一种表征红树植物抗盐能力的方法
1.本发明属于植树造林增汇和生态修复技术领域,具体涉及一种利用电生理信息表征红树植物抗盐能力的方法。
背景技术:
2.植物的生长发育及整个生命活动如耐盐性、物质代谢、水分状况和信号传导等都有质子和介电电荷的传输和分离。植物生理过程中的各种变化,如光合作用、呼吸、蒸腾、物质流、能量代谢和植物生长都与电信号直接或间接相关。叶片是植物最重要的功能器官,对光能利用、能量代谢等过程最为敏感,在植物的生长发育中起着至关举足轻重的作用。完全展开叶的叶片均是成熟的叶片,它们的细胞均具有中心液泡,在叶肉细胞里,液泡和细胞质占据了细胞内绝大部分空间,它们的吸水方式主要是渗透性吸水。无论是细胞还是细胞器,它们的外部均有细胞膜包被,磷脂双分子层是构成细胞膜的基本支架。在电镜下可分为三层,即在膜的靠内外两侧各有一条厚约2.5nm的电子致密带(亲水部分),中间夹有一条厚2.5nm的透明带(疏水部分)。因此,细胞(器)可以看成是一个同心球的电容器,只不过这种电容器因膜上的外周蛋白质和内在蛋白质变得具有兼有电感器和电阻器作用的复杂电容器。因此,植物叶片细胞的电生理特性紧密地与植物叶片的能量代谢相关。此外,细胞膜在稳定细胞内环境方面起着至关重要的作用。细胞代谢所消耗的能量几乎占细胞所消耗总能量的60%。细胞膜中的表面蛋白和结合蛋白与盐分转运能力关系最为密切,因此,细胞的盐分转运能力反映在膜蛋白的组成和含量上。
3.植物通过叶片的蒸腾作用,将约95%的水分耗散,叶细胞中剩余的5%水分可用于支持其生长。这被植物所利用的宝贵的5%胞内水,在生理过程中发挥重要作用。植物细胞在受到非生物或生物胁迫时,会产生结构和成分的变化,离子通透性也随之发生变化,电学参数也发生相应的变化。因此,受到环境影响的胞内水的水分代谢显然也反映到电生理参数的变化上。
4.红树林具有广泛的经济和生态效益,在控制沿海地区土壤盐分、减少海水入侵等方面发挥着重要作用。不同红树植物具有不同的抗盐能力和抗盐机制。虽然能够通过检测植物的生长、光合以及抗氧化能力等来反映红树林的抗盐能力,但是这些方法,或者需要较长的时间和繁杂的步骤,或者是破坏性的不具有动态和实时性,因此,开发出一种快速、在线定量检测不同环境下不同植物的抗盐能力的方法,对红树林的造林和管理具有重要意义。
5.红树植物的抗盐能力,取决于在高盐度条件下,植物调节内部盐浓度和防止离子达到有毒水平的能力。因此,植物将盐分排出细胞的盐分排出能力、将高浓度稀释低浓度的盐分稀释能力以及植物超滤能力将决定着红树植物的抗盐能力。现有技术没有很好的参数来描述不同植物在植物的盐分排出能力、盐分稀释能力和植物超滤能力上的差异,甚至常见的光合参数也不例外。
6.lcr可以测定叶片的生理电阻、生理电容、生理阻抗等电生理指标,电生理参数有
可能表征红树植物盐分排出能力、植物盐分稀释能力以及植物超滤能力以及植物总抗盐能力。
技术实现要素:
7.有鉴于此,本发明提供了一种利用电生理信息表征红树植物抗盐能力的方法,用生物物理指标来表征红树植物盐分排出能力、植物盐分稀释能力、植物超滤能力以及植物总抗盐能力,实现快速、实时、在线、无损、动态监测红树植物的抗盐性。
8.本发明是通过以下技术手段实现上述技术目的的。
9.一种表征红树植物抗盐能力的方法:
10.由植物叶片固有生理容抗ixc、固有生理感抗ixl、固有生理电阻ir以及固有生理阻抗 iz,获取植物叶片盐分排出能力c1,所述c1=a1irn+b1ixcn+c1ixln+d1izn,其中a1、b1、c1和d1均为常数,且a1+b1+c1+d1=1,irn、ixcn、ixln和izn分别由1/ir、1/ixc、1/ixl和 1/iz归一化得到;
11.根据所述植物叶片固有生理电容icp、植株叶片比有效厚度d、植物叶片胞内水分相对持水量iwhc和植物叶片盐分运输能力stc,获取植物盐分稀释能力c2,所述 c2=a2icpn+b2dn+c2iwhcn+d2stcn,其中a2、b2、c2和d2均为常数,且a2+b2+c2+d2=1,icpn、 dn、iwhcn和stcn分别由icp、d、iwhc和stc归一化得到;
12.由植物叶片胞内水分利用效率iwue、植物胞内水分相对持水时间iwht、单位面积的相对盐分通量usf和植物叶片盐分转移率str,获取植物超滤能力c3,所述c3=a3iwuen+ b30.25 iwhtn+c3usfn+d3strn,其中a3、b3、c3和d3均为常数,且a3+b3+c3+d3=1,iwuen、 iwhtn、usfn和strn分别由1/iwue、iwht、1/usf和str归一化得到;
13.再依据所述c1、c2以及c3,获取植物总抗盐能力t:其中l、m、n均为常数。
14.进一步的技术方案,所述ixc=y1+k1,其中y1、k1为植物叶片的生理容抗随夹持力变化模型中的参数;所述ixl=y2+k2,其中y2、k2为植物叶片的生理感抗随夹持力变化模型中的参数;所述ir=y3+k3,其中y3、k3为植物叶片的生理电阻随夹持力变化模型中的参数;所述iz=y4+k4,其中y4、k4为植物叶片的生理阻抗随夹持力变化模型中的参数。
15.更进一步的技术方案,所述其中f为测试频率,π是圆周率;
16.更进一步的技术方案,所述u为测试电压,h为植物叶片的生理电容随夹持力变化模型参数;所述所述stc=usf
×
str,usf为植物叶片单位面积的相对盐分通量。
17.更进一步的技术方案,所述所述iwht=icp
×
iz;所述其中:ir为植物的固有生理电阻,ixc为植物的固有生理容抗,ixl为植物的固有生理感抗;所述vt是植物叶片胞内水分转移速率。
18.进一步的技术方案,所述植物叶片盐分排出能力 c1=0.25*irn+0.25*ixcn+0.25*
ixln+0.25*izn。
19.更进一步的技术方案,所述植物盐分稀释能力c2=0.25*icn+0.25*dn+0.25*iwhcn+0.25*stcn。
20.更进一步的技术方案,所述植物超滤能力c3=0.25*iwuen+0.25*iwhtn+0.25*usfn+0.25*strn。
21.更进一步的技术方案,所述植物总抗盐能力
22.进一步的技术方案,所述生理容抗、生理感抗、生理电阻、生理阻抗随夹持力变化模型分别根据不同夹持力下待测叶片的生理电容、生理电阻和生理阻抗进行构建的。
23.更进一步的技术方案,所述不同夹持力下待测叶片的生理电容、生理电阻和生理阻抗的测定,是选取生长期的待测植株上不同叶位的待测叶片进行饱水处理后进行的。
24.本发明的有益效果为:
25.本发明不仅可以用电生理指标表征红树植物的盐分排出能力,而且可以用电生理指标表征红树植物的盐分稀释能力和超滤能力,为研究红树植物的抗盐机制提供了独特的技术手段,为因地制宜、适时适地造林的物种选择、配置和红树林管理提供了技术支撑,为红树林分类管理提供了便利,与传统的植物抗盐性的检测方法相比,本发明可以快速、实时、在线、无损、动态定量监测红树植物的综合抗盐能力,测定的结果具有可比性;本发明简便、快速,适用性广。
具体实施方式
26.下面结合具体实施例对本发明作进一步的说明,但本发明的保护范围并不限于此。
27.本发明的基本原理为:
28.植物叶片的生理电容c随夹持力f变化模型为:
[0029][0030]
其中:u为测试电压,δh为系统(由细胞组成的植物叶片系统)的内能,d为植物叶片的比有效厚度,f为夹持力;令(1)式可变形为:
[0031]
c=x0+hf(2)
[0032]
这里x0和h为模型参数,因此
[0033]
植物叶片的生理容抗、生理感抗、生理电阻、生理阻抗随夹持力变化模型均可根据现有技术推到出来,在此不再赘述。植物叶片的生理容抗、生理感抗、生理电阻、生理阻抗随夹持力变化模型可分别表示为:
[0034]
植物叶片的生理容抗随夹持力变化模型:
[0035][0036]
其中y1、k1和b1为模型的参数;
[0037]
植物叶片的生理感抗随夹持力变化模型为:
[0038]
[0039]
其中y2、k2和b2为模型的参数;
[0040]
植物叶片的生理电阻随夹持力变化模型为:
[0041][0042]
其中y3、k3和b3为模型的参数;
[0043]
植物叶片的生理阻抗随夹持力变化模型为:
[0044][0045]
其中y4、k4和b4为模型的参数。
[0046]
当夹持力f=0,则可将植物叶片固有生理容抗ixc表示为:ixc=y1+k1,将植物叶片固有生理感抗ixl表示为:ixl=y2+k2,将植物叶片固有生理电阻ir表示为:ir=y3+k3,将植物叶片固有生理阻抗iz表示为:iz=y4+k4。
[0047]
依据固有生理容抗ixc计算固有生理电容icp的方法为:其中f为测试频率,π是圆周率,本实施例中π取3.1416。
[0048]
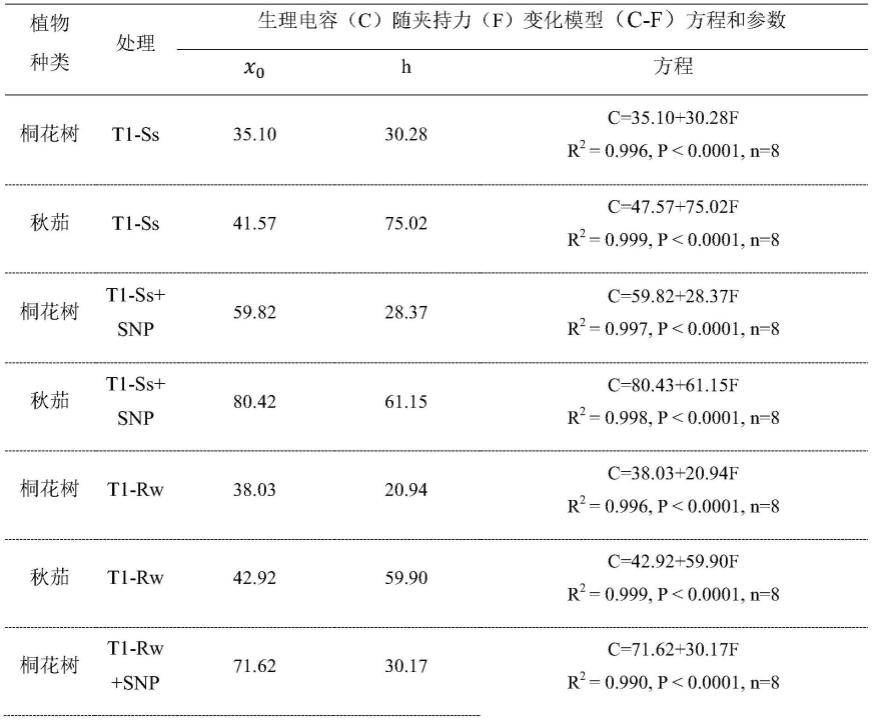
由于细胞(器)是球形结构,细胞的生长和体积的增长紧密相关,同一种植物器官尤其是叶片,细胞的体积与其内的液胞体积大小成正相关,而液胞的主要成分则是水分;因此,植物细胞的电容可借用同心球形电容器的计算公式:
[0049][0050]
其中,π是圆周率等于3.1416,c为同心球形电容器的电容,ε为电解质的介电常数,r1、 r2分别为外球和内球的半径。在细胞(器)中,r
2-r1可作为膜的厚度,r1≈r2,同一植物组织和器官的同一类细胞(器),膜的厚度一定,ε一定,因此细胞(器)的体积与细胞的电容c存在以下关系:
[0051][0052]
(8)式中,α为常数,且对于同一植物组织和器官的同一类细胞(器)来说,α的值一定,又由于细胞(器)尤其是展开叶叶片的细胞(器),体积与持水量成正比,也即细胞的持水量与成正比,因此,可以用表征植物叶片的持水量,依据固有生理电容icp计算植物叶片胞内水分相对持水量iwhc的方法则为:
[0053]
植物叶片的比有效厚度d,代表细胞的生长,胞内水分相对持水量iwhc支撑植物细胞生长,则可表征为叶片胞内水分利用效率iwue,计算方法则为:
[0054]
依据欧姆定律可知:电流iz=u/z,其中u为测定电压,iz为生理电流,z为阻抗;同时,电流又等于电容乘以电压在时间上的微分,经过积分变换,时间t则是电容量与阻抗的乘积,因此依据固有生理电容icp和植物叶片固有生理阻抗iz,获得基于电生理参数的植物胞内水分相对持水时间iwht的计算公式则为:iwht=icp
×
iz。依据叶片胞内水分相对持水量iwhc 和植物胞内水分相对持水时间iwht,则可以计算出胞内水分转移速率,其计算公式为:
[0055]
胞内水分转移速率和盐分转移率str在概念上相同,因此,植物叶片盐分转移率str 等于植物叶片胞内水分转移速率vt。
[0056]
细胞膜蛋白包括引起生物组织感抗的蛋白质-结合蛋白质(内在蛋白质)和引起生物组织容抗的蛋白质尤其是表面蛋白质(外周蛋白质),它们与盐分转运关系最为密切;因此,植物叶片单位面积的相对盐分通量usf可以用(9)表示:
[0057][0058]
其中:ir为植物的固有生理电阻,ixc为植物的固有生理容抗,ixl为植物的固有生理感抗;
[0059]
植物叶片盐分运输能力stc:stc=usf
×
str。
[0060]
盐分流出能力与生物电流有关。生物电流越大,与电子相反的阳离子如钠离子流出的就越多。因此,基于电阻的盐分流出能力ir=u/ir、基于容抗的盐分流出能力i
xc
=u/ixc、基于感抗的盐分流出能力i
xl
=u/ixl、基于阻抗的盐分流出能力i
iz
=u/iz,其中,u代表测定时施加的电压;由于植物叶片生理电容、生理电阻和生理阻抗测定时,采用的是并联方式,因此u相同;因此将1/ir、1/ixc、1/ixl和1/iz定义为植物细胞中各电器元件(电阻、容抗、感抗和阻抗)的绝对盐分流出能力,再将1/ir、1/ixc、1/ixl和1/iz的值分别归一化到 (0,1)之间,分别定义为irn、ixcn、ixln和izn;植物叶片盐分排出能力c1则为: c1=a1irn+b1ixcn+c1ixln+d1izn,其中a1、b1、c1和d1均为常数,且a1+b1+c1+d1=1,本实施例中,a1、b1、c1和d1的取值均为0.25。
[0061]
增加细胞及细胞器的体积可以稀释细胞内盐分,这种增加细胞及细胞器的体积,维持细胞内持盐能力可定义为植物盐分稀释能力,可通过icp、d、iwhc和stc来确定。将icp、 d、iwhc和stc分别归一化到(0,1)之间,分别定义为icpn、dn、iwhcn和stcn;植物盐分稀释能力c2则为:c2=a2icpn+b2dn+c2iwhcn+d2stcn,其中a2、b2、c2和d2均为常数,且a2+b2+c2+d2=1,本实施例中,a2、b2、c2和d2的取值均为0.25。
[0062]
红树植物具有超滤能力,使大多数盐分并不随蒸腾作用转移到体内。超滤能力与盐在细胞膜中的流速和流动时间有关。1/iwue代表盐溶液通量,iwht代表植物胞内水分相对持水时间,1/usf代表溶质通量,str代表盐分转移率。因此,可以用1/iwue、iwht、1/usf 和str定义红树植物的超滤能力。将1/iwue、iwht、1/usf和str的值分别归一化到(0,1) 之间,定义为iwuen、iwhtn、usfn和strn;植物超滤能力c3则为:c3=a3iwuen+b30.25 iwhtn+c3usfn+d3strn,其中a3、b3、c3和d3均为常数,且a3+b3+c3+d3=1,本实施例中, a3、b3、c3和d3的取值均为0.25。
[0063]
则植物总抗盐能力t为:其中l、m、n均为常数;本实施例中令c1、 c2和c3的权重相同(即l=m=n,则植物总抗盐能力t为:
[0064]
实施例
[0065]
在江苏大学农业工程学院温室内选取来自泉州桐青红树林科技有限公司提供的两种不同的红树林物种桐花树和秋茄幼苗;将待测植物培养在待测环境下,以处于生长期的植株为测定材料,这些植物分别进行如下的盐盐胁迫处理30天:
[0066]
在盐胁迫阶段(ss),采用不同的nacl和硝普钠(snp)处理,即t1,nacl(100mm); t2,snp(0.01mm)+nacl(100mm);t3,nacl(200mm);t4,snp(0.01mm)+nacl (200mm);t5,nacl(400mm);t6,snp(0.01mm)+nacl(400mm)。
[0067]
随后又进行复水处理(rw)30天,处理设置如下:t1,nacl(100mm);t2,snp(0.01mm) +nacl(100mm);t3,nacl(100mm);t4,snp(0.01mm)+nacl(100mm);t5,nacl (200mm);t6,snp(0.01mm)+nacl(200mm)。
[0068]
从不同的待测红树植物植株的新鲜枝条上选取第四叶位和第五叶叶片作为实验材料,放 到蒸馏水中浸泡30分钟;吸干叶片表面水,立即将待测叶片夹在测定装置平行电极板之间, 设置测定电压、频率,通过改变铁块的质量来设置不同的夹持力,并联模式测定不同夹持力 下的植物叶片生理电容、生理电阻、生理阻抗,并计算生理容抗、生理感抗;根据红树植物 叶片生理电容计算生理容抗,其中xc为植物叶片生理容抗,c为植物叶片生理电 容,f为测试频率,π是圆周率,本实施例中π取3.1416;依据红树植物叶片生理阻抗、生理 电阻和生理容抗,计算植物叶片生理感抗,其中xl为植物叶片生理感抗, xc为植物叶片生理容抗,z为植物叶片生理阻抗,r为植物叶片生理电阻;随后构建红树植 物叶片的生理电容、生理容抗、生理感抗、生理电阻、生理阻抗随夹持力变化模型,获得各 模型的各个参数;表1-5是桐花树和秋茄随机样本数据构建的模型;其他数据模型省略。
[0069]
表1桐花树和秋茄随机样本数据构建的生理电容(c)随夹持力(f)变化模型(c-f)及参数
[0070][0071]
[0072]
表2桐花树和秋茄随机样本数据构建的生理容抗(xc)随夹持力(f)变化模型(xc-f)及参数
[0073][0074]
表3桐花树和秋茄随机样本数据构建的生理感抗(xl)随夹持力(f)变化模型(xl-f)及参数
[0075]
[0076][0077]
表4桐花树和秋茄随机样本数据构建的生理电阻(r)随夹持力(f)变化模型(r-f)及参数
[0078][0079]
表5桐花树和秋茄随机样本数据构建的生理阻抗(z)随夹持力(f)变化模型(z-f)及参数
[0080]
[0081][0082]
依据生理电容随夹持力变化模型中的参数,获取红树植物叶片比有效厚度d;依据生理容抗、生理感抗、生理电阻、生理阻抗随夹持力变化模型,获取红树植物叶片的固有生理容抗 ixc、固有生理感抗ixl、固有生理电阻ir和固有生理阻抗iz;依据红树植物叶片固有生理容抗ixc计算植物叶片固有生理电容icp,如表6-7。
[0083]
表6桐花树不同处理下的红树植物叶片比有效厚度d、固有生理电阻(ir)、固有生理容抗(ixc)、固有生理感抗(ixl)、固有生理阻抗(iz)以及固有生理电容(icp)
[0084][0085]
表中,数值表示平均值
±
标准差,n=8。同列小写字母不同表示duncan检测显著差异 (p《0.05)。
[0086]
表7秋茄不同处理下的红树植物叶片比有效厚度d、固有生理电阻(ir)、固有生理容抗(ixc)、固有生理感抗(ixl)、固有生理阻抗(iz)以及固有生理电容(icp)
[0087][0088]
[0089]
表中,数值表示平均值
±
标准差,n=8。同列小写字母不同表示duncan检测显著差异 (p《0.05)。
[0090]
依据不同红树植物不同处理的植物叶片的比有效厚度d、植物叶片固有生理电容icp和植物叶片固有生理阻抗iz,分别计算植物叶片胞内水分相对持水量iwhc、叶片胞内水分利用效率iwue、植物胞内水分相对持水时间iwht,如表8-9;同时也计算出不同红树植物不同处理的叶片胞内水分转移速率vt,得出植物叶片盐分转移率str,如表10-11。依据植物叶片固有生理容抗ixc、植物叶片固有生理感抗ixl、植物叶片固有生理电阻ir以及植物叶片盐分转移率str,获取植物叶片单位面积的相对盐分通量usf和植物叶片盐分运输能力 stc;如表10-11。
[0091]
表8桐花树不同处理下的红树植物叶片胞内水分相对持水量iwhc、叶片胞内水分利用效率iwue以及植物胞内水分相对持水时间iwht
[0092][0093]
表中,数值表示平均值
±
标准差,n=8。同列小写字母不同表示duncan检测显著差异 (p《0.05)。
[0094]
表9秋茄不同处理下的红树植物叶片胞内水分相对持水量iwhc、叶片胞内水分利用效率iwue以及植物胞内水分相对持水时间iwht
[0095][0096][0097]
表中,数值表示平均值
±
标准差,n=8。同列小写字母不同表示duncan检测显著差异 (p《0.05)。
[0098]
表10桐花树不同处理下的红树植物叶片单位面积的相对盐分通量usf、盐分转移率str和叶片盐分运输能力stc
[0099][0100]
表中,数值表示平均值
±
标准差,n=8。同列小写字母不同表示duncan检测显著差异 (p《0.05)。
[0101]
表11秋茄不同处理下的红树植物叶片单位面积的相对盐分通量usf、片盐分转移率str和叶片盐分运输能力stc
[0102][0103]
表中,数值表示平均值
±
标准差,n=8。同列小写字母不同表示duncan检测显著差异 (p《0.05)。
[0104]
依据植物叶片固有生理容抗ixc、植物叶片固有生理感抗ixl、植物叶片固有生理电阻 ir以及植物叶片固有生理阻抗iz,获取植物叶片盐分排出能力c1,如表12-13;依据植物叶片固有生理电容ic、植株叶片比有效厚度d、植物叶片胞内水分相对持水量iwhc和植物叶片盐分运输能力stc,获取植物盐分稀释能力c2,如表12-13;依据植物叶片胞内水分利用效率iwue、植物胞内水分相对持水时间iwht、单位面积的相对盐分通量usf和盐分转移率str,获取植物超滤能力c3,如表12-13;依据植物叶片盐分排出能力c1、植物盐分稀释能力c2以及植物超滤能力c3获取植物总抗盐能力t,如表12-13。
[0105]
表12桐花树不同处理下的红树植物叶片盐分排出能力c1、植物盐分稀释能力c2以及植物超滤能力c3获取植物总抗盐能力t
[0106][0107]
表13秋茄不同处理下的红树植物叶片盐分排出能力c1、植物盐分稀释能力c2以及植物超滤能力c3获取植物总抗盐能力t
[0108][0109]
本发明的实施效果如下:
[0110]
从表12和表13中可以看出,桐花树和秋茄的盐分排出能力c1、植物盐分稀释能力c2、植物超滤能力c3以及总抗盐能力具有明显不同。在低盐水平下,硝普钠(snp)能显著提高桐花树和秋茄的盐分排出能力c1、植物盐分稀释能力c2、植物超滤能力c3以及总抗盐能力。在中盐水平,硝普钠(snp)对桐花树的盐分排出能力c1、植物盐分稀释能力c2、植物超滤能力c3以及总抗盐能力有促进作用,而对秋茄的盐分排出能力c1、植物盐分稀释能力c2、植物超滤能力c3以及总抗盐能力影响不大。在高盐水平下,硝普钠(snp)能显著提高桐花树的盐分排出能力c1、植物盐分稀释能力c2、植物超滤能力c3以及总抗盐能力,而对秋茄的盐分排出能力c1、植物盐分稀释能力c2、植物超滤能力c3以及总抗盐能力影响不大。
[0111]
在中盐水平下,中盐至低盐的复水降低桐花树和秋茄的盐分排出能力c1、植物盐分稀释能力c2、植物超滤能力c3以及总抗盐能力。在高盐水平下,高盐至中盐的复水对桐花树的植物盐分稀释能力c2、植物超滤能力c3和总抗盐能力有些微提高,但高盐至中盐的复水对秋茄的盐分排出能力c1有极大的提高,对秋茄的植物盐分稀释能力c2、植物超滤能力c3和总抗盐能力也具有较大的促进作用。
[0112]
在硝普钠(snp)存在下,中盐至低盐的复水降低了桐花树的盐分排出能力c1、植物超滤能力c3以及总抗盐能力,但中盐至低盐的复水对秋茄的植物超滤能力c3有显著的促进作用,对秋茄的盐分排出能力c1和植物盐分稀释能力c2有负面作用,对秋茄的总抗盐能力没有显著影响。
[0113]
在硝普钠(snp)存在下,高盐至中盐的复水只降低了桐花树的盐分排出能力c1,对植物盐分稀释能力c2以及总抗盐能力影响不大,但高盐至中盐的复水降低了秋茄的植物超滤能力c3以及总抗盐能力,对秋茄的盐分排出能力c1和植物盐分稀释能力c2影响不大。
[0114]
总之,硝普钠(snp)在低盐水平下大大促进红树植物的抗盐能力;在中盐及高盐水平下,硝普钠(snp)仅对桐花树的抗盐能力的改善有帮助,而不能改善秋茄的抗盐能力,甚至还逆转高盐至中盐的复水效果。此外,高盐至中盐的复水对秋茄的抗盐能力有较大的提高,但对桐花树的抗盐能力影响较小。
[0115]
上述结果表明两种红树林物种在不同盐水平下的耐盐机制和适应策略是不同的。在不同盐水平下硝普钠对桐花树的抗盐能力均有促进作用,这与硝普钠能增强植物的抗氧化能力及桐花树的生物泌盐特性有关;高盐至中盐的复水情况下,硝普钠对秋茄的抗盐能力具有抑制作用,这是较强的抗氧化能力抑制盐分的物理排出对冲高盐至中盐的复水提高的盐分物理排出的结果。这些与实际情况相符合,同时,我们可以看出,桐花树具有较强的生物泌盐特性,而秋茄也有相对应的较强的物理泌盐特性,生物泌盐和物理泌盐共同发挥
作用提高红树植物对盐分的适应性。
[0116]
所述实施例为本发明的优选的实施方式,但本发明并不限于上述实施方式,在不背离本发明的实质内容的情况下,本领域技术人员能够做出的任何显而易见的改进、替换或变型均属于本发明的保护范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1