对黑茶藨子长管蚜生物型NR:1具有抗性的生菜植物的制作方法
对黑茶藨子长管蚜生物型nr:1具有抗性的生菜植物
1.序列表的并入
2.包含文件名“semb042usp1_st25.txt”的序列表的全部内容通过引用并入本文,其大小为41.2千字节(在中测量)并在 2019年9月25日创建并包含95个序列。
技术领域
3.本发明涉及用于产生对生菜蚜虫黑茶藨子长管蚜(nasonoviaribisnigri)生物型nr:1表现出增强的抗性的生菜植物的方法和组合物。
背景技术:
4.宿主植物的抗性是农业,特别是粮食作物生产领域中的重要性状。尽管在各种生菜物种中已经鉴定了赋予害虫抗性的基因座,但是由于缺乏与该基因座连锁的特异性标记,阻碍了将这些基因座引入栽培品系的努力。在植物育种中使用标记辅助选择(mas)使得基于与目标性状连锁的遗传标记来选择植物成为可能。然而,即使已经表征了与性状相关的基因,用于鉴定或追踪植物中的所需性状的准确标记也常常不可获得。这些困难由于以下因素而进一步复杂化:如多基因遗传或数量遗传、上位性以及对所需表型的表达的遗传背景的不完全了解。
技术实现要素:
5.一方面,本发明提供了一种良种莴苣(lactuca sativa)植物,其在染色体8上包含来自刺莴苣(lactuca serriola)的至少第一重组染色体区段,其中所述重组染色体区段包含相对于缺少所述重组染色体区段的植物赋予对黑茶藨子长管蚜生物型nr:1的抗性的等位基因。在一些实施方案中,所述第一重组染色体区段在染色体8上包含选自由标记基因座m1(seq id no:26)、标记基因座m2(seq id no:16)、标记基因座m4(seq id no:46)、标记基因座m5(seq id no:11)、标记基因座m7(seq id no:21)、标记基因座m8(seq id no:41)、标记基因座m10(seq id no:36)和标记基因座m11(seq id no:31) 组成的群组的标记基因座。在其他实施方案中,所述黑茶藨子长管蚜生物型nr:1抗性等位基因位于公共莴苣参考基因组lsat_salinas_v7 的染色体8上的106,984,777 bp与136,545,853 bp之间。在某些实施方案中,植物对于所述重组染色体区段是纯合的。
6.另外,本发明提供了良种莴苣植物的植物部分,其在染色体8上包含来自刺莴苣的至少第一重组染色体区段,其中所述重组染色体区段包含相对于缺少所述重组染色体区段的植物赋予对黑茶藨子长管蚜生物型nr:1的抗性的等位基因。在某些实施方案中,所述植物部分是细胞、种子、根、茎、叶、头、花或花粉。在进一步的实施方案中,本发明提供了良种莴苣植物的种子,其在染色体8上包含来自刺莴苣的至少第一重组染色体区段,其中所述重组染色体区段包含相对于缺少所述重组染色体区段的植物赋予对黑茶藨子长管蚜生物型 nr:1的抗性的等位基因。
7.本发明还提供了一种良种莴苣植物,其在染色体8上包含来自刺莴苣的至少第一重组染色体区段,其中所述第一重组染色体区段包含相对于缺少所述第一重组染色体区段的植物赋予对黑茶藨子长管蚜生物型nr:1的抗性的等位基因,且其中所述植物在染色体4上还包含第二重组染色体区段,其中所述第二重组染色体区段包含相对于缺少所述第二重组染色体区段的植物赋予对黑茶藨子长管蚜生物型 nr:1的抗性进一步改进的等位基因。在一些实施方案中,所述第二重组染色体区段在染色体4上包含选自由标记基因座m13(seq id no:61)、标记基因座m14(seq id no:66)、标记基因座m15(seq id no: 67)、标记基因座m16(seq id no:68)、标记基因座m17(seq id no: 69)、标记基因座m18(seq id no:70)、标记基因座m19(seq id no: 75)、标记基因座m20(seq id no:76)、标记基因座m21(seq id no: 81)、标记基因座m22(seq id no:86)和标记基因座m23(seq id no: 91)组成的群组的标记。
8.另外,本发明提供了良种莴苣植物的植物部分,其在染色体8上包含来自刺莴苣的至少第一重组染色体区段,其中所述第一重组染色体区段包含相对于缺少所述第一重组染色体区段的植物赋予对黑茶藨子长管蚜生物型nr:1的抗性的等位基因,且其中所述植物在染色体4上还包含第二重组染色体区段,其中所述第二重组染色体区段包含相对于缺少所述第二重组染色体区段的植物赋予对黑茶藨子长管蚜生物型nr:1的抗性改进的等位基因,并且其中所述植物部分包含所述第一和所述第二重组染色体区段。在某些实施方案中,所述植物部分是细胞、种子、根、茎、叶、头、花或花粉。在进一步的实施方案中,本发明提供了良种莴苣植物的种子,其在染色体8上包含来自刺莴苣的至少第一重组染色体区段,其中所述第一重组染色体区段包含相对于缺少所述第一重组染色体区段的植物赋予对黑茶藨子长管蚜生物型nr:1的抗性的等位基因,且其中所述植物在染色体4上还包含第二重组染色体区段,其中所述第二重组染色体区段包含相对于缺少所述第二重组染色体区段的植物赋予对黑茶藨子长管蚜生物型 nr:1的抗性进一步改进的等位基因。在仍进一步的实施方案中,包含所述第一和所述第二重组染色体区段的所述植物的种子的代表性样品已经以atcc登录号pta-126067保藏。
9.另一方面,本发明提供了一种产生对黑茶藨子长管蚜生物型nr:1 具有改进抗性的良种莴苣植物的方法,其包括使在所述植物的基因组中侧接有染色体8上的标记基因座m5(seq id no:11)和标记基因座m4(seq id no:46)的重组染色体区段内的黑茶藨子长管蚜生物型nr:1抗性等位基因渗入所述植物内,其中相对于缺少所述等位基因的植物,所述渗入的黑茶藨子长管蚜生物型nr:1抗性等位基因赋予所述植物对黑茶藨子长管蚜生物型nr:1的抗性。在一些实施方案中,所述渗入包括:a)将包含所述重组染色体区段的植物与自身或与不同基因型的第二莴苣植物杂交,以产生一种或多种子代植物;以及 b)选择包含所述重组染色体区段的子代植物。在其他实施方案中,选择子代植物包括检测包含标记基因座m1(seq id no:26)、标记基因座m2(seq id no:16)、标记基因座m4(seq id no:46)、标记基因座m5(seq id no:11)、标记基因座m7(seq id no:21)、标记基因座m8(seq id no:41)、标记基因座m10(seq id no:36) 或标记基因座m11(seq id no:31)的核酸。在一些实施方案中,子代植物是f
2-f6子代植物。在其他实施方案中,所述渗入包括回交,标记辅助选择或测定对黑茶藨子长管蚜生物型nr:1的所述抗性。在进一步的实施方案中,所述回交包括2-7代回交。在其他实施方案中,所述植物进一步包含在所述植物的基因组中侧接有染色体8上
的标记基因座m5(seq id no:11)和标记基因座m4(seq id no:46)或染色体4上的标记基因座m13(seq id no:61)和标记基因座 m23(seq id no:91)的重组染色体区段内的第二渗入的黑茶藨子长管蚜生物型nr:1抗性等位基因。本发明进一步提供了可通过本文提供的方法获得的莴苣植物。
10.本发明还提供了选择对黑茶藨子长管蚜生物型nr:1表现出抗性的莴苣植物的方法,其包括:a)将权利要求1的莴苣植物与自身或与不同基因型的第二莴苣植物杂交,以产生一种或多种子代植物;以及 b)选择包含所述黑茶藨子长管蚜生物型nr:1抗性等位基因的子代植物。在一些实施方案中,选择所述子代植物包括检测与所述黑茶藨子长管蚜生物型nr:1抗性等位基因遗传连锁的标记基因座。在进一步的实施方案中,选择所述子代植物包括检测在所述植物的基因组中侧接有染色体8上的标记基因座m5(seq id no:11)和标记基因座 m4(seq id no:46)的染色体区段内的标记基因座或与所述染色体区段遗传连锁的标记基因座。在其他实施方案中,选择子代包括检测包含标记基因座m1(seq id no:26)、标记基因座m2(seq id no:16)、标记基因座m4(seq id no:46)、标记基因座m5(seq id no:11)、标记基因座m7(seq id no:21)、标记基因座m8(seq id no:41)、标记基因座m10(seq id no:36)或标记基因座m11(seq id no:31) 的核酸。在一些实施方案中,所述子代植物是f
2-f6子代植物。在其他实施方案中,产生所述子代植物包括回交。
附图说明
11.图1:示出了在染色体8上鉴定的与黑茶藨子长管蚜生物型nr:1 抗性qtl相关的标记的遗传位置的概况。
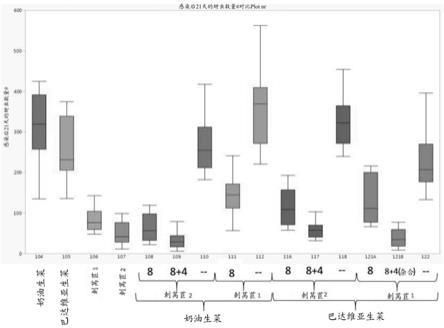
12.图2:示出了黑茶藨子长管蚜生物型nr:1抗性测定的结果。通过 qtl定位鉴定出的抗性基因座从两个不同的刺莴苣品系(显示为“刺莴苣1”和“刺莴苣2”)渗入两个不同的良种生菜品种(奶油生菜 (butterhead)和巴达维亚生菜(batavia))中,并向植物接种了固定数量的蚜虫。抗性表示为感染后(dpi)21天每棵植物上计数的蚜虫数量。除非另有说明,否则渗入在测试植物中是固定的。
具体实施方式
13.莴苣(l.sativa)是属于莴苣属和菊科的物种。此物种在商业上称为生菜。生菜主要作为叶类蔬菜种植,供新鲜市场消费,且通常分为七个主要栽培品种组,每个组具有多个品种:(i)叶、散叶、刈叶或成束生菜;(ii)直立生菜/cos;(iii)卷心生菜或脆头生菜;(iv)奶油生菜;(v) 法国脆生菜(summercrisp)或巴达维亚生菜;(vi)嫩茎莴苣(celtuce)或茎用生菜(stem);以及(vii)油籽生菜。生菜与几种其他莴苣物种密切相关,包括但不限于野生物种刺莴苣(l.serriola)。与莴苣相比,刺莴苣 (也称为带刺的生菜)被认为是在温带和亚热带地区发现的一种侵袭性田间作物杂草。
14.生菜是一种高价值作物,在世界范围内具有重要的经济意义。通常,种植者努力生产需要最少加工并且可以直接食用的生菜。这就要求生菜头在收获时必须没有昆虫。蚜虫是生菜作物的主要害虫,因为它们的生命周期短并且能够传播植物病毒。黑茶藨子长管蚜 (mosley)(n.ribisnigri)是全世界生菜中主要的蚜虫物种。收获时蚜虫的存在使菜头和沙拉包装无法销售,给种植者造成了巨大的经济损失。由于这种蚜虫更喜欢以生菜头的内叶
为食,因此在某些生菜类型中,菜头的封闭性质使其难以施用农药到达进食部位。此外,消费者越来越喜欢不含农药的作物。因此,有必要鉴定和开发对黑茶藨子长管蚜具有宿主植物抗性的栽培品种。
15.直到2007年左右,来自野生生菜物种毒莴苣(lactuca virosa)(l. virosa)的nr基因被广泛用作栽培生菜品种中对黑茶藨子长管蚜的抗性的有效机制。此时,在欧洲出现了能够感染含有nr基因的生菜品种的蚜虫群体的报道,这表明黑茶藨子长管蚜的新型抗性突破生物型。虽然nr基因对先前表征的生物型(nr:0)是有效的,但发现对新生物型无效。这种新的生物型被正式认定为nr:1,并造成了整个欧洲的生菜作物损失,包括西班牙、法国、德国、荷兰和英国。
16.本发明通过提供对黑茶藨子长管蚜生物型nr:1具有抗性的良种莴苣植物代表了本领域的显著进步。这样的植物可被称为黑茶藨子长管蚜生物型nr:1抗性生菜品种的植物。进一步提供了产生这种黑茶藨子长管蚜生物型nr:1抗性生菜植物、品系和品种的方法。本文还公开了与促成黑茶藨子长管蚜生物型nr:1抗性的数量性状基因座 (qtl)连锁的分子标记。通过使用本文描述的此类标记和方法,本领域技术人员可以增加生菜植物中黑茶藨子长管蚜生物型nr:1抗性的程度,并选择植物以增加对黑茶藨子长管蚜生物型nr:1抗性的易感性。在特定的实施方案中,所述方法在包含一个或多个促成在刺莴苣中发现的黑茶藨子长管蚜生物型nr:1抗性的qtl的生菜植物上进行。
17.已经在多种莴苣物种中鉴定了黑茶藨子长管蚜生物型nr:1抗性来源。例如,对毒莴苣登录物的研究在毒莴苣登录号pi273597中鉴定了染色体6和7上的黑茶藨子长管蚜生物型nr:1抗性qtl(wo 2016/066748)。但是,这些qtl所赋予的抗性仅在毒莴苣植物而非莴苣植物中评估。此外,wo 2011/058192报告了刺莴苣衍生的对.黑茶藨子长管蚜生物型nr:1的抗性是单基因和显性的,而相同的发明人在wo 2012/066008和wo 2012/065629中报告了刺莴苣衍生的对.黑茶藨子长管蚜生物型nr:1的抗性是单基因和隐性的。此外,对于任何公开的刺莴苣衍生的抗性,未提供在莴苣背景下的遗传信息、遗传 /分子标记或抗性谱。
18.本发明所代表的一项显著进步在于,在一个实施方案中,其提供了由染色体8上的新型qtl所赋予的生菜植物中的黑茶藨子长管蚜生物型nr:1抗性以及包含所述qtl的来自刺莴苣的新型重组染色体区段,以及其生产方法。在另一个实施方案中,本发明提供了由与染色体8上的新型qtl一起存在的染色体4上的新型qtl所赋予的生菜植物中的改进的黑茶藨子长管蚜生物型nr:1抗性,以及包含所述 qtl的来自刺莴苣的新型重组染色体区段,包括其生产方法。令人惊讶地发现,qtl可以组合使用以获得增加的抗性。本文提供了新基因座的新型标记,从而允许在新品种的开发过程中基因座被准确地渗入和追踪。因此,本发明允许衍生自刺莴苣的黑茶藨子长管蚜生物型nr:1抗性基因座渗入潜在的任何所需的良种生菜品种中。
19.在某些实施方案中,本文提供了植物,其在染色体8上包含渗入的黑茶藨子长管蚜生物型nr:1抗性等位基因,其中所述等位基因相对于不包含所述等位基因的植物赋予对黑茶藨子长管蚜生物型nr:1 的抗性。在进一步的实施方案中,提供了植物,其在染色体8和4上包含渗入的黑茶藨子长管蚜生物型nr:1抗性等位基因的组合。
20.在一些实施方案中,渗入的黑茶藨子长管蚜生物型nr:1抗性等位基因被定义为位于侧接有染色体8上的标记基因座m5(seq idno:11)和标记基因座m4(seq id no:46)的来
自刺莴苣的重组染色体区段内。在其他实施方案中,这样的区段可以包含以下中的一个或多个:标记基因座m2(seq id no:16)、标记基因座m7(seq id no: 21)、标记基因座m1(seq id no:26)、标记基因座m11(seq id no: 31)、标记基因座m10(seq id no:36)和标记基因座m8(seq id no: 41)。标记基因座m5在公共莴苣参考基因组lsat_salinas_v7的 106,984,777 bp处包含从t到c的snp变化,标记基因座m2在公共莴苣参考基因组lsat_salinas_v7的110,784,917 bp处包含从a到t 的snp变化,标记基因座m7在公共莴苣参考基因组lsat_salinas_v7 的112,532,048 bp处包含从c到t的snp变化,标记基因座m1在公共莴苣参考基因组lsat_salinas_v7的113,983,446 bp处包含从t到 c的snp变化,标记基因座m11在公共莴苣参考基因组 lsat_salinas_v7的122,770,672 bp处包含从c到t的snp变化,标记基因座m10在公共莴苣参考基因组lsat_salinas_v7的124,352,100 bp处包含从t到c的snp变化,标记基因座m8在公共莴苣参考基因组lsat_salinas_v7的132,833,792 bp处包含从c到g的snp变化,并且标记基因座m4在公共莴苣参考基因组lsat_salinas_v7的 136,545,853 bp处包含从t到g的snp变化。生菜的公共基因组可从例如lgr.genomecenter.ucdavis.edu处获得,并且本领域技术人员将理解如何将本申请中首次提供的标记序列定位在公共基因组的任何版本(或以后的版本)上。
21.尽管莴苣植物可以在所有指示的标记处包含供体(刺莴苣)等位基因,但是染色体8上的qtl区间侧接的标记基因座m5(seq id no: 11)和标记基因座m4(seq id no:46)的有利等位基因是轮回亲本等位基因。对于间质标记基因座m2(seq id no:16)、标记基因座 m7(seq id no:21)、标记基因座m1(seq id no:26)、标记基因座 m11(seq id no:31)、标记基因座m10(seq id no:36)和标记基因座m8(seq id no:41),有利的等位基因是来自供体亲本的等位基因。
22.在其他实施方案中,本发明提供了植物,其在染色体8上包含来自刺莴苣的新型重组染色体区段以及在染色体4上包含来自刺莴苣的新型重组染色体区段。出人意料的是,该组合提供对黑茶藨子长管蚜生物型nr:1的累加抗性。进一步提供了产生包含改进抗性的这种植物的方法。在一些实施方案中,渗入的黑茶藨子长管蚜生物型nr:1 抗性等位基因被定义为位于侧接有标记基因座m13(seq id no:61) 和标记基因座m23(seq id no:91)的来自刺莴苣的重组染色体区段内的染色体4上。在其他实施方案中,这样的区段可以包含以下一个或多个:标记基因座m14(seq id no:66)、标记基因座m15(seq idno:67)、标记基因座m16(seq id no:68)、标记基因座m17(seqid no:69)、标记基因座m18(seq id no:70)、标记基因座m19(seqid no:75)、标记基因座m20(seq id no:76)、标记基因座m21(seqid no:81)和标记基因座m22(seq id no:86)。标记基因座m13 在公共莴苣参考基因组lsat_salinas_v7的309,028,468 bp处包含从g 到t的snp变化,标记基因座m14在公共莴苣参考基因组 lsat_salinas_v7的317,543,051 bp处包含从c到t的snp变化,标记基因座m15在公共莴苣参考基因组lsat_salinas_v7的324,002,441 bp处包含从c到t的snp变化,标记基因座m16在公共莴苣参考基因组lsat_salinas_v7的331,652,666 bp处包含从a到t的snp变化,标记基因座m17在公共莴苣参考基因组lsat_salinas_v7的 341,160,568 bp处包含从c到t的snp变化,标记基因座m18在公共莴苣参考基因组lsat_salinas_v7的348,314,352 bp处包含从a到g 的snp变化,标记基因座m19在公共莴苣参考基因组lsat_salinas_v7 的357,158,000 bp处包含从a到t的snp变化;标记基因座m20在公共莴苣参考基因组lsat_
salinas_v7的361,400,802 bp处包含从c到 t的snp变化,标记基因座m21在公共莴苣参考基因组 lsat_salinas_v7的365,781,913 bp处包含从c到t的snp变化,标记基因座m22在公共莴苣参考基因组lsat_salinas_v7的371,266,283 bp处包含从c到t的snp变化,并且标记基因座m23在公共莴苣参考基因组lsat_salinas_v7的373,021,175 bp处包含从c到t的snp 变化。生菜的公共基因组可从例如lgr.genomecenter.ucdavis.edu处获得,并且本领域技术人员将理解如何将本申请中首次提供的标记序列定位在公共基因组的任何版本(或以后的版本)上。
23.表2还指示了存在于snp位置处的供体(刺莴苣)等位基因的核苷酸(因此,轮回亲本等位基因的核苷酸是针对snp位置所示的替代物)。尽管莴苣植物在所有指示的标记处都可能含有供体等位基因,但在染色体4上qtl区间侧接的标记基因座m13(seq id no:61) 和标记基因座m23(seq id no:91)的有利等位基因优选为轮回亲本等位基因。对于间质标记基因座m14(seq id no:66)、标记基因座 m15(seq id no:67)、标记基因座m16(seq id no:68)、标记基因座m17(seq id no:69)、标记基因座m18(seq id no:70)、标记基因座m19(seq id no:75)、标记基因座m20(seq id no:76)、标记基因座m21(seq id no:81)和标记基因座m22(seq id no:86),有利的等位基因是来自供体亲本的等位基因。
24.在某些实施方案中,本发明提供了产生或选择对黑茶藨子长管蚜 nr:1表现出抗性的生菜植物的方法,其包括:a)将本文提供的生菜植物与自身或与不同基因型的第二生菜植物杂交,以产生一种或多种子代植物;以及b)选择包含黑茶藨子长管蚜生物型nr:1抗性等位基因的子代植物。在一些实施方案中,本发明的方法包括通过检测包含标记基因座m1(seq id no:26)、标记基因座m2(seq id no:16)、标记基因座m4(seq id no:46)、标记基因座m5(seq id no:11)、标记基因座m7(seq id no:21)、标记基因座m8(seq id no:41)、标记基因座m10(seq id no:36)或标记基因座m11(seq id no:31) 的核酸来选择子代植物。
25.因为遗传上不同的植物品系可能难以杂交,所以使用常规育种方法将黑茶藨子长管蚜生物型nr:1抗性基因座和/或等位基因渗入栽培品系可能需要过大的分离群体用于子代筛选,且结果不确定。因此,标记辅助选择(mas)是将赋予对黑茶藨子长管蚜生物型nr:1的抗性的基因座有效渗入良种栽培品种所必需的。本发明首次通过提供改进和有效的标记以用于检测与黑茶藨子长管蚜生物型nr:1抗性相关的基因型来实现有效的mas,而无需为了观察表型而使大的植物群体生长至成熟。
26.i.生菜中与对黑茶藨子长管蚜生物型nr:1的抗性相关的基因组区域、基因座和多态性
27.本发明提供了与用于在植物育种期间追踪渗入的多态性核酸和连锁标记一起,一个或多个与对黑茶藨子长管蚜生物型nr:1的抗性相关的基因座向生菜中的新型渗入。
28.诸位发明人已经鉴定了超过20种对黑茶藨子长管蚜生物型nr:1 具有抗性的刺莴苣登录物。可以筛选任何已知的刺莴苣登录物的抗性,并用作本文所述的渗入片段的来源。由于刺莴苣是一种野生物种,因此也可以从其最初发现的地区(如欧洲、亚洲和北非)收集登录物。另外,刺莴苣登录物可获自基因库,包括荷兰瓦赫宁根(wageningen) 的荷兰遗传资源中心(cgn)及美国农业部(usda)的国家植物种质系统(the national plant germplasm system of the us department ofagriculture)。另外,以atcc登录号pta-126067保藏的种子可以用作染色体8上的重组染色体区段以及染色体4上的重组渗入的来
源。
29.在一个实施方案中,本发明提供了用于从刺莴苣的任何其他登录物获得赋予对黑茶藨子长管蚜生物型nr:1的抗性的基因座的材料和方法。使用本文所列出的信息,包括但不限于本文提供的多态性标记,可以将来自刺莴苣的对黑茶藨子长管蚜生物型nr:1的抗性渗入莴苣品种中,而没有在其他方面与刺莴苣相关的不良农艺特性。
30.使用本发明的改进的遗传标记和测定,本发明人能够成功地鉴定出赋予生菜植物对黑茶藨子长管蚜生物型nr:1的抗性的新型渗入。在某些实施方案中,本发明提供了生菜植物,其在染色体8上的标记基因座m5(seq id no:11)与标记基因座m4(seq id no:46)之间,和/或在染色体4上的标记基因座m13(seq id no:61)与标记基因座m23(seq id no:91)之间包含供体dna。
31.ii.与对黑茶藨子长管蚜生物型nr:1的抗性相关的基因组区域的渗入
32.标记辅助渗入涉及将由一个或多个标记限定的染色体区域从第一遗传背景转移到第二遗传背景。含有渗入的基因组区域的杂交后代可通过来自第一遗传背景的所需渗入的基因组区域所特有的标记与第二遗传背景所特有的连锁和非连锁标记的组合来鉴定。
33.本发明提供了用于鉴定和追踪本文公开的一个或多个基因组区域从黑茶藨子长管蚜生物型nr:1抗性植物渗入到栽培品系中的新型准确标记。本发明进一步提供了用于在植物育种过程中鉴定和追踪本文公开的新型渗入的标记,包括表1和2中列出的标记。
34.在本发明的任何基因组区间内或与之连锁的标记可用于多种育种工作,包括将与害虫抗性相关的基因组区域渗入所需的遗传背景中。例如,与本文所述的害虫抗性相关的标记的40cm、20cm、15cm、 10cm、5cm、2cm或1cm内的标记可用于与害虫抗性表型相关的基因组区域的标记辅助渗入。
35.还提供了生菜植物,其包含一个或多个与所需表型相关的渗入区域,其中至少10%、25%、50%、75%、90%或99%的剩余基因组序列携带轮回亲本种质所特有的标记。还提供了包含渗入区域的生菜植物,该渗入区域包含与本文提供的及与害虫抗性表型相关的基因组区域和标记紧密连锁或相邻的区域。
36.iii.对黑茶藨子长管蚜生物型nr:1具有抗性的生菜品种的开发
37.对于大多数育种目标,商业育种者使用“栽培”,“栽培类型”或“良种”的种质。此种质较容易育种,因为在评估园艺性能时通常表现良好。已经开发了多种栽培的生菜类型,包括莴苣,其在农业上是良种并且适于商业栽培。生菜栽培品种组包括但不限于cos、刈叶生菜、茎生菜(或芦笋)、奶油生菜、脆头生菜(或卷心或甘蓝)、拉丁(latin) 和油籽生菜组(de vries,gen.resources and crop evol.44:165-174, 1997)。然而,缺乏等位基因多样性可以抵消栽培种质提供的性能优势。育种者通常会接受这种折衷,因为用栽培材料进行育种比用遗传多样的来源进行育种进展更快。
38.相反,当栽培种质与非栽培种质杂交时,育种者可以获得来自非栽培类型的新型等位基因。然而,由于与不同品系之间的杂交相关的育性问题以及来自非栽培亲本的负连锁阻力,该方法提出了巨大的难题。在生菜植物中,非栽培类型(如刺莴苣)可以提供与抗病性相关的等位基因。然而,这些非栽培类型的园艺品质可能较差。
39.在避免遗传连锁的有害基因座或遗传力低的问题的同时,将所需抗性基因从非栽培品系渗入良种栽培品系的过程是一个漫长且通常艰巨的过程。在部署源自野生近缘种的
基因座时,通常期望引入最小或截短的渗入,该渗入提供所需性状但没有有害影响。为了帮助渗入,可靠的标记测定优于表型筛选。通过简化关键属性的遗传学以允许集中于数量性状如害虫抗性的遗传增益来进一步取得成功。此外,通过获得mas的准确标记,可以极大地促进从非栽培品系渗入基因组区域的过程。
40.因此,本领域技术人员将理解,本发明提供的基因座、多态性和标记允许将本文中鉴定的任何基因组区域追踪并引入任何遗传背景。另外,本文公开的与害虫抗性相关的基因组区域可以从一种基因型渗入到另一种基因型,并使用mas来追踪。因此,发明人关于与害虫抗性相关的准确标记的发现将促进具有有益表型的生菜植物的开发。例如,可以使用本发明的标记对种子进行基因分型,以选择包含与害虫抗性相关的所需基因组区域的植物。此外,mas允许鉴定对于所需渗入纯合或杂合的植物。
41.物种间杂交也可导致重组受到抑制,并且植物的育性或繁殖力低。例如,已观察到大麦中的番茄线虫抗性基因mi、mla和mlg基因;小麦中的yr17和lr20基因;葡萄藤中的run1基因以及花生中的rma基因的重组受到抑制。减数分裂重组是经典育种所必需的,因为它能够在遗传背景之间转移有利的基因座,去除有害的基因组片段,以及累加在遗传上紧密连锁的性状。因此,受抑的重组迫使育种者扩大子代筛选的分离群体,以便获得所需的遗传组合。
42.大量群体的表型评估是耗时的,资源密集的并且在每种环境下均不可再现。标记辅助选择提供了一种可行的替代方案。设计成检测独特多态性(如snp)的分子测定是通用的。然而,它们可能无法在单次测定中区分生菜物种内部和之间的基因座。染色体的结构重排(如缺失)会损害合成标记的寡核苷酸的杂交和延伸。在发生复制事件的情况下,多个拷贝在单个反应中无区别地扩增。因此,准确和高度预测性标记的开发和验证对于成功的mas育种程序至关重要。
43.iv.标记辅助育种和基因工程技术
44.可用于实施本发明的遗传标记包括但不限于限制性片段长度多态性(rflp)、扩增片段长度多态性(aflp)、简单序列重复(ssr)、简单序列长度多态性(sslp)、单核苷酸多态性(snp)、插入/缺失多态性 (indel)、可变数量串联重复(vntr)和随机扩增多态性dna(rapd)、同工酶和本领域技术人员已知的其他标记。作物植物中的标记发现和开发为应用于标记辅助育种活动提供了初始框架(美国专利公布号: 2005/0204780、2005/0216545、2005/0218305和2006/00504538)。所得的“遗传图谱”是所表征的基因座(多态性核酸标记或可鉴定其基因座的任何其他基因座)彼此相对位置的表示。
45.包含少至单个核苷酸变化的多态性可以通过多种方式测定。例如,可以通过电泳技术进行检测,包括单链构象多态性(orita等人 (1989)genomics,8(2),271-278)、变性梯度凝胶电泳(myers(1985)ep 0273085)或切割片段长度多态性(life technologies,inc.,gaithersburg, md),但dna测序的广泛应用性通常使直接对扩增产物进行简单测序变得容易。一旦已知多态性序列差异,就可以设计用于子代测试的快速测定,通常涉及特定基因座的某些版本的pcr扩增(pasa; sommer等人(1992)biotechniques 12(1),82-87)或多个特定基因座的 pcr扩增(pamsa;dutton和sommer(1991)biotechniques,11(6), 700-7002)。
46.多态性标记充当用于测定植物以确定品系或品种的同一性程度的有用工具(美国专利号6,207,367)。这些标记形成了用于确定与表型的相关性的基础,并且可用于驱动遗
传增益。在本发明方法的某些实施方案中,多态性核酸可以用于在生菜植物中检测与害虫抗性相关的基因型,鉴定具有与害虫抗性相关的基因型的生菜植物,以及选择具有与害虫抗性相关的基因型的生菜植物。在本发明方法的某些实施方案中,多态性核酸可用于产生生菜植物,该生菜植物在其基因组中包含与害虫抗性相关的渗入的基因座。在本发明的某些实施方案中,多态性核酸可用于育种包含与害虫抗性相关的一个或多个基因座的子代生菜植物。
47.遗传标记可以包括“显性”或“共显性”标记。“共显性”标记揭示存在两个或更多个基因座(每个二倍体个体两个)。“显性”标记揭示仅存在单个基因座。标记优选以共显性方式遗传,以便容易检测到在二倍体基因座上存在两个基因座或在三倍体或四倍体基因座上存在多个基因座,并且它们没有环境变化,即,其遗传力为1。标记基因型通常在二倍体生物的每个基因座处包含两个标记基因座。每个基因座的标记等位基因组成可以是纯合或杂合的。纯合性是基因座上的两个基因座都表征为相同核苷酸序列的情况。杂合性是指基因座上的两个基因座不同的情况。
48.用于确定遗传多态性的存在或不存在(即,用于基因分型)的基于核酸的分析可以用于育种程序以进行鉴定、选择、渗入等。用于遗传多态性分析的多种遗传标记是可获得的,并且是本领域技术人员已知的。该分析可用于选择包含或连锁至与生菜植物中的害虫抗性连锁或相关的遗传标记的基因、基因部分、qtl、基因座或基因组区域。
49.如本文所用,核酸分析方法包括但不限于基于pcr的检测方法 (例如,taqman测定)、微阵列方法、基于质谱的方法和/或核酸测序方法(包括全基因组测序)。在某些实施方案中,通过使用核酸扩增方法可有利于dna、rna或cdna样品中多态性位点的检测。具体地,此类方法增加跨多态性位点或包括位于其远端或近端的位点和序列的多核苷酸的浓度。此类扩增的分子通过凝胶电泳、荧光检测方法或其他手段可容易检测。
50.一种实现这种扩增的方法采用聚合酶链式反应(pcr)(mullis等人 (1986)cold spring harbor symp.quant.biol.51:263-273;欧洲专利 50,424;欧洲专利84,796;欧洲专利258,017;欧洲专利237,362;欧洲专利201,184;美国专利4,683,202;美国专利4,582,788;和美国专利4,683,194),使用能够与以其双链形式定义多态性的近端序列杂交的引物对。也可以使用基于质谱的dna分型方法。此类方法在美国专利6,613,509和6,503,710以及见于其中的参考文献中公开。
51.可通过本领域中熟知的多种有效方法来检测或分型dna序列中的多态性,所述有效方法包括但不限于以下专利中公开的那些:美国专利号5,468,613、5,217,863;5,210,015;5,876,930;6,030,787; 6,004,744;6,013,431;5,595,890;5,762,876;5,945,283;5,468,613; 6,090,558;5,800,944;5,616,464;7,312,039;7,238,476;7,297,485; 7,282,355;7,270,981和7,250,252,这些专利全部以引用方式整体并入本文。然而,本发明的组合物和方法可与任何多态性分型方法结合使用以检测基因组dna样品中的多态性。所用的这些基因组dna 样品包括但不限于直接从植物分离的基因组dna、克隆的基因组 dna或扩增的基因组dna。
52.例如,可通过与基因座特异性寡核苷酸(aso)探针杂交来检测dna序列中的多态性,如美国专利号5,468,613和5,217,863中所公开的。美国专利号5,468,613公开了基因座特异性寡核苷酸杂交,其中核酸序列中的单个或多个核苷酸变异可通过以下过程在核酸中
检测到,在所述过程中扩增含有核苷酸变异的序列,将其点涂在膜上并用标记的序列特异性寡核苷酸探针处理。
53.靶核酸序列也可通过例如如美国专利号5,800,944中所公开的探针连接方法来检测,在所述探针连接方法中将感兴趣的序列扩增并与探针杂交,之后进行连接,以检测探针的标记部分。
54.微阵列也可用于多态性检测,其中寡核苷酸探针组以重叠的方式组装以表示单个序列,使得靶序列在一个点处的差异将导致部分探针杂交(borevitz等人.,genome res.13:513-523(2003);cui等人., bioinformatics 21:3852-3858(2005))。在任何一个微阵列上,预期将存在多个靶序列,其可表示基因和/或非编码区,其中每个靶序列是由一系列重叠寡核苷酸而不是单个探针来表示。此平台提供对多个多态性的高通量筛选。美国专利6,799,122;6,913,879;和6,996,476中描述了通过基于微阵列的方法对靶序列的分型。
55.用于检测snp和indel的其他方法包括单碱基延伸(sbe)方法。 sbe方法的实例包括但不限于美国专利号6,004,744;6,013,431; 5,595,890;5,762,876;和5,945,283中公开的那些。
56.在用于检测多态性的另一种方法中,可以通过美国专利号 5,210,015;5,876,930;和6,030,787中公开的方法来检测snp和indel,其中寡核苷酸探针具有共价连接到所述探针的5'和3'端的5'荧光报告染料和3'淬灭染料。当探针完整时,报告染料与淬灭染料的接近导致报告染料荧光受到抑制,例如通过forster型能量转移。在pcr期间,正向引物和反向引物与侧接多态性的靶dna的特异性序列杂交,而杂交探针与扩增的pcr产物内的含有多态性的序列杂交。在随后的 pcr循环中,具有5
’→
3'核酸外切酶活性的dna聚合酶裂解探针并将报告染料与淬灭染料分离,从而导致报告物的荧光增加。
57.在另一个实施方案中,可使用核酸测序技术对感兴趣的一个或多个基因座直接测序。用于核酸测序的方法是本领域中已知的,并且包括由以下公司提供的技术:454life sciences(branford,ct)、 agencourt bioscience(beverly,ma)、applied biosystems(foster city, ca)、li-cor biosciences(lincoln,ne)、nimblegen systems(madison, wi)、illumina(san diego,ca)和visigen biotechnologies(houston, tx)。此类核酸测序技术包括如以下的形式:平行珠阵列、连接法测序、毛细管电泳、电子微芯片、“生物芯片”、微阵列、平行微芯片和单分子阵列。
58.已经开发了各种基因工程技术,并且可以被本领域技术人员用来将性状引入植物中。在要求保护的发明的某些方面,通过将单个遗传基因座或转基因改变或引入其品种或祖先的基因组中来将性状引入生菜植物。将基因和多核苷酸修饰、缺失或插入植物的基因组dna 中的基因工程方法是本领域中公知的。
59.在本发明的特定实施方案中,可以通过植物基因组的位点特异性修饰来产生改良的生菜品系。基因工程方法包括,例如,利用序列特异性核酸酶,如锌指核酸酶(参见,例如,美国专利申请公布号 2011-0203012);工程化或天然的大范围核酸酶;tale-核酸内切酶(参见,例如,美国专利号8,586,363和9,181,535);和rna指导的核酸内切酶,如crispr/cas系统的那些(参见,例如,美国专利号8,697,359 和8,771,945以及美国专利申请公布号2014-0068797)。因此,本发明的一个实施方案涉及利用核酸酶或任何相关蛋白来进行基因
组修饰。此核酸酶可以在供体模板dna内异源性地提供以进行模板化基因组编辑,或在单独的分子或载体中提供。重组dna构建体还可包含编码一个或多个指导rna的序列,以便将核酸酶引导至待修饰的植物基因组内的位点处。用于改变或引入单个遗传基因座的其他方法包括,例如,利用单链寡核苷酸以在植物基因组中引入碱基对修饰(参见,例如,sauer等人,plant physiol,170(4):1917-1928,2016)。
60.用于定点改变或引入单个遗传基因座的方法是本领域中公知的,并且包括利用序列特异性核酸酶(如上述的)或切割基因组dna以在遗传基因座处产生双链断裂(dsb)或切口的蛋白质和指导rna的复合物的那些方法。如本领域中公知的,在修复由核酸酶引入的dsb 或切口的过程中,供体模板、转基因或表达盒多核苷酸可以整合到 dsb或切口位点处的基因组中。通过同源重组或非同源末端连接 (nhej),待整合的dna中同源臂的存在可促进在修复过程中插入序列在植物基因组中的采用和靶向。
61.在本发明的另一个实施方案中,遗传转化可用于将选择的转基因插入本发明的植物中,或替代地,可用于制备可通过回交引入的转基因。用于转化植物的方法是本领域技术人员公知的并且适用于许多作物物种,包括但不限于电穿孔、微粒轰击、农杆菌介导的转化以及原生质体直接摄取dna。
62.为了通过电穿孔进行转化,可以使用松散的组织,如细胞或胚性愈伤组织的悬浮培养物,或者替代地,可以直接转化未成熟的胚胎或其他组织化的组织。在此项技术中,通过以受控方式将所选细胞暴露于果胶降解酶(果胶酶)或机械损伤的组织,可以部分降解所选细胞的细胞壁。
63.用于将转化的dna区段递送至植物细胞的有效方法是微粒轰击。在这种方法中,颗粒被核酸包被并通过推力递送到细胞中。示例性颗粒包括由钨、铂和优选地金组成的那些颗粒。对于轰击,将悬浮液中的细胞在过滤器或固体培养基上浓缩。替代地,可以将未成熟的胚胎或其他靶细胞排列在固体培养基上。将待轰击的细胞放置在大颗粒阻挡板下方的适当距离处。
64.通过加速将dna递送到植物细胞中的方法的一个说明性实施方案是生物弹射颗粒递送系统,其可用于将dna包被的颗粒或细胞通过筛网(如不锈钢或nytex筛网)推进到靶细胞覆盖的表面上。筛网分散颗粒,以使得它们不会以大聚集体形式递送到受体细胞。微粒轰击技术是广泛适用的,并且可用于转化几乎任何植物物种。
65.农杆菌介导的转移是用于将基因座引入植物细胞的另一种广泛适用的系统。该技术的一个优势是可以将dna引入整个植物组织,从而无需从原生质体再生完整的植物。现代农杆菌转化载体能够在大肠杆菌以及农杆菌中复制,从而允许方便的操作(klee等人,nat. biotechnol.,3(7):637-642,1985)。此外,用于农杆菌介导的基因转移的载体的最新技术进步已经改善了基因和限制位点在载体中的排列,以促进能够表达各种多肽编码基因的载体的构建。所描述的载体具有侧接有启动子和聚腺苷酸化位点的便利的多接头区域,以用于直接表达插入的多肽编码基因。另外,含有装甲和卸甲的ti基因的农杆菌可以用于转化。
66.在农杆菌介导的转化有效的那些植物株系中,由于基因座转移的简便性和确定性,它是一种选择的方法。使用农杆菌介导的植物整合载体将dna引入植物细胞是本领域中公知的(fraley等人,nat. biotechnol.,3:629-635,1985;美国专利号5,563,055)。
67.植物原生质体的转化也可以使用基于磷酸钙沉淀、聚乙二醇处理、电穿孔的方法
以及这些处理的组合来实现(参见,例如potrykus 等人,mol.gen.genet.,199:183-188,1985;omirulleh等人,plant mol. biol.,21(3):415-428,1993;fromm等人,nature,312:791-793,1986; uchimiya等人,mol.gen.genet.,204:204,1986;marcotte等人,nature, 335:454,1988)。在choi等人(plant cell rep.,13:344-348,1994)和ellul 等人(theor.appl.genet.,107:462-469,2003)中例示了植物的转化和外源遗传元件的表达。
68.v.定义
69.提供了以下定义来更好地限定本发明并指导本领域普通技术人员实践本发明。除非另外说明,否则术语应按照相关技术领域的一般技术人员的常规用法来理解。
70.如本文所用,术语“植物”包括植物细胞、植物原生质体、可再生生菜植物的组织培养的植物细胞、植物愈伤组织、植物块以及在植物或植物部分(如花粉、花、种子、叶、茎等)中完整的植物细胞。
71.如本文所用,术语“群体”意指具有共同亲本衍生的植物的遗传异质集合。
72.如本文所用,术语“品种”和“栽培品种”意指通过其遗传谱系和性能可以从同一物种内的其他品种中鉴定出的一组类似植物。
73.如本文所用,“等位基因”是指染色体上给定基因座处的基因组序列的两种或更多种替代形式之一。
[0074]“数量性状基因座(qtl)”是编码影响表型的表达度的至少第一基因座的染色体位置。
[0075]
如本文所用,“标记”意指可用于区分生物体的可检测特征。此类特征的实例包括但不限于遗传标记、生物化学标记、代谢物、形态特征和农学特征。
[0076]
如本文所用,术语“表型”意指可受基因表达影响的细胞或生物体的可检测特征。
[0077]
如本文所用,术语“基因型”意指植物的特定等位基因组成。
[0078]
如本文所用,“良种”或“栽培”品种是指由育种并针对优异农学性能进行选择所产生的任何品种。“良种植物”是指属于良种品种的植物。许多良种品种是生菜育种领域的技术人员可获得且已知的。“良种群体”是在给定作物物种(如生菜)的农学上优异的基因型方面可用于代表现有技术水平的良种个体或品种的分类。类似地,“良种种质”或良种种质株系是农学上优异的种质。
[0079]
如本文所用,术语“渗入的”当参考遗传基因座使用时,是指已引入到新的遗传背景中(如通过回交)的遗传基因座。遗传基因座的渗入可通过植物育种方法和/或分子遗传学方法来实现。此类分子遗传学方法包括但不限于各种植物转化技术和/或提供同源重组、非同源重组、位点特异性重组的方法和/或提供基因座替代或基因座转化的基因组修饰。
[0080]
如本文所用,术语“重组”或“重组的”在染色体区段的情境下是指在构型上包含一个或多个遗传基因座的重组dna序列,其中所述一个或多个遗传基因座例如由于减数分裂期间同源染色体之间的重组事件而在自然界中不存在。
[0081]
如本文所用,术语“连锁的”当在核酸标记和/或基因组区域的情境下使用时,意指标记和/或基因组区域位于同一连锁群或染色体上,使得它们在减数分裂时倾向于一起分离。
[0082]
如本文所用,“耐受基因座”意指与对疾病或害虫的耐受性或抗性相关的基因座。例如,在一个实施方案中,根据本发明的耐受基因座可控制对黑茶藨子长管蚜生物型nr:1
的耐受性或易感性。
[0083]
如本文所用,植物中的“耐受性”或“改进的耐受性”是指植物在疾病条件下或在害虫侵染下例如通过维持产量而表现良好的能力。耐受性还可以是指植物在疾病条件下或在害虫侵染下维持植物活力表型的能力。耐受性是一个相对术语,表示与在类似疾病条件下或在相似害虫压力下生长的不同(耐受性较差)植物(例如,不同植物品种)相比,“耐受性”植物更能够保持表现。本领域技术人员将了解,植物对疾病或害虫条件的耐受性变化很大,并且可代表耐受性更高或耐受性更低的表型谱。然而,通过简单观察,本领域技术人员一般可确定不同植物、植物品种或植物科在疾病或害虫条件下的相对耐受性,且此外,还将识别“耐受性”的表型等级。
[0084]
如本文所用,植物中对疾病或害虫条件的“抗性”或“改进的抗性”是植物比非抗性或抗性较低的植物更能减少疾病或害虫负担的指示。抗性是一个相对术语,表示“抗性”植物与在类似疾病条件或害虫压力下生长的不同(抗性较低)植物(例如,不同植物品种)相比,更能够减轻疾病负担或害虫负担。本领域技术人员将了解,植物对疾病条件或害虫侵染的抗性变化很大,并且可代表抗性更高或抗性更低的表型谱。然而,通过简单观察,本领域技术人员一般可确定不同植物、植物品种或植物科在疾病条件或害虫压力下的相对抗性,且此外,还将识别“抗性”的表型等级。
[0085]
如本文所用,“抗性等位基因”意指与对害虫侵染的耐受性或抗性相关的核酸序列。
[0086]“序列同一性”和“序列相似性”可以通过使用全局或局部比对算法对两个核苷酸序列进行比对来确定。当通过例如程序gap或 bestfit或emboss程序“needle”(使用默认参数)进行最佳比对具有至少某一最小百分比的序列同一性时,序列可被称为“基本上相同”或“基本上相似”。这些程序使用needleman和wunsch全局比对算法在两个序列的整个长度上进行比对,从而最大化匹配数并最小化缺口数。通常,使用默认参数,其中缺口产生罚分=10,且缺口延伸罚分=0.5(对于核苷酸和蛋白质比对)。对于核苷酸,使用的默认评分矩阵是dnafull(henikoff&henikoff,pnas 89:10915-10919;1992)。序列比对和序列同一性百分比的计分可以例如使用计算机程序来确定,如可在万维网上的ebi.ac.uk/tools/psa/emboss_needle/处获得的 emboss。替代地,可以通过搜索数据库(如fasta、blast等)来确定序列相似性或同一性,但是应当检索命中并成对比对来比较序列同一性。如果序列同一性百分比为至少85%、90%、95%、98%、99%或更高(例如,至少99.1、99.2、99.3、99.4、99.5、99.6、99.7、99.8、 99.9(通过emboss“needle”使用默认参数来确定,即缺口产生罚分=10,缺口延伸罚分=0.5,对于核酸使用评分矩阵dnafull)),则两个核酸序列具有“大体序列同一性”。标记有时可能会表现出变异,尤其是在探针无法识别的区域。
[0087]
术语“约”用于指示一个值包括用于确定所述值的装置或方法的误差的标准偏差。除非明确指明仅指代替代方案或替代方案相互排斥,否则权利要求书中所用的术语“或”用于意指“和/或”,但本公开支持仅指代替代方案和“和/或”的定义。当在权利要求书中与词语“包含 (comprising)”或其他开放语言结合使用时,除非明确指出,否则词语“一个/种(a/an)”表示“一个(种)或多个(种)”。术语“包含(comprise)”、“具有(have)”和“包括(include)”是开放式连系动词。这些动词中的一个或多个的任何形式或时态,如“包含(comprises)”、“包含(comprising)”、“具有(has)”、“具有(having)”、“包括(includes)”和
“
包括(including)”也是开放式的。例如,“包含”、“具有”或“包括”一个或多个步骤的任何方法不限于仅具有那一个或多个步骤并且还涵盖其他未列出的步骤。类似地,“包含”、“具有”或“包括”一个或多个性状的任何植物不限于仅具有这一个或多个性状并且还覆盖其他未列出的性状。
[0088]
vi.保藏信息
[0089]
保藏物是由至少625粒生菜品系ja_bag-ja19-0689的种子组成,所述生菜品系包含来自刺莴苣的基因渗入,如本文所述。保藏在美国典型培养物保藏所(american type culture collection(atcc), 10801university boulevard,manassas,va.20110-2209usa)中完成。保藏物被指定atcc登录号pta-126067,并且保藏日期为2019年7 月24日。在本申请未决状态期间,享有保藏物权利的人可以请求获得所述保藏物。所述保藏物将保存在作为公共保藏处的atcc保藏处,为期30年或最近一次请求后的5年或者专利的可实施寿命(以较长者为准),如果在此期间不能存活,则将被替换。申请人并不放弃对根据本专利或任何其他形式的品种保护(包括植物品种保护法(美国法典第7篇2321及以下))授予的权利的任何侵犯。
[0090]
实施例
[0091]
实施例1.生菜中黑茶藨子长管蚜生物型nr:1抗性的作图
[0092]
在大规模筛选刺莴苣品系的过程中鉴定出超过20种对黑茶藨子长管蚜生物型nr:1具有抗性的登录物。将这些登录物的子集与易感的良种莴苣品系杂交,产生作图群体。使用非选择抗性测定的变型,测试了从这些杂交获得的f2群体对黑茶藨子长管蚜生物型nr:1的抗性。使用具有3个块和4个复制物的随机完整块设计(总共12个植物 /科)。在每次复制中均包括亲本品系和病理对照,总共16株植物/对照。首先将种子播种在岩棉平地中,并在4周后将其移植到装有土壤的8.5cm盆中。在第6周时,每株植物都接种了4个相似大小的具有生物型nr:1的蚜虫,并用穿孔袋覆盖,以使蚜虫限制在它们所放置的植物上。通过计数接种后14天植物上的蚜虫数量对试验进行评分。另外,从用于这些测定的植物中获取组织以用于对超过2000个标记基因座的基因分型。在该初始标记集合中,选择了超过900个标记来绘制赋予对黑茶藨子长管蚜生物型nr:1的抗性的遗传区域。最初的作图揭示了染色体8上的一个区域,该区域解释了对黑茶藨子长管蚜生物型nr:1的抗性的约28%表型变异。标记m1(seq id no: 26)被鉴定为最接近qtl峰的标记。
[0093]
在进一步的作图实验中,将数种不同的刺莴苣登录物与奶油生菜品种的良种莴苣品系杂交,并在染色体8上发现了两个重叠基因组区域。对于一组登录物,鉴定了标记m3(seq id no:1)与m4(seq idno:46)之间的29cm区域,而对于另一组登录物,鉴定了标记 m5(seq id no:11)与m6(seq id no:51)之间的27cm区域。这些区域在标记m4(seq id no:46)与m5(seq id no:11)之间重叠。标记m1(seq id no:26)位于标记m4(seq id no:46)与m5(seq idno:11)之间的基因组区域内,由此证实了在初始作图实验中鉴定出的qtl峰。因此,赋予黑茶藨子长管蚜生物型nr:1抗性的基因组区域位于染色体8上的标记m4(seq id no:46)与m5(seq id no:11) 之间。另外的标记m2(seq id no:16)、m7(seq id no:21)、m8(seqid no:41)、m9(seq id no:6)、m10(seq id no:36)和m11(seq id no:31)被鉴定为在侧接有标记m4(seq id no:46)和m5(seq idno:11)的区域内。表1示出了染色体8上与黑茶藨子长管蚜生物型 nr:1抗性qtl相关的标记,其可用于追踪和选择基因座。
[0094]
表1.染色体8上用于追踪刺莴苣衍生的对黑茶藨子长管蚜生物型nr:1的抗性的标记。
[0095][0096]
除了染色体8上的qtl,还鉴定了染色体4上的qtl。qtl最初被定位到染色体上位于标记m12(seq id no:56)与m23(seq idno:91)之间的区域。为了精细绘制染色体4上的基因座,开发了标记m13(seq id no:61)、m14(seq id no:66)、m15(seq id no: 67)、m16(seq id no:68)、m17(seq id no:69)、m18(seq id no: 70)、m19(seq id no:75)、m20(seq id no:76)、m21(seq id no: 81)和m22(seq id no:86)。将qtl进一步精细绘制为位于标记 m13(seq id no:61)与m22(seq id no:86)之间。表2示出了染色体4上与黑茶藨子长管蚜生物型nr:1抗性qtl相关的标记,其可用于追踪和选择基因座。
[0097]
表2.染色体4上用于追踪刺莴苣衍生的对黑茶藨子长管蚜生物型nr:1的抗性的标记。
[0098][0099]
此外,对于所有表型抗性植物,确定染色体8和染色体4上的 qtl区域具有刺莴苣来源,而当植物是表型易感时,染色体8和染色体4上的qtl区域中的基因组区域具有莴苣来源。这证实了黑茶藨子长管蚜生物型nr:1抗性刺莴苣登录物确实是在刺莴苣
×
莴苣作图群体中观察到的抗性表型的供体。
[0100]
实施例2.当渗入不同的莴苣背景中时,由在染色体4和8上鉴定的基因座赋予的抗
性的验证
[0101]
为了确定在刺莴苣中鉴定的黑茶藨子长管蚜生物型nr:1抗性基因座的功效,对莴苣植物进行使用固定数量的蚜虫的非选择测定,其中将染色体8上的基因座渗入植物中,或将染色体8上的基因座和染色体4上的基因座都渗入植物中。莴苣植物为巴达维亚生菜或奶油生菜品种。巴达维亚生菜背景的植物也包含nr基因,该基因提供对黑茶藨子长管蚜生物型nr:0的抗性,而奶油生菜植物则没有。使用两种不同的刺莴苣登录物作为抗性供体,以研究抗性刺莴苣登录物之中对黑茶藨子长管蚜生物型nr:1抗性的一致性。使用具有5个复制物和3-4个植物/复制物的随机完整设计。每次复制均放置易感(巴达维亚生菜和奶油生菜亲本品系)和抗性(用作供体的两个刺莴苣登录物) 对照。起初将种子播种在岩棉平地中,并在第5周时将其移植到装有土壤的12cm盆中。在第8周时,使植物接种5个相似大小的具有生物型nr:1的蚜虫,并用穿孔袋覆盖,以使蚜虫限制在植物上。在接种后21天通过计数每株植物上的蚜虫数量对试验进行一次评分。将抗性确定为接种后21天每株植物上存在的蚜虫数量,其中低的蚜虫计数表示高水平的抗性。
[0102]
观察到,当渗入两种莴苣生菜类型中时,染色体8上抗性基因座的纯合分布赋予了对黑茶藨子长管蚜生物型nr:1的稳健抗性(图2)。此外,染色体4上抗性基因座以纯合或杂合构型的存在进一步提高了两种莴苣生菜类型中对染色体8上的基因座所赋予的对黑茶藨子长管蚜生物型nr:1的抗性。从两种刺莴苣登录物渗入到两种莴苣生菜类型中的基因座所赋予的抗性是相当的(图2)。这些结果证实,可以从多种刺莴苣供体获得相同水平的抗性。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1