杨树钙调素结合蛋白PdeCAMBP在调控植物器官形成和生物量中的应用
本发明属于生物,具体涉及杨树钙调素结合蛋白pdecambp在调控植物器官形成和生物量中的应用。
背景技术:
1、木本植物之所以在茂密的森林中能够率先占领生态位,占领更大的生存空间,是因为其具有发达的维管系统,植物的维管系统不仅为植物提供了强有力的支撑作用,也是植物营养和水分等物质运输的重要场所。植物的维管系统发育主要分为初生生长、次生生长和初生生长向次生生长转换三个阶段。初生生长使植物纵向生长,是新植物器官,如叶片、花和分枝等组织的主要来源,同时,初生生长发育阶段提供的可以持续分裂的分生组织细胞是次生生长的细胞来源,在次生生长阶段,分生组织细胞分裂得到的细胞进一步分化,向内分化形成木质部,向外分化形成韧皮部,使植物进行径向的加粗。植物的维管系统对植物本身的生存具有重要意义,由叶片的光合作用产生的营养物质通过叶片的维管系统输送到植物的根、茎、花、果实等各个部位,是植物得以进行物质积累和繁衍的重要营养运输系统,而同时,茎的维管系统,尤其是木本植物茎的木质部,是木材、生物能源及化石燃料的主要来源,同时,森林也是陆地上最大的碳储库,因此,研究树木的维管系统发育,培育优良的林木,是解决能源危机和环境危机的重要途经之一。
技术实现思路
1、本发明要解决的技术问题是如何调控植物新生器官的形成及与生物量相关的性状,以培育优良林木。
2、为了解决上述技术问题,本发明首先提供了一种蛋白质,该蛋白质的名称为pdecambp-2,来源于杂交杨“南林895”(populus deltoides×p.euramericana),所述pdecambp-2蛋白质是如下a)或b)或c)或d)所示的蛋白质:
3、a)氨基酸序列是序列4所示的蛋白质;
4、b)在序列4所示的蛋白质的n端和/或c端连接标签得到的融合蛋白质;
5、c)将序列4所示的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的具有相同功能的蛋白质;
6、d)与序列4所示的氨基酸序列具有75%或75%以上的同一性且具有相同功能的蛋白质。
7、上述b)所述蛋白质中,所述标签是指利用dna体外重组技术,与目的蛋白一起融合表达的一种多肽或者蛋白质,以便于目的蛋白的表达、检测、示踪和/或纯化。所述标签可为flag标签、his标签、mbp标签、ha标签、myc标签、gst标签和/或sumo标签等。
8、上述c)所述蛋白质中,所述一个或几个氨基酸残基的取代和/或缺失和/或添加为不超过10个氨基酸残基的取代和/或缺失和/或添加或不超过9个氨基酸残基的取代和/或缺失和/或添加或不超过8个氨基酸残基的取代和/或缺失和/或添加或不超过7个氨基酸残基的取代和/或缺失和/或添加或不超过6个氨基酸残基的取代和/或缺失和/或添加或不超过5个氨基酸残基的取代和/或缺失和/或添加或不超过4个氨基酸残基的取代和/或缺失和/或添加或不超过3个氨基酸残基的取代和/或缺失和/或添加或不超过2个氨基酸残基的取代和/或缺失和/或添加或不超过1个氨基酸残基的取代和/或缺失和/或添加。
9、上述d)所述蛋白质中,所述同一性是指氨基酸序列的同一性。可使用国际互联网上的同源性检索站点测定氨基酸序列的同一性,如ncbi主页网站的blast网页。例如,可在高级blast2.1中,通过使用blastp作为程序,将expect值设置为10,将所有filter设置为off,使用blosum62作为matrix,将gap existence cost,per residue gap cost和lambdaratio分别设置为11,1和0.85(缺省值)并进行检索一对氨基酸序列的同一性进行计算,然后即可获得同一性的值(%)。
10、上述a)或b)或c)或d)所述蛋白质可人工合成,也可先合成其编码基因,再进行生物表达得到。
11、为了解决上述技术问题,本发明又提供了与pdecambp-2蛋白质相关的生物材料。
12、本发明提供的与pdecambp-2蛋白质相关的生物材料为下述a1)至a8)中的任一种:
13、a1)编码pdecambp-2蛋白质的核酸分子;
14、a2)含有a1)所述核酸分子的表达盒;
15、a3)含有a1)所述核酸分子的重组载体;
16、a4)含有a2)所述表达盒的重组载体;
17、a5)含有a1)所述核酸分子的重组微生物;
18、a6)含有a2)所述表达盒的重组微生物;
19、a7)含有a3)所述重组载体的重组微生物;
20、a8)含有a4)所述重组载体的重组微生物。
21、上述生物材料中,a1)所述核酸分子为如下1)或2)或3)所示的基因:
22、1)其编码序列是序列5或序列6所示的dna分子;
23、2)与1)限定的核苷酸序列具有75%或75%以上同一性,且编码pdecambp-2蛋白质的dna分子;
24、3)在严格条件下与1)或2)限定的核苷酸序列杂交,且编码pdecambp-2蛋白质的dna分子。
25、其中,所述核酸分子可以是dna,如cdna、基因组dna或重组dna;所述核酸分子也可以是rna,如mrna或hnrna等。
26、本领域普通技术人员可以很容易地采用已知的方法,例如定向进化和点突变的方法,对本发明的编码pdecambp-2蛋白质的核苷酸序列进行突变。那些经过人工修饰的,具有与本发明分离得到的pdecambp-2核苷酸序列75%或者更高同一性的核苷酸,只要编码pdecambp-2蛋白质且具有相同功能,均是衍生于本发明的核苷酸序列并且等同于本发明的序列。
27、这里使用的术语“同一性”是指与天然核酸序列的序列相似性。“同一性”包括与本发明的编码序列1所示的氨基酸序列组成的蛋白质的核苷酸序列具有75%或更高,或80%或更高,或85%或更高,或90%或更高,或95%或更高同一性的核苷酸序列。同一性可以用肉眼或计算机软件进行评价。使用计算机软件,两个或多个序列之间的同一性可以用百分比(%)表示,其可以用来评价相关序列之间的同一性。
28、上述生物材料中,a2)所述的含有编码pdecambp-2蛋白质的核酸分子的表达盒(pdecambp-2基因表达盒)是指能够在宿主细胞中表达pdecambp-2的dna,该dna不但可包括启动pdecambp-2转录的启动子,还可包括终止pdecambp-2转录的终止子。进一步的,所述表达盒还可包括增强子序列。可用于本发明的启动子包括但不限于:组成型启动子;组织、器官和发育特异的启动子及诱导型启动子。合适的转录终止子包括但不限于:农杆菌胭脂碱合成酶终止子(nos终止子)、花椰菜花叶病毒camv 35s终止子、tml终止子及豌豆rbcs e9终止子。
29、可用现有的表达载体构建含有所述pdecambp-2基因表达盒的重组载体。所述植物表达载体包括双元农杆菌载体和可用于植物微弹轰击的载体等。如pahc25、pbin438、pcambia1302、pcambia2300、pcambia2301、pcambia1301、pcambia1300、pbi121、pcambia1391-xa或pcambia1391-xb(cambia公司)等。所述植物表达载体还可包含外源基因的3′端非翻译区域,即包含聚腺苷酸信号和任何其它参与mrna加工或基因表达的dna片段。所述聚腺苷酸信号可引导聚腺苷酸加入到mrna前体的3′端,如农杆菌冠瘿瘤诱导(ti)质粒基因(如胭脂碱合成酶基因nos)、植物基因(如大豆贮存蛋白基因)3′端转录的非翻译区均具有类似功能。使用本发明的基因构建植物表达载体时,还可使用增强子,包括翻译增强子或转录增强子,这些增强子区域可以是atg起始密码子或邻接区域起始密码子等,但必需与编码序列的阅读框相同,以保证整个序列的正确翻译。所述翻译控制信号和起始密码子的来源是广泛的,可以是天然的,也可以是合成的。翻译起始区域可以来自转录起始区域或结构基因。为了便于对转基因植物细胞或植物进行鉴定及筛选,可对所用植物表达载体进行加工,如加入可在植物中表达的编码可产生颜色变化的酶或发光化合物的基因(gus基因、萤光素酶基因等)、抗生素的标记基因(如赋予对卡那霉素和相关抗生素抗性的nptii基因,赋予对除草剂膦丝菌素抗性的bar基因,赋予对抗生素潮霉素抗性的hph基因,和赋予对氨甲喋呤抗性的dhfr基因,赋予对草甘磷抗性的epsps基因)或是抗化学试剂标记基因等(如抗除莠剂基因)、提供代谢甘露糖能力的甘露糖-6-磷酸异构酶基因。从转基因植物的安全性考虑,可不加任何选择性标记基因,直接以逆境筛选转化植株。
30、上述生物材料中,所述载体可为质粒、黏粒、噬菌体或病毒载体。
31、上述生物材料中,所述微生物可为酵母、细菌、藻或真菌;所述细菌可为农杆菌,具体可为gv3101菌株。
32、为了解决上述技术问题,本发明还提供了上述pdecambp-2蛋白质或上述相关生物材料的新用途。
33、本发明提供了上述pdecambp-2蛋白质或上述相关生物材料在如下b1)-b8)任一种中的应用:
34、b1)调控(如促进)植物生长发育;
35、b2)调控(如促进)植物器官形成或产生;
36、b3)调控(如增加)植物器官数量;
37、b4)调控(如增加)植物生物量;
38、b5)调控(如增加)植物株高;
39、b6)调控(如增加)植物主根数量;
40、b7)调控(如增加)植物叶片数量;
41、b8)培育器官数量和/或生物量增加的转基因植物。
42、为了解决上述技术问题,本发明还提供了pdecambp-1蛋白质和pdecambp-2蛋白质的新用途。
43、本发明提供了pdecambp-1蛋白质和pdecambp-2蛋白质在如下c1)-c13)任一种中的应用:
44、c1)调控(如促进)植物生长发育;
45、c2)调控(如促进)植物器官形成或产生;
46、c3)调控(如增加)植物器官数量;
47、c4)调控(如增加)植物生物量;
48、c5)调控(如增加)植物主根数量;
49、c6)调控(如增加)植物主根长度;
50、c7)调控(如增加)植物叶片数量;
51、c8)调控(如增加)植物株高;
52、c9)调控(如增加)植物节间数量;
53、c10)调控(如降低)植物节间长度;
54、c11)调控(如增加)植物干重;
55、c12)培育器官数量和/或生物量增加的转基因植物;
56、c13)培育树形变化的转基因植物(如定向培育树形符合实际生产需求的林木)。
57、为了解决上述技术问题,本发明还提供了抑制植物中pdecambp-1蛋白质和pdecambp-2蛋白质活性的物质或降低植物中pdecambp-1蛋白质和pdecambp-2蛋白质含量的物质的新用途。
58、本发明提供了抑制植物中pdecambp-1蛋白质和pdecambp-2蛋白质活性的物质或降低植物中pdecambp-1蛋白质和pdecambp-2蛋白质含量的物质在如下d1)-d13)任一种中的应用:
59、d1)抑制植物生长发育;
60、d2)抑制植物器官形成或产生;
61、d3)减少植物器官数量;
62、d4)降低植物生物量;
63、d5)减少植物主根数量;
64、d6)减少植物主根长度;
65、d7)减少植物叶片数量;
66、d8)降低植物株高;
67、d9)减少植物节间数量;
68、d10)增加植物节间长度;
69、d11)减少植物干重;
70、d12)培育器官数量减少和/或生物量减少和/或节间长度增加的转基因植物;
71、d13)培育树形变化的转基因植物(如定向培育树形符合实际生产需求的林木)。
72、上述应用中,所述抑制植物中pdecambp-1蛋白质和pdecambp-2蛋白质活性的物质或降低植物中pdecambp-1蛋白质和pdecambp-2蛋白质含量的物质可为干扰植物中pdecambp-1蛋白质编码基因和pdecambp-2蛋白质编码基因表达的物质或敲除植物中pdecambp-1蛋白质编码基因和pdecambp-2蛋白质编码基因的物质。
73、进一步的,所述干扰植物中pdecambp-1蛋白质编码基因和pdecambp-2蛋白质编码基因表达的物质可为干扰植物中pdecambp-1蛋白质编码基因和pdecambp-2蛋白质编码基因表达的mirna或含有所述mirna的表达载体。
74、所述敲除植物中pdecambp-1蛋白质编码基因和pdecambp-2蛋白质编码基因的物质可为敲除植物中pdecambp-1蛋白质编码基因和pdecambp-2蛋白质编码基因的crispr/cas9系统,该crispr/cas9系统包括靶向pdecambp-1基因和pdecambp-2基因的sgrna或含有所述sgrna的表达盒或含有所述sgrna的表达载体。
75、更进一步的,所述干扰植物中pdecambp-1蛋白质编码基因和pdecambp-2蛋白质编码基因表达的mirna具体为taataggggtgactgtctcga。
76、所述靶向pdecambp-1基因的sgrna中的靶序列具体为aatcagcaagaactttggcg和tgggagaaacgttcagcttc,含有所述sgrna的表达盒的核苷酸序列具体如序列7和序列8所示。
77、所述靶向pdecambp-2基因的sgrna中的靶序列具体为gaagataatagctacgaggc和gcagcaagctcacaacattc,含有所述sgrna的表达盒的核苷酸序列具体如序列9和序列10所示。
78、为了解决上述技术问题,本发明还提供了一种培育器官数量和/或生物量增加的转基因植物的方法。
79、本发明提供的培育器官数量和/或生物量增加的转基因植物的方法为如下x1)或x2):
80、x1)包括提高受体植物中pdecambp-2的表达量和/或活性,得到转基因植物的步骤;所述转基因植物的器官数量和/或生物量高于所述受体植物;
81、x2)包括提高受体植物中pdecambp-1和pdecambp-2的表达量和/或活性,得到转基因植物的步骤;所述转基因植物的器官数量和/或生物量高于所述受体植物。
82、上述方法中,所述x1)中,所述转基因植物的器官数量和/或生物量高于所述受体植物体现在如下m1)-m3)中任一种;
83、m1)所述转基因植物的主根数量高于所述受体植物;
84、m2)所述转基因植物的叶片数量高于所述受体植物;
85、m3)所述转基因植物的株高高于所述受体植物。
86、进一步的,所述x1)中,所述提高受体植物中pdecambp-2蛋白质的表达量和/或活性的方法为在受体植物中过表达pdecambp-2蛋白质。
87、更进一步的,所述过表达的方法为将pdecambp-2蛋白质的编码基因导入受体植物。
88、为了解决上述技术问题,本发明最后提供了一种培育器官数量减少和/或生物量减少和/或节间长度增加的转基因植物的方法。
89、本发明提供的培育器官数量减少和/或生物量减少和/或节间长度增加的转基因植物的方法为如下y1)或y2):
90、y1)包括降低受体植物中pdecambp-2蛋白质的表达量和/或活性,得到转基因植物的步骤;所述转基因植物的器官数量和/或生物量低于所述受体植物;
91、y2)包括降低受体植物中pdecambp-1蛋白质和pdecambp-2蛋白质的表达量和/或活性,得到转基因植物的步骤;所述转基因植物的器官数量和/或生物量低于所述受体植物。
92、上述方法中,所述y2)中,所述转基因植物的器官数量和/或生物量低于所述受体植物体现在如下n1)-n6)中任一种;
93、n1)所述转基因植物的主根数量小于所述受体植物;
94、n2)所述转基因植物的主根长度小于所述受体植物;
95、n3)所述转基因植物的叶片数量小于所述受体植物;
96、n4)所述转基因植物的株高小于所述受体植物;
97、n5)所述转基因植物的节间数量小于所述受体植物;
98、n6)所述转基因植物的干重小于所述受体植物。
99、进一步的,所述y2)中,所述降低受体植物中pdecambp-1蛋白质和pdecambp-2蛋白质的表达量和/或活性的方法为将干扰受体植物中pdecambp-1蛋白质编码基因和pdecambp-2蛋白质编码基因的表达的物质或敲除受体植物中pdecambp-1蛋白质编码基因和pdecambp-2蛋白质编码基因的物质导入受体植物中。
100、更进一步的,所述干扰受体植物中pdecambp-1蛋白质编码基因和pdecambp-2蛋白质编码基因表达的物质可为干扰受体植物中pdecambp-1蛋白质编码基因和pdecambp-2蛋白质编码基因表达的mirna或含有所述mirna的表达载体。
101、所述敲除受体植物中pdecambp-1蛋白质编码基因和pdecambp-2蛋白质编码基因的物质可为敲除受体植物中pdecambp-1蛋白质编码基因和pdecambp-2蛋白质编码基因的crispr/cas9系统,该crispr/cas9系统包括靶向pdecambp-1基因和pdecambp-2基因的sgrna或含有所述sgrna的表达盒或含有所述sgrna的表达载体。
102、在本发明的具体实施例中,所述干扰受体植物中pdecambp-1蛋白质编码基因和pdecambp-2蛋白质编码基因表达的mirna为taataggggtgactgtctcga。
103、所述靶向pdecambp-1基因的sgrna中的靶序列为aatcagcaagaactttggcg和tgggagaaacgttcagcttc,含有所述sgrna的表达盒的核苷酸序列如序列7和序列8所示。
104、所述靶向pdecambp-2基因的sgrna中的靶序列为gaagataatagctacgaggc和gcagcaagctcacaacattc,含有所述sgrna的表达盒的核苷酸序列如序列9和序列10所示。
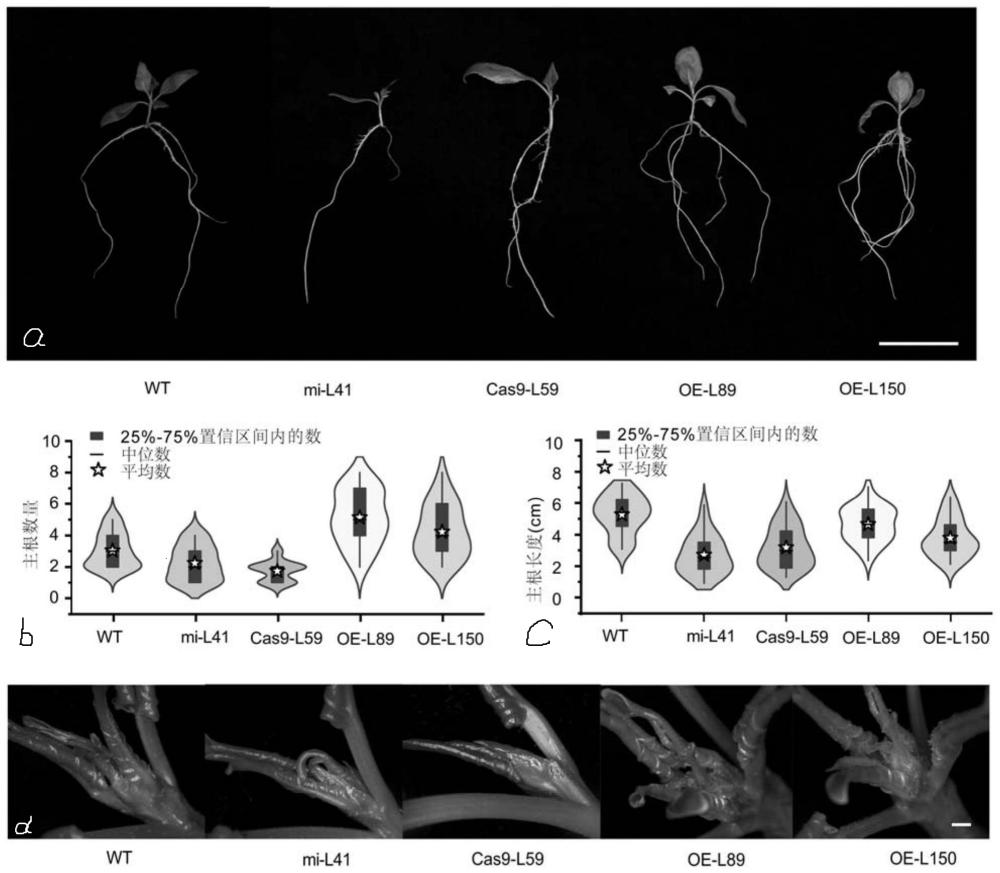
105、上述任一所述应用或方法中,所述器官(或新生器官)可为根和/或叶,所述根具体可为主根,所述叶具体可为新生的幼嫩叶片。
106、上述任一所述应用或方法中,所述节间长度可为第1节至第5节的总长度或第1节长度或第2节长度或第3节长度或第4节长度或第5节长度。
107、上述任一所述应用或方法中,所述pdecambp-1蛋白质的氨基酸序列如序列1所示。
108、上述任一所述应用或方法中,所述植物包括水稻、小麦、大麦、玉米、大豆、马铃薯、豆类、燕麦和粟等在内的粮食作物;包括拟南芥、大白菜、萝卜、辣椒、草莓、番茄、西瓜、黄瓜、卷心菜、甜瓜、西葫芦、韭菜、洋葱和胡萝卜等在内的蔬菜作物;包括人参、烟草、棉花、芝麻、甘蔗、甜菜、野生芝麻、花生和油菜籽等在内的经济作物;包括苹果、梨、枣、桃、猕猴桃、葡萄、橘子、柿子、李子、杏和香蕉等在内的水果;包括玫瑰、剑兰、大丁草、康乃馨、菊花、百合和郁金香等在内的花卉;包括黑麦草、红三叶草、鸭茅、紫花苜蓿、高酥油草和多年生黑麦草等在内的饲料作物;包括杨树、柳树、桦树、槐树、榆树、水杉、云杉、山毛榉、槭树、橡树、苦楝、水曲柳、金莲木、铁线子、紫檀、黄檀、柚木、白蜡、枫树等在内的用以制造木材的木本植物。进一步的,所述植物为双子叶植物。更进一步的,所述双子叶植物为双子叶木本植物。在本发明的具体实施例中,所述双子叶木本植物为杨树。
109、通过实验证明,将序列6所示的pdecambp-2cds序列与pcabia2300-3×flag融合的重组载体转化杨树,得到的转基因杨树在正常条件下与野生型杨树相比,株高增加了10.33%,主根数量增加了39.6%,且新产生的幼嫩叶片数量明显增多。而利用microrna干扰技术同时降低pdecambp-1和pdecambp-2基因的表达量或者利用crispr敲除技术同时敲除pdecambp-1和pdecambp-2基因时,得到的转基因杨树在正常条件下与野生型杨树相比,主根数量分别减少了26.3%和42.8%,主根长度分别减少了48.9%和39.6%,新产生的幼嫩叶片数量减少,株高分别降低了12.7%和25.3%,节间数量分别减少了21.5%和52%,干重分别降低了62.7%和62.9%,节间长度增加了82.31%和363.7%。与现有技术相比,本发明的有益效果如下:本发明既可通过提高植物pdecambp-1和/或pdecambp-2基因表达量来增加植物新器官的数量和生物量;亦可通过降低植物pdecambp-1和/或pdecambp-2基因表达量,或敲除pdecambp-1和/或pdecambp-2基因来减少植物新器官的产生,定向培育目标性状的植物,对人为控制植物器官形成与数量、节间数量和节间长度之间的平衡及生物量具有重要的指导意义。
- 还没有人留言评论。精彩留言会获得点赞!