基于基因密码子扩展技术细胞内编码膦配体设计金属酶
背景技术
0、
背景技术:
1、磷(p)是生物体的基本元素,其以磷酸盐、磷酸酯或者磷酸酐的形式(主要为+5价氧化态)执行许多基本生物功能1。然而,在非生物系统中磷通常采用-3、0、+3或者+5的氧化态。通常包含3个p-c键以及p原子上一个孤对电子的膦配体(p(iii))对于设计和合成可用于众多重要反应,如suzuki,heck,stille,negishi,buchwald-hartwig偶联以及不对称氢化反应的均相催化剂特别有用2。这部分是由于膦配体稳定过渡金属低氧化态,如pd(0),有利于金属中心氧化加成,还原消除和配体交换的独特能力1a。此外,staudinger连接,重要的生物正交反应,也发生在膦和叠氮之间3。
2、为了弥补非生物膦配体与蛋白化学之间的巨大差异,科学家们已经成功地将膦配体及其金属复合物在体外插入蛋白中,并创造了新的人工金属酶(arms);这是通过共价修饰蛋白侧链或者通过高亲和力锚的非共价连接来实现的4。arms是显著的混合催化剂,其将蛋白架构的卓越分子识别与人发明的过渡金属或有机催化剂的独特且多功能的反应性结合5。然而,由于磷配体的氧敏感性,研究人员目前主要通过将已有的膦—过渡金属催化剂和蛋白直接偶联和构建arms,该策略可以追溯到1978年wilson和whitesides最早将生物素化的膦-铑催化剂整合入亲和素架构6。kamer和laan团队在惰性气氛中进行了蛋白半胱氨酸介导的膦配体修饰4a,7。最近,ward和同事报道了使用生物素化的膦-钯复合物的链霉素基arms4b。o’hagan和shaw团队将马来酰亚胺修饰的膦-铑复合物锚定入乳酸乳球菌菌多药抗性调节因子(lmrr)蛋白架构。
3、尽管在体外进行了这些开创性的工作,构建精简的体内平台以实现有氧条件下膦配体位点特异性遗传整合仍然是艰巨的挑战。在过去二十年中,有力的遗传密码扩展策略(gce)已经使得许多非经典氨基酸(ncaas)在多种生物体中通过翻译过程被整合入蛋白中,从而产生新的功能蛋白9。通过该策略,我们设想可以创造出结合膦-金属化学和蛋白设计的新人工金属酶。
4、为了实现这一目标,必须克服三个挑战:首先,多数膦配体非常大以至于无法遗传整合入蛋白中。迄今为止,由于氨酰trna合成酶的底物口袋体积限制,基因编码的非天然氨基酸最多含有三个芳香环9a。其次,大多数膦配体容易被空气氧化,而进行氨酰trna合成酶定向进化以促进非天然氨基酸遗传整合的过程通常在有氧条件下进行;因此,需要膦配体保护策略。最后,由于大多数生物实验室不配备手套操作箱,金属偶联最好能在有氧条件下进行。
技术实现思路
1、为了实现上述目标,一方面,本申请提供了一种基于基因密码子扩展技术细胞内编码膦配体设计金属酶的方法,其特征在于,所述方法中使用结构如下所述的膦配体p3bf:
2、
3、另一方面,本申请提供了一种用于金属蛋白设计的基因编码磷配体p3bf,其结构为:
4、
5、另一方面,本申请提供了一种酪氨酰trna合成酶,其序列为seq id no.1。
6、另一方面,本申请提供了上述磷配体p3bf在制备金属蛋白中的应用。
7、进一步地,上述应用中使用上述酪氨酰trna合成酶。
8、另一方面,本申请提供了向金属蛋白中引入金属元素的方法,包括将上述磷配体p3bf遗传整合入蛋白,以及金属偶联的步骤。
9、进一步地,使用基因密码子扩展技术将上述磷配体p3bf遗传整合入蛋白,优选通过使用上述酪氨酰trna合成酶将上述磷配体p3bf遗传整合入蛋白,将上述磷配体p3bf遗传整合入蛋白的过程优选在大肠杆菌中进行。
10、进一步地,所述金属蛋白为人工酶。
11、进一步地,所述蛋白为lmrr蛋白,优选同二聚体lmrr蛋白。
12、进一步地,所述lmrr蛋白氨基酸序列为seq id no.11,其中15位处的f为p3bf。
13、进一步地,所述lmrr蛋白核苷酸编码序列为seq id no.8。
14、进一步地,所述金属元素为钯,优选为钯环二聚体。
15、进一步地,所述金属偶联包括单锅脱硼步骤,优选所述单锅脱硼步骤包括将含有p3bf的蛋白与1当量以上,优选1当量的钯环二聚体孵育。
16、进一步地,所述孵育在dmf水溶液中,有氧条件下进行。
17、另一方面,本申请提供了金属蛋白,所述金属蛋白包含膦配体-金属复合物。
18、进一步地,所述金属蛋白为人工酶。
19、进一步地,所述膦配体-金属复合物为p3f金属复合物,所述p3f为:
20、
21、进一步地,所述人工酶中包括lmrr蛋白,优选同二聚体lmrr蛋白人工酶载体。
22、进一步地,所述lmrr蛋白氨基酸序列为seq id no.11,其中15位处的f为p3f。
23、进一步地,所述lmrr蛋白核苷酸编码序列为seq id no.8。
24、本申请还提供了一种包含磷配体的超级折叠荧光蛋白,其氨基酸序列为:seq idno:5-7,其中第17、3或者9位置处的f为p3bf。
25、进一步地,所述包含磷配体的超级折叠荧光蛋白,其编码核苷酸序列选自:seq idno.2-4。
26、本申请中的简写或者名称对应的化学式如下,这些名称和化学式以及对应化学式的其他命名可以交换使用,表示相同的意思。
27、p3bf:
28、
29、p3f:
30、
31、钯环(cpd)二聚体:
32、
33、p3f-cpd:
34、
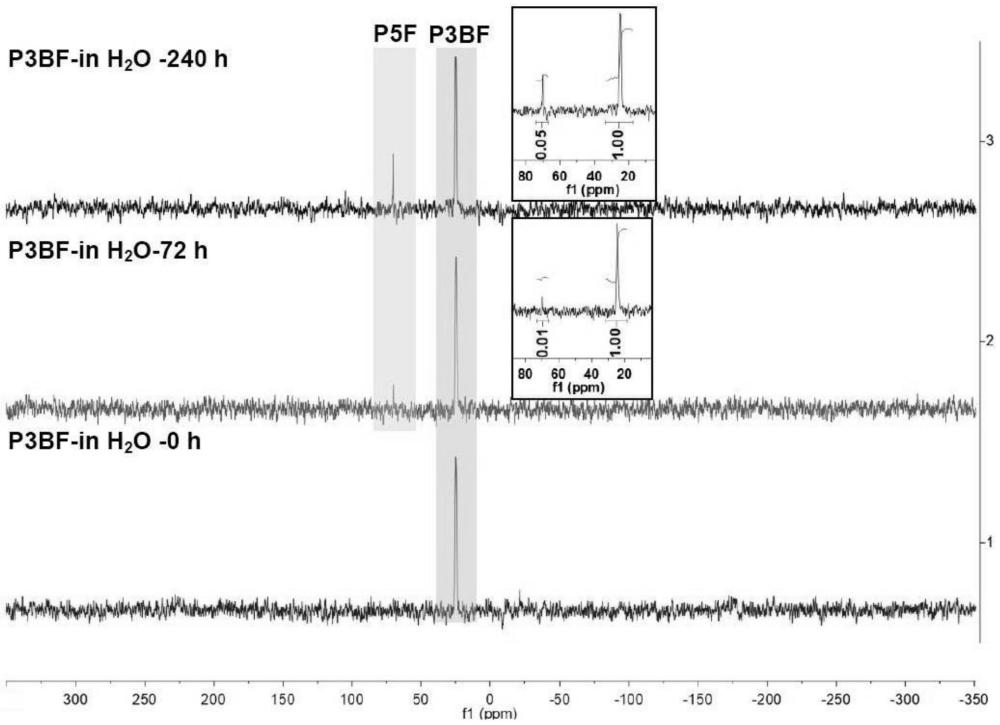
35、本申请开发了一种有效的策略以在大肠杆菌中将p3bf,一种含有硼烷保护的膦的非天然氨基酸定点整合入蛋白。p3bf显示出良好的生物相容性并在生理环境中保持稳定。这是第一个关于膦配体遗传纳入蛋白的报道。我们以的分辨率解析了含有p3bf的sfgfp的三维结构,这是第一种含p-b键的蛋白结构。重要的是,我们开发了在有氧条件下水溶液中实现高效单锅脱硼和钯配位的简单方法。这个有前途的简便平台能赋予蛋白以膦配体的非凡化学性质。强有力的基因工程和定向进化策略9f,28现在可以与独特的膦-金属化学无缝结合以超出主要配位条件精细调整金属环境,并能够合理设计许多新人工金属酶29。
技术特征:
1.一种基于基因密码子扩展技术细胞内编码膦配体设计金属酶的方法,其特征在于,所述方法中使用结构如下所述的膦配体p3bf:
2.一种用于金属蛋白设计的基因编码磷配体p3bf,其特征在于,所述磷配体p3bf结构为:
3.一种酪氨酰trna合成酶,其特征在于,所述酪氨酰trna合成酶氨基酸序列为seq idno.1。
4.根据权利要求2所述的磷配体p3bf在制备金属蛋白中的应用。
5.根据权利要求4中所述的应用,其中使用根据权利要求3所述的酪氨酰trna合成酶。
6.向蛋白中引入金属元素的方法,其特征在于,所述方法包括将根据权利要求2所述的磷配体p3bf遗传整合入蛋白,以及金属偶联的步骤。
7.根据权利要求6所述的方法,其中使用基因密码子扩展技术将磷配体p3bf遗传整合入蛋白;优选通过使用根据权利要求3所述的酪氨酰trna合成酶将磷配体p3bf遗传整合入蛋白;将磷配体p3bf遗传整合入蛋白的过程优选在大肠杆菌中进行。
8.根据权利要求6或7所述的方法,其中所述蛋白为人工酶。
9.根据权利要求6-8任一项所述的方法,所述蛋白为lmrr蛋白,优选同二聚体lmrr蛋白。
10.根据权利要求9所述的方法,其中所述lmrr蛋白氨基酸序列为seq id no.11,其中15位处的f为p3bf。
11.根据权利要求10所述的方法,其中所述lmrr蛋白核苷酸编码序列为seq id no.8。
12.根据权利要求7-11任一项所述的方法,其中所述金属元素为钯,优选为钯环二聚体。
13.根据权利要求7-12任一项所述的方法,其中所述金属偶联包括单锅脱硼步骤,优选所述单锅脱硼步骤包括将含有p3bf的蛋白与1当量以上,优选1当量的钯环二聚体孵育。
14.根据权利要求13所述的方法,所述孵育在dmf水溶液中,有氧条件下进行。
15.一种金属蛋白,其特征在于,所述金属蛋白包含膦配体-金属复合物。
16.根据权利要求15所述的金属蛋白,其中所述金属蛋白为人工酶。
17.根据权利要求15或16所述的金属蛋白,其中所述膦配体-金属复合物为p3f金属复合物,所述p3f为:
18.根据权利要求16或17所述的金属蛋白,其中所述人工酶中包括lmrr蛋白,优选同二聚体lmrr蛋白人工酶载体。
19.根据权利要求18所述的金属蛋白,所述lmrr蛋白氨基酸序列为seq id no.11,其中15位处的f为p3f。
20.根据权利要求19所述的金属蛋白,所述lmrr蛋白核苷酸编码序列为seq id no.8。
技术总结
本申请提供了一种基于基因密码子扩展技术细胞内编码膦配体设计金属酶的方法和其中使用的磷配体P3BF,以及使用基因密码子扩展技术将该磷配体P3BF纳入蛋白,进而向蛋白中引入金属的方法。本申请还提供了单锅方法以在水和有氧条件下进行磷配体P3BF的脱硼和钯配位。磷配P3BF显示出良好的生物相容性并在生理环境中保持稳定。本申请的膦配体和相应方法能够用于设计和制备金属蛋白,大幅度简化反应过程和条件要求。
技术研发人员:王江云,胡诚,刘晓红
受保护的技术使用者:中国科学院生物物理研究所
技术研发日:
技术公布日:2024/4/22
- 还没有人留言评论。精彩留言会获得点赞!