包含与细胞因子融合用于nk细胞衔接的nkp46结合位点、癌抗原结合位点的多特异性蛋白质的制作方法
提供了经由与多种受体相互作用而结合并特异性地重定向效应细胞以裂解感兴趣的靶细胞的多特异性蛋白质。这些蛋白质可用于治疗疾病,特别是癌症或感染性疾病。
背景技术:
1、白介素2(il2或il-2)是作用于由nk细胞表达的细胞因子受体的多能细胞因子的一个示例。il-2主要由活化的t细胞,尤其是cd4+t辅助性细胞产生,并且在辅助b细胞、t细胞和nk细胞的增殖和分化中起作用。il-2还是treg功能和存活所必需的。在真核细胞中,人il-2(uniprot:p60568)被合成为具有20个残基信号序列的153个氨基酸的前体肽,从而产生具有seq id no:404的氨基酸序列的成熟分泌的il-2。白介素2具有四个反向平行的两亲性α螺旋。这四个α螺旋是形成其功能所必需的四级结构。在大多数情况下,il-2通过三种不同受体起作用:白介素2受体α(il-2rα;cd25)、白介素2受体β(il-2rβ;cd122)和白介素2受体γ(il-2rγ;cd132)。il-2rβ和il-2rγ是il-2信号传导所必需的,虽然il-2rα(cd25)不是信号传导所必需的,但可赋予il-2与受体的高亲和力结合。由il-2rα、β和γ的组合形成的三聚体受体(il-2αβγ)是il-2高亲和力受体(kd约10pm),二聚体受体(il-2βγ)是中等亲和力受体(kd约1nm)。
2、免疫细胞表达二聚体或三聚体il-2受体。二聚体受体在细胞毒性cd8+t细胞和自然杀伤细胞(nk)上表达,而三聚体受体主要在活化淋巴细胞和cd4+cd25+foxp3抑制性调节性t细胞(treg)上表达。因为静息效应t细胞和nk细胞在细胞表面上不具有cd25,所以它们对il-2相对不敏感。treg细胞在体内始终表达最高水平的cd25。由于通常存在于组织中的低浓度的il-2,il-2优先激活表达高亲和力受体复合物(cd25:cd122:cd132)的细胞,因此在正常情况下,il-2将优先刺激treg细胞增殖。
3、il-15、il-12、il-7、il-27、il-18、il-21和ifn-α共享与il-2的受体结合、复合物组装和信号传导的许多方面。例如,il-15、il-21和il-7像il-2一样都经由共同的γ链受体(cd132)作用于nk细胞。il-15与il-15受体(il-15r)结合,该受体由三个亚基:il-15rα、cd122和cd132组成。这些亚基中的两者(即cd122和cd132)与il-2的受体共享,但il-2受体具有附加亚基(cd25)。il-15rα(cd215)以非常高的亲和力特异性地结合il15,并且能够独立于其他亚基而结合il-15。il-21是i型细胞因子的另一个示例,并且其il-21受体(il-21r)已被证明与il-2/il-15受体共同γ链(cd132)形成异源二聚体受体复合物。
4、nk细胞具有介导抗肿瘤免疫的潜力。然而,当il-2与ifn-α一起施用时,nk细胞已被证明通过它们的过度活化以及分泌多种炎性细胞因子而在小鼠中引起毒性(rothschilds等人,oncoimmunology.,2019年,第8卷第5期:)。此外,nk细胞还被证明引起细胞因子il-15的毒性,该细胞因子也通过il-2rβy进行传导信号(参见引用以下文献的wo2020247843:guo等人,j immunol.,2015年,第195卷第5期:第2353-64页)。
5、由细胞因子诸如il-2介导的免疫毒性的一个潜在解决方案是将该细胞因子与肿瘤特异性抗体融合或缔合。然而,据发现,虽然il-2与抗肿瘤抗体在体内抗肿瘤中确实有协同作用,但在相同分子中包含il-2和抗肿瘤抗原抗体没有呈现出疗效或毒性优势。il-2部分完全控制生物分布,这解释了以下观察结果:当与抗体组合时,识别无关抗原的免疫细胞因子的表现与肿瘤特异性免疫细胞因子相当(tzeng等人,proc natl acad sci usa.,2015年3月17日,第112卷第11期:第3320-332页)。
6、关于细胞因子对nk细胞的影响的研究通常集中在单一细胞因子或简单的组合。最近,已经报道了单独和组合的il-15、il-18、il-21和ifn-α,以及它们与il-2协同作用的潜力,并且非常低浓度的先天性和适应性共同γ链细胞因子与同样低浓度的il-18协同作用以驱动快速和有效的nk细胞的cd25和ifn-γ表达(nielsen等人,front immunol.,2016年,第7卷:第101页)。然而,向人类施用细胞因子涉及毒性,这使得与细胞因子的组合治疗具有挑战性。此外,对nk细胞中的细胞因子受体信号传导途径与其他活化受体之间的潜在协同作用或相互作用知之甚少。因此,需要在疾病、特别是癌症的治疗中动员nk细胞的新方法。
技术实现思路
1、本发明源于对功能性多特异性蛋白质的这种发现:这些功能性多特异性蛋白质与nk细胞上的nkp46和细胞因子受体结合,并且任选地进一步结合nk细胞上的cd16a,并且还与靶细胞(例如癌细胞)上的感兴趣的抗原(例如癌抗原)结合。这些多特异性蛋白质能够增加nk细胞对表达该感兴趣的抗原的靶细胞(例如,导致疾病的细胞、癌细胞)的细胞毒性。
2、据信多特异性蛋白质在nk细胞表面以顺式与nkp46和细胞因子受体(并且任选地进一步通过cd16a)结合的能力导致特别有利的细胞表面受体信号传导,继而导致nk细胞的有效抗肿瘤应答。
3、已知il2-介导的免疫毒性由nk细胞驱动。然而,本公开的带有il-2部分的多特异性蛋白质表现出强的nk细胞介导的抗肿瘤活性而没有免疫毒性。与单独的细胞因子相比,在多特异性蛋白质中掺入细胞因子使细胞因子-受体介导的活性在nk细胞中的效力增加两个数量级。同时,当结合nkp46和cd16a两者并且包含细胞因子(il-2变体)(该细胞因子基本上保留了对其在野生型il-2的nk细胞上的受体的亲和力)时,多特异性蛋白质示出了用于诱导肿瘤细胞的细胞毒性的ec50,这远低于该蛋白质诱导nk细胞增殖的ec50。因此,多特异性蛋白质在它们开始诱导显著的细胞因子-受体信号传导和/或nk细胞增殖的浓度下,甚至当细胞因子被减毒时,在激活nk细胞介导的细胞毒性方面将是高度有效的。这可以解释所观察到的毒性缺乏,因为所使用(例如施用)的蛋白质的浓度(例如剂量)可以是高度有效的,但仍保持低于引起nk细胞介导的免疫毒性的水平。因此,这些蛋白质代表了一种通用平台,该通用平台能够容易地整合不同细胞因子和细胞因子的不同减毒水平。
4、当将该蛋白质制成掺入cd122 abd(其体现为失去与cd25的结合的il2变体)时,与天然il-2相比,多特异性蛋白质在诱导treg细胞中的信号传导和/或活化的能力方面表现出4-log降低。当与前述的和天然il-2相比诱导nk细胞中的信号传导的能力的2-log增加组合时,与在tregs中相比,多特异性蛋白质在诱导nk细胞中的细胞因子受体信号传导的效力方面具有1,000,000倍的增加。因此,结合cd122的多特异性蛋白质可以这样的浓度使用:其中它们介导nk细胞增殖和/或浸润,同时尽量减少可能对抗肿瘤应答造成负面影响的treg增殖。
5、尽管所采用的蛋白质被设计成仅以单价方式结合其在肿瘤细胞上的靶标,但这些蛋白质表现出高度有利的体内功效。通过避免由与肿瘤细胞上的靶抗原的多价结合引起的亲和力增加,与肿瘤抗原、nkp46(和cd16a)和细胞因子受体的单价结合使多特异性蛋白质易于调节。据观察,每种abd(即,肿瘤抗原abd、cd16a abd、nkp46 abd和细胞因子受体abd)对体内nk细胞介导的抗肿瘤活性的增强做出了不同贡献。将具有单价肿瘤抗原结合的示例性蛋白质掺入具有亲和力的细胞因子,该亲和力使得这些蛋白质与nk细胞上的该细胞因子的受体结合,并且该亲和力不高于由nkp46 abd赋予nkp46的亲和力。抗肿瘤细胞活性高于相当的常规抗体,即使常规抗体是以二价结合的,因此通常对其靶标具有相当高的结合亲和力。
6、在体内,多特异性蛋白质显著增加肿瘤中的nk细胞浸润;与共享相同抗肿瘤vh/vl对的金标准adcc诱导性抗体奥妥珠单抗(obinutuzumab)的1.3倍增加相比,以及与缺乏cd122 abd的相同多特异性蛋白质的1.6倍增加相比,多特异性蛋白质导致肿瘤中表达nkp46的nk细胞的9倍增加。
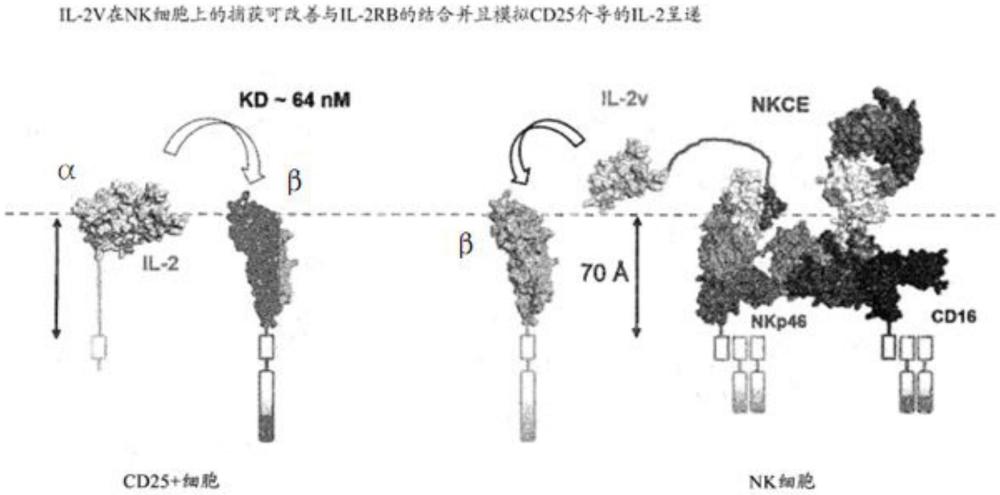
7、这些示例使用变体il-2细胞因子(il-2v),该细胞因子经修饰以降低对其在t细胞(cd25)上的受体的亲和力,但基本上保留了对其在nk细胞上的受体(cd122和/或cd132)的基本上全部的亲和力(与野生型il-2相当)。结合nkp46的结构域(以包含在fab或scfv中的vh/vl对为例)、结合cd16的fc结构域和细胞因子在该蛋白质内彼此相邻串联放置,各自仅通过短的柔性肽接头与相邻元件(即,nkp46 abd、fc结构域或细胞因子)分开。多特异性蛋白质的这些构型被设计为呈递相应抗原结合结构域以便允许nkp46和细胞因子受体在相同细胞表面平面上的共衔接(即,在nkp46中,细胞因子受体(和另外的cd16a)以顺式结合)。此外,这些示例使用结合cd16a的fc结构域,表明与cd16a结合不会对肿瘤和靶向nk的生物分布造成负面影响,而是导致nkp46、cd16a和细胞因子受体的三重共衔接,并且继而允许掺入一种细胞因子,该细胞因子保留了对其在nk细胞上的受体的结合亲和力。通过掺入多特异性蛋白质抗nkp46vh/vl结构域而在kd的低纳摩尔范围(kd为约15nm)内赋予对nkp46的结合亲和力,可使用保留了对其在nk细胞上的受体的良好亲和力(任选地基本上全部的结合亲和力)的细胞因子,从而足以介导nk细胞中的有效信号传导。通常,细胞因子(诸如本文所述的细胞因子)通常具有与其在nk细胞上的受体结合的亲和力,该亲和力不强于多特异性蛋白质对nkp46的亲和力(亲和力可确定为kd)。
8、考虑到这些结果,据信将细胞因子(例如1型细胞因子(诸如il-2、il-15、il-21、il-7、il-27或il-12细胞因子、il-18细胞因子)或1型干扰素(例如ifn-α、ifn-β))靶向带有nkp46的nk细胞表面,从而促进向细胞因子的受体(例如il2/15βγ、il-21r、il-7ra、il-27ra、il-12r、il-18r、ifnar)的顺式呈递,如图1所示的细胞因子il2和细胞因子受体复合物il2βγ)。如本文所示,紧邻结合cd16a的abd或结合nkp46的abd(并且在其c末端侧上)放置的il2v允许发生三重受体顺式呈递(il2v通过具有少至五个氨基酸残基的接头肽连接至相邻结构域)。有利地,使用二聚体fc结构域nas、结合cd16a的abd提供了fcrn,继而赋予足够长的半衰期以在体内诱导肿瘤浸润和nk细胞的增殖。
9、针对nk细胞上的nkp46的多特异性蛋白质具有以下优点:这些蛋白质允许使用和/或测试一系列细胞因子,而不需要降低对其在nk细胞上的受体(例如cd122)的结合亲和力。因此,可对细胞因子进行或不进行修饰以减弱或降低对其受体的结合亲和力。因此,针对nk细胞上的nkp46的多特异性蛋白质可利用以野生型形式存在的几种细胞因子中的任一种,特别是在细胞因子对其在nk细胞上的受体的活性没有显著降低的情况下,以及/或者在细胞因子对其受体的亲和力不强于nkp46 abd对nkp46的亲和力的情况下。因此,在任何实施方案中,可将细胞因子abd(例如多特异性蛋白质内的细胞因子部分)指定为对其在nk细胞上的受体具有结合亲和力和/或活性(例如诱导信号传导),该结合亲和力和/或活性与细胞因子的野生型形式的结合亲和力和/或活性相比没有显著降低。任选地,细胞因子部分在nk细胞上的其受体(例如cd122)处诱导信号转导,该信号转导是用细胞因子的野生型形式观察到的信号转导的至少70%或80%。因此,在任何实施方案中,可将细胞因子abd(例如多特异性蛋白质内的细胞因子部分)指定为对其在nk细胞上的受体具有亲和力,该亲和力与细胞因子的野生型形式的亲和力相比没有显著降低。在一些实施方案中,细胞因子部分对其在nk细胞上的受体在(例如cd122)具有结合亲和力,该结合亲和力在细胞因子的野生型形式的结合亲和力的3-log、2-log或1-log内(例如,细胞因子部分具有与细胞因子受体结合的kd,该kd比所观察到的细胞因子的野生型形式的kd高不超过3-log、2-log或1-log)。亲和力可以是与重组受体蛋白质结合的kd,如使用spr所测定的。可将细胞因子的信号传导或受体结合亲和力指定为当掺入在其他方面等价的多特异性蛋白质中时。
10、因此,高疗效与低免疫毒性的组合是治疗分子的一个特殊优势,该治疗分子组合了结合个体nk细胞上的nkp46、细胞因子受体(例如cd122)和另外的cd16a中的每一者的能力,特别是对于具有长的体内半衰期的治疗剂而言。具体地,结合fcrn的fc结构域的掺入允许足够长的半衰期,从而允许nk细胞在体内的肿瘤部位处增殖和累积。
11、多特异性蛋白质是特别有利的,因为其不仅在增强nk细胞活性(例如nk细胞增殖、活化、细胞毒性和/或细胞因子释放,包括通过肿瘤浸润的nk细胞)方面具有高效力,而且还具有低免疫毒性,如通过细胞因子il-6和tnf-α的低全身增加或释放所证明的。本公开提供了使用蛋白质形式的示例,这些蛋白质形式允许nkp46与细胞因子受体(例如cd122)和cd16a结合结构域之间有足够的距离以允许所有三种受体与单个nk细胞结合,从而提供组合的nk细胞受体活化。重要的是,在单个细胞上的组合结合可以解释表达nkp46和/或表达cd16的效应细胞(例如,nk细胞)的最小的脱靶免疫毒性并且没有自相残杀,因为多特异性蛋白质除了与nkp46和/或cd16+效应细胞表面上的细胞因子受体(例如,cd122)结合之外,还与至少一种活化受体结合。
12、多特异性蛋白质具有进一步的优势,因为它们能够增强nkp46+cd16+nk细胞和nkp46+cd16a-nk细胞两者的活性和/或增殖。如本文所示,在不存在与cd16a结合的情况下,与nkp46和cd122的组合双重结合展示出nk细胞活性的明显增强作用。在健康个体中,cd16-群体占总nk细胞群体的5%-15%,而在一些癌症患者中,cd16-nk细胞的比例大大增加,占总nk细胞群体的多达50%。此外,已经显示肿瘤微环境通过诱导cd16a从细胞表面脱落或促进cd16a+nk细胞向cd16-nk细胞的转化来影响cd16a+nk细胞的表型。此外,由于cd16a多态性,一些个体具有cd16a中的突变(例如在cd16a的残基158处),这导致介导adcc的能力降低。克服例如可能发生于肿瘤环境中的cd16a缺陷,同时增加肿瘤中nkp46+nk细胞的数量和活化,这是特别有利的。此外,多特异性蛋白质不需要经由nkg2d进行结合或信号传导,并且可用于在具有nk和/或t细胞的患者中增强nk细胞活性,这些nk和/或t细胞的特征在于所活化受体nkg2d的相对低水平的表面表达,例如已知是胃癌和前列腺癌中的普遍或共同特征。
13、尤其提供了一种多特异性蛋白质,其包含:(a)与人nkp46多肽结合的结合nkp46的结构域,(b)结合感兴趣的抗原(例如肿瘤相关抗原或癌抗原;由靶细胞表达呈递的感兴趣的抗原)的结合结构域,(c)与人cd16a多肽结合的任选的结合cd16a的结构域(例如fc二聚体),和(c)与nk细胞上表达的人细胞因子受体多肽(例如受体,诸如如cd122(il2/15rβ)、il-21r、il-7ra、il-27ra、il-12r、il-18r、ifnar(ifnar1和/或ifnar2))结合的抗原结合结构域。还提供了一种多特异性蛋白质,其包含:与人nkp46多肽结合的结合nkp46的结构域,结合感兴趣的抗原(例如肿瘤相关抗原或癌抗原;由靶细胞表达呈递的感兴趣的抗原)的结合结构域,与人fcrn结合(并且任选地进一步与人cd16a多肽结合)的fc结构域(例如fc结构域二聚体),以及与人细胞因子受体多肽(例如cd122(il2/15rβ)、il-21r、il-7ra、il-27ra、il-12r、il-18r、ifnar(ifnar1和/或ifnar2))结合的抗原结合结构域。结合细胞因子受体的抗原结合结构域可以是变体细胞因子,其与其野生型形式相比具有这样的修饰:该修饰降低了与非nk细胞(例如t细胞、treg细胞)上发现的受体对应物的结合。
14、在任何实施方案中,可将与人nkp46多肽结合的abd和与人细胞因子受体结合的abd指定为被构造成能够采用膜平面结合确认。
15、在任何实施方案中,可将多特异性蛋白质指定为能够与nk细胞表面上的nkp46和细胞因子受体以及任选的另外的cd16a相互作用、结合或共衔接。
16、在任何实施方案中,任选地将与人nkp46多肽结合的abd和与人细胞因子受体结合的abd以及任选的另外的fc结构域指定为在多特异性蛋白质内串联定位或连接(例如,相对于多特异性蛋白质的n末端和c末端)。
17、可将结合nkp46的abd(例如fab、单可变结构域或scfv)任选地指定为通过ig衍生的结构域接头(例如来自铰链结构域或者重链或轻链恒定结构域的肽)或非ig衍生的结构域接头连接至结合cd16a的结构域(例如fc结构域),任选地其中该结构域接头是柔性多肽接头。可将结合细胞因子受体的abd任选地指定为包含野生型或变体细胞因子,该野生型或变体细胞因子通过结构域接头(任选的柔性多肽接头)连接至多特异性蛋白质的其余部分或nkp46 abd。可将细胞因子任选地指定为定位在多特异性蛋白质上的结合nkp46的结构域和结合cd16a的结构域两者的c末端,并且任选地进一步,细胞因子经由具有15个、10个或5个残基或更少个残基的肽接头连接至多特异性蛋白质的其余部分(或例如,其结构域,nkp46abd)。可将结合nkp46的结构域和结合cd16a的结构域任选地指定为在多特异性蛋白质上彼此相邻放置并且任选地通过具有15个、10个或5个残基或更少个残基的长度的肽接头(例如免疫球蛋白衍生的接头(诸如铰链衍生的接头)、非免疫球蛋白衍生的接头、柔性接头)彼此连接。
18、在一个实施方案中,将与人nkp46多肽结合的abd定位成与蛋白质内的fc结构域相邻(或在其多肽链上),并且其中将与人nkp46多肽结合的abd和fc结构域中的一者定位成与结合人细胞因子受体的abd相邻,任选地进一步地,其中结合人细胞因子的abd通过具有20个或少于20个氨基酸残基、任选地少于15个氨基酸残基、任选地少于10个氨基酸残基、任选地介于5个和15个残基之间、任选地介于5个和10个残基之间、任选地介于3个和5个残基之间的接头肽连接至与人nkp46多肽结合的abd或fc结构域。
19、在任何方面,结合细胞因子受体的abd可以是人细胞因子多肽,例如cd122(il2/15rβ)、il-21r、il-7ra、il-27ra、il-12r、il-18r、ifnar(ifnar1和/或ifnar2)。结合细胞因子受体的abd可以任选地是人细胞因子多肽(例如il-2、il-15、il-21),其经修饰(例如通过引入氨基酸修饰)以降低对其所结合的细胞因子受体的结合亲和力,任选地其中对不在nk细胞表面表达的受体或者也在非nk细胞(例如t细胞、treg细胞)表面表达的受体的结合亲和力被选择性地降低。例如,结合人细胞因子受体的abd可以是变体细胞因子,与未修饰的或野生型细胞因子多肽相比,该变体细胞因子表现出降低的对t细胞上存在的细胞因子受体的结合亲和力。
20、如本文进一步所述的,当细胞因子具有多于一个受体作为其天然结合的配偶体并且这些受体中的一个受体在非nk细胞上表达时,细胞因子多肽与其野生型细胞因子对应物相比可经修饰以减少与在非nk细胞(例如treg细胞、t细胞)上表达的此类受体的结合。
21、在一个实施方案中,以掺入nkp46-1 vh/vl对的cdr的蛋白质为例,结合nkp46的结构域与nkp46多肽的d1/d2结合部结合。基于与结合nkp46的结构域复合的nkp46的x射线晶体研究,据信结合nkp46的多肽和il-2部分的结合部定位在距细胞表面约70埃处,这对应于cd122的细胞因子结合位点距细胞表面的预测距离。如图1c所示,与nkp46多肽的d1/d2结合部结合以及/或者与结合到nkp46-1的区域或表位结合可提供nkp46 abd在距nk细胞表面一定距离处的定位,该定位允许细胞因子受体诸如cd122的最佳衔接。继而,长度缩短的结构域接头(例如介于2个和5个残基之间,介于2个和10个残基之间;3个、4个、5个、6个、7个、8个、9个或10个残基)可用于细胞因子和nkp46 abd或多特异性蛋白质的其余部分之间,而效力没有任何降低。当结合nkp46上的其他结构域时,可使用更长的结构域接头,例如介于5个和15个残基、介于10个和15个残基之间或更多个残基。因此,本文描述的具有有限数量的非常短的接头的蛋白质具有以下优点:具有最少的非天然(或非免疫球蛋白衍生的)氨基酸序列。
22、在一个实施方案中,多特异性蛋白质包含结合nkp46的结构域或其部分,该结合nkp46的结构域或其部分任选地经由结构域接头与结合细胞因子受体的结构域(例如与在nk细胞表面表达的受体结合的细胞因子)融合。
23、在一个实施方案中,多特异性蛋白质包含结合nkp46的结构域或其部分,该结合nkp46的结构域或其部分任选地经由结构域接头与结合细胞因子受体的结构域(例如与在nk细胞表面表达的受体结合的细胞因子)融合。在一个实施方案中,结合nkp46的结构域的部分包括单一可变结构域(例如,与第一恒定结构域融合的第一可变结构域),该单一可变结构域与互补可变结构域(例如,与第二恒定区融合的第二可变结构域)一起形成结合nkp46的abd(例如,fab)。
24、在一个实施方案中,多特异性蛋白质包含:(i)第一多肽链,该第一多肽链从n末端至c末端包含结合nkp46的结构域或其部分,该结合nkp46的结构域或其部分包括可变结构域、人ch1或cl恒定结构域、任选的结构域接头,以及野生型或变体il-2、il-15、il-21、il-7、il-27、il-12、il-18、ifn-α或ifn-β多肽;和
25、(ii)第二多肽链,该第二多肽链从n末端至c末端包含可变结构域和人ch1或cl恒定结构域,该可变结构域与(i)的可变结构域缔合以形成结合nkp46的结构域;
26、其中(i)和(ii)的恒定结构域中的一者是ch1并且另一者是cl,使得(i)和(ii)的恒定结构域通过ch1-cl二聚化缔合。在一个实施方案中,该蛋白质还包含二聚体fc结构域和结合感兴趣的抗原的abd。
27、在一个实施方案中,提供了一种包含nkp46 abd细胞因子单位的蛋白质。在一个实施方案中,该蛋白质是多特异性蛋白质,该多特异性蛋白质包含vh和vl以及nkp46 abd-细胞因子单位(和任选的另外的cd16aabd(例如二聚体fc结构域)),该vh和该vl缔合以形成结合癌抗原或其他感兴趣的抗原的abd。nkp46 abd-细胞因子单元包含nkp46 abd,该nkp46 abd任选地经由结构域接头与结合在nk细胞表面表达的受体的细胞因子融合。
28、在本文的任何实施方案中,多特异性蛋白质因此可包含nkp46 abd-细胞因子单元,该nkp46 abd-细胞因子单元由两条多肽链缔合形成并且具有以下结构之一:
29、
30、或
31、
32、或
33、
34、其中:
35、va-2和vb-2各自是vh结构域或vl结构域,其中va-2和vb-2中的一者是vh并且另一者是vl,并且其中va-2和vb-2形成结合nkp46的abd;
36、ch1是人免疫球蛋白ch1结构域,并且cl是人轻链恒定结构域;
37、(ch1或cl)a和(ch1或cl)b中的一者是ch1并且另一者是cl,从而形成(ch1/cl)对;
38、l、l1和l2各自是氨基酸结构域接头,其中l、l1和l2可以是不同的或相同的,其中l1是将nkp46 abd-细胞因子单元连接至多特异性蛋白质(例如,包含vh和vl的蛋白质,并且该vh和该vl缔合形成结合癌抗原的abd)的其余部分的接头;并且cyt是与nk细胞上存在的细胞因子受体结合的细胞因子多肽或其部分,任选地其中cyt是野生型或变体人il-2、il-15、il-21、il-7、il-27、il-12、il-18、ifn-α或ifn-β多肽。可将cyt指定为具有游离的c末端(没有另外的结构域或氨基酸序列在cyt的游离末端与其融合)。任选地,可将链1指定为具有游离的c末端。可将“-”指定为表示经由共价键(例如肽键)与其他氨基酸残基连接。
39、在一个实施方案中,nkp46 abd-细胞因子单位包含:(i)第一多肽链,该第一多肽链从n末端至c末端包含结合nkp46的结构域或其部分,该结合nkp46的结构域或其部分包括可变结构域、人ch1或cl恒定结构域、任选的结构域接头和野生型或变体il-2、il-15、il-21、il-7、il-27、il-12、il-18、ifn-α或ifn-β多肽,该可变结构域包含与seq id no:3、5、7、9、11、13、112、113、115、116、117、119、120、121、123、124、125、127、128、129或236-313中任一项的氨基酸序列至少80%、90%、95%、98%或99%相同的氨基酸序列,该野生型或变体多肽包含与seq id no:404-439中任一项的氨基酸序列或其至少40、50、60、80或100个连续氨基酸的片段至少80%、90%、95%、98%或99%相同的氨基酸序列;和
40、(ii)第二多肽链,该第二多肽链从n末端至c末端包含可变结构域和人ch1或cl恒定结构域,该可变结构域与(i)的可变结构域缔合以形成结合nkp46的结构域,其中该可变结构域包含与seq id no:4、6、8、10、12、14、114、118、122、126、130或314-403中任一项的氨基酸序列至少80%、90%、95%、98%或99%相同的氨基酸序列。
41、在一个实施方案中,nkp46 abd-细胞因子单位包含:(i)第一多肽链,该第一多肽链从n末端至c末端包含结合nkp46的结构域或其部分,该结合nkp46的结构域或其部分包括可变结构域、人ch1或cl恒定结构域、任选的结构域接头和野生型或变体il-2、il-15、il-21、il-7、il-27、il-12、il-18、ifn-α或ifn-β多肽,该可变结构域包含与seq id no:4、6、8、10、12、14、114、118、122、126、130或314-403中任一项的氨基酸序列至少80%、90%、95%、98%或99%相同的氨基酸序列,该野生型或变体多肽包含与seq id no:404-439中任一项的氨基酸序列或其至少40、50、60、80或100个连续氨基酸的片段至少80%、90%、95%、98%或99%相同的氨基酸序列;和
42、(ii)第二多肽链,该第二多肽链从n末端至c末端包含可变结构域和人ch1或cl恒定结构域,该可变结构域与(i)的可变结构域缔合以形成结合nkp46的结构域,其中该可变结构域包含与seq id no:3、5、7、9、11、13、112、113、115、116、117、119、120、121、123、124、125、127、128、129或236-313中任一项的氨基酸序列至少80%、90%、95%、98%或99%相同的氨基酸序列。
43、在其中nkp46 abd-细胞因子单元置于蛋白质的一条多肽链上的另一个实施方案中,nkp46 abd-细胞因子单元可具有以下结构:
44、–l1–va-2–l2–vb-2–l3–cyt
45、其中:
46、va-2和vb-2各自是vh结构域或vl结构域,其中va-2和vb-2中的一者是vh并且另一者是vl,并且其中va-2和vb-2形成结合nkp46的abd;
47、l1、l2和l3各自是氨基酸结构域接头,其中l1、l2和l3可以是不同的或相同的,其中l1是将nkp46 abd-细胞因子单元连接至多特异性蛋白质(例如,包含vh和vl的蛋白质,并且该vh和该vl缔合形成结合癌抗原的abd)的其余部分的接头;并且cyt是与nk细胞上存在的细胞因子受体结合的细胞因子多肽或其部分,任选地其中cyt是野生型或变体人il-2、il-15、il-21、il-7、il-27、il-12、il-18、ifn-α或ifn-β多肽。可将l1、l2和l3各自独立地指定为具有15个、10个或5个残基或更少个残基的长度。
48、在本文任何实施方案的一个方面,细胞因子或细胞因子受体abd(作为游离细胞因子或掺入到多特异性蛋白质中)结合其受体,如通过spr测定的,其中结合亲和力(kd)为1μm或更低、200nm或更低、100nm或更低、50nm或更低、或25nm或更低。在一个实施方案中,细胞因子或细胞因子受体abd结合其受体,如通过spr测定的,其中结合亲和力(kd)为1nm或高于1nm、任选地高于10nm、任选地高于15nm。在一个实施方案中,细胞因子或细胞因子受体abd结合其受体,如通过spr测定的,其中结合亲和力(kd)介于约1nm至约200nm之间、任选地介于约1nm至约100nm之间、任选地介于约10nm至约1μm之间、任选地介于约10nm至约200μm之间、任选地介于约10nm至约100nm之间、任选地介于约15nm至约1μm之间、或任选地介于约15nm至约200nm之间。
49、在一个实施方案中,细胞因子是野生型细胞因子或其片段或变体,该细胞因子具有野生型细胞因子对应物诱导nk细胞中信号传导的能力的至少80%,任选地其中通过使分离的细胞因子部分与nk细胞接触并测量nk细胞中的stat磷酸化来评估信号传导。在一个实施方案中,细胞因子是野生型细胞因子或其片段,该细胞因子与野生型细胞因子对应物相比保留了对nk细胞上存在的其细胞因子受体的亲和力的至少70%、80%或90%。在一个实施方案中,细胞因子是变体细胞因子,其中该细胞因子与野生型细胞因子对应物相比保留了对nk细胞上存在的其细胞因子受体的亲和力的至少70%、80%或90%。在一个实施方案中,细胞因子不包含显著降低细胞因子对nk细胞上存在的细胞因子受体的亲和力的突变。在一个实施方案中,多特异性蛋白质(或当包含在多特异性蛋白质中时的细胞因子)在nk细胞中表现出用于细胞因子途径信号传导的ec50,其低于单独使用其野生型细胞因子对应物观察到的ec50。在一个实施方案中,多特异性蛋白质(或当包含在多特异性蛋白质中时的细胞因子)在nk细胞中表现出用于细胞因子途径信号传导的ec50,其低于单独使用细胞因子或在相当结构但缺乏nkp46abd和/或cd16 abd的蛋白质中观察到的ec50。任选地,用于nk细胞中细胞因子途径信号传导的ec50低到至少1/10或1/100,任选地,其中通过使相应的细胞因子或多特异性蛋白质与nk细胞接触并测量nk细胞中的stat磷酸化来评估细胞因子途径信号传导。
50、在一个实施方案中,多特异性蛋白质被构造成使得fc结构域(或结合cd16的结构域)、结合nkp46的结构域和结合细胞因子受体的结构域各自能够结合它们的相应nkp46、cd16a或细胞因子受体结合配偶体,当此类结合配偶体一起存在于细胞(例如nk细胞)的表面时。在一些实施方案中,多特异性蛋白质的特征可在于通过与nkp46单价结合(例如多特异性蛋白质仅包含一个nkp46 abd)、与感兴趣的抗原单价(或任选地二价)结合、与cd16a单价结合(例如多特异性蛋白质仅包含一个fc结构域二聚体),以及与细胞因子受体单价结合(例如多特异性蛋白质仅包含一个细胞因子受体abd)。
51、在一个实施方案中,多特异性蛋白质通过以下方式来构造:例如通过多特异性蛋白质内的结构域的布置或构型,任选地通过使用一个或多个结构域接头,这些结构域接头当处于拉伸构型时具有18埃(5个氨基酸残基)、36埃(10个残基)或54埃(15个残基)的最大潜在长度,使得结合nkp46的结构域和结合细胞因子受体的结构域以及当存在并能够结合cd16时的结合cd16的结构域可呈现膜平面结合构象,从而使得nkp46、cd16a和细胞因子受体中的每一者结合在nk细胞的表面。
52、多特异性蛋白质因此可被构造成使得结合细胞因子受体的结构域拓扑地置于多聚体蛋白质内,置于多特异性蛋白质内结合nkp46的结构域和结合cd16a的结构域两者的末端(例如c末端)。例如,可将细胞因子受体abd置于多特异性蛋白质的多肽链的c末端,使得该定位允许细胞因子受体abd在多聚体蛋白质内为拓扑c末端)。nkp46 abd和cd16a abd(例如,二聚体fc结构域)可在蛋白质中彼此拓扑地相邻定位,任选地经由短结构域接头彼此连接。nkp46 abd、cd16a abd和细胞因子(或其部分)因此可在蛋白质中(或在其多肽链上)串联连接或定位。有利地,该蛋白质包含二聚体fc结构域(fc结构域例如被指定为由置于单独的多肽链上的两个fc单体组成)。因此,在多特异性蛋白质中,nkp46 abd、二聚体fc结构域和细胞因子有利地彼此拓扑地相邻定位(在多特异性蛋白质的拓扑结构内)。在一个实施方案中,结合nkp46的结构域(或其部分,例如vh或vl)和fc结构域单体(或结合cd16a的结构域)彼此相邻地置于相同多肽链上,例如相邻的结合nkp46的结构域和结合cd16a的结构域可由结构域接头分开,但不需要任何居间蛋白质结构域(例如不需要与抗原结合的居间结构域),并且细胞因子部分可置于其c末端。
53、例如,在具有特别有利的nk细胞增强活性的优选多特异性蛋白质中,该多特异性蛋白质包含由定位在不同多肽链上的第一和第二fc结构域单体组成的fc结构域二聚体(其经由ch3-ch3缔合进行二聚化)。第一fc结构域单体可在其c末端与抗nkp46 abd(或其部分)融合,并且抗nkp46abd(或其部分)继而在其c末端与细胞因子融合。抗nkp46 abd的部分可以是例如((vh或vl)-ch1)单元或((vh或vl)-cl)单元,其中abd是fab。图2a至图2c、图2e、图2g至图2j、图2l、图2m示出了示例性结构域构型,其中抗nkp46 abd和细胞因子相对于n末端和c末端彼此拓扑地相邻并且经由一个fc结构域单体融合。图2d、图2f、图2k和图2n示出了其中抗nkp46 abd和细胞因子各自与不同fc结构域单体的c末端融合的结构域构型。
54、在任何实施方案中,可将结合细胞因子受体的结构域(细胞因子受体abd)、结合nkp46的结构域(nkp46 abd)和结合cd16的结构域(cd16 abd)指定为置于构成多特异性蛋白质的一条或多条多肽链内,使得这些结构域以其中它们在多聚体(例如异源多聚体)蛋白质上彼此相邻或从n末端至c末端串联的构型取向。结构域可任选地由结构域接头分开,例如具有5个至20个残基的连接肽,该连接肽本身不与预先确定的抗原结合。
55、在任何实施方案中,可将多特异性蛋白质指定为例如通过多特异性蛋白质内的结构域的布置或构型来构造,使得nkp46 abd和细胞因子受体abd(例如细胞因子部分)具有呈现它们在多特异性蛋白质分子内的fc结构域二聚体的相同侧或面的位置的能力,以便增强以膜平面结合构象结合nkp46、cd16a和细胞因子受体的能力。这种构型可容易地在本公开的异源二聚体、异源三聚体或异源四聚体蛋白质中的任一者中以任何方式实施,例如通过将nkp46 abd(或其部分,如果abd由两条多肽链缔合而成)和细胞因子受体abd(或其部分,如果abd由两条多肽链缔合而成)与fc结构域单体之一一起定位在相同多肽链上。
56、多特异性蛋白质可具有式(x1)-l1-(x2)-l2-(x3),其中x1和x2中的一者是nkp46abd(例如,fab、scfv、vhh)或其部分(例如,该部分可以是vh或vl、vh-ch1、vh-cl、vl-cl、vl-ch1)并且另一者是fc二聚体或其部分(例如,fc单体),并且x3是细胞因子,其中l1和l2各自是任选的结构域接头。细胞因子定位在其(或其部分)所在的多肽链的c末端。l1经由共价键(例如肽键)连接x1和x2。l2经由共价键(例如肽键)连接x2和x3。可将x1至x3任选地指定为从蛋白质的拓扑n末端至c末端排列。多特异性蛋白质还可包含结合感兴趣的抗原的abd,例如连接在n末端oif x1或定位在蛋白质的拓扑结构内的n末端。
57、如本文所证明的,应当理解,不同的元件x1、x2和x3(以及进一步结合感兴趣的抗原的abd)可各自容易地分布到蛋白质内的两条或更多条不同多肽链上。在一个实施方案中,多特异性蛋白质可任选地被表征为包含第一多肽链,该第一多肽链包含式(x1)-l1-(x2)-l2-(x3),其中x1和x2中的一者是nkp46 abd或其部分并且另一者是fc单体,并且x3是细胞因子或其部分,其中l1和l2各自是任选的结构域接头。因此,细胞因子可定位在其(或其部分)所在的多肽链的c末端。多特异性蛋白质还可包含结合感兴趣的抗原的abd,其中abd(或其部分)置于第一多肽链上,或置于与第一多肽链(或与蛋白质的任何其他链)缔合(例如二聚化)的单独多肽链上。然后,蛋白质可包含一条、两条或更多条附加多肽链,这些多肽链提供了以下项的互补结构域:nkp46 abd(当nkp46 abd是abd的一部分时)、fc单体(以便形成fc二聚体)、细胞因子(其中该细胞因子是一种细胞因子的一部分)和/或结合感兴趣的抗原的abd。因此,此类附加多肽链可经由非共价相互作用和任选地进一步的共价相互作用与蛋白质的第一或其他多肽链缔合(例如二聚化)。
58、在另一个实施方案中,多特异性蛋白质可任选地被表征为包含:(i)第一多肽链,该第一多肽链包含式(x1)-l1-(x2a),其中x1是第一fc单体并且x2a是nkp46 abd的第一部分;和(ii)第二多肽链,该第二多肽链包含式(x2b)-l2-(x3),其中x2b是nkp46 abd的第二部分(例如vh或vl、vh-ch1、vh-cl、vl-cl、vl-ch1),该第二部分与x2a缔合形成nkp46abd,并且x3是细胞因子或其部分。l1和l2各自是结构域接头。多特异性蛋白质还可包含结合感兴趣的抗原的abd,其中abd(或其部分)置于第一多肽链上(例如x1的n末端),或置于与第一多肽链(或与蛋白质的任何其他链)缔合(例如二聚化)的单独多肽链上。多特异性蛋白质还可包含结合cd16a的abd,任选地,abd是二聚体fc结构域;可将结合cd16a(或其部分)的abd置于第一多肽链上(例如x1的n末端),并且当abd是二聚体fc结构域时,可将fc单体之一置于第一多肽链上,并且可将第二fc单体置于与第一多肽链缔合(例如二聚化)的单独多肽链上。
59、在一个实施方案中,结合nkp46的结构域结合nkp46,使得当与细胞表面处的nkp46结合时,多特异性蛋白质的结合nkp46的结构域距细胞膜约70埃。在一个此类实施方案中,以掺入nkp46-1 vh/vl对的cdr的蛋白质为例,结合nkp46的结构域与nkp46多肽的d1/d2结合部结合。任选地,与野生型nkp46多肽相比,结合nkp46的结构域显示与nkp46突变体2(在残基k41、e42和e119处具有突变)和突变体supp7(在残基y121和y194处具有突变)的结合减少。在一个实施方案中,与野生型nkp46多肽相比,nkp46抗原结合结构域可被表征为显示与具有突变k41、e42、e119、y121和y194中的一者、两者、三者、四者或五者的nkp46突变体多肽的结合减少。
60、在一个实施方案中,结合nkp46的结构域结合nkp46,使得当与细胞表面处的nkp46结合时,多特异性蛋白质的结合nkp46的结构域距细胞膜小于约70埃,任选地小于约50埃。在一个此类实施方案中,以掺入nkp46-4 vh/vl对的cdr的蛋白质为例,结合nkp46的结构域与nkp46多肽的d1结构域结合,与d1/d2结合部相比,该nkp46多肽的d1结构域被定位成更接近nk细胞膜。任选地,与野生型nkp46多肽相比,结合nkp46的结构域显示与nkp46突变体6(在残基r101和v102处具有突变)和突变体supp6(在残基e104和l105处具有突变)的结合减少。在一个实施方案中,与野生型nkp46多肽相比,nkp46抗原结合结构域可被表征为显示与具有突变r101、v102、e104和l105中的一者、两者、三者或四者的nkp46突变体多肽的结合减少。在一个实施方案中,多特异性蛋白质包含具有至少10个氨基酸残基的结构域接头,该结构域接头位于在d1结构域内结合的结合nkp46的结构域与细胞因子之间。
61、在另一个实施方案中,以掺入nkp46-3 vh/vl对的cdr的蛋白质为例,结合nkp46的结构域与nkp46多肽的d2结构域结合,与d1/d2结合部相比,该nkp46多肽的d2结构域被定位成更接近nk细胞膜。任选地,与野生型nkp46多肽相比,结合nkp46的结构域显示与nkp46突变体19(在残基i135和s136处具有突变)和突变体supp8(在残基p132和e133处具有突变)的结合减少。在一个实施方案中,与野生型nkp46多肽相比,nkp46抗原结合结构域可被表征为显示与具有突变i135、s136、p132和e133中的一者、两者、三者或四者的nkp46突变体多肽的结合减少。在一个实施方案中,多特异性蛋白质包含具有至少10个氨基酸残基的结构域接头,该结构域接头位于在d2结构域内结合的结合nkp46的结构域与细胞因子之间。
62、在本文的任何实施方案中,多特异性蛋白质的特征可在于仅具有一个细胞因子受体结合结构域。
63、在本文的任何实施方案中,多特异性蛋白质的特征可在于仅具有一个结合nkp46的结构域。
64、某些示例性异源多聚体蛋白质可包含以下结构1a或1b的一般结构域组织,其中cd16 adc(例如fc结构域)和nkp46 abd在蛋白质内彼此紧邻,并且nkp46 abd紧邻细胞因子受体abd(体现为细胞因子(cyt)),并且其中nkp46 abd插置在cd16 abd与cyt之间:
65、(cd16 abd)(nkp46 abd)(cyt)(结构1a)
66、或
67、(fc结构域二聚体)(nkp46 abd)(cyt) (结构1b)
68、其中nkp46 abd和cd16abd(例如fc结构域二聚体)通过结构域接头连接,并且nkp46 abd和cyt通过结构域接头连接。
69、nkp46 abd可以方便地是fab、单域抗体或scfv。cd16 abd可以是fc结构域、fc结构域二聚体、人igg1亚型的fc结构域。cyt可以是例如il-2、il-15、il-21、il-7、il-27、il-12、il-18、ifn-α或ifn-β)多肽,任选地其中该多肽是与野生型人细胞因子对应物相差至少一个残基的变体细胞因子。
70、结构1a的蛋白质可包含与靶细胞上的感兴趣的抗原结合的abd(抗原abd),该抗原abd置于cd16 abd(例如fc结构域二聚体)的末端(例如n末端),如在具有以下结构1c或1d的异源多聚体蛋白质中:
71、(抗原abd)n(cd16 abd)(nkp46 abd)(cyt) (结构1c)
72、或
73、(抗原abd)n(fc结构域二聚体)(nkp46 abd)(cyt) (结构1d)其中“n”为1或2,并且抗原abd和cd16 abd(例如fc结构域二聚体)通过接头连接,任选地其中接头是免疫球蛋白铰链多肽,其中cd16 abd和nkp46 abd通过接头连接,并且nkp46 abd和cyt通过接头连接。当“n”为2时,结构1d也可表示为结构1e:
74、(抗原abd)
75、(fc结构域二聚体)(nkp46 abd)(cyt) (结构1e)
76、(抗原abd)
77、在任何实施方案中,多特异性蛋白质可被表征为与nkp46多肽和细胞因子受体中的每一者以单价结合,并且能够指导表达nkp46的nk细胞裂解表达感兴趣的抗原的靶细胞。有利地,在一个实施方案中,在存在nk细胞和靶细胞的情况下,多特异性蛋白质可结合(i)靶细胞上的感兴趣的抗原、(ii)nk细胞上的nkp46、(iii)nk细胞上的cd16a和(iv)nk细胞上的细胞因子受体(例如cd122、il-21r、il-7ra、il-27ra、il-12r、il-18r、ifnar),当与靶细胞和nk细胞上的这些蛋白质结合时,可通过nkp46(该蛋白质充当nkp46激动剂)和细胞因子受体(该蛋白质充当细胞因子受体激动剂)诱导nk细胞中的信号传导和/或nk细胞的活化,从而促进nk细胞的活化和/或靶细胞的裂解,特别是经由nkp46传递的活化信号。
78、在一个实施方案中,在存在nk细胞和靶细胞的情况下,多特异性蛋白质可诱导nk细胞中的细胞因子受体途径信号传导的细胞毒性(如通过stat信号传导评估的)和/或nk细胞的活化,其中这种细胞毒性、活化和/或信号传导比在不存在靶细胞的情况下使多特异性蛋白质与nk细胞接触时所观察到的细胞毒性、活化和/或信号传导更大(例如,低到至少1/100或1/1000的ec50值)。
79、任选地,多特异性蛋白质可结合nk细胞上的nkp46和cd122(例如,该蛋白质包含il2或il15部分,任选地与cd25的结合减少的经修饰的或变体il2或il15),并且当与nkp46和cd122两者结合时,可通过nkp46和cd122两者诱导nk细胞中的信号传导。任选地,多特异性蛋白质可结合nk细胞上的nkp46、cd16a和cd122,并且当与nkp46、cd16和cd122结合时,可通过nkp46、cd16a和cd122诱导nk细胞中的信号传导。可通过测量stat5来评估细胞因子受体信号传导,任选地其中所观察到的信号传导大于使用其中结合nkp46的结构域被对照abd(例如,其不与测定系统中存在的任何蛋白质结合)取代的比较蛋白质所观察到的信号传导。
80、任选地,多特异性蛋白质可结合nk细胞上的nkp46和il-21r(例如,该蛋白质包含il21部分),并且当与nkp46和il-21r两者结合时,可通过nkp46和il-21r两者诱导nk细胞中的信号传导。任选地,多特异性蛋白质可结合nk细胞上的nkp46、cd16a和il-21r,并且当与nkp46、cd16a和il-21r结合时,可通过nkp46、cd16a和il-21r诱导nk细胞中的信号传导。任选地,通过测量stat3来评估细胞因子信号传导,其中所观察到的信号传导大于使用其中结合nkp46的结构域被对照abd(例如,其不与测定系统中存在的任何蛋白质结合)取代的比较蛋白质所观察到的信号传导。
81、任选地,多特异性蛋白质可结合nk细胞上的nkp46和il-18r(例如,该蛋白质包含il18部分),并且当与nkp46和il-18r两者(il-18rα和/或il-18rβ)结合时,可通过nkp46和il-18r两者诱导nk细胞中的信号传导。任选地,多特异性蛋白质可结合nk细胞上的nkp46、cd16a和il-18r,并且当与nkp46、cd16a和il-18r结合时,可通过nkp46、cd16a和il-18r诱导nk细胞中的信号传导。任选地,通过测量stat3来评估细胞因子信号传导,其中所观察到的信号传导大于使用其中结合nkp46的结构域被对照abd(例如,其不与测定系统中存在的任何蛋白质结合)取代的比较蛋白质所观察到的信号传导。
82、任选地,多特异性蛋白质可结合nk细胞上的nkp46和il-7r(例如il-7rα(cd127)和/或cd132)(例如,该蛋白质包含il-7部分),并且当与nkp46和il-7r两者结合时,可通过nkp46和il-7rα两者诱导nk细胞中的信号传导。任选地,多特异性蛋白质可结合nk细胞上的nkp46、cd16a和il-7r,并且当与nkp46、cd16a和il-7r结合时,可通过nkp46、cd16a和il-7r诱导nk细胞中的信号传导。任选地,通过测量stat5来评估细胞因子信号传导,其中所观察到的信号传导大于使用其中结合nkp46的结构域被对照abd(例如,其不与测定系统中存在的任何蛋白质结合)取代的比较蛋白质所观察到的信号传导。
83、任选地,多特异性蛋白质可结合nk细胞上的nkp46和il-27r(例如il-27rα和/或gp130)(例如,该蛋白质包含il-27部分),并且当与nkp46和il-27r两者结合时,可通过nkp46和il-27r两者诱导nk细胞中的信号传导。任选地,多特异性蛋白质可结合nk细胞上的nkp46、cd16a和il-27r,并且当与nkp46、cd16a和il-27r结合时,可通过nkp46、cd16a和il-27r诱导nk细胞中的信号传导。经由nkp46和/或cd16a的信号传导可通过nk细胞活化的标志物(例如在实施例中使用的标志物、cd69表达等)来评估。任选地,通过测量stat1来评估细胞因子信号传导,其中所观察到的信号传导大于使用其中结合nkp46的结构域被对照abd(例如,其不与测定系统中存在的任何蛋白质结合)取代的比较蛋白质所观察到的信号传导。
84、任选地,多特异性蛋白质可结合nk细胞上的nkp46和il-12r(例如il-12rβ1和/或il-12rβ2)(例如,该蛋白质包含il-27部分),并且当与nkp46和il-12r两者结合时,可通过nkp46和il-12r两者诱导nk细胞中的信号传导。任选地,多特异性蛋白质可结合nk细胞上的nkp46、cd16a和il-12r,并且当与nkp46、cd16a和il-12r结合时,可通过nkp46、cd16a和il-12r诱导nk细胞中的信号传导。任选地,通过测量stat4来评估细胞因子信号传导,其中所观察到的信号传导大于使用其中结合nkp46的结构域被对照abd(例如,不与测定系统中存在的任何蛋白质结合)取代的比较蛋白质所观察到的信号传导。
85、任选地,多特异性蛋白质可结合nk细胞上的nkp46和ifnar,并且当与nkp46和ifnar(ifnar1和/或ifnar2)两者结合时,可通过nkp46和ifnar两者诱导nk细胞中的信号传导。例如,多特异性蛋白质可包含ifn-α或ifn-β部分)任选地,多特异性蛋白质可结合nk细胞上的nkp46、cd16a和ifnar,并且当与nkp46、cd16a和ifnar两者结合时,可通过nkp46、cd16a和ifnar诱导nk细胞中的信号传导。任选地,通过测量stat(例如,stat1、stat2或ifn调节因子(irf)-9)来评估细胞因子信号传导,其中所观察到的信号传导大于使用其中结合nkp46的结构域被对照abd(例如,其不与测定系统中存在的任何蛋白质结合)取代的比较蛋白质所观察到的信号传导。
86、经由nkp46和/或cd16a的信号传导可通过nk细胞活化的标志物(例如在实施例中使用的标志物、cd69表达等)来评估。
87、在一些实施方案中,多特异性蛋白质包含人fc结构域的至少一部分,例如足以使得fc结构域与人fcrn多肽结合的部分,任选地其中如通过spr评估的所述fcrn结合亲和力在常规人igg1抗体的fcrn结合亲和力的1-log内。
88、多特异性蛋白质有利地能够有效地动员cd16+和cd16-nk细胞两者(所有nk细胞是nkp46+)。
89、在一个方面,多特异性蛋白质包含两条或更多条多肽链,即,该多特异性蛋白质包含多链蛋白质(也称为多聚体蛋白质)。例如,多特异性蛋白质或多链蛋白质可以是异源二聚体、异源三聚体或异源四聚体,或者可包含多于四条多肽链。
90、任何抗原结合结构域(例如结合感兴趣的抗原(例如肿瘤抗原)、nkp46或细胞因子受体的abd)可完整包含在单一多肽链上,例如作为scfv或单抗原结合结构域(诸如sdab或纳米抗体、vnar或vhh结构域)或蛋白质模块)。另选地,抗原结合结构域可由置于单独多肽链上的两个或更多个蛋白质结构域组成,使得当两个或更多个互补蛋白质结构域(例如作为vh/vl对)缔合在多聚体蛋白质中时,抗原结合结构域结合其靶标。
91、多特异性蛋白质可以单价或多价方式结合感兴趣的抗原(例如癌抗原)。当多特异性蛋白质以单价结合感兴趣的抗原、以单价结合nk46并且以单价结合细胞因子受体时,可将多特异性蛋白质指示为具有1:1:1构型。当多特异性蛋白质以二价结合感兴趣的抗原、以单价结合nk46并且以单价结合细胞因子受体时,可将多特异性蛋白质可指示为具有2:1:1构型。不同的1:1:1和2:1:1构型的代表性示例示于图2中。
92、在任何方面,多特异性蛋白质可被表征为具有这样的结构:其中一个或多个抗原结合结构域(abd)的运动自由度(链内结构域运动)或灵活性增加,例如与常规人igg抗体的abd相比。在一个实施方案中,提供了一种多特异性蛋白质,其包含这样的结构:该结构允许第一抗原结合结构域的抗原结合位点和第二抗原结合结构域的抗原结合位点分开一定距离,该距离导致增强的功能,例如多特异性蛋白质诱导nkp46信号传导和裂解靶细胞的能力,例如任选地小于80埃的距离。其中abd具有更大的灵活性和/或以优化的距离分开的多特异性蛋白质可增强裂解性nkp46-靶突触的形成,从而增强nkp46介导的信号传导。这种灵活性和/或运动结构域可通过使用接头(例如,基于柔性氨基酸的接头)而容易地实现,这些接头将结合nkp46的结构域与fc结构域(例如,fc结构域二聚体,或更一般地,多特异性蛋白质的其余部分)分开。
93、在任何方面,多特异性蛋白质可被表征为具有增加的抗原结合结构域的运动自由度(例如,与常规人igg抗体(例如人igg1抗体)的abd相比)。此类蛋白的一个示例是含有多聚体fc结构域的蛋白质(例如异源二聚体或异源三聚体),其中抗原结合结构域(例如结合nkp46的abd)经由柔性接头与fc结构域连接或融合。接头可通过赋予弯曲能力从而潜在地减小接头处的abd与fc结构域之间(或两个abd之间)的角度来提供一个或多个abd的灵活性或运动自由度。任选地,两个抗原结合结构域(并且如果附加的abd存在于多特异性蛋白质中,则任选地更多个抗原结合结构域)经由接头(通常为柔性肽接头)与fc结构域连接或融合。任选地,具有增加的运动自由度的蛋白质允许蛋白质采取这样的构象:其中nkp46结合位点与感兴趣的抗原结合位点之间的距离小于在其中两个结合结构域都是fab的蛋白质中观察到的距离,或者小于在全长抗体中观察到的距离。
94、abd可经由结构域接头连接至fc结构域单体(或其ch2或ch3结构域)。接头可以是多肽接头,例如包含至少5个残基、至少10个残基、至少15个残基、至少20个残基或更多个残基的长度的肽接头。在其他实施方案中,接头包含介于2个至4个残基之间、介于2个至5个残基之间、介于2个至6个残基之间、介于2个至8个残基之间、介于5个至10个残基之间、介于2个至15个残基之间、介于4个至15个残基之间、介于3个至15个残基之间、介于5个至15个残基之间、介于10个至15个残基之间、介于4个至20个残基之间、介于5个至20个残基之间、介于2个至20个残基之间、介于10个至30个残基之间、或介于10个至50个残基之间的长度。任选地,接头包含源自抗体恒定区的氨基酸序列,例如来自ch1或cκ结构域的氨基酸序列(例如来自ch1或cκ结构域的n末端序列),或源自铰链的氨基酸序列。任选地,接头包含氨基酸序列rtva。任选地,接头是柔性接头,其主要或仅由甘氨酸和/或丝氨酸残基组成,例如包含氨基酸序列(gxs)n,其中g为1、2、3或4,并且n为1至10、1至6或1至4的整数。任选地,接头包含1个至20个或1个至10个另外的氨基酸残基。
95、在一个实施方案中,提供了具有多肽链1、2和3的异源三聚体:
96、
97、其中:
98、va-1、vb-1、va-2和vb-2各自是vh结构域或vl结构域,其中va-1和vb-1结合感兴趣的抗原(任选地,其中va-1和vb-1中的一者是vh并且另一者是vl,并且其中va-1和vb-1形成结合感兴趣的抗原的第一抗原结合结构域(abd)),其中va-2和vb-2中的一者是vh并且另一者是vl,并且其中va-2和vb-2形成结合nkp46的第二abd;
99、ch1是人免疫球蛋白ch1结构域,并且cl是人轻链恒定结构域;
100、(ch1或cl)a和(ch1或cl)c中的一者是ch1并且另一者是cl,从而形成(ch1/cl)对;
101、(ch1或cl)b和(ch1或cl)d中的一者是ch1并且另一者是cl,从而形成(ch1/cl)对;
102、铰链是免疫球蛋白铰链区或其部分;
103、l1和l2各自是氨基酸结构域接头,其中l1和l2能够是不同的或相同的;
104、ch2和ch3分别是人免疫球蛋白ch2和ch3结构域;并且
105、cyt是与nk细胞上存在的细胞因子受体结合的细胞因子多肽或其部分,任选地其中cyt是野生型或变体人il-2、il-15、il-21、il-7、il-27、il-12、il-18、ifn-α或ifn-β多肽。在可使用相同结构域和结构域接头制备的另一个实施方案中,提供了具有多肽链1、2和3的异源三聚体:
106、
107、在一个实施方案中,提供了具有多肽链1和2的异源二聚体:
108、
109、其中:
110、va-1、vb-1、va-2和vb-2各自是vh结构域或vl结构域,其中va-1和vb-1结合感兴趣的抗原(任选地,其中va-1和vb-1中的一者是vh并且另一者是vl,并且其中va-1和vb-1形成结合感兴趣的抗原的第一抗原结合结构域(abd)),其中va-2和vb-2中的一者是vh并且另一者是vl,并且其中va-2和vb-2形成结合nkp46的第二abd;
111、ch1是人免疫球蛋白ch1结构域,并且cl是轻链恒定结构域;
112、(ch1或cl)a和(ch1或cl)b中的一者是ch1并且另一者是cl,从而形成(ch1/cl)对;
113、铰链是免疫球蛋白铰链区或其部分;
114、l1、l2和l3各自是氨基酸结构域接头,其中l1、l2和l3能够是不同的或相同的;
115、ch2和ch3分别是人免疫球蛋白ch2和ch3结构域;并且
116、cyt是与nk细胞上存在的细胞因子受体结合的细胞因子多肽或其部分,任选地其中cyt是野生型或变体人il-2、il-15、il-21、il-7、il-27、il-12、il-18、ifn-α或ifn-β多肽。
117、本公开还提供了如本文进一步所述的另外的异源二聚体、异源三聚体和异源四聚体多特异性分子和结构域排列。在一个方面,多特异性蛋白质是具有如图2a至图2n中任一者所示的结构或结构域排列的异源多聚体、异源二聚体、异源三聚体、异源四聚体。
118、在本文所述的任何实施方案的一个方面,可将abd(例如,抗nkp46abd、结合感兴趣的抗原或肿瘤抗原的abd)指定为包含免疫球蛋白重链可变结构域(vh)和免疫球蛋白轻链可变结构域(vl),其中每个vh和vl包含三个互补决定区(cdr-1至cdr-3)。在本文所述的任何实施方案的一个方面,可将vh指定为具有人vh结构域的氨基酸序列。在本文所述的任何实施方案的一个方面,可将vl指定为具有人vl结构域的氨基酸序列。
119、在任何实施方案的一个方面,vh区包含与由人v基因组的基因编码的氨基酸序列具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列,该人v基因组选自ighv1-18、ighv1-2、ighv1-24、ighv1-3、ighv1-45、ighv1-46、ighv1-58、ighv1-69、ighv1-69-2、ighv1-69d、ighv1-8、ighv2-26、ighv2-5、ighv2-70、ighv2-70d、ighv3-11、ighv3-13、ighv3-15、ighv3-20、ighv3-21、ighv3-23、ighv3-23d、ighv3-30、ighv3-30-3、ighv3-30-5、ighv3-33、ighv3-43、ighv3-43d、ighv3-48、ighv3-49、ighv3-53、ighv3-62、ighv3-64、ighv3-64d、ighv3-66、ighv3-7、ighv3-72、ighv3-73、ighv3-74、ighv3-9、ighv3-nl1、ighv4-28、ighv4-30-2、ighv4-30-4、ighv4-31、ighv4-34、ighv4-38-2、ighv4-39、ighv4-4、ighv4-59、ighv4-61、ighv5-10-1、ighv5-51、ighv6-1、ighv7-4-1、ighv1-38-4、ighv1/or15-1、ighv1/or15-5、ighv1/or15-9、ighv1/or21-1、ighv2-70、ighv2/or16-5、ighv3-16、ighv3-20、ighv3-25、ighv3-35、ighv3-38、ighv3-38-3、ighv3/or15-7、ighv3/or16-10、ighv3/or16-12、ighv3/or16-13、ighv3/or16-17、ighv3/or16-20、ighv3/or16-6、ighv3/or16-8、ighv3/or16-9、ighv4-61、ighv4/or15-8、ighv7-81和ighv8-51-1。任选地,vh区包含有包含来自所述基因的氨基酸序列(例如,cdr和/或人框架区,例如根据kabat编号)的vh。在任何实施方案的一个方面,vh区包含与seq id no:236-313的氨基酸序列具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列。
120、在任何实施方案的一个方面,vl区包含与由人v基因组的基因编码的氨基酸序列具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列,该人v基因组选自igkv1-12、igkv1-13、igkv1-16、igkv1-17、igkv1-27、igkv1-33、igkv1-39、igkv1-5、igkv1-6、igkv1-8、igkv1-9、igkv1-nl1、igkv1d-12、igkv1d-13、igkv1d-16、igkv1d-17、igkv1d-33、igkv1d-43、igkv1d-8、igkv2-24、igkv2-28、igkv2-29、igkv2-30、igkv2-40、igkv2d-26、igkv2d-28、igkv2d-29、igkv2d-30、igkv2d-40、igkv3-11、igkv3-15、igkv3-20、igkv3d-11、igkv3d-15、igkv3d-20、igkv3d-7、igkv4-1、igkv5-2、igkv6-21、igkv6d-21、igkv1-37、igkv1/or2-0、igkv1/or2-108、igkv1d-37、igkv1d-42、igkv2d-24、igkv3-7、igkv3/or2-268、igkv3d-20、igkv6d-41、iglv1-36、iglv1-40、iglv1-44、iglv1-47、iglv1-51、iglv10-54、iglv2-11、iglv2-14、iglv2-18、iglv2-23、iglv2-8、iglv3-1、iglv3-10、iglv3-12、iglv3-16、iglv3-19、iglv3-21、iglv3-22、iglv3-25、iglv3-27、iglv3-9、iglv4-3、iglv4-60、iglv4-69、iglv5-37、iglv5-39、iglv5-45、iglv5-52、iglv6-57、iglv7-43、iglv7-46、iglv8-61、iglv9-49、iglv1-41、iglv1-50、iglv11-55、iglv2-33、iglv3-32、iglv5-48和iglv8/or8-1。任选地,vl区包含有包含来自所述基因的氨基酸序列(例如,cdr和/或人框架区,例如根据kabat编号)的vl。在任何实施方案的一个方面,vl区包含与seqid no:314-403的氨基酸序列具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列。
121、在本文所述的任何实施方案的一个方面中,abd包含scfv或fab,其中该scfv包含vh、结构域接头和vl,该vh包含与选自seq id no:3、5、7、9、11、13、112、113、115、116、117、119、120、121、123、124、125、127、128、129、132、134、136、138、140、142、144、146、148、150、152、154和236-313中任一项的序列至少90%相同的氨基酸序列,该vl包含与选自seq idno:4、6、8、10、12、14、114、118、122、126、130、133、135、137、139、141、143、145、147、149、151、153、155和314-403中任一项的序列至少90%相同的氨基酸序列;并且其中该fab包含一个vh、一个vl、一个人ch1结构域和一个人cl结构域,该一个vh包含与选自seq id no:3、5、7、9、11、13、112、113、115、116、117、119、120、121、123、124、125、127、128、129、132、134、136、138、140、142、144、146、148、150、152、154和236-313中任一项的序列至少90%相同的氨基酸序列,该一个vl包含与选自seq id no:4、6、8、10、12、14、114、118、122、126、130、133、135、137、139、141、143、145、147、149、151、153、155和314-403中任一项的序列至少90%相同的氨基酸序列,该一个人ch1结构域包含与seq id no:156至少90%相同的氨基酸序列,该一个人cl结构域包含与seq id no:159至少90%相同的氨基酸序列,其中该vh与该ch1结构域或该cl结构域中的一者融合,并且该vl与该ch1结构域或该cl结构域中的另一者融合。
122、在本文所述的任何实施方案的一个方面,il2包含与seq id no:404-417中任一项的il-2多肽或与其至少40个、50个、60个、70个、80个或100个氨基酸残基的连续序列具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列。任选地,il2还包含2、3、4、5个或更多个减少与cd25结合的氨基酸取代,例如在本公开的任何残基处的取代。
123、在本文所述的任何实施方案的一个方面,il15包含与seq id no:418中任一项的il-15多肽或与其至少40个、50个、60个、70个、80个或100个氨基酸残基的连续序列具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列。
124、在本文所述的任何实施方案的一个方面,il12包含与seq id no:438和/或439中任一项的il-12多肽或与其至少40个、50个、60个、70个、80个或100个氨基酸残基的连续序列具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列。
125、在本文所述的任何实施方案的一个方面,il7包含与seq id no:435中任一项的il-7多肽或与其至少40个、50个、60个、70个、80个或100个氨基酸残基的连续序列具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列。
126、在本文所述的任何实施方案的一个方面,il27包含与seq id no:436和/或437中任一项的il-21多肽或与其至少40个、50个、60个、70个、80个或100个氨基酸残基的连续序列具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列。
127、在本文所述的任何实施方案的一个方面,il21包含与seq id no:420或421中任一项的il-27多肽或与其至少40个、50个、60个、70个、80个或100个氨基酸残基的连续序列具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列。
128、在本文所述的任何实施方案的一个方面,il18包含与seq id no:422中任一项的il-18多肽或与其至少40个、50个、60个、70个、80个或100个氨基酸残基的连续序列具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列。
129、在本文所述的任何实施方案的一个方面,ifn-α与seq id no:423-433中任一项的ifn-α多肽或与其至少40个、50个、60个、70个、80个或100个氨基酸残基的连续序列具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列。
130、在本文所述的任何实施方案的一个方面,ifn-β与seq id nos:434中任一项的ifn-α多肽或与其至少40个、50个、60个、70个、80个或100个氨基酸残基的连续序列具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列。
131、在本文所述的任何实施方案的一个方面,fc结构域包含与seq id no:160-165中任一项的fc多肽或与其至少40个、50个、60个、70个、80个或100个氨基酸残基的连续序列具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列。
132、在本文所述的任何实施方案的一个方面,ch1、ch2和ch3结构域分别包含与seq idno:156、157或158的ch1多肽或与其至少40个、50个、60个、70个、80个或100个氨基酸残基的连续序列具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列。
133、在本文所述的任何实施方案的一个方面,ck或cl结构域包含与seq id no:159中任一项的ck多肽或与其至少40个、50个、60个、70个、80个或100个氨基酸残基的连续序列具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列。
134、在本文所述的任何实施方案的一个方面,铰链结构域包含与seq id no:166-170中任一项的ck多肽具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列。
135、在本文所述的任何实施方案的一个方面,多特异性蛋白质包含:
136、(a)与感兴趣的抗原结合的abd,其中该abd包含
137、vh、结构域接头和vl,该vh包含与选自seq id no:132、134、136、138、140、142、144、146、148、150、152、154和236-313中任一项的序列至少70%、80%或90%相同的氨基酸序列,该vl包含与选自seq id no:133、135、137、139、141、143、145、147、149、151、153、155和314-403中任一项的序列至少90%相同的氨基酸序列;和
138、(b)与人nkp46多肽结合的abd,其中该abd包含vh、结构域接头和vl,该vh包含与选自seq id no:3、5、7、9、11、13、112、113、115、116、117、119、120、121、123、124、125、127、128、129和236-313中任一项的序列至少70%、80%或90%相同的氨基酸序列,该vl包含与选自seq id no:4、6、8、10、12、14、114、118、122、126、130和314-403中任一项的序列至少90%相同的氨基酸序列;
139、(c)包含两个fc结构域单体多肽的fc结构域二聚体,其中每个fc结构域单体多肽包含与选自seq id no:160-165的序列至少70%、80%或90%相同的氨基酸序列;和
140、(d)细胞因子多肽,其包含与选自seq id no:404-436的序列或其至少40个、50个、60个、70个、80个或100个氨基酸残基的连续序列至少70%、80%或90%相同的氨基酸序列,该细胞因子多肽经由结构域接头与该多特异性蛋白质的多肽链中的一者的c末端融合。
141、在本文所述的任何实施方案的一个方面,铰链结构域包含与seq id no:166-170中任一项的ck多肽具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列。
142、在本文所述的任何实施方案的一个方面,多特异性蛋白质包含:
143、(a)与感兴趣的抗原结合的abd,其中该abd包含scfv或fab,
144、a.其中该scfv包含vh、结构域接头和vl,该vh包含与选自seq id no:132、134、136、138、140、142、144、146、148、150、152、154和236-313中任一项的序列至少90%相同的氨基酸序列,该vl包含与选自seq id no:133、135、137、139、141、143、145、147、149、151、153、155和314-403中任一项的序列至少90%相同的氨基酸序列;并且
145、b.其中该fab包含一个vh、一个vl、一个人ch1结构域和一个人cl结构域,该一个vh包含与选自seq id no:132、134、136、138、140、142、144、146、148、150、152、154和236-313中任一项的序列至少90%相同的氨基酸序列,该一个vl包含与选自seq id no:133、135、137、139、141、143、145、147、149、151、153、155和314-403中任一项的序列至少90%相同的氨基酸序列,该一个人ch1结构域包含与seq id no:156至少90%相同的氨基酸序列,该一个人cl结构域包含与seq id no:159至少90%相同的氨基酸序列,其中该vh与该ch1结构域或该cl结构域中的一者融合,并且该vl与该ch1结构域或该cl结构域中的另一者融合,
146、(b)与人nkp46多肽结合的abd,其中该abd包含scfv或fab,
147、a.其中该scfv包含vh、结构域接头和vl,该vh包含与选自seq id no:3、5、7、9、11、13、112、113、115、116、117、119、120、121、123、124、125、127、128、129和236-313中任一项的序列至少90%相同的氨基酸序列,该vl包含与选自seq id no:4、6、8、10、12、14、114、118、122、126、130和314-403中任一项的序列至少90%相同的氨基酸序列;并且
148、b.其中该fab包含一个vh、一个vl、一个人ch1结构域和一个人cl结构域,该一个vh包含与选自seq id no:3、5、7、9、11、13、112、113、115、116、117、119、120、121、123、124、125、127、128、129和236-313中任一项的序列至少90%相同的氨基酸序列,该一个vl包含与选自seq id no:4、6、8、10、12、14、114、118、122、126、130和314-403中任一项的序列至少90%相同的氨基酸序列,该一个人ch1结构域包含与seq id no:156至少90%相同的氨基酸序列,该一个人cl结构域包含与seq id no:159至少90%相同的氨基酸序列,其中该vh与该ch1结构域或该cl结构域中的一者融合,并且该vl与该ch1结构域或该cl结构域中的另一者融合,
149、(c)包含两个fc结构域单体多肽的fc结构域二聚体,其中每个fc结构域单体多肽包含与选自seq id no:160-165的序列至少或90%相同的氨基酸序列;和
150、(d)细胞因子多肽,其包含与选自seq id no:404-436的序列或其至少40个、50个、60个、70个、80个或100个氨基酸残基的连续序列至少90%相同的氨基酸序列,该细胞因子多肽经由结构域接头与该多特异性蛋白质的多肽链中的一者的c末端融合。
151、在一个实施方案中,多特异性蛋白质包含:包含与本文所述异源三聚体蛋白质的第一链的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽,包含与本文所述异源三聚体蛋白质的第二链的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽,以及包含与本文所述异源三聚体蛋白质的第三链的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽。在一个实施方案中,多特异性蛋白质包含:包含与本文所述异源二聚体蛋白质的第一链的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽,以及包含与本文所述异源二聚体蛋白质的第二链的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽。
152、在一个实施方案中,多特异性蛋白质包含:包含本文所述异源三聚体蛋白质的第一链的氨基酸序列的多肽,包含本文所述异源三聚体蛋白质的氨基酸序列的多肽,以及包含本文所述异源三聚体蛋白质的第三链的氨基酸序列的多肽。在一个实施方案中,多特异性蛋白质包含:包含本文所述异源二聚体蛋白质的第一链的氨基酸序列的多肽,以及包含本文所述异源二聚体蛋白质的第二链的氨基酸序列的多肽。
153、在一个实施方案中,多特异性蛋白质包含:包含与seq id no:175的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽,包含与seq id no:176的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽,以及包含与seq id no:177的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽。
154、在一个实施方案中,多特异性蛋白质包含:包含与seq id no:193的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽,包含与seq id no:185的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽;以及包含与seq id no:194的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽。
155、在一个实施方案中,多特异性蛋白质包含:包含与seq id no:199的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽,包含与seq id no:200的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽;以及包含与seq id no:201的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽。
156、在一个实施方案中,多特异性蛋白质包含:包含与seq id no:209的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽,包含与seq id no:210的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽;以及包含与seq id no:211的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽。
157、在一个实施方案中,多特异性蛋白质包含:包含与seq id no:212的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽,包含与seq id no:213的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽;以及包含与seq id no:214的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽。
158、在一个实施方案中,多特异性蛋白质包含:包含与seq id no:215的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽,包含与seq id no:216的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽;以及包含与seq id no:217的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽。
159、在一个实施方案中,多特异性蛋白质包含:包含与seq id no:218的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽,包含与seq id no:219的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽;以及包含与seq id no:220的氨基酸序列具有至少80%、90%或95%序列同一性的氨基酸序列的多肽。
160、在一个方面,本发明提供了分离的多特异性异源三聚体蛋白质,其包含第一多肽链、第二多肽链和任选的第三多肽链,该第一多肽链包含与本文公开的t5、t6、t25或t26蛋白质的第一多肽链的序列至少50%、60%、70%、80%、85%、90%、95%、98%或99%相同的氨基酸序列;该第二多肽链包含与本文公开的相应t5、t6、t25或t26蛋白质的第二多肽链的序列至少50%、60%、70%、80%、85%、90%、95%、98%或99%相同的氨基酸序列;该第三多肽链包含与本文公开的t5、t6、t25或t26蛋白质的第三多肽链的序列至少50%、60%、70%、80%、85%、90%、95%、98%或99%相同的氨基酸序列。在一个实施方案中,cdr被排除在考虑用于计算序列同一性的序列之外。在一个实施方案中,vh和/或vl可变区域被排除在考虑用于计算多肽链序列同一性的序列之外。任选地,每个vh区包含与seq id no:3、5、7、9、11、13、112、113、115、116、117、119、120、121、123、124、125、127、128、129、132、134、136、138、140、142、144、146、148、150、152、154或236-313中任一项的氨基酸序列具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列。任选地,每个vl区包含与seq id no:4、6、8、10、12、14、114、118、122、126、130、133、135、137、139、141、143、145、147、149、151、153、155或314-403中任一项的氨基酸序列具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列。
161、在本文所述的任何实施方案的一个方面,提供了编码第一多肽链和/或第二多肽链和/或第三多肽链和/或第四多肽的重组核酸。在本文所述的任何实施方案的一个方面,本发明提供了包含编码第一多肽链和/或第二多肽链和/或第三多肽链的核酸的重组宿主细胞,任选地其中该宿主细胞以至少1mg/l、2mg/l、3mg/l或4mg/l的产率(纯化之前或之后的最终生产率或浓度)产生根据本发明的多聚体或其他蛋白质。还提供了核酸试剂盒或核酸组,其包含编码根据本发明的第一多肽链的重组核酸、编码根据本发明的第二多肽链的重组核酸,以及任选的编码根据本发明的第三多肽链的重组核酸。还提供了制备根据本发明的二聚体、三聚体和四聚体蛋白质的方法。
162、在另一个实施方案中,本公开提供了新型变体il-2多肽,其特别适用于抗原结合蛋白,特别是具有fc结构域的抗原结合蛋白,更进一步地,其中该fc结构域经修饰以降低或消除结合cd16a和/或其他fcγ受体的能力。本文提供了包含氨基酸取代t41a的经修饰的或变体il-2多肽,其中残基的编号是相对于seq id no:404的il-2多肽而言的。在一个实施方案中,本文提供了包含至少两个、三个、四个、五个、六个或更多个氨基酸取代的经修饰的或变体il-2多肽,其中该多肽包含取代t41a。在一个实施方案中,本文提供了包含至少三个氨基酸取代的经修饰的或变体il-2多肽,其中与人野生型il-2多肽相比,该多肽包含残基r38、f42和t41处的取代,其中残基数量是相对于seq id no:404的il-2多肽而言的。在一个方面,与人野生型il-2多肽相比,经修饰的或变体il-2多肽包含至少三个氨基酸取代,其中该多肽包含以下取代:r38a、f42k和t41x,其中x是任何氨基酸残基。在一个方面,与人野生型il-2多肽相比,经修饰的或变体il-2多肽包含至少三个(例如3个、4个、5个、6个或更多个)氨基酸取代,其中该多肽包含以下取代:r38a、f42k和t41a。在一个方面,经修饰的或变体il2包含seq id no:408的氨基酸序列。在一个方面,经修饰的或变体il2包含与seq idno:408的il-2多肽或与其至少40个、50个、60个、70个、80个或100个氨基酸残基的连续序列具有至少约80%、85%、90%、95%、97%、98%或99%同一性的氨基酸序列。还提供了包含上述经修饰的或变体il-2多肽的蛋白质或多肽。在一个实施方案中,提供了与异源氨基酸序列或多肽融合(例如,在其n末端)的经修饰的或变体il-2多肽。在一个实施方案中,提供了结合感兴趣的抗原(例如,癌抗原或免疫细胞(任选的nk细胞)上的受体)的多肽或蛋白质(例如含fc结构域的蛋白质、抗原结合蛋白、多特异性蛋白质或抗体),该多肽或蛋白质包含经修饰的或变体il-2多肽,任选地其中il-2多肽经由结构域接头与多肽或蛋白质融合,任选地其中il-2多肽经由结构域接头与蛋白质或多肽的c末端融合。在一个实施方案中,包含变体il-2的蛋白质或多肽包含人fc结构域,更进一步地,其中该fc结构域经修饰以降低或消除结合cd16a和/或其他fcγ受体的能力。在一些实施方案中,相对于野生型il-2,经修饰的il-2具有降低的对cd25的结合亲和力。在一些实施方案中,相对于野生型il-2,经修饰的il-2具有降低的对静息或活化的t细胞的活性。
163、任何方法可进一步被表征为包括描述于本技术中的任何步骤,特别包括描述于“本发明的具体实施方式”中的步骤。本发明还涉及鉴定、测试和/或制备本文所述的蛋白质的方法。本发明还涉及可通过本发明的任何方法获得的多特异性蛋白质。本公开还涉及含有本文公开的多特异性蛋白质中的至少一种的药物或诊断制剂。本公开还涉及在治疗或诊断方法中使用本发明的多特异性蛋白质的方法。
164、本发明的这些和另外的有利方面和特征可在本文的其他地方进一步描述。
- 还没有人留言评论。精彩留言会获得点赞!