一种类风湿性关节炎标志物及其下游通路抑制剂的应用
1.本发明涉及生物医药领域,更具体地,涉及一种类风湿性关节炎标志物及其下游通路抑制剂的应用。
背景技术:
2.持续性滑膜炎是类风湿性关节炎(ra)的临床特征之一,它是影响全球约1%人口的最常见的全身性自身免疫性疾病。尽管在过去十年中,靶向治疗策略已被广泛应用于ra的治疗中,但由于很大一部分患者对当前的治疗方法耐药或难治,临床上仍存在未满足的需求。因此,现有技术亟需更好地了解其发病机制,以获取相关的标志物或靶向位置,促进新型靶向治疗药物、治疗方案的形成。
技术实现要素:
3.本发明旨在克服上述现有技术的至少一种不足,提供一种类风湿性关节炎标志物及其下游通路抑制剂的应用,标志物便于类风湿性关节炎的预测、诊断,其下游通路抑制剂能为类风湿性关节炎提供新的有效治疗策略。
4.本发明的一个目的在于提供一种类风湿性关节炎标志物,包括年龄相关b细胞和/或tph细胞。在本发明的一个以上实施例中,通过动物实验以及ra患者和spa患者、oa患者、健康对照的比较,发现年龄相关b细胞在ra患者中数量、百分比呈现高水平,即通过年龄相关b细胞或其增长趋势,便于实现类风湿性关节炎(ra)患者的预测、诊断。在本发明的一个以上实施例中,还发现了年龄相关b细胞与t
ph
细胞正相关,即通过t
ph
细胞,也有利于类风湿性关节炎的预测和诊断。
5.本发明的又一个目的在于提供检测年龄相关b细胞和/或t
ph
细胞(外周t辅助细胞)的试剂在制备预测、诊断类风湿性关节炎的产品中的应用。相应的,检测年龄相关b细胞、t
ph
细胞或其组合的试剂,能用于制备预测、诊断类风湿性关节炎的产品中。所述产品包括预测、诊断类风湿性关节炎的试剂盒。
6.本发明的又一个目的在于提供年龄相关b细胞下游通路抑制剂在制备预防、治疗类风湿性关节炎的产品中的应用。在本发明的一个以上实施例中,发现abcs诱导fls激活,且在抑制abcs通过tnf-α介导的erk1/2和jak-stat1通路时,可抑制相应的fls激活。
7.进一步地,年龄相关b细胞下游通路抑制剂包括tnf-α抑制剂、erk1/2抑制剂和/或jak-stat1通路抑制剂。
8.进一步地,tnf-α抑制剂包括抗tnf-α;和/或,erk1/2抑制剂包括sch772984;和/或,jak-stat1通路抑制剂包括巴瑞替尼;和/或,stat1激活抑制剂包括氟达拉滨。
9.本发明的再一目的在于提供一种治疗类风湿性关节炎的药物,包括年龄相关b细胞下游通路抑制剂。
10.进一步地,年龄相关b细胞下游通路抑制剂包括tnf-α抑制剂、erk1/2抑制剂、jak-stat1通路抑制剂。
11.进一步地,tnf-α抑制剂包括抗tnf-α;和/或,erk1/2抑制剂包括sch772984;和/或,jak-stat1通路抑制剂包括巴瑞替尼;和/或,stat1激活抑制剂包括氟达拉滨。
12.进一步地,还包括药学上可接受的辅料。
13.本发明的再一目的在于提供一种年龄相关b细胞下游通路抑制剂在制备抑制fls激活的产品中的应用。
14.与现有技术相比,本发明的有益效果为:提供了一种类风湿性关节炎的标志物,通过对标志物的检测并比较其与健康样本,便于预测和诊断类风湿性关节炎的存在与否。同时,通过对具体年龄相关b细胞机制的研究,发现通过阻断年龄相关b细胞介导的下游通路,能够降低fls的活化,有利于应用类风湿性关节炎中实现新的治疗策略,从而有效治疗以持续性滑膜炎为主要临床特征的类风湿性关节炎,满足现有技术中临床上的需求,促进开发新型靶向治疗方案。
附图说明
15.图1显示abcs在cia小鼠中扩增;(a-b)第42天,wt和cia小鼠脾脏、关节和血液中cd11c+t-bet+abcs的代表性facs图(a)和量化(b);所有数据代表2-3个独立实验;数据表示为平均值
±
sem;统计显着性由学生t检验确定,*p《0.05,**p《0.01。
16.图2显示ra患者abcs的表型特征;(a)ra患者血液中abc的代表性facs图;(b)cd11c-b细胞(蓝线)和abcs(红线)上t-bet、mhc-ii、cd86、cd80、cd138、bcma、il-13rα1、il-4rα和il-13rα2表达的代表性直方图;(c)abcs和cd11c-b细胞中细胞因子的mrna表达;数据表示为平均值
±
sem;统计显着性由学生t检验确定,*p《0.05,***p《0.001,****p《0.0001。ns,没有意义。
17.图3显示ra患者abcs增加,且与疾病活动度相关。(a)ra(n=67)、健康对照(hc,n=29)、spa(n=23)和oa患者(n=25)中abcs的比例。(b)按das28分组的ra患者中abcs的比例。(c)abc与tjc、sjc和das28比例之间的相关性。(d-e)具有阳性或阴性抗ccp(d)和rf(e)的ra中abc的比例。(f)ra患者血液和滑膜液中abc的代表性facs图和定量。(g)通过免疫荧光对ra患者滑膜组织中的cd20和cd11c进行共染色;绿色表示cd20,红色表示cd11c,黄色表示双阳性细胞。(h)ra、hc、spa和oa患者中tph的比例。(i)cd4
+
t细胞中tph比例与cd19
+
b细胞中abcs比例的相关性。所有数据代表2-3次独立实验。数据表示为平均值
±
sem。统计显着性由单因素方差分析确定,然后是多组比较的bonferroni多重比较检验或学生t检验,*p《0.05,**p《0.01,***p《0.001,****p《0.0001,ns,没有意义。
18.图4ra中abcs的转录组图谱。(a)与幼稚和记忆b细胞相比,显示abcs上调和下调基因的维恩图。(b)条形图显示与幼稚b细胞相比,abcs上调基因中最丰富的通路。(c)显示与趋化性、迁移、粘附和中性粒细胞活化相关基因的热图。(d)与cd11c-b细胞相比,abc上ccr2和cxcr3的mrna表达。数据表示为平均值
±
sem。统计显着性由学生t检验确定,*p《0.05,****p《0.0001。
19.图5abcs来源的tnf-α可诱导fls的激活。(a)abcscm处理的fls或单独fls上icam-1和vcam-1表达的代表性直方图;(b)fls、fls+cd11c-b和fls+abc细胞上清液中的il-6、mmp-1、mmp-3和mmp-13水平。(c)fls、fls+abcs和fls+abcscm上清液中的il-6、mmp-1、mmp-3和mmp-13水平。(d)细胞计数珠阵列检测到的abcscm和幼稚b细胞培养基中ifn-α、ifn-γ、
tnf-α、il-17a和il-1β水平。(e)fls、fls+abcscm、fls+abcscm+阿达木单抗和fls+abcscm+抗il-1β抗体的上清液中il-6、mmp-1、mmp-3和mmp-13水平。数据表示为平均值
±
sem。统计显着性由单因素方差分析确定,然后是bonferroni的多组比较多重比较检验,**p《0.01,***p《0.001,****p《0.0001。
20.图6abcs促进fls中干扰素刺激基因的上调。(a)火山图显示,与单独的fls相比,共培养的fls中具有显着上调的差异表达基因(fold change》2;fdr《0.05)或下调(fold change《0.5;fdr《0.05)。(b)条形图显示,与单独的fls相比,共培养的fls的上调基因中的最高富集通路。(c)基因集富集分析(gsea)显示,与单独的fls相比,共培养的fls中富集了干扰素α和γ反应通路。(d)热图显示与ⅰ型和ⅱ型干扰素信号相关的基因。
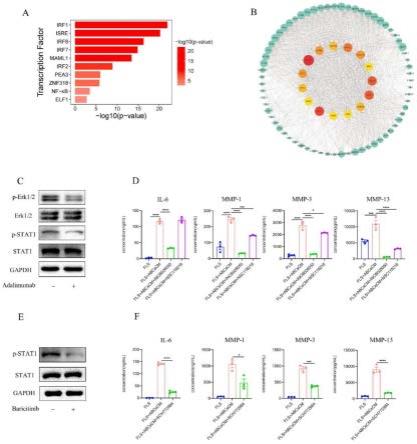
21.图7显示erk1/2和jak-stat1通路的阻断阻止了abcs诱导的fls活化。(a)基于msigdb基因集中已知靶基因的转录因子预测。(b)与单独的fls相比,abcscm处理fls中上调基因的蛋白质-蛋白质相互作用分析。(c)在阿达木单抗存在或不存在的情况下,用abcscm处理的fls中stat1、erk1/2、stat1磷酸化和erk1/2磷酸化的蛋白质印迹分析。(d)单独的fls、abcscm处理的fls、abcscm处理并添加10μm baricitinib(incb028050)的fls和abcscm处理并添加10μm fludarabine(nsc118218)的fls上清液中,il-6、mmp-1、mmp-3和mmp-13的水平。(e)在存在或不存在10μm baricitinib的情况下,用abcscm处理的fls中,stat1和stat1磷酸化的蛋白质印迹分析。(f)单独的fls、abcscm处理的fls和abcscm处理并添加了10μm erk1/2抑制剂(sch772984)的fls的上清液中,il-6、mmp-1、mmp-3和mmp-13的水平。数据表示为平均值
±
sem。统计显着性由单因素方差分析确定,然后是bonferroni的多组比较多重比较检验,*p《0.05,***p《0.001,****p《0.0001。
22.图8显示il-21和tlr7激动剂促进小鼠abcs分化。(a)小鼠脾脏中abcs的门控策略;(b
–
c)通过抗igm、抗cd40、il-21和tlr激动剂r837刺激wt和cia小鼠脾脏cd23+b细胞,获得cd11c+t-bet+abcs的代表性facs图(b)和定量(c)。象限中的数字表示每个象限中细胞的百分比。(d-f)显示cd11c和t-bet(d)表达的代表性直方图,以及显示cd11c(e)和t-bet(f)的平均荧光强度(mfi)的统计图。所有数据代表2-3次独立实验。数据表示为平均值
±
sem。统计显着性由学生t检验确定,*p《0.05、**p《0.01、***p《0.001和****p《0.0001。
23.图9显示对照和ra中abc的表型特征。(a)ra患者循环中abcs的门控策略。(b)hc(黑线)和ra-abc(红线)中t-bet、mhc-ii、cd86、cd80、cd138、bcma、il-13rα1、il-4rα和il-13rα2表达的代表性直方图。所有数据代表2-3个独立实验。
24.图10显示abcs与疾病活动和临床参数的相关性分析。(a)ra、hc、spa和oa患者的abcs数量。(b)按das28分组的ra患者的abcs数量。(c)abcs数量与tjc、sjc和das28之间的相关性。(d-e)抗ccp(d)和rf(e)阳性或阴性的ra患者中abcs数。(f)abcs数量与ra患者年龄之间的相关性。(g-j)男性和女性患者之间的abcs数量(g)、病程《2年或≥2年(h)、是否使用皮质类固醇(i)以及是否使用生物制剂(j);(k)接受mtx、lef、hcq和ssz治疗的患者中的abc数量。(l)abc和tph细胞数量之间的相关性。所有数据代表2-3次独立实验。数据表示为平均值
±
sem。统计显着性由单因素方差分析确定,然后是多组比较的bonferroni多重比较检验或学生t检验。*p《0.05,**p《0.01,****p《0.0001,n。s.,不重要。
25.图11显示ra中abcs的转录组谱。(a)火山图显示与具有幼稚或记忆b细胞的abcs相比,差异表达基因显着上调(倍数变化》2;fdr《0.05)或下调(倍数变化《0.5;fdr《0.05);(b)
基因本体生物过程分析,突出与记忆b细胞相关的abcs中上调基因相关的生物学网络;(c)热图显示与t细胞活化、趋化性、迁移和粘附途径相关的基因。
26.图12显示fls的识别;(a)fls的形态鉴定。(b)fls中cd55表达。
27.图13显示abcscm处理的fls中stat1和erk1/2的激活。(a)单独的fls和用abcscm处理15分钟、30分钟和1小时的fls中,stat1和stat1磷酸化的蛋白质印迹分析;(b)单独fls和用abcscm处理1h、2h和3h的fls中,erk1/2和erk1/2磷酸化的蛋白质印迹分析。
具体实施方式
28.应该指出,以下详细说明都是例示性的,旨在对本技术提供进一步的说明。除非另有指明,本文使用的所有技术和科学术语具有与本技术所属技术领域的普通技术人员通常理解的相同含义。
29.需要注意的是,这里所使用的术语仅是为了描述具体实施方式,而非意图限制根据本技术的示例性实施方式。如在这里所使用的,除非上下文另外明确指出,否则单数形式也意图包括复数形式,此外,还应当理解的是,当在本说明书中使用术语“包含”和/或“包括”时,其指明存在特征、步骤、操作、器件、组件和/或它们的组合。
30.现结合具体实例对本发明作进一步的说明,以下实施例仅是为了解释本发明,但不构成对本发明的限制。在以下实施例中所用到的试验样本及试验过程包括以下内容(如果实施例中未注明的实验具体条件,通常按照常规条件,或按照试剂公司所推荐的条件;下述实施例中所用的试剂、耗材等,如无特殊说明,均可从商业途径得到)。实施例1
31.一、方法材料
32.1、患者和健康人
33.外周血和血清样本来自67名ra(类风湿关节炎)患者、25名oa(骨关节炎)患者和23名spa(脊柱关节炎)患者。使用超声引导抽吸方式从ra患者受影响膝关节中获取滑液。并招募了29名性别和年龄相匹配的健康人。ra患者在诊断时符合2010年acr/eular分类标准并提供了书面知情同意书。
34.2、原代成纤维细胞样滑膜细胞的分离
35.膝关节滑膜组织样本取自医院接受全膝关节置换术的ra患者。在rpmi-1640培养基中,用0.5mg/ml胶原酶
ⅷ
(sigma)和dnaseⅰ(roche)消化滑膜组织,在培养箱摇床中孵育1小时,分离原发性fls(成纤维细胞样滑膜细胞)。在传代之前,将解离的细胞置于补充有10%胎牛血清(fbs),1%青霉素-链霉素和25mm hepes的rpmi-1640中生长至汇合。通过流式细胞术评估cd55的表达,并使用3至5次之间的传代进行共培养实验。
36.3、胶原诱导的关节炎模型
37.c57b6/l小鼠购自杭州紫园实验动物技术有限公司,并在适当条件下饲养在无特定病原体的动物室内。实验中使用8-12周龄的雄性小鼠。cia的建立采用现有技术常见方法。在第42天处死小鼠并进行分析。所有实验均按照动物设施的规章制度进行,并经动物伦理委员会批准。
38.4、流式细胞仪
39.按照常规方法处理小鼠脾脏和后爪以获得单细胞悬液。通过心脏穿刺将血液收集
到含有乙二胺四乙酸(edta)的管中。裂解红细胞并用荧光染料偶联的抗体对白细胞进行染色。使用lymphoprep通过密度梯度离心,分离人外周血单个核细胞(pbmc)。收获滑液单核细胞(sfmc)。对表面分子进行染色,并在4℃避光孵育30分钟。对于细胞内染色,细胞在表面染色后用foxp3染色缓冲液组(thermo fisher scientific)固定。用于流式细胞术的所有荧光染料偶联抗体列于补充表2中。数据是在lsrfortessa流式细胞仪(bd biosciences)上采集的,并使用flowjo软件(treestar)进行分析。
40.5、细胞分选
41.用抗cd19、抗cd27和抗cd11c抗体对人pbmc单细胞悬液进行染色。b细胞在facsariaⅲ(bd biosciences)上分为cd19
+
cd11c-cd27-(naive)、cd19
+
cd11c-cd27
+
(memory)和cd19
+
cd27-cd11c
+
(abcs)。分选后纯度大于90%。
42.6、abc体外分化
43.按照制造商的说明,用生物素化的抗cd23(bd bioscience)和链霉亲和素微珠(miltenyi biotec)富集小鼠脾脏cd23
+
b细胞,然后,在存在或不存在50ng/ml il-21(peprotech)和1μg/ml tlr7激动剂咪喹莫特(invitrogen)的情况下,通过5μg/ml f(ab')2anti-mouse igm(jackson immunoresearch laboratories)、5μg/ml anti-mouse cd40(bio x cell)在rmpi-1640培养基中进行刺激,
44.7、定量实时pcr
45.使用rneasy plus mini kit(qiagen)从细胞中分离总rna。使用fastking gdna dispelling rt supermix(天根,中国)合成cdna。在quantstudio q5实时pcr系统上使用quantinova sybr green pcr kit(qiagen,hilden,germany)进行定量实时pcr。使用2
δδct
方法计算基因表达,并将其标准化为gapdh表达。其中涉及的对应基因引物包括:gapdh:forward:5
’‑
ggagcgagatccctccaaaat-3’;reverse:5
’‑
ggctgttgtcatacttctcatgg-3’。t-bet:forward:5
’‑
gtccaacaatgtgacccagat-3’;reverse:5
’‑
acctcaacgatatgcagccg-3’。il-4:forward:5
’‑
agatcatcggcattttgaacg-3’;reverse:5
’‑
tttggcacatccatctccg-3’。il-17a:forward:5
’‑
tcccacgaaatccaggatgc-3’;reverse:5
’‑
ggatgttcaggttgaccatcac-3’。il-21:forward:5
’‑
tagagacaaactgtgagtggtca-3’;reverse:5
’‑
gggcatgttagtctgtgtttctg-3’。tnf-α:forward:5
’‑
gaggccaagccctggtatg-3’;reverse:5
’‑
cgggccgattgatctcagc-3’。ifn-γ:forward:5
’‑
tcggtaactgacttgaatgtcca-3’;reverse:5
’‑
ccatctttggaaggttcaggttg-3’。ccr2:forward:5
’‑
ccacatctcgttctcggtttatc-3’;reverse:5
’‑
cagggagcaccgtaatcataatc-3’。cxcr3:forward:5
’‑
ccacctagctgtagcagacac-3’;reverse:5
’‑
agggctcctgcgtagaagtt-3’。
46.8、滑膜组织的免疫荧光染色
47.将冷冻的滑膜组织切片置于烘箱中,37℃下孵育10-20分钟,并用多聚甲醛固定30分钟。洗涤3次后,将载玻片浸入edta抗原修复缓冲液(ph 8.0)中,并置于微波炉中20分钟。将载玻片在室温下冷却10分钟并用蒸馏水洗涤3次。使用3%bsa封闭非特异性结合30分钟。从载玻片上去除封闭溶液,并将一抗添加到含有0.1%tween 20的pbs中。载玻片与兔抗人cd20(克隆ep459y,abcam)和兔抗人cd11c(gb11059,servicebio)在湿盒中于4℃孵育过夜,然后与hrp标记的山羊抗兔二抗孵育(gb23303,servicebio)。采用酪胺信号扩增(tsa)技术进行多色染色。洗涤后,用fitc-tsa(g1231,servicebio)和cy3-tsa(g1223,servicebio)溶
signaling technology,usa)和第二抗兔igg-hrp(cell signaling technology,usa)的抗体探测膜。gapdh(d16h11,cell signaling technology,usa)用作凝胶上样对照。印迹使用聚偏二氟乙烯(pvdf)膜显影,并在bio-rad图像系统上成像。
56.13、统计分析
57.所有数据均用mean
±
sd表示。两组比较采用无配对双尾student’s t检验,单因素方差分析(one-way anova),多组比较采用graphpad prism软件进行bonferroni多重比较检验。p值《0.05为有统计学意义。
58.二、cia小鼠中abc的扩增
59.现有技术报道,abc在自身免疫易感小鼠(例如自发性狼疮小鼠)中扩增,表明abc在疾病发展中有致病作用。为了评估abc在关节炎小鼠中的分布,在第42天,通过流式细胞术评估了cia小鼠血液、脾脏和关节中的abc。与在先研究一致,小鼠abc被定义为b220
+
cd11c
+
t-bet
+
(图8a)。结果显示,与wt对照组相比,cia小鼠脾脏和血液中abc的比例和数量增加(图1a,b)。cia小鼠关节中abc的数量也高于wt小鼠,尽管差异无统计学意义(p=0.09;图1a,b)。即结果显示,abc在关节炎小鼠中扩增,不仅在脾脏和外周血中扩增,在发炎的关节中也有扩增。
60.三、il-21和tlr7促进cia小鼠的abcs分化
61.据现有技术报道,abcs可以通过刺激il-21和tlrs而产生。因此,本发明人研究了这些信号是否可以驱动关节炎小鼠的abc分化。与预期的一样,添加il-21,将导致cia来源的脾脏cd23
+
b细胞中cd11c
+
t-bet
+
abcs比例显著高于健康小鼠(图8b,c)。然而,单独添加tlr7激动剂咪喹莫特并不影响它们的分化(数据未显示)。值得注意的是,il-21、tlr7组合刺激引起的abcs产生是高于单独的il-21刺激的(图8b,c)。一致地,经il-21刺激后,与野生型对照组相比,cia小鼠cd23
+
b细胞中cd11c和t-bet的表达(图8d-f)(mfi评估)升高。总之,cia小鼠的b细胞在体外暴露于il-21和tlr7信号后,表现出更强的产生abcs的能力。
62.四、ra(类风湿性关节炎)患者中存在促炎表型的abcs
63.接下来,为了证实ra中abcs(年龄相关b细胞)的存在,并探讨其表型特征,本发明人评估了ra患者外周血中的abcs。如现有技术所揭示的,人类abc被定义为cd19
+
cd27-igd-cd21-cd11c
+
(如图2a和图9a)。与cd11c-b细胞相比,来自ra患者的abc表现显著更高的转录因子t-bet表达水平,且在小鼠和人类中都有所描述(图2b)。类似的,在ra abcs中观察到cd86和mhc-ii的更高表达,但未观察到cd80的更高表达(图2b),这与现有技术在狼疮小鼠中的发现一致。对于浆细胞标志物,abcs中bcma和cd138表达没有显着上调(图2b)。由于已报道il-4信号传导在体外下调abc形成,本发明人研究了abcs中的il-4和il-13受体表达。结果发现,与cd11c-b细胞相比,abcs中的il-13rα1表达上调;而il-4rα和il-13rα2的表达在两个群体之间是相当的(图2b),这与之前在小鼠中的发现一致。
64.为了了解来自健康对照的abc与ra中存在的abc相比如何,检查了健康个体中的表型标志物表达。如图9b所示,所有标志物的表达模式是可比较的(通过流式细胞术检查),表明这些细胞在健康个体中的相似性。为了进一步评估abc与其生物学行为相关的表型特征,对ra患者的abcs和cd11c-b细胞进行了分类,并通过qpcr检查了几种细胞因子的表达水平。结果显示,促炎细胞因子,如il-21、il-17和tnfα,不包括ifn-γ,在abc中升高(图2c),表明这些细胞存在促炎表型。而且,abc中的il-4表达也高于cd11c-b细胞il-4表达(图2c)。
65.五、ra患者abcs增加,且与疾病活动度相关
66.对ra患者abcs的进一步检查表明,ra患者abcs的数量和百分比高于spa、oa和健康对照组(图3a和图10a)。值得注意的是,活动性ra患者的abc频率较高,其das28高于3.2(图3b和图10b)。一致地,abcs与如压痛关节计数(tjc)、肿胀关节计数(sjc)和das28一类临床参数呈正相关,表明其与疾病的发病机制相关(图3c和图10c)。本发明人进一步根据血清中是否存在类风湿因子(rf)和抗环瓜氨酸肽(ccp)以对ra患者进行分层。结果显示,抗ccp阳性患者的abcs高于抗ccp阴性患者,而在rf阳性和阴性患者之间,观察到的abc数量相当(图3d、e和图10d、e)。与之前显示abc与年龄呈正相关的一项研究相反,本发明人没有发现abc的数量与年龄、性别或疾病持续时间之间存在任何相关性(图10f-h)。接下来,本发明人检测在ra患者的炎症部位是否也能发现abc。值得注意的是,与同一患者的血液相比,滑膜液中abc的比例高出10倍以上(图3f)。此外,ra患者的滑膜组织中存在cd20
+
cd11c
+
abcs(图3g),表明这些细胞可能具有迁移到炎症组织中的能力。
67.为了探索药物对abcs的潜在影响,根据接受的不同治疗对患者进行分组。然而,在接受不同治疗的患者之间观察到类似数量的abcs,包括皮质类固醇、csdmard(甲氨蝶呤、来氟米特、羟氯喹、柳氮磺胺吡啶)和生物制剂(图10i-k)。
68.最近,已经确定一种新的外周辅助t细胞亚群(t
ph
)在ra患者中表现为扩增。重要的是,t
ph
细胞可以通过分泌il-21诱导b细胞向浆细胞分化;其中,il-21是一种细胞因子,已被证实是abcs产生的关键调节因子。因此,本发明人评估了ra患者中abcs和t
ph
细胞之间的关联。结果证实,与spa和健康对照组相比,ra患者循环中t
ph
细胞的频率升高(图3h)。此外,abcs的比例和数量与t
ph
细胞呈正相关(图3i和图10l)。最近的一项研究表明,在体外条件下,滑膜t
ph
细胞通过提供il-21和ifn-γ,使b细胞分化向abc表型倾斜;可以合理地推测:t
ph
细胞扩增可能促进ra患者b细胞向abc分化。
69.六、转录组分析揭示了abcs的不同特征
70.为了更好地了解ra abcs与幼稚、记忆b细胞的关系,基于cd19、cd27和cd11c的表达状况对b细胞进行分类,并采用基于rna的测序(rna-seq)对幼稚b细胞(cd19
+
cd11c-cd27-)、记忆b细胞(cd19
+
cd11c-cd27
+
)和abc(cd19
+
cd27-cd11c
+
)的转录组进行比较。如维恩图所示,与幼稚、记忆b细胞相比,abcs显示出不同的基因表达模式(图4a)。总体而言,与幼稚b细胞相比,abcs中有1505个基因被上调,498个基因被下调。相比之下,abcs和记忆b细胞显示出相似的转录谱,与记忆b细胞相比,abcs中只有283个基因上调,153个基因下调(fold change》2,fdr《0.05;图11a)。值得注意的是,abcs-vs-记忆b细胞中的大多数上调基因(198/283)在abcs-vs-幼稚b细胞中也上调,表明这些基因是abcs特异性的。与先前在sle中的发现一致,abcs的一些关键表型标志物通过mrna表达得到了证实,例如itgax(cd11c),tbx21(t-bet),fcrl5,il-21r的表达升高和cd27、cd38的表达降低(数据未显示)。然而,尽管sle中的abcs被确定为浆细胞的前体,但本发明人并没有发现浆细胞标志物的表达上调,例如prdm1和xbp1。接下来,本发明人通过过度表示分析将degs映射到已知的基因本体生物学过程。值得注意的是,与幼稚b细胞相比,abcs的中性粒细胞活化、白细胞迁移和粘附相关基因显着上调(图4b,c)。且当将abcs与记忆b细胞进行比较时,观察到了类似的结果(图11b和c)。进一步地,通过qpcr证实了ccr2和cxcr3在abcs中的上调表达(图4d)。结合发现的abcs在ra患者的炎症关节中存在并扩增,这些数据揭示了ra中abcs的独特特征,并表明循
环abcs能够迁移到炎症关节以促进疾病的发病机制。
71.七、abcs通过不依赖细胞接触的方式诱导fls的激活
72.众所周知,fls及其与免疫细胞的相互作用在ra滑膜炎的进展中起着重要作用。长期以来,研究人员一直关注t细胞,尤其是cd4
+
t细胞与fls之间的相互关系,而b细胞与fls之间的串扰则鲜为人知。最近,一项研究表明,tnfα刺激增加了b细胞粘附fls的能力。为了确定abcs是否可以在体外与fls相互作用,对ra患者的abcs进行分类,并将它们与fls共培养3天。通过形态和cd55表达情况(12a,b),来自ra患者滑膜组织的fls证实了该点。首先,本发明人通过流式细胞术检查了icam-1和vcam-1在fls中的表达,且后者已被证明有助于b细胞粘附到fls。与证明cd4
+
t细胞和fls之间相互作用的发现相似,在abcs存在下,icam-1和vcam-1在fls中上调,表明fls被激活(图5a)。接下来,本发明人测量了上清液中的il-6、mmps水平(fls激活的标志)。值得注意的是,与单独的fls或与cd11c-b细胞共培养的fls相比,fls与abc共培养的上清液中il-6、mmp-1、mmp-3和mmp-13的水平显着升高(图5b)。为了进一步阐明abcs诱导的fls的激活是否依赖于细胞接触,用abcs条件培养基(abcscm)处理fls。值得注意的是,在abcscm和与fls共培养的abcs之间观察到相当数量的il-6和mmps产生,这表明细胞接触对于abcs激活fls是不必要的(图5c)。值得注意的是,在与fls共培养的abcscm和abcs细胞之间,il-6和mmps的产生量相当,这表明abcs细胞激活fls是不需要细胞接触的(图5c)。因此,在共培养实验中,使用abcscm代替abcs以进一步研究其机制。
73.八、abcscm衍生的tnf-α会促进fls中干扰素刺激基因的上调
74.上述数据表明,abcs以不依赖于细胞接触的方式作用于fls,通过上调粘附分子和增加促炎细胞因子的产生,以诱导活化和侵袭性的表型。为了研究源自abc的fls激活介质,使用细胞计数珠阵列量化了abcscm中的几种炎性细胞因子和ifn。值得注意的是,abcscm中的tnf-α和il-1β水平显着升高;而在abcscm和幼稚b细胞培养基之间,il-17、ifn-α和ifn-γ水平相当(图5d)。添加抗tnf-α可下调il-6、mmp-1、mmp-3和mmp-13的产生,而添加抗il-1β抗体则不下调,表明abcscm衍生的tnf-α负责fls的激活(图5e)。
75.为了进一步探索fls激活的机制,通过rna-seq以比较用或不用abcscm处理fls的转录组。如图6a所示,与单独的fls相比,用abcscm处理的fls中共有144个基因显着上调,而3个基因下调(fold change》2,fdr《0.05)。值得注意的是,在abcscm处理的fls中,上调的富集通路包括与干扰素(ifn)信号传导(i型和ii型ifn信号传导)相关的通路(图6b-d)。这与之前的一项研究一致,该研究表明fls中tnf诱导的趋化因子表达是依赖于i型ifn的。
76.为了预测可能靶向deg的转录因子(tf),基于msigdb基因集中的已知靶基因进行了tf富集分析。在前10个富集的tf中,与单独的fls相比,在abcscm处理的fls中stat1、irf1和irf7显着上调(图7a)。随后,进行蛋白质-蛋白质相互作用(ppi)分析,将排名前15位且程度最高的基因视为枢纽基因,包括stat1、irf7和irf1(图7b)。根据tf预测、ppi和差异表达分析的p值对基因进行排序,最终选择stat1作为排名靠前的基因进行进一步确认。
77.九、阻断erk1/2和jak-stat1通路可以阻止abcs诱导的fls激活
78.基于以上结果,本发明人进一步验证abcscm处理的fls是否诱导了stat1。图13a显示,磷酸化stat1的表达在abcscm刺激后15分钟增加,并且抗tnf-α抗体可以降低这种增加(图7c和图13a)。值得注意的是,用特异性抑制剂抑制stat1可使上清液中mmp-1、mmp-3和mmp-13的水平降低至fls
+
abcscm组的一半,而il-6的产生不受影响,这表明stat1部分调节
mmp1、mmp3和mmp13,而不调节il-6(图7d)。这可以通过先前的研究来解释,先前研究表明stat1磷酸化是响应il-6信号传导而发生的。为了进一步评估是否janus激酶(jak)控制了由abcscm诱导的fls激活,在jak抑制剂baricitinib存在下重复了该实验。结果显示,使用jak抑制剂可以显著抑制磷酸化stat1的表达以及il-6、mmp-1、mmp-3和mmp-13的产生(图7d,e)。
79.除此之外,还有其他途径,已被报道调节fls中mmp的产生,如mapk。abcscm处理诱导fls中的perk1/2,添加中和tnf-α抗体后,该诱导下调(图7c和图13b)。同时,抑制perk1/2可显著降低mmp-1、mmp-3、mmp-13和il-6的产生(图7f),表明abcscm衍生的tnf-α诱导的perk1/2可调节il-6、mmp1、mmp-3和mmp-13。
80.总之,结果表明,abcs通过tnf-α介导的erk1/2和jak-stat1通路诱导fls的激活。
81.总体上,本实施例中,发明人发现abcs在cia小鼠中以及ra患者的循环和炎症关节中增加。转录组分析揭示了,在raabcs中,具有上调趋化性和迁移相关基因表达的独特转录谱。实际上,我们发现abcs在滑膜液中的频率比在外周血中高10倍以上,这表明它们是从循环中募集到炎症关节的。此外,我们发现循环abcs与疾病活动度呈正相关,类似于sle中的发现。这些数据表明abcs是ra发病机制中的关键贡献者,并且是监测疾病活动的敏感指标。
82.abcs的形成受先天和适应性信号的调节。对小鼠系统的早期研究揭示了先天信号、tlr7和tlr9在促进abc的分化和生成中起到关键作用。除了先天性刺激之外,人们认为il-21和ifn-γ等细胞因子在控制小鼠和人类的abcs生成方面发挥重要作用。最近,一项研究通过研究具有明确先天免疫缺陷的患者,证实了bcr和t细胞衍生的il-21在abcs体内扩增中起到重要作用,表明体内炎症环境也参与调节abcs分化。在本实施例中,本发明人发现与非关节炎对照组相比,在cia小鼠中,il-21和tlr7激动剂的共同刺激会驱动更多的abcs生成。值得注意的是,本发明人发现患者中abcs的数量与外周t
ph
的数量呈正相关;外周t
ph
是一种独特的辅助t细胞亚群,已被确定其能通过产生il-21以促进b细胞分化为浆细胞。这与最近在幼年特发性关节炎(jia)中的发现一致,该发现表明jia患者关节中t
ph
细胞的扩张,并揭示了滑膜t
ph
频率与原位abcs呈正相关。jia患者的滑膜t
ph
细胞通过提供il-21和ifn-γ使b细胞在体外向abcs表型分化。因此,认为t
ph
细胞通过ra患者分泌细胞因子(如il-21和ifn-γ)促进b细胞分化为abcs。
83.为了进一步描述abcs在关节炎环境中的效应潜能,通过体外共培养实验研究了abcs和fls之间的串扰。本研究表明,abcs与fls相互作用,在不依赖细胞接触的方式下,通过增加il-6和mmps的分泌来诱导活性表型。进一步的研究证实,abcscm衍生的tnf-α参与fls的激活。接下来,进行rna-seq以更深入地了解fls如何被abcscm激活。值得注意的是,abcscm促进了fls中干扰素刺激基因的上调。此外,我们测试了激活的fls中的erk1/2和stat1磷酸化,并验证了阻断erk1/2和jak-stat1通路在预防abcscm诱导的fls激活中的作用。该结果具有临床意义,因为像baricitinib一类的jak抑制剂已被批准用于活动性ra的临床治疗。即本实施例研究结果表明,abcs通过tnf-α介导的erk1/2和jak-stat1通路诱导fls的激活,从而导致滑膜炎的慢性发生。
84.显然,本发明的上述实施例仅仅是为清楚地说明本发明技术方案所作的举例,而并非是对本发明的具体实施方式的限定。凡在本发明权利要求书的精神和原则之内所作的任何修改、等同替换和改进等,均应包含在本发明权利要求的保护范围之内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1