茶树在不同营养元素缺乏条件下荧光定量PCR内参基因及应用
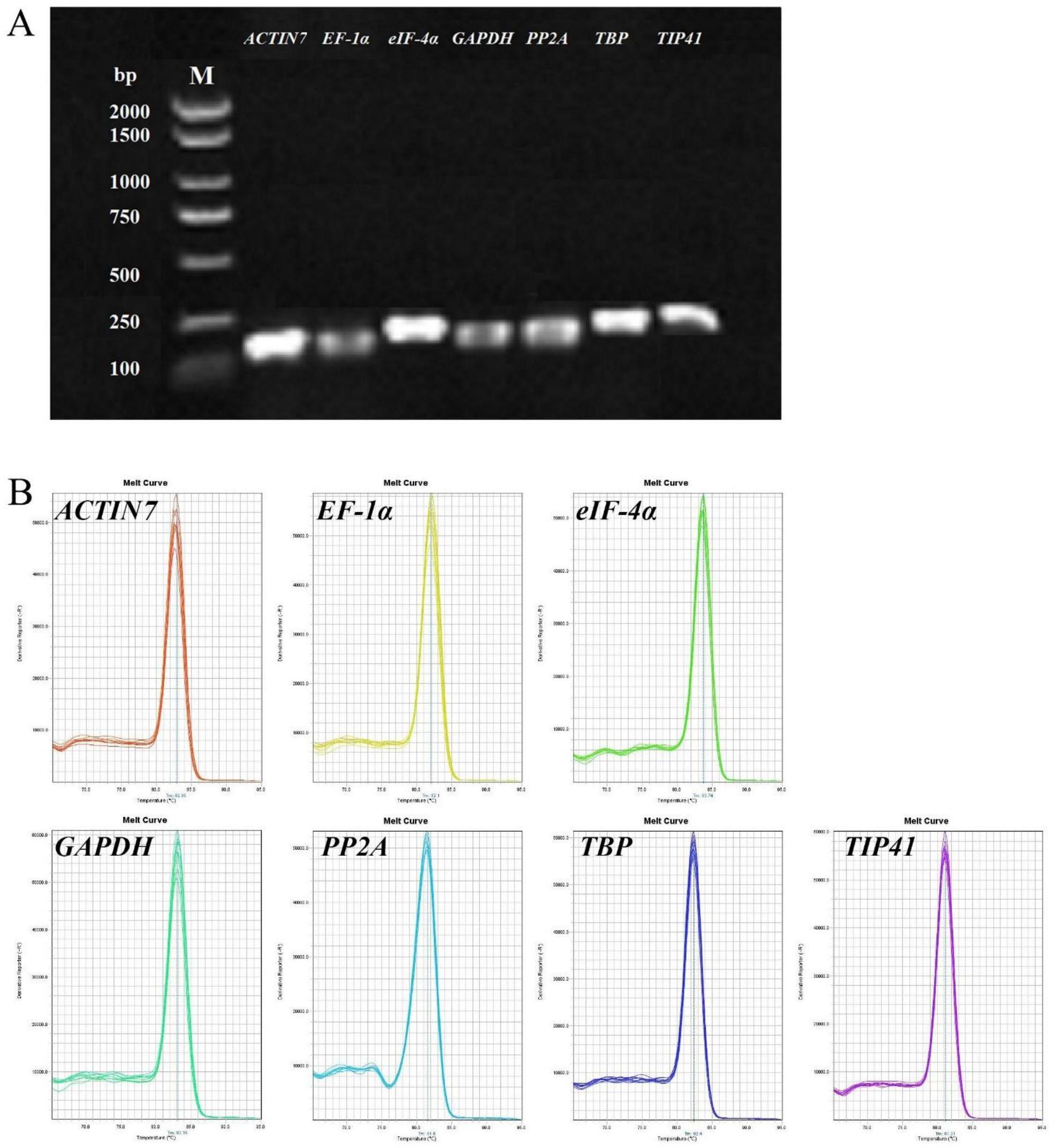
本发明属于分子生物学领域,涉及一种茶树荧光定量pcr内参基因及应用,尤其涉及一种茶树在不同营养元素缺乏条件下荧光定量pcr内参基因及应用。
背景技术:
1、茶树(camellia sinensis)是最重要的木本经济作物之一,在其生命周期中容易受到非生物和生物胁迫的影响。茶树的qrt-pcr内参基因选择已在茶树的不同器官、叶片发育阶段、各种非生物胁迫(即寒冷、干旱、重金属和氮处理)和生物胁迫(橘二叉蚜感染、茶小绿叶蝉感染和茶尺蠖感染)中有所研究。茶树qrt-pcr分析中最常用的内参基因包括csactin1,csef1,csgapdh1,cs18srrna(18s recombinant rna),cstip41,csclathrin(clathrin adaptor complex subunit),cstbp1和cssand1(sand family protein)。然而,没有关于不同营养缺乏条件下茶树中最适内参基因选择的研究。
2、实时荧光定量pcr(quantitative real-time reverse-transcriptase pcr,qrt-pcr)是一种简单、灵敏和准确的基因表达定量方法,已被广泛用于评估基因的转录水平。然而,这个“金标准”将受到不同质量的rna、pcr扩增效率和选用的内参基因的影响。因此,选择一个或多个合适的内参基因来规范目的基因表达水平至关重要。
3、在植物中,qrt-pcr的内参基因通常是稳定表达的基因,其中常用的内参基因包括:肌动蛋白基因(actin)、延伸因子基因(elongation factor-1a,ef-1α)、真核转录起始因子基因(eukaryotic translation initiation factor 4α,eif-4α)、3-磷酸-甘油醛脱氢酶基因(glyceraldehyde-3-phosphate dehydrogenase,gapdh)和tata结合蛋白基因(tata-box binding protein,tbp)。然而,每个特定内参基因的转录水平在不同的植物物种、实验条件以及植物组织和发育阶段并不总是稳定的。例如,在中国大白菜(brassicarapa l.ssp.pekinensis)中,最稳定的内参基因之一被确定为gapdh,但它在中国无头大白菜中不是一个合适的选择。当暴露于不同的非生物胁迫条件下,pp2a(proteinphosphatase 2a,pp2a)在高粱(sorghum)中最稳定;杉木(cunninghamia lanceolata)中gapdh是最稳定的内参基因;然而,gapdh是白花前胡(peucedanum praeruptorum dunn)中最不稳定的基因。此外,actin是毛白杨(populus tomentosa)经历初级和次级生长的茎段中的稳定内参基因,但它是杨树不同发育阶段中最不稳定的内参基因。此外,即使在同一物种的不同组织中,内参基因也并不总是稳定表达的。例如:actin2在猕猴桃成熟叶,柱头臂和果肉中具有高稳定性,但actin3在猕猴桃腋芽中具有高稳定性。因此,有必要在特定的实验条件下选择茶树中qrt-pcr稳定表达的内参基因。
技术实现思路
1、为了解决背景技术中存在的上述技术问题,本发明提供了一种茶树荧光定量pcr内参基因及应用,对茶树在不同营养元素缺乏的条件下茶树不同组织间的表达稳定性进行评价,以期筛选出不同缺素处理下稳定表达的内参基因,为后续茶树营养吸收和代谢相关的功能基因研究提供内参。
2、为了实现上述目的,本发明采用如下技术方案:
3、一种茶树荧光定量pcr内参基因,所述茶树荧光定量pcr内参基因是肌动蛋白基因actin7、延伸因子基因ef-1α、真核转录起始因子基因eif-4α、3-磷酸-甘油醛脱氢酶基因gapdh、蛋白磷酸酶基因pp2a、tata结合蛋白基因tbp和tip41家族蛋白基因tip41中至少一种。
4、作为优选,本发明所采用的内参基因actin7的核苷酸序列如序列表中seq id no:1所示,所述内参基因ef-1α的核苷酸序列如序列表中seq id no:2所示;所述内参基因eif-4α的核苷酸序列如序列表中seq id no:3所示;所述内参基因gapdh的核苷酸序列如序列表中seq id no:4所示;所述内参基因pp2a的核苷酸序列如序列表中seq id no:5所示;所述内参基因tbp的核苷酸序列如序列表中seq id no:6所示;所述内参基因tip41的核苷酸序列如序列表中seq id no:7所示。
5、作为优选,本发明所采用的引物是用于扩增肌动蛋白基因actin7的第一引物对、用于扩增延伸因子基因ef-1α的第二引物对、用于扩增真核转录起始因子基因eif-4α的第三引物对、用于扩增3-磷酸-甘油醛脱氢酶基因gapdh的第四引物对、用于扩增蛋白磷酸酶基因pp2a的第五引物对、用于扩增tata结合蛋白基因tbp的第六引物对或用于扩增tip41家族蛋白基因tip41的第七引物对;
6、所述第一引物对的上游引物以及下游引物分别是:cagaccgtatgagcaaggaa以及gcttagggatgcgaggatag;
7、所述第二引物对的上游引物以及下游引物分别是:caagcgtgtcatcgagagat以及ataccacgttcacgttcagc;
8、所述第三引物对的上游引物以及下游引物分别是:tgagaaggttatgcgagcac以及gcaacatgtcaaacacacga;
9、所述第四引物对的上游引物以及下游引物分别是:gactggagaggtggaagagc以及agccattccagtcaatttcc;
10、所述第五引物对的上游引物以及下游引物分别是:caacatgttcgctctgcttt以及gggaaaggaatattggcaga;
11、所述第六引物对的上游引物以及下游引物分别是:aagggatccaaagacgacag以及tgaaatccttgaatttggca;
12、所述第七引物对的上游引物以及下游引物分别是:cgaaagagcccattctcttc以及acgtgtgtccctcaatctca。
13、如前所记载的茶树荧光定量pcr内参基因作为茶树在营养元素缺乏条件下荧光定量pcr内参基因的应用。
14、如前所记载的茶树荧光定量pcr内参基因作为茶树在六种营养元素铵态氮、硝态氮、磷、钾、钙、镁中一种或多种以上缺乏条件下荧光定量pcr内参基因的应用。
15、如前所记载的茶树荧光定量pcr内参基因作为茶树新梢在六种营养元素铵态氮、硝态氮、磷、钾、钙、镁中一种或多种以上缺乏条件下荧光定量pcr内参基因的应用。
16、作为优选,本发明所采用的茶树荧光定量pcr内参基因作为茶树新梢在铵态氮元素缺乏(-nh4+-n)下荧光定量pcr内参基因的应用时,所述茶树荧光定量pcr内参基因是蛋白磷酸酶基因pp2a和/或肌动蛋白基因actin7;优选的,所述茶树荧光定量pcr内参基因是蛋白磷酸酶基因pp2a和肌动蛋白基因actin7;
17、所述茶树荧光定量pcr内参基因作为茶树新梢在硝态氮元素缺乏(-no3--n)下荧光定量pcr内参基因的应用时,所述茶树荧光定量pcr内参基因是肌动蛋白基因actin7和/或延伸因子基因ef-1α;优选的,所述茶树荧光定量pcr内参基因是肌动蛋白基因actin7和延伸因子基因ef-1α;
18、所述茶树荧光定量pcr内参基因作为茶树新梢在磷元素缺乏(-p)下荧光定量pcr内参基因的应用时,所述茶树荧光定量pcr内参基因是延伸因子基因ef-1α、蛋白磷酸酶基因pp2a和tata结合蛋白基因tbp中至少一种;优选的,所述茶树荧光定量pcr内参基因是延伸因子基因ef-1α、蛋白磷酸酶基因pp2a和tata结合蛋白基因tbp;
19、所述茶树荧光定量pcr内参基因作为茶树新梢在钾元素缺乏(-k)下荧光定量pcr内参基因的应用时,所述茶树荧光定量pcr内参基因是真核转录起始因子基因eif-4α和/或肌动蛋白基因actin7;优选的,所述茶树荧光定量pcr内参基因是真核转录起始因子基因eif-4α和肌动蛋白基因actin7;
20、所述茶树荧光定量pcr内参基因作为茶树新梢在钙元素缺乏(-ca)下荧光定量pcr内参基因的应用时,所述茶树荧光定量pcr内参基因是3-磷酸-甘油醛脱氢酶基因gapdh和/或延伸因子基因ef-1α;优选的,所述茶树荧光定量pcr内参基因是3-磷酸-甘油醛脱氢酶基因gapdh和延伸因子基因ef-1α;
21、所述茶树荧光定量pcr内参基因作为茶树新梢在镁元素缺乏(-mg)下荧光定量pcr内参基因的应用时,所述茶树荧光定量pcr内参基因是延伸因子基因ef-1α和/或真核转录起始因子基因eif-4α;优选的,所述茶树荧光定量pcr内参基因是延伸因子基因ef-1α和真核转录起始因子基因eif-4α。
22、如前所记载的茶树荧光定量pcr内参基因作为茶树根部在六种营养元素铵态氮、硝态氮、磷、钾、钙、镁中一种或多种以上缺乏条件下荧光定量pcr内参基因的应用。
23、作为优选,本发明所采用的茶树荧光定量pcr内参基因作为茶树根部在铵态氮元素缺乏(-nh4+-n)下荧光定量pcr内参基因的应用时,所述茶树荧光定量pcr内参基因是延伸因子基因ef-1α、蛋白磷酸酶基因pp2a和tata结合蛋白基因tbp中至少一种;优选的,所述茶树荧光定量pcr内参基因是延伸因子基因ef-1α、蛋白磷酸酶基因pp2a和tata结合蛋白基因tbp;
24、所述茶树荧光定量pcr内参基因作为茶树根部在硝态氮元素缺乏(-no3--n)下荧光定量pcr内参基因的应用时,所述茶树荧光定量pcr内参基因是肌动蛋白基因actin7和/或tata结合蛋白基因tbp;优选的,所述茶树荧光定量pcr内参基因是肌动蛋白基因actin7和/或tata结合蛋白基因tbp;
25、所述茶树荧光定量pcr内参基因作为茶树根部在磷元素缺乏(-p)下荧光定量pcr内参基因的应用时,所述茶树荧光定量pcr内参基因是tata结合蛋白基因tbp、延伸因子基因ef-1α和tip41家族蛋白基因tip41中至少一种;优选的,所述茶树荧光定量pcr内参基因是tata结合蛋白基因tbp、延伸因子基因ef-1α和tip41家族蛋白基因tip41;
26、所述茶树荧光定量pcr内参基因作为茶树根部在钾元素缺乏(-k)下荧光定量pcr内参基因的应用时,所述茶树荧光定量pcr内参基因是延伸因子基因ef-1α和/或tip41家族蛋白基因tip41;优选的,所述茶树荧光定量pcr内参基因是延伸因子基因ef-1α和tip41家族蛋白基因tip41;
27、所述茶树荧光定量pcr内参基因作为茶树根部在钙元素缺乏(-ca)下荧光定量pcr内参基因的应用时,所述茶树荧光定量pcr内参基因是tata结合蛋白基因tbp和/或延伸因子基因ef-1α;优选的,所述茶树荧光定量pcr内参基因是tata结合蛋白基因tbp和延伸因子基因ef-1α;
28、所述茶树荧光定量pcr内参基因作为茶树根部在镁元素缺乏(-mg)下荧光定量pcr内参基因的应用时,所述茶树荧光定量pcr内参基因是tip41家族蛋白基因tip41、肌动蛋白基因actin7和真核转录起始因子基因eif-4α中至少一种;优选的,所述茶树荧光定量pcr内参基因是tip41家族蛋白基因tip41、肌动蛋白基因actin7和真核转录起始因子基因eif-4α。
29、如前所记载的茶树优选为鄂茶10号。
30、本发明的优点是:
31、本研究分析了7个内参基因actin7,ef-1α,eif-4α,gapdh,pp2a,tbp和tip41在茶树新梢和根部在六种营养元素缺乏条件下[即:铵态氮元素缺乏(-nh4+-n)、硝态氮元素缺乏(-no3--n)、磷元素缺乏(-p)、钾元素缺乏(-k)、钙元素缺乏(-ca)、镁元素缺乏(-mg)]qrt-pcr归一化过程中的表达稳定性,借助genorm、normfinder和bestkeeper三个程序对内参基因稳定性进行评价。结果表明:(1)pp2a和actin7在-nh4+-n的新梢中稳定表达,ef-1α,pp2a和tbp在-nh4+-n根部和-p新梢中稳定表达;(2)actin7和ef-1α或actin7和tbp在-no3--n的新梢和根部中稳定表达;(3)tbp,ef-1α和tip41在-p的根部中稳定表达;(4)eif-4α和actin7在-k的新梢中稳定表达;ef-1α和tip41在-k的根部中稳定表达;(5)gapdh和ef-1α在-ca的新梢中稳定表达;tbp和ef-1α在-ca的根部中稳定表达;(6)ef-1α和eif-4α在-mg的新梢中稳定表达;tip41,actin7和eif-4α在-mg的根部中稳定表达。因此,各内参基因的表达稳定性差异很大。本研究首次系统对不同营养缺乏条件下茶树标准化内参基因的表达进行分析。研究结果将为茶树营养生物学的进一步研究提供重要依据。
- 还没有人留言评论。精彩留言会获得点赞!