一种通过操纵m6ARNA甲基化酶调控生物合成基因表达的方法及其应用
本发明属于医药,涉及表观遗传学和代谢工程领域,特别涉及到通过调控酿酒酵母中rna的n6-甲基腺嘌呤(m6a)修饰重塑酿酒酵母代谢网络的代谢工程新方法及其在构建以乙酰辅酶a和芳香族氨基酸为前体的化合物(如萜类化合物和酪醇)高产酵母工程菌中的应用。
背景技术:
1、近年来,代谢工程在规模化生产生物燃料、精细化学品、药用化合物等方面取得了突破性进展。例如,高产青蒿酸的工程化酿酒酵母的建立,产率高达25g/l(paddon c j,etal.nature,2013,496:528-532);建立高产檀香挥发油所有8种主要成分的工程酿酒酵母,总产量达到1.6g/l(zha w,et al.acs synthetic biololy,2020,9:449-456);用工程化的mannheimia succiniciproducens生产琥珀酸,产率为每小时38.6g/l(lee j w,etal.metabolic engineering,2016,38:409-417)。
2、目前代谢工程中所使用的优化策略主要集中以下几个方面。包括关键酶的改造和过表达、启动子和转录终止子的工程化、转运蛋白的工程化、调控因子和调控蛋白的工程化、辅因子供给的调控、细胞器的工程化以及酶表达的细胞器区室化改造(lian j,etal.metabolic engineering,2018,50:85-108;arendt p,et al.metabolic engineering,2017,40:165-175;kim j e,et al.metabolic engineering,2019,56:50-59;farhi m,etal.metabolic.engineering,2011,13:474-481;yuan j,and ching cb.metabolic.engineering,2016,38:303-309)。以上策略的运用推动了代谢工程领域的发展。但是,许多化合物,尤其是复杂化合物的高产体系的构建仍面临巨大挑战。而且以上策略的迭代使用,造成细胞负担过重,不利于细胞的生长繁殖和化合物产量的提高。造成这些困难的根本原因是目前对生物体复杂的代谢调控网络并不完全了解。
3、rna的n6-甲基腺嘌呤(m6a)修饰在真核生物的基因表达调控中扮演重要角色。ime4是酿酒酵母中的m6a甲基化酶,调控细胞分裂等相关基因的表达。但是,m6a对生物体代谢网络的系统作用以及在代谢工程中的应用,尚无报道(bushkin g g,et al.naturecommunications,2019,10:3414;agarwala s d,et al.plos genetics,2012,8:e1002732)。
技术实现思路
1、本发明的首要目的在于提供一种通过操纵m6a rna甲基化酶调控生物合成基因表达的方法,以及提供所述方法在构建酿酒酵母工程菌中的应用。
2、本发明的另一目的在于提供一种酿酒酵母工程菌株,以及其应用。
3、为实现上述目的,本发明通过以下技术方案予以实现:
4、本发明技术方案的第一方面是提供了一种通过操纵m6a rna甲基化酶调控生物合成基因表达的方法,其特征在于,在酿酒酵母内过表达m6a rna甲基化酶基因ime4。
5、进一步的,将ime4基因内源性启动子替换为强启动子ppgk1。
6、所述的酿酒酵母优选为by4741(购买自美国英杰生命技术有限公司(invitrogen))。
7、本发明技术方案的第二方面是提供了一种通过操纵m6a rna甲基化酶调控生物合成基因表达的方法在代谢工程中的应用,其特征在于,所述的应用包括酿酒酵母工程菌株的构建。
8、进一步的,所述的应用包括以乙酰辅酶a和芳香族氨基酸为前体的化合物酿酒酵母工程菌株的构建。
9、进一步的,所述的化合物包括萜类化合物或芳香族化合物。
10、进一步的,所述的萜类化合物包括紫穗槐-4,11-二烯、广藿香醇,檀香烯和檀香醇,所述的芳香族化合物包括酪醇、色醇、对香豆酸、咖啡酸、阿魏酸、柚皮素、圣草酚、紫杉叶素和白藜芦醇。
11、本发明技术方案的第三个方面是提供了一种酿酒酵母工程菌株,其特征在于,所述的菌株过表达m6arna甲基化酶基因ime4。
12、进一步的所述的酿酒酵母工程菌株包括以下菌株:
13、(1)以酿酒酵母by4741为出发菌株,过表达ime4基因,构建的酪醇和色醇酿酒酵母工程菌株pime-1;
14、(2)以酿酒酵母by4741为出发菌株,过表达ime4基因,导入紫穗槐-4,11-二烯的生物合成途径基因,构建的紫穗槐-4,11-二烯酿酒酵母工程菌株z6;
15、(3)以酿酒酵母by4741为出发菌株,过表达ime4基因,导入广藿香醇的生物合成途径基因,构建的一种广藿香醇酿酒酵母工程菌株z7;
16、(4)以菌株wl-2(已申请中国专利,申请号:201911180415.0,名称:一种高产檀香烯和檀香醇的酿酒酵母工程菌及其构建方法与应用,已公开)为出发菌株,过表达ime4基因,构建的α-檀香烯/z-α-檀香醇酿酒酵母工程菌株z8;
17、(5)以酿酒酵母by4742(购买自美国英杰生命技术有限公司(invitrogen))为出发菌株,过表达ime4基因,单拷贝导入对香豆酸的生物合成途径基因,构建的两种对香豆酸酿酒酵母工程菌株z9和z10;
18、(6)以酿酒酵母by4742为出发菌株,过表达ime4基因,多拷贝导入对香豆酸的生物合成途径基因,构建的两种对香豆酸酿酒酵母工程菌株z11和z12;
19、(7)以工程菌株yc1031(该菌株已在yang j,et al.metabolic engineering,2020,59:44-52中公开)为出发菌株,过表达ime4基因,构建的两种咖啡酸/阿魏酸酿酒酵母工程菌z13和z14;
20、(8)以工程菌株yt1021(该菌株已在yang j,et al.metabolic engineering,2020,59:44-52中公开)为出发菌株,过表达ime4基因,构建的一种柚皮素/圣草酚/紫杉叶素酿酒酵母工程菌z15;
21、(9)以酿酒酵母by4742为出发菌株,过表达ime4基因,导入白藜芦醇的生物合成途径基因,构建的两种白藜芦醇工程菌株z16和z17。
22、进一步的,所述的酿酒酵母工程菌株,其特征在于,各酿酒酵母工程菌株构建方法如下所示:
23、(1)所述的菌株pime-1以酿酒酵母by4741为出发菌株,将by-1的ime4基因内源性启动子替换为ppgk1启动子;
24、(2)所述菌株z6以pime-1为出发菌株,过表达ads基因,ads基因整合到酿酒酵母染色体ndt80位点;
25、(3)所述菌株z7以pime-1为出发菌株,过表达pts基因,pts基因整合到酿酒酵母染色体ndt80位点;
26、(4)所述菌株z8以wl-2为出发菌株,将ime4内源性启动子替换为ppgk1启动子;
27、(5)所述菌株z9以酿酒酵母by4742为出发菌株,过表达hatal、aro4k229l和aro7g141s基因,hatal、aro4k229l和aro7g141s基因整合到酿酒酵母染色体ndt80位点,将ime4内源性启动子替换为ppgk1启动子;
28、(6)所述菌株z10以酿酒酵母by4742为出发菌株,过表达hatal、aro4k229l和aro7g141s基因,hatal、aro4k229l和aro7g141s基因整合到酿酒酵母染色体ndt80位点,将ime4内源性启动子替换为ptef2启动子;
29、(7)所述菌株z11以酿酒酵母by4742为出发菌株,过表达hatal、aro4k229l和aro7g141s基因,hatal、aro4k229l和aro7g141s基因整合到酿酒酵母染色体ty2位点,将ime4内源性启动子替换为ppgk1启动子;
30、(8)所述菌株z12以酿酒酵母by4742为出发菌株,过表达hatal、aro4k229l和aro7g141s基因,hatal、aro4k229l和aro7g141s基因整合到酿酒酵母染色体ty2位点,将ime4内源性启动子替换为ptef2启动子;
31、(9)所述菌株z13以yc1031为出发菌株,将ime4内源性启动子替换为ppgk1启动子;
32、(10)所述菌株z14以yc1031为出发菌株,将ime4内源性启动子替换为ptef2启动子;
33、(11)所述菌株z15以yt1021为出发菌株,将ime4内源性启动子替换为ptef2启动子;
34、(12)所述菌株z16以酿酒酵母by4742为出发菌株,,过表达hatal、aro4k229l和aro7g141s基因,hatal、aro4k229l和aro7g141s基因整合到酿酒酵母染色体ty2位点,过表达at4cl2和vvvst基因,at4cl2和vvvst基因整合到酿酒酵母染色体met17位点,将ime4内源性启动子替换为ptef2启动子;
35、(13)所述菌株z17以酿酒酵母by4742为出发菌株,,过表达hatal、aro4k229l和aro7g141s基因,hatal、aro4k229l和aro7g141s基因整合到酿酒酵母染色体ty2位点,过表达at4cl2和vvvst基因,at4cl2和vvvst基因整合到酿酒酵母染色体met17位点,将ime4内源性启动子替换为ppgk1启动子。
36、进一步的,所述的酿酒酵母工程菌的构建方法,包括如下步骤:
37、(1)模块的构建
38、(a)构建his3-ppgk1片段,命名为模块1;
39、(b)构建ura3-pgal10-ads-tcyc1片段,命名为模块2;
40、(c)构建ura3-pgal10-pts-tcyc1片段,命名为模块3;
41、(d)构建kileu2-ptdh3-hatal-ttdh2-ppgk1-aro4k229l-tadh1-ptef1-aro7g141s-tcyc1片段,命名为模块4;
42、(e)构建leu2-ptef1-vvvst-tcyc1-ppgk1-at4cl2-tadh1片段,命名为模块5;
43、(f)构建leu2-ppgk1片段,命名为模块6;
44、(g)构建his3-ptef2片段,命名为模块7。
45、(2)构建菌株pime-1
46、将步骤(1)中得到的模块1整合到by4741染色体pime4位点,得到菌株pime-1。
47、(3)构建菌株z1
48、将步骤(1)中得到的模块2整合到by4741染色体ndt80位点,得到菌株z1。
49、(4)构建菌株z2
50、将步骤(1)中得到的模块3整合到by4741染色体ndt80位点,得到菌株z2。
51、(5)构建菌株z3
52、将步骤(1)中得到的ptdh3-hatal-ttdh2-ppgk1-aro4k229l-tadh1-ptef1-aro7g141s-tcyc1和ura3整合到by4742染色体ndt80位点,得到菌株z3。
53、(6)构建菌株z4
54、将步骤(1)中得到的模块4整合到by4742染色体ty2位点,得到菌株z4。
55、(7)构建菌株z5
56、将步骤(1)中得到的模块5整合到z4染色体met17位点,得到菌株z5。
57、(8)构建菌株z6
58、将步骤(1)中得到的模块2整合到pime-1染色体ndt80位点,得到菌株z6。
59、(9)构建菌株z7
60、将步骤(1)中得到的模块3整合到pime-1染色体ndt80位点,得到菌株z7。
61、(10)构建菌株z8
62、将步骤(1)中得到的模块6整合到wl-2染色体pime4位点,得到菌株z8。
63、(11)构建菌株z9
64、将步骤(1)中得到的模块1整合到z3染色体pime4位点,得到菌株z9。
65、(12)构建菌株z10
66、将步骤(1)中得到的模块7整合到z3染色体pime4位点,得到菌株z10。
67、(13)构建菌株z11
68、将步骤(1)中得到的模块1整合到z4染色体pime4位点,得到菌株z11。
69、(14)构建菌株z12
70、将步骤(1)中得到的模块7整合到z4染色体pime4位点,得到菌株z12。
71、(15)构建菌株z13
72、将步骤(1)中得到的模块1整合到yc1031染色体pime4位点,得到菌株z13。
73、(16)构建菌株z14
74、将步骤(1)中得到的模块7转化整合到yc1031染色体pime4位点,得到菌株z14。
75、(17)构建菌株z15
76、将步骤(1)中得到的模块7整合到yt1021染色体pime4位点,得到菌株z15。
77、(18)构建菌株z16
78、将步骤(1)中得到的模块7整合到z5染色体pime4位点,得到菌株z16。
79、(19)构建菌株z17
80、将步骤(1)中得到的模块1整合到z5染色体pime4位点,得到菌株z17。
81、步骤(1)所述的ads、pts、hatal、aro4k229l,aro7g141s,at4cl2和vvvst的核苷酸序列分别如seq id no.1~7所示。
82、步骤(1)所述的筛选标记his3、ura3、kileu2和leu2的核苷酸序列如seq id no.8~11所示。
83、步骤(1)所述的启动子ppgk1、pgal10、ptdh3、ptef1和ptef2的核苷酸序列如seq id no.12~16所示。
84、步骤(1)所述的终止子tcyc1、ttdh2和tadh1的核苷酸序列如seq id no.17~19所示。
85、本发明技术方案的第四方面是提供了上述任一项的酿酒酵母工程菌株在制备化合物中的应用,其特征在于:
86、(1)菌株pime-1经发酵培养,从发酵液中可制备酪醇和色醇;
87、(2)菌株z6经发酵培养,从发酵液中可制备紫穗槐-4,11-二烯;
88、(3)菌株z7经发酵培养,从发酵液中可制备广藿香醇;
89、(4)菌株z8经发酵培养,从发酵液中可制备α-檀香烯和z-α-檀香醇;
90、(5)菌株z9、z10、z11或z12经发酵培养,从发酵液中可制备对香豆酸;
91、(6)菌株z13或z14经发酵培养,从发酵液中可制备咖啡酸和阿魏酸;
92、(7)菌株z15经发酵培养,从发酵液中可制备柚皮素、圣草酚和紫杉叶素;
93、(8)菌株z16或z17经发酵培养,从发酵液中可制备白藜芦醇。
94、与现有技术相比,本技术具有以下有益效果:
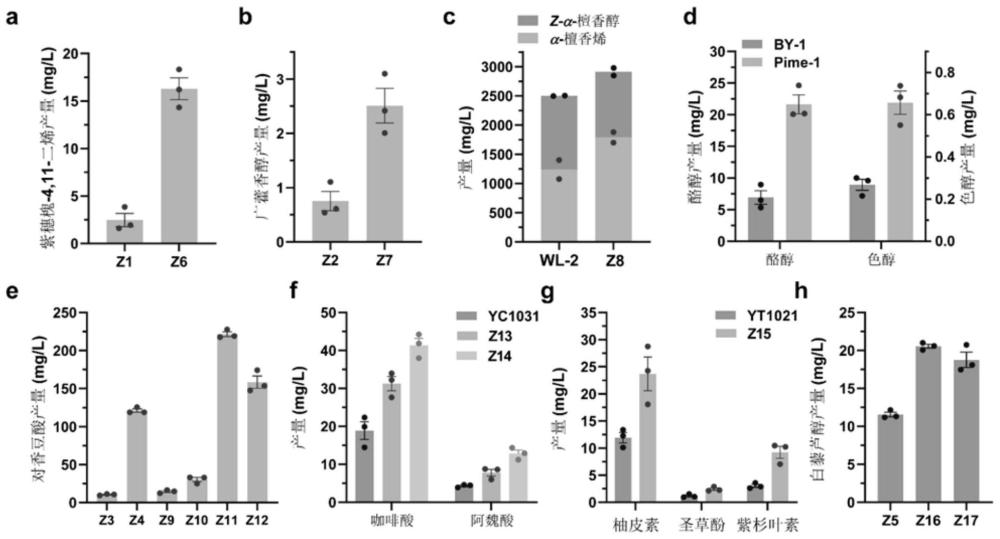
95、本发明提供了一种通过操纵m6a rna甲基化酶调控生物合成基因表达的新方法,在酿酒酵母内过表达m6a甲基化酶基因ime4,能够显著改变糖酵解、丙酮酸代谢、芳香族氨基酸合成、泛酸和辅酶a合成、辅因子合成等途径中关键基因的表达水平(图1)。提供的所述方法在构建产以乙酰辅酶a和芳香族氨基酸为前体化合物的酵母工程菌中的应用,表明过表达m6a rna甲基化酶ime4能够使紫穗槐-4,11-二烯(amorpha-4,11-diene)产量提高5.6倍,达到16.3mg/l;广藿香醇(patchoulol)产量提高2.3倍,达到2.5mg/l;α-檀香烯(α-santalene)和z-α-檀香醇(z-α-santalol)总产量提高16.4%,达到2.9g/l;酪醇(tyrosol)产量提高2.1倍,达到21.6mg/l;色醇(tryptophol)产量提高1.5倍,达到0.66mg/l;对香豆酸(p-coumaric acid)产量最大提高82.9%,达到221.4mg/l;咖啡酸(caffeic acid)产量最大提高1.2倍,达到41.4mg/l;阿魏酸(ferulic acid)产量最大提高2.0倍,达到12.8mg/l;柚皮素(naringenin)产量提高了98.7%,达到了23.7mg/l;圣草酚(eriodictyol)产量提高1.1倍,达到了2.4mg/l;紫杉叶素(taxifolin)产量提高2.0倍,达到9.2mg/l;白藜芦醇(resveratrol)的产量最大提高77.9%,达到20.6mg/l(图1)。
96、表1.过表达ime4基因使代谢途径基因的表达水平发生显著性改变的基因列表
97、
98、
99、
100、
101、
- 还没有人留言评论。精彩留言会获得点赞!