一种苦味受体多个基因敲除的小鼠模型构建方法及应用与流程
本发明涉及一种苦味受体多个基因敲除的小鼠模型构建方法及应用,特别涉及一种使用crispr/cas9技术对苦味受体mtas2r105基因簇进行基因突变/基因敲除,进而利用金葡菌滴鼻感染建立苦味受体基因敲除小鼠肺部感染模型的方法,属于生物医学与动物模型。
背景技术:
1、现代生物学将人类味觉分为五种基础味觉,即咸味、酸味、苦味、甜味和鲜味,这五种味觉信息是通过味觉受体及其下游信号传导通路的协调运作而传递的。前两种味觉信息是通过味蕾细胞上的离子通道或门控离子通道进行传导的;后三种味觉信息则依赖于两大类g-蛋白偶联受体(gpcrs)家族,即t1rs和t2rs。t1rs编码甜味和鲜味的受体蛋白,t2rs则编码苦味的受体蛋白。人t2rs家族则包括~25种苦味受体,小鼠则包括~35种苦味受体(stephen d.roper&nirupa chaudhar,nat rev neurosci.2017 18(8):485–497.doi:10.1038/nrn.2017.68;chandrashekar c et al.,nature.2006nov16;444(7117):288-94.doi:10.1038/nature05401;chaudhari n&roper sd.j cell biol.2010 9;190(3):285-96.doi:10.1083/jcb.201003144)。
2、最初的研究仅仅将这些味觉受体局限在舌上味蕾中,近年的研究显示这些味觉受体(包括t1rs和t2rs)在体内存在广泛的分布,在体内很多种上皮组织均存在特异性表达(finger te,kinnamon sc.f1000 biol rep.2011;3:20.doi:10.3410/b3-20)。尽管苦味受体众多,但是转基因小鼠研究显示苦味受体的表达存在组织特异性。例如mtas2r105基因簇在气管、肠道、睾丸、舌下腺、肾脏(近端肾小管和肾小球)存在表达(feng l.,mol humreprod.2013 19(6):349-60.doi:10.1093/molehr/gat009),mtas2r131则在上呼吸道、下呼吸道(气管、肺部)、肠道、睾丸、卵巢、胸腺、脑区存在表达,而在肾脏、心脏不表达(voigta,chem senses.2012nov;37(9):897-911.doi:10.1093/chemse/bjs082),mtas2r143/mtas2r135/mtas2r126在心脏、胃肠道、呼吸道、胸腺、雌性生殖道上皮表达(lu p et al.,jcell physiol.2021 236(9):6407-6423.doi:10.1002/jcp.30315)。这就说明苦味受体基因在不同的组织器官可能存在不同的表达谱及不同的生物学功能。
3、苦味受体t2rs在呼吸系统的分布及生物学功能研究较详细。t2rs在人和小鼠呼吸道平滑肌细胞和呼吸道上皮细胞中均存在表达。苦味化合物可通过激活呼吸道平滑肌细胞t2rs来影响平滑肌的扩张反应,也可通过激活呼吸道上皮细胞t2rs而增加胞内ca2+浓度,最终导致上皮细胞动力纤毛摆动频率加强而排出有毒物质(lee rj,j clininvest.2014mar;124(3):1393-405.doi:10.1172/jci72094)。最近的系列研究揭示了苦味受体和细菌感染、固有免疫之间的关系。在人上呼吸道(包括鼻窦粘膜上皮细胞)表达苦味受体ht2r38,革兰氏阴性菌感染上呼吸道后细菌分泌酰基高丝氨酸环内酯(ahl),ahl可以有效激活表达在鼻窦纤毛细胞的苦味受体ht2r38,继而触发胞内ca2+信号,通过激活一氧化氮合成酶而促进一氧化氮(no)的产生。一氧化氮一方面可以激活蛋白硫酸化酶g(pkg),磷酸化纤毛蛋白而增加纤毛摆动频率;另一方面它渗透到鼻腔液中直接杀死细菌。甜味受体ht1r2/ht1r3可以拮抗苦味受体ht2rs的上述作用(lee rj,cohen na.cell mol lifesci.2015jan;72(2):217-36.doi:10.1007/s00018-014-1736-7)。进一步研究发现ahl诱导粘膜上皮细胞产生no的能力和苦味受体ht2r38的基因型相关。来自苦味受体ht2r38基因型avi/avi(对苯硫脲ptc不敏感)的粘膜上皮细胞,缺乏ahl诱导no产生的能力,而基因型pav/pav(对苯硫脲ptc超敏感)的粘膜上皮细胞具有强烈的ahl诱导no产生的能力,这种表型仅被发现在上呼吸道的粘膜上皮细胞,表明苦味受体参与上呼吸道感染的固有免疫反应。临床上已发现人苦味受体ht2r38基因型和上呼吸道细菌感染易感性相关(patel nn et al.,j pathog.2018oct 1;2018:9541987.doi:10.1155/2018/9541987.ecollection 2018;leerj,cohen na.am j rhinol allergy.2013jul-aug;27(4):283-6.doi:10.2500/ajra.2013.27.3911),对奎宁的敏感性和患者鼻窦炎的状态相关(workman ad.,frontimmunol.2018mar 28;9:624.doi:10.3389/fimmu.2018.00624.ecollection 2018)。在具有严重耐药性的哮喘儿童患者中观察到了苦味受体基因表达水平特异性,呼吸道苦味受体基因特异性上调为哮喘治疗药物提供一个全新的靶点(orsmark-pietras c et al.,eurrespir j.2013jul;42(1):65-78.doi:10.1183/09031936.00077712;ren y et al.,frontpharmacol.2022jun 1;13:916508.doi:10.3389/fphar.2022.916508.ecollection2022)。
4、最近研究发现苦味受体家族在人外周血白细胞,包括淋巴细胞、单核细胞巨噬细胞和中性粒细胞、肥大细胞均存在表达。苦味物质(氯喹和苯甲地那铵)可以抑制lps诱导的人外周血白细胞分泌细胞因子的能力,包括il-1b,il-2,il-4,il-5,il-10,il-17,g-csf,gm-csf,ifn-c和pge2等。苦味物质可抑制ige受体介导的人原代肥大细胞组胺和pgd2的释放。在人t、b、nk细胞上存在味觉受体(tas1rs/tas2rs),其中40-50%的外周血淋巴细胞表达htas2r43/31和htas2r38,苦味受体表达水平的敲低可以减弱外周血淋巴细胞对糖精的趋化能力。苦味受体ht2r38在中性粒细胞的表达被进一步定位在亚细胞器-脂肪滴上,被认为是革兰氏阴性菌分泌的酰基高丝氨酸环内酯(ahl)的作用受体,从而通过苦味受体把细菌感染直接和固有免疫反应相互联系。在铜绿假单胞菌感染过程中,htas2r138受体可激活过氧化物酶体增殖物激活γ受体(pparg),促进中性粒细胞中脂滴(ld)的降解,最终加速ahl-12从中性粒细胞中的清除过程,增加细胞的成活几率。苦味受体在人肺巨噬细胞存在一定的表达。脂多糖lps刺激可以增加很多苦味受体的表达水平,奎宁和苯甲地那铵可以抑制lps刺激的细胞因子的分泌水平。苦味化合物和细菌tas2r激动剂通过百日咳毒素(ptx)敏感性、磷脂酶c依赖性和肌醇三磷酸受体依赖性钙释放途径增加胞内钙离子浓度。钙信号可通过enos和nnos诱导低水平no的产生。no通过pkg途径增加胞内cgmp浓度,从而增强吞噬作用。苦味受体激活同时也降低camp浓度。camp浓度的降低也有助于增强吞噬作用。此外,巨噬细胞与气道上皮细胞的共培养模型研究结果表明上皮细胞产生的no也能显著增强巨噬细胞的吞噬作用。
5、人及小鼠肠道也表达多种苦味受体。现在的研究显示,肠道中的苦味受体有许多生物学功能。例如螺旋锥虫感染激活肠道tuft细胞的苦味受体信号通路,刺激细胞因子il-25的释放,启动ii型免疫反应。苦味受体ht2r38基因型和人群结直肠癌发病有关,在苦味受体ht2r38基因型avi/avi人群中,苦味物质不能激活t2rs而调控肠激素的释放,诱发保护性生理反应的能力减弱,从而失去有效清除肠道内毒素的能力。肠道内分泌细胞苦味受体信号可调控肠道激素(例如glp-1、pyy、生长素ghrelin等)的释放以及胆汁酸稳态平衡,影响胃肠动力、食欲和血糖。靶向苦味受体也许可以通过影响肠道激素的释放及胆汁酸代谢而改善代谢综合征的多种临床特征。
6、此外,前期研究发现小鼠肾脏中肾小球、近端肾小管上皮细胞和集合管上皮细胞均存在苦味受体(包括mtas2r105基因簇)的表达,苦味化合物苯硫脲(ptc)、中药提取物黄连碱和黄柏碱均可通过激活苦味受体信号传导通道有效诱导肾小管上皮细胞和集合管上皮细胞内钙离子浓度的升高。tas2rs在甲状腺中表达,苦味物质可通过激活t2rs来调控腺体细胞的功能及t3/t4的产生。
7、综上所述,苦味受体在动物体内分布广泛且存在重要的生物学功能。小鼠mtas2r105基因簇有几个特点:第一、苦味受体mtas2r105可识别目前苦味化合物库中35%的苦味物质(lossow k et al.,j biol chem.201615;291(29):15358-77.doi:10.1074/jbc.m116.718544);第二、mtas2r105基因簇包含mtas2r107/106/104/105/114五个基因,和人tas2r10基因同源,tas2r10基因已被发现和人的免疫细胞功能相关,tas2r10可能是宿主对多种感染性疾病诱发的免疫反应的关键因素之一;第三、mtas2r105基因簇中五个基因在味蕾、肾脏、肠道、肺脏中的表达水平不一样,可能存在不同的调控机制;第四、mtas2r104、mtas2r105、mtas2r114三个基因非常相近,但是感知的苦味物质不一样,说明存在不同的功能。目前尚缺乏相关动物模型研究其重要的生物学功能,尤其是在感染性疾病中这些受体发挥什么样的生物学功能。因此,为了方便从整体动物水平上研究苦味受体基因的功能和能够全面研究苦味受体信号的生物学功能,基于苦味受体mtas2r105基因簇和g蛋白gnat3建立苦味受体mtas2r105基因簇和gnat3的多基因敲除小鼠很有必要。
技术实现思路
1、本发明旨在为研究苦味受体基因的功能和能够全面研究苦味受体信号的生物学功能提供一种动物模型。
2、为了实现上述目的,本发明采取了以下技术方案:
3、本发明的第一方面,提供一种苦味受体多个基因敲除的小鼠模型构建方法,包括以下步骤:
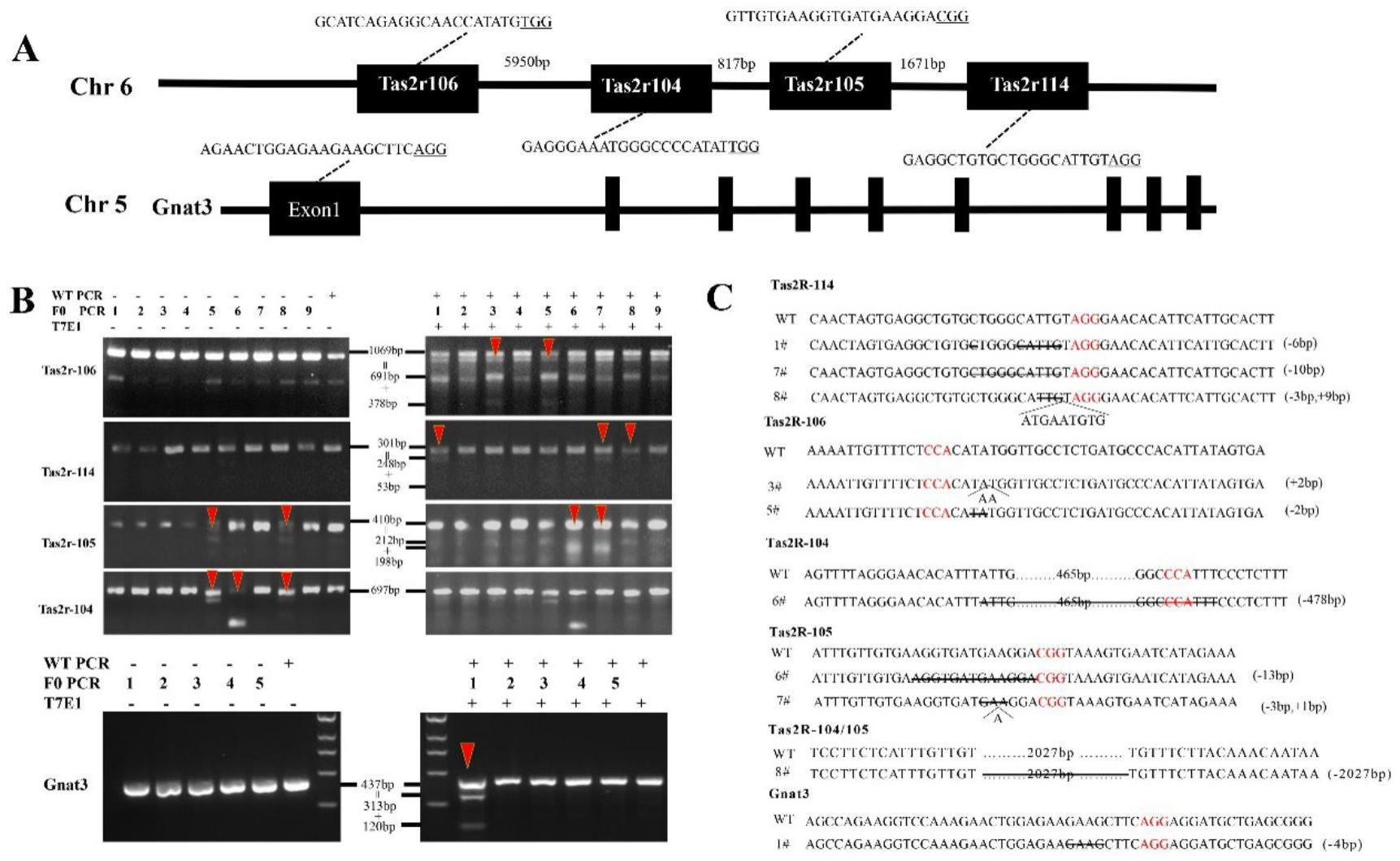
4、步骤1:分别合成针对苦味基因tas2r104、tas2r105、tas2r106、tas2r114、或基因gnat3的sgrna及cas9 mrna;
5、步骤2:通过显微共注射的方法将步骤1中合成的四种tas2r苦味基因sgrna的组合、gnat3基因sgrna分别和cas9 mrna混合后注射入小鼠原核期受精卵中;
6、步骤3:将注射后的受精卵移植入假孕母icr雌鼠的输卵管壶腹膨大部位,待小鼠出生后,通过t7e1酶切法及pcr扩增进行初步鉴定;
7、步骤4:对初步鉴定正确的小鼠产物进行t载体连接、转化、挑取单克隆进行测序确认;
8、步骤5:测序正确的基因突变小鼠为f0代子鼠,将f0子代小鼠和野生型c57bl/6小鼠连续级进杂交5代以上,通过基因型鉴定获得杂合子小鼠;
9、步骤6:将杂合子小鼠互交,获得四种tas2r苦味基因中的一个或多个tas2r基因敲除小鼠和gnat3基因敲除小鼠。
10、优选地,所述步骤6中获得了五个品系的基因敲除小鼠tas2r104-/-/tas2r105-/-、tas2r114-/-/tas2r105-/-、tas2r104-/-/tas2r105-/-/tas2r114-/-、tas2r106-/-和gnat3-/-。
11、优选地,还包括步骤7:将步骤6获得的tas2r基因敲除小鼠和gnat3基因敲除小鼠进行杂交获得相应的双基因敲除小鼠模型。
12、更优选地,将tas2r基因敲除小鼠tas2r104-/-/tas2r105-/-、tas2r114-/-/tas2r105-/-、tas2r104-/-/tas2r105-/-/tas2r114-/-或tas2r106-/-与gnat3-/-进行杂交获得相应的双基因敲除小鼠模型。
13、优选地,所述步骤6或7获得的小鼠模型进一步通过细菌感染制备细菌感染小鼠模型。
14、优选地,所述步骤1中合成sgrna及cas9 mrna所用的引物序列如seq id no:8-13所示。
15、优选地,所述步骤3中鉴定所用的pcr扩增引物序列如seq id no:12-23所示。
16、本发明的第二方面,提供本发明第一方面所述的小鼠模型构建方法构建得到的小鼠模型在制备感染性或传染性疾病模型中的应用。
17、本发明的第三方面,提供本发明第一方面所述的小鼠模型构建方法构建得到的小鼠模型在研究苦味受体基因的功能和作用机制、苦味类药物研制和研究苦味类化合物的作用机制中的应用;
18、本发明的第四方面,提供一种用于构建苦味受体多个基因敲除小鼠模型的组合物或试剂盒,所述组合物或试剂盒包括:
19、用于合成sgrna的引物组,其包括:用于扩增sgrna骨架的引物对,序列如seq idno:8-9所示;苦味基因tas2r104、tas2r105、tas2r106、tas2r114、基因gnat3对应的sgrna引物,序列如seq id no:3-7所示;以及用于扩增含t7启动子的sgrna的引物对,序列如seq idno:10-11所示;
20、和/或,用于合成cas9 mrna的引物对,序列如seq id no:12-13所示;
21、和/或,用于小鼠基因型鉴定的pcr引物组,其包括:用于扩增苦味基因tas2r104的引物对,序列如seq id no:14-15所示;用于扩增苦味基因tas2r105的引物对,序列如seqid no:16-17所示;用于扩增苦味基因tas2r106的引物对,序列如seq id no:18-19所示;用于扩增苦味基因tas2r114的引物对,序列如seq id no:20-21所示;以及用于扩增基因gnat3的引物对,序列如seq id no:22-23所示。
22、本发明提供了一种获得多种苦味受体基因敲除/突变小鼠品系的制备方法。小鼠苦味受体家族含有35个成员,其中小鼠tas2r105基因簇含有五个苦味受体基因,本发明采用crispr/cas9基因编辑的方法一次获得多个苦味受体基因敲除/突变小鼠,再通过级进杂交、横交固定的育种方法获得五个品系。结果也说明tas2r104/tas2r105/tas2r114三个基因可以共线性遗传。具体说,本发明提供了一套苦味受体信号相关研究的基因敲除/突变小鼠。包含(1)五个苦味受体基因敲除/突变小鼠;(2)苦味受体信号传导通路关键分子-g蛋白gnat3基因突变小鼠;(3)苦味受体和gnat3双基因突变小鼠。这些小鼠为整体动物水平研究苦味受体基因的功能以及苦味类药物研制和优化提供一个有用的模型。整体分析苦味受体在感染性疾病发生发展过程中的生物学功能,以及在宿主和病原微生物之间相互作用中发挥的作用成为可能。
23、本发明提供了一种测试苦味物质或苦味化合物的方法及工具。本发明包含的基因敲除/突变小鼠可以用来测试或区分苦味化合物的信号传导路径。例如具体实施途径中使用这些小鼠发现苦味受体tas2r105/tas2r114可能是葫芦素b的细胞膜受体,gnat3是其重要的信号传导通路元件。葫芦素b具有抗肿瘤、抗化学致癌、保肝、提高机体免疫力等多种生物活性。本发明使得在动物整体水平上深入研究葫芦素b的作用机制成为可能。
24、本发明提供的苦味受体基因敲除/突变小鼠有望模拟人苦味受体基因突变后的表型研究。tas2r105基因簇同人tas2r10苦味受体同源,生物信息学研究发现人tas2r10被发现和感染性疾病、传染性疾病发生发展过程中宿主的免疫反应相关。具体实施案列中,我们也发现基因敲除/突变小鼠肺部细菌载量比野生型小鼠显著增多。免疫荧光染色结果也显示基因敲除/突变小鼠肺组织ki67阳性细胞显著增加,预示肺组织的更新明显增强。该模型为以人tas2r10为药物靶点,研究治疗细菌感染肺炎的苦味类药物研制提供一套较佳的小动物模型工具。
25、另外,本发明还提供一整套金葡菌感染基因敲除/突变小鼠肺炎模型制备及分析技术方法。这种方法包括:第一金葡菌培养、菌落测定、滴鼻感染造模,使用细菌涂板测定模型肺组织cfu菌落数;第二设计特异性引物使用实时定量pcr技术检测金葡菌毒素基因的表达水平;第三全肺组织石蜡包埋、切片及he染色,然后对组织病理切片全片扫描后进行病理评分,综合评价肺组织病理变化;第四肺组织oct包埋、冰冻切片制备,再使用一套抗体进行免疫组化/免疫荧光染色分析,包括免疫细胞抗体cd68、ly6g、cd3等,细胞增殖标记物ki67,支气管上皮标志物zo-1等。
26、本发明与现有技术相比,具有如下有益效果:
27、目前研究发现苦味受体遗传突变和人多种疾病相关,包括内分泌失调、直肠癌发病、上呼吸道感染、男性生殖不育等(calvo ss,egan jm.nat rev endocrinol.2015apr;11(4):213-27.doi:10.1038/nrendo.2015.7.;martens,k.;steelant,b.;bullens,d.m.a..cells 2021,10,2889.https://doi.org/10.3390/cells 10112889;j,fendler w,pietruszewska w.int.j.mol.sci.2020,21,5156;doi:10.3390/ijms21145156);因此,对苦味受体和g蛋白gnat3基因进行特异性的基因突变/敲除,在动物整体水平影响苦味受体信号的传递,进一步理解苦味受体信号在人类疾病发生发展过程中的作用机制,本发明提供的模型有望用于筛选治疗相关疾病的苦味类药物。
- 还没有人留言评论。精彩留言会获得点赞!