一种具有情绪化表现的机器人的制作方法
一种具有情绪化表现的机器人
1.技术领域 本发明涉及人工智能技术领域的一种机器人。
2.
背景技术:
机器人包括硬件系统和控制系统,控制系统通过电子电路和软件(程序)控制硬件机构实现各种功能动作,目前的控制系统采用人工智能技术,能够实现对视觉、听觉信息的识别和处理,包括对人类语言的感知、识别和执行,但对于同时面对多种信息的输入、识别、反射和多路输出控制之间的整合和平衡处理方面,尤其是包含有情绪调控方面,并无能够模拟其工作的相关技术。
3.
技术实现要素:
本发明目的是公开一种机器人,能够形成对外部指令的“情绪化”状态表示,尤其是还能够模拟对外部指令产生不合逻辑的“情绪化”反应。以此也模拟或演示人脑情绪反应的工作机制。
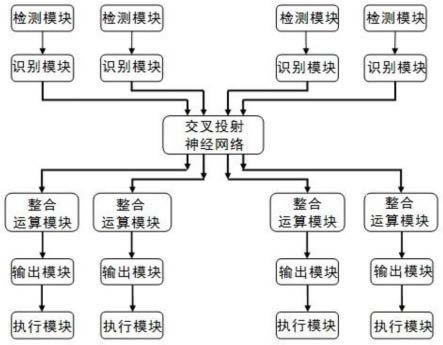
4.本发明的机器人包括硬件系统和控制系统,硬件系统包括机械结构和执行模块,控制系统包括至少一个的外部信息反应通道和至少一个的内部信息反应通道。
5.一个外部信息反应通道包括有:检测模块,用于检测接收来自机器人系统外部的某种外部信息,比如声音、影像、机械振动、气味等信息的一种;识别模块,用于对所接收到的外部信息进行识别;外部信息整合运算模块,用于对识别后的外部信息进行整合和运算,(比如数据运算或逻辑运算);输出模块,用于将外部信息整合运算模块所输出的运算结果,转化为输出信号,并连接到硬件系统的执行模块。
6.一个内部信息反应通道包括有:检测模块,用于检测接收来自机器人系统自身的某种内部信息,比如温度、压力、电压等的一种;识别模块,用于对所接收到的内部信息进行识别;内部信息整合运算模块,用于对识别后的内部信息进行整合和运算,(比如数据运算或逻辑运算);输出模块,用于将内部信息整合运算模块所输出的运算结果,转化为输出信号,并连接到硬件系统的执行模块。硬件系统的执行模块用于接收并执行外部信息反应通道输出模块和内部信息反应通道输出模块的各种输出信号。
7.本发明的技术特征在于:在外部信息反应通道的识别模块与外部信息整合运算模块之间,和内部信息反应通道的识别模块与内部信息整合运算模块之间,连接有一个交叉投射神经网络。
8.该交叉投射神经网络具有至少一个外部信息调制神经通道和至少一个内部信息调制神经通道。一个外部信息调制神经通道的输入端和输出端,分别连接到一个外部信息反应通道的识别模块的输出端、和外部信息整合运算模块的输入端;一个内部信息调制神经通道的输入端和输出端,分别连接到一个内部信息反应通道的识别模块的输出端、和内部信息整合运算模块的输入端。
9.在神经网络内部,一个外部信息调制神经通道包括有一个神经输入层(外部信息神经输入层)和一个神经输出层(外部信息神经输出层),神经输入层的输入端作为外部信息调制神经通道的输入端,神经输出层的输出端作为外部信息调制神经通道的输出端;神经输入层的输出端连接神经输出层的输入端。
10.同样的,一个内部信息调制神经通道包括有一个神经输入层(内部信息神经输入层)和一个神经输出层(内部信息神经输出层),神经输入层的输入端作为内部信息调制神
经通道的输入端,神经输出层的输出端作为内部信息调制通道的输出端;神经输入层的输出端连接神经输出层的输出端。
11.所述的交叉投射是指外部信息调制神经通道的外部信息神经输入层的每一个输出端,还同时投射到内部信息调制神经通道的内部信息神经输出层的各个输入端,(每一个输出端的信号作为这些输入端的输入信号)。同样的内部信息调制神经通道的内部信息神经输入层的每一个输出端,还同时投射到外部信息调制神经通道的外部信息神经输出层的各个输入端,(每一个输出端的信号作为这些输入端的输入信号)。
12.具体到神经元的连接,外部信息神经输入层和外部信息输出层各自包括有多个神经元,两者神经元数量相等且一一对应。外部信息神经输入层的每一个神经元的输出端(轴突输出端),只投射到外部信息神经输出层的相对应的一个神经元的输入端(树突输入端),通过突触(具有突触可塑性的模拟突触)与输入端进行连接。(注意这一点跟现有神经网络是不一样的,现有的神经网络是广泛性的互相投射,而本发明在同样的外部或内部神经通道里面,只有一个神经元的输出端投射到对应的一个神经元的输入端)。
13.同样的,内部信息神经输入层和内部信息输出层各自包括有多个神经元,两者神经元数量相等且一一对应。内部信息神经输入层的每一个神经元的输出端(轴突输出端),只投射到内部信息神经输出层的相对应的一个神经元的输入端(树突输入端),通过突触(具有突触可塑性的模拟突触)与输入端进行连接。
14.具体到神经元的所述的交叉投射,便是外部信息神经输入层的每一个神经元的输出端(轴突输出端),还同时投射到外部信息神经输出层的各个(所有的、每一个)神经元的输入端(树突输入端),通过突触(具有突触可塑性的模拟突触)与输入端进行连接。同样的,内部信息神经输入层的每一个神经元的输出端(轴突输出端),投射到外部信息神经输出层的各个(所有的、每一个)神经元的输入端(树突输入端),通过突触(具有突触可塑性的模拟突触)与输入端进行连接。(注意这一点跟现有神经网络是不一样的,本发明只是在不同的外部信息神经通道或内部信息神经通道之间才存在互相投射)。
15.本发明还可在内部信息整合运算模块或输出模块的输出端,设置一条连接外部信息整合运算模块的调制通道,当内部信息反应通道出现特定输出时,同时调节外部信息整合运算模块的输出信号。
16.本发明具有对外部指令的响应执行,对内部参数的监测和显示,并分别有多个层次的情绪反应:
17.1、合乎逻辑的情绪反应:在一般工作状态下,负责“外部信息检测—控制输出”的外部信息反应通道根据所输入的外部信息比如指令去控制执行部件,在正常逻辑下完成指令控制,比如:指令输入“加速”则加快执行部件比如电机转速以加快动作;同时负责“内部信息检测—状态显示”的内部信息反应通道能够检测机器内部的某些参数,在正常逻辑下进行状态显示,比如:当检测到电机温度偏高,则在显示器上显示“痛苦”的情绪表现,并启动散热风扇和降低电机转速。这能够模拟正常的情绪反应,这种反应是合乎逻辑的,其实质仍属于正常的输入—输出自动控制,当电机温度降下来,则会停止“痛苦”的表示和恢复正常电机转速。这种情绪反应属于通过预先的设定进行工作,且能够合乎逻辑的进行解释,通过目前的现有技术也可以实现这一层次的情绪模拟反应。
18.2、不合逻辑的“闹情绪”反应:如果不断发出某种口令比如“加速”,使电机较长时
间处在快速工作状态,导致电机温度已经偏高,内部信息反应通道也已经显示“痛苦”情绪,这时还不断发出“加速”指令,则开始通过神经网络的学习机制形成情绪反应并形成记忆。形成情绪记忆时,当接收到口令“加速”,则不管电机温度是否偏高,(即使停止工作一段时间后电机温度已经降低为常温),它仍然输出“痛苦”反应,并拒绝加快电机转速,反而继续降低电机转速和启动散热风扇。这是一种不合逻辑的反应,模拟的是人的情绪处于懊恼的状态时,对外部指令有违情理的对抗性的闹情绪反应,所以即使电机常温,只要接收到“加速”指令,它都直接输出“痛苦”情绪反应,而且对抗性的反过来降低电机转速。
19.3、“情绪化”反应的记忆效应和“去情绪化”(恢复功能):闹情绪的反应会由于记忆效应而存在一定时间,在记忆效应的有效期内始终存在情绪化表现。但能够通过多次输入“去情绪化”的指令比如“减速”,使电机不断降温,不断减弱直至消除记忆效应,这类似于人类的通过安抚来消除情绪化。
20.本发明的机器人在进行多种信息输入和多路控制输出的基础上,还能够模拟人类情绪反应以实现更复杂或更有趣的功能。本发明不同于现有技术通过电路或软件来设定情绪反应,它除了能够模拟正常的“合乎逻辑”的情绪反应,还能够模拟更有意思的“不合逻辑的闹情绪”反应,而且这种情绪化反应不是预先设定的,而是在工作中通过反复的口令-执行-反馈这个过程而得到的“经验”,这更加符合人类闹情绪的形成机制。本发明还能够通过多次输入“去情绪化”的指令来减弱直至消除情绪化效应,类似于人类的通过安抚来消除情绪化。当然人脑的情绪机制是复杂的,本发明只是进行某种程度的模拟。
21.附图说明 图1是本发明的控制系统的原理方框图。图2是一个实施例的原理方框图。图3是交叉投射神经网络的一种内部结构示意图。图4是交叉投射神经网络的另一种内部结构示意图。
22.图5是典型的生物进化树的示意图。图6是肠腔动物神经系统的示意图。图7是线形动物神经系统的示意图。图8是线形动物神经系统等效的信息处理架构的示意图。图9是节肢动物神经系统的神经投射示意图。图10是节肢动物神经系统神经投射示意图的另一种表示结构。图11是节肢动物神经系统等效的信息处理架构方框图。图12是鱼类神经系统的神经投射示意图。图13是鱼类神经系统等效的信息处理架构的方框图。图14是鱼类神经系统的多向调节神经回路与其他神经回路的神经投射关系示意图。图15是图14神经系统所等效的信息处理架构方框图。图16是爬虫类动物神经系统的神经投射关系示意图。图17是鸟类动物的自主神经中枢的神经投射关系示意图。图18是鸟类动物神经系统的神经投射关系示意图。图19是哺乳动物神经系统的视觉信号相关核团的神经投射示意图。图20是哺乳动物神经系统的听觉信号相关核团的神经投射示意图。图21是哺乳动物神经系统的运动神经中枢的神经投射示意图。图22是哺乳动物神经系统的海马环路与端脑皮质等核团的神经投射示意图。图23是哺乳动物神经系统的自主神经中枢的神经投射示意图。图24是哺乳动物神经系统的脑内多向调节神经中枢与其他神经回路的神经投射示意图。图25是人脑总体的系统框架示意图。图26是人脑主要核团的神经投射示意图。图27是图26的神经投射示意图所等效的主要信息处理架构的方框图。
23.具体实施方式 以下为对本发明的原理和具体实施的叙述。
24.本发明的机器人包括硬件系统和控制系统,硬件系统包括机械结构和执行模块,执行模块用于接收并执行控制系统的各种输出信号来实现各种功能,比如运动、显示和声
音输出等,这属于现有技术。本发明的创新在于其控制系统,它模仿了人脑对机体外部信息和内部信息的整合,以及情绪化反应的功能。
25.根据后面所附的关于人脑工作框架的研究资料。人脑的系统框架可视为两个大的信息处理系统和一个小的脑内多向调节系统。
26.一个是机体内部信息处理系统,也即自主神经系统,包括内脏调节神经中枢、化学和内分泌调节神经中枢。主要由孤束核、臂旁核、下丘脑、上丘脑等核团构成,用于接收来自机体内部各种传感器传入的机体内部信息,经过整合处理后通过交感神经和副交感神经输出,对各个内脏器官、血压、血糖、体温等等和免疫内分泌系统的工作进行调节,内部信息的整合处理过程一般是不可知觉的且在无意识下进行的。
27.另一个是机体外部信息处理系统,包括感觉神经中枢、思维神经中枢和运动神经中枢。主要由脑干的几个网状核团、丘脑、海马环路、基底环路、小脑和端脑皮质等构成,由于接收来自视觉、听觉、触觉等各种感觉器官传入的机体外部信息,经过整合处理后输出对机体运动(包括说话和书写)进行控制,思维神经中枢对这一过程进行决策和控制,同时也完成对各种中间信息的再整合。
28.而边缘系统的杏仁体和脑干的中缝大核、中缝背核、蓝斑核、被盖腹侧区(vta)等若干调制性核团则构成一个小而重要的脑内多向调制系统,对内部信息处理系统和外部信息处理系统各个神经环路的工作进行相互调节,使彼此的工作状态能够协调和适应,并由此产生了情绪、欲望、动机、成瘾等等有趣而神秘的宏观表现,人类的情绪中枢正是这一调节系统的组成部分。
29.本发明模仿了人脑的这一系统框架,如图1,本发明的控制系统包括至少一个外部信息反应通道和至少一个内部信息反应通道。(图1是内外各两个信息反应通道)。
30.一个外部信息反应通道包括有:外部信息检测模块,用于检测接收来自机器人系统外部的外部信息,比如听觉(声)、视觉(光)、触觉(机械振动)、嗅觉(化学)等信息的一种;识别模块,用于对所接收到的外部信息进行识别;外部信息整合运算模块,用于对识别后的外部信息进行整合和运算,(比如数据运算或逻辑运算);输出模块,用于将外部信息整合运算模块所输出的运算结果,转化为输出信号,并连接到硬件系统的执行模块。执行模块包括多种装置,比如电机、显示屏、喇叭等,用于接收并执行外部信息反应通道输出模块和内部信息反应通道输出模块的各种输出信号。所述的外部信息是指来自机器人外部的信息,比如视频传感器接收的视频影像,声音传感器接收的音频信息,化学传感器接收的化学信息比如气味,外部信息类似于人类通过眼睛看到的影像,通过耳朵听到的声音,通过鼻子嗅到的气味等等。
31.一个内部信息反应通道包括有:内部信息检测模块,用于检测接收来自机器人系统自身的一种内部信息,比如温度、压力、电压等的一种;内部信息识别模块,用于对所接收到的内部信息进行识别;内部信息整合运算模块,用于对识别后的内部信息进行整合和运算,(比如数据运算或逻辑运算);内部信息输出模块,用于将内部信息整合运算模块所输出的运算结果,转化为输出信号,并连接到执行模块。所述的内部信息是指来自机器人自身各种部件的各种物理信息,比如电机温度,电池的电压和电量,液压系统的压力等等。内部信息类似于人类所感觉接收的来自机体内部器官的血压、血糖、体温等等信息。
32.外部信息反应通道的外部信息识别模块和外部信息整合运算模块,内部信息反应
通道的识别模块和内部信息整合运算模块,既可以采用传统的电子技术来实现,也可以采用神经网络来实现其信号识别、信息整合与运算的功能,比如采用ann神经网络来识别声音指令和整合多位信号等等,这些都是现有技术。
33.本发明的特征在于:在外部信息反应通道的识别模块与外部信息整合运算模块之间,和内部信息反应通道的识别模块与内部信息整合运算模块之间,连接有一个交叉投射神经网络。
34.该交叉投射神经网络具有至少一个外部信息调制神经通道和至少一个内部信息调制神经通道。一个外部信息调制神经通道的输入端和输出端,分别连接到一个外部信息反应通道的外部信息识别模块的输出端、和外部信息整合运算模块的输入端;一个内部信息调制神经通道的输入端和输出端,分别连接到一个内部信息反应通道的识别模块的输出端、和内部信息整合运算模块的输入端。
35.图2的方框图是一个简单的实施例。外部信息反应通道和内部信息反应通道都只有一个通道。其中外部信息反应通道用于对声音进行信息处理,采用声音接收(麦克风)作为检测模块,识别模块能够识别声音指令,并输出多位的并行信号,每一位输出信号代表一种指令,比如“加速”、“减速”、“停止”等。然后这些并行信号输入到交叉投射神经网络,从交叉投射神经网络返回的并行信号再输入整合与运算模块,对这些并行信号进行整合或/和运算,得到控制输出信号,再由输出模块放大或缓冲后,输出并控制执行模块比如电机的运转,以“加速”、“减速”、“停止”等来完成某种机械动作。内部信息反应通道用于对电机温度进行信息处理,采用温度检测(温度传感器)作为检测模块,识别模块能够识别温度值,将温度值分为几个区域,并输出多位的并行信号,每一位输出信号代表一种温度范围,比如“小于50度”、“大于50度而小于80度”、“大于80度”等。然后这些并行信号输入到交叉投射神经网络,从交叉投射神经网络返回的并行信号再输入整合与运算模块,对这些并行信号进行整合或/和运算,得到控制输出信号,再由输出模块放大或缓冲后,输出并控制执行模块的工作,比如分别用“愉快”、“平静”、“痛苦”等显示来表现“小于50度”、“大于50度而小于80度”、“大于80度”等状态的情绪反映,并在“痛苦”时启动散热风扇的工作,对电机进行散热来“减轻这种痛苦”。还可在内部信息整合与运算模块或输出模块的输出端,设置一条连接外部信息整合与运算模块的调制通道,当内部信息反应通道出现“痛苦”情绪时,同时调节外部信息整合与运算模块的输出信号,比如直接降低电机转速。
36.交叉投射神经网络的结构如图3(对于ann神经网络)或者图4(对于snn脉冲神经网络)。在神经网络内部,一个外部信息调制神经通道包括有一个神经输入层(外部信息神经输入层)和一个神经输出层(外部信息神经输出层),神经输入层的输入端作为外部信息调制神经通道的输入端,神经输出层的输出端作为外部信息调制神经通道的输出端;神经输入层的输出端连接神经输出层的输入端。
37.同样的,一个内部信息调制神经通道包括有一个神经输入层(内部信息神经输入层)和一个神经输出层(内部信息神经输出层),神经输入层的输入端作为内部信息调制神经通道的输入端,神经输出层的输出端作为内部信息调制通道的输出端;神经输入层的输出端连接神经输出层的输出端。
38.所述的交叉投射是指外部信息调制神经通道的外部信息神经输入层的每一个输出端,还同时投射到内部信息调制神经通道的内部信息神经输出层的各个输入端,(每一个
输出端的信号作为这些输入端的输入信号)。同样的内部信息调制神经通道的内部信息神经输入层的每一个输出端,还同时投射到外部信息调制神经通道的外部信息神经输出层的各个输入端,(每一个输出端的信号作为这些输入端的输入信号)。
39.具体到神经元的连接,外部信息神经输入层和外部信息输出层各自包括有多个神经元,两者神经元数量相等且一一对应。外部信息神经输入层的每一个神经元的输出端(轴突输出端),只投射到外部信息神经输出层的相对应的一个神经元的输入端(树突输入端),通过突触(具有突触可塑性的模拟突触)与输入端进行连接。(注意这一点跟现有神经网络是不一样的,现有的神经网络是广泛性的互相投射,而本发明在同样的外部或内部神经通道里面,只有一个神经元的输出端投射到对应的一个神经元的输入端)。
40.同样的,内部信息神经输入层和内部信息输出层各自包括有多个神经元,两者神经元数量相等且一一对应。内部信息神经输入层的每一个神经元的输出端(轴突输出端),只投射到内部信息神经输出层的相对应的一个神经元的输入端(树突输入端),通过突触(具有突触可塑性的模拟突触)与输入端进行连接。
41.具体到神经元的所述的交叉投射,便是外部信息神经输入层的每一个神经元的输出端(轴突输出端),还同时投射到外部信息神经输出层的各个(所有的、每一个)神经元的输入端(树突输入端),通过突触(具有突触可塑性的模拟突触)与输入端进行连接。同样的,内部信息神经输入层的每一个神经元的输出端(轴突输出端),投射到外部信息神经输出层的各个(所有的、每一个)神经元的输入端(树突输入端),通过突触(具有突触可塑性的模拟突触)与输入端进行连接。(注意这一点跟现有神经网络是不一样的,本发明只是在不同的外部信息神经通道或内部信息神经通道之间才存在互相投射)。
42.在交叉投射神经网络中,由于外部信息神经通道和内部信息神经通道之间存在交叉投射,如果两个神经元的活动经常具有相关性,那么它们之间的突触联系会出现突触传递效能的增强,最终使得它们的活动具有记忆效应。所以,这一交叉投射神经网络的作用是,在工作过程中,如果经常有一个外部信息跟一个内部信息存在相关性活动,比如经常存在一个外部信息(比如“加速”指令)出现后总是伴随着一个内部信息(比如电机温度大于80度)的出现,(这种情况就是在电机已经工作很长时间,温度已经很高时,还不断喊出“加速”指令),从神经元层面说,就是经常存在一个外部信息神经通道的神经元出现活动后便伴随着一个内部信息神经通道的神经元也出现活动,那么,它们之间的联系会产生增强,(突触可塑性增强),使得它们之间建立起记忆效应。而一旦它们之间建立了记忆效应,其结果是,在该外部信息神经通道的神经元再次由于接收到输入信号(“加速”指令)而出现活动时,即使该内部信息神经通道的神经元没有接收到输入信号,(电机由于散热而温度下降,温度没有大于80度),这时也会由于与外部信息神经通道对应神经元的联系的记忆增强作用,而使该内部信息神经通道的神经元出现活动,并输出对应的信号。从神经网络的输入和输出看,便是当该外部信息神经通道由于再次接收到对应输入信号(“加速”指令),即使对应的内部信息神经通道没有接收到输入信号(温度大于80度),但由于存在记忆效应,该内部信息神经通道也会再次出现输出信号,显示“痛苦”的情绪反映。这种“痛苦”的情绪反映不是真正的电机状态“温度没有大于80度”,而是对之前频繁得到“加速”指令的一种非正常反映,这模拟了人类那种非理性的情绪化的情绪。这种“闹情绪”的反应会由于记忆效应而存在一定时间,需要通过多次输入反向指令“减速”,使电机不断降温,减弱直至消除记忆效应来完
成,类似于人类的通过安抚来消除情绪化。
43.本发明的神经网络包含神经元和突触,这可以在计算机上通过软件来模拟和实现,就如同当前的各种ann人工智能神经网络。也可以采用电子元件构成的模拟神经元和模拟突触来实现,就如同当前的snn脉冲神经网络。一般来说,模拟神经元包括有输入端(模拟树突输入端)、运算模块和轴突输出端(模拟轴突输出)。模拟突触包括有输入端(模拟突触前膜)、运算模块和输出端(模拟突触后膜),根据其突触传递效能是否具有可塑性,模拟突触又可分为模拟固定突触、模拟可塑性突触、和模拟预置性突触。其中模拟固定突触用于模拟信息输入和输出通道上的突触,还有胆碱能神经对其他神经元投射的突触,其一般没有突触传递可塑性;模拟可塑性突触用于模拟海马体、杏仁体等核团的氨基酸能神经之间的突触,具有长时程可塑性,尤其是时间依赖的突触可塑性(stdp可塑性);模拟预置性突触模拟的是一些端脑皮质区域的锥体神经元之间的突触,其在开始时尚未形成有效突触连接(或突触尚未形成有效传递效能),在两个神经元之间反复出现具有关联性的动作电位发放动作后,在它们之间才慢慢形成突触连接(或其突触开始产生传递效能),预置性突触模拟的是端脑皮质的锥体神经元之间通过突触重构形成的长期记忆(或称皮质记忆或永久记忆)。模拟神经元、各种模拟突触、以及它们之间的具体连接技术属于现有技术,可使用现有的各种相关技术,也可参考申请人之前提交的申请号为2014106066977,2018107602170和2017100121546的中国专利申请所公开的相关技术。
44.附带资料:关于人脑的形成及其工作框架。以下部分以生物进化的几个基本原则为基础,分析神经系统在生物进化过程中如何适应机体的进化而进行演变,推演出当前阶段的人脑进行信号处理的系统框架,这也是本发明设计所依据的基本原理。至于大脑在思维及运动控制方面的工作,可参考申请人之前提交的申请号为2015107802671和2015101775882的中国专利申请的说明书中所附的资料。
45.一、脑科学研究的现状和关键问题。
46.1.1脑科学研究的不同层面。
47.从大的方面看,对脑的研究有三个层面。细胞层面:描述神经细胞、突触等的分子生物学的工作原理及工作过程;神经环路层面:描述人脑各个核团或区域之间的神经投射及信息处理;功能层面:描述人脑在宏观上可观察的功能及现象。我们的目的是以这三个层面来分析整个人脑的工作原理:以细胞层面的分子生物学为基础,分析信息在人脑及其各个神经环路的处理过程,并以此解释各种脑功能或现象。
48.借鉴电子技术,如果将大脑视为一个具有多路输入和输出的智能控制装置,那么细胞层面是指构成装置的各种电子元件,神经环路是连接电子元件的电路图,功能层面便是其控制功能。我们的目的是以各种元件的工作特性为基础,分析装置内部的电路图及其工作原理,并以此解释它如何实现各种控制功能。
49.1.2我们目前的研究现状。细胞层面上,通过各种分子实验和计算神经科学,我们对神经元、突触、受体的结构、特性和功能了解得不错,相当于基本弄明白控制装置里面各种电子元件的原理和工作特性。
50.功能层面上,通过各种现象观察和心理实验,我们也对大脑各个模块的高级功能有足够的认识,相当于对控制装置的信号输入输出关系和各种控制功能也很清楚。
51.我们还通过很多神经科学方面的实验发现一些相关性结论。比如发现大脑表现某
种功能时与哪些神经核团或者哪些神经细胞的什么活动有关,或者发现某个核团或神经束出现异常时大脑会表现出哪些功能异常,甚至能够确定某个具体的神经环路便是与某个脑功能对应的。这相当于发现控制装置完成某种控制功能时其内部哪些电子元件的信号会出现什么变化,或者发现摘掉某个元件或切断某条连线会导致出现什么控制功能异常,甚至能够确定了某个局部电路便是实现某个控制功能。
52.1.3我们面临的关键问题在于:我们未能建立整个大脑的系统模型及其工作机制,以便能够由上而下的普适性的在各个神经环路中去描述微观上神经细胞的活动如何产生宏观上大脑的高级功能和现象。相当于我们未能整体描绘出控制装置的电路图(即使是电路原理方框图)并描述其工作原理,以便以此去普适性的分析控制装置的各种输入输出及其逻辑和控制功能是如何实现的。显然,解决这一问题也能够给人工智能尤其是通用人工智能的发展带来极大的启示。
53.这一问题主要包括两方面:一是未能准确和完整的描绘出大脑中各种核团的神经联系,二是未能描述出各个神经环路进行信息处理的机制。其中第二点是更主要的,因为即使从解剖上我们已经描绘出一些神经环路的神经投射关系,却未能弄清它们是怎样进行信息处理的。这相当于我们很清楚那些电子元件是什么,也描绘出它们之间的一些连接关系,但就是无法解释它们是怎么工作的。
54.1.4我们的研究方向错了吗?
55.从技术手段看,我们似乎研究得够深了,已经更深入到细胞内部的工作,了解各种神经递质及其受体通道的具体结构和详细工作过程,这绝对不是以前说的“盲人摸象”,而是拿着电刀和放大镜等工具在解剖并观察大象的各个部位,按理说是应该能够建立出大象的模型并进行各种合理解释了,但实际没有。
56.既然看起来我们目前应该能够弄明白人脑的工作原理了,而事实上又是没有,那么,一定有什么地方不对劲。我们的研究方向错了吗?
57.大脑是一个极度复杂的系统,如果总是由下而上的根据一些实验现象去寻找现象背后的机制,得到的往往只是一些局部的细节性的因果关系,容易使我们一叶障目,或者只是得到一些相关性结论而无法获得其深层的原理。我们是不是需要反思,对于大脑,这种试图采用归纳法通过实验观察来寻找机制的研究方向是否是正确的?
58.我们是否更应该从某些基本原则出发,分析在生物进化过程中,神经系统如何出现,又如何演变以适应机体及环境的变化?分析从简单动物进化到哺乳动物和人这一过程,中枢神经会演变为怎样的系统架构?以此通过演绎法推导出当前阶段的人脑“应该存在”怎么样的模型,并建立理论框架。这个模型和理论可以普适性的解释大脑各个功能模块和神经环路的工作,还能够以此进行一些推论,而这些推论可以设计实验来验证。
59.申请人试图从这样的思路来分析和建立人脑的模型和系统框架,而且依赖的仅仅是几个基本原则和一些已经得到大家公认的动物学和脑科学的知识。或许我们还可以发现,这一演绎过程及其结果会是如此之顺畅和谐,体现了“宇宙中的规律应该是简单而美的”。
60.二、几个基本原则及前提。
61.2.1我们先来确定几个基本原则。作为科研人员,大体对以下这几个原则没有异议:
62.原则0:脑是自然演化的结果。就是说,没有非自然力量的设计或干预,不会去考虑任何非自然因素,一切都是漫长进化和演变的结果。这是最基本的原则。
63.原则1:生物的进化是不可逆的。(不可逆律)。进化树上沿着树干到分支的进化是不可逆的。
64.原则2:神经系统是与机体的进化相适应的。(相关律)。至于两者是谁适应谁,申请人的观点是先有机体的变化,然后才是神经系统的适应性改变,神经系统是适应和服务机体的。但不管到底谁适应谁,它们两者是互相适应的。
65.原则3:根据原则1和原则2可以得到推论:神经系统的演变也是不可逆的。神经系统在演变过程产生的新构造,是从老构造的基础上演变产生的,并跟机体的进化相对应。
66.2.2我们还需要确认几个前提。我们需要假设以下这些前提是可信任的:
67.前提1:目前生物学方面对生物进化过程,包括从肠腔动物到节肢动物到鱼类到爬行动物到哺乳动物和人这一进化路径的研究结果和描述是基本正确的,至少相关教材是基本正确的。我们不排除现有研究可能存在某些细节性的不完善甚至错误,这可以不管,但从进化树的基干到哺乳动物和人,其各主要阶段的描述是正确且充分的,不会也不应该存在某条不为人知的从腔肠动物独立向人的进化途径。
68.前提2:目前神经生物学方面对于神经元、突触、神经递质及受体的工作的研究结果和描述是基本正确的,至少相关教材是基本正确的。我们可以不管分子层面以下怎么工作,只需确定神经元和突触怎么样工作,轴突末梢释放哪些神经递质,这些神经递质通过什么受体进入突触前和突触后,引起什么变化。这相当于确认电阻、电容、晶体管等这些元件的工作特性,并以此为基础来分析电路的工作原理,而无需涉及这些元件在微观层面的物理运动包括量子力学方面的机制。
69.前提3:目前神经解剖学方面对于中枢神经的主要核团之间的神经联系的研究结果和描述是基本正确的,至少相关教材是基本正确的。事实上目前对中枢神经的解剖资料也很难保证就都是准确和正确的,在一些细节,不同文献或教材都存在描述的差异甚至矛盾。但从大的方面看,目前对中枢神经的解剖结构和神经联系是基本正确的。这相当于确认所观察到的各个电子元件及其连接关系的描述是正确的。
70.前提4:确认目前对于脑功能的研究结果和描述是基本正确的,至少相关教材是基本正确的。主要包括:中枢神经实现了对各种感觉器官的感觉信息的接收和处理,中枢神经的运动输出实现了对机体运动的控制,包括说话和写字这些特殊的运动,等等。
71.根据上述这几个原则和前提,我们从初等动物到人的进化进程,来分析神经系统如何从简单的网状神经系统演变到极度复杂的人脑,进而推演出当前阶段的人脑的系统框架,以及它大体上是怎样工作的。
72.三、脑的形成及进化。
73.在动物学教材和一些文献中,有从腔肠动物到哺乳动物的神经系统演变的内容,但只是描述其可观察的形态上的进化结果,而我们更需要的是分析和描述在这个过程中,神经系统的具体构成及其连接关系是如何跟随机体的进化而进行相适应的演变,其信号处理的框架及流程又发生了怎样的变化。
74.按照原则1和原则2,中枢神经系统的形成和进化是跟随生物体的进化过程而相应变化的,它是动物处理各种外界信息及机体内部信息、并对自身各种活动进行控制的神经
组织的集合。那么我们按照图5的生物进化树,来对几个关键阶段进行分析。(需要注意的是生物进化是在不断进行的,某个进化分支的末端和始端,比如高等爬虫类和原始爬虫类会有很大差别,所以这里只能是选择某个阶段的一些有代表性的重要特征来进行分析。而且,重点不是在考究什么时期出现演变,而是分析神经系统在怎么样演变)。
75.3.1第一阶段:从简单细胞到节肢动物。
76.这阶段的中枢神经都很简单,但既然是描述中枢神经的演化过程,就不得不提及,而且它们是中枢神经的初始结构,未来高度复杂的人脑也是以此为基础逐步演变形成的。
77.3.1.1当生命从一些分子团进化为单细胞生物,这时没有也无需反射系统的,有些可以通过一些神经肽对外界的刺激产生应激反应。
78.简单的多细胞动物比如海绵,实质是一群细胞在协同完成吞噬和消化功能,顶多就是细胞之间存在相互影响。(虽然有文献称海绵已出现神经细胞,但未能阐述这些神经细胞的工作及功能)。
79.3.1.2腔肠动物为二胚层结构,有了明确的细胞分化,一些细胞分别分化为感觉细胞和肌细胞,二者之间出现神经节细胞,感觉细胞的冲动信号通过神经细胞传导到肌细胞引起肌细胞动作,带动身体运动,如图6的左侧。但感觉信号在神经细胞之间的传递是幅散传导、没有方向性的,其结果是一旦出现感觉细胞被刺激,则终端的肌细胞一起产生同步动作,一起收缩或一起放松,“牵一发而动全身”,所以其神经系统如同简单的开关控制电路,输入信号直接控制负载动作,其信息处理等效于图6的右侧。
80.3.1.3线形动物和环节动物具有三胚层结构,其感觉细胞包括有触觉、光觉和化学觉(嗅觉),中胚层形成肌肉结构,感觉细胞和肌肉细胞之间存在神经细胞形成神经反射弧,通过感觉细胞的信号来控制肌肉结构以产生定向运动。
81.由于存在触觉、光觉和嗅觉,这些不同感觉信息对机体运动的控制可能会存在矛盾,比如嗅觉吸引机体向前,而触觉需要机体退避。但在同一时刻你不能既前进又后退,所以,作为一种适应(原则3),在处理不同感觉信号的神经细胞之间,进化产生一些中间神经细胞交叉投射构成简单的神经网络,如图7左侧的中间网络,通过神经细胞之间的自我抑制和互相抑制,对不同感觉信号进行协调和控制,决定以哪一种感觉传入为主来控制肌肉运动,这实质上也是最原始最简单的一种“注意”控制和决策机制。(请注意图7只是一种用来说明投射关系的简化示意图,显然动物的神经投射比这复杂多了,存在多个感觉传入通道和运动控制通道并进行交叉投射)。
82.另外,既然机体能够产生定向运动,那么就需要至少两组肌肉在进行不同步的动作,所以,作为适应,也就需要有多组不同的神经反射弧来控制不同的肌肉,而且需要在不同反射弧之间产生一些中间神经细胞,通过交叉投射构成简单的神经网络,如图7右边的中间网络,来协调控制不同肌肉进行不同步的动作,比如一组收缩另一组放松,使身体产生定向运动。
83.在环节动物比如蚯蚓,这些简单的神经网络聚集在头部,形成了简单的神经中枢。这些交叉投射的神经网络将是人脑的初始形态,在后面将会看到,高度复杂的人脑正是以此为基础进行演变的。在未来的进化中,作为最古老的结构,图7左侧中间的交叉神经网络将最终形成人脑脑干网状结构中跟感觉和思维控制相关的核团,比如内侧区的巨细胞网状核,图7右侧中间的交叉神经网络将最终形成脑干网状结构中跟运动协调控制相关的核团,
比如外侧区的脚桥被盖网状核。
84.上面描述的神经反射弧及其交叉投射网络,用于对机体外部信息进行感知和整合处理,并输出运动信号去控制肌肉动作来使机体产生反应,形成一个外部信息处理系统。如图8的信息处理方框图。
85.另外,由于线形动物和环节动物分化出消化、排泄、生殖系统,所以相应的也会形成另一类神经反射弧,用于对机体各内脏系统的内部信息进行感知和整合处理,并输出信号去调节这些内脏系统的工作,形成一个机体内部信息处理系统,其结构和工作跟外部信息处理系统是类似的。(为叙述和条理清晰,以下主要叙述外部信息处理系统的演变和工作)。
86.3.1.4节肢动物开始复杂起来了。由于在陆地广阔的范围进行生活和活动,需要及时感知多样多变的复杂环境并及时做出更快捷有效的反应,其身体进化形成头、胸、腹和附肢,具有触觉、味觉、嗅觉、听觉、视觉和平衡觉等感觉器官来感受各种外部刺激,也能够通过多条附肢的协调控制来使身体产生灵活的有明确目的的运动,机体内部也具有呼吸、循环、消化、生殖、排泄等内脏系统及内分泌系统,所以,作为适应,其神经系统也出现较大变化。
87.节肢动物的感觉器官进一步进化,能够感受更丰富更细节的感觉信息。比如嗅觉器官能够感受不同嗅觉信息,眼不止能感受光线强弱,而且能够感受形状及空间运动,于是其神经系统也进行相适应的演变,图7中感觉传入节点的神经元数量明显增加,这些神经元集中在一起形成神经核团,如图9左侧的感觉核,里面具有多组神经元进行互相投射,其功能是接收感觉信息并进行整合处理。(这种结构还没有像人脑的神经核团那样边界清晰,也许还不能称为核团,但又找不到更合适的词,就还是称为核团)。对应的,图7左侧中间,那些与不同感觉传入的神经细胞进行交叉投射、对各种感觉信息进行协调控制的交叉神经网络也相应增大,形成一个对感觉核的神经元活动进行协调控制的网状结构的核团,如图9左侧中间的网状核,网状核内部的神经投射类似于图7左侧中间的神经网络。
88.感觉核和底层的网状核构成原始的感觉神经中枢。感觉信息传入后会同时传递到感觉核和网状核,其中网状核进行信息的粗和大方面的处理,尤其是进行不同感觉信息的竞争和协调,而感觉核在网状核的协调控制下对具体的细节性的感觉信息进行整合、判断和输出。
89.同时,由于机体出现更多的结构和肌肉来实现更灵活的身体运动,图7中的运动输出神经元数量也相应增加,并形成多条对不同肌肉进行运动控制的神经输出通路,这些运动神经元集中在一起形成另外一些核团,如图9右侧的运动核,里面也具有多组神经元进行互相投射,其功能是对运动信息进行整合处理并输出运动控制信号。同样的,图7右侧的中间,那些与运动神经元进行交叉投射、对各种运动信息进行协调控制的中间神经也相应增大,形成一个对运动核的神经元活动进行协调控制的网状结构的核团,如图9右侧中间的网状核,其内部的神经投射结构类似于图7的右侧中间的神经网络。
90.运动核和底层的网状核团构成原始的运动神经中枢。其中网状核团进行运动信息的粗和大方面的控制,包括在时间上协调决策那些肌肉启动(或停止)收缩(或放松)动作,而运动核在网状核的协调控制下对进行具体运动信息尤其是空间信息的整合和输出。
91.另外,在图9中,左侧的感觉核向右侧的运动核进行交叉投射,构成一个简单的中
间神经网络,根据感觉核输出的各种信息,整合形成控制信号并向运动核进行输出。在未来的进化中,这些中间神经网络将率先演变为皮质结构,比如鱼脑的嗅叶的旧皮质。
92.感觉核、运动核和网状核集中在头部构成脑,其中感觉核因为跟头部前方的嗅觉器官和视觉器官联系密切而处在上层(前部)的位置形成前脑,而在进化最早期形成的网状核处在底层的位置形成中脑。为了表示这种上层和底层的位置关系,将图9画为另一种表示方式,如图10。在未来的进化中,这些感觉核将演变成为丘脑上跟各种感觉信息处理相关的几个感觉核团,运动核将演变成为丘脑上跟运动信息处理相关的几个运动核团,而网状核将演变为脑干网状结构的几个核团。
93.这些神经核团及其神经投射构成新的一个对机体外部信息进行整合处理以及对机体运动进行协调控制的外部信息处理系统,其信息处理方框图如图11。用于对机体内部各内脏器官进行感觉和反馈调节的内部信息处理系统也会发生相应进化,脑干的自主神经系统初步成型,但神经元数量较少,这里从略。
94.3.2第二阶段:脊椎动物。动物进化的另一条途径是向鱼和爬行动物等脊椎动物进化。
95.3.2.1间脑的形成。在圆口类,脑的分化尚不明显,前脑、中脑、后脑还是连在一块。由于嗅觉在动物早期的活动中显得非常重要,所以处理嗅觉的神经细胞率先增多,成为一条较为独立的神经回路。但由于嗅觉信息无法进一步精细化,所以嗅觉相关的神经回路在后期的进化中反而没有太大的演变。相反,由于视觉和听觉信息能够不断精细和丰富,视觉信息从只涉及亮度变化进化到涉及亮度、颜色、形状、运动变化等,听觉信息从只涉及响度进化到涉及响度、频率和音色等,所以视觉和听觉的相关神经回路在后期出现巨大变化。在下来的内容中,将主要是以视觉和听觉来分析相关神经回路的演变情况。
96.进化到硬骨鱼类,随着视觉、触觉、听觉等感觉信息精细化和更加丰富,图10中的感觉核的神经细胞进一步分化,形成初级核和联络感觉核,演变为图12的左侧部分。(这里的初级核和联络感觉核都还是处在间脑的位置,相当于未来的人脑的丘脑膝状体和枕核这些,而跟感觉皮质初级区和联络区无关)。
97.这些核团的神经细胞数量增加,体积膨大,向前突出形成间脑(丘脑)。随着呼吸、循环、消化等内脏系统还有内分泌系统的进一步完善,对内脏器官和内分泌的工作进行反馈调节的自主神经系统也相应的发展,神经细胞数量增大,这些主要处于延脑,还有在间脑形成脑垂体和松果体等核团。而底层的网状核由于只是负责信息在类别和粗大方面的整合决策,神经元数量增加不明显,仍然停留在中脑的位置。
98.鱼的机体及脑还有一些更显著和重要的进化:
99.3.2.2灵活的机体运动控制和基底神经环路的形成。首先,鱼能够依靠鱼鳍(包括胸鳍、背鳍、腹鳍、尾鳍等)在立体的水空间里实现灵活多变又准确的各种游动,既能够让鱼稳定的静止停留在水中,也能够以不同速度游动、转向和减速制动,这需要通过肌肉对这些鱼鳍(还有鳔)进行精准的控制。
100.比如,鱼的胸鳍(相当于陆生动物的前肢)具有平衡、运动、控制运动速度和运动方向的多种机能:当鱼在水里静止不动时,胸鳍用于控制身体平衡使其不致摇摆不定;当鱼缓慢游动时,胸鳍起到类似船桨的划水作用;当鱼高速行进时,胸鳍收起紧贴鱼体减少阻力;而当胸鳍举起时,则可起到减速和制动作用;当胸鳍一侧贴近鱼体,另一侧举起,则起到协
助鱼体拐弯前进的作用。而且,这些运动动作都是能够控制其力度和幅度的,使得运动动作能够精准和圆滑进行。
101.单就一对胸鳍便具有如此复杂和精细的作用,那么,如何通过对胸鳍相关肌肉的动作控制,来实现对这些鱼鳍的运动(举起或收起)进行不同力度、不同幅度、不同角度的精准控制,显然这不止需要对运动进行空间位置的目标控制,还需要存在一个神经环路来对运动动作进行更精准的过程控制。
102.我们注意鱼脑在这一阶段的变化,一个是后脑出现一个凸起,这便是小脑的雏形,用于静止、平衡和肌肉微调控制的。另一个显著变化便是新出现的旧纹状体也即苍白球,苍白球与丘脑运动核之间构成一个“丘脑运动核
→
苍白球
→
丘脑运动核”的闭环控制环路,正是用于对鱼鳍相关肌肉的运动进行精准的闭环控制,包括对相配合工作的肌肉(类似主动肌和拮抗肌)的动作进行协调,对运动的幅度进行精准控制,在运动的末端进行精准的减速和制动。(部分文献指出鱼类出现的纹状体的功能是嗅感,申请人不认同这一观点,鱼已经存在嗅脑和简单的端脑在处理嗅觉)。这个神经环路是在图10的基础上,与丘脑运动核配合形成的,如图12的右侧部分。该环路也即是未来的人脑基底节中由苍白球和纹状体等构成的进行运动控制的基底运动环路,所以这里直接使用这个名称。基底运动环路包含有多条神经通路,每条神经通路对应控制一条肌肉的运动,其输入端接收丘脑运动核输出的运动信息,经过苍白球的整合后,重新输出到丘脑运动核,对其神经活动进行协调控制,从而实现对运动进行精细的过程控制,也使得多条肌肉能够协调动作,协同控制各条鱼鳍的精准运动。在鱼,由于运动信息较少,基底运动环路以苍白球为主,纹状体还不够显著,传统上也有把苍白球称为旧纹状体。(在传统教材中,人脑基底核不止包括控制运动的基底神经环路,还包括其他一些其他功能的核团)。
103.3.2.3能够对多种感觉信息进行整合和记忆,海马环路的形成。对于鱼,不止感觉信息和机体运动更加丰富,如何对各种感觉信息与运动输出之间的关系进行平衡、判别和决策也变得更复杂。比如,鱼能够依赖嗅觉识别各种食物、环境和危险(天敌),也能够依靠视觉来识别不同食物、环境和危险,尤其是活动的食物和天敌。这些不同的感觉信息对于鱼的生存具有不同的重要性,如何进行整合、判别和决策,已经无法依赖原有简单且固定的底层网状核来实现。
104.而且,鱼的生活环境变得更复杂,鱼的寿命也变得更长,这都使得鱼的一生所碰到的食物和天敌(危险)等信息变得更复杂和多样化,鱼已经无法依赖先天形成的固定的神经投射关系来对不同感觉信息进行判别和决策,而需要在后天通过经历来学习和记忆这些重要的经验。(鱼在觅食、回避危险、识别环境方面都存在长时记忆,在养殖行业中定时投食也能够被鱼所记忆)。
105.所以,鱼的神经系统出现相应的演变来适应上述这些需要。如图12的中间部分,一个演变是嗅球及丘脑感觉核与丘脑的运动核之间交叉投射的那部分中间神经网络的神经元数量增大,集中形成一个比较复杂的神经网络结构,其输入端接受嗅觉和其他感觉信息,其输出投射到右侧的运动核,起到在感觉输入和运动输出之间进行比较、判别和决策的简单“思维”的作用。在鱼,这一中间网络结构相当于嗅叶,是构成“嗅脑”的一部分,并与纹状体同时构成了鱼的端脑。这一中间网络结构的神经细胞开始在端脑外部形成一层薄薄的简单的皮质,也即“旧皮质”。在未来的进一步演变中,它对应于人类的“内嗅区”。另一个演变
是形成对中间信息进行整合和形成长时记忆的中间神经环路,这一神经环路的输入端连接感觉核和内嗅区的输出,接受已经被整合的各种中间信息,经过一些交叉投射且具有长时记忆效应(长时程突触可塑性)的神经网络的整合后,重新投射到中间核团也即是端脑,这一新进化的具有长时记忆功能的中间神经环路便是海马环路。海马环路也与底层的网状结构存在往返互相投射,在未来的进化中,与海马环路互相投射的底层结构也即是人脑基底前脑的隔核和斜角带核,它们控制着海马环路的工作,尤其是控制着海马环路形成长时记忆的工作。
106.在这一阶段,海马环路的作用是对已经被识别和整合的中间信息的空间(位置)关系和时间(时序)关系进行再整合并形成记忆,再重新投射回到端脑,通过将这些被海马环路整合和记忆的信息再整合到端脑的神经网络中,影响其在后的判别和决策。端脑和海马环路互相配合,更完善的实现感觉信息与运动输出之间的整合、判别和决策过程。其工作分工和工作过程如下:当多种感觉信息从感觉核传递到端脑的神经网络时,先是由端脑根据之前形成的联结关系进行判别决策,并向后面的运动核输出运动信息。比如,新出现具有某种新嗅觉和新形状的小物体,端脑的神经网络输出动作信息进行捕食。但这种捕食的动作输出可能是正确的,(食物,吞下),也可能是错误的,(不是食物,吐出),如果相同情况被重复几次,则这些感觉信息(气味、形状)及其运动输出信息(可吞食或不可吞食)的关系会通过海马环路进行整合和记忆,并将这些新形成的信息返回整合到中间神经网络上。那么在此之后,当该感觉信息(气味、形状)再次出现、从感觉核投射到端脑时,便可直接进行判别,做出吞食或不吞食的合理决策。
107.至此,对于处理感觉信息和机体运动的外部信息处理系统,鱼脑形成如图12的神经投射关系,其等效于图13的信息处理架构。
108.由于鱼所需整合和记忆的中间信息很少,所以海马环路不发达,只有少量的神经细胞构成,但这仍然使鱼具备了整合各种中间信息及形成长时记忆的功能。对于鱼,嗅觉是最重要的感觉,所以鱼脑中与嗅觉处理的神经细胞占了很大一部分,这导致从解剖上看到的,鱼的端脑主要就是嗅脑。但实际上,嗅脑的次级中枢,也就是未来的人脑中被称为“内嗅区”的皮质区,并非只是处理嗅觉信息,而是连接并与海马体配合工作,整合各种次级感觉信息及中间信息并形成长时记忆。所以鱼的嗅脑的次级皮质区实际上是鱼对各种信息进行整合、判别和决策的大脑,只是由于嗅觉信息占了极大比例而其他感觉信息占比很少,所以被视为“嗅脑”。在未来的进化中,随着视觉、听觉等其他感觉信息的极度丰富和复杂,相关核团的体积显著膨大成为丘脑和大脑皮质中重要的结构,而嗅觉由于信息复杂程度没有明显增加,嗅觉相关传导路没有增大,反而被包围在大脑内部成为不起眼的结构。而内嗅区和海马体,由于各种感觉信息和中间信息的丰富,体积也明显增大,成为边缘系统的主要结构。
109.3.2.4脑内多向调节神经回路(包括未来人脑的情绪神经中枢)的形成。
110.另一方面,当生物进化到鱼这一阶段,由于机体对复杂环境进行各种反应的需要,机体的内部信息处理系统与外部信息处理系统彼此之间需要存在互相联系和互相影响。比如当身体为快速捕食或快速逃离危险时,需要机体内脏器官提供更多的氧气和营养,所以这时需要通过自主神经系统对呼吸和循环系统进行调节;而当机体感到内部营养储备不足(感觉饥饿)时,需要反射到决策和运动神经,做出降低活动强度或进行觅食的行动。
111.于是相应的进化形成这样一个在外部信息处理系统和内部信息处理系统之间进行交叉投射和互相调制的神经回路。其一方面接收机体外部信息处理系统的信号传入,(包括处理嗅觉视觉听觉等信号的各种感觉核团、和对中间信息进行整合处理的各种中间核团的信号传入),另一方面接收机体内部信息处理系统的信号传入,(包括处理内脏和内分泌等机体内部信号的神经核团、和处理这些信号的相关中间核团的信号传入),经过一个神经回路的整合和反射后,其输出端又反过来向各个神经核团进行投射,调制它们的活动。其神经投射关系如图14,所等效的信息处理方框图如图15。
112.这一神经回路的生物学意义是对脑内各种神经回路的工作进行多向调节,使各种神经活动处在合适和协调的状态,尤其是协调内外两个信息处理系统的神经活动,使机体的内脏和内分泌系统的工作状态,与机体当前所处的环境或者机体所处的活动状态能够更好的互相适应,以便更有效进行捕食或避险等活动,从而有利于动物的生存。(比如:鱼受惊吓后能够更快逃离危险现场)。在鱼这一阶段,由于这个神经回路所处理的信息较少,其中间神经网络的神经元也不多,所以还没有形成分界明显的神经核团。在未来的进化中,这一神经回路所处理的信息会更加复杂,其神经元数量也会增加,形成了分界明显的以杏仁体为主的几个核团,并产生一些有趣的功能和表现,最终形成了包括人脑负责情绪和动机的功能复杂的情绪中枢。(鱼的这一神经回路的调制功能是已经存在,但其神经结构还不显著,无法描述其具体神经投射关系,所以在这里先提一下,具体结构及环路在后面再进一步描述)。
113.旧纹状体、海马环路、还有脑内多向调节神经回路这些是在丘脑的基础上形成的,而丘脑的后方又是中脑、后脑和脊髓,所以这些新结构只能往丘脑前方挤出,与嗅脑一起形成早期的端脑。在未来的进化中,由于大脑皮质的体积高度增大,基底核、嗅脑和海马环路这些结构反被包围在大脑半球的中间。
114.3.2.5爬虫类动物的进化。爬虫类的神经系统在鱼的基础上进一步演变。但从鱼到两栖动物到爬行动物,感觉器官和机体运动有一定发展,而智力没有显著的差别,所以其脑的变化没有那么显著,主要是处理感觉信息和运动信息的神经细胞数量增加导致的变化。
115.间脑及端脑的变化。到爬行动物这阶段,感觉器官尤其是视觉器官和听觉器官进一步精细化,能够接收和感知更多更细节的感觉信息。比如对于视觉,除了强度还能够接收不同颜色、形状和空间的信息,通过对这些不同物理性质的视觉信息进行整合,来识别不同物体或环境。所以,随着感觉的精细化,感觉信息变得更加丰富,丘脑的感觉核神经细胞数量迅速增加,体积显著膨大,而且在丘脑的初级感觉核与联络感觉核之间出现更多交叉投射的中间神经细胞,以便整合、识别和记忆这些不同性质且更精细的感觉信息,这些中间神经细胞向外延伸挤出,形成一层凸出在丘脑前方的较薄的皮质,这就是早期的端脑感觉皮质。如图16的左侧。感觉皮质与丘脑感觉核存在双向投射,皮质内部也存在交叉投射和局部神经回路,还接受来自底层核团的调制,所以进化形成更复杂的多层皮质结构。
116.由于丘脑初级感觉核与联络感觉核之间互相联系的神经细胞被挤出形成感觉皮质,所以在丘脑这里,初级感觉核和联络感觉核之间逐渐失去神经联系,最终形成彼此分离的不同核团,初级感觉核变成了感觉传入至感觉皮质之间的一个中继核团。比如对于视觉,丘脑的初级感觉核最终形成人脑视觉传导路的中继核也即外侧膝状体,而次级感觉核形成丘脑的枕核。(把外侧膝状体称为中继核似乎不妥,它不只是传导中继,而是在进行视觉信
息的初级处理)。
117.同时,爬行动物因为四肢和关节的出现,相应的控制四肢运动的肌肉数量增加,机体运动的灵活度和复杂度也增加,这相应的导致运动信息的复杂化和精细化,也使丘脑运动核的神经细胞数量增大,且与前后核团之间出现很多交叉投射的中间神经细胞来整合运动信息。这些中间神经细胞也是向外延伸挤出,凸出在丘脑前方,形成早期的端脑运动皮质。如图16的右侧。跟感觉皮质一样的原因,运动皮质也是形成多层结构。于是,原有由丘脑的运动核向脑干的运动输出核输出运动信号的神经投射便逐渐改为由运动皮质向运动输出核输出运动信号的投射。
118.感觉皮质和运动皮质都是从丘脑向外挤出,一起凸出在丘脑外面,并与丘脑分离,构成一层简单的薄薄的端脑皮质。这些感觉皮质和运动皮质也与旧皮质(主要是嗅叶皮质)产生神经联系,并一起构成爬行类动物的端脑。在未来向哺乳动物和人的进化中,感觉皮质和运动皮质会进一步分化,它们与旧皮质之间的神经联系(图16画了两个圈圈的地方)也会进一步增多和复杂,并形成更多的端脑新皮质区,比如感觉联络区、运动前区和联合皮质等。
119.基底环路及小脑的变化。在鱼便形成的对运动过程进行精细控制的基底环路,其环路本身的结构和功能没有变化,但由于从丘脑的次级运动核挤出形成运动皮质,那么基底环路原来接受丘脑运动核输出信息的输入端口也就变为更多的是从运动皮质接受运动输出信息。如图16的右侧。而且由于运动信息的丰富,这一输入端口的神经元数量增加,体积增大,与苍白球分离,成为一个更明显的核团,也即新纹状体,包括尾状核和壳核。另外,由于维持机体运动状态及肌肉微调的信息增加,小脑的神经元数量也相应增加,体积进一步膨大。(小脑回路的内容略)。
120.海马环路的变化。爬行类动物和鱼比较在智力上没有产生代际的显著变化,嗅叶和海马回路也没有显著的变化,只是由于所需处理的中间信息存在一定程度的增加,嗅叶及海马环路的神经元数量增加,体积会有一些增大。但相对迅速增大的丘脑和基底核来说,海马环路由于体积增大不明显反而逐渐被包围在中间,并在未来的进化中逐渐被挤压在大脑半球的内侧面,形成海马结构。
121.自主神经中枢方面。为了与脱离水环境相适应,爬行动物使用肺呼吸,生殖系统、排泄系统(肾功能)都得到加强,等等,这方面都会促进自主神经中枢也进行相适应的演化,交感神经和副交感神经更加发达。但总的说,这一阶段没有产生代际变化,其架构基本不变。
122.3.2.6鸟类的进化。鸟类在中枢神经的显著变化是自主神经中枢和纹状体的发展。
123.自主神经中枢进一步完善。鸟类的活动范围更大,面对的环境变化更复杂,所以鸟类开始保持体温恒定,双呼吸和双循环,其内脏系统和内分泌系统更加复杂和功能强大,对内脏和内分泌的感觉传入和反馈调节更加细化。相应的促使鸟类的自主神经系统得到明显增强,处理内脏和免疫内分泌信息的神经细胞数量明显增加,成为分界较明显的上丘脑和下丘脑(丘脑下部)。其自主神经中枢的神经投射示意图如图17,(省略与其他神经中枢的联系)。由于鸟脑的体积较小,自主神经中枢的传入核团和输出核团在解剖上大多还不够明显,但这些核团的神经细胞和功能是存在的,并在哺乳动物和人脑中进一步增大和显著。
124.视觉和纹状体高度发达。鸟类在空间飞行,这需要其视觉高度发达,机体运动也高
度灵活和精细化。相适应的,鸟脑的视叶和丘脑发达,出现视丘,纹状体和小脑高度发达,以便与复杂运动相适应。另一方面,鸟的叫声很丰富,这也促进其纹状体高度发达。(动物的叫声其实质是一种特殊的运动输出)。
125.3.2.7中枢神经至此所完成的演变结果。至此,动物进化到脊椎动物的高级爬虫类和鸟类,中枢神经进行相适应的演变,形成如图18所示的神经投射关系。(图中省略“脑内多向调节神经回路”与其他核团的投射关系,具体可参考图14)。其等效的信息处理系统框架与图15基本相同。
126.可以看到,动物的神经系统进化到鸟脑,对机体内部和外部信息的接收反应和整合处理已经很全面了,也产生了对两者进行相互调节的神经回路,形成一个比较完善的系统架构。在未来,我们的人脑正是以这个系统架构为核心,通过进一步优化完善而形成的。
127.对于鱼和爬虫类这一阶段的脑进化,有些教材或文献会有“丘脑开始有一些纤维向上投射到端脑皮质”这样的表述。申请人觉得这种表述不恰当,在这个阶段之前本来就还没有端脑皮质,何来开始有神经纤维投射到端脑皮质?不是“丘脑开始有一些神经纤维向上投射到端脑皮质”,而是“丘脑的一些神经元数量显著增加,体积膨大被挤出,向上凸出形成了端脑皮质”。
128.3.3第三阶段:哺乳动物和人。进化到哺乳动物,生物出现较大的代际变化,机体的机能和活动能力都有显著提高,脑也快速进行相适应的变化。最突出的是端脑高度发达,成为体积最大的结构,而丘脑、下丘脑、纹状体、海马体这些结构相对的体积变化没有那么大,成为被端脑包围的皮质下核团。
129.3.3.1感觉中枢的变化。随着感觉器官的精致,各种感觉信息变得更加精细和丰富,信息量剧增,于是各种感觉传入通路在丘脑这里的交叉投射的中间神经元剧增,丘脑感觉核团和感觉皮质进一步分化,这些中间神经元被挤出到端脑外表面形成新皮质,并分化为初级感觉区和次级感觉区,连接丘脑原有核团和新皮质的轴突便形成神经投射纤维也即丘脑与皮质之间的白质。但各感觉传入与脑干网状结构、以及脑干网状结构与丘脑之间,除了会有一定数量的增加,其连接方式和工作过程基本不变。
130.比如视觉传入路,虽然处理视觉的基本要素(颜色、线条、运动方向等)并没有多大变化,但由于视觉信息的精细化导致视觉信息量剧增,(对于人脑,尤其是文字的出现),所以在丘脑这里,对视觉进行初级处理的核团包括外侧膝状体(lgb)和枕核(p)的神经元和体积变化不大,但它们之间相互投射的对视觉具体信息(物体、画面、文字等)进行处理的中间神经元数量剧增,于是这部分神经元剧烈膨大挤出构成端脑的视觉皮质,其中外侧膝状体与枕核之间的中间神经元挤出形成端脑的初级视皮质,枕核与板内核之间的中间神经元向上挤出形成端脑的联络视区,连接它们之间的轴突便构成丘脑外侧膝状体与端脑初级视区之间的视觉传递路,还有丘脑枕核与联络视区之间的联络纤维。其视觉信号相关的神经投射如图19。(对于人脑,联络视皮质还分化出专门用于文字阅读的皮质分区也即“阅读中枢”,运动皮质还分化出专门用于书写的皮质分区也即“书写中枢”)。
131.同样的在听觉传入通路,虽然处理声音的基本要素(频率、响度、相位等)没有多大变化,但由于听觉信息的精细化导致视觉信息量剧增,(对于人脑,尤其是语言的出现),所以在丘脑这里,对听觉进行初级处理的核团包括内侧膝状体(mgb)与枕核或丘脑后区(不确定)的神经元和体积变化不大,但对听觉具体信息进行处理的神经元数量剧增,于是这部分
神经元剧烈膨大挤出构成端脑的听觉皮质,(包括初级听觉区和联络区),连接它们之间的轴突便构成丘脑内侧膝状体与初级视皮质之间的视觉传递路,还有丘脑枕核或丘脑后区与听觉联络区之间的联络纤维。其听觉信号相关的神经投射如图20。(对于人脑,端脑皮质还分化出专门用于语言处理的皮质分区也即“听话中枢”和“说话中枢”)。
132.其他感觉信息的处理具有同样的演变。比如躯体感觉,处理感觉初级信息的丘脑腹后核(vp)体积没有显著增长,但处理躯体感觉具体信息的神经元数量剧增而膨大挤出构成端脑皮质的躯体感觉区,连接它们之间的轴突便构成丘脑腹后核与端脑躯体感觉区之间的联系纤维。但显然,由于人的味觉、嗅觉、温度觉等感觉并不精细和敏感,所以这些感觉传导路的相关核团和皮质区域并不显著。比如,由于人类的嗅觉没有演变得更灵敏精细(甚至可能变差),所以用于处理嗅觉具体信息的皮质神经元,仍停留在内嗅区而没有膨大凸出到大脑新皮质。(内嗅区仍然是端脑新皮质通过扣带回连接海马环路的信息联络端口)。
133.3.3.2运动神经中枢的变化。从爬行动物到哺乳动物到人,骨骼肌数量并无显著增加,控制这些肌肉运动的神经环路数量没有大的变化,所以从锥体系运动输出端发射到机体各骨骼肌的运动神经纤维并没有显著变化,从纹状体到苍白球这部分基底神经环路的核团体积没有很大变化。不过在进化过程中,四肢或其他部位(比如人用于控制发声的喉头肌肉群,还有特殊的比如大象的鼻子)可进行的动作变得非常复杂和精细,对应的运动信息(尤其是精细控制运动空间位置的信息)急剧增加,所需的神经元数量也就急剧增加,所以从丘脑运动核团的腹前核(va)和腹外侧核(vl)连接锥体系输出端的神经元、还有锥体系连接基底环路端口的神经元数量急剧增加,于是这部分神经元剧烈膨大挤出构成端脑的运动皮质,(包括运动前区和运动区),连接它们之间的轴突便构成丘脑腹前核和腹外侧核与运动皮质、还有基底神经环路与运动皮质之间的联系纤维。基底环路的核团分化也更加清晰,形成纹状体、苍白球、黑质和底丘脑这些独立的核团,但对比大脑皮质的急剧增大,基底环路这些核团的神经元数量和体积变化相对不大,于是被包围在大脑半球中间成为基底核。演变后的运动神经中枢的神经投射如图21。
134.另外,由于肌肉运动的高度精细化,用于协调控制肌肉精细运动和平衡的另一个神经环路及其核团的集合也即小脑,其神经元数量也急剧增长,不过由于肌肉数量没有显著增长,控制这些肌肉所需的神经环路没有显著增长,所以小脑与大脑相连接的神经纤维数量也就没有显著增长,于是演变的结果就是:小脑的神经元数量及体积急剧增长,但小脑与大脑的连接端口没有显著增大,导致小脑膨大鼓出成为一个相对大脑独立存在的组件。更典型的比如大象的小脑神经元及体积增长远大于大脑,这可能跟它用于控制鼻子的精细动作和空间平衡有关。
135.3.3.3端脑新皮质及海马环路的变化。
136.联结在感觉中枢与运动中枢之间的神经结构包括两部分:一个是直接联结在感觉皮质与运动皮质之间的中间神经网络,也即爬行类动物的旧皮质,对感觉信息与运动输出进行整合、判别和决策,另一个是海马环路,对各种中间信息进行再整合处理,包括将信息通过联结构成新的信息和实现这些信息的储存记忆,再将这些信息整合到中间网络中影响其下一次判别和决策。在哺乳动物,随着感觉信息和运动信息的急剧增加,需要整合处理的这些中间信息也暴增,于是旧皮质中的部分中间神经元数量暴增,体积急剧膨大,向外凸出成为端脑新皮质(联合皮质)。联合皮质仍然与感觉皮质和运动皮质之间进行互相联络,它
们与丘脑之间相互连接的轴突便形成神经投射纤维,也即连接丘脑(尤其是丘脑板内核群)与联合皮质之间的白质。对于海马环路,其输入输出端口、也即与端脑各个皮质区相互联络的那部分神经元数量显著增加形成内嗅区和扣带回,而由于该环路的核心海马体只是对中间信息进行再整合,神经元数量和核团体积变化不大,反而被包围在中间并被挤压成为一个形状怪异的结构,整个海马环路也变成一个被大脑包围在中间的皮质下环路。海马环路与端脑新皮质、扣带回、内嗅区等核团的相关神经投射如图22。
137.但海马环路与联合皮质的功能及关系仍然不变:当感觉信息从感觉中枢传递到端脑联合皮质时,先是由联合皮质根据之前形成的联结关系进行判别、整合和决策,需要时可以向后面的运动中枢输出运动控制信息。而感觉皮质和联合皮质输出的中间信息会投射到海马环路,通过海马环路对这些信息进行再整合,依赖长时程突触可塑性(ltp和ldp)实现对信息的空间位置和时序关系的整合和长时程记忆(或者称海马记忆),然后这些经过再整合的信息会重新投射回到联合皮质,影响在后的判别和决策。而如果这些信息记忆被多次输入、刷新或使用,则联合皮质上的相关神经元被多次协同激活,会依赖突触重构来形成时效性更长和更稳定的皮质记忆,(也有称为永久记忆)。而一旦在联合皮质上形成更直接和更稳定的皮质记忆,则原有通过海马环路建立的中间通道便会由于失用而逐渐失效,其神经元会由于其他新信息的输入而建立起新的中间通道,用于对新信息进行整合和记忆。
138.3.3.4自主神经中枢的变化。
139.相对于爬行动物,哺乳动物的呼吸、消化和循环系统更加稳定,尤其是生殖系统出现巨大变化,而且对免疫内分泌和对体液内各种物质浓度的调节会更加精细和复杂,所以自主神经中枢进一步发达,上丘脑、下丘脑、以及孤束核、臂旁核等相关核团分化成为独立的核团。其自主神经中枢的神经投射大概如图23。(由于所掌握的自主神经中枢相关解剖资料可能不够详细,不确定图23是准确的)。总体而言,机体内部信息处理系统这部分的变化还是远远没有外部信息处理系统那么显著,所以自主神经中枢的神经元数量和核团体积也就增大不够显著,被体积剧增的端脑包围在底部和中间。而且由于机体内部的感觉信息没有进一步精细化,上丘脑、下丘脑这里用于整合处理内部信息的神经元没有更多的分化增加,所以也就没有出现神经元迅速增加以致被挤出形成新皮质的情况。
140.3.3.5脑内多向调节神经中枢的变化。
141.脑内多向调节神经中枢也就是人类跟情绪相关神经中枢。从鱼开始,进化形成一个神经环路来对机体内外部信息及运动的各个神经中枢进行多向调节,协调各个中枢的工作状态以配合机体各系统更好的工作。从爬行动物到哺乳动物到人,这一脑内调节神经中枢的投射环路基本没有变化,同样是接受感觉中枢、思维中枢、运动中枢、内脏神经中枢、免疫内分泌神经中枢的多种传入,经过整合后通过多条调节通路,反过来向前述这些神经中枢进行弥漫性的调节。
142.由于哺乳动物的机体内外部两个信息处理系统所处理的信息急剧增加,那么这个脑内多向调节神经中枢所接收处理和输出调节的信息也相应的显著增加,其进行信息接收整合处理的核心核团的神经元数量显著增加,体积膨大,形成具有多个亚核结构的杏仁体。而其输出调节也变得多样化,一方面由杏仁体直接向各个神经中枢和神经环路进行投射去直接调节它们的工作,另一方面杏仁体也与脑干几个调制性核团存在互相联系,这些调制性核团以da、5-ht、ne等单胺类神经递质工作,它们向大脑其他核团和区域进行广泛的投
射,对其他核团或环路进行各种不同效果的调制。其脑内多向调节神经中枢与其他神经回路的神经投射如图24。从人脑看,脑干这几个调制性神经核团的作用不局限于情绪相关,其功能是对脑内各个神经环路的工作进行多向的反馈和调节,所以申请人更倾向于把以杏仁体为主的情绪中枢和脑干这些调制性核团,还有下丘脑的部分调制性输出,一起归类为一个脑内多向调节中枢。
143.而且,在人,由于感觉信息和中间信息(尤其是语言文字这些特殊信息)的高度丰富,杏仁体的输入输出端与感觉中枢的感觉皮质、以及思维中枢的联合皮质相连接的这部分神经元数量剧增,这也形成了另外一块联合皮质(主要分布在前额叶)。而由于语言的产生,对这一内部调节所产生的多种不同的机体感受,人类称之为“情绪”并采用各种丰富的词汇来描述和记忆这些感受,这反过来可能促进了这一神经环路的演化,包括增加了这一神经环路输入输出端口的神经元数量,以及强化了这一神经环路的调节作用。语言和文字能够直接使人情绪产生很大波动,正是这一演化的结果。
144.通过上面第3.3.1至3.3.5的演变,将图19至图24结合起来,构成了人脑主要核团的神经投射关系的示意图,如图26。与其等效的信息处理系统框架如图27。(其中语言相关的皮质分区是人脑独有的,其他哺乳动物的没有这些相关分区或者规模极小)。从大的方面看,其等效于图25的系统框架。
145.这就是我们的人脑。如果忽略语言相关部分的差异,这也是高级哺乳动物的脑。
146.3.3.6人脑相比其他哺乳动物脑的特异性。按以上分析,人脑跟哺乳动物脑在整体架构上是基本一样的,但为什么两者在智力上存在显著差异?
147.从结构形态上看,人脑相比其他哺乳动物的脑具有更发达的端脑新皮质,尤其是更多的皮质联络区和联合皮质区,这使其具有更多的神经元来识别和处理机体外部信息,并能够将这些信息整合为更丰富的各种中间信息(比如语言和文字),还具有更强大的对这些中间信息进行再次整合处理的能力(尤其是思维能力),这是智力差异的生物学基础。那又是什么造成了这种差异?其中最关键的特异性在哪里?
148.既然神经系统的演变应该是跟机体进化相适应的,我们可观察到两者在机体及功能上存在两点显著的差异:1、人直立行走,前肢进化为手,能够进行更灵活的动作,2、人形成了语言,那么人脑高水平的智力会跟这两点相关。虽然有“劳动创造了人”的说法,但申请人认为语言的形成显然更关键。理由是前肢进化为双手,能够进行更灵活的操作,跟这一进化相关的更多是运动神经中枢,包括运动皮质、基底环路和小脑,尤其促进了运动皮质和小脑的发达,但对智力不存在显著的促进作用。一个例子,大象的鼻子也很灵活,也能够进行很多灵活操作,所以大象的小脑特别发达,并因此造成整个大象脑的神经元总量也极其庞大,但这并没有促进其形成更高水平的智力。
149.而语言和文字的形成,在人脑产生了更重要的变化:在感觉中枢(感觉联络区)上,有更多的神经元来实现对语言和文字的识别,这些神经元形成了端脑新皮质的更多的联络性区域,也即所谓的“听觉性语言中枢”和“视觉性语言中枢”;在运动中枢(运动前区)上,有更多神经元来实现对语言和文字的运动性输出,这些神经元形成了所谓的“运动性语言中枢(说话中枢)”和“书写中枢”;在思维中枢的联合皮质和海马环路上,也有更多神经元来实现对语言和文字相关信息的再整合,以记忆、整合、重组和输出语言和文字的相关信息。这些都在结构方面导致端脑皮质的显著增大。
150.在功能方面,正是这些丰富的语言词汇,使人类能够对自然界更多的事物和事件进行定义、识别、描述和记忆,形成更丰富更具体更精确的天量信息,而且信息可以在群体间进行学习、传承和积累。这些信息在人脑中被接收和识别、整合和重组、记忆和输出等等,产生了复杂的思维活动,最终导致人脑跟其他动物脑出现显著的智力差异。其他动物由于缺乏丰富的语言,能够建立和识别的基本的信息要素太少,它们无法进行更复杂的对信息进行再整合的高级智力活动。一个简单实验,我们可以自己给选出100个词作为信息的基本要素,然后试着只能使用这100个词来对所观察到的世界进行识别、描述、记忆和思考,便会发现这种状态下所表现的智力是很低的。而对于大多动物,它们的情况还不如这样子。
151.所以说语言的形成是造成人脑跟其他动物脑存在显著差异的最重要原因。当然,进一步往前面看,也可能正是因为人类长期的直立行走,使得气管通道和咽喉的生理结构产生变化,喉咙部的肌肉所能够进行的动作控制出现更高水平的精细化,并且形成了与运动中枢的直接神经连接,这才产生了语言这种需要高度精细控制的特殊运动输出。
152.至于促使人类语言形成的根本因素到底是喉咙结构先出现变异还是大脑先出现语言相关神经环路?这个目前尚无定论。申请人是这个观点:机体的进化在先,然后才有神经系统的适应性改变,神经系统是适应和服务机体的。而且,从解剖上看,其他哺乳动物比如猫狗的中枢神经,也是存在与发声相关的神经环路和皮质区域,只是相关神经元数量很少,所以只能发出简单的少量的不同叫声,而无法形成丰富的语言。这说明,喉咙部位的肌肉与运动中枢之间的发声相关神经环路是从哺乳动物就存在的,但只有人类的喉咙结构和肌肉群产生变化,能够进行更精细的运动控制,才能够发出更复杂和更丰富的不同发音,于是相关神经环路和皮质分区的神经元数量也进行适应性改变,促成人类语言的形成。
153.四、人脑的系统框架和工作机制简述。
154.4.1传统上有这样的观点:人脑是随着生物的进化而从低级到高级进化发展,当人脑进化出端脑等高级的新结构后,之前的老结构便处于从属地位,而端脑等新皮质便处于主导,去控制皮质下中枢的工作。更有观点将端脑皮质称为高级中枢,是进行思考、决策、智能等高级脑功能的结构,而将中脑以下结构甚至将皮质下结构都称为生命中枢,认为只是维持生命活动的结构。这些观点出现在各种文献甚至教材上,是传统的主流观点。
155.申请人强烈反对这些观点。认为这些观点是割裂了人脑各个结构之间在信号处理上的相互联系,实质上也是割裂生物进化各阶段之间的前后联系。既然生物的进化是连续的且不可逆的(原则1),大脑又是与机体的进化相适应的(原则2),那么,脑的进化自然也应该是连续且不可逆的,不管人脑的结构如何复杂,功能如何强大,它也是从低等动物的神经系统连续演变而来的(原则3)。
156.比如,在爬行虫阶段,脑对外部信息的整合处理仍然以间脑和嗅叶为主,但已经有部分联结它们的中间神经元数量明显增加,被挤出并向外凸出,成为一些薄薄的简单的端脑皮质,用于对间脑和嗅叶的信息进行再整合处理。而进入哺乳动物阶段,由于中间信息的丰富和复杂,用于整合处理中间信息的端脑皮质的神经元数量显著增加,体积迅速膨大,才发展成为硕大的端脑新皮质。端脑新皮质并非突兀出现,而是在原有基础上进一步发展而来的。那种把人脑简单的划分为“爬行脑”、“哺乳脑”和“皮质脑”的观点,是割裂人脑各个功能结构的相互联系,也是割裂它们在进化过程的前后联系,这相当于认为哺乳动物是在爬行动物基础上独立增加一些新结构,认为人是在一般哺乳动物的基础上独立增加一些新结
构,而没有正确认识到后者其实只是在前者的基础上进一步演变和完善,是不可分割。
157.当人脑进化形成新的结构(端脑新皮质)时,它既不是改变脑的原有系统框架,也不会改变跟老结构(皮质下核团)之间主导与从属的工作联系,新结构只是在原有框架基础上的延伸和扩展,看似庞大且神秘的端脑新皮质其实只是由于所处理的信息量急剧增加而导致丘脑中某些节点(核团)的神经元数量也急剧增加,这些神经元及其联系纤维向外膨大和扩展形成新皮质,并在脑颅有限的空间内由于凸出和挤压而形成端脑各种回及其褶皱的皮质结构,但它们实质上只是皮质下的脑干网状结构和丘脑的延伸,其工作仍然受脑干网状结构和丘脑的控制。脑的演变不会也不应存在“从低等动物进化到人时,端脑成为主导,去控制皮质下核团工作”的情况,否则原有的神经投射关系和信号处理流程会发生混乱甚至逆转,也导致在两个进化节点的中间过程会有互相矛盾的神经投射关系和信号处理流程同时存在的混乱情况,这既不应该存在也不符合进化原则。
158.4.2如图25,人脑的系统框架可视为两个大的信息处理系统和一个小的脑内多向调节系统。
159.一个是机体内部信息处理系统,也即自主神经系统,包括内脏调节神经中枢、化学和内分泌调节神经中枢。主要由孤束核、臂旁核、下丘脑、上丘脑等核团构成,用于接收来自机体内部各种传感器传入的机体内部信息,经过整合处理后通过交感神经和副交感神经输出,对各个内脏器官、血压、血糖、体温等等和免疫内分泌系统的工作进行调节,(部分内分泌方面的分泌工作是直接在上丘脑和下丘脑直接实现),这一内部信息的整合处理过程一般是不可知觉的且在无意识下进行的。
160.另一个是机体外部信息处理系统,包括感觉神经中枢、思维神经中枢和运动神经中枢。主要由脑干的几个网状核团、丘脑、海马环路、基底环路、小脑和端脑皮质等构成,由于接收来自视觉、听觉、触觉等各种感觉器官传入的机体外部信息,经过整合处理后输出对机体运动(包括说话和书写)进行控制,思维神经中枢对这一过程进行决策和控制,同时也完成对各种中间信息的再整合。
161.而边缘系统的杏仁体和脑干的中缝大核、中缝背核、蓝斑核、被盖腹侧区(vta)等若干调制性核团则构成一个小而重要的脑内多向调制系统,对内部信息处理系统和外部信息处理系统各个神经环路的工作进行相互调节,使彼此的工作状态能够协调和适应,并由此产生了情绪、欲望、动机、成瘾等等有趣而神秘的宏观表现,人类的情绪中枢正是这一调节系统的组成部分。
162.4.3能够被我们所感知和意识到的主要是图27上部的机体外部信息处理系统的活动,也是图25和图26的上部分,(一些机体内部感觉比如痛觉也能够被感知)。感觉中枢由各个感觉传入通路、脑干网状结构一部分、丘脑若干核团(外侧膝状体lgb、内侧膝状体mgb、腹后核vp、枕核p,等等)、和感觉皮质等构成,用于对感觉信息进行整合、识别、记忆等处理。运动中枢由肌梭感觉传入通路、脑干被盖网状核、丘脑的腹前核va、腹外侧核vl、基底运动环路、小脑环路、运动皮质和锥体输出通路等等构成,用于对运动信息进行反馈、整合和记忆,并输出去控制机体运动和说话书写等特殊运动。思维中枢由脑干网状结构另一部分、丘脑板内核群、部分丘脑网状核、联合皮质和海马环路等构成,其实质是连接感觉中枢与运动中枢之间的中间神经网络,用于对它们所处理的中间信息进行再整合处理,其神经活动形成了记忆、思维和意识等宏观表现。在没有外部信息传入或外部信息没有引起注意时,思维中
枢的联合皮质和海马环路能够与丘脑和脑干网状结构等构成往返投射环路,进行高节律的神经活动,对中间信息进行再整合,比如“在内心进行”的思考。
163.各种外部信息尤其是视觉、听觉、触觉信息通过感觉器官传入中枢神经后,一方面投射到脑干网状结构,进行大的粗的整体方面的竞争和协调处理,另一方面投射到丘脑中继核团,对信息的各种物理性质相关参数进行初步处理,然后再传递到“因为凸出和挤压而形成”的端脑新皮质。新皮质在脑干网状结构和丘脑的控制下,对信息各种具体参数进行更精细的处理,由感觉皮质完成具体内容的整合和识别处理,由联合皮质和海马环路完成对中间信息的重组,需要时再由运动皮质整合输出运动信息,最后由锥体输出进行运动控制(包括机体运动和说话写字),并由基底环路和小脑进行精细控制。
164.4.4不管端脑新皮质看起来有多么庞大、复杂和神秘,人脑的工作仍然是以最早形成的底层的脑干网状结构为控制核心,以中间层的间脑(丘脑、基底环路、边缘系统部分核团等)为联络节点,而处在表层的端脑皮质只是在脑干网状结构和间脑的控制下进行具体的信息处理。当然,脑干网状结构—丘脑—端脑皮质之间是双向投射的,脑干网状结构通过丘脑上行控制皮质的神经活动,反过来端脑皮质的神经活动也下行向丘脑和脑干网状结构进行反馈,影响它们的神经活动。而思维、智力、决策、注意力、意识这些,其实质是人脑在进行信息整合处理时所表现出来的宏观功能或现象,只是由于“人类使用自己定义的信息要素(语言)来描述信息的形成和处理过程”和“人类通过大脑对大脑的工作进行自我认识和理解”的原因,显得神秘和不好理解。
165.以一个作战体系来说明:“司令部”这种作战系统的核心和指挥中枢仍然是早期诞生的少数元老(脑干网状结构),它接收各种外部情报的简报,(各种感觉信息的非特异性传入),接收前沿阵地战况的简报,(皮质神经活动通过丘脑的下行反馈),经过协调后发出各种重要指令,(上行发放的激励和控制信号)。
166.丘脑则是司令部与前沿阵地之间的“联络所”,它负责将各种外部情报中转给前沿阵地,(感觉传入通过丘脑特异性中继核团向感觉皮质的投射),负责将司令部的指令分发到各前沿阵地并控制其执行,(脑干网状结构通过丘脑非特异性核团向皮质的上行激励),同时将前沿阵地反馈的战况汇总归纳为简报向司令部汇报,(丘脑尤其是丘脑网状核下行向脑干网状结构的反馈投射)。
167.而端脑皮质是“前沿战斗阵地”,虽然士兵众多战斗激烈(神经元数量巨大,神经活动高度活跃),但其实只是在联络所的控制下执行具体的战斗任务,(包括思维在内的各种信息处理活动),并将战斗情况反馈给联络所,(皮质下行向丘脑的反馈投射)。不能看到前沿战斗阵地士兵众多占地庞大战斗激烈就认为它占主导地位,认为是它在控制那些人员少地方小的机构。
168.理解这一点,将会有助于我们更好的理解人脑的整体框架及工作机制,尤其是认清端脑的神经活动是受皮质下核团的神经活动所控制的,不能割裂皮质与皮质下核团的关系去分析端脑皮质的工作,相反,应该由下而上、由整体至局部、由抽象至具体的从皮质下核团的投射关系和工作流程去分析皮质上的神经活动。这将能够更好的分析理解感觉中枢、思维中枢、运动中枢、自主神经中枢、情绪中枢及其各个神经环路的工作,有助于分析理解思维、意识、情绪和注意力控制的形成机制。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1