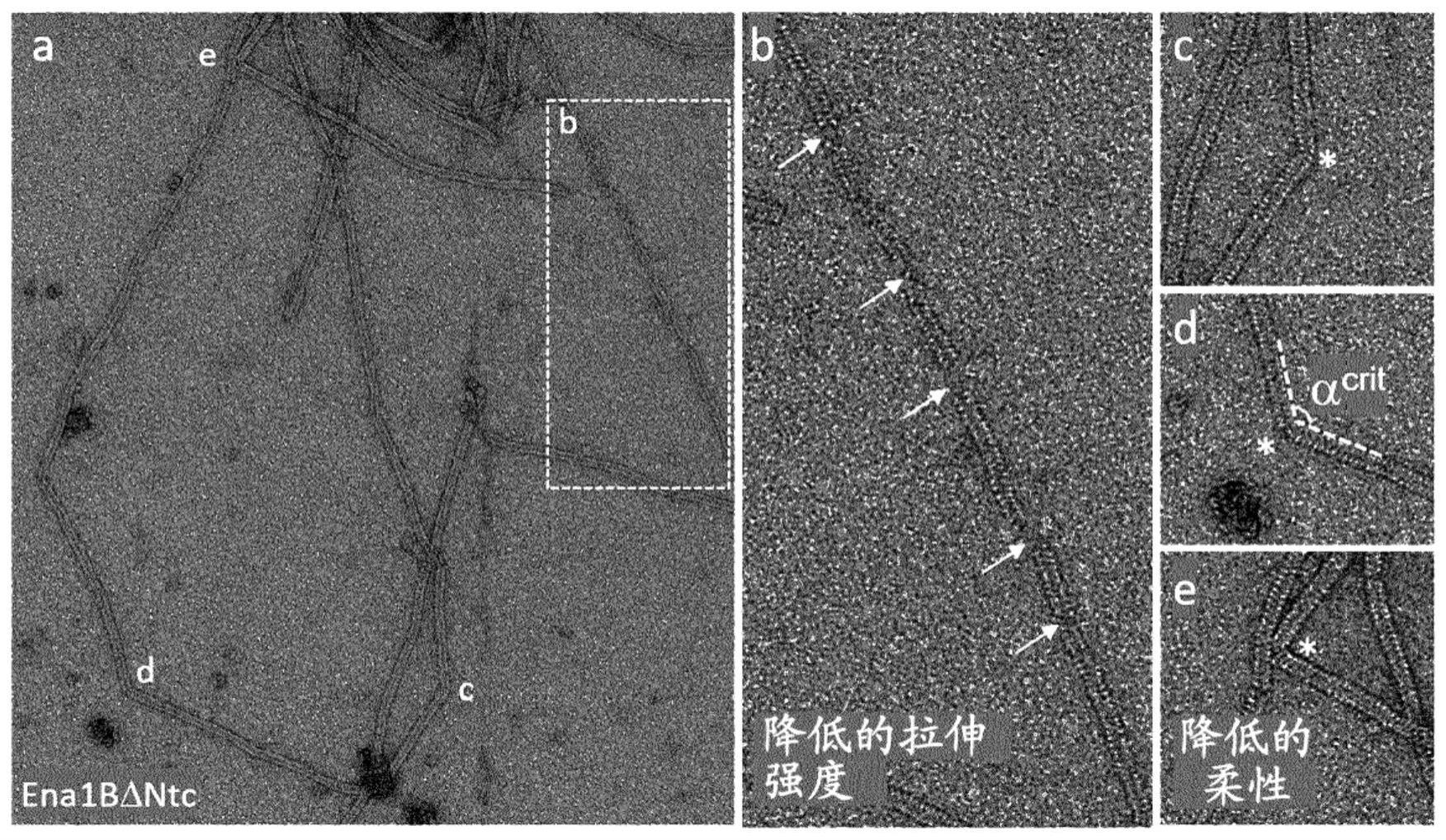
1、本发明基于从食物中毒爆发菌株蜡样芽孢杆菌nvh-0075/95中分离的内生孢子附属物(ena)的遗传和结构基础的解析,这揭示了两种主要形态的蛋白纤维,s型和l型纤维。通过使用冷冻电子显微镜和三维螺旋重建表明,芽孢杆菌内生孢子附属物(ena)形成一类新的革兰氏阳性菌毛,其特征在于具有果冻卷拓扑结构的亚基,形成通过β-片层扩张(augmentation)横向堆叠的多聚体。此外,ena纤维通过凭借其n-末端蛋白亚基肽的延伸的桥接多聚体的二硫化物交联而纵向稳定,从而产生高度耐热、耐旱和耐化学损伤的柔性菌毛(参见图2)。三维结构允许推断ena纤维由迄今为止功能未知的含有细菌duf3992结构域的蛋白质的蛋白质家族、和每个家族成员的保守n-末端区域组成,在本文中首次被注释为“ena”蛋白。通过分析缺乏编码潜在ena蛋白亚基的基因的突变体,证实了s型和l型纤维组分的遗传标识。系统发育分析表明,s型ena纤维由双顺反子操纵子编码,该操纵子独特存在于蜡样芽孢杆菌群的菌种亚组,并揭示了在不同生态型和致病型之间存在限定的ena进化枝,这些ena基因具有编码ena蛋白的共性,其特征在于具有至少两个保守半胱氨酸残基和间隔区域的n-末端区域(参见图8),然后是duf3992结构域,以允许自组装成本文定义的折叠结构,从而产生多聚或纤维组装体。在体内,在ena操纵子中编码的亚基对于ena的组装是相互依赖的。令人惊讶的是,重组表达的ena蛋白可以单独自组装成蛋白质纳米纤维,其与体内的ena具有相似的特性和结构。因此,ena代表了一类新的菌毛,其特别适应细菌孢子遇到的恶劣条件,通过揭示遗传和结构基础,本文确立了如何产生修饰的孢子、或修饰和工程化改造的ena原体或多聚体的见解,以提供适用于下一代生物材料的蛋白质组装体,例如圆盘或螺旋。
2、本发明的第一方面涉及一种具有自组装特性的蛋白质,其特征在于其氨基酸序列属于pfam13157类,即,特征在于其序列中存在duf3992结构域,并且还需要匹配ena蛋白的三维结构折叠,如本文所示,具体地是ena1b的折叠(具有seq id no:8中描述的序列),具有高度显著的相似性分数,定义为6或更高、6.5或更高、或优选n/10-4或更高的dali z分数,其中n是所述蛋白质序列的氨基酸数。在一个实施方案中,所述自组装蛋白亚基由细菌来源蛋白质提供,所述细菌来源蛋白质包含选自代表在本技术中鉴定的ena蛋白序列的seq idno:1-80、seq id no:145和seq id no:146的氨基酸序列,或与seq id no:1-80、seq idno:145或seq id no:146中的任一序列具有至少60%、或至少70%、或至少80%、或至少90%同一性的任何原核同源物,其中%同一性是在序列的全长窗口上计算的。事实上,本文描述的与本文公开的ena1b折叠匹配的结构要求通常仍然代表具有与seq id no:8的结构参考序列具有甚至低于60%同一性的同源物的细菌蛋白质,因为细菌ena家族被进一步分类在不同的成员中,如下所述。因此,一个实施方案涉及包含duf3992结构域的分离的自组装蛋白,如通过与表1所示的其隐马尔可夫模型(hidden markov model)比对所确定的,并且其中所述蛋白质亚基具有与ena1b结构匹配的三维(预测)折叠,如本文所定义的折叠相似性分数为6.5或更高,并且其中ena1b对应于seq id no:8,并且其中ena1b参考结构对应于如本文在表2中提供的坐标,并且放置于pdb7a02。
3、在一个具体的实施方案中,本文所指的自组装蛋白质涉及所述ena蛋白家族,如上文所定义,和/或由seq id no:1-80、seq id no:145或seq id no:146中描述的氨基酸序列提供,其中分别提供了代表性示例:芽孢杆菌ena1a(seq id no:1-7)、ena1b(seq id no:8-14)、ena1c(seq id no:15-20)、芽孢杆菌ena2a(seq id no:21-28、seq id no:145)、ena2b(seq id no:29-37)、ena2c(seq id no:38-48、seq id no:146),以及不同类型的其他芽孢杆菌ena3(seq id no:49-80)蛋白,或其任何一种的细菌直系同源物,所述同源物与seq idno:1-80、seq id no:145或seq id no:146所示的任意序列具有至少80%的同一性。通过图16-19中描绘的多序列比对,显示了ena家族成员的序列保守区域和水平。
4、另一个实施方案涉及如本文所述的所述自组装蛋白质,其是工程化改造的自组装蛋白质,其中如本文所述的ena折叠和hmm谱与如本文所述的ena1b折叠和duf3992谱相匹配,但其是“工程化改造的”或“修饰的”,进一步包括例如但不限于至少一种修饰,包括异源n-或c-末端标签、和/或空间阻断,蛋白质序列变体可以包含一个或多个与天然或野生型ena序列相比的突变,或可以包含肽或支架的插入、或数个氨基酸的缺失、或可以作为ena蛋白的分开部分提供,例如“分裂的”部分,在共同孵育时组装。
5、本发明的第二个方面涉及一种蛋白质多聚体,其包含或含有至少七个所述自组装蛋白质亚基,优选7个至最多十二个亚基,其是非共价连接的。更具体地,所述多聚体由七个、八个、九个、十个、十一个、十二个、13个、14个、15个、16个、17个、18个、19个、20个或更多个如本文定义的自组装ena蛋白亚基组成,其通过β-片层扩张非共价堆叠(remaut和waksman,2006中描述的蛋白质-蛋白质相互作用原理)。在一个具体的实施方案中,如本文所述的所述多聚体可进一步包含共价连接,例如其由所述多聚体的不同蛋白质亚基之间的cys连接提供(在合适的条件下)。在一个实施方案中,所述多聚体“原样”存在,即,不作为细丝或纤维群存在,因此,是非天然存在的多聚体组装体。具体地,本文定义为ena蛋白质的所述自组装蛋白质亚基还可在如本文可互换使用的其n-末端区域或n-末端连接子中包含至少两个保守的半胱氨酸残基,用于与其他多聚体形成分子间二硫键桥。在一个具体的实施方案中,多聚体组装体包含来自ena蛋白家族的七个至十二个蛋白质亚基,所述蛋白质亚基如本文进一步定义,或由seq id no:1-80、seq id no:145、或seq id no:146中描述的氨基酸序列提供,其分别提供了代表性示例:芽孢杆菌ena1a(seq id no:1-7)、ena1b(seq idno:8-14)、ena1c(seq id no:15-20)、芽孢杆菌ena2a(seq id no:21-28、seq id no:145)、ena2b(seq id no:29-37)、ena2c(seq id no:38-48、seq id no:146),以及不同类型的其他芽孢杆菌ena3(seq id no:49-80)蛋白质,或其细菌直系同源物,所述直系同源物与seqid no:1-80、seq id no:145或seq id no:146所示的任意序列具有至少80%的同一性。一个具体的实施方案涉及所述多聚体,其具有7至12个蛋白质亚基,具有与本文所述相同的自组装蛋白质。或者,多聚体包含至少7个蛋白质亚基,其中至少一个所述蛋白质亚基是工程化改造的自组装ena蛋白,如本文所定义并且涉及非天然存在的ena蛋白。在一个具体实施方案中,所述多聚体包含至少7个、优选最多12个ena蛋白亚基,其中至少一个亚基是工程化改造的ena蛋白,其在n-和/或c-末端包含空间阻断,从而防止多聚体进一步组装成纤维(图14)。在一个具体实施方案中,所述n-或c-末端空间阻断是异源n-和/或c-末端标签。在一个具体的实施方案中,形成诸如空间阻断的所述异源n-和/或c-末端标签或延伸最少为1、2、3、4、5个,优选6个或更多个氨基酸残基。某些实施方案涉及所述多聚体,其中所述ena蛋白亚基可以是相同或不同的自组装ena蛋白,其中的至少一个被工程化改造以包含异源n-和/或c-末端标签。或者,所述至少一个工程化改造的ena蛋白亚基可以是ena突变蛋白变体,或者可以是作为融合蛋白的ena蛋白,或者在暴露的环处包含插入的肽或蛋白质结构域,如图15所示例和描述,并在实施例部分概述。
6、一个具体的实施方案涉及如本文所述的所述多聚体,其是同源多聚体或异源多聚体,更具体地涉及由6个或7至12个亚基组成的多聚体,优选涉及由7个亚基组成的七聚体,或由9个亚基组成的九聚体,二者都可能形成盘状多聚体,或分别由10、11或12个亚基组成的十聚体、十一聚体或十二聚体,从而形成螺旋轮或弧的β-螺旋桨结构(图14)。
7、另一个实施方案涉及所述自组装蛋白质亚基,或自组装的含有duf3992的蛋白质亚基或ena蛋白亚基或工程化改造的ena蛋白亚基的多聚体,其包含n-末端区域或n-末端连接子(ntc)区域,其中存在氨基酸残基共有基序zxnccxmc,其中x是任何氨基酸,n是1或2,m是10-12,并且z优选是leu、ile、val或phe,并且优选地其中c-末端区域或c-末端接收区域包含共有基序gx2/3cx4y,其中g是甘氨酸,x是任何氨基酸(2或3个残基),y是酪氨酸,因此存在于蛋白质亚基的所述n-和c-末端区域基序中的半胱氨酸(c)可以形成二硫键桥,用于将一个多聚体纵向连接到另一个多聚体(最终导致组装成s纤维,如图14a;图16-17所示)。另一个替代实施方案涉及工程化改造的自组装蛋白质亚基或多聚体,其包含如本文所定义的具有基序zxnccxmc的n-末端连接区域,但具有较短的n-末端间隔区域,其中m为7至9,或具有较长的n-末端间隔区域,其中m为13-16。与其中所述间隔区域的m为10至12的多聚体组装的纤维相比,所述工程化改造的多聚体在自组装时将产生具有较低的柔性或增加的刚性的纤维。另一个替代实施方案涉及由所述ena蛋白亚基构成的所述自组装蛋白质亚基或多聚体,所述ena蛋白亚基包含n-末端区域或n-末端连接区域,其中存在氨基酸残基共有基序zxnc(c)xmc,其中x是任何氨基酸,n是1或2,m是10-12,并且z优选是leu、ile、val或phe,c是cys并且(c)是任选的cys,这意味着这些ena蛋白(最终在本文中进一步分类为ena3蛋白)的所述基序中存在一个或2个半胱氨酸,并且优选地,其中c-末端区域或c-末端接收区域包含共有基序s-z-n-y-x-b,其中z是leu或ile,b是phe或tyr,x是任何氨基酸,因此存在于蛋白质亚基的所述n-末端和c-末端区域基序中的半胱氨酸(c)可以形成二硫键桥,用于纵向连接一个多聚体与另一个多聚体(最终导致组装成l-纤维,如图14b;图19)。
8、本发明的另一方面涉及产生的包含至少两个如本文所述的所述多聚体的蛋白质纤维,其中不阻碍所述多聚体通过二硫键纵向交联,更具体地不阻碍所述多聚体通过至少一个二硫键,优选地通过两个或更多个二硫键纵向交联。所述二硫键可以在多聚体的一个或多个亚基的n-末端区域或n-末端连接子的半胱氨酸残基的侧链与多聚体(其构成纵向形成的蛋白质纤维的前一层)的一个或多个亚基的n-和/或c-末端区域中存在的一个或多个半胱氨酸残基之间形成。所述蛋白质纤维可以是重组生产的纤维。
9、在另一个实施方案中,所述蛋白质纤维是工程化改造的蛋白质纤维,其包含至少两个多聚体,其中至少一个多聚体是如本文所定义的工程化改造的多聚体,或其中至少一个多聚体包含至少一个如本文所定义的工程化改造的ena蛋白。在一个优选的实施方案中,蛋白质纤维包含多聚体,其中蛋白质亚基包含与本文所述相同的自组装蛋白质亚基,和/或由相同的ena蛋白组成。
10、本发明的另一个方面涉及包含启动子或调节序列元件的嵌合基因构建体,所述启动子或调节序列元件可操作地连接至dna元件,所述dna元件包含如本文定义的(工程化改造的)自组装蛋白质(优选ena蛋白)的编码序列。更具体地,所述编码序列可以编码蛋白质,所述蛋白质包含如seq id no:1-80;seq id no:145,或seq id no:146所示的ena蛋白,或与seq id no:1-80、seq id no:145、或seq id no:146中任意具有至少80%氨基酸同一性的包含ena1/2a、ena1/2b、ena1/2c或ena3a的所述ena家族成员的任何功能同源物,或可以编码如本文所定义的其工程化改造的ena蛋白形式。在具体的实施方案中,所述启动子或调节元件与其可操作地连接的编码序列是异源的,并且任选地是诱导型启动子,如本领域已知的。
11、另一个实施方案涉及用于表达如本文所述的嵌合基因、或用于表达如本文所述的多聚体或蛋白质组装体的自组装原体的宿主细胞。另一个实施方案涉及修饰的形成孢子的细胞或细菌,其包含如本文所述的嵌合基因,或工程化改造的ena基因或编码工程化改造的ena蛋白的基因。另一个实施方案涉及修饰的细菌孢子,特别是修饰的芽孢杆菌内生孢子,其包含和/或展示ena蛋白或其工程化改造的形式、或如本文所述的多聚体,或具有蛋白质纤维,特别是工程化改造或修饰的蛋白质纤维、重组产生的纤维或孢子,如本文所述。
12、在本发明的另一方面,提供了修饰的表面或固体支持物,所述表面包含本文所述的ena蛋白、多聚体组装体或蛋白质纤维,或其任何的工程化改造形式。所述修饰的表面通过所述ena蛋白、多聚体或纤维与所述表面共价连接组成,并且可以是细胞或人工表面,特别是任何材料类型的固体表面。因此,所述修饰的表面可用作蛋白质纤维外延生长的成核剂,例如当所述修饰的表面暴露于或接触ena蛋白溶液时,其中所述ena蛋白优选以单体或寡聚形式存在。
13、进一步的实施方案涉及蛋白质膜,其包含如本文所述的工程化改造的ena蛋白纤维和/或ena蛋白纤维,所述膜优选地是本领域已知的薄膜。或者,本文公开了一种水凝胶,其包含本文所述的工程化改造的蛋白质纤维和/或本文所述的ena蛋白纤维。另一个实施方案涉及包含工程化改造的蛋白质纤维的纳米线,所述工程化改造的蛋白质纤维被纺成更粗的线状束。
14、本发明的最后一个方面涉及重组产生如本文所述的蛋白质组装体的方法,更具体地涉及重组产生ena蛋白、多聚体和纤维状组装体、或修饰的表面(特别是如本文所述的孢子表面或合成表面)的方法。
15、一个实施方案描述了一种产生如本文所述的自组装的含有duf3992结构域的单体或多聚体的方法,所述方法包括以下步骤:
16、a)在宿主细胞中表达如本文所述的嵌合基因构建体,或使用如本文所述的宿主细胞,其中自组装的蛋白质亚基任选地包含n-和/或c-末端标签,和(任选地)
17、b)纯化自组装的含有duf3992结构域的蛋白质或多聚体,后者是在表达的蛋白质亚基寡聚化后形成的。
18、另一个实施方案提供了一种重组产生自组装的包含duf3992结构域或ena蛋白的方法,所述蛋白质在纤维组装或外延生长中被阻滞或至少受到阻碍,因此涉及一种重组产生纤维生长被阻断的工程化改造的ena蛋白的方法,包括如上所述的方法,其中n-和/或c-末端标签的长度为至少1个、优选至少6个、更优选至少9个或15个氨基酸,以空间阻断蛋白质亚基或多聚体纵向形成纤维的自组装。在进一步的实施方案中,所述n-或c-末端标签的长度为至少6个氨基酸,以可逆地阻止或阻碍纵向刚性纤维形成的蛋白质亚基或多聚体的自组装。在所述情况下,n-或c-末端标签可以是可去除的标签,例如,通过包括用于通过蛋白酶去除标签的蛋白酶识别序列,以及逆转亚基和多聚体组装的空间阻断。
19、另一个实施方案涉及生产如本文所述的蛋白质纤维的方法,包括上述方法的步骤a)和b),其中所述n-和/或c-末端标签作为可去除或可切割的标签存在,所述方法进一步包括步骤c),其中去除或切掉n-和/或c-末端标签以允许将形成的多聚体进一步自组装成蛋白质纤维。或者,步骤c)可以在纯化步骤b)之前进行。此外,提供了一种生产如本文所述的修饰的表面的方法,所述方法包括步骤a)、b)和/或c)(或反之,c)和/或b)),还包括步骤d),其中通过将(工程化改造的)ena蛋白、多聚体或纤维展示或共价连接到所述表面来修饰所述表面。
20、最后,蛋白组装体,例如本文所述的纤维,可以在细胞内产生,如重组产生ena蛋白纤维的方法中所述,包括以下步骤:
21、a)在宿主细胞中表达如本文所述的嵌合基因构建体,或使用如本文所述的宿主细胞,或表达如本文所述的ena蛋白或工程化改造的ena蛋白,其中蛋白质亚基不具有空间阻断,所以自组装的蛋白质由野生型或工程化改造的自组装ena蛋白组成,带有游离的n-末端连接子,和(可选地)
22、b)分离ena蛋白组装体,例如,分离在细胞质内表达的蛋白质亚基寡聚化后形成的纤维或多聚体。
- 还没有人留言评论。精彩留言会获得点赞!