磷酸转位器的制作方法
发明领域
本发明涉及可用于从细菌中输出磷酸化的化合物的多肽。该多肽包含磷酸转位器(phosphatetranlocator)和微生物膜整合蛋白。本发明还涉及产生和输出磷酸化的产物的细菌方法。
发明背景
通常使用化学合成来生产高价值的化学化合物。然而,这些方法可能成本高昂并且导致低收率。此外,由于使用高温、高压和有机溶剂产生的废料,它们对环境有重大影响。
许多目标化合物在自然界中是通过在生物体(例如细菌)中天然存在的酶产生的。因此,近年来,利用细菌代谢产生这些所需化合物引起了人们的极大兴趣。已经开发了一些方法,其中微生物可以经工程化以产生有价值的化学物质,作为化学合成的可持续的并且环境友好的替代物。
非常期望产生的一类化合物是磷酸化的化合物,例如磷酸化的糖。这些化合物的生产成本非常高,例如,磷酸二羟基丙酮(dhap)的当前价格在每克1200-6000欧元之间。许多磷酸化的糖在许多生物体,例如细菌中,通过糖酵解过程自然产生。但是,这些化合物用于细菌代谢中,因此通常不在细菌内积聚,并且由于细菌膜上存在负电荷而不能输出。工程化细菌也存在产生过量的这些化合物的问题,例如,如果发生过量的dhap,这将在细胞内部降解为甲基乙二醛,其是有毒的并导致细胞死亡。
本发明已经开发了一种多肽,其可以在细菌中表达,以将这些高价值的磷酸化的化合物输出跨过整个细菌膜。因此,这实现了磷酸化的化合物的细菌生产。这种方法将在这些产品的生产中节省大量成本。
发明概述
本发明基于以下发现:可以对细菌细胞进行基因工程化以输出磷酸化的化合物。这样,这些基因工程化的细菌可用于以比传统合成路线更经济有效和环境友好的方式生产高价值的磷酸化的化合物。
为了允许输出磷酸化的化合物,已经开发了工程化的多肽。该多肽包含磷酸转位器和微生物膜整合蛋白。磷酸转位器天然地存在于植物的质体中,并且用于将光合作用的产物从质体中运出用于代谢途径中。出乎意料的是,转位器和微生物膜整合蛋白的组合允许磷酸转位器在细菌膜内表达,其中它将识别并输出磷酸化的化合物。
该工程化的多肽可以进一步与其他基因工程化方法组合以改善通过细菌产生磷酸化的化合物。例如,可以产生基因敲除,其除去产生降解磷酸化的化合物的酶所需要的基因。使用这种方法,细菌产生更高水平的磷酸化的产物,然后将其输出。
在正常情况下,磷酸化的化合物的积累对细胞有毒。在本发明中,由于转位器的存在,这些产物得到输出,并因此不会对细胞产生负面影响。此外,一旦产物得到输出,就可以容易地从培养基中提取它们。
因此,本发明的一个方面涉及一种包含磷酸转位器和微生物膜整合蛋白的多肽。
本发明的第二方面涉及一种核酸序列,其编码包含磷酸转位器和微生物膜整合蛋白的多肽。
本发明的第三方面涉及一种载体,其包含编码包含磷酸转位器和微生物膜整合蛋白的多肽的核酸序列。
本发明的第四方面涉及一种经基因修饰以输出磷酸化的化合物的细菌。
本发明的第五方面涉及一种制备磷酸化的化合物的方法,包括培养如上定义的细菌的步骤。
附图说明
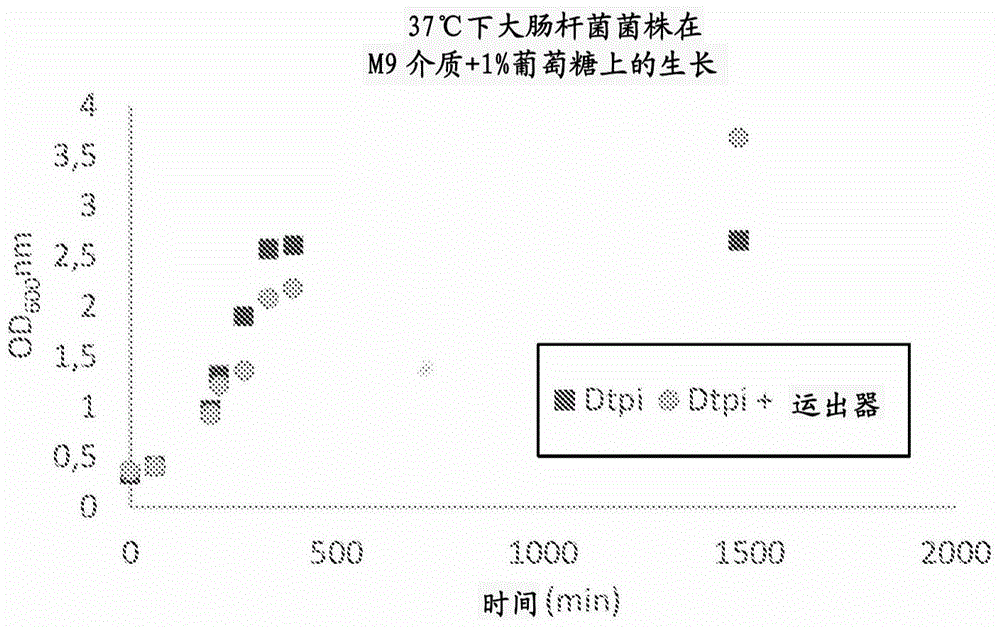
图1显示了两种细菌培养物的光密度(od)与时间的关系。已经对第一细菌培养物进行了基因工程化以敲除编码磷酸三糖异构酶dtpi(δtpi)的基因。已经对第二细菌培养物进行了基因工程化以敲除编码磷酸三糖异构酶的基因并用表达磷酸三糖/磷酸转位器和膜整合蛋白(称为mistic)的融合蛋白(dtpi+转运器(transporter))的质粒转化。od是细胞生长的量度。
图2显示了在表达磷酸三糖/磷酸转位器+mistic的融合蛋白的大肠杆菌(e.coli)细胞的细菌细胞培养物上清液以及未经基因工程化的比较的对照培养物中检测到的dhap的浓度。dhap的浓度通过实施例3中所述的酶促测定法来确定。
图3显示了使用气相色谱质谱(gcms)分析的dhap和介质对照的色谱图。上图显示了含dhap的对照样品的色谱图。计数v采集时间为5.54分钟时可以看到一个大的峰。下图显示了m9介质的对照样品的色谱图。计数v采集时间为大约5.54min时,不存在峰。
图4显示了其中已生长出对照大肠杆菌菌株的m9介质的色谱图。在5.4min时没有峰表明该样品中不存在dhap。
图5显示了m9介质的色谱图,其中已生长了表达磷酸三糖/磷酸转位器+mistic的融合蛋白的大肠杆菌菌株。在5.38min处看到一个大峰。这与图3上图中显示的dhap对照样品的计数v采集时间一致,因此该样品中存在dhap。
图6显示了通过高效液相色谱质谱(hplc-ms)对大肠杆菌培养物上清液中的甘油磷酸-3-磷酸检测。
图6-a显示了包含由c-13同位素标记的标准甘油-3-磷酸的对照样品的色谱图。在7min处看到一个大峰。
图6-b显示了其中已生长出对照大肠杆菌菌株的m9介质的色谱图。在7min处没有峰,表明该样品中不存在甘油-3-磷酸。
图6-c显示了其中已经生长出表达磷酸三糖/磷酸转位器+mistic的融合蛋白的大肠杆菌菌株的m9介质的色谱图。在7min处看到一个大峰。这与图6-a的上图中显示的甘油-3-磷酸对照样品的计数v采集时间一致,因此该样品中存在甘油-3-磷酸。
图7显示了通过质谱检测来自表达来自拟南芥(arabidospisthaliana)的木酮糖-5-磷酸转位器的δtkta的培养基中的木酮糖5-磷酸。在7.830min处看到一个峰,表明该样品中存在木酮糖5-磷酸。
发明详述
在本发明的第一方面,提供了一种多肽,其包含磷酸转位器和细菌膜整合蛋白。
如本文所用,术语“磷酸转位器”是指可以横跨膜上转移磷酸化的化合物的膜蛋白。这些磷酸转位器通常在光合作用真核生物以及其他含有质体的生物体中的质体中发现。质体是由内膜和外膜围绕的细胞器并且具有许多不同类型的蛋白质转运器,以移动化合物(例如蛋白质、氨基酸、核酸、肽、糖和磷酸化的糖)进出细胞器。这些转位器可以被称为质体磷酸转位器,并且可以通过它们的优选底物来分类。质体磷酸酯转位器的实例包括磷酸三糖/磷酸转位器、葡萄糖-6-磷酸/磷酸转位器、磷酸烯醇-丙酮酸/磷酸转位器或木酮糖5-磷酸/磷酸转位器。
根据本发明的第一方面的磷酸转位器可以是质体磷酸转位器,其中磷酸转位器天然地在质体的膜中表达。优选地,磷酸转位器选自包括以下项的组:磷酸三糖/磷酸转位器、葡萄糖-6-磷酸/磷酸转位器、磷酸烯醇-丙酮酸/磷酸转位器或木酮糖5-磷酸/磷酸转位器。在一个优选的实施方案中,磷酸转位器是磷酸三糖/磷酸转位器。磷酸三糖/磷酸转位器可以具有seqidno1的氨基酸序列。
如本文所用,术语“膜整合蛋白(membraneintegratingprotein)”是指可以在有或没有伴侣蛋白的情况下将蛋白寻址和/或折叠进入生物膜的蛋白序列。当氨基序列短于一百个氨基酸并且更具体地为约十个氨基酸时,该膜整合蛋白也可以是膜整合肽。优选地,该膜整合肽是指蛋白质进入生物膜的周知的寻址序列,也称为信号肽。
如本文所用,术语“膜整合蛋白”还指可以折叠进入生物膜的整合膜蛋白。如本文所用,术语“膜”是指充当生物体中边界的脂质双层。膜整合蛋白将附着在生物膜上。该蛋白质可以是跨越生物膜的整个脂质双层的多中心蛋白,或者可以是附着在膜上但不跨越整个脂质双层的单中心蛋白。
这种膜整合蛋白质可以是微生物的。特别地,它可以来源于细菌、真菌或古细菌。优选地,膜整合蛋白来源于芽孢杆菌属细菌,最优选地来源于枯草芽孢杆菌(bacillissubtilis)。膜整合蛋白质可以自主折叠进入膜,特别是细菌膜。通过自主折叠,膜整合蛋白不需要转位子即可将其自身插入膜中。
特别优选的膜整合蛋白是已知为mistic的蛋白。mistic是一种整合的膜蛋白,其自动折叠进入膜,并在整个芽孢杆菌中均保守。根据本发明的实施方案,微生物膜整合蛋白可包含氨基酸序列seqidno2。
为了连接磷酸转位器和微生物膜整合蛋白,它们可以通过使用接头或间隔子序列制备为融合蛋白。接头序列是一系列氨基酸残基,其将磷酸转位器和膜整合蛋白分开。可以选择该系列氨基酸以提供柔性接头序列。通常,柔性接头序列富含甘氨酸。可以选择一系列氨基酸以在两个蛋白质结构域中产生刚性接头或防止两个蛋白质结构域的相互作用。通常,刚性接头序列富含脯氨酸并且可以形成螺旋结构。在高度优选的实施方案中,接头序列具有氨基酸序列sssnnnnnlgsngsg。
根据本发明的第二方面,提供了编码如上定义的多肽的核苷酸序列。该核苷酸序列可以编码磷酸转位器和微生物膜整合蛋白。任选地,该核苷酸序列也可以编码合适的接头序列以附接磷酸转位器和微生物膜整合蛋白。在高度优选的实施方案中,核苷酸序列包含seqidno4。
本领域技术人员将理解,本文描述的所有核酸和氨基酸序列具有同源物、等同物和衍生物。因此,本发明还涵盖在其整个长度上具有与本文所述的核酸和氨基酸序列基本相同的序列的核酸分子。
本领域技术人员将理解,本文定义的核酸和氨基酸序列还可包括本文举例说明的那些特定核酸分子的变体。这些可能自然发生,例如由于菌株变异。例如,包括添加、取代和/或删除。本领域技术人员还将意识到,鉴于遗传密码的简并性,与本文示例的特定核酸分子的变异将是可能的。优选地,变体在其整个长度上与本文所述的核酸序列具有基本上的同一性。
如本文所用,具有“基本上的同一性”的核酸序列优选地与所述序列具有至少80%、90%、91%、92%、93%、94%、95%、96%、97%、98%、98.1%、98.2%、98.3%、98.4%、98.5%、98.6%、98.7%、98.8%、98.9%、99%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%或99.9%的同一性。期望地,术语“基本上的同一性”表示与现有技术的核酸序列相比,所述序列与本文所述的任何序列具有更高的同一性程度。
可以对核苷酸序列进行密码子优化,以在特定表达宿主如细菌中表达。核苷酸序列还可以包含表达所编码的多肽所需的另外的核苷酸序列,例如启动子序列、核糖体结合位点和用于蛋白质的可诱导表达的核苷酸序列。合适的序列将是本领域技术人员已知的。
为了表达所编码的多肽,可以将根据本发明第二方面的核苷酸序列插入合适的表达宿主中。这样,本发明的第三方面是包含如上所定义的核苷酸序列的载体。
载体是将目标核酸序列递送到合适的宿主中以表达由该核酸序列编码的蛋白质的媒介物。合适的载体是本领域已知的,合适的载体的非限制性实例包括质粒、病毒载体、酵母人工染色体和细菌人工染色体。
可以将载体转化到合适的宿主中,例如细菌细胞、酵母细胞、昆虫细胞或哺乳动物细胞。为了将载体引入宿主细胞中,可以使用多种转化、转染和转导方法。技术人员将能够基于所使用的载体的类型和宿主细胞的类型来确定最合适的方法。
根据本发明的第四方面,提供了一种经基因修饰以输出磷酸化的化合物的细菌。特别地,细菌可以经基因修饰以表达如上定义的多肽。为了表达如上定义的多肽,可用包含编码多肽的核酸序列的载体转化细菌,所述多肽包含磷酸转位器和微生物膜整合蛋白。
如本文所用,术语“基因修饰”和“基因工程化”可互换使用。如本文所用,这些术语是指通过改变遗传物质来改变生物体的特性,这可以通过引入、缺失或修饰遗传物质来实现。
一类特别感兴趣的磷酸化的化合物是磷酸化的糖。通常,糖可以具有通式cn(h2o)m,并且磷酸化的糖具有附接的另外的磷酰基。可以由经基因修饰的细菌输出的磷酸化的糖的非限制性实例包括磷酸二羟基丙酮(dhap)、脱氧木糖-5-磷酸、木酮糖-5-磷酸酯、葡萄糖-6-磷酸、果糖-6磷酸、果糖-1,6-二磷酸、3-甘油醛-3-磷酸、甘油-3-磷酸、磷酸烯醇丙酮酸、2-磷酸甘油酸、3-磷酸甘油酸、1,3-二磷酸甘油酸。
细菌可以经基因修饰以表达膜转运器,其中膜转运器将磷酸化的化合物转移跨过细菌膜。转运器可以经由许多不同的机制输出化合物,所述机制例如扩散、易化扩散、初级主动转运或次级主动转运。合适的膜转运器的实例包括载体蛋白和通道蛋白、α型通道、β-桶状孔蛋白、单向运输蛋白、同向运输蛋白、反向运输蛋白、磷酸转位器、烟酰胺核糖核苷转运器。
在一个实施方案中,细菌经基因修饰以表达磷酸转位器,优选地,其中磷酸转位器选自包括以下项的组:磷酸三糖/磷酸转位器、葡萄糖-6-磷酸/磷酸转位器、磷酸烯醇-丙酮酸/磷酸转位器或木酮糖5-磷酸/磷酸转位器,更优选地,其中磷酸转位器是磷酸三糖/磷酸转位器。
为了实现磷酸化的化合物的输出,优选的是组成型表达转位器或转位器与微生物膜整合蛋白的融合蛋白。这样,编码转位器或转位器与微生物膜整合蛋白的融合蛋白的核酸序列可以受组成型启动子如galp启动子、t7启动子、cmv启动子的控制。
磷酸盐转位器允许输出由细菌产生的磷酸化的化合物。为了增强由细菌产生的目标磷酸化的化合物的量,可以对细菌进行进一步的基因工程化。可以对细菌进行基因修饰,以减少磷酸化的化合物的代谢。通过减少磷酸化的产物的代谢,这将增加可用于输出的磷酸化的产物的量。这可以通过多种方式来实现,例如通过使负责代谢目标化合物的基因失活。使基因失活可能涉及目标基因的全部或部分缺失。有多种方法可用于基因失活,这是本领域技术人员已知的。基因失活技术的非限制性实例包括质粒介导的同源重组、线性dna介导的同源重组和插入失活。还可以对细菌进行基因修饰以增加磷酸化的化合物的产生。这可以通过在代谢途径中某些蛋白质的过表达来实现。蛋白质过表达的方法是本领域已知的,并且包括使用可诱导表达系统,其可以利用可诱导启动子、非编码rna和核糖开关。这些系统将在可诱导启动子、非编码rna或核糖开关的控制下增强由基因编码的蛋白质的表达。
在一个优选的实施方案中,细菌经基因工程化以输出dhap。为了输出dhap,细菌经基因修饰以表达磷酸三糖/磷酸转位器。转位器任选地作为与微生物膜整合蛋白的融合蛋白产生。在优选的实施方案中,细菌表达seqidno3的氨基酸序列。磷酸三糖/磷酸转位器的表达可以通过用编码该转位器的载体转化细菌来实现。载体可以包含seqidno4的核苷酸序列。
任选地,可以进行基因工程化以降低磷酸三糖异构酶活性。这可以通过使负责产生磷酸三糖异构酶的基因失活来实现,磷酸三糖异构酶是一种将dhap转化为甘油醛-3-磷酸的酶。使用这种方法,dhap将不会用于细菌代谢,因此dhap将开始积累,并因此经由磷酸三糖/磷酸转位器输出。在该实施方案中,由于细菌不使用dhap,因此所产生的dhap将被输出并因此可以实现更高的收率。
许多细菌菌株适合经工程化以输出磷酸化的化合物。这些包括革兰氏阴性细菌,例如假单胞菌(pseudomonas)、克雷伯菌(klebsiella)、肠杆菌(enterobacter)、幽门螺杆菌(helobacter)、沙门氏菌。在一个优选的实施方案中,大肠杆菌(escherichiacoli)是经过基因工程化的。
根据本发明的第五方面,提供了一种制备磷酸化的化合物的方法,包括培养如上所述经基因工程化的细菌的步骤。
细菌培养条件可能取决于所用的细菌菌株并且可以针对特定细菌进行优化。本领域技术人员将能够优化培养条件用于最大的蛋白质表达或用于目标磷酸化的化合物的最大产率。合适的培养条件包括约20℃至约45℃,优选地约30℃至约37℃,最优选地约37℃的培养温度。在培养细菌细胞的同时,可以搅拌容器以使培养物通气,可以以约100rpm至约250rpm,优选地约180rpm至约220rpm进行搅拌。
根据本发明的第五方面,可以使用标准培养方案,包括首先在固体琼脂平板上产生目标工程化细菌的单个菌落,然后产生接种有单个菌落的液体介质起始培养物,起始培养物可以然后用于接种更大体积的液体介质。最终培养物体积可以在约500毫升至约2000升,优选地在约500毫升至约500升,更优选地在约500毫升至50升的范围内。
许多不同的介质适合于培养细菌。如本文所用,术语“介质”是指可以在其中生长微生物的固体、液体或半固体支持物,例如凝胶。介质可以补充有另外的组分以改善细菌生长,例如氨基酸、糖、乳糖、葡萄糖、甘露糖、甘油、钙、镁、铁、磷酸盐、硫酸盐、氯化物、乙酸盐、柠檬酸盐等。介质还可以补充有抗生素或抗微生物剂,例如氨苄青霉素、四环素、卡那霉素、链霉素、羧苄青霉素、红霉素和氯霉素。可以使用这些抗微生物剂来选择性地培养特定细菌,其中细菌包含特定的抗生素抗性基因。可用于培养细菌的培养基的非限制性实例包括溶菌肉汤(lysogenybroth,lb)、极好肉汤(terrificbroth,tb)、2xyt肉汤,超级最佳肉汤(superoptimalbroth,sob)、具有降解物抑制的超级最佳肉汤(superoptimalbrothwithcataboliterepression,soc)、最小盐介质(m9)、nzcym肉汤、nz胺肉汤。
根据第五方面,细菌可以是革兰氏阴性细菌,例如假单胞菌、克雷伯菌、肠杆菌、幽门螺杆菌。在一个优选的实施方案中,将经过基因工程化的大肠杆菌细菌用于生产磷酸化的化合物。
培养基因工程化的细菌时,磷酸化的化合物经由磷酸转位器输出通过细菌膜。因此,培养基中存在目标化合物。在根据本发明的第五方面的实施方案中,该方法进一步包括从培养基中提取磷酸化的化合物的步骤。提取过程可以以各种方式进行,但是关键步骤是将细菌细胞从液体培养基中分离出来,这可以通过离心、过滤或沉淀来实现。一旦从细菌细胞中分离出了液体培养物,就可以使用标准技术,例如柱色谱法、高效液相色谱法、离子交换色谱法、薄层色谱法和反相色谱法纯化目标化合物。
根据本发明的第五方面的方法可以用于制造许多磷酸化的化合物。根据第五方面,该方法可以用于制造磷酸化的糖,例如磷酸二羟基丙酮(dhap)、脱氧木酮糖-5-磷酸、木酮糖-5-磷酸、葡萄糖-6-磷酸、果糖-6-磷酸、果糖-1,6-二磷酸、甘油醛-3-磷酸、甘油-3-磷酸、磷酸烯醇丙酮酸、2-磷酸甘油、3-磷酸甘油、1,3-二磷酸甘油酸。在一个优选的实施方案中,该方法用于制造dhap。
该方法可用于制造各种高价值的磷酸化的化合物。这些化合物可用作合成其他高价值化合物(如药物)的前体。特别地,该方法可用于生产dhap,然后可将其转化为甲基乙二醛。在一个实施方案中,根据本发明的第五方面的方法进一步包括将dhap转化为甲基乙二醛的步骤。dhap可以通过热降解转化为甲基乙二醛。为了将dhap转化为甲基乙二醛,可以将细菌培养物在大约25℃和大约45℃之间,更优选地在大约35℃和大约40℃之间,最优选地在37℃孵育。为了将dhap转化为甲基乙二醛,可以首先从细菌培养物中提取并纯化dhap,或者可以在从细菌培养物中提取并纯化之前将dhap转化为甲基乙二醛。
下列实施例示例说明了本发明。
实施例1-细菌培养
在基因操作期间和从甘油原液(30%(v/v),保持在-80℃)获取的细胞进行第一个起始培养期间,将细胞培养在溶菌(lb)培养基上,并铺板在lb琼脂上。向介质中添加1.5%的微生物琼脂制备lb琼脂。培养物的生长在m9基本介质中进行,该介质含有浓度为20g/l的(d)-葡萄糖以及18g/lna2hpo4·12h2o、3g/lkh2po4、0.5g/lnacl、2/lgnh4cl、0.5g/lmgso4·7h2o、0.015g/lcacl2·2h2o、0.010g/lfecl3、0.006g/l盐酸硫胺、1.2mg/lmnso4·h2o、1.2mg/lcucl2·2h2o。3-(n-吗啉)丙磺酸(mops)溶液(ph7)用于在过滤器灭菌后(merckmilliporeexpressplus)将m9基本介质缓冲至最终浓度为20g/l。将介质过滤灭菌或高压灭菌。需要时,将抗生素以100μg/ml的氨苄西林浓度添加到介质中。所有化学品均购自sigma-aldrich。
从冷冻的甘油储备液中接种起始培养物(在50ml试管中的10mllb介质(bdfalcon)),并培养过夜。将来自这些培养物的细胞用于在250ml摇瓶中以od~0.2接种50ml葡萄糖基础介质。所有培养物均在200rpm的旋转摇床(inforsht)上于37℃或30℃进行。通过使用分光光度计监测od600来跟踪细胞生长,并取出培养上清液用于酶测试和hplc分析。
实施例2-质粒和菌株构建
在整个研究中,大肠杆菌k-12substr.mg1655用作亲本菌株。δtpi缺失突变体通过根据miller[miller等人,1992]修改的噬菌体转导方法产生。噬菌体裂解物从keio保藏的菌株制备[baba等人,2006],这些菌株携带单个缺失。
通过genscript化学地合成了携带磷酸三糖/磷酸转位器的质粒。骨架是具有氨苄青霉素抗性基因的puc57质粒。转位器基因tpt已针对大肠杆菌进行了密码子优化,并且与在枯草芽孢杆菌中发现的称为mistic的膜整合蛋白产生融合。转位器和mistic蛋白的融合蛋白的氨基酸序列示于seqidno3,并且相对应的核苷酸序列示于seqidno4。向序列中添加规范核糖体结合位点(rbs)和来自galp的组成型启动子。在5'和3'末端分别添加了两个限制性位点kpni和hindiii。
使用标准方案进行转化[sambrook等人,1989]。使用来自[chung等人,1989]的一步方案制备化学感受态细胞(chimio-competentcell)。使用50μg/ml氨苄青霉素在lb平板上于37℃过夜来选择转化的细胞。
为了确定基因工程化对大肠杆菌细胞生长的影响,在m9基本介质中进行了生长测定法。根据实施例1中所述的方法培养大肠杆菌δtpi突变体,并随时间测量培养物的od(光密度)。在相同条件下,也培养用包含磷酸三糖/磷酸转位器的质粒转化的大肠杆菌δtpi突变体,并随时间测量od。该实验的结果显示在图1中,并证明δtpi和转位器表达的组合导致更高的生长水平,如较高的od值所示。由于dhap的输出,有可能实现生长水平的提高。当dhap不输出并且不被磷酸三糖异构酶降解时,dhap将转化为对细胞有毒的甲基乙二醛。
seqidno1-来自拟南芥的磷酸三糖/磷酸转位器(tpt)的氨基酸序列。
mesrvllratanvvgipklrrpigaihrqfstassssfsvkpiggigeganlisgrqlrpillldssainggekreilkpvkaaaaeggdtagdakvgflakypwlvtgffffmwyflnvifnilnkkiynyfpypyfvsvihlfvgvvycliswsvglpkrapidsnllkvlipvavchalghvtsnvsfaavavsfthtikalepffnaaasqfimgqsipitlwlslapvvlgvamasltelsfnwlgfisamisnisftyrsifskkamtdmdstnvyayisiialfvcippaiivegpkllnhgfadaiakvgmtkfisdlfwvgmfyhlynqlatntlervaplthavgnvlkrvfvigfsivifgnkistqtgigtgiaiagvamysiikakieeekrqgkka
seqidno2-来自枯草芽孢杆菌的mistic的氨基酸序列
mkvtsdekeqlsaaidrmnegldvfiqfynesekdepliqleddtaklmkearelygqdklnetlnaiikqilsislseegkkk
seqidno3-mistic和tpt由带下划线的接头序列连接的融合蛋白的氨基酸序列
mkvtsdekeqlsaaidrmnegldvfiqfynesekdepliqleddtaklmkearelygqdklnetlnaiikqilsislseegkkksssnnnnnlgsngsgmesrvllratanvvgipklrrpigaihrqfstassssfsvkpiggigeganlisgrqlrpillldssainggekreilkpvkaaaaeggdtagdakvgflakypwlvtgffffmwyflnvifnilnkkiynyfpypyfvsvihlfvgvvycliswsvglpkrapidsnllkvlipvavchalghvtsnvsfaavavsfthtikalepffnaaasqfimgqsipitlwlslapvvlgvamasltelsfnwlgfisamisnisftyrsifskkamtdmdstnvyayisiialfvcippaiivegpkllnhgfadaiakvgmtkfisdlfwvgmfyhlynqlatntlervaplthavgnvlkrvfvigfsivifgnkistqtgigtgiaiagvamysiikakieeekrqgkka
seqidno4-大肠杆菌密码子优化的核苷酸序列,编码通过接头序列连接的mistic和tpt的融合蛋白
实施例3-酶促测定法
为了确定输出到介质中的dhap的量,使用酶促测定法分析了来自细菌培养物的上清液。dhap浓度如下确定。磷酸三糖异构酶(tpi)将一分子的dhap转化为一分子的甘油醛-3-磷酸(gap)。然后通过nadh的形成来测量gap脱氢酶(gapdh)对gap的还原作用。nadh在340nm处吸收。反应混合物250μl在1ml中包含1utpi、0.1ugapdh、50mmhepes(ph7.98)、1mm二硫苏糖醇(dtt)、1mmnad+、2mm硫胺素焦磷酸和200μl培养上清液。反应于37℃进行10分钟。通过分光光度计于22℃在340nm处进行测定法。
图2显示了在其中培养δtpitpt菌株的培养基和其中培养对照菌株的培养基上进行的测定法结果。在δtpitpt菌株的培养基中检测到的dhap量是对照的约4.5倍高。
实施例4-对磷酸化的产物的分析
为了确认对由细菌产生的产物的鉴别,进行了结构分析。使用与agilent6490三重四极式lc/ms系统耦合的agilent1260infinityiilc系统分析dhap。在等温条件下(35℃)使用sequantzichilic(merck,100x2.1mm,3.5μm膜厚,100a°),并使用二元梯度进行色谱法。溶剂a为10mm的碳酸氢铵水溶液,ph9.0(使用氢氧化铵溶液调节ph),溶剂b为乙腈。过滤后的样品(通过0.45μm过滤器)的注射体积为5μl并且使用0.5ml/min的流速。表i和表ii中列出了用于分离的梯度和三重四极式检测器条件。仪器控制、数据采集、处理和报告生成是通过agilentmasshunterworkstationdataacquisition完成的。在去离子水(d.i.水,18.2mω-cm-1,millipore)-乙腈溶液中制备用于单个分析物的所有储备标准溶液。注射的标准是浓度约为0.1ng/ml。
表1-hplc梯度和参数
表2.mrm操作模式下的三重四极式检测器条件
在其他实施方案中,如以下实施例中所述,该方法可用于生产和输出甘油-3-磷酸。为了输出甘油-3-磷酸,细菌经基因修饰以表达磷酸三糖/磷酸转位器。任选地,制得转位器为与微生物膜整合蛋白的融合蛋白。在优选的实施方案中,细菌表达seqidno3的氨基酸序列。磷酸三糖/磷酸转位器的表达可以通过用编码该转位器的载体转化细菌来实现。载体可以包含seqidno4的核苷酸序列。
实施例5-通过tpt转位器输出甘油-3-磷酸
为了评估磷酸三糖转位器是否允许输出甘油-3-磷酸,将dtpi菌株如实施例1中所述在m9介质中培养,但是用甘油和木糖代替葡萄糖。在固定相期间,取出培养基并通过质谱分析。如图6所示,甘油-3-磷酸存在于培养基中,表明甘油-3-磷酸通过转位器输出。
为了增加产量,用携带磷酸三糖转位器基因的质粒转化来自keio保藏的甘油3-磷酸脱氢酶(glpd)突变体[baba等人,2006]。使用标准方案进行转化[sambrook等人,1989]。使用来自[chung等人,1989]的一步方案制备化学感受态细胞。使用50μg/ml氨苄青霉素在lb平板上于37℃过夜来选择转化的细胞。
该菌株在含有1%木糖和1%甘油的m9介质上培养。在那种情况下,在固定相期间在培养基中存在甘油-3-磷酸为33.47mg/l,而在相同条件下培养没有转位器的简单突变体时,情况并非如此。
这表明在glpd突变体中tpt转位器的表达允许甘油-3-磷酸的外部产生。
在其他实施方案中,如以下实施例中所述,该方法可用于生产和输出木酮糖-5-磷酸。为了输出木酮糖-5-磷酸,细菌经基因修饰以表达木糖-5-磷酸转位器。任选地,制得转位器为与微生物膜整合蛋白的融合蛋白。可以通过用编码该转位器的载体转化细菌来实现木糖-5-磷酸转位器的表达。载体可以包含seqidno5的核苷酸序列。
实施例6-通过来自拟南芥的木糖-5-磷酸转位器输出木酮糖-5-磷酸
在整个研究中,大肠杆菌k-12substr.mg1655用作亲本菌株。δtkta缺失突变体通过根据miller[miller等人,1992]修改的噬菌体转导方法产生。噬菌体裂解物从keio保藏的菌株制备[baba等人,2006],这些菌株携带单个缺失。
通过genscript化学地合成携带木酮糖-5-磷酸/磷酸转位器的质粒(eicks等人,2002)。骨架是具有氨苄青霉素抗性基因的puc57质粒。转位器基因xpt针对大肠杆菌进行了密码子优化,并且与在枯草芽孢杆菌中发现的称为mistic的膜整合蛋白产生融合。转位器和mistic蛋白的融合蛋白的核酸序列示于seqidno5。向序列中添加规范核糖体结合位点(rbs)和来自galp的组成型启动子。在5'和3'末端分别添加了两个限制性位点bamhi和xbai。
使用标准方案进行转化[sambrook等人,1989]。使用来自[chung等人,1989]的一步方案制备化学感受态细胞。使用50μg/ml氨苄青霉素在lb平板上于37℃过夜来选择转化的细胞。
seqidno5-大肠杆菌密码子优化的核苷酸序列,编码融合蛋白mistic和转位器基因xpt
ctggataactttacgggcatgcataaggctcgtataatatattcaggaggtaaataatgatcagcctgaacctgagcccgagcctgaacccgggtctgctgcacaagacccgtacctgccaacaaccgacccgtctgagcgcgctgctggttaccaacccgaaaccgttcaaccatcgtcacccgctgggcctgagcccgatcccgaacctgcagattcgtgatgtgagcgcgaagccgctgctgagcctgaccaacccggagagcagcagcggttttagccgtaaaccgcgtagcattgcggcggttggcagcagcgacagcaacccggatgagaagagcgacctgggcgaggcggaaaagaaagaaaagaaagcgaaaaccctgcagctgggtattgttttcggcctgtggtacttccaaaacatcgtgttcaacatcttcaacaagaaagcgctgaacgttttcccgtatccgtggctgctggcgagcttccaactgtttgcgggcagcatctggatgctggtgctgtggagctttaagctgtacccgtgcccgaagattagcaaaccgttcatcattgcgctgctgggtccggcgctgtttcacaccatcggccacattagcgcgtgcgttagcttcagcaaggttgcggtgagctttacccacgtgatcaaaagcgcggagccggttttcagcgtgatttttagcagcctgctgggtgatagctacccgctggcggtttggctgagcatcctgccgattgtgatgggctgcagcctggcggcggttaccgaagtgagcttcaacctgggtggcctgagcggtgcgatgatcagcaacgttggcttcgtgctgcgtaacatttatagcaagcgtagcctgcagagctttaaagagatcgacggtctgaacctgtacggctgcatcagcattctgagcctgctgtacctgttcccggttgcgatctttgtggaaggtagccactgggttccgggctaccacaaggcgattgcgagcgtgggtaccccgagcaccttctatttttgggttctgctgagcggcgtgttctaccacctgtataaccagagcagctaccaagcgctggatgagatcagcccgctgaccttcagcgttggtaacaccatgaaacgtgtggttgtgatcattagcaccgttctggtgtttcgtaacccggtgcgtccgctgaacgcgc
tgggtagcgcgattgcgattttcggcacctttctgtatagccaagcgaccgcgaagaaaaagaaaattgaagtgggtggcgacaagaaaaac
为了确定基因工程化对大肠杆菌细胞生长的影响,在m9基本介质中进行了生长测定法。根据实施例1中所述的方法培养通过xpt-puc19质粒转化的大肠杆菌δtkta突变体,不同之处在于用木糖和甘油代替葡萄糖。在固定相期间取出培养物上清液,并通过质谱分析。
图7表明在培养基中检测到木酮糖-5-磷酸的量约为5mg/l,并且检测到dhap。这表明在突变体中xpt转位器的表达允许木酮糖-5-磷酸的外部产生。
序列表
<110>bioc3
<120>多肽
<130>bio-pat001wo
<160>5
<170>patentin版本3.5
<210>1
<211>410
<212>prt
<213>拟南芥
<400>1
metgluserargvalleuleuargalathralaasnvalvalglyile
151015
prolysleuargargproileglyalailehisargglnpheserthr
202530
alaserserserserpheservallysproileglyglyileglyglu
354045
glyalaasnleuileserglyargglnleuargproileleuleuleu
505560
aspserseralaileasnglyglyglulysarggluileleulyspro
65707580
vallysalaalaalaalagluglyglyaspthralaglyaspalalys
859095
valglypheleualalystyrprotrpleuvalthrglyphephephe
100105110
phemettrptyrpheleuasnvalilepheasnileleuasnlyslys
115120125
iletyrasntyrpheprotyrprotyrphevalservalilehisleu
130135140
phevalglyvalvaltyrcysleuilesertrpservalglyleupro
145150155160
lysargalaproileaspserasnleuleulysvalleuileproval
165170175
alavalcyshisalaleuglyhisvalthrserasnvalserpheala
180185190
alavalalavalserphethrhisthrilelysalaleugluprophe
195200205
pheasnalaalaalaserglnpheilemetglyglnserileproile
210215220
thrleutrpleuserleualaprovalvalleuglyvalalametala
225230235240
serleuthrgluleuserpheasntrpleuglypheileseralamet
245250255
ileserasnileserphethrtyrargserilepheserlyslysala
260265270
metthraspmetaspserthrasnvaltyralatyrileserileile
275280285
alaleuphevalcysileproproalaileilevalgluglyprolys
290295300
leuleuasnhisglyphealaaspalailealalysvalglymetthr
305310315320
lyspheileseraspleuphetrpvalglymetphetyrhisleutyr
325330335
asnglnleualathrasnthrleugluargvalalaproleuthrhis
340345350
alavalglyasnvalleulysargvalphevalileglypheserile
355360365
valilepheglyasnlysileserthrglnthrglyileglythrgly
370375380
ilealailealaglyvalalamettyrserileilelysalalysile
385390395400
glugluglulysargglnglylyslysala
405410
<210>2
<211>84
<212>prt
<213>枯草芽孢杆菌
<400>2
metlysvalthrseraspglulysgluglnleuseralaalaileasp
151015
argmetasngluglyleuaspvalpheileglnphetyrasngluser
202530
glulysaspgluproleuileglnleugluaspaspthralalysleu
354045
metlysglualaarggluleutyrglyglnasplysleuasngluthr
505560
leuasnalaileilelysglnileleuserileserleusergluglu
65707580
glylyslyslys
<210>3
<211>509
<212>prt
<213>合成
<400>3
metlysvalthrseraspglulysgluglnleuseralaalaileasp
151015
argmetasngluglyleuaspvalpheileglnphetyrasngluser
202530
glulysaspgluproleuileglnleugluaspaspthralalysleu
354045
metlysglualaarggluleutyrglyglnasplysleuasngluthr
505560
leuasnalaileilelysglnileleuserileserleusergluglu
65707580
glylyslyslysserserserasnasnasnasnasnleuglyserasn
859095
glyserglymetgluserargvalleuleuargalathralaasnval
100105110
valglyileprolysleuargargproileglyalailehisarggln
115120125
pheserthralaserserserserpheservallysproileglygly
130135140
ileglygluglyalaasnleuileserglyargglnleuargproile
145150155160
leuleuleuaspserseralaileasnglyglyglulysarggluile
165170175
leulysprovallysalaalaalaalagluglyglyaspthralagly
180185190
aspalalysvalglypheleualalystyrprotrpleuvalthrgly
195200205
phephephephemettrptyrpheleuasnvalilepheasnileleu
210215220
asnlyslysiletyrasntyrpheprotyrprotyrphevalserval
225230235240
ilehisleuphevalglyvalvaltyrcysleuilesertrpserval
245250255
glyleuprolysargalaproileaspserasnleuleulysvalleu
260265270
ileprovalalavalcyshisalaleuglyhisvalthrserasnval
275280285
serphealaalavalalavalserphethrhisthrilelysalaleu
290295300
gluprophepheasnalaalaalaserglnpheilemetglyglnser
305310315320
ileproilethrleutrpleuserleualaprovalvalleuglyval
325330335
alametalaserleuthrgluleuserpheasntrpleuglypheile
340345350
seralametileserasnileserphethrtyrargserilepheser
355360365
lyslysalametthraspmetaspserthrasnvaltyralatyrile
370375380
serileilealaleuphevalcysileproproalaileilevalglu
385390395400
glyprolysleuleuasnhisglyphealaaspalailealalysval
405410415
glymetthrlyspheileseraspleuphetrpvalglymetphetyr
420425430
hisleutyrasnglnleualathrasnthrleugluargvalalapro
435440445
leuthrhisalavalglyasnvalleulysargvalphevalilegly
450455460
pheserilevalilepheglyasnlysileserthrglnthrglyile
465470475480
glythrglyilealailealaglyvalalamettyrserileilelys
485490495
alalysileglugluglulysargglnglylyslysala
500505
<210>4
<211>1530
<212>dna
<213>合成
<400>4
atgaaagttacctccgacgaaaaagaacaattatcggcagccatcgaccgcatgaatgag60
ggtcttgacgttttcattcagttctataacgagagtgaaaaagatgagccgctgatccaa120
ctggaggatgacaccgctaaactcatgaaggaagcacgtgaattgtatggccaggacaaa180
ttgaacgagaccctgaacgccatcattaaacaaattctgagtattagcctgagcgaagaa240
ggaaaaaaaaaatcttcttcaaacaataacaacaaccttgggtcaaatggtagcggaatg300
gagagccgtgtgctgttgcgtgcgacagctaatgtcgtcggcattccgaaactgcgtcgc360
ccgatcggtgccatccaccgtcaattttcaacggccagctcgagctcgtttagcgttaaa420
ccaattggcggcatcggcgaaggcgcaaacctgatcagcggccgccagttacgcccgatt480
cttctgcttgattcttcagcaatcaatgggggcgaaaagcgcgaaattctgaagcccgta540
aaagcggcagcggcggaaggcggcgacacggctggggacgcgaaagtcgggtttttggcg600
aaatatccgtggctggtcaccggcttcttttttttcatgtggtatttccttaacgtgatc660
tttaacattctgaacaaaaaaatctataattactttccgtatccgtacttcgtatctgtg720
atccacctctttgtaggcgttgtgtattgtttaattagctggagcgtgggcctgccgaaa780
cgtgctccaattgactctaacctgctgaaagtccttattccggtggcggtgtgccatgcc840
ctcggccatgtaaccagcaacgtttctttcgcggcagttgccgtttcgtttacacacact900
atcaaagcattggagccattttttaatgccgcggcgtcgcaatttatcatgggccagtct960
atcccaattaccctgtggctgagtctggcaccggtcgtgcttggtgttgctatggcatct1020
ctgacagagctgtcatttaactggctgggctttattagcgccatgattagcaatatcagc1080
tttacgtaccgttcaatcttttccaaaaaagcaatgacggacatggattctactaatgtg1140
tatgcatacattagcattatcgccttattcgtctgcatcccgccagctatcattgtagag1200
ggcccgaaattgttaaaccatgggtttgcagatgcgattgcaaaagtgggcatgacgaaa1260
tttatttcagatctgttctgggtgggtatgttctaccacctctacaaccagcttgcgact1320
aacactctggaacgcgtggcgccgctgacccatgctgtgggtaatgtgttgaaacgtgtt1380
ttcgtcattggtttctctatcgtcattttcgggaacaaaattagtacccaaacgggtatt1440
ggcactggcatcgccatcgcaggcgttgcgatgtacagcattatcaaagcaaaaatcgaa1500
gaagaaaaacgccaagggaaaaaagcatag1530
<210>5
<211>1307
<212>dna
<213>合成
<400>5
ctggataactttacgggcatgcataaggctcgtataatatattcaggaggtaaataatga60
tcagcctgaacctgagcccgagcctgaacccgggtctgctgcacaagacccgtacctgcc120
aacaaccgacccgtctgagcgcgctgctggttaccaacccgaaaccgttcaaccatcgtc180
acccgctgggcctgagcccgatcccgaacctgcagattcgtgatgtgagcgcgaagccgc240
tgctgagcctgaccaacccggagagcagcagcggttttagccgtaaaccgcgtagcattg300
cggcggttggcagcagcgacagcaacccggatgagaagagcgacctgggcgaggcggaaa360
agaaagaaaagaaagcgaaaaccctgcagctgggtattgttttcggcctgtggtacttcc420
aaaacatcgtgttcaacatcttcaacaagaaagcgctgaacgttttcccgtatccgtggc480
tgctggcgagcttccaactgtttgcgggcagcatctggatgctggtgctgtggagcttta540
agctgtacccgtgcccgaagattagcaaaccgttcatcattgcgctgctgggtccggcgc600
tgtttcacaccatcggccacattagcgcgtgcgttagcttcagcaaggttgcggtgagct660
ttacccacgtgatcaaaagcgcggagccggttttcagcgtgatttttagcagcctgctgg720
gtgatagctacccgctggcggtttggctgagcatcctgccgattgtgatgggctgcagcc780
tggcggcggttaccgaagtgagcttcaacctgggtggcctgagcggtgcgatgatcagca840
acgttggcttcgtgctgcgtaacatttatagcaagcgtagcctgcagagctttaaagaga900
tcgacggtctgaacctgtacggctgcatcagcattctgagcctgctgtacctgttcccgg960
ttgcgatctttgtggaaggtagccactgggttccgggctaccacaaggcgattgcgagcg1020
tgggtaccccgagcaccttctatttttgggttctgctgagcggcgtgttctaccacctgt1080
ataaccagagcagctaccaagcgctggatgagatcagcccgctgaccttcagcgttggta1140
acaccatgaaacgtgtggttgtgatcattagcaccgttctggtgtttcgtaacccggtgc1200
gtccgctgaacgcgctgggtagcgcgattgcgattttcggcacctttctgtatagccaag1260
cgaccgcgaagaaaaagaaaattgaagtgggtggcgacaagaaaaac1307
- 还没有人留言评论。精彩留言会获得点赞!