一种无细胞催化合成亚精胺的生物方法
本发明涉及一种生物法合成亚精胺的方法,属于生物工程。
背景技术:
1、亚精胺(spermidine)是一种天然多功能多胺,存在于所有生物中,是一种重要的生理活性物质。医学上,亚精胺具有延长动物寿命的功效,并抵消与年龄相关的疾病,如心血管疾病,神经退行性疾病和癌症等。农业上,植物和微生物的生理过程中起关键作用的重要内源性调节剂,可以提高果实产量与质量。
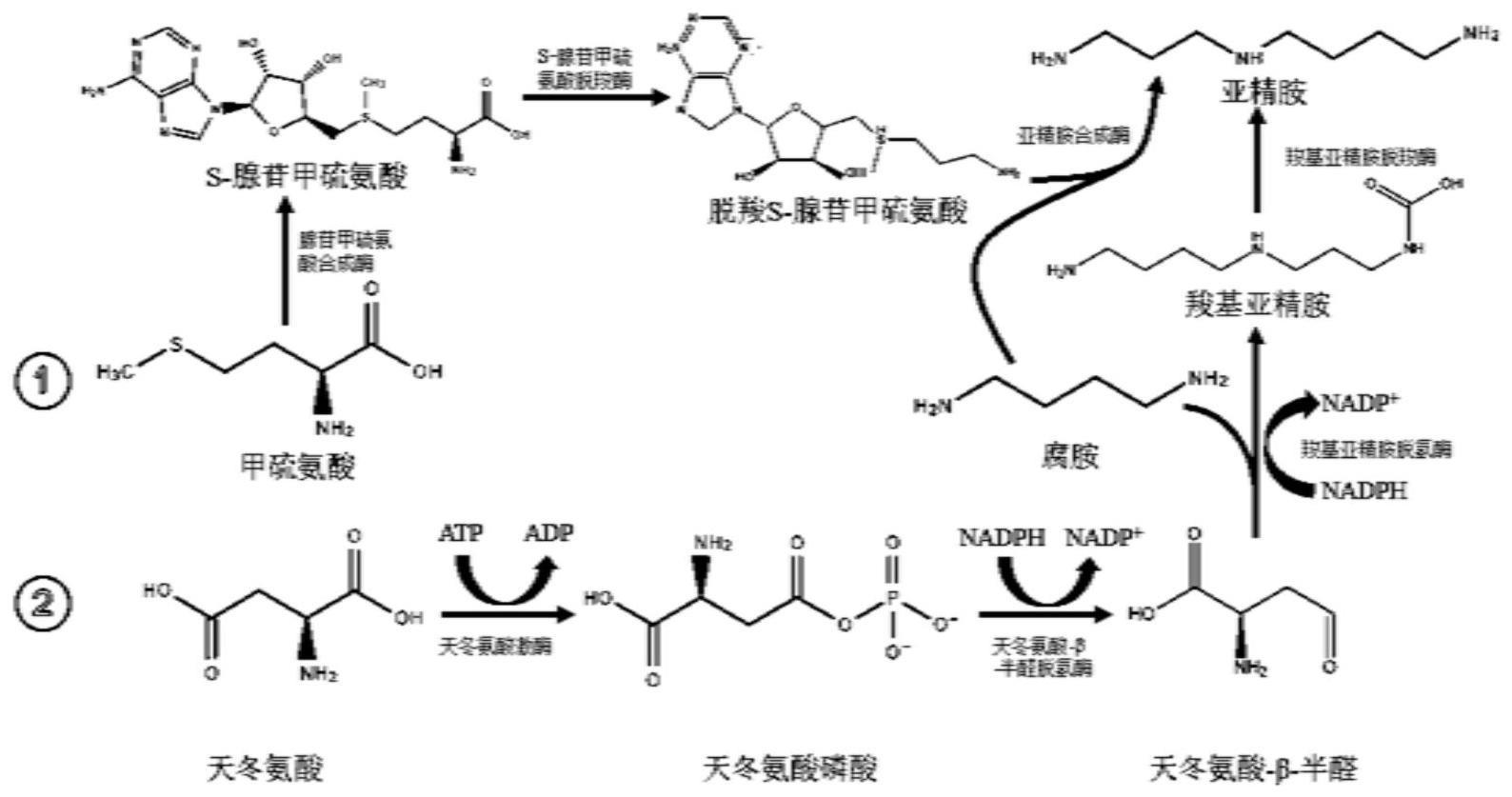
2、生物体中主要有两种途径合成亚精胺(如图1):一是脱羧s-腺苷甲硫氨酸与腐胺在亚精胺合成酶的作用下合成亚精胺,该途径涉及腺苷甲硫氨酸合成酶、s-腺苷甲硫氨酸脱羧酶、亚精胺合成酶。腺苷甲硫氨酸合成酶(sams)的作用是催化甲硫氨酸合成s-腺苷甲硫氨酸,s-腺苷甲硫氨酸脱羧酶(samdc)的作用是催化s-腺苷甲硫氨酸脱羧生成脱羧s-腺苷甲硫氨酸,亚精胺合成酶(srm)的作用是转移脱羧s-腺苷甲硫氨酸的氨基丙基到丁二胺上形成亚精胺,但途径中涉及的底物sam价格昂贵,产业化难度高;二是天冬氨酸-β-半醛和腐胺在羧基亚精胺脱氢酶(casdh)和羧基亚精胺脱羧酶(casdc)的催化下合成亚精胺,该途径涉及天冬氨酸激酶(ak)、天冬氨酸-β-半醛脱氢酶(asadh)、casdh和casdh。天冬氨酸经ak的催化作用生产天冬氨酸-β-磷酸,天冬氨酸-β-磷酸在asadh的催化作用下生成天冬氨酸-β-半醛,casdh将其与腐胺催化生成羧基亚精胺,最后在casdc的催化作用下生产亚精胺,因底物天冬氨酸的价格低廉,是一种新的替代合成途径。
3、然而,在方法二中,天冬氨酸经ak催化合成天冬氨酸磷酸反应过程的吉布斯自由能小于零(-2.2kj/mol),天冬氨酸磷酸经asadh催化形成天冬氨酸-β-半醛反应过程的吉布斯自由能小于零(-7.1kj/mol),导致二者的转化率低,这两个限速步骤仍是当前亟需解决的关键难题。
技术实现思路
1、本发明通过高通量筛选得到ak和天冬氨酸-β-半醛脱氢酶的突变株,通过过表达天冬氨酸与腐胺合成亚精胺途径中的四种关键酶,使其以天冬氨酸与腐胺为底物来实现亚精胺的高效生产。
2、技术方案
3、本发明提供了一种无细胞催化合成亚精胺的生物方法,是以天冬氨酸与腐胺为底物合成亚精胺,天冬氨酸被天冬氨酸激酶以atp为辅酶催化生成天冬氨酸-β-磷酸和adp,天冬氨酸-β-磷酸被天冬氨酸-β-半醛脱氢酶以nadph或nadp为辅酶催化生成天冬氨酸-β-半醛和nadp(nad);羧基亚精胺脱氢酶以nadph或nadp为辅酶将天冬氨酸-β-半醛和腐胺合成酶羧基亚精胺,辅酶再生为nadp(nad),最后羧基亚精胺脱羧酶将羧基亚精胺脱羧生成亚精胺;在以上过程中,由多聚磷酸盐激酶实现atp的再生、由葡萄糖脱氢酶实现nadph(nadp)的再生。
4、所述天冬氨酸激酶来源于萎缩芽孢杆菌nx-12,其氨基酸序列如seq.1所示。经突变筛选后,天冬氨酸激酶突变体酶活达到228.5u/mg,转化率达98.8%。天冬氨酸激酶突变体(命名为aspkinase-m),其氨基酸序列如seq.2所示,包含以下差别:
5、将如seq id no.1所示氨基酸序列的第34位精氨酸替换为天冬氨酸,第68位酪氨酸替换为亮氨酸,第121位丙氨酸替换为精氨酸,220位谷氨酸替换为精氨酸,第333位精氨酸替换为天冬氨酸。
6、所述的天冬氨酸-β-半醛脱氢酶来源于萎缩芽孢杆菌nx-12,其氨基酸序列如seq.3所示。经突变筛选后,天冬氨酸-β-半醛脱氢酶突变体酶活达到89.2u/mg,转化率达95.6%。天冬氨酸-β-半醛脱氢酶突变体(重新命名为ecasen-m),所述的氨基酸序列如seq.4所示,包含以下差别:
7、将如seq id no.3所示氨基酸序列的第28位丙氨酸替换为天冬氨酸,第75位苯丙氨酸替换为甘氨酸,第155位天冬酰胺替换为色氨酸,第223位赖氨酸替换为丙氨酸,第235位组氨酸替换为天冬氨酸,第356位苏氨酸替换为丙氨酸。
8、所述羧基亚精胺脱氢酶可以来自于porphyromonas catoniae atcc 51270、porphyromonas crevioricanisatcc 33277、runella slithyformis dsm 19594或porphyromonas gingivalis atcc 33277。所述羧基亚精胺脱氢酶的氨基酸序列可以是ncbi上accession no.为ls483447.1、cp002859.1、erj96771.1或eri79986.l的序列。编码所述羧基亚精胺脱氢酶的基因的核苷酸序列可以是ncbi上accession no.为cp072333region:566074...3567270、cp025931region:466768...467985或cp011996region:1326074…1327291的序列。
9、所述羧基亚精胺脱羧酶可以来自于butyrivibrio crossotus strain dsm 2876,eubacterium eligens atcc 27750或ruminococcus gauvreauii strain dsm 19829。所述羧基亚精胺脱羧酶的氨基酸序列可以是ncbi上accession no.为cp102270.1,cp001104.1,cp102290.1的序列。编码所述羧基亚精胺脱氢酶的基因的核苷酸序列可以是ncbi上accession no.为cp102270region264434…265564,cp001104region 1476074…1477300或cp102267region450849…451970的序列。
10、本发明通过spy-tag和spy-catcher分别实现天冬氨酸激酶与多聚磷酸盐激酶、天冬氨酸-β-半醛脱氢酶与葡萄糖脱氢酶、羧基亚精胺脱氢酶与葡萄糖脱氢酶的体外自组装,从而实现atp,nadph的再生,来提高催化效率。
11、将spycatcher和spytag分别与天冬氨酸激酶(aspkinase-m)与多聚磷酸盐激酶(polykinase)进行融合,通过胞内共表达,以实现双酶自组装,产生高效的辅酶再生循环系统。
12、将spycatcher和aspkinase-m两个基因分别连接到petduet-1载体上,得到重组质粒petduet-1-spycatcher-aspkinase-m并将其转入escherichia coli bl21(de3);将spytag和polykinase两个基因分别连接到pacycduet-1载体上,得到重组质粒pacycduet-1-spytag-polykinase并将其转入escherichia coli bl21(de3),将重组菌培养后接种到含有终浓度100μg/ml卡那霉素的lb培养基中培养,诱导进行蛋白的表达,得到重组蛋白液spytag-polykinase和spycatcher-aspkinase-m,分别取蛋白液500μl,加入离心管中,37℃组装30min。
13、将spycatcher和spytag分别与天冬氨酸-β-半醛脱氢酶(ecasen-m)与葡萄糖脱氢酶(gdh)进行融合,通过胞内共表达,以实现双酶自组装,产生高效的辅酶再生循环系统。将spycatcher和ecasen-m两个基因分别连接到petduet-1载体上,得到重组质粒petduet-1-spycatcher-ecasen-m并将其转入escherichia coli bl21(de3);将spytag和gdh两个基因分别连接到pacycduet-1载体上,得到重组质粒pacycduet-1-spytag-gdh并将其转入escherichia coli bl21(de3),将重组菌培养后接种到含有终浓度100μg/ml卡那霉素的lb培养基中培养,诱导进行蛋白的表达,得到重组蛋白液spytag-gdh和spycatcher-ecasen-m,分别取蛋白液500μl,加入离心管中,37℃组装30min。
14、将spycatcher和spytag分别与羧基亚精胺脱氢酶(caspermidh-3)与葡萄糖脱氢酶(gdh)进行融合,通过胞内共表达,以实现双酶自组装,产生高效的辅酶再生循环系统。将spycatcher和caspermidh-3两个基因分别连接到petduet-1载体上,得到重组质粒petduet-1-spycatcher-caspermidh-3并将其转入escherichia coli bl21(de3);将spytag和gdh两个基因分别连接到pacycduet-1载体上,得到重组质粒pacycduet-1-spytag-gdh并将其转入escherichia coli bl21(de3),将重组菌培养后接种到含有终浓度100μg/ml卡那霉素的lb培养基中培养,诱导进行蛋白的表达,得到重组蛋白液spytag-gdh和spycatcher-caspermidh-3,分别取蛋白液500μl,加入离心管中,37℃组装30min。
15、所述葡萄糖脱氢酶来自于bacillus subtilisatcc 13952。或者,所述葡萄糖脱氢酶的氨基酸序列是ncbi上accession no为wp_013351020.1序列。或者,所述葡萄糖脱氢酶的核苷酸序列是ncbi上accession no为:nz_cp009748region:386154..38693。
16、所述多聚磷酸盐激酶为来自于sinorhizobium meliloti。或者,所述多聚磷酸盐激酶,其氨基酸序列是ncbi上accession no.为wp_010968631.1的序列。或者,多聚磷酸盐激酶的核苷酸序列是ncbi上accession no.为nc_003047regio n:complement(564142..565044)的序列。
17、所述的体外无细胞催化体系包括:复合多酶体系1-1000mg/l,l-天冬氨酸0.01-100g/l,腐胺0.01-100g/l,葡萄糖0.01-100g/l,六聚偏磷酸钠0.01-150g/l,atp 0.01-10g/l,nadph 0.01-10g/l;反应条件:ph 6-9,温度25-40℃,时间2-36h。
18、本发明还提供一种用于蛋白表达菌株构建的方法:通过spy-tag和spy-catcher分别实现天冬氨酸激酶与多聚磷酸盐激酶、天冬氨酸-β-半醛脱氢酶与葡萄糖脱氢酶、羧基亚精胺脱氢酶与葡萄糖脱氢酶的体外自组装,使用petduet-1分别对天冬氨酸激酶与多聚磷酸盐激酶、天冬氨酸-β-半醛脱氢酶与葡萄糖脱氢酶、羧基亚精胺脱氢酶与葡萄糖脱氢酶进行连接,转化escherichiacolibl21(de3),进行表达纯化,形成偶联atp或nadph循环系统的复合酶系;使用pet-28a做为羧基亚精胺脱羧酶的载体,转化至escherichiacolibl21(de3)进行表达纯化。诱导表达纯化方法:将重组大肠杆菌按体积比4%的量转接到lb发酵培养基(蛋白胨10g/l,酵母粉5g/l,nacl 10g/l)中,当细胞od600达到0.6-0.8后,加入终浓度为0.5mm的iptg,在16℃诱导表达16h。诱导表达结束后,4℃、800rpm、20分钟离心收集细胞。破碎后histag标签法纯化酶,得到纯酶后测定活性。
19、有益效果
20、本发明通过筛选得到天冬氨酸激酶和天冬氨酸-β-半醛脱氢酶的突变株,并实现了无细胞催化亚精胺的合成。本发明选用的氧化还原酶类均能高效使用nad(nadp)为辅酶、反应过程无产物反馈抑制。
21、本发明提供的亚精胺合成方法,原料易于得到且价格便宜,时空产率高,能够实现亚精胺的高效合成,具有良好的工业化应用前景。
- 还没有人留言评论。精彩留言会获得点赞!