酿酒酵母工程菌、构建方法及其应用
1.本发明涉及生物技术与水产养殖领域,特别涉及酿酒酵母工程菌、构建方法及其应用。
背景技术:
2.作为人类不可或缺的食物和经济来源,水产养殖业在世界经济体系中地位显著。中国是水产养殖大国,水产业已日渐成为我国农业经济的增长点,甲壳类则是水产动物中最大的群体。近年来甲壳类动物病害日趋严重,给我国水产养殖业带来巨大的经济损失。
3.甲壳类动物缺乏后天获得的特异性免疫功能,但是它们有比较完善的先天免疫系统,能够迅速识别和有效清除入侵的微生物。甲壳动物的先天免疫主要包括细胞免疫和体液免疫。其中,细胞免疫反应由血细胞介导,主要包括吞噬作用、结节的形成和包被作用等。而体液免疫反应主要依赖于血淋巴,产生凝集反应,酚氧化酶原的激活所导致的黑化反应,以及产生抗菌肽等。病原体被机体有效识别是启动其先天免疫反应的第一步,对无脊椎动物而言,区分自己和非己物质主要是由模式识别受体(pattern recognition receptors,prrs)完成的,它能够识别外来微生物等病原表面特异的、保守的病原相关分子模式(pathogen-associated molecular patterns,pamps),凝集素既是一种模式识别受体,它具有病原识别功能,其表面所含有特异性糖基决定簇受体,能够根据外源物表面的糖基组成来区分异己,促进机体内吞噬细胞对异物的吞噬,从而诱导机体产生有效的免疫防御应答。
4.目前对甲壳类动物免疫蛋白的研究,大多采用e.coli原核表达目的蛋白的方式,该种方式容易出现蛋白包涵体而导致活性降低,而酿酒酵母(saccharomyces cerevisiae)是一种较为优良的真核基因表达体系,其蛋白产物具有较好的表达活性和可溶性并广泛用于生物生产。然而,目前对罗氏沼虾凝集素基因在酿酒酵母中表达的报道少之又少。而作为食品级安全酿酒酵母,其在水产养殖中作用显著。首先酵母菌可以为虾蟹等水产养殖动物提供营养,促进营养物质的吸收,其次可以抑制病原菌,调节肠道微生态平衡,此外,酵母菌还可以用来调节养殖水质。因此,如何实现甲壳类动物来源的凝集素lectin的酵母活性表达并将此工程菌株作为饲料免疫菌剂用于水产养殖并能有效提高虾机体生长及免疫力是目前亟需解决的问题。
技术实现要素:
5.发明目的:本发明针对上述问题,本发明构建了酿酒酵母工程菌将罗氏沼虾(macrobrachium rosenbergii)重要模式识别受体-lectin进行了活性表达。本发明还提供了该酿酒酵母工程菌的构建方法及其应用,本发明将重组蛋白菌株作为饲料免疫菌剂投喂于罗氏沼虾,经过生长指标和免疫指标测定及病原菌侵染实验验证其有效促进了虾机体的生长及免疫力的提高。
6.技术方案:本发明所述的酿酒酵母工程菌,将如seq id no.1所示的基因片段
mrlectin克隆到表达载体中,获得重组表达载体,将重组表达载体与酿酒酵母菌株整合,获得酿酒酵母工程菌。
7.本发明采用食品级安全生物-酿酒酵母进行目的蛋白的真核表达,大大减少了包涵体的产生,实现了蛋白的活性表达,同时,本身作为水产养殖常用益生菌的酿酒酵母,活性蛋白表达菌株更易应用于水产养殖领域,对于提高虾蟹动物的机体免疫力,从而促进水产养殖绿色健康稳定发展具有重要意义。
8.作为本发明的一种优选实施方式,所述表达载体为phac181。
9.作为本发明的一种优选实施方式,重组表达载体整合至酿酒酵母菌株中gal1启动子下游。
10.本发明通过酿酒酵母真核表达系统对罗氏沼虾凝集素(lectin)基因进行表达,构建一种表达罗氏沼虾凝集素的酿酒酵母工程菌。通过将罗氏沼虾超凝集素mrlectin的cdna cds区域克隆至表达质粒phac181上得到测序正确的重组质粒phac181-mrlectin,然后设计同源重组引物,利用同源重组技术将目的基因mrlectin整合至酿酒酵母宿主菌株的gal1启动子下游。该工程菌株在半乳糖的诱导下,可活性大量表达目的蛋白mrlectin。
11.本发明所述的酿酒酵母工程菌的构建方法,包括以下步骤:
12.(1)将如seq id no.1所示的mrlectin基因片段克隆到表达质粒phac181上,得到重组表达质粒phac181-mrlectin;
13.(2)对质粒phac181-mrlectin进行扩增,得到含有mrlectin基因的核苷酸片段,将该核苷酸片段同源重组整合至酿酒酵母菌株中gal1启动子下游。
14.作为本发明的一种优选实施方式,步骤(2)中,所述扩增引物如seq id no.2和seq id no.3所示。
15.本发明还提供了一种重组质粒,所述重组质粒含有如seq id no.1所示的基因片段。
16.本发明进一步提供了所述的酿酒酵母工程菌表达蛋白的方法,包括以下步骤:将上述的酿酒酵母工程菌的菌株单菌落接种于sd-leu培养基中过夜培养,培养物第二天转接于ypg培养基中,经半乳糖诱导培养后,提取菌株蛋白。
17.本发明进一步提供了一种含有上述的酿酒酵母工程菌的饲料。
18.作为本发明的一种优选实施方式,所述酿酒酵母工程菌在饲料中的添加量为0.2~0.5%。作为本发明的一种优选实施方式,所述酿酒酵母工程菌在饲料中的添加量为0.3%。
19.本发明进一步提供了含有上述的酿酒酵母工程菌在饲料免疫菌剂中的应用。
20.本发明所述的饲料免疫菌剂可用于提高虾的免疫能力。
21.除非另有说明,本发明中“%”为质量百分比。
22.有益效果:(1)本发明构建的酿酒酵母工程菌,可以实现罗氏沼虾凝集素mrlectin的大量表达、无包涵体存在的活性表达;本发明工程菌发酵得到的mrlectin具有活力,诱导培养6h后每ml发酵液可得到0.064mg蛋白。(2)本发明构建成功的酿酒酵母工程菌mrlectin,实现了罗氏沼虾凝集素mrlectin的大量表达,将工程菌株大量培养后浓缩复配至颗粒饲料中,投喂后进行病原菌感染,发现该工程菌株可有效降低嗜水气单胞菌侵染的死亡率,与其他组别对比死亡率降低25.16%,而投喂复配该酿酒酵母工程菌株的饲料颗粒
组中的罗氏沼虾,其机体免疫指标酶活力(sod,cat,acp,akp)及免疫基因(sod,cat,acp,akp,lectin)表达量均显著高于其他组别。(3)本发明构建成功的酿酒酵母工程菌株mrlectin有效促进虾机体免疫力,目前新型饲料蛋白源与免疫饲料益生菌剂的研制与开发己成为促进现代水产养殖业健康发展的关键技术,本发明的酵母工程菌不仅可以更进一步对甲壳类动物重要免疫基因的蛋白功能进行研究,也将为开发健康安全的新型免疫饲料益生菌剂提供科学依据及新方法。
附图说明
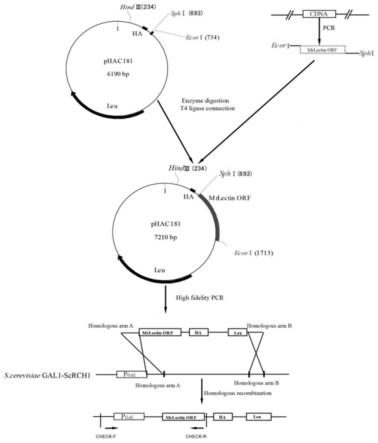
23.图1为酿酒酵母工程菌株构建策略图。
24.图2为罗氏沼虾凝集素mrlectin cdna扩增条带(990bp)。
25.图3为将罗氏沼虾凝集素mrlectin cdna克隆至载体phac181上时,菌落pcr验证阳性转化子;其中,a图中line 21为正确的转化子;b图中line 2、6、7为正确的转化子。
26.图4为将罗氏沼虾凝集素mrlectin cdna克隆至载体phac181上时,阳性转化子的酶切验证,图中line 10、12、13、14、15、16为正确的转化子;
27.图5为高保真酶primestar gxl酶对重组质粒phac181-mrlectin进行扩增(5348bp)。
28.图6为检测引物检测phac181-mrlectin与含gal1启动子的菌株同源重组,同源重组成功的条带为1276bp;其中,a图中line2为正确转化子;b图中line7、9为正确的转化子。
29.图7为western blot检测到gal-phac181-mrlectin蛋白表达,目的蛋白为42kd。
30.图8为对饲喂一个月不同饲料的罗氏沼虾进行病原菌(嗜水气单胞菌)感染实验并统计死亡率,pbs为注射对照组,结果表明,副溶血弧菌感染组中,饲喂复配有酿酒酵母-mrlectin菌株颗粒饲料的罗氏沼虾死亡率最低。
31.图9为嗜水气单胞菌后0、2、6、12、24和36小时罗氏沼虾肝胰腺中相对免疫酶活性的变化,注射pbs组为对照;其中,a图为akp酶活力变化结果图;b为acp酶活力变化结果图;c图为sod酶活力变化结果图;d图为cat酶活力变化结果图;图中bar值代表六次测定的平均值,具有标准误差,星号表示显着差异(*p《0.05,**p《0.01)。
32.图10为在副溶血性弧菌感染罗氏沼虾0、2、6、12、24和36h后,rt-pcr检测罗氏沼虾肝胰腺中免疫基因表达量,β-肌动蛋白为内参基因;其中,a图为lectin基因表达量;b图为akp基因表达量;c图为acp基因表达量;d图为sod基因表达量;e图为cat基因表达量;bar值代表六次测定的平均值,具有标准误差,星号表示显着差异(*p《0.05,**p《0.01)。
具体实施方式
33.la培养基的组成:蛋白胨10g/l、酵母提取物5g/l、nacl 10g/l、琼脂12g/l,调ph为7.0,amp抗生素100mg/ml(120℃,20min)。
34.sd-leu培养基的组成:含有酵母氮源(无aa)1.7g/l、硫酸铵5g/l、葡萄糖20g/l、10xaa mix(-ura,-leu,-his)100ml,100xura 10ml,100xhis 10ml,琼脂20g/l(120℃,20min)。
35.ypg培养基的组成:d-半乳糖20g/l、蛋白胨20g/l、酵母提取物10g/l(115℃,20min)。
36.实施例1:酿酒酵母基因工程菌的构建(基因克隆与同源重组)
37.通过构建高表达质粒phac181-mrlectin,具体方法为用trizol法提取罗氏沼虾组织rna,反转录试剂盒将rna反转录为cdna(图2)。
38.利用高保真酶将目的基因片段mrlectin进行pcr扩增,扩增出的mrlectin基因片段cds区域(不包含终止密码子)共990bp,如seq id no.1所示。然后将seq id no.1所示片段克隆到高表达质粒phac181多克隆位点上(图1),将连接载体转入大肠杆菌感受态细胞transt1中,涂布于la平板上37℃过夜培养。la平板上得到的阳性转化子进行菌落pcr及酶切验证阳性转化子(图3和图4所示),选择正确的重组子进行dna测序,验证序列没有发生突变,得到重组高表达质粒phac181-mrlectin。
39.根据构建好的重组高表达质粒phac181-mrlectin设计同源重组引物(如seq id no.2和seq id no.3所示),利用高保真primestar gxl酶对质粒phac181-mrlectin进行扩增,扩增成功的长片段为5348bp(图5),将扩增成功的长片段整合至酿酒酵母菌株中gal1启动子下游(图1),得到gal-phac181-mrlectin工程菌株。
40.f1:caaatgtaataaaagtatcaacaaaaaattgttaatatacctctatactttaacgtcaaggagaaaaaacccggatctcaaaatgtggcattcaaggggc(seq id no.2);
41.r1:tatggacgaggtaataaggaaactcagaaccagaatagtggcatgagctctccaatttaacatatttgccattagtgacccgatgataagctgtcaaacatg(seq id no.3);
42.基因同源重组(整合)的具体步骤为:
43.1.挑活化的gal1-scrch1菌株单菌落,接种到3ml ypd培养基,30℃220rpm培养过夜。
44.2.取300ul过夜培养物接种到4.7ml 2
×
ypd中,30℃培养4~5h(达到od600=0.6-1.0);
45.3.将5ml菌液分3管,室温离心4000rpm,1min,弃上清,再用1ml水重悬合并到1个ep管中,3000rpm离心1min,弃上清。
46.4.加100ul的0.1m liac溶液,吹吸混匀,12000rpm离心10s,弃上清。
47.5.重复步骤4。
48.6.依次加入下列物质,加入后温和混匀:
[0049][0050][0051]
7.混匀,30℃水浴中温育30min;
[0052]
9. 42℃水浴中温育30min(热激);
[0053]
10. 3000rpm离心除去上清,用1ml的无菌水重悬菌体,3000rpm离心1min弃上清,预留100ul菌液铺在选择平板(sd-leu)上,30℃培养3~5天。
[0054]
11.将长出的转化子在sd-leu平板上进行划线纯化。单菌落接种于ypd培养基中过夜培养,提取转化子基因组,用检测引物df、dr进行pcr检测,有目的片段扩增出的转化子即为同源重组成功的菌株gal1-phac181-mrlectin。
[0055]
同源重组的检测引物为df、dr,序列如下所示:
[0056]
df:cctggccccacaaaccttc(seq id no.4);
[0057]
dr:taggttgtatctgctgacc(seq id no.5);
[0058]
用检测引物(如seq id no.4和seq id no.5所示)对同源重组后的阳性转化子进行菌落pcr检测,目的条带扩增成功者(1276bp)则为同源重组成功的质粒(图6)。
[0059]
菌落pcr配制体系及步骤:
[0060][0061]
将上述体系混合均匀,短暂离心,使溶液聚集于管底,pcr反应程序采用:95℃预变性30s,95℃变性5s、60℃退火、72℃延伸30s,40个循环,72℃10min,4℃保存。
[0062]
实施例2:工程菌株蛋白表达与western blot检测
[0063]
上述同源重组成功的gal1-phac181-mrlectin菌株单菌落接种于sd-leu培养基5ml中过夜培养,培养物第二天转接于45mlypg培养基中,经半乳糖诱导培养6h(od检测为1.2-1.5)后,提取菌株蛋白。提取的蛋白用于western blot检测。
[0064]
具体步骤为:
[0065]
(一)细胞中总蛋白的提取
[0066]
1、挑取单菌落于所需液体培养基(sd-leu培养基)中30℃过夜培养至饱和;
[0067]
2、取5ml过夜培养的菌液加入45ml新鲜的液体培养基(ypg培养基)并且在30℃摇床里振荡培养约6h(od=1.2-1.5)(转速为220rmp);
[0068]
3、以8000rpm/1min收集菌体,去掉上层的液体;
[0069]
4、用预冷双蒸水重悬菌体,并于离心后去掉上清液;
[0070]
5、加入与菌体等量并预冷的peb(protein extraction buffer),peb中加入100
×
pmsf;
[0071]
6、加入与菌体等体积的酸洗玻璃珠;
[0072]
7、在振荡混合器上振荡ep管10
×
30s,每次振荡后放冰上1min;
[0073]
8、12000rpm离心10min,吸取上清液于冰上保存,弃去沉淀;
[0074]
9、用考马斯亮蓝法检测浓度(od
595
),将样品浓度调整一致;
[0075]
10、加入5
×
sb 95℃煮5min。
[0076]
(二)考马斯亮蓝法检测蛋白浓度
[0077]
1、标准蛋白溶液的制备:称量bsa固体10mg溶解于1ml的水中,得到10mg/ml的溶液作为标准贮液。用标准贮液配制系列稀释的标准蛋白溶液,其浓度分别为1.2mg/ml、1.0mg/ml、0.8mg/ml、0.6mg/ml、0.4mg/ml、0.2mg/ml、0.1mg/ml。
[0078]
2、待测蛋白溶液的制备:将待测样品稀释至浓度在0.1mg/ml-1.2mg/ml之间。
[0079]
3、4μl染液+200μl蛋白溶液,混匀,静置3min。按照核酸蛋白分析仪的“protein”程
序测定蛋白浓度。
[0080]
在本实验中,测得的酿酒酵母表达的mrlectin蛋白浓度为16ug/ul。换算可得蛋白浓度为0.064mg/ml,即每ml发酵液得到0.064mg蛋白。
[0081]
(三)sds-page
[0082]
1.装置组装;2.制胶;3.蛋白样品制备;4、上样,必须点预染marker,电泳。浓缩胶80v,分离胶100v。
[0083]
(四)转印(浸渍法)。
[0084]
(五)封闭:加入封闭液封闭,用摇床轻轻摇动1h-4h。
[0085]
(六)加入一抗:1、轻轻转移掉封闭液;2、加入一抗杂交液,室温2h或4℃过夜。
[0086]
(七)加入二抗1、轻轻转移掉一抗杂交液(可重复使用),用tbst洗3次,每次10min;2、加入二抗杂交液,室温轻摇2h;3、用tbst洗膜3次,每次10min。
[0087]
(八)反应。
[0088]
(九)曝光显影—暗光下操作
[0089]
显影后,依据蛋白marker相比,检测到有gal1-phac181-mrlectin目的蛋白表达,分子量为42kd(如seq id no.6所示),这些菌株为成功构建的酿酒酵母mrlectin免疫蛋白工程菌株(图7)。
[0090]
实施例3:酿酒酵母mrlectin工程菌株有效提高虾机体免疫力
[0091]
(1)酵母工程菌株的扩大培养及浓缩
[0092]
将上述构建成功的酿酒酵母mrlectin工程菌株进行实验室扩大培养。先将工程菌株用500ml三角瓶培养(扩大培养的培养基为:ypg培养基),然后扩大培养至1l,3l,5l,将培养后的工程菌株离心过滤,菌体置于室温自然干燥。阴干后的菌体与虾颗粒饲料混合,待其风干后,将混有酵母工程菌株的饲料投喂罗氏沼虾,同时设定对照组,观察含有重组工程菌株的饲料与普通饲料在投喂时对罗氏沼虾的影响。投喂期为一个月。
[0093]
(2)罗氏沼虾机体病原菌感染及死亡率测定
[0094]
从本地水产市场中购买体格大小均一(约20g左右)的罗氏沼虾300只。分组对其投喂不同饲料:实验组:普通虾颗粒饲料(巴大饲料,南通)中复配有酵母mrlectin工程菌株(0.3%);对照组一:普通虾颗粒饲料中复配有空载的酿酒酵母菌株;对照组二:普通虾颗粒饲料。投喂量为其体重的4%,每天早晚投喂2次,共投喂4周。1个月后用嗜水气单胞菌3.0
×
105cfu 50ul对虾体进行注射感染,同时设定50μlpbs注射作为对照组。在副溶血弧菌感染后的0,2,6,12,24和36h统计累积死亡率,结果表明,饲喂4周后,饲喂酿酒酵母mrlectin颗粒的罗氏沼虾死亡率最低(累积死亡率值为66.9%),普通饲料饲喂组死亡率最高,在感染后36h达到92.06%,饲料中混有酵母空载饲料组死亡率高于重组蛋白组小于普通饲料组(图8),这说明重组酵母mrlectin工程菌株可有效降低病原菌侵染后的死亡率。
[0095]
(3)罗氏沼虾肝胰腺组织免疫酶活力及免疫基因表达量测定
[0096]
在嗜水气单胞菌感染后的0,2,6,12,24和36h取肝胰腺组织进行免疫指标的测定,包括免疫酶活力及免疫基因的表达量变化测定。其中,测定的免疫酶活力包括:碱性磷酸酶(akp);酸性磷酸酶(acp);超氧化物歧化酶(sod)以及过氧化氢酶(cat)。
[0097]
具体步骤为:将0.1g左右的肝胰腺置于1ml 0.6%的生理盐水中进行匀浆处理(冰上操作),匀浆后的肝组织样品4℃,3500g离心10min,上清液用于免疫酶活力的测定。
[0098]
碱性磷酸酶(akp)的测定:
[0099]
碱性磷酸酶分解磷酸苯二钠,产生游离酚和磷酸,酚在碱性溶液中与4-氨基安替吡啉作用经铁氰化钾氧化生成红色醌衍生物,根据红色深浅,利用可见光分光光度计测其吸光度,则可以测定酶活力的高低。结果表明:饲喂酿酒酵母mrlectin组的罗氏沼虾的akp活性从感染后2小时开始增加,并在12小时达到最高值75.33u/g,酿酒酵母空载饲喂组与普通颗粒饲喂组相似,其酶活性水平低于目标蛋白组(图9中a图所示)。
[0100]
酸性磷酸酶(acp)的测定:
[0101]
酸性磷酸酶分解磷酸苯二钠,产生游离酚和磷酸,酚在碱性溶液中与4-氨基安替吡啉作用经铁氰化钾氧化生成红色醌衍生物,根据红色深浅,利用可见光分光光度计测其吸光度,可以测定酶活力的高低。结果表明:饲喂酿酒酵母mrlectin组的罗氏沼虾,其6h acp活性明显高于其他两组,在12h最高达到52.6u/g。空载酿酒酵母颗粒饲料和普通颗粒饲料饲喂组显示相似的结果,其acp酶活性水平低于目标蛋白组,并且在此期间没有明显变化(图9中b图所示)。
[0102]
超氧化物歧化酶(sod)的测定:
[0103]
通过黄嘌呤及黄嘌呤氧化酶反应系统产生超氧阴离子自由基(o
2-.
),后者氧化羟胺形成亚硝酸盐,在显色剂的作用下呈现紫红色,用可见光分光光度计测其吸光度。当被测样品中含sod时,则对超氧阴离子自由基有专一性的抑制作用,使形成的亚硝酸盐减少,比色时测定管的吸光值低于对照管的吸光值,通过计算公式可求出被测样品中的sod活力。结果表明:酿酒酵母mrlectin颗粒饲喂组sod活性在嗜水气单胞菌攻击后2h开始明显高于两个对照组,然后在12时达到最高值(30.3u/g,p《0.05)(图9中c图所示)。
[0104]
过氧化氢酶(cat)的测定
[0105]
实验采用钼铵酸比色法测定过氧化氢酶,其原理是过氧化氢酶催化h2o2分解生成h2o和o2,酶促反应剩余的h2o2与钼铵酸生成稳定的黄色复合物,其颜色深浅与酶活性成反比,405nm下进行比色测定,计算酶活性。结果表明:在饲喂酿酒酵母mrlectin的组中,cat活性在感染后2h和12h显着增加,其中12h达到最高值为11.56u/g,而饲喂酿酒酵母空载颗粒和普通饲料颗粒组低于靶蛋白组观察到的酶活力水平(图9中d图所示)。
[0106]
以上酶活力数据表明,在受到病原菌嗜水气单胞菌浸染后,饲喂复配有酵母工程菌mrlectin组中的罗氏沼虾表现了更强的免疫酶活力,说明该组中虾机体免疫力高于其他两个组。
[0107]
(4)rt-pcr检测罗氏沼虾肝胰腺组织免疫基因表达量变化
[0108]
通过实时定量pcr(rt-pcr)对感染嗜水气单胞菌的肝胰腺组织中的免疫基因(lectin,akp,acp,sod,cat)进行测定,检测酿酒酵母mrlectin工程菌株对免疫基因表达量的影响。具体步骤为:
[0109]
根据trizol试剂说明书,进行组织裂解并提取总rna。
[0110]
(1)将采集的虾体肝胰腺组织样品中加1mltrizol,于2ml匀浆器中充分快速研磨,室温下放置5min,使其充分裂解。此时样品可放入-70℃长期保存。12,000g离心5min,收集上清于新的离心管中,弃沉淀。
[0111]
(2)按200μl氯仿/mltrizol加入氯仿,振荡混匀15s,室温放置12min,4℃,12,000g离心15min。
[0112]
(3)吸取上层水相(约500-600μl),至另一离心管中。
[0113]
(4)按0.5ml异丙醇/ml trizol加入异丙醇混匀,室温放置8min。
[0114]
(5)4℃12,000g离心10min,弃上清,此时rna沉于管底。
[0115]
(6)按1ml 75%乙醇/ml trizol加入75%乙醇(depc处理过),温和振荡离心管,轻轻悬浮沉淀。
[0116]
(7)4℃8,000g离心5min,弃上清。
[0117]
(8)室温晾干或真空干燥5-10min。
[0118]
(9)用20-50μl depc-h2o溶解rna样品,55-60℃,5-10min,充分溶解rna。
[0119]
cdna反转录
[0120]
按下列组份配制rt反应液(反应液配制在冰上进行)。
[0121][0122]
反转录反应条件:37℃15min,85℃5s。
[0123]
免疫相关基因的荧光定量rt-pcr检测
[0124]
按照荧光定量pcr试剂盒sybr premix ex tagtm kit的说明书进行荧光定量pcr操作。pcr反应体系如下:
[0125][0126]
将上述体系混合均匀,短暂离心,使溶液聚集于管底,rt-pcr反应程序采用二步法:95℃预变性30s,95℃变性5s、60℃退火、延伸30s,40个循环。根据绘制的熔解曲线,摸索cdna模板的浓度。基因相对表达采用2-δδct
法(δδct=(ct目的基因-ct管家基因)实验组-(ct目的基因-ct管家基因)对照)计算,在使用ct方法之前,确定目的基因和管家基因的扩增效率基本一致。pbs刺激组作为空白对照,同时用肌动蛋白β-actin基因作为内参基因,数据分析采用t-检验,当p《0.01时被判定为显著性差异。
[0127]
结果表明:罗氏沼虾的免疫基因的表达量与对照组相比,lectin基因表达量在6h,12h,24h时明显增加,但在12h表达量达到最高值(51folds,p《0.01)(图10中a图)。akp基因表达量在24h达到最大值(15folds,p《0.01)(图10中b图)。acp基因表达量在2h时明显增加,24h表达量增加至最高值(52folds,p《0.01,但与两个对照组相比,酿酒酵母mrlectin饲喂组基因表达量高于两个对照饲喂组(图10中c图)。sod基因表达量在嗜水气单胞菌感染后的
12h至24h内上调(图10中d图)。cat基因在6h至12h时表达量上调并在12h达到最大峰值(33folds,p《0.01)(图10中e图)。根据上述研究结果,说明饲喂复配有酿酒酵母mrlectin工程菌株的颗粒饲料组中的罗氏沼虾在机体受到嗜水气单胞菌侵染后,其体内的免疫基因表达量更高,显著高于复配酿酒酵母空载颗粒饲料组和普通颗粒饲料组,这说明酿酒酵母mrlectin工程菌株有效增强了罗氏沼虾机体免疫能力,可在受到病原菌侵染时,机体可以表达更多的免疫基因来参与先天免疫。因此该酿酒酵母重组mrlectin菌株是具有较好潜力的免疫菌株,可提高甲壳动物机体免疫水平,增加对甲壳动物病原-嗜水气单胞菌的抵抗能力。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1