一种剪接辅助因子AtU2AF65b在植物花期调控中的应用的制作方法
本发明涉及分子生物学和生物技术领域,提供了一种剪接辅助因子atu2af65b在植物花期调控中的应用。
背景技术:
开花是高等植物由营养生长向生殖生长转换的一个重要生长发育过程。影响植物开花的因素有很多,其中光照(光周期途径)和温度(春化途径)是两个主要的外部因素,赤霉素(ga途径)和一些自主性因子(自主途径)是主要的内部因素,这些因素通过调控开花基因ft(floweringlocust)、fd(floweringlocusd)、soc1(suppressorofoverexpressionofconstans1)和fd(floweringlocusd)等多个基因的表达,在茎尖启动花发育过程,使茎尖分生组织向花(花絮)分生组织转化,实现对植物开花时间的精确调控(fornaraetal.,2010;ribonietal.,2013;ribonietal.,2016)。
细胞核中的基因转录形成的pre-mrna需要将内含子剪接后形成成熟的mrna,mrna转运到细胞质后翻译形成蛋白质起作用。内含子的剪接通过剪接体以及多种剪接因子完成。剪接体由蛋白质和rna组成的snrnp复合体u1、u2、u4、u5和u6等组成。哺乳动物中剪接辅助因子u2af由u2af65和u2af35两个亚基组成异二聚体,负责招募u2snrnp到内含子的特定位点参与pre-mrna的剪接(shaoetal.,2014)。u2af65包含2个功能结构域-rna结合结构域(rrm)和n-端富含精氨酸/丝氨酸的rs结构域。rs结构域与premrna的分支点(branchpoint,bp)交叉连接参与pre-mrna剪接(valcárceletal.1996)。u2af65含有3个rrm结构域,前2个rrm使u2af65与pre-mrna结合形成复合物,并协同作用招募u2复合体结合到pre-mrna的多聚嘧啶序列(polypyrimidinetrac,ppt)上游的分支位点;u2af65c-的第3个rrm具有特殊的结构特征,主要参与蛋白质之间的相互作用(kielkopfetal.2004)。u2af65与u2af35以复合体的形式起作用,但二者的功能并不完全相同,参与剪接的基因也各不相同(shaoetal.,2014)。u2af65和u2af35的突变导致不同的可变剪接事件(hastingsetal.2007);u2af35突变能够延迟hela细胞的生长,但u2af65的突变不能引起lela细胞的生长的迟缓。有实验表明u2af35是一个组成型剪接因子,可能参与所有pre-mrna内含子的剪接(wuetal.1999);而u2af65b仅参与某些特定基因的剪接。体外实验发现,u2af65b并非参与每个内含子ppt的结合,并且u2af65b的功能可以被其他蛋白代替(shaoetal.,2014)。最近的研究结果显示u2af65的敲除并没有引起大范围的pre-mrna剪接缺陷,而只是引起部分基因的剪接发生变化,表明u2af65参与的pre-mrna的剪接具有非常高的特异性(hastingsetal.2007)。
目前,人们对植物的u2af了解较少。基因序列比对显示,拟南芥中存在2个u2af65的同源蛋白(atu2af65a和atu2af65b)和2个u2af35的同源蛋白(atu2af35a和atu2af35b)。atu2af35a和atu2af35b基因表达下调的突变体在长日照和短日照条件下开花时间延迟,突变体的株型叶片形态、花型以及角果的形态都发生了变化(wangandbrendel,2006)。拟南芥剪接因子sf1与atu2af65a和atu2af65b都存在互作关系,该基因的突变体sf1在长日照下花期提前,但突变体的植株长的非常矮小(jiangetal.2014),难以应用。因此,有必要寻找仅影响植物开花时间而对其他生长发育过程无影响的基因。
技术实现要素:
本发明的发明人针对上述现有技术的情况,提供了一种剪接辅助因子atu2af65b在植物花期调控中的应用,所述的剪接辅助因子atu2af65b来源于拟南芥,通过提高该基因表达水平可用于延迟植物开花,而降低该基因表达水平使一年生植物提早开花或多年生植物缩短童期,可应用于基因工程改良植物的生长进程中。
本发明所述剪接辅助因子atu2af65b,其核苷酸序列如seqidno.1所示,所编码的氨基酸序列如seqidno.2所示。
具体应用是可以通过提高该基因表达水平用于延迟植物开花;也可以通过降低该基因表达水平使一年生植物提早开花或多年生植物缩短童期;但两者均不影响植株其他方面的生长发育。
可见本发明所提供的atu2af65b基因在植物花期控制中具有非常大的应用价值,可用于解决育种工作中某些植物父、母本不同期开花而造成的杂交授粉的难题;用于多年生植物缩短童期,提早结果;也可用于花卉的花期调控,使花卉四季均衡生产和反季节供花,以解决花卉市场淡旺季矛盾。本发明为改良植物的生长进程、调控植物的花期提供了一种有效的技术手段,具有广泛的应用前景和经济价值。
附图说明
图1为拟南芥atu2af65b的2个突变体atu2af65b-1和atu2af65b-2中t-dna的插入位点、鉴定基因型的引物位置及突变体纯合体鉴定结果;
图2为突变体atu2af65b-1和atu2af65b-2中atu2af65b基因的表达结果;
图3为野生型拟南芥中扩增atu2af65b基因编码区的结果;
图4为atu2af65b同源重组到表达载体并转化大肠杆菌后的pcr鉴定;
图5为含有atu2af65b基因的表达载体转化到农杆菌gv3101后的pcr鉴定;
图6为超表达atu2af65b转基因拟南芥的pcr鉴定结果;
图7为atu2af65b基因在野生型拟南芥茎尖的表达量远高于幼苗、花和种子;
图8在ms培养基上生长7天的野生型、atu2af65b-1和atu2af65b-2拟南芥幼苗;
图9为移苗后在温室(长日照条件下)生长3周的野生型、atu2af65b-1和atu2af65b-2拟南芥;
图10为移苗后在温室长日照条件下生长4周的野生型、atu2af65b-1和atu2af65b-2拟南芥植株;
图11为移苗后在温室长日照条件下开始抽薹时野生型、atu2af65b-1和atu2af65b-2突变体的一个完整植株的叶片;
图12为在长日照条件下的野生型、atu2af65b-1和atu2af65b-2突变体的花和花序;
图13为移苗后在温室长日照条件下生长6周的野生型、atu2af65b-1和atu2af65b-2突变体植株;
图14为在长日照条件下的野生型、atu2af65b-1和atu2af65b-2突变体的角果;
图15为移苗后在温室短日照条件下生长8周的野生型、atu2af65b-1和atu2af65b-2拟南芥;
图16为移苗后在温室短日照条件下生长15周的野生型、atu2af65b-1和atu2af65b-2拟南芥;
图17为长日照下生长2周的野生型、突变体atu2af65b-1与atu2af65b-2和超表达atu2af65b拟南芥的生长情况;
图18为长日照下生长4周的野生型、突变体atu2af65b-1与atu2af65b-2和超表达atu2af65b拟南芥抽薹情况;
图19为长日照下生长5周的野生型、突变体atu2af65b-1与atu2af65b-2和超表达atu2af65b拟南芥抽薹、开花情况;
图20为长日照下野生型、突变体atu2af65b-1与atu2af65b-2和超表达atu2af65b植株开花时间和抽薹时叶片数量统计结果。
具体实施方式
以下实施例中进一步定义本发明,根据以上的描述和这些实施例,本领域技术人员可以确定本发明的基本特征,并且在不偏离本发明精神和范围的情况下,可以对本发明作出各种改变和修改,以使其适用各种用途和条件。除特殊注明外,本发明所采用的均为本领域现有技术;
下述实施例中所用的材料如下:
atu2af65b基因的t-dna插入突变体atu2af65b-1(wiscdslox321f12)和atu2af65b-2(salk_055049)是将野生型拟南芥中的atu2af65b基因通过t-dna插入进行敲除突变得到的,其余基因没有变化。这两个突变体种子均购自abrc。拟南芥材料均为哥伦比亚生态型(col)。植物生长条件:温度20-22℃,光强为120μmolm-2s-1。长日照为16h光照、8h暗期;短日照为8h光照、16h暗期。
实施例1atu2af65b基因的突变体atu2af65b-1和atu2af65b-2中纯合体的鉴定
(1)取温室生长2周左右的拟南芥,从植株上剪取一至两个叶片5-10mg放入离心管中,研磨后加400μl提取缓冲液(200mmtris-hclph7.5,250mmnacl,25mmedta,0.5%sds),常规方法提取基因组dna。
(2)以基因组dna为模板,分别用引物lp和rp、lb和rp进行聚合酶链式反应(pcr)扩增。
pcr扩增的反应体系为:2×estaqmastermix10μl,上游引物lp1/lb1(20μmol/l)0.5μl,下游引物rp1(20μmol/l)0.5μl,模板1.0μl,加ddh2o至总体积为20μl。
pcr扩增的反应条件为:首先95℃预变性5min;94℃变性30s、56℃退火30s、72℃延伸1min,进行30个循环;最后72℃延伸10min;
电泳检测pcr结果,lp和rp引物扩增不出来而lb和rp引物能扩增出特异性条带的植株为纯合体;lp和rp引物以及lb和rp引物都能扩增出特异性条带的植株为杂合体。lb和rp引物扩增不出来而lp和rp引物能扩增出特异性条带的植株为野生型。同时设置野生型拟南芥(wt)基因组dna为对照。
鉴定atu2af65b-1基因型的引物位置见图1a,引物序列分别为:
上游引物lp1:5′-cgttcaacccagtagaagacc-3′;其核苷酸序列如seqidno.3所示,
下游引物rp1:5′-tggatgatgagttacgaaggc-3′;其核苷酸序列如seqidno.4所示,
上游引物lb1(p745):5′-aacgtccgcaatgtgttattaagttgtc-3′,其核苷酸序列如seqidno.5所示,引物由上海生物工程有限公司合成。
结果如图1b所示,m为dna分子量标准dl2000;野生型拟南芥基因组dna(样品1,wt)为模板、lp1和rp1引物扩增得到的产物长度为1.1kb,lb1与rp1扩增不出片段,说明所用引物以及pcr程序合适。用这两对引物分别鉴定atu2af65b-16株(样本2-7)突变体材料,如图1b所示,2、3、7为杂合体,4为纯合体;5、6为野生型。
鉴定atu2af65b-2基因型的引物位置见图1a,引物序列分别是:
上游引物lp2:5′-tgggtcatgatctggattagg-3′;其核苷酸序列如seqidno.6所示,
下游引物rp2:5′-tattggccgcaagtctatacg-3′;其核苷酸序列如seqidno.7所示,
上游引物lb2(lbb1.3):5′-attttgccgatttcggaac-3′,其核苷酸序列如seqidno.8所示;
如图1c所示,m为dna分子量标准dl2000;以野生型拟南芥基因组dna(样品8)为对照,用lp2和rp2引物扩增得到1.2kb的片段,lb2与rp2扩增不出片段,说明所用引物以及pcr程序合适。用这两对引物分别鉴定atu2af65b-26株(样本9-14)突变体,10为杂合体,其余均为纯合体。
实施例2
在实施例1的基础上对突变体atu2af65b-1和atu2af65b-2纯合体中atu2af65b基因表达水平进行检测
(1)取在ms培养基上生长两周的突变体纯合体和野生型拟南芥植物各10mg,用总rna提取试剂盒(购自天根生化科技(北京)有限公司)分别提取野生型和突变体的总rna,方法参照说明书进行。用反转录酶(北京全式金生物技术有限公司)将1ugrna反转录成cdna。
(2)以cdna为模板,用atu2af65b基因特异性引物,通过反转录pcr检测野生型和突变体中atu2af65b转录本的量(按照天根生化科技(北京)有限公司的通用型一步法rt-pcr试剂盒说明书进行)。引物序列为:
上游引物(atu2af65bf):5′-atgatgagttacgaaggcaacggag-3′,其核苷酸序列如seqidno.9所示;
下游引物(atu2af65br):5′-tcagtcttcgtagtcgccttgg-3′,其核苷酸序列如seqidno.10所示。
tubulin2基因为内参,引物序列为:
上游引物(tubulin2f):5′-ggtatccaggtcggaaatgc-3′,其核苷酸序列如seqidno.11所示;
下游引物(tubulin2r):5′-tcccgtagtcaacagaaagt-3′,其核苷酸序列如seqidno.12所示。(上海生物工程有限公司合成)
pcr扩增的反应体系为:2×estaqmastermix25μl,上游引物(20μmol/l)1μl,下游引物(20μmol/l)1μl,模板2.0μl,加ddh2o至总体积为50μl。
pcr扩增的反应条件为:首先95℃预变性5min;94℃变性40s、58℃退火30s、72℃延伸1min,进行28个循环;最后72℃延伸10min;
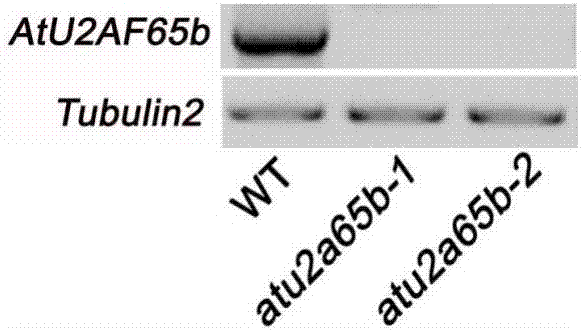
电泳检测rt-pcr结果。图2结果显示野生型拟南芥中有atu2af65b基因的转录本,但atu2af65b-1和atu2af65b-2中几乎没有atu2af65b转录产物。
实施例3
超表达atu2af65b载体的构建和转基因植株的鉴定
(1)以实施例2中野生型拟南芥的cdna为模板,用实施例2中相同atu2af65b基因特异性引物(atu2af65bf和atu2af65br)扩增atu2af65b编码区全长。引物序列为:
上游引物(atu2af65bf):5′-atgatgagttacgaaggcaacggag-3′,其核苷酸序列如seqidno.9所示;
下游引物(atu2af65br):5′-tcagtcttcgtagtcgccttgg-3′,其核苷酸序列如seqidno.10所示;
pcr扩增的反应体系为:高保真dna聚合酶0.5μl,5×dna聚合酶缓冲液10μl,dntp1μl,上游引物(20μmol/l)1.5μl,下游引物(20μmol/l)μl,加ddh2o至总体积为50μl。pcr扩增条件同实施例2。
电泳检测pcr产物长度,marker为dna分子量标准dl2000。图3显示pcr扩增的片段长度为1.8kb左右,与atu2af65b编码区实际长度1770bp相近。
(2)将pcr产物用胶回收试剂盒(天根生化科技(北京)有限公司)进行回收,方法参照说明书进行。将回收的产物连接到gateway入门载体(pcr8/gw/topotacloningkit,invitrogen,方法参照说明书进行),转化大肠杆菌感受态细胞(dh5α)后,经50μg/ml壮观霉素抗生素筛选,对阳性菌落测序(生工生物工程(上海)股份有限公司)。
用质粒提取试剂盒(购自天根生化科技(北京)有限公司)提取测序正确的质粒,通过lrclonase(购自invitrogen)将质粒同源重组到pmdc43双元表达载体并转化大肠杆菌感受态细胞dh5α(天根生化科技(北京)有限公司),方法参照说明书进行。用50μg/ml卡那霉素抗生素筛选,提取阳性菌落质粒(购自天根生化科技(北京)有限公司的质粒提取试剂盒),方法参照说明书进行。利用载体上35s启动子序列的特异性上游引物(35sf)和atu2af65b特异性的下游引物(atu2af65br)进行pcr鉴定,电泳检测pcr产物长度。所用引物序列分别为:
上游引物(35sf):5′-gaaggtggctcctacaaatgcca-3′,其核苷酸序列如seqidno.13所示;
下游引物(atu2af65br):5′-tcagtcttcgtagtcgccttgg-3′,其核苷酸序列如seqidno.10所示;
pcr扩增的反应体系为:2×estaqmastermix10μl,上游引物(20μmol/l)0.5μl,下游引物(20μmol/l)0.5μl,模板1.0μl,加ddh2o至总体积为20μl。pcr扩增条件同实施例2。
图4为电泳检测结果,marker为dna分子量标准dl2000,1为空载体质粒的阴性对照,2和3为转化成功的阳性菌落的质粒pcr结果,pcr产物长度约为2.2kb,说明该基因被同源重组到表达载体并成功转换入大肠杆菌。
(3)常规方法将质粒转化农杆菌菌株gv3101的感受态细胞。50μg/ml的卡那霉素和50μg/ml利福平筛选,提取菌落的质粒(天根生化科技(北京)有限公司的质粒提取试剂盒),利用该实施例(2)相同的引物和pcr条件进行pcr鉴定。图5显示电泳结果,marker为dna分子量标准dl2000,1-3为阳性菌落的质粒pcr结果,4为大肠杆菌阳性菌落质粒的pcr对照(阳性对照),5为空载体质粒的阴性对照,说明成功将构建好的表达载体转化到农杆菌。
(4)通过农杆菌介导拟南芥的常规方法转化野生型拟南芥,每隔一周转化一次,共转化三次。收获的种子在ms培养基上进行抗生素筛选(50μg/ml卡那霉素),将有抗性的植株移栽到营养土中杂温室培养。常规方法提取生长3周左右拟南芥叶片的基因组dna为模板,用该实施例2相同的引物和pcr条件进行pcr鉴定,电泳检测pcr产物长度。图6中,marker为dna分子量标准dl2000,1为野生型拟南芥基因组的pcr结果(阴性对照),2为表达载体质粒的pcr结果(阳性对照),3-8为6个具有抗生素抗性株系的pcr鉴定结果,说明这6个株系均为超表达atu2af65b的拟南芥。
实施例4
atu2af65b基因的表达模式检测
(1)分别提取生长2周的野生型拟南芥植株和茎尖、生长6-8周的花和角果的总rna,分别取2ug的rna用反转录酶(北京全式金生物技术有限公司)将rna反转录成cdna。
(2)用同实施例2相同的引物(atu2af65b基因特异性引物:atu2af65bf和atu2af65br;内参基因tubulin2特异性引物:tubulin2f和tubulin2r)和相同的pcr反应体系以及反应条件,通过反转录pcr检测这几种组织器官中atu2af65b的转录水平。引物序列为:
上游引物(atu2af65bf):5′-atgatgagttacgaaggcaacggag-3′,其核苷酸序列如seqidno.9所示;
下游引物(atu2af65br):5′-tcagtcttcgtagtcgccttgg-3′,其核苷酸序列如seqidno.10所示。
tubulin2基因为内参,引物序列为实施例2的tubulin2f和tubulin2f
上游引物(tubulin2f):5′-ggtatccaggtcggaaatgc-3′,其核苷酸序列如seqidno.11所示;
下游引物(tubulin2r):5′-tcccgtagtcaacagaaagt-3′,其核苷酸序列如seqidno.12所示;(上海生物工程有限公司合成)。
电泳检测rt-pcr结果。结果如图7所示,该基因在生长2周的拟南芥的茎尖表达量远高于幼苗、花和角果,说明该基因的主要功能是参与花期调控。
实施例5
野生型拟南芥和突变体植株在长日照下的生长发育表型进行观察和统计(根据参考文献,抽薹时花的发育已经完成。本实施例中以抽薹时间为开花统计的参数)
(1)首先,将野生型拟南芥(wt)和突变体纯合体(atu2af65b-1,atu2af65b-2)种子播种于0.5ms固体培养基上,22℃、长日照(16h光照/8h暗期)垂直培养1周左右。观察植株生长状况。结果如图8所示,突变体与野生型的植株的根长无明显差别。
(2)同时将野生型拟南芥和突变体atu2af65b的幼苗移栽到植物营养基质中,每个营养杯中移栽一株野生型拟南芥和一株突变体,设置100个重复。观察在长日照条件下(22℃)生长的野生型拟南芥和突变体植株的叶片、叶片形态和叶色等的差异。图9显示温室生长3周左右的野生型拟南芥和突变体植株间叶片、叶片形态和叶色无明显差异。
(3)观察同一个营养杯中生长的野生型拟南芥和突变体atu2af65b植株抽薹时间的差异。温室长日照条件下生长4周的突变体植株atu2af65b-1和atu2af65b-2都已经抽薹,但同一个营养杯中生长的野生型拟南芥(箭头)还没有或刚开始抽薹(图10a和b)。
对100个重复实验的统计结果显示野生型拟南芥在22℃长日照条件下叶片数量达13片时开始抽薹,突变体叶片数约为11片开始抽薹(图10c)。野生型拟南芥需要生长28天开始抽薹,突变体atu2af65b-1和atu2af65b-2各需要22天和21天开始抽薹(图10d)。突变体抽薹时间比野生型提早6-7天,二者间差异显著。
(4)将开始抽薹时的叶片从植株上剥离,观察叶片形态和叶色,图11显示野生型拟南芥和突变体atu2af65b-1和atu2af65b-2除叶片数量有差异外,叶片形态等均无明显差异。
(5)观察同一个营养杯中温室生长的盛花期的野生型拟南芥和突变体atu2af65b的花絮和花,均没有明显差异(图12)。
(6)观察温室长日照条件下生长6周的野生型拟南芥和atu2af65b突变体。突变体atu2af65b-1和atu2af65b-2比野生型拟南芥提早进入结果期(图13),角果形态无明显差异(图14)
实施例6
野生型拟南芥和突变体植株在短日照下生长发育表型进行观察和统计,开花时间的统计标准参照实施例5进行。
(1)将移栽到植物培养基质的野生型拟南芥和突变体atu2af65b的幼苗在短日照条件下(8h光照/16h暗期,22℃)培养。每个营养杯中同样种植一株野生型拟南芥和一株突变体,设置100个重复。观察野生型拟南芥和突变体植株的叶片、叶片形态和叶色等的差异。图15显示生长8周左右的野生型拟南芥和突变体植株间叶片、叶片形态和叶色没有明显差异。
(2)进一步观察短日照条件下野生型拟南芥和突变体atu2af65b植株抽薹时间的差异。生长15周的突变体植株atu2af65b-1和atu2af65b-2都已经抽薹,但同一个营养杯中生长的野生型拟南芥还都没有抽薹(箭头)(图16a和b)。对100个重复实验的统计结果显示野生型拟南芥在短日条件下抽薹时叶片数量约56片,突变体叶片数约为40片(图16c),突变体比野生型拟南芥少16个叶片,差异显著。野生型拟南芥需要生长115天开始抽薹,突变体atu2af65b-1和atu2af65b-2需要95天开始抽薹(图16d)。突变体抽薹时间提早20天,与野生型拟南芥差异显著。
实施例7
野生型拟南芥、突变体和超表达转基因拟南芥植株在长日照下生长发育表型进行观察和统计,开花时间的统计标准参照实施例5进行。
(1)将野生型(wt)、突变体(atu2af65b-1和atu2af65b-2)和三个超表达株系(oe1、oe2和oe3)的幼苗种于同一容器中,在完全相同的条件下下生长,并设计100个重复。观察在长日照条件下(22℃)植株生长发育情况。图17a和b显示温室生长2周左右的野生型拟南芥、突变体植株和超表达植株间叶片大小、形态和叶色无明显差异。但超表达株系oe1和oe2中atu2af65b基因的表达量明显升高,oe3中atu2af65b基因的表达量与野生型相近(图17c)。
(2)进一步观察这些植株抽薹时间的差异。温室长日照条件下生长4周的突变体植株atu2af65b-1和atu2af65b-2都已经开花并且结果,同一个营养杯中生长的野生型拟南芥抽薹并开始开花,但超表达株系仅oe3(其atu2af65b基因的表达量与野生型相近)抽薹开花(图18,大箭头),oe1和oe2即将抽薹或刚抽薹(atu2af65b基因的表达量明显比野生型高)(图18,小箭头),结果说明atu2af65b基因的表达量与开花时间成负相关。
(3)温室长日照条件下生长5周的超表达植株开始抽薹(图19)。
(4)对100个重复实验的统计结果显示野生型拟南芥在抽薹时叶片数量约14片,atu2af65b表达量明显升高的oe1和oe2植株抽薹时叶片约17片(图20a)。野生型拟南芥需要生长28天左右开始抽薹,oe1和oe2植株植株生长38天左右开始抽薹(图20b),超表达植株抽薹时间比野生型植株晚10天左右,差异显著。
由上述结果分析表明,本发明所涉及的atu2af65b基因主要在茎尖表达,是一个新的植物特异性花期调控基因。提高atu2af65b基因的表达水平能延迟植物抽薹和开花;降低该基因的表达水平能使植物抽薹和开花时间提前,尤为重要的是上调和下调该基因的表达水平都不改变植物的生长势、株型、叶型和花型等性状,为改良植物的生长进程、调控植物的花期提供了一种有效的技术手段,具有广泛的应用前景和经济价值。
<110>山东农业大学
<120>一种剪接辅助因子atu2af65b在植物花期调控中的应用
<160>13
<210>1
<211>1770
<212>dna
<213>拟南芥(arabidopsisthaliana(l.))
<400>1
atgatgagttacgaaggcaacggagatggtgtcgctatctccaccgaaaaccacaacgag60
aactacatatctctagagagctcgccttttcatgaagactccaagtctcgagaatctcat120
gatcttaaaaaggattcttcgaaaatcagtgagaaagacaatgaaaatggacgtgataaa180
gacggaaacaaagatcgtgatcgtgagaaggatcgtgatagggagaagagcagagacaga240
gatagggagaaaagcagagatagagatagggatagggagagaagtaaggataggcaacga300
gaccgtcatcatagagatcgtcatagggaccgtagcagagaacgtagtgagaaaagagat360
gatttggatgatgatcatcatcgcagaagtcgagaccgtgataggaggaggtctcgcgat420
cgagacagagaggtgagacacaggcgacggtcacgctctcggtcaaggagccgctctgaa480
cgtaggtcaaggtcagaacatagacataagtctgaacatagatcacgctcacggtcgcga540
tcaaggtcaaagagcaagaggagaagtggatttgatatggcgcctcctgatatgttagct600
gctactgctgttgctgcagcaggccaggttcctagtgtgccaactactgctactattcca660
gggatgttctcaaacatgtttccaatggtccctggtcagcaactaggtgcgcttcctgta720
ctgccagttcaggcaatgactcaacaggcaactaggcatgctcggcgagtctatgttggt780
ggccttccaccaactgctaatgaacagtcggtgtcaacgttctttagccaagtgatgtcg840
gccattgggggaaacactgctgggccaggtgatgcggtggtcaatgtttatataaaccac900
gaaaagaagtttgcttttgtagagatgagatctgttgaggaggctagtaacgcaatggca960
ttagacggcattatattagagggagttcctgtaaaggtgaggaggccaactgactataac1020
ccgtctcttgctgcaactcttggtccgagccagcctaatcccaacctcaacttaggggct1080
gttggattgtcttcagggtctactggtgggcttgagggtcccgaccgcatttttgtgggc1140
ggacttccctattacttcacagaggtacagatcagggagctcttggagtcttttgggccc1200
ctaagaggtttcaacttggtcaaagacagggagactggaaattcgaagggatatgcattc1260
tgtgtttaccaagatccgtcagtaacagatattgcatgtgccgctctaaacgggattaag1320
atgggtgataagacacttacagttaggcgtgcaatccagggtgcaattcaacctaaaccc1380
gagcaagaagaagtcttactttatgcccaacaacagattgctttgcagaggcttatgttc1440
caaccaggaggtactcccaccaagattgtctgtttgactcaagtggttacagctgatgat1500
cttagagacgatgaagaatatgcagagataatggaggacatgcgacaagaaggtggaaaa1560
ttcggtaacttagtgaatgttgtgattccgaggcctaatccagatcatgacccaacacca1620
ggagttgggaaggttttcttagagtatgcggatgtggatggctcatcaaaggccagatcg1680
gggatgaatggaagaaaatttggaggaaaccaagtagtggctgtgtattaccccgaagac1740
aagtatgcccaaggcgactacgaagactga1770
<210>2
<211>589
<212>prt
<213>拟南芥(arabidopsisthaliana(l.))
<400>2
metmetsertyrgluglyasnglyaspglyvalalaileserthr
151015
gluasnhisasngluasntyrileserleugluserserprophe
202530
hisgluaspserlysserarggluserhisaspleulyslysasp
354045
serserlysileserglulysaspasngluasnglyargasplys
505560
aspglyasnlysaspargaspargglulysaspargaspargglu
657075
lysserargaspargaspargglulysserargaspargasparg
808590
asparggluargserlysaspargglnargasparghishisarg
95100105
asparghisargaspargserarggluargserglulysargasp
110115120
aspleuaspaspasphishisargargserargaspargasparg
125130135
argargserargaspargasparggluvalarghisargargarg
140145150
serargserargserargserargsergluargargserargser
155160165
gluhisarghislyssergluhisargserargserargserarg
170175180
serargserlysserlysargargserglypheaspmetalapro
185190195
proaspmetleualaalathralavalalaalaalaglyglnval
200205210
proservalprothrthralathrileproglymetpheserasn
215220225
metpheprometvalproglyglnglnleuglyalaleuproval
230235240
leuprovalglnalametthrglnglnalathrarghisalaarg
245250255
argvaltyrvalglyglyleuproprothralaasngluglnser
260265270
valserthrphepheserglnvalmetseralaileglyglyasn
275280285
thralaglyproglyaspalavalvalasnvaltyrileasnhis
290295300
glulyslysphealaphevalglumetargservalglugluala
305310315
serasnalametalaleuaspglyileileleugluglyvalpro
320325330
vallysvalargargprothrasptyrasnproserleualaala
335340345
thrleuglyproserglnproasnproasnleuasnleuglyala
350355360
valglyleuserserglyserthrglyglyleugluglyproasp
365370375
argilephevalglyglyleuprotyrtyrphethrgluvalgln
380385390
ilearggluleuleugluserpheglyproleuargglypheasn
395400405
leuvallysasparggluthrglyasnserlysglytyralaphe
410415420
cysvaltyrglnaspproservalthraspilealacysalaala
425430435
leuasnglyilelysmetglyasplysthrleuthrvalargarg
440445450
alaileglnglyalaileglnprolysprogluglnglugluval
455460465
leuleutyralaglnglnglnilealaleuglnargleumetphe
470475480
glnproglyglythrprothrlysilevalcysleuthrglnval
485490495
valthralaaspaspleuargaspaspgluglutyralagluile
500505510
metgluaspmetargglngluglyglylyspheglyasnleuval
515520525
asnvalvalileproargproasnproasphisaspprothrpro
530535540
glyvalglylysvalpheleuglutyralaaspvalaspglyser
545550555
serlysalaargserglymetasnglyarglyspheglyglyasn
560565570
glnvalvalalavaltyrtyrprogluasplystyralaglngly
575580585
asptyrgluasp
589
<210>3
<211>21
<212>dna
<213>人工序列
<400>3
cgttcaacccagtagaagacc21
<210>4
<211>21
<212>dna
<213>人工序列
<400>4
tggatgatgagttacgaaggc21
<210>5
<211>28
<212>dna
<213>人工序列
<400>5
aacgtccgcaatgtgttattaagttgtc28
<210>6
<211>21
<212>dna
<213>人工序列
<400>6
tgggtcatgatctggattagg21
<210>7
<211>21
<212>dna
<213>人工序列
<400>7
tattggccgcaagtctatacg21
<210>8
<211>19
<212>dna
<213>人工序列
<400>8
attttgccgatttcggaac19
<210>9
<211>25
<212>dna
<213>人工序列
<400>9
atgatgagttacgaaggcaacggag25
<210>10
<211>22
<212>dna
<213>人工序列
<400>10
tcagtcttcgtagtcgccttgg22
<210>11
<211>20
<212>dna
<213>人工序列
<400>11
ggtatccaggtcggaaatgc20
<210>12
<211>20
<212>dna
<213>人工序列
<400>12
tcccgtagtcaacagaaagt20
<210>13
<211>23
<212>dna
<213>人工序列
<400>13
gaaggtggctcctacaaatgcca23
- 还没有人留言评论。精彩留言会获得点赞!