氟西汀在制备诱导听觉皮层可塑性的药物中的应用的制作方法
本发明属于生命科学/医学领域;本发明涉及提高脑感觉系统可塑性,维持脑健康的方案,具体涉及氟西汀在诱导听觉皮层可塑性中的应用。
背景技术:
大脑发育的早期,脑结构发育还未发生,神经元具有幼儿时期的突触结构和不成熟的胞外基质,此时的脑具有极高可塑性,我们把这个时期称为大脑发育的“关键期”。在听觉系统,普遍认为,声音的感知依赖于初级听皮层的正常发育。大脑发育“关键期”时,初级听皮层对声音的频率信息,时间信息,强度信息的编码区域受到环境声音的塑造,这种声音引起的皮层结构变化能持续至成年。“关键期”结束后,不注意的或是被动的声音环境已不足以引起听皮层的变化。可塑性调制需要调动更多的大脑功能,比如注意力,奖赏和新奇刺激。
多数研究认为,脑内兴奋性与抑制性之间的平衡是控制脑可塑性的关键。比如:将成年大鼠放入黑暗的环境中能诱导出类似于幼儿脑的视觉可塑性。我们发现,氟西汀能改变脑内兴奋性与抑制性之间的平衡,使听皮层出现类似“关键期”的可塑性。
所有的人都认可大脑对人体的重要性,但如果没有明显的眩晕疼痛,几乎没有人注意到自己的大脑是否健康。随着人类人口的增长和环境污染问题的不断发展,诸多影响大脑健康的不良因素成为了人类急需面对的问题。有研究显示,低浓度铅污染会损伤大脑听觉系统对方位信息的分辨能力。生活在机场旁的儿童长期受到噪音的污染,会引起大脑认知功能下降。通过学习和训练大脑可以部分改善发生变化的大脑功能,这是由于大脑能够受到环境经验进一步自我塑造,我们称之为脑可塑性。如何通过改变脑可塑性让大脑变得更加健康成为人类日益增长的迫切需求。
目前诱导听皮层可塑性可使用基底神经节电刺激或听觉行为训练。基底神经节电刺激可促进中脑基底区域的胆碱能释放,改变皮层兴奋性与抑制性比率,提高可塑性,但需要手术埋藏刺激电极,创伤较大。特定声刺激与动物的行为相联合并通过训练的方式同样可诱导听皮层的可塑性变化,但需花费大量的时间(zhouetal.,2010;zhangetal.,2013;zhuetal.,2016;liuetal.,2018)。
氟西汀(fluoxetine,flx)是一类选择性五羟色胺再摄取抑制剂,这类药物可以选择性的抑制突触前膜对五羟色胺(5-ht)的回收,调节脑内五羟色胺能神经递质,改变大脑兴奋与抑制性比率。
技术实现要素:
为了克服现有技术中存在的缺陷,本发明利用氟西汀对脑兴奋性的调节作用,建立一种改变听觉皮层可塑性新策略。
“噪音理论”一直被认为控制着中枢神经系统的发育。关键期内,随着抑制性神经系统的逐渐发育,过多的兴奋性噪音被抑制性神经递质祛除,同时大脑可塑性伴随抑制性神经系统的发育完善降低至成年水平。因此,皮层兴奋性与抑制性的比率被认为是“重启”关键期可塑性的重要开关。本发明对慢性氟西汀用药后大鼠听皮层抑制性相关蛋白的检测发现,听皮层抑制性神经系统pv蛋白及其胞外基质pnn的表达水平降低,提示氟西汀能有效调节皮层兴奋性与抑制性之间的比率。对比幼年组动物,发现这种变化向幼年脑方向发展。
目前,能够改变听皮层兴奋性与抑制性比率的办法有很多,但是能够成功诱导听皮层的方法并不多。使用基底神经节刺激促进胆碱能的释放,能有效提高听皮层可塑性。这种方法需要在基底神经节部位埋藏一个刺激电极,使用电流才能驱动胆碱能递质的释放,因而对大脑创伤极大。同样的,白噪音同样可以在成年期诱导可塑性,但是长时间的噪音暴露会引起动物不适。氟西汀本身是临床抗癫痫用药,给予溶解于动物的饮用水的氟西汀即可引起皮层兴奋性与抑制性比率的变化,诱导出类似关键期的可塑性。药品来源方便,操作简单,实用化程度高。
本发明提出了氟西汀在制备诱导听觉皮层可塑性的药物中的应用。
大脑在发育关键期时,非常容易受到环境因素的影响,这是由于此时的大脑具有非常高的可塑性。关键期之后的大脑可塑性变得很低,提高大脑的可塑性,接近于幼年时期的程度,本发明中称之为“诱导”。
本发明提出了氟西汀在制备改善、提高听觉皮层可塑性的药物中的应用。
本发明还提出了氟西汀在制备预防和/或治疗和/或改善听觉障碍性疾病或听觉功能异常的药物中的应用。
其中,所述听觉障碍性疾病包括老龄化引起的听力减退;重金属,药物或噪音(如中等强度噪音引起的听觉神经系统问题)等环境引起的听觉异常;由遗传因素引起的神经性耳聋;认知系统病变引起的听感觉异常等等均属于本发明权利要求的保护范围。
其中,所述氟西汀改变听皮层神经元频率选择特性,和/或增大选择性神经元数量,和/或增大选择性神经元表达面积。
其中,所述氟西汀改变听皮层频率拓扑结构。
其中,所述氟西汀弱化听皮层抑制性神经系统功能。
其中,所述氟西汀改变大鼠听皮层抑制性神经递质系统相关蛋白表达,促进胞外基质pnn类蛋白向幼年方向发展。
本发明还提出了一种改善、提高听觉皮层可塑性的药物组合物或试剂盒,所述药物组合物或试剂盒包括有效量的氟西汀;药物组合物或试剂盒说明书(包括氟西汀用药剂量、用药剂量与用药周期)。
本发明还提出了氟西汀联合感觉训练、丰富环境在制备改善、提高听觉皮层可塑性的药物中的应用。
本发明还提出了所述氟西汀联合感觉训练、丰富声环境在制备改善、提高听觉皮层可塑性的药物中、在制备预防和/或治疗和/或改善听觉障碍性疾病或听觉功能异常的药物中的应用。其中,丰富环境指的是动物的饲养环境空间大,内置物体丰富而新奇,成员较多,不仅提供多感官刺激和运动的机会,而且赋予了相互间社交行为可能。丰富声环境是指将声音刺激作为一种新的元素加入传统意义的丰富环境当中。丰富环境与特定声音刺激组合,已经证明能够改变大鼠的听觉皮层频率调谐能力。
本发明还提出了一种改善、提高听觉皮层可塑性的方法,所述方法包括配制浓度为200mg/l的氟西汀,溶解于动物的日常饮水中,给予有需要的个体4周时间。
优选地,所述氟西汀的浓度为200mg/l。
本发明中使用200mg/l浓度氟西汀溶液连续喂食大鼠4周,利用在体电生理技术验证了氟西汀药物能够诱导听皮层类似关键期可塑性。
本发明还提出了一种基于听皮层频率地形图检测听皮层可塑性的方法,在使用慢性氟西汀用药的同时给与7khz纯音刺激,采用电生理技术,检测待测对象(如大鼠)听皮层频率地形图,基于检测获得的所述待测对象听皮层频率地形图,判断氟西汀是否明显增大了7khz选择性神经元数量与表达面积。
在使用慢性氟西汀用药的第4周,同时给与7khz纯音刺激;采用电生理技术,绘制待测对象听皮层频率地形图;同时设置对照组;检测待测对象7khz选择性神经元数量、代表区面积。在一具体实施方式中,本发明在使用慢性氟西汀用药的第4周,同时给与7khz纯音刺激;采用电生理技术,绘制大鼠听皮层频率地形图;对照正常动物,发现记录到的7khz选择性神经元数量比正常动物多,并且其代表区面积比正常动物大。由于纯音刺激并不能引起成年期动物的听皮层频率代表区扩张,氟西汀诱导的类似发育关键期可塑性在纯音刺激下能够被电生理技术检测到。这种策略常被应用于听皮层可塑性的检测中。
其中,所述氟西汀引起了pv中间神经元和胞外基质pnn的变化,慢性氟西汀用药可通过改变抑制性神经系统及胞外基质的变化来调节听皮层可塑性。
本发明还提出了一种通过慢性氟西汀用药改变抑制性神经系统及胞外基质的变化、调节听皮层可塑性的调制方法。
本发明还提出了一种改善、提高听觉皮层可塑性的系统,所述系统包括氟西汀药物、氟西汀给药剂量和给药周期。
在一具体实施方式中,本发明使用大鼠作为实验对象,参考2008年意大利科学家maffie实验室给出的适合剂量,将氟西汀粉末配制为浓度为0.2mg/l的氟西汀口服溶液(氟西汀采购自西格玛(sigma)公司,货号phr1394),喂食8周龄成年大鼠4周,发现氟西汀能够有效提高听觉皮层可塑性,通过分子生物学对脑内相关蛋白的检测,发现此时大脑的诸多生化指标,包括pv蛋白在听皮层的表达、pnn蛋白在皮层的表达向幼年脑发展。由此,本发明认为,氟西汀能诱导听觉皮层类似关键期的可塑性。
本发明的有益效果包括:(1)无创性,由于氟西汀已是临床常用的抗抑郁药物,其毒理药理研究基本完善,因此,将氟西汀用于诱导听觉皮层可塑性对动物本身不会造成伤害。(2)成本低,本发明是建立在对现阶段神经可塑性原理的基础上,创造性地提出了一种新型的对听觉皮层可塑性的诱导方案,无需任何复杂的实验仪器或设备。(3)应用前景广,本发明在诸多神经系统问题中都具有普遍适用性,使用氟西汀药物诱导听觉皮层可塑性,配合良好的听感觉环境,可以改善多种环境因素引起的听觉功能异常,比如低浓度铅暴露引起的听觉方位分辨异常,中等强度噪音引起的听觉神经系统问题等。
本发明中,涉及的相关术语,如下:
所述听觉皮层可塑性是指脑可塑是指大脑可以为环境和经验所修饰,具有在外界环境和经验的作用下塑造大脑结构和功能的能力,故听觉皮层可塑性,特指在听觉皮层发生的脑可塑性过程。
类似关键期的可塑性是指大脑在发育关键期时,非常容易受到环境因素的影响,这是由于此时的大脑具有非常高的可塑性。关键期之后的大脑可塑性变得很低,通过本发明上述方法提高成年期大脑可塑性,并发现这种可塑性的状态与关键期类似,所以称为类似关键期的可塑性。
所述“慢性氟西汀用药”是指长时期(4周)的氟西汀用药。
所述神经可塑性原理是指发生在大脑可塑性调节过程中的神经分子机制。比如:抑制性神经系统的发育成熟是引起关键期可塑性的降低的主要原因,那么改变兴奋性与抑制性之间的比率既有可能提高脑可塑性。
附图说明
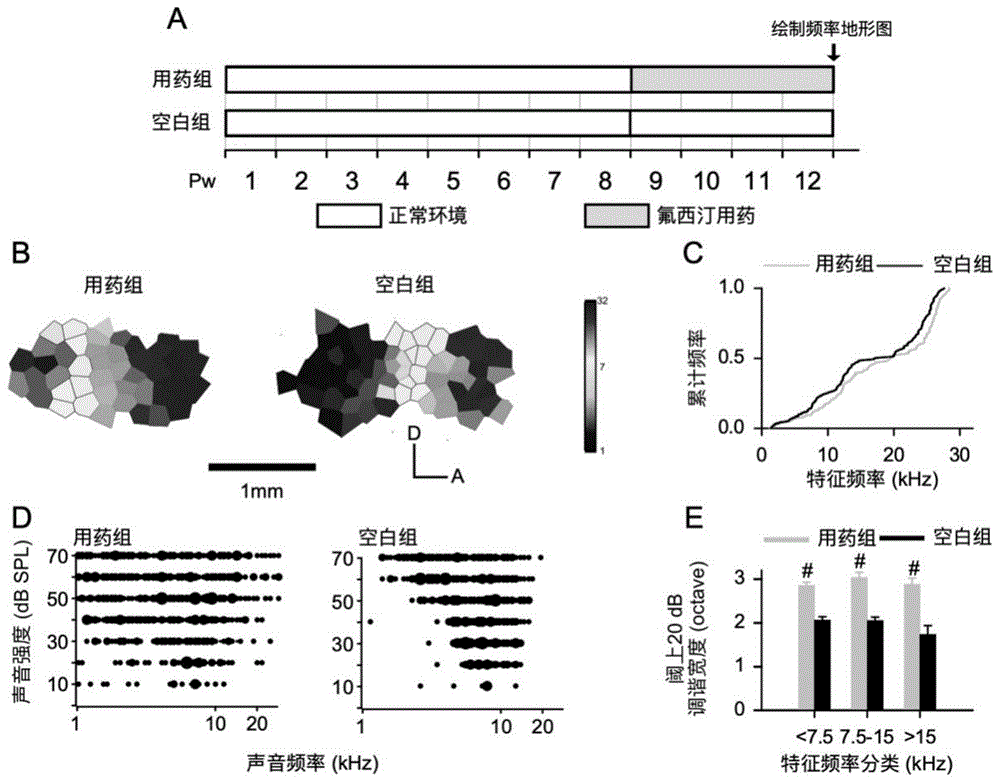
图1:慢性氟西汀用药后大鼠听皮层与空白组大鼠的对比。图1a,实验时间线。图1b,两组大鼠初级听皮层频率拓扑结构图。图1c,两组大鼠初级听皮层神经元特征频率分布累计函数曲线。图1d,两组大鼠听皮层神经元的频率感受野示例。图1e,两组大鼠听皮层神经元频率调谐曲线阈上20的调谐带宽(bw20)对比(数据表示为平均值±标准误,p<0.01用#表示)。
图2:慢性氟西汀用药大鼠同时给与7khz纯音暴露。图2a,实验时间线。
图2b,听皮层频率拓扑结构图;氟西汀用药后纯音暴露1周,大鼠听皮层7khz代表区域过量表达。图2c,听皮层频率分布散点图。图2d,听皮层各频率段神经元代表面积变化百分比(数据表示为平均值±标准误,p<0.05用*表示,p<0.001用+表示)。
图3:氟西汀用药后纯音暴露的效果。图3a,实验时间线。图3b,三组大鼠听皮层频率拓扑结构图(图中黑色粗线框选区域为7khz频率代表区域)。图3c,听皮层各频率代表面积变化百分比(数据表示为平均值±标准误,p<0.05表示为*)。
图4:氟西汀用药后大鼠听皮层小白蛋白(pv)和胞外基质蛋白(pnn)的表达及共定位分布情况。图4a,小白蛋白(pv)与胞外基质蛋白(pnn)免疫荧光标记示意图,其中左图为小白蛋白(pv),中图为胞外基质蛋白(pnn),右图为两类标记在空间上的相对位置。该图用以示意pv阳性神经元和pnn阳性神经元的形态。图4b,三组大鼠听皮层小白蛋白(pv)与胞外基质蛋白(pnn)表达分布示意图,其中红色标记为小白蛋白(pv),绿色标记为胞外基质蛋白(pnn)。图4c,小白蛋白(pv)与胞外基质蛋白(pnn)免疫荧光标记数量统计;上图统计了听皮层各层小白蛋白阳性标记的密度;中图统计了听皮层神经元胞外基质蛋白(pnn)阳性标记的密度,其中用药组大鼠听皮层胞外基质表达量在4-6层明显低于空白组大鼠;下图统计了小白蛋白(pv)与胞外基质蛋白(pnn)共标神经元密度,其中第四层用药组共标神经元密度略低于空白组大鼠(数据表示为平均值±标准误,p<0.05表示为*,p<0.01表示为#)。
其中,pv+是指听皮层表达pv的神经元;
pnn+是指表达pnn蛋白的神经元;
pv+pvv是指既表达pv又表达pnn的共标神经元;
l2/3是指皮层2,3细胞层;
l4是指皮层第4细胞层;
l5/6是指皮层第5,6细胞层;
pd11是指出生后11天的幼年组;
pv+cell是指pv阳性神经元,即表达pv蛋白的神经元;
pnn+cell是pnn阳性神经元,即表达pnn蛋白的神经元;
pv+/pnn+cell是指既表达pv又表达pnn的神经元;
ii+iii是指2,3细胞层;
iv是指第四细胞层;
v+vi是指5,6细胞层。
具体实施方式
结合以下具体实施例和附图,对本发明作进一步的详细说明,本发明的保护内容不局限于以下实施例。在不背离发明构思的精神和范围下,本领域技术人员能够想到的变化和优点都被包括在本发明中,并且以所附的权利要求书为保护范围。实施本发明的过程、条件、试剂、实验方法等,除以下专门提及的内容之外,均为本领域的普遍知识和公知常识,本发明没有特别限制内容。
实施例1、氟西汀改变听皮层神经元频率选择特性。
(1.1)实验动物与分组
(1.1.1)氟西汀溶液
将200mg氟西汀溶解于1l蒸馏水中,即可得到氟西汀溶液。
(1.1.2)实验动物
用药组:6只年龄8周的大鼠,使用氟西汀溶液喂食。
空白组:6只年龄8周的大鼠,使用正常蒸馏水喂食。
图1a为本实施例实验时间线,两组大鼠同时在动物房正常环境中饲养8周后,用药组大鼠喂食氟西汀药物4周,空白组大鼠继续使用正常水喂食4周,开展本实施例下述实验。
(1.2)在体细胞外记录技术绘制初级听皮层神经元特征频率分布
称量大鼠体重,使用2%戊巴比妥钠溶液麻醉,麻醉剂量为45mg/kg。等大鼠麻醉状态稳定,施气管插管术。用手指在大鼠颈部摸找喉软骨,用两个镊子分离肌肉,暴露气管,用手术刀切开气管,插入插管,手术线缝合。再暴露颞侧头骨,使用眼眶固定方式固定大鼠头部,打开电热毯,维持大鼠体温。然后施脑脊液引流术,用手术刀尖,轻轻戳破枕骨大孔上的粘膜,流出清亮液体,释放脑脊液。在枕骨大孔外放置一个吸水棉,稳定颅内压。最后用牙科电钻,打开颞叶骨窗,用细钨丝针挑开硬脑膜,生理盐水润湿,等待电生理记录。
本实施例使用tdt(tucker-davistechnologies)公司生产的tdtsystemiii记录系统,神经信号通过双通道金属电极记录(电阻2mω,通道间距240μm),记录过程由brainware软件控制实现。在体视显微镜观察下,缓慢操纵微操,让电极尖端碰到脑面,同时将深度记录仪归零。按照每秒3-4μm的速度操作微操将电极插入脑面。在350-600μm范围内寻找稳定放电。在找到稳定放电后,brainware软件中打开记录。声音刺激包括频率0-30khz,以0.1octave为梯度共49个频率的纯音,声强为0-70dbspl共8种声强,组合共392个声音刺激。同时记录下每一个声音刺激下神经元的反应数,软件绘制频率感受野,即可建立频率调谐曲线。使用ccd相机对记录位置进行拍照,在图形软件中标记记录位点。结合记录位点特征频率参数,在matlab软件中绘制听皮层神经元特征频率分布图。
(1.3)实验结果
听皮层是脑对听信息处理的高级中枢,听皮层结构和功能的改变影响着个体的听觉功能。一个正常大鼠的听皮层神经元特征频率分布具有头侧向尾侧,高频向低频的分布特点。
本发明检测了两组大鼠初级听皮层神经元特征频率分布,由初级听皮层频率拓扑结构图结果发现,用药组大鼠(氟西汀用药组)听皮层神经元分布与空白组大鼠相比,差异并不明显(图1b)。对所有记录到的听皮层神经元的特征频率进行统计,可以看出用药组大鼠(氟西汀用药组)和空白组大鼠相比,初级听皮层频率分布累计函数曲线并无差异(图1c,用药组vs空白组,p>0.9999,kolmogorov-smirnovtest),说明氟西汀并没有引起大鼠初级听皮层神经元特征频率分布的变化。
听皮层神经元对不同强度和不同频率组合声音的电反应,可反应听皮层神经元的功能。图1d为两组听皮层神经元频率感受野示意图,即图中黑色区域。频率感受野表示听皮层神经元对声音频率和声音强度的感受范围。对频率感受野进行描边即可得到频率调谐曲线,该曲线可得神经元的特征反应频(characteristicalfrequency,cf),最低反应阈值(minimumthreshold,mt)和调谐宽度(bandwidth,bw)等属性。比较两组大鼠听皮层神经元频率调谐曲线阈上20db的调谐宽度(bw20),发现用药组大鼠听皮层神经元bw20明显高于空白组大鼠(图1d)。本发明对记录到的神经元按照特征频率分类,发现在低,中,高三个频率段,用药组大鼠听皮层神经元调谐带宽bw20均显著宽于空白组大鼠(图1e,用药组vs空白组,cf<7.5khz,p<0.0001;7.5khz<cf<15khz,p<0.0001;cf>15khz,p<0.0001,t-testbofforonicorrection)。该实验结果说明,氟西汀用药后,大鼠的听皮层神经元频率选择特性降低。由于皮层兴奋性与抑制性的比例是控制大脑可塑性的关键,本实施例通过给与大鼠氟西汀4周后,大鼠听皮层神经元频率调谐曲线显著变宽,这说明听皮层兴奋性与抑制性之间比率升高,表明本实施例成功诱导了皮层可塑性。
实施例2、氟西汀改变听皮层频率拓扑结构图
(2.1)实验动物和分组与实施例1相同
(2.1.1)氟西汀溶液
将200mg氟西汀溶解于1l蒸馏水中,即可得到氟西汀溶液。
(2.1.2)实验动物
氟西汀+纯音暴露组(实验组):6只8周龄大鼠,使用氟西汀溶液喂食4周,在喂食第4周将大鼠置于纯音暴露箱中暴露1周,即为氟西汀+纯音暴露组,同时也是本实施例的实验组。
纯音暴露对照组:为了验证纯音暴露对成年大鼠听皮层的影响,设立纯音暴露对照组。将6只11周龄大鼠放入纯音暴露箱中暴露1周,为纯音暴露对照组。
氟西汀用药对照组:为了验证在没有纯音诱导的条件下,氟西汀喂食对皮层频率分布变化的影响,设立氟西汀用药对照组。将6只8周龄大鼠喂食氟西汀4周,即为氟西汀用药对照组。
空白组:6只12周龄的正常大鼠。
图2a为本实施例实验时间线,按照各组的喂食方式饲养造模,在大鼠12周龄时开展本实施例以下实验。
(2.2)声音暴露装置
声音暴露装置为一个自制的隔音箱,内置喇叭,自动灯光和通风系统。使用的声音为7khz纯音,一个纯音为50ms,15个纯音组成一串,时长1s,每串纯音间隔1s。纯音文件使用adobeauditioncc制作,使用dvd刻录机刻录在光盘中。用dvd播放机播放,经功率放大器,由喇叭发出。声音强度为65dbspl.
(2.3)实验结果
大鼠听皮层特征频率神经元的排列具有头侧向尾侧,高频向低频的分布方式。通常把听皮层的这种编码方式称作听皮层的频率筑构。听皮层的频率分布图随可塑性的改变发生变化,使用7khz纯音刺激处于脑发育“关键期”的大鼠24h,听皮层7khz特征神经元的表达数量显著提高。这种对听皮层频率分布图的检测被广泛应用于听觉可塑性研究当中(kilgardmpandmerzenichmm.,1998)。
本实施例使用在体电生理技术对各组大鼠听皮层频率分布做图,结果发现,与空白组大鼠相比,氟西汀+纯音暴露组(实验组)听皮层中神经元对于7khz纯音的表达面积显著增大(图2b)。对各组大鼠记录到的神经元位置信息进行标准化做图,得到听皮层频率分布散点图,发现氟西汀+纯音暴露组(实验组)记录到的7khz神经元数量显著增多(图2c)。计算各组大鼠听皮层各频率段神经元代表面积变化百分比(用每组测量到的7khz平均表达面积减去空白组7khz平均表达面积),由图2d可见,氟西汀+纯音暴露组(实验组)大鼠听皮层7khz表达面积显著高于其余三组(氟西汀+纯音暴露组vs空白组;p=0.0015,t-testbofforonicorrection)(图2d)。以上结果提示在纯音诱导的条件下,氟西汀提高了听皮层功能的可塑性;而在没有纯音诱导的条件下,氟西汀喂食并不会引起听皮层频率分布变化;同样的,纯音暴露对成年大鼠听皮层不具有影响。
实施例3为了进一步验证氟西汀用药对听皮层功能可塑性的影响
(3.1)实验动物与分组
(3.1.1)氟西汀溶液
将200mg氟西汀溶解于1l蒸馏水中,即可得到氟西汀溶液。
(3.1.2)实验动物
用药后纯音暴露组:6只8周龄大鼠,使用氟西汀溶液喂食4周,然后置于7khz纯音暴露箱中进行纯音暴露。
用药后7周纯音暴露组:6只8周龄大鼠,使用氟西汀溶液喂食4周,停药7周再置于7khz纯音暴露箱中进行纯音暴露。
空白组:6只12周龄的正常大鼠。
图3a为本实施例实验时间线,按照各组的喂食方式饲养造模,开展本实施例以下实验。
(3.2)实验过程及结果
对三组大鼠绘制频率地形图,得到三组大鼠听皮层频率拓扑结构图,将两组大鼠(用药后纯音暴露组、用药后7周纯音暴露组)频率地形图与空白组对照,检测氟西汀增强听皮层可塑性的效果,由图3b可看出,氟西汀停药后马上给与纯音暴露(即用药后纯音暴露组)仍可引起7khz纯音表达面积增大。对所有记录到的听皮层神经元分布面积进行了进一步统计发现,氟西汀用药停药后马上给与纯音暴露(即用药后纯音暴露组)仍可显著影响听皮层频率构筑(图3c,用药后纯音暴露组vs空白组,p=0.034,t-testbofforonicorrection)。而氟西汀用药4周停药7周后才给与纯音(即用药后7周纯音暴露组)并不能引起听皮层7khz纯音表达面积的变化。
将11-13日龄大鼠置于7khz纯音环境中暴露一天,就会引起个体成年后听皮层7khz相关神经元表达量上升,相关频率表达面积增大。听皮层神经元的频率筑构同样会因兴奋性与抑制性平衡的改变而发生变化;有研究显示,抑制性中间神经元被认为控制着皮层频率筑构的完整性和频率代表区之间的边界(casanovaetal.,2002)。
此外,使用细胞外多通道记录技术检测听皮层频率拓扑分布图。结果发现,相对于空白组,用药后纯音暴露组经过纯音暴露后,听皮层7khz神经元表达面积发生明显扩张;而相对于空白组,用药后7周纯音暴露组在用药4周停药7周后再放入7khz纯音环境,就不会观察到类似的现象(图3b)。
实施例4、氟西汀改变大鼠听皮层抑制性神经递质系统相关蛋白表达变化
(4.1)实验动物与分组
(4.1.1)氟西汀溶液
将200mg氟西汀溶解于1l蒸馏水中,即可得到氟西汀溶液。
(4.1.2)实验动物
用药组:6只年龄8周的大鼠,在动物房使用氟西汀溶液喂食4周,开展本实施例以下实验。
空白组:6只年龄8周的大鼠,在动物房使用正常蒸馏水喂食4周,开展本实施例以下实验。
幼年组:6只年龄11天的幼年大鼠。为了解听皮层内相关蛋白质表达与幼年大鼠之间的关系,本实施例加入了一组由6只出生11天的大鼠组成的幼年组作为对照,即幼年组。
(4.2)免疫荧光(小白蛋白(pv)和胞外基质蛋白(pnn)免疫荧光双标染色方法)
为进一步确定氟西汀对听皮层内生物化学指标的影响,本发明采用免疫印迹法特异性标记听皮层小白蛋白(pv)和胞外基质蛋白(pnn)。
对大鼠利用心脏灌流的方法,先使用生理盐水进行灌流,再使用4%多聚甲醛灌注固定。然后剪下鼠头,取出鼠脑,浸泡在4%多聚甲醛中6小时,后梯度脱水。梯度脱水后用包埋剂包埋,放入-80℃冰箱快速冷冻保存。切片时,提前两小时转入冰冻切片机(leica),设置-20℃工作温度,设定切片厚度30μm,切片位置参照脑图谱定位,进行切片。将脑片浸泡在原位杂交保护液中,放入-20℃冰箱中保存备用。
把脑片从原位杂交保护液中取出,用漂洗板在pbs中漂洗3次,每次10min,共30min。用1ml或2mlep管,配置封闭液10%羊血清和tritonpbs,分装封闭液,每一个脑片浸泡在100μl封闭液内,37℃漂片封闭1.5h。配置鼠源anti-pv(sigma)1:1000,anti-wfa(vectorlabs)1:200,1%羊血清和tritonpbs。将1抗分装,每张脑片100μl浸泡,放入烘箱37℃孵育6h,后转移到4℃冰箱中过夜保存。次日,将脑片挑进漂洗板中,pbs漂洗40min。待漂洗快结束时,配置荧光二抗(546nm,mouse,sigma)1:300,1%羊血清和tritonpbs。分装二抗,每张脑片100μl,将脑片放入二抗中37℃孵育2h。取出后把脑片放入漂洗板中,pbs漂洗45min。最后贴片,滴加带有dapi的封片剂使脑片保持潮湿。盖上盖玻片并用指甲油密封边缘,干燥4℃保存。
(4.3)实验结果
本实施例使用免疫荧光技术标记了神经元相关蛋白pv、pnn(图4a)。从形态来看,pv标记下的神经元形态是一个实心的纺锤体(图4a左图),pnn标记下的神经元包绕在pv标记之外(图4a右图),呈放射状形态(图4a中图)。直观的看,相较于空白组,用药组大鼠听皮层pv和pnn蛋白的标记数量明显低于空白组大鼠(图4b)。对两类标记蛋白进行计数统计发现用药组大鼠胞外基质蛋白(pnn)在听皮层第4-6层表达显著低于空白组大鼠(图4c,用药组vs空白组,layeriv:p<0.01;layerv/vi:p<0.05,t-testbofforonicorrection)。
同时本实施例加入幼年组大鼠与用药组和空白组大鼠相比较,发现用药组大鼠在氟西汀用药后,其听皮层胞外基质(pnn)的表达趋势向幼年方向发展。
由此可见,听皮层抑制性中间神经元相关蛋白表达量降低是氟西汀提高听皮层可塑性的诱因。
“关键期”大脑结构的发育常伴随抑制性神经元由丘脑向皮层的迁移与锚定,这同时被认为是皮层神经网络成熟的重要节点(blueandparnavelas1983)。有研究显示,抑制性中间神经元髓鞘的形成与神经元网络的功能性成熟具有密切联系(ishiguroetal.1991;brückneretal.2000)。这样的现象已经在中枢神经系统中多个部位得到证实,包括视觉皮层,脊髓背角运动神经元,桶皮层和小脑(dityatevetal.2007;mcraeetal.2007)。胞外基质(pnns)是一类围绕在中枢神经系统多数神经元胞体外的一类网状结构,由胞外基质分子组成(ecm,extracellularmatrix),包括透明质烷(ha),链接蛋白,软骨素表面蛋白(cspgs,chondroitinsulfateproteo-glycans)和黏蛋白(tn-r,tenascin-r),在中枢神经系统神经元胞体周围及近端树突外大量表达(
听皮层抑制性中间神经元多数表达pv,pv阳性中间神经元通过改变细胞的内在属性和突触的输出属性参与调节经验依赖的可塑性(devillers-sidanietal.,2008;zhouetal.,2011;ouelletanddevillers-sidani,2014)。神经可塑性的诱导和表达依赖于神经元与神经元之间和神经元与周围环境的相互联系。pv阳性中间神经元周围常包裹有称为pnns的胞外基质,其中包含有硫酸软骨素蛋白多糖(cspgs)(deepaetal.,2006)。pv阳性中间神经元和pnns的形成与成熟都发生在关键期关闭的时期(takesianandhensch,2013)。这说明pnns通过促进突触稳定性与突触重排过程参与调解关键期可塑性(pizzorussoetal.,2002)。更直接的证据证明,药理学方法处理硫酸软骨素蛋白多糖(cspgs)可增强皮层可塑性和学习能力(gogollaetal.,2009)。因此,本发明同时考察了pv阳性中间神经元和胞外基质pnns的表达。结果发现,氟西汀用药组表达在pv阳性中间神经元细胞周围的胞外基质pnns同样降低。结果提示,氟西汀处理引起了听皮层pv阳性中间神经元的活动变化,进一步提高了听皮层的可塑性。
本发明的结果证实了氟西汀针对情绪疾病上的病理生理应用之外的一种潜在应用。氟西汀能够为一些特殊行业工作者提供脑感觉功能增强的解决方案,比如配合训练可以提高潜艇官兵的听感觉功能,更好的分辨声呐回波。同样的,使用氟西汀诱导的脑听皮层可塑性有可能改善环境引起的大脑听感觉功能退化。
以上所述仅为本发明的较佳实施例而已,并不用以限制本发明,凡在本发明的精神和原则之内所作的任何修改、等同替换和改进等,均应包含在本发明的保护范围之内。
- 还没有人留言评论。精彩留言会获得点赞!