针对癌症的基于PTP的疫苗的制作方法
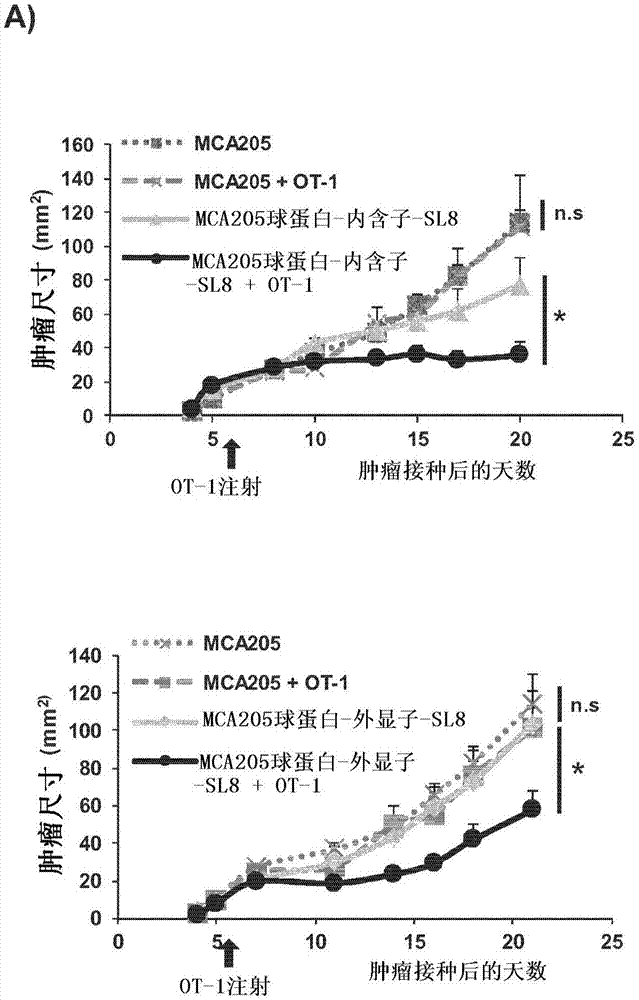
本发明涉及医药领域,并且通常用于治疗和预防领域。本发明更具体地涉及由具有7至50个氨基酸的肽组成的先行翻译产物(“ptp”),涉及含有这种ptp的微泡,涉及含有所述ptp的组合物,特别是疫苗组合物,并且涉及刺激受试者的免疫反应、优选针对肿瘤抗原的免疫反应的方法。
背景技术:
疫苗接种的主要目标是诱导可以控制人类的病毒感染性疾病和癌症的有效免疫反应。免疫系统分为两类:一方面是先天免疫系统,并且另一方面是适应性免疫系统。针对感染或转化细胞的细胞免疫反应需要活化适应性免疫系统。这种活化只能通过刺激抗原特异性细胞毒性t淋巴细胞如cd8+t细胞、b细胞和t辅助性t细胞如cd4+t细胞来实现。实际上,细胞毒性cd8+t细胞能够检测在其细胞表面呈递抗原的病毒感染细胞或癌细胞,所述抗原结合于mhci类分子。因此这种识别具有t细胞对感染细胞或肿瘤细胞的直接细胞毒性作用。然而,针对这些不同状态的适当免疫反应需要通过专职性抗原呈递细胞(papc)如树突状细胞和巨噬细胞对cd8+t细胞的活化,所述专职性抗原呈递细胞吸收外部肽物质以通过称为交叉呈递的过程将所述外部肽物质呈递在其mhci类分子上。直接和交叉呈递路径是检测和消除对宿主造成威胁的细胞的基本过程。这个过程还依赖于t辅助性t细胞,所述细胞识别来源于与mhcii类分子结合的外源蛋白质的13-20个氨基酸的短肽形式的抗原。数年前,据假设,直接呈递给mhci类限制途径的肽来源不是来源于全长蛋白质的降解,而是来源于所谓的缺陷型核糖体产物或drip。此后,进一步的研究支持了这一观点,即使尚未了解i类路径的肽的实际来源。发明人已经显示爱巴病毒(epsteinbarrvirus)的潜在蛋白质ebna1影响mrna翻译以抑制抗原呈递,并且以这种方式避免了它的检测。此外,他们观察到mrna翻译的速率与抗原呈递密切相关。另外,一些mhci类结合肽已被描述为由隐蔽性翻译(cryptictranslation)产生,其是指由非常规翻译机制在细胞中合成的多肽。这些可以是由内含子、内含子/外显子连接区、5'和3'非翻译区或交替翻译阅读框编码的肽。所有这些观察结果使得焦点从蛋白质降解转移至mrna翻译作为抗原产生的关键过程。最近,发明人已经显示,无论肽是由内含子表达还是由外显子表达,抗原呈递都是等效的,并产生所谓的先行翻译产物(ptp),所述先行翻译产物通过不同于产生全长蛋白质的规范事件的翻译事件产生。前述结果得到了以下事实的支持:如果从细胞核到细胞质的mrna输出被阻断,则外显子和内含子编码的肽的抗原呈递显著增加。总体而言,发明人已经展示,用于mhci类路径的抗原肽在很大程度上来源于mrna翻译事件,该mrna翻译事件与产生全长蛋白质的mrna翻译事件不同且无关,并且在早期扫描新合成的mrna期间在细胞核区室中发生(apcher,millot等人,2013,apcher,daskalogianni等人,2015)。这些ptp可能构成难以捉摸的drip。它们可能在mrna剪接发生之前产生,这例如解释了免疫系统可如何“耐受”组织依赖性替选性剪接产物。如今,旨在改善宿主免疫介导的肿瘤识别和破坏的癌症免疫疗法中的治疗性疫苗接种正成为新的研究热点。但在1996年,在临床试验中已经测试了来源于mage-1蛋白的i类结合合成表位作为基于肽的疫苗。然而,使用合成表位作为疫苗的同一组和其它组在黑素瘤患者中看不到任何有益的临床反应。接着,使用针对不同癌症的其它短肽,再次未在患者中展示任何有益的t细胞反应。接着,使用多种肽疫苗,特别是针对黑素瘤,但没有任何突破。最近,已经显示利用合成的长肽(超过20个氨基酸)的免疫更具免疫原性,并且具有比用短肽观察到的免疫更好的抗肿瘤生长作用。含有最小短抗原表位的两种肽之间的这些差异可见于以下事实:较长肽由于其三级结构而可以被保护不受细胞外降解,并且见于以下事实:它们太长而不能与任何细胞系的mhci类分子直接结合。此外,使用较长肽作为疫苗的益处在于它们需要被内化并且需要被papc中的蛋白酶体适当加工,然后在细胞表面被呈递并活化cd8+t细胞。此外,较长肽具有更好的机会含有数个表位,所述表位可以诱导不同cd8+t细胞的活化,并因此诱导多重免疫反应。已经研究了免疫细胞或肿瘤细胞分泌的外来体(exosome)在肿瘤免疫疗法中的潜力。外来体来源于多泡体(mvb)中的腔内(intralumenal)囊泡,并且选择性并入了特殊蛋白质。外来体是直径为30至100nm的囊泡。已经假设肿瘤来源外来体可能含有肿瘤抗原,因此可以用作癌症疫苗的肿瘤抗原来源。此外,许多研究小组报道,来源于树突状细胞(dc)的外来体可能是用于诱导特异性抗肿瘤免疫的适用且有效的药剂。然而,越来越多的证据表明来源于肿瘤的外来体是不完美的,因为它们可能在不同路径中通过不同作用诱导肿瘤免疫逃避,如通过抑制dc的分化或通过负调节nk细胞(valenti,huber等人,2006;clayton,mitchell等人,2008;whiteside,mandapathil等人,2011)。发明人现在在本文中描述了一种疫苗组合物,所述疫苗组合物包含由内含子或外显子序列产生的ptp,所述ptp优选与含有ptp的微泡、通常地外来体组合,所述疫苗组合物能够诱导针对肿瘤的适当cd8+t细胞免疫反应,从而使得完全抑制肿瘤生长,优选破坏肿瘤。技术实现要素:本发明涉及用于改善抗原特异性免疫反应的产品和方法,特别是在癌症治疗和预防领域。本发明基于如下意外发现:先行翻译产物(“ptp”),其由具有7至50个氨基酸的肽组成,通常包含至少一个mhci类表位,优选包含至少一个mhci类表位和至少一个mhcii类表位,能够在罹患癌症的受试者中诱导针对表达这种肽的肿瘤的有效、优选持续的免疫反应。因此,本发明的第一目的涉及一种用作受试者的疫苗、优选癌症疫苗的先行翻译产物(“ptp”),所述先行翻译产物由通常为5kda或更小且具有7至50个氨基酸的肽组成。ptp通常由选自以下的序列表达:内含子、3'或5'非翻译区(utr)、lncrna(长非编码rna)、mirna(微小rna)、基因间序列以及其组合。ptp优选包含至少一个mhci类表位和/或至少一个mhcii类表位。本发明的第二目的涉及一种微泡,通常是外来体或等效的肿瘤来源微泡,如黑素体,所述微泡包含至少一种ptp(优选数种ptp),通常为本文所述的ptp,所述ptp优选包含至少一个mhci类表位和/或至少一个mhcii类表位。本发明的第三目的涉及一种组合物,特别是疫苗组合物,所述组合物包含至少一种ptp(优选数种ptp)和/或微泡,通常为本文所述的外来体或肿瘤来源微泡,以及药学上可接受的载体或赋形剂。优选的疫苗组合物至少包含本文所述的第一ptp、微泡以及药学上可接受的载体或赋形剂。优选地,微泡包含至少一种由具有7至50个氨基酸的肽组成的第二ptp,所述第二ptp优选包含至少一个mhci类表位和/或至少一个mhcii类表位,所述微泡任选地包含第一ptp。本发明还涉及一种编码ptp的核酸序列,其根据本发明用作疫苗;并且涉及包含这种核酸序列和药学上可接受的载体或赋形剂的组合物,特别是疫苗组合物。在一个优选方面,本文所述的疫苗组合物是用于人类的。本发明还涉及这种ptp、微泡、核酸或组合物用于预防或治疗受试者的癌症的用途。本发明的另一个目的涉及一种针对特定抗原、优选肿瘤抗原在受试者中产生免疫反应或向受试者接种疫苗的方法,所述方法包含向所述受试者注射来源于所述抗原的根据本发明的ptp、包括所述ptp的微泡或包括所述ptp的疫苗组合物。本发明的另一个目的涉及一种预防或治疗受试者的癌症的方法,所述方法包含向所述受试者注射根据本发明的ptp、优选来源于由受试者的癌性肿瘤表达的多肽的ptp、包括所述ptp的微泡或包括所述ptp的疫苗组合物。具体实施方式主要组织相容性复合物(mhc)i类抗原呈递路径使得免疫系统可区别自身与非自身。尽管已对抗原肽的加工进行了广泛的研究,但对其起源知之甚少。发明人揭示称为先行翻译产物(“ptp”)的一类独特的肽在mrna翻译的先行轮次中产生,并且提供直接呈递至mhci类路径的抗原肽底物的主要来源。他们已展示,mhci类路径的大部分底物在mrna成熟的早期步骤期间通过细胞核内的非规范翻译机制并且在内含子被剪接掉之前合成(apcher等人,2013)。这个机制与产生全长蛋白质的机制无关。发明人还展示,与全长蛋白质相比,ptp是mhci类交叉呈递路径的肽的更好来源,并且现在在本文中揭示,这些ptp,特别是当与微泡组合时,可以用作疫苗,特别是用于有效预防或治疗癌症。因此,本发明的第一目的涉及一种用作受试者的疫苗、优选癌症疫苗的先行翻译产物(“ptp”),所述先行翻译产物由具有7至50个氨基酸残基或8至50个氨基酸残基的肽组成。先行翻译产物(“ptp”)在本文中被定义为衍生自非剪接mrna的肽,所述肽由内含子、外显子、3'和5'非翻译区(utr)、lncrna(长非编码rna)、mirna(微小rna)和/或基因间序列表达。优选地,ptp由具有7至50个氨基酸的肽组成,所述肽由选自以下的序列表达:i)内含子、3'或5'非翻译区(utr)、lncrna(长非编码rna)、mirna(微小rna)、基因间序列以及其组合,ii)内含子、lncrna(长非编码rna)、mirna(微小rna)、基因间序列以及其组合,或iii)内含子、lncrna(长非编码rna)、mirna(微小rna)和基因间序列。ptp通过不同于产生全长蛋白质的规范事件的翻译事件产生,该翻译事件在早期扫描新合成的mrna期间在细胞核区室中发生(apcher,millot等人,2013;apcher,daskalogianni等人,2015)。这些ptp优选不是源自病毒或细菌。它们通常由7至50个氨基酸残基的序列组成并且具有5kda或更小、通常3kda或更小的原子质量。ptp优选由7至30个氨基酸残基如7至27个氨基酸残基的序列组成。ptp可以例如包含7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49或50个氨基酸残基。ptp通常包含mhci类表位。从肿瘤细胞的细胞核区室纯化的ptp[在本文中也被鉴定为“肿瘤相关ptp”(ta-ptp)]通常引发针对肿瘤细胞所来自于的肿瘤的特异性抗肿瘤cd8+t细胞反应。根据本发明的优选ptp至少包含mhci类表位和/或mhcii类表位,所述mhcii类表位引发来自免疫系统的长效cd4+t细胞反应,从而延长抗肿瘤cd8+t细胞反应。本发明的ptp可以获得自或纯化自(并且称为“来源于”)任何蛋白质、多肽或抗原,针对所述蛋白质、多肽或抗原将会使用标准生物化学方法在待治疗/疫苗接种的受试者中引发特异性免疫反应。在肿瘤的情况下,ptp提取涉及用清洁剂或盐裂解肿瘤细胞,接着提取5kda或更小、优选3kda或更小的肽,并通过标准色谱方法包括柱上阴离子或疏水色谱法和/或亲和色谱法对其进行纯化。在本发明的另一个实施方式中,可以用柠檬酸盐磷酸盐缓冲液(ph3.3)从肿瘤细胞表面洗脱来源于ptp的抗原表位。可以通过质谱法分析抗原表位,并且可以进行肽从头测序。该分析方法确实允许在不使用序列数据库的情况下从串联质谱(ms/ms)中推导出肽的氨基酸序列。在鉴定来自内含子、外显子、3'和5'utr、lncrna、mirna或基因间区域的表位后,可以合成含有不同mhci类和/或ii类表位的新ptp。在本发明的一个具体实施方式中,本发明的ptp包含至少一个mhci类表位和/或至少一个mhcii类表位。优选地,本发明的ptp包含至少一个mhci类表位和至少一个mhcii类表位。在优选实施方式中,ptp是活化cd4+t细胞和/或cd8+t细胞cd8+t细胞的ptp。本文所述的特定ptp是选自以下任一项的ptp:seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:5、seqidno:6、seqidno:7、seqidno:8、seqidno:12、seqidno:13、seqidno:14、seqidno:15、seqidno:16、seqidno:17、seqidno:18、seqidno:19、seqidno:20、seqidno:21、seqidno:22和seqidno:23(参考表1和表2以及序列表)。在优选实施方式中,在受试者中用作癌症疫苗的ptp是来源于受试者的癌症肿瘤的ptp(“肿瘤相关ptp”或“ta-ptp”)。发明人已将这种ta-ptp鉴定为活化cd8+t细胞的ptp。在另一个优选实施方式中,将这种ptp与相应的全长蛋白质或多肽(即由相同mrna规范翻译的蛋白质或多肽)或其抗原组合用作癌症疫苗。本发明的另一个目的是一种编码本文所限定的ptp的核酸序列(dna或mrna),所述核酸序列在受试者中用作疫苗。本发明的另一个目的涉及一种微泡,通常是外来体或等效的肿瘤来源微泡,所述微泡包含/表达如本文所述的至少一种ptp。外来体是内体来源的囊泡,其在晚期内体多泡体与质膜融合后分泌于细胞外环境中(garin等人,2001;thery等人,2002)。已经显示来自各种组织类型的细胞分泌外来体,如树突状细胞、b淋巴细胞、肿瘤细胞和肥大细胞。来源于肿瘤细胞的外来体在本文中被鉴定为肿瘤来源微泡。从黑素瘤细胞获得的外来体或肿瘤来源微泡在本文中被鉴定为“黑素体”。不同来源的外来体展现分立的蛋白质和脂质部分组(thery等人,1999;thery等人,2001)。它们特别含有参与抗原呈递和免疫调节的蛋白质,指示外来体在细胞-细胞通讯中起作用,导致调节免疫反应。实际上,来自用肿瘤抗原来源的肽脉冲处理的树突状细胞(dc)的外来体在使用匹配肿瘤的动物模型中引发抗肿瘤反应(wolfers等人,2001,zitvogel等人,1998)。产生、纯化或使用外来体来用于治疗目的或作为研究工具的方法已描述于例如wo99/03499、wo00/44389和wo97/05900中,所述专利通过引用并入本文中。本领域已经描述了重组外来体,其来源于用编码重组蛋白质的质粒转染的细胞。这些重组外来体含有质粒编码的重组蛋白(wo00/28001)。在wo03/016522中描述了操作外来体的蛋白质含量和呈现抗原、佐剂和标志物以用于治疗目的或作为研究工具的方法。因此,发明人在本文中描述了一种微泡,通常是来源于肿瘤细胞的微泡,所述微泡包含/表达本文所定义的用作受试者的疫苗、优选癌症疫苗的ptp。在一个具体实施方式中,这种微泡包含数种ptp,特别是数种具有不同长度和任选具有不同来源即来源于不同(非剪接)mrna的ptp。肿瘤细胞产生的肿瘤来源微泡可以根据本领域已知的技术如通过离心、色谱法等收集和/或纯化。wo00/44389和us09/780,748中已经描述优选技术,所述专利通过引用并入本文中。发明人还在本文中描述了一种制备含有/表达本文所述的ptp的功能化的微泡/外来体/黑素体的方法,所述方法包含:-提供编码ptp的嵌合基因构建体;-将所述构建体引入微泡/外来体/黑素体产生细胞中来产生功能化的微泡/外来体/黑素体,所述功能化的微泡/外来体/黑素体含有/表达所述ptp,通常在其表面呈递所述ptp,以及-收集和/或纯化所述功能化的微泡/外来体/黑素体。由这些细胞产生的微泡可以根据本领域已知的技术如通过离心、色谱法等收集和/或纯化。wo00/44389和us09/780,748中已经描述优选技术,所述专利通过引用并入本文中。发明人还在本文中描述了一种产生本文所述的ptp的方法,所述方法包含:-提供编码ptp的嵌合基因构建体;-将所述构建体引入微泡/外来体/黑素体产生细胞中来产生功能化的微泡/外来体/黑素体,所述功能化的微泡/外来体/黑素体含有/表达所述ptp,通常在其表面呈递所述ptp,-收集和/或纯化所述功能化的微泡/外来体/黑素体,以及-从所述功能化的微泡/外来体/黑素体回收和/或纯化所述多肽或其片段。在本发明的上下文中,术语“包含/表达”来源于ptp的抗原表位或ptp的微泡(外来体或黑素体)表示含有来源于ptp的这种抗原表位或附着于其膜的ptp的微泡。来源于ptp的抗原表位可以暴露在微泡外,并且ptp通常包含在微泡内(即附着于膜的内侧或悬浮在微泡内)。通常,微泡使得ptp可有效传送至树突状细胞,并使得可在树突状细胞表面有效交叉呈递ptp和来源于ptp的抗原表位。本发明还涵盖包含上述嵌合基因构建体的载体,以及包含上述嵌合基因构建体或载体的重组细胞。载体可以是质粒、噬菌体、病毒、人造染色体等。典型实例包括质粒,如来源于市售质粒的质粒,特别是puc、pcdna、pbr等。其它优选载体来源于病毒,如复制缺陷型逆转录病毒、腺病毒、aav、杆状病毒或牛痘病毒。本领域技术人员可以根据应该使用载体的重组宿主细胞来调整所述载体的选择。在这方面,优选使用可以转染或感染哺乳动物细胞的载体。实际上,优选的重组宿主细胞是哺乳动物细胞。这些可以是原代细胞或已建立的细胞系。说明性实例包括纤维母细胞、肌肉细胞、肝细胞、免疫细胞等,以及其祖细胞或前体细胞。最优选的哺乳动物细胞是产生外来体的哺乳动物细胞。这些包括例如肿瘤细胞、树突状细胞、b和t淋巴细胞或肥大细胞。本发明的微泡可以单独用作疫苗。在一个优选实施方式中,将这种微泡与靶细胞或组织(例如肿瘤)表达的全长蛋白质或多肽和/或与至少一种ptp、通常与数种ptp组合使用,所述ptp衍生自对应于所述全长蛋白质或多肽的非剪接mrna。本发明的另一个目的涉及一种组合物,特别是疫苗组合物,优选癌症疫苗,所述组合物包含本文所述的产物,通常至少一种ptp,对应于ptp全长的蛋白质或多肽和/或本文所述的微泡(外来体或黑素体)以及药学上可接受的载体或赋形剂。本发明的优选组合物包含不同长度的数种ptp。本发明的另一种优选组合物包含活化cd4+t细胞和/或cd8+t细胞的ptp。当存在时,微泡通常包括(含有或表达)ptp,例如与组合物中存在的ptp相同的ptp,以及任选地,(至少一种)不同的ptp。优选的疫苗组合物包含至少一种本文所述的第一ptp、微泡以及药学上可接受的载体或赋形剂。优选地,微泡包含至少一种由具有7至50个氨基酸的肽组成的第二ptp,所述第二ptp优选包含至少一个mhci类表位和/或至少一个mhcii类表位,所述微泡任选地包含第一ptp。微泡可以是微泡的组合物,所述组合物包含表达所需ptp的重组微泡和来源于待治疗受试者的天然微泡,例如来源于待治疗受试者的肿瘤的微泡(肿瘤来源微泡)。在一个优选实施方式中,微泡是活化cd8+t细胞的微泡,通常为外来体或肿瘤来源微泡,如黑素体,所述微泡天然表达活化具有肿瘤的受试者的cd8+t细胞的ptp。在本发明的另一个不同的实施方式中,组合物是疫苗组合物,其包含编码本文所定义的ptp的核酸序列(dna或mrna)或基因构建体。基因疫苗接种可以使用如下物质进行:各种病毒载体,如牛痘病毒、痘病毒、腺病毒、腺相关病毒等;非病毒载体,如与各种脂质或肽组合物相关的核酸序列;或使用纯(例如,裸露或换言之不含任何转染促进剂的)核酸。疫苗接种可以通过各种注射途径进行,包括肌肉内、静脉内、皮下或皮内注射。各种载体递送装置或技术可以用于基因疫苗接种,包括基因枪或电穿孔。还可以使用用载体体外转染的细胞系使受试者免疫化。选择用于释放大量外来体的细胞系将是特别有利的。优选的癌症疫苗包含肿瘤相关的ptp以及外来体,优选肿瘤来源微泡,和/或对应于ptp全长的蛋白质或多肽,以及药学上可接受的载体或赋形剂。另一种优选的癌症疫苗包含ptp和微泡,所述ptp和微泡两者均来源于待接种疫苗的受试者的肿瘤,所述癌症疫苗优选还包含至少一种不同的ptp和/或表达相同ptp和/或至少一种不同ptp的外来体,以及药学上可接受的载体或赋形剂。另一种更优选的癌症疫苗还包含对应于ptp全长的蛋白质或多肽。可用于本发明上下文的药学上可接受的赋形剂、媒剂或载体是例如盐水、稀释剂、等渗或缓冲溶液如甘露醇20%,任选与稳定剂如等基因白蛋白或任何其它稳定性蛋白质、甘油等组合。合适佐剂的实例包括cpg寡脱氧核苷酸、凋亡诱导因子(aif)、热休克蛋白(hsp)、toll样受体(tlr)如tlr3激动剂(聚i:c)以及细胞因子和趋化因子如il-7、il-12、il-15和粒细胞巨噬细胞集落刺激因子(gm-csf)。本发明还涉及本文所述的本发明产品(ptp、微囊泡、核酸)的用途,其用于制备组合物,特别是疫苗组合物,用于预防或治疗受试者的疾病,特别是癌症。典型的疫苗组合物供使用于人类。本发明的目的还涉及一种针对具体靶标、优选肿瘤抗原或癌症/肿瘤细胞在受试者中产生免疫反应的方法,通常是向受试者接种疫苗的方法,所述方法包含向所述受试者注射来源于所述靶标的根据本发明的ptp、包括所述ptp的根据本发明的微泡或根据本发明的疫苗组合物。本发明的另一个目的是一种预防或治疗受试者的癌症的方法,所述方法包含向所述受试者注射根据本发明的ptp、优选来源于由受试者的癌性肿瘤表达的蛋白质或多肽的ptp、包括所述ptp的根据本发明的微泡或根据本发明的疫苗组合物。如本文所用,“治疗”是指试图改变经受治疗的受试者的自然进程的治疗性干预,并且可以被执行以用于预防(防治)或治疗目的。治疗的期望效果包括但不限于预防疾病的发生或复发、减轻症状、减少疾病的任何直接或间接病理后果、降低疾病进展的速度、减轻或缓和疾病状态以及缓解或改善预后。在优选实施方式中,本发明的组合物和方法用于延迟癌症的发展或减缓癌症的进展,通常是肿瘤生长的进展。通常,治疗将诱导受试者免疫系统的治疗反应,通常是cd4+和/或cd8+t细胞反应。诱导t细胞反应在本文中是指引发针对某种抗原的t细胞反应。在所述诱导之前,所述t细胞反应不存在,或低于检测水平或不起作用。在本文中,增强t细胞反应是指与所述增强之前针对某种抗原的t细胞的总体作用相比,使得所述t细胞的总体作用更高和/或更有效。例如,在所述增强后,可以产生更多针对所述抗原的t细胞。结果,额外产生的t细胞的作用提高了针对所述抗原的总体作用。或者,所述增强可以包含针对所述抗原的t细胞的作用的增加。所述t细胞可以例如与所述抗原更强和/或更快地反应。当然,所述增强的结果可以是产生额外的t细胞以及增加所述t细胞的作用。或者,所述增强可以仅包含产生额外的t细胞或增加t细胞的作用。所述治疗,通常是疫苗,旨在用于受试者。术语“受试者”或“个体”是指动物,通常是哺乳动物。哺乳动物的实例包括人类和非人类动物,诸如但不限于驯养动物(例如牛、绵羊、猫、狗和马)、非人类灵长类动物(如猴子)、兔子以及啮齿动物(例如小鼠和大鼠)。该治疗优选旨在用于有需要的人类,无论其年龄或性别。尤其考虑罹患癌症的受试者,或被认为“具有产生这种癌症的风险”的受试者,其中必须预防这种情况。患者通常具有肿瘤。除非在本公开中另外说明,否则肿瘤是癌性或恶性肿瘤。癌症或肿瘤可以是任何类型的癌症或瘤变。肿瘤通常是实体肿瘤,特别是来源于上皮、神经外胚层或间充质的实体肿瘤。其可以选自黑素瘤、肉瘤、癌瘤、淋巴瘤以及小儿肿瘤(神经胶质瘤),例如选自黑素瘤或肉瘤。在治疗的情形下,本发明适用于原发性肿瘤或继发性侵袭、局部区域或远端转移,并且在预防的情形下,是为了避免继发性恶性中枢神经系统累及,如观察到从黑素瘤、肺癌、肾癌、乳癌和结肠癌侵袭(转移)。在本发明的疫苗组合物中,ptp的存在量足以引发给定受试者的免疫系统针对期望标靶(病原体、靶细胞)的治疗反应,例如cd8+t细胞反应,通常为cd4+t细胞反应,优选cd4+和cd8+t细胞治疗反应,并且预防或治疗、通常控制疾病,优选癌症。当受试者是哺乳动物,优选人类时,疫苗组合物通常包含0.1至10毫克/千克体重的ptp,以及任选地,0.1至5毫克/千克体重的微泡。本文描述的能够诱导治疗性免疫反应的产品可以体内施用于有需要的任何哺乳动物受试者,特别是人类受试者。可以通过各种途径进行施用,如通过全身注射,例如静脉内、肌肉内、腹膜内、肿瘤内、皮下等。本领域技术人员可以通过如elisa、elispot、迟发型超敏反应、细胞内细胞因子染色和/或细胞外细胞因子染色的技术容易地确定治疗性免疫反应的检测。通过以下附图和实施例进一步说明本发明。然而,这些实施例和附图不应以任何方式解释为限制本发明的范围。附图图1:先行翻译产物(ptp)在肿瘤排斥中的作用。a)向小鼠皮下注射mca205wt肿瘤细胞或用编码球蛋白-内含子-sl8的质粒转染的mca205,该质粒编码球蛋白-外显子-sl8或卵白蛋白。在第6天或第4天,来自每个组的一半小鼠接受静脉内ot1细胞。评定肿瘤尺寸随时间的变化直到第20天。数据以平均值±sem的形式提供。*p<0.05(非配对学生t检验(unpairedstudentttest))。b)向小鼠皮下注射b16f10wt肿瘤细胞或用编码球蛋白-内含子-sl8的质粒转染的b16f10,该质粒编码球蛋白-外显子-sl8或卵白蛋白。在第3天,来自每个组的一半小鼠接受静脉内ot1细胞。评定肿瘤尺寸随时间的变化直到第19天。数据以平均值±sem的形式提供。*p<0.05(非配对学生t检验)。c)向小鼠静脉内注射2.106个标记有cfse的ot1细胞。3h后,腹膜内注射5.106个hek细胞wt或由质粒球蛋白-内含子-sl8或球蛋白-外显子-sl8或ova转染的hek细胞。3天后,收集来自淋巴结和脾脏的细胞,并且分析了cd8细胞中的cfse表达。点图表示在不同小鼠中获得的结果。图2:所有ptp:用于癌症疫苗的肽的来源。用125μg(ptpx1)、64μg(ptpx1/2)、32μg(ptpx1/4)ptp或用8μg(siinfekl1/25)在cpg+聚i:c(阴性对照)中乳化的siinfekl表位(mca-205-ova的阳性对照和mca-205wt细胞的阴性对照)向6只小鼠的组接种疫苗。15天后,在右侧腹用50.103个表达卵白蛋白的mca-205活细胞(a)和在左侧腹中用50.103个mca-205wt活细胞(b)皮下攻击小鼠。每7天测量各肿瘤细胞系的肿瘤生长。各线表示每组中6只小鼠的以面积(mm2)计的肿瘤尺寸。图3:来自肉瘤细胞系的特异性ptp:用于癌症疫苗的肽的来源。用125μg(ptp-hisx1)、64μg(ptp-hisx1/2)、32μg(ptp-hisx1/4)ptp或用8μg(siinfekl1/25)在cpg+聚i:c(阴性对照)中乳化的siinfekl表位(阳性对照)向6只小鼠的组接种疫苗。15天后,在右侧腹用50.103个表达卵白蛋白的mca-205活细胞(a)和在左侧腹中用50.103个mca-205wt活细胞(b)皮下攻击小鼠。每7天测量各肿瘤细胞系的肿瘤生长。各线表示每组中6只小鼠的以面积(mm2)计的肿瘤尺寸。图4:ptp加外来体:新型癌症疫苗。a)通过facs分析从mca205-球蛋白-内含子-sl8细胞纯化的外来体中cd9和cd81的表达。浅灰色为纯外来体,深灰色为wt外来体,并且黑色为球蛋白-内含子-sl8-外来体。b)左图:通过从mca205wt或mca205-球蛋白-内含子-sl8纯化的外来体脉冲处理bmdc(骨髓树突状细胞)。将其收集并且与ot1细胞一起培养。进行elisa来检测il-2m。数据以平均值±sem的形式提供。右图:在bmdc不存在的情况下将外来体添加到ot1细胞中。通过elisa评估在至少18h后上清液中产生的mil-2的量。数据可以表示为平均值±sem。c)使用25d1抗体对mca205细胞和外来体进行的siinfekl表达的facs分析。左图,短划线为纯mca205细胞,浅灰色为wtmca205并且白色为表达球蛋白-内含子-sl8构建体的mca205细胞。右图,浅灰色为纯外来体,深灰色为从mca205细胞纯化的外来体并且黑色为从表达球蛋白-内含子-sl8构建体的mca205细胞纯化的外来体。d)用64μg(ptp-hisx1/2)、32μg(ptp-hisx1/4)肿瘤来源ptp或用64μg(ptp-hisx1/2)、32μg(ptp-hisx1/4)肿瘤来源ptp加15μg含有ptp的肿瘤来源外来体或作为阳性对照的8μg(siin1/25)在cpg+聚i:c中乳化的siinfekl表位向6只小鼠的组接种疫苗。15天后,在右侧腹中用50.103个表达卵白蛋白的mca-205活细胞皮下攻击小鼠。每7天测量各肿瘤细胞系的肿瘤生长。各线表示每组中6只小鼠的以面积(mm2)计的肿瘤尺寸。图5:添加cd4表位以改善ptp癌症疫苗。用64μg(ptp-hisx1/2)肿瘤来源ptp或用64μg(ptp-hisx1/2)肿瘤来源ptp加1mg纯化的卵白蛋白或作为阳性对照的8μg(siin1/25)在cpg+聚i:c中乳化的siinfekl表位向6只小鼠的组接种疫苗。15天后,在右侧腹用50.103个表达卵白蛋白的mca-205活细胞和在左侧腹中用50.103个mca-205wt活细胞皮下攻击小鼠。每7天测量各肿瘤细胞系的肿瘤生长。各线表示每组中6只小鼠的以面积(mm2)计的肿瘤尺寸。图6:说明卵白蛋白cdna和β-球蛋白基因的内含子序列中sl8抗原表位的不同位置的图。图7:来自黑素瘤细胞系的特异性ptp:用于癌症疫苗的肽的来源。用32μg或16μgptp-his或用8μg在cpg+聚i:c(阴性对照)中乳化的siinfekl表位(阳性对照)向6只小鼠的组接种疫苗。十五天后,在右侧腹用30.103个表达卵白蛋白的b16f10活细胞以及基质胶(a)和在左侧腹中用30.103个b16f10wt活细胞(b)皮下攻击小鼠。每3-4天测量各肿瘤细胞系的肿瘤生长。各线表示每组中6只小鼠的以面积(mm2)计的平均肿瘤尺寸。图8:来自黑素瘤细胞系的ptp加外来体。用16μgptp-his、用15μg来源于b16f10细胞的外来体或用在cpg+聚i:c(阴性对照)中乳化的16μgptp-his连同15μg外来体向6只小鼠的组接种疫苗。十五天后,在右侧腹用30.103个表达卵白蛋白的b16f10活细胞以及基质胶(a)和在左侧腹中用30.103个b16f10wt活细胞(b)皮下攻击小鼠。每3-4天测量各肿瘤细胞系的肿瘤生长。各线表示每组中6只小鼠的以面积(mm2)计的平均肿瘤尺寸。图9:来自黑素瘤细胞系的ptp加黑素体。a)通过从b16f10-球蛋白-内含子-sl8细胞纯化的黑素体脉冲处理bmdc。接着将bmdc与sl8特异性cd8+t细胞杂交瘤(b3z)一起共培养16h,并通过测量β-半乳糖苷酶来估计t细胞活化。b和c)用32μgptp-his或用30μg在cpg+聚i:c(阴性对照)中乳化的来源于b16f10细胞的黑素体向6只小鼠的组接种疫苗。十五天后,在右侧腹用30.103个表达卵白蛋白的b16f10活细胞以及基质胶(b)和在左侧腹中用30.103个b16f10wt活细胞(c)皮下攻击小鼠。每3-4天测量各肿瘤细胞系的肿瘤生长。各线表示每组中6只小鼠的以面积(mm2)计的平均肿瘤尺寸。在本申请中,各种参考文献描述了本发明所属领域的现有技术。这些参考文献的公开内容在此通过引用并入本公开中。本发明的其它特征和优点在以下实验部分(参考图1至6)中给出,所述实验部分应被视为说明性的而非限制本申请的范围。实验部分实施例1-先行翻译产物(ptp)与外来体的组合:一种新型癌症疫苗。材料与方法细胞培养在37℃和5%co2下,在标准条件下,将mca205小鼠肉瘤细胞系在rpmi1640培养基(lifetechnologies)中在1%谷氨酰胺、1%丙酮酸、1%非必需氨基酸和10%fbs(lifetechnologies)存在下培养。在37℃和5%co2下,将b16f10(来自c57bl/6j小鼠的同种同基因细胞)在含有10%fcs、2mml-谷氨酰胺和100iu/ml青霉素/链霉素的dmem中培养。根据制造商纯化ptp的方案(ozyme),使用jetprime用yfp-球蛋白-内含子-sl8-his质粒转染mca205和b16f10细胞。对于肿瘤排斥实验,制备稳定的mca205-ova和稳定的b16f10-ova细胞。在标准条件下在rpmi1640中培养稳定mca205-ova。在标准条件下在dmem中培养稳定表达卵清蛋白的稳定b16f10-ova细胞。动物研究:c57bl/6j小鼠获自harlan。ot1c57bl/6j小鼠由cerfe(c.daviaud)慷慨提供,并在gustaveroussy动物设施中繁殖。7周时,在c57bl/6j小鼠右侧腹皮下接种0.1×106个mca205或b16f10肿瘤细胞。对于mca205,当肿瘤达到约20mm2的大小时,向小鼠静脉内注射0.1×106个ot1细胞。在b16f10模型中,在肿瘤接种后3天静脉内接种0.2×106个ot1细胞。所有动物实验均遵照法国和欧洲法律和法规进行。ptp-his纯化将转染的mca205或b16f10肿瘤细胞在10ml6m盐酸胍、0.01mtris/hcl(ph8.0)、5mm咪唑和10mmβ-巯基乙醇中超声处理。接着,温育裂解物并与ni2+-nta-琼脂糖珠粒(qiagen)一起在室温下旋转4h。在室温下将珠粒依次用8ml以下缓冲液中的每一种洗涤5min:6m盐酸胍、0.01mtris/hcl(ph8.0)和10mmβ-巯基乙醇;6m尿素、0.01mtris/hcl(ph8.0)和10mmβ-巯基乙醇;6m尿素、0.01mtris/hcl(ph6.8)、10mmβ-巯基乙醇和0.2%tritonx-100;6m尿素、0.01mtris/hcl(ph6.8)和10mmβ-巯基乙醇;6m尿素,0.01mtris/hcl(ph6.8)、10mmβ-巯基乙醇和0.1%tritonx-100。接着通过在室温下在400mm咪唑、0.15mtris/hcl(ph6.8)、30%甘油,0.72mβ-巯基乙醇和5%sds中温育珠粒20min来洗脱ptp。在室温下使用透析管mwco0.5kd(vwr)将洗脱液在pbs中透析过夜。最后,通过bradford分析(thermofisher)定量洗脱液。所有ptp纯化使mca205肿瘤细胞裂解,接着在10ml6m盐酸胍、0.01mtris/hcl(ph8.0)、5mm咪唑和10mmβ-巯基乙醇中超声处理。纯化裂解产物并使用3kda离心过滤器(merckmillipore)浓缩多肽。将该柱在3000g下离心90min.,这可纯化ptp定义中的小多肽。在室温下使用透析管mwco0.5kd(vwr)将下半部分在pbs中透析过夜。最后,通过bradford分析(thermofisher)定量洗脱液。从实体肿瘤中提取肽通过用0.22μm细胞滤过器破碎材料在冰上进行实体肿瘤分解。通过添加组织重量10倍的1×sds缓冲液(0.125mtris-hcl(ph6.8)、2%十二烷基硫酸钠、10%甘油、5%2-巯基乙醇)进行增溶。将分解的组织在70℃下温育并以1400rpm摇动10min。接着,将其在室温下在13200g下离心5min,以沉积并去除固体组织。使用d-tubetm透析器(merckmillipore)纯化并浓缩分子量截留为5kda的肽。在3000g下离心1小时30分钟。最后,使用bca蛋白质分析试剂盒(pierce)测量肽浓度。疫苗接种根据以下组制备mca205细胞的疫苗:ptp-hisx1(128μg)、ptp-hisx1/2(64μg)、ptp-hisx1/4(32μg)-/+外来体、外来体(从mca转染细胞纯化(15μg))、ptp-hisx1/2(64μg)-/+(1mg/50μl/小鼠)卵白蛋白(calbiochem)、所有ptpx1(128μg)、所有ptpx1/2(64μg)、所有ptpx1/4(32μg)、cpg(20μg)(invivogen)和聚(i:c)(50μg)(invivogen)、pbs(达300μl)。在注射前2小时制备疫苗并保持在冰上。在接种疫苗之前,用3%异氟烷麻醉c57bl/6小鼠。将疫苗皮下注射到腿(150μl/腿)和足垫(50μl/脚)中。两周后,皮下注射50×103个mca205肿瘤细胞(右侧腹)和mca205ova肿瘤细胞(左侧腹)。每周一次测量肿瘤尺寸直至其达到300mm2。根据以下组制备b16f10细胞的疫苗:32μg或16μgptp-his、外来体(从b16f10转染细胞纯化,15μg)、黑素体(从b16f10细胞纯化,30μg)或用8μgsiinfekl表位(阳性对照)、cpg(20μg)(invivogen)和聚(i:c)(50μg)(invivogen)。在注射前2小时制备疫苗并保持在冰上。在接种疫苗之前,用3%异氟烷麻醉c57bl/6小鼠。将疫苗皮下注射到腿(150μl/腿)和足垫(50μl/脚)中。两周后,皮下注射30×103个b16f10肿瘤细胞(右侧腹)和b16f10ova肿瘤细胞(左侧腹)。每周一次测量肿瘤尺寸直至其达到300mm2。结果先行翻译产物(ptp)在肿瘤排斥中的作用:在过去十年中,已经提出ptp和drip是内源性mhci类路径的肽的主要来源。为了精确定义ptp在介导特异性cd8+t细胞抗肿瘤免疫反应中的作用,发明人用两种不同的肿瘤模型接种了单独的c57bl/6小鼠:稳定表达其不同构建体的mca肉瘤模型和b16f10黑素瘤模型(参见图6)。对于mca模型,将105个肿瘤细胞皮下注射到c57bl/6小鼠中。接着使肿瘤生长至约20mm2。此时,将105个ova特异性tcr-转基因cd8+ot-1原初t细胞过继转移至小鼠。接着每两天监测并记录肿瘤生长。14天后,发明人观察到过继转移ot-1t细胞阻止了mca肿瘤的发展,这些肿瘤在球蛋白-内含子或球蛋白-外显子环境中独立地稳定表达siinfekl/sl8表位(图1a,下图和上图)。并且,正如预期的那样,过继转移ot-it细胞不能阻止sl8阴性mca肿瘤的生长(图1a,下图和上图),从而证实cd8+t细胞对肿瘤细胞系表达的抗原的特异性识别,并证实ptp在诱导抗肿瘤反应中的特异性作用。对于b16f10模型,将105个肿瘤细胞皮下注射到c57bl6小鼠中。接着,3天后,将105个ova特异性tcr-转基因ot-1原初细胞过继转移至小鼠。与mca模型类似,过继转移ot-it细胞一方面阻止在内含子或外显子序列中稳定表达siinfekl/sl8表位的b16f10肿瘤的发展(图1b,上图和下图),并且另一方面,不阻止sl8阴性b16f10肿瘤的生长(图1b,上图和下图),从而再次支持ptp诱导特异性抗肿瘤反应的观点。此外,为了最终得出结论ptp可以促进体内模型中的交叉引发(crosspriming),用不同构建体转染hek-293细胞并皮下注射到cd45.1同类系c57bl/6小鼠中,该小鼠在3小时前接受用cfse染色的ot-i原初cd8+t细胞。如果由外显子和/或内含子序列表达的ptp有助于交叉引发,则预期会观察到cfse荧光随时间减少,证实cd8+ot-it细胞的增殖。如图1c所示,接种3天后,与hek-293细胞仅用空载体转染并且cd8+ot-1t细胞在接种相同时间后没有增殖的阴性对照相比,ptp诱导了cd8+ot-it细胞分裂。由于hek-293细胞来源于人类,所以它们不能直接将来自ptp的抗原直接呈递至鼠cd8+ot-1t细胞。因此,cd8+t细胞的增殖只能通过ptp的交叉引发发生,从而支持肿瘤排斥结果。这些结果展示ptp可以通过促进特异性抗原肿瘤排斥而在体内诱导特异性免疫反应。此外,这些结果显示ptp除了用作内源路径的抗原肽的主要来源外,还可能是mhci类外源路径的外源肽的来源。肿瘤多肽:用于癌症疫苗的肽的来源与此同时,并且为了证实携带mhci类表位的多肽作为癌症疫苗的主要来源的特异性作用,发明人从wt肿瘤细胞系中纯化了ptp。出于这个目的,将mca205wt肿瘤细胞系裂解,并且将ptp定义中的所有5kda或小于5kda的多肽纯化并且用作小鼠的疫苗,如之前关于来自发明人的相关构建体的ptp所述。用不同浓度的ptp或用佐剂自身(阴性对照)向不同的6只小鼠的组接种疫苗。2周后,将表达或不表达卵白蛋白构建体的50.104个mca205肿瘤细胞系皮下注射到小鼠的右侧腹(mca-205球蛋白-内含子-sl8)和左侧腹(mca-205wt)中。发明人的数据指示,从肿瘤细胞系的细胞核区室纯化的5kda或小于5kda的多肽可以诱导相同肿瘤的缺陷,而与肿瘤是否表达发明人的特定模型表位无关(图2a和2b)。与此同时,纯化来自在小鼠中生长数周的实体肿瘤的多肽。将实体肿瘤分解,接着以5kda的截止值纯化含有ptp的多肽。将纯化的多肽用作小鼠的疫苗,两周后用从其纯化得到这些多肽的相同肿瘤细胞系攻击所述小鼠。发明人的数据指示,从实体肿瘤纯化的多肽可以诱导相同肿瘤的缺陷,而与肿瘤是否表达发明人的特定模型表位无关。这些实验表明由不同长度的肿瘤来源多肽组成的疫苗对肿瘤生长的特异性作用,并支持ptp可用作疫苗来引发特异性抗肿瘤t细胞反应的观点。ptp:用于癌症疫苗的肽的来源在这个研究中,在将从肉瘤mca205和黑素瘤b16f10细胞系纯化的ptp用作疫苗之前,通过质谱法对其进行分析,来更仔细地研究构成疫苗的不同多肽的性质。如表1所示,疫苗由不同长度的不同多肽组成。表1:来源于mca205细胞所产生的外来体的肽的质谱分析。突出显示了对应于衍生自内含子序列的siinfekl肽的肽。sl8表位是在细胞表面由ova特异性tcr-转基因原初cd8+ot-1t细胞识别的表位,并因此诱导特异性cd8+t细胞增殖和肿瘤排斥。在该研究的前述部分中,发明人研究了借助于特异性cd8+t细胞的对肿瘤细胞系的肿瘤排斥,所述肿瘤细胞系表达其ptp,而在该研究的这一部分中,发明人旨在展示用肿瘤来源ptp以预防方式接种疫苗的小鼠与未接种疫苗的小鼠相比显示肿瘤生长缺陷,从而支持ptp可以作为疫苗诱导肿瘤生长缺陷和特异性cd8+t细胞免疫抗肿瘤反应的假设。出于该目的,用不同浓度的ptp或用佐剂自身(阴性对照)或用在相同佐剂中乳化的sl8表位(阳性对照)向不同的6只小鼠的组接种疫苗。从之前用球蛋白-内含子-sl8-his构建体转染的小鼠肿瘤细胞系纯化这些ptp。2周后,将来自转染的mca205肿瘤细胞系的50.104个细胞皮下注射到小鼠的右侧腹中,所述转染的mca205肿瘤细胞系表达与纯化的ptp相同的ptp。在小鼠的左侧腹中,类似地接种50.104个野生型mca205肿瘤细胞。发明人的数据指示,ptp可以从表达ptp的肿瘤细胞系而非野生型(wt)肿瘤细胞系诱导肿瘤生长缺陷(图3a和3b),从而证实ptp疫苗的特异性抗肿瘤作用。所有这些实验都表明ptp对肿瘤生长的特异性作用,并支持ptp可以以预防和治疗方式用作小鼠的疫苗来引发特异性抗肿瘤t细胞反应的观点。ptp和外来体:新型癌症疫苗发明人最近展示,与全长蛋白质相比,ptp是mhci类交叉呈递路径的肽的更好来源。发明人现在报道ptp在储存于囊泡中时允许更好的交叉呈现。实际上,大多数细胞可释放的亚细胞部分在小于400nm时称为微泡或外来体(30-100nm)。遵循这一观点,发明人假设ptp转移是通过由供体细胞分泌并由骨髓树突状细胞(bmdc)内化的外来体介导的。来自mca205细胞系的外来体根据前述报道纯化。为了证实纯化的物质是外来体,进行了facs分析。图4a揭示了不同表面蛋白cd9和cd81的存在,所述cd9和cd81是外来体的常用标志物,从而证实来自不同细胞系的纯化微泡是外来体。接着,将这些外来体直接在bmdc上脉冲处理。图4b(左图)显示已吞噬来源于mca205肿瘤的外来体的bmdc能够活化cd8+ot-1t细胞。由于外来体的纯化部分来自mca肿瘤细胞系,即内源表达kb分子的细胞系,所以发明人想知道来自这种细胞系的外来体是否可以直接活化cd8+ot-1t细胞。将来源于mca的外来体直接在cd8+ot-1t细胞上脉冲处理。添加外来体后未观察到t细胞的活化(图4b,右图)。事实上,发明人通过fac分析使用抗kb抗体研究了mhci类kb分子的表达,正如预期的那样,他们可以在小鼠细胞系的细胞表面上检测到kb分子,并且同时,他们不能在外来体的细胞表面检测到kb分子(图4c),从而支持了mca外来体不能靠自身活化cd8+ot-1t细胞的事实。因此,疫苗设计的下一步是在基于ptp的癌症疫苗中包括从其纯化得到ptp的相同肿瘤细胞系的外来体。出于这个目的,发明人将来自mca-205-球蛋白-内含子-sl8的纯化ptp与来自相同肿瘤的外来体一起在佐剂中温育数小时。接着,在外来体(15μg)存在或不存在下用不同浓度的ptp或用佐剂自身(阴性对照)或用在相同佐剂中乳化的sl8表位(阳性对照)向不同的6只小鼠的组接种疫苗。发明人的数据指示,与仅由肿瘤来源ptp组成的疫苗(图4d,方形线)相比,由肿瘤来源ptp和肿瘤来源外来体组成的疫苗更好地从表达sl8表位作为ptp的肿瘤细胞系(mcaova肿瘤细胞)诱导肿瘤生长缺陷(图4d,交叉线)。添加cd4表位来改善ptp癌症疫苗从以上结果,发明人已经显示并入ptp中并在外来体中发现的mhci类肽在小鼠中诱导特异性抗肿瘤反应。然而,疫苗接种的主要目标,并且特别是在癌症治疗中的主要目标是避免其复发。并且为了避免这种复发,必须诱导长效免疫。众所周知,cd4+t细胞可以激发和延长特异性抗肿瘤cd8+t细胞的寿命,并进一步诱导肿瘤位点处的专职性抗原呈递细胞(papc)的累积。这种累积可以是有益的,因为与全长蛋白质相比,由肿瘤产生的ptp是papc呈递的mhci类路径的更好来源。出于所有这些原因,使用由ptp以及来自相同基因的全长蛋白质组成的疫苗。分别用单独的ptp或其与蛋白质卵白蛋白的组合或用佐剂自身(阴性对照)或用在相同佐剂中乳化的sl8表位(阳性对照)向不同的6只小鼠的组接种疫苗。发明人的数据指示,与仅由肿瘤来源ptp组成的疫苗(图5,方形线)相比,由肿瘤来源ptp与全长蛋白质的组合组成的疫苗更好地从表达sl8表位作为ptp的肿瘤细胞系(mcaova肿瘤细胞)诱导肿瘤生长缺陷(图5,交叉线),并且对于wt肿瘤细胞系的生长未能发现作用,从而证实ptp-全长蛋白质疫苗的特异性抗肿瘤作用,并且支持如下观点:为了获得针对转化细胞的更好的免疫反应,优选在疫苗中组合活化cd8+和cd4+t细胞的肽来诱导更好并且长效的抗肿瘤免疫反应。实施例2-针对黑素瘤的疫苗针对黑素瘤的基于ptp的疫苗:发明人在实施例1中显示,从肉瘤细胞系如mca205纯化的ptp可用作小鼠的疫苗来以预防方式引发特异性抗肿瘤t细胞反应。为了扩展发明人的ptp适用作抗癌症疫苗的观点,他们研究了其它类型的癌症。出于这个目的,发明人从黑素瘤细胞系如鼠b16f10细胞系纯化了ptp。接着,用不同浓度的ptp、用佐剂自身(阴性对照)或用在相同佐剂中乳化的sl8表位(阳性对照)向不同的6只小鼠的组接种疫苗。从先前用发明人的球蛋白-内含子-sl8-his构建体转染的小鼠b16f10肿瘤细胞系纯化这些ptp。2周后,将来自转染的b16f10肿瘤细胞系的50.104个细胞皮下注射到小鼠的右侧腹中,所述转染的b16f10肿瘤细胞系表达与纯化的ptp相同的ptp。在小鼠的左侧腹中,类似地接种50.104个野生型b16f10肿瘤细胞。发明人的数据指示,ptp可以从表达ptp的黑素瘤肿瘤细胞系而非野生型(wt)黑素瘤肿瘤细胞系诱导肿瘤生长缺陷,从而证实ptp疫苗的特异性抗肿瘤作用(图7a和7b)。所有这些实验都表明ptp对任何肿瘤生长亚型的特异性作用,并支持ptp可在预防和治疗策略中用作小鼠的疫苗来引发特异性抗肿瘤t细胞反应的观点。针对黑素瘤的基于ptp-外来体的疫苗:发明人先前已经报道肿瘤来源外来体含有ptp,并且这些外来体可以与ptp结合以用作癌症疫苗,所述ptp自身是从肿瘤细胞系纯化得到。根据前述报道纯化来自表达球蛋白-内含子-sl8构建体的b16f10细胞系的外来体。发明人将来自b16f10-球蛋白-内含子-sl8的纯化ptp与来自相同肿瘤的外来体一起在佐剂中温育数小时。接着,在外来体(15μg)存在或不存在下用不同浓度的ptp或用佐剂自身(阴性对照)或用在相同佐剂中乳化的sl8表位(阳性对照)向不同的6只小鼠的组接种疫苗。发明人的数据指示,与仅由肿瘤来源外来体组成的疫苗(图8a,圆形黑线)相比,由肿瘤来源ptp和肿瘤来源外来体组成的疫苗从表达sl8表位作为ptp的肿瘤细胞系(b16f10ova肿瘤细胞)诱导更好的肿瘤生长缺陷(图8a,方形线)。根据这些结果,与发明人用来源于mca205的外来体所见相比,来自黑素瘤细胞系的含有ptp的肿瘤来源外来体刺激弱的特异性免疫反应。事实上,肿瘤来源ptp与肿瘤来源外来体的组合更有效地诱导肿瘤生长缺陷,即使这种作用不足以诱导完全的肿瘤排斥。发明人的数据还指示,从黑素瘤细胞系纯化的ptp和外来体的组合可以从表达ptp的肿瘤细胞系而非野生型(wt)肿瘤细胞系诱导弱的肿瘤生长缺陷(比较图8a和8b),从而证实基于ptp-外来体的疫苗的特异性抗肿瘤作用。针对黑素瘤的基于ptp-黑素体的疫苗:由于来自黑素瘤细胞系的外来体诱导弱的肿瘤生长缺陷,所以发明人假设由黑素瘤细胞系释放的其它囊泡可以具有与肉瘤外来体对肿瘤生长的作用相同的作用。黑素瘤细胞系不仅能够分泌外来体,还能够分泌黑素体。事实上,黑素细胞专门产生黑色素,所述黑色素储存在称为黑素体的细胞器中(raposo和marks,2007)。黑素体是一种组织特异性溶酶体相关细胞器(raposo和marks,2007),基于形态学和着色水平分为两个主要成熟期(watabe,kushimoto等人,2005)。不成熟的(i期和ii期)黑素体缺乏色素并且位于中心细胞质中;其被称为“成熟前黑素体”。成熟的重度着色黑素体(iii期和iv期)或“成熟黑素体”在远端树突中占主导地位,所述远端树突是其分泌的主要位点。为了理解如下观点:ptp转移可以由黑素瘤供体细胞分泌并且由骨髓树突状细胞(bmdc)内化的黑素体介导,如发明人对于肉瘤外来体所报道,根据前述报道纯化来自b16f10细胞系的分泌的黑素体。接着,将这些黑素体直接在bmdc上脉冲处理。图9a显示吞噬来源于b16f10肿瘤的黑素体的bmdc能够活化b3z杂交瘤细胞系,所述bmdc特异性识别细胞表面的mhci类kb/siinfekl复合物。发明人接下来测试了发明人是否可以在这些分泌的黑素体内检测到相应的ptp。出于这个目的,发明人在b16f10细胞中表达了构建体,在所述构建体中将6xhis-标签紧邻sl8表位插入内含子中。接着,发明人使用镍琼脂糖珠粒从纯化和超声处理的分泌的iv期黑素体中富集ptp,并将这些级份进行lc-ms/ms质谱分析。表2显示携带或不携带sl8表位的不同肽片段。值得指出的是,需要富集步骤以产生足够浓度的内含子衍生ptp来通过ms分析检测。这与先前的观察结果一致,显示虽然ptp是内源路径的优异底物,但ptp仍是稀有产物。肽序列肽长度,a.a.肽来源leynynshnvyimadk(seq06)16yfp-球蛋白agytmvhltpeek(seq12)13yfp-球蛋白sampegyvqer(seq02)11yfp-球蛋白savtalwgk(seq13)9yfp-球蛋白vnvdevggealgr(seq01)13yfp-球蛋白dhmvllefvtaagitlgmdelyk(seq14)23yfp-球蛋白fegdtlvnr(seq03)9yfp-球蛋白geelftgvvpilveldgdvnghk(seq07)23yfp-球蛋白aevkfegdtlvnrielk(seq15)17yfp-球蛋白gidfkedgnilghk(seq16)14yfp-球蛋白tiffkddgnyk(seq17)11yfp-球蛋白siinfek(seq05)7鸡卵白蛋白fsvsgegegdatygk(seq04)15yfp-球蛋白savtalwgkvnvdevggealgr(seq18)22yfp-球蛋白kagytmvhltpeek(seq19)14yfp-球蛋白yqtslyk(seq20)7yfp-球蛋白fsvsgegegdatygkltlk(seq21)19yfp-球蛋白fegdtlvnrielk(seq22)13yfp-球蛋白aevkfegdtlvnr(seq23)13yfp-球蛋白表2:来源于由球蛋白-内含子-sl8构建体转染的b16f10细胞所产生的黑素体的肽的质谱分析。突出显示对应于衍生自内含子序列的siinfekl肽的肽。此外,发明人在本文上述基于ptp的癌症疫苗中包括来自b16f10黑素细胞的纯化的分泌的iv期黑素体。出于该目的,分别用30μg分泌的iv期黑素体、用16μg从b16f10-球蛋白-内含子-sl8纯化的ptp和用佐剂自身(阴性对照)向不同的6只小鼠的组进行疫苗接种。发明人的数据指示,与仅由肿瘤来源ptp组成的疫苗相比,由肿瘤来源iv期黑素体组成的疫苗从表达sl8表位作为ptp的肿瘤细胞系(b16f10ova肿瘤细胞)诱导更好的肿瘤生长缺陷(图9b)。发明人的数据还指示,黑素体可以从表达ptp的肿瘤细胞系而非野生型(wt)肿瘤细胞系诱导肿瘤生长缺陷(比较图9b和9c),从而证实含有黑素体的基于ptp的疫苗的特异性抗肿瘤作用。讨论如果疫苗的主要目标是降低转化细胞逃避宿主免疫系统的机会,则发明人在本研究中证实:i)早期由不同于产生全长蛋白质的规范事件的翻译事件产生的多肽(ptp)可以用作特异性和强健的癌症疫苗,ii)这些多肽和携带类似多肽的外来体的组合可以是作为癌症疫苗的更强大的组合,以触发针对转化细胞的广泛t细胞库,以及iii)对于长效免疫反应,需要cd8和cd4ptp的组合。已经在临床试验中测试了来源于mage-1蛋白的i类结合合成表位作为基于单一肽的疫苗。接着,之后使用针对不同癌症的其它短肽。然而,在所有这些研究中,使用单一的合成表位作为疫苗,预期结果都不如希望得一样好,因为其能够在黑素瘤患者中看到任何有益的临床反应。这些结果可以通过以下事实来解释:短肽可以直接结合多种类型的细胞,而不仅仅是可以活化特异性cd8+t细胞的papc。更糟糕的是,当短肽与mhci类分子结合到非专职性细胞时,其可能诱导耐受免疫反应。此外,由于这些肽很短,所以其可能具有任何三级结构,因此进行快速降解。出于所有这些原因,发明人的基于ptp的癌症疫苗似乎是比使用短肽更好的策略。不同的理由是i)发明人的ptp已显示由不同长度、长于6个氨基酸、优选至少7或8个氨基酸的肽组成,ii)它们已显示是内源以及外源mhci类路径的肽的主要来源,iii)它们需要被papc捕获并适当加工以到达mhci类路径并呈递在细胞表面,iv)它们可由mhci类表位以及mhcii类表位组成。如果疫苗的主要目标是诱导针对癌症的长效免疫应答,则最后一个理由非常重要。事实上,发明人正在设计一种癌症疫苗来避免任何类型癌症的任何复发。他们的疫苗是一种治疗性疫苗,其中ptp和外来体需要从已经患有癌症的患者中纯化。发明人的疫苗的目标是具有完全肿瘤排斥并且不会复发。出于这些目的,其疫苗需要基于细胞毒性cd8+t细胞的活化诱导快速免疫反应,但是为了长效反应,该疫苗还需要诱导记忆反应。在该特定情况下,记忆反应将基于cd4+t细胞的作用。事实上,当发明人从wt肿瘤细胞系纯化ptp时(图2a),他们观察到的肿瘤生长抑制优于他们仅使用特别地来自他们的工程化模型构建体的ptp的时候,在所述工程化模型构建体中仅使用一种mhci类表位(图3a)。解释是,他们相信在来自wt肿瘤细胞系的纯化ptp中,不仅纯化出含有mhci类表位的多肽,而且纯化出含有mhcii类表位的多肽。这一观察结果得到以下事实的支持:当他们将从他们的工程化构建体纯化的ptp与全长卵白蛋白混合时,这种组合疫苗的作用比他们仅使用ptp自身时有效得多(图5)。尽管cd4+t细胞与papc之间存在cd40-cd40l相互作用,但cd4+t细胞已显示出是维持记忆cd8+t细胞所必要的,这再次证实了papc在疫苗接种成功中的重要和特定作用。根据一系列报道,已经发现肿瘤来源外来体是通过作用于不同途径诱导肿瘤免疫逃避的免疫抑制剂,例如通过抑制dc的分化或通过负调节nk细胞,但是据报道它们还具有通过诱导特异性肿瘤免疫反应而产生的免疫刺激作用。已显示它们通常含有肿瘤抗原,因此被用作用于癌症疫苗的肿瘤抗原的新型来源。从发明人的研究结果来看,含有ptp的肿瘤-外来体刺激特异性免疫反应。事实上,肿瘤来源ptp和肿瘤来源外来体的组合与肿瘤来源ptp自身相比更有效地诱导肿瘤排斥(图4)。该结果可以通过以下事实来解释:发明人已经成功地从外来体纯化由他们的工程化构建体产生的ptp,并且外来体也可能含有来自肿瘤自身的mhcii类表位。根据一系列报道,黑素体可以从黑素细胞转移到角质形成细胞。事实上,一系列研究报道了例如黑素体是负责将黑色素从黑素细胞转移到邻近角质形成细胞的囊泡。但对于发明人而言更重要地,最近还显示黑素瘤细胞可以通过细胞间转移借助于分泌的黑素体获得mhcii类抗原。这里,发明人报道,不仅可以转移mhcii类表位而且可以转移mhci类表位。事实上,发明人发现分泌的黑素体含有ptp,所述ptp可以从黑素瘤细胞系转移至bmdc,从而活化特异性cd8+t细胞。此外,发明人还报道了黑素体可以是用于黑素瘤癌症疫苗的基剂。发明人显示,在已经接种黑素瘤细胞系的小鼠中注射的黑素体可以诱导重要的肿瘤生长缺陷,从而支持ptp与黑素体的组合可以用作合适的黑素瘤癌症疫苗的观点。考虑到适当的肿瘤免疫反应取决于特异性cd8细胞毒性t细胞的募集和活化,以及cd4+t细胞是这些过程所必需的事实,以及适当的抗cd8肿瘤免疫反应不能在抗肿瘤-cd4+t细胞不存在下建立长效t细胞记忆的事实,他们的结果显示了在基于ptp的黑素瘤癌症疫苗中使用黑素体的重要性,其中分泌的黑素体不仅含有mhcii类还含有mhci类表位。参考文献-apcher,s.,c.daskalogianni和r.fahraeus(2015)."pioneertranslationproductsasanalternativesourceformhc-iantigenicpeptides(先行翻译产物作为mhc-i抗原肽的可替选来源)."molimmunol.-apcher,s.,g.millot,c.daskalogianni,a.scherl,b.manoury和r.fahraeus(2013)."translationofpre-splicedrnasinthenuclearcompartmentgeneratespeptidesforthemhcclassipathway(核区室中预剪接rna的翻译产生用于mhci类途径的肽)."procnatlacadsciusa110(44):17951-17956.-clayton,a.,j.p.mitchell,j.court,s.linnane,m.d.mason和z.tabi(2008)."humantumor-derivedexosomesdown-modulatenkg2dexpression(人肿瘤衍生的外来体下调nkg2d表达)."jimmunol180(11):7249-7258.-garin,j.,diez,r.,kieffer,s.,dermine,j.f.,duclos,s.,gagnon,e.,sadoul,r.,rondeau,c.和desjardins,m.thephagosomeproteome:insightintophagosomefunctions(吞噬蛋白质组:了解吞噬体功能).jcellbiol152,165-80.,2001-raposo,g.和m.s.marks(2007)."melanosomes--darkorganellesenlightenendosomalmembranetransport(黑素体-黑暗的细胞器启发内体膜运输)."natrevmolcellbiol8(10):786-797.-raposo,g.和m.s.marks(2007)."melanosomes-darkorganellesenlightenendosomalmembranetransport黑素体-黑暗的细胞器启发内体膜运输)."natrevmolcellbiol8(10):786-797.-thery,c.,boussac,m.,veron,p.,ricciardi-castagnoli,p.,raposo,g.,garin,j.和amigorena,s.proteomicanalysisofdendriticcell-derivedexosomes:asecretedsubcellularcompartmentdistinctfromapoptoticvesicles(树突细胞衍生的外泌体的蛋白质组学分析:不同于凋亡小泡的分泌的亚细胞区室).jimmunol166,7309-18.,2001-thery,c.,regnault,a.,garin,j.,wolfers,j.,zitvogel,l.,ricciardi-castagnoli,p.,raposo,g.和amigorena,s.molecularcharacterizationofdendriticcell-derivedexosomes.selectiveaccumulationoftheheatshockproteinhsc73(树突细胞衍生的外来体的分子特征,选择性积累热休克蛋白hsc73).jcellbiol147,599-610,1999-theryc.,zitvogell.和amigorenas.exosomes:composition,biogenesisandfunction(外来体:组成、生物产生和功能.naturereview2(2002)569-valenti,r.,v.huber,p.filipazzi,l.pilla,g.sovena,a.villa,a.corbelli,s.fais,g.parmiani和rivoltini(2006)."humantumor-releasedmicrovesiclespromotethedifferentiationofmyeloidcellswithtransforminggrowthfactor-beta-mediatedsuppressiveactivityontlymphocytes(人肿瘤释放的微泡促进骨髓细胞的分化与转化生长因子-β介导的对t淋巴细胞的抑制活性)."cancerres66(18):9290-9298.-watabe,h.,t.kushimoto,j.c.valencia和v.j.hearing(2005)."isolationofmelanosomes.(黑素体的分离)"currprotoccellbiol第3章:第3单元(14)-whiteside,t.l.,m.mandapathil,m.szczepanski和m.szajnik(2011)."mechanismsoftumorescapefromtheimmunesystem:adenosine-producingtreg,exosomesandtumor-associatedtlrs(肿瘤逃离免疫系统的机制:产生腺苷的treg、外来体以及肿瘤相关的tlr)."bullcancer98(2):e25-31.-wolfers,j.,lozier,a.,raposo,g.,regnault,a.,thery,c.,masurier,c.,flament,c.,pouzieux,s.,faure,f.,tursz,t.,angevin,e.,amigorena,s.和zitvogel,l.tumor-derivedexosomesareasourceofsharedtumorrejectionantigensforctlcross-priming(肿瘤来源的外来体是ctl交叉引发的共享肿瘤排斥抗原的来源.natmed7,297-303.,2001-zitvogel,l.,regnault,a.,lozier,a.,wolfers,j.,flament,c.,tenza,d.,ricciardi-castagnoli,p.,raposo,g.和amigorena,s.eradicationofestablishedmurinetumorsusinganovelcell-freevaccine:dendriticcell-derivedexosomes(使用新型无细胞疫苗根除已建立的小鼠肿瘤:树突细胞衍生的外来体).natmed4,594-600,1998序列表<110>法国古士塔柏罗斯学院(institutgustaveroussy)<120>针对癌症的基于ptp的疫苗<130>b2195pc00<160>23<170>patentinversion3.5<210>1<211>13<212>prt<213>人工序列<220><223>ptp<400>1valasnvalaspgluvalglyglyglualaleuglyarg1510<210>2<211>11<212>prt<213>人工序列<220><223>ptp<400>2seralametprogluglytyrvalglngluarg1510<210>3<211>9<212>prt<213>人工序列<220><223>ptp<400>3phegluglyaspthrleuvalasnarg15<210>4<211>15<212>prt<213>人工序列<220><223>ptp<400>4pheservalserglygluglygluglyaspalathrtyrglylys151015<210>5<211>7<212>prt<213>人工序列<220><223>ptp<400>5serileileasnpheglulys15<210>6<211>16<212>prt<213>人工序列<220><223>ptp<400>6leuglutyrasntyrasnserhisasnvaltyrilemetalaasplys151015<210>7<211>23<212>prt<213>人工序列<220><223>ptp<400>7glyglugluleuphethrglyvalvalproileleuvalgluleuasp151015glyaspvalasnglyhislys20<210>8<211>4<212>prt<213>人工序列<220><223>ptp<400>8serileileasn1<210>9<211>1161<212>dna<213>原鸡(gallusgallus)<400>9atgggctccatcggtgcagcaagcatggaattttgttttgatgtattcaaggagctcaaa60gtccaccatgccaatgagaacatcttctactgccccattgccatcatgtcagctctagcc120atggtatacctgggtgcaaaagacagcaccaggacacaaataaataaggttgttcgcttt180gataaacttccaggattcggagacagtattgaagctcagtgtggcacatctgtaaacgtt240cactcttcacttagagacatcctcaaccaaatcaccaaaccaaatgatgtttattcgttc300agccttgccagtagactttatgctgaagagagatacccaatcctgccagaatacttgcag360tgtgtgaaggaactgtatagaggaggcttggaacctatcaactttcaaacagctgcagat420caagccagagagctcatcaattcctgggtagaaagtcagmcaaatggaattatcagaaat480gtccttcagccaagctccgtggattctcaaactgcaakggttctggttaatgccattgtc540ttcaaaggactgtgggagaaagcatttaaggatgaagacacacaagcaatgcctttcaga600gtgactgagcaagaaagcaaacctgtgcagatgatgtaccagattggtttatttagagtg660gcatcaatggcttctgagaaaatgaagatcctggagcttccatttgccagtgggacaatg720agcatgttggtgctgttgcctgatgaagtctcaggccttgagcagcttgagagtataatc780aactttgaaaaactgactgaatggaccagttctaatgttatggaagagaggaagatcaaa840gtgtacttccctcgcatgaagatggaggaaaaatacaacctcacatctgtcttaatggct900atgggcattactgacgtgtttagctcttcagccaatctgtctggcatctcctcagcagag960agcctgaagatatctcaagctgtccatgcagcacatgcagaaatcaatgaagcaggcaga1020gaggtggtagggtcagcagaggctggagtggatgctgcaagcgtctctgaagaatttagg1080gctgaccatccattcctcttctgtatcaagcacatcgcaaccaacgccgttctcttcttt1140ggcagatgtgtttccccttaa1161<210>10<211>1204<212>dna<213>智人(homosapiens)<400>10atggtgcacctgactgatgctgagaaggctgctgtctctggcctgtggggaaaggtgaac60tccgatgaagttggtggtgaggccctgggcaggttggtatccaggttacaaggcagctca120caagaagttgggtgcttggagacagaggtctgctttccagcaggcactaactttcagtgt180cccctgttatgtttccctttttaggctgctggttgtctacccttggacccagaggtactt240tgatagctttggagacctatcctctgcctctgctatcatgggtaatgccaaagtgaaggc300ccatggcaagaaagtgataactgcctttaacgagggcctgaatcacttggacagcctcaa360gggcacctttgccagcctcagtgagctccactgtgacaagctccatgtggatcctgagaa420cttcagggtgagtctgatgggcacctcgtttccttcccctggctattctgctcaaccttt480ctatcagaaggaaaggggaagcgattctagggagcagtctccatgactgtgtgtggagtg540ttgacaagagtttggatattttattctctactcagaatcgctgctccctctcactctgtt600ctgtgttgtcatttcctctttctttggtaagcttttaatttccagttgcattttactaaa660ttaattaagctggttatttacttcccatcctgatatcagcttcccctcctcctttcatcc720cagtccttctctctctcttctctctttctctaatcctttcctttccctcactccatttct780tcttctttgatctacttttgtttgtctttttaaatattgccttgtaacttgctcagagga840caaggaagatatgtccctgtttcttctcatagctctcaagaatagtagcataattggctt900ttatgccagggtgacaggggaagaatatattttacatataaattctgtgtgacataggat960cttataataatttgtcagtagtttaaggttgcaaacaaatgtctttgtaaataagcctgc1020agtatctggtatttttgctctacagttatgttgatggttcttccatcttcccacactcct1080gggcaatatgatcgtgattgtgctgggccaccacctgggcaaggatttcacccccgctgc1140acaggctgccttccagaaggtgatggctggagtggccactgccctggctcacaagtacca1200ctaa1204<210>11<211>8<212>prt<213>人工序列<220><223>对照肽<400>11serileileasnpheglulysleu15<210>12<211>13<212>prt<213>人工序列<220><223>ptp<400>12alaglytyrthrmetvalhisleuthrprogluglulys1510<210>13<211>9<212>prt<213>人工序列<220><223>ptp<400>13seralavalthralaleutrpglylys15<210>14<211>23<212>prt<213>人工序列<220><223>ptp<400>14asphismetvalleuleugluphevalthralaalaglyilethrleu151015glymetaspgluleutyrlys20<210>15<211>17<212>prt<213>人工序列<220><223>ptp<400>15alagluvallysphegluglyaspthrleuvalasnargilegluleu151015lys<210>16<211>14<212>prt<213>人工序列<220><223>ptp<400>16glyileaspphelysgluaspglyasnileleuglyhislys1510<210>17<211>11<212>prt<213>人工序列<220><223>ptp<400>17thrilephephelysaspaspglyasntyrlys1510<210>18<211>22<212>prt<213>人工序列<220><223>ptp<400>18seralavalthralaleutrpglylysvalasnvalaspgluvalgly151015glyglualaleuglyarg20<210>19<211>14<212>prt<213>人工序列<220><223>ptp<400>19lysalaglytyrthrmetvalhisleuthrprogluglulys1510<210>20<211>7<212>prt<213>人工序列<220><223>ptp<400>20tyrglnthrserleutyrlys15<210>21<211>19<212>prt<213>人工序列<220><223>ptp<400>21pheservalserglygluglygluglyaspalathrtyrglylysleu151015thrleulys<210>22<211>13<212>prt<213>人工序列<220><223>ptp<400>22phegluglyaspthrleuvalasnargilegluleulys1510<210>23<211>13<212>prt<213>人工序列<220><223>ptp<400>23alagluvallysphegluglyaspthrleuvalasnarg1510当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1