表达人源化抗体的非人转基因动物及其用途
本申请为分案申请,原申请的申请日为2009年11月30日,申请号为200980151815.7(pct/gb2009/002781),发明名称为“表达人源化抗体的非人转基因动物及其用途”。
发明领域
本发明涉及使用转基因非人哺乳动物响应抗原攻击而衍生和选择各种功能性亲和力成熟四聚体免疫球蛋白(包含重链和轻链)库的改进方法及其用途。
具体地讲,本发明涉及经过工程改造使得其产生小鼠内源κ和/或λ轻链免疫球蛋白的能力大大降低或者轻链与重链复合的能力降低、消除或被阻断的非人哺乳动物,优选小鼠。本发明的非人哺乳动物产生小鼠功能性内源重链的能力也下降。因此,其形成包含由所述突变基因座产生的重排重链和轻链的功能性免疫球蛋白四聚体的能力已大大降低或消除。本发明还披露了产生这类哺乳动物的方法以及利用这类哺乳动物使用免疫球蛋白重链和轻链转基因产生人四聚体抗体和杂交四聚体抗体的方法。
在下面的说明中,所有氨基酸残基的位置号根据由kabat等人((1991),美国公共卫生署(uspublichealthservices)出版物第91-3242号)设计的编号系统给出。
发明背景
抗体
抗体的结构是本领域众所周知的。大多数天然抗体是四聚体,包含2条重链和2条轻链。重链通过大致位于每条重链中部的铰链结构域之间的二硫键彼此连接。轻链在铰链结构域的n端侧与各重链缔合。每条轻链通常通过接近铰链结构域的二硫键与其对应的重链结合。
当抗体分子正确折叠时,每条链折叠成许多通过较为线性的多肽序列连接的独特的球状结构域。例如,轻链折叠成可变(vl)结构域和恒定(cκ或cλ)结构域。重链具有单个可变结构域vh、第一恒定结构域(ch1)、铰链结构域和两个或三个另外的恒定结构域。重链的恒定结构域和铰链结构域一起形成通常所称的抗体重链恒定区。重(vh)链和轻(vl)链可变结构域的相互作用导致形成抗原结合区(fv)。重链的ch1结构域和轻链的cκ或cλ结构域有利于重链和轻链的相互作用。一般而言,抗原结合既需要vh又需要vl,但研究已表明重链二聚体和氨基端片段在轻链不存在时保持活性(jaton等(1968)biochemistry,7,4185-4195)。通常循环λ轻链的比例低下,可能占血浆中复合成四聚体免疫球蛋白的全部轻链的2-5%(goldsby等(2003)immunology,第5版,w.h.freeman&cony)。
体外操纵重链免疫球蛋白基因以构建新的抗体最早披露于二十世纪八十年代。大多数早期抗体工程研究使用针对充分表征的抗原产生的重排小鼠免疫球蛋白μ基因(igm)。该抗体的特征是已知存在于vh结构域的抗原结合特异性,因为不相关轻链的装配和分泌显示保持抗原结合(neuberger和williams(1986)phil.trans.r.soc.lond.,a317,425-432)。使用该系统显示,可使用小鼠抗原特异性vh结合结构域获得包含与小鼠抗原特异性vh结构域融合的人ε恒定效应子区的新抗体。所得杂交ige保持抗原特异性,并显示ige的预期效应子活性(neuberger等(1985)nature,314,268-270)。重链工程的其它文献实例包括:编码保持抗磷酸胆碱活性的小鼠vh人/iga或igg抗体融合体的杂交小鼠-人抗体基因(morrison等(1984)pnas,81,6851-6855);包含小鼠vh和人igg(γ3)恒定区的抗癌相关抗原17-1a抗体(sun等(1987)pnas,84,214-218);以及包含人igg(γ1)恒定区和小鼠vh结构域的抗人t细胞抗体(抗cd7)(参见heinrich等(1989)j.immunol.,143,3589-97)。
正常人b细胞在14号染色体上含有单个免疫球蛋白重链基因座,通过重排从中产生编码重链的基因。在小鼠中,重链基因座位于12号染色体上。正常的重链基因座包含多个v基因区段、许多d基因区段和许多j基因区段。大多数vh结构域由v基因区段编码,但是每个vh结构域的c末端由d基因区段和j基因区段编码。vdj在b细胞中重排,接着亲和力成熟,为每个vh结构域提供其抗原结合特异性。对从包含单个v区段的重链免疫球蛋白得到的正常h2l2四聚体进行的序列分析表明,响应抗原攻击的多样性主要由vdj重排和体细胞高变的组合引起(xu和davies(2000)immunity,13,37-45)。超过50个的人v基因区段存在于人基因组中,其中只有39个具有功能。在正常的二倍体产抗体b细胞中,各细胞从一组重链和轻链抗体基因座产生抗体四聚体(h2l2)。由于称为等位基因排斥的过程所致,另一组基因座的使用是非生产性的(singh等(2003)j.exp.med.,197,743-750及其中的参考文献)。
现可从响应抗原攻击的转基因小鼠中得到全人抗体(h2l2)。这类转基因小鼠一般包含单个人重链免疫球蛋白基因座和分离的人轻链免疫球蛋白基因座。相应的内源小鼠重链、κ轻链和任选λ轻链基因座编码序列缺失或部分缺失。因此,在低本底的包含人重链和小鼠λ轻链的小鼠/人抗体的情况下只产生包含κ轻链的人抗体(wo90/04036;wo93/12227;wo98/24893;us5877397、us5814318和us6162963)。已实现缺失所有内源鼠重链和轻链免疫球蛋白基因的区段以完全排除内源重链和轻链基因的表达,但技术上的难度仍然存在,特别是如果剔除所有λ轻链编码序列被认为是必需的。已经通过完全缺失所有功能性v和j基因区段以及λ基因座的c1、c2和c3恒定区而达到剔除鼠λ轻链编码序列,产生具有沉默λ轻链基因座的小鼠(参见ep1399559)。
一种不同的方法是通过破坏编码鼠重链的基因的膜外显子(membraneexon)来限制小鼠b细胞发育和免疫球蛋白分泌。因此,虽然内源鼠重链基因由于在响应抗原攻击时被转录并进行vdj重排而具有功能,但是因为igm从不在前b细胞的细胞表面上表达,所以进一步的发育受到抑制,导致对抗原攻击非生产性的应答(kitamura等(1991)nature,350,423-426),即使内源小鼠κ和λ轻链基因两者仍保持结构完整和功能性也是如此(tuaillon(2000)molecularimmunology,37,221-231)。
在内源小鼠重链和轻链基因座保持功能的情况下,任何其它引入的免疫球蛋白重链转基因也受等位基因排斥的调节,因此一些b细胞只功能性表达小鼠重链和轻链基因座,而其它b细胞只功能性表达人重链基因座和小鼠轻链基因座(nussenzweig等(1987)science,236,816-819)。在任何单个非人转基因动物中存在高度多样化的表达抗体的b细胞群,所述抗体响应不同抗原攻击而来源于可能所有免疫球蛋白基因座。采用已确立的杂交瘤技术,利用hat选择进行随后的抗原特异性抗体的选择(davis等(1982)j.immunol.methods,50,161-171)无法区分表达一种重链免疫球蛋白基因座的杂交瘤与表达另一种重链免疫球蛋白基因座的杂交瘤。
免疫球蛋白重链转基因中存在的调节元件包含必需的组织特异性增强子元件以确保以拷贝数依赖性方式进行b细胞特异性表达。5’内含增强子和3’lcr的存在确保了转基因在b细胞成熟的所有阶段都有活性(guglielmi等(2003)biochim.biophys.acta,1642,181-190)。在转基因座中加入重链和轻链特异性lcr确保不仅表达是b细胞特异性的,而且不论整合入基因组的位点都进行表达(wo90/10077,mills等(1997)j.exp.med.,186,845-858和pettersson等(1997)immunobiol.,198,236-248))。因此,假若存在lcr,则每个转基因不论其在基因组中的位置都具有功能。在存在于转基因的lcr部分缺失的情况下,转基因周围的染色质在任何时间点只是部分对转录开放,导致位置效应镶嵌表达(mosaicexpression),因此转基因跨靶组织的表达水平受限(festenstein等(1996)science,23,271(5252):1123-5;milot等(1996)cell,87(1),105-14)。
在小鼠背景中产生人免疫球蛋白的备选方法是用人的同源基因区段置换鼠免疫球蛋白基因区段。因此,如果只是小鼠v、d和j基因区段被人同源物置换,则在抗原攻击后可形成包含人vh和vl结构域和小鼠恒定(效应子)区的功能性小鼠/人杂交抗体(wo94/04667)。如果所有鼠基因区段都被人同源物置换,则在抗原攻击后将产生完整的人免疫球蛋白(us6596541)。观察到该方法的一个优势是,如果只交换编码区,则所得到的转基因保留所有小鼠调节元件,从而确保了对抗原攻击的最大程度的应答。该方法根据转基因的最终结构,提供高血清效价的高亲和力人抗体或小鼠/人杂交抗体。然而,实际上,通过同源重组置换小鼠基因组中所有单独的v、d和j区段是一项长时间的艰巨任务。同样,构建包含所有39个功能性人v、d和j区段与恒定(效应子)区的重链转基因在技术上非常费力。
因此,在本领域中仍需要不依赖于基因组dna大量区段缺失或多重缺失的方法,这允许(i)简化小鼠的产生,该小鼠在响应抗原攻击时以b细胞特异性的方式表达内源重链和轻链免疫球蛋白基因的能力大大降低,或者在抗原攻击后在b细胞中表达内源免疫球蛋白重链和/或轻链基因,但是编码缺乏装配成为功能性免疫球蛋白四聚体的能力的免疫球蛋白重链和轻链蛋白,导致对抗原攻击的非生产性应答;(ii)用于构建并b细胞特异性表达多个重链转基因座的可再现的简化方法,所述重链转基因座尽管可总共包含所有39个人v基因区段,但各自包含优选的较小的v基因区段群,各与所有d和j基因区段以及一些或全部恒定(效应子)区组合,其功能性表达是抗原依赖性的,并且最终由等位基因排斥确定;以及(iii)具有这样的能力:选择排除表达残余内源小鼠免疫球蛋白的杂交瘤,并选出表达和分泌装配的包含存在于重链转基因座上的完整v区段库的免疫球蛋白四聚体的杂交瘤,或者选出表达存在于与另一重链转基因座相对的重链转基因座上的一亚组v基因区段的杂交瘤。
发明详述
根据本发明第一个方面,提供含有内源λ轻链基因座、内源κ轻链基因座和内源重链基因座的非人哺乳动物,每个基因座可重排使得在抗原攻击后在b细胞中形成并表达免疫球蛋白重链和轻链基因,但是所述基因座已经突变使得形成包含从所述突变基因座产生的重排重链和轻链的功能性免疫球蛋白四聚体的能力已大大降低或消除。
在非人哺乳动物中,可通过将移码突变、多肽编码序列和/或一个或多个终止密码子引入某内源基因座或各内源基因座中,使内源λ轻链基因座、内源κ轻链基因座和内源重链基因座的至少一个已发生突变。
突变优选为插入小于50个核苷酸。
在非人哺乳动物中,可通过剔除某基因座或各基因座的部分或全部lcr,使内源λ轻链基因座、内源κ轻链基因座和内源重链基因座的至少一个的表达已大大减少。
优选引入位于内源κ轻链基因座上。
备选地,引入位于内源λ轻链基因座上。
在另一备选中,引入位于内源κ轻链基因座和内源λ轻链基因座上。
引入还可位于内源重链基因座上。
在进一步的备选中,引入可位于内源κ轻链基因座和内源重链基因座上。
在又进一步的备选中,引入可位于内源λ轻链基因座和内源重链基因座上。
在更进一步的备选中,引入可位于内源λ轻链基因座、内源κ轻链基因座和内源重链基因座上。
优选在内源κ轻链基因座上有如上限定的引入,并且在内源λ基因座上有部分或完整的如上限定的lcr缺失。
如有需要,可使内源重链基因座突变,使得发生重链基因重排、mrna转录和蛋白质合成,但是阻断b细胞活化。
优选如上定义的非人哺乳动物包含含有一个或多个异源κ轻链基因座和相关b细胞特异性调节元件的转基因。
非人哺乳动物还可包含含有一个或多个异源λ轻链基因座和相关b细胞特异性调节元件的转基因。
在如上定义的非人哺乳动物中,含有异源轻链基因座的转基因可包含显性选择标记基因。
如上定义的非人哺乳动物还可包含含有一个或多个异源重链基因座和相关b细胞特异性调节元件的转基因。
如有需要,非人哺乳动物可包含两个或更多个含有两个或更多个不同的异源重链基因座和相关b细胞特异性调节元件的转基因。
在非人哺乳动物中,含有异源重链基因座的某转基因或每个转基因可包含显性选择标记基因。
在非人哺乳动物中,每个异源重链基因座可包含ctcf结合位点。
优选非人哺乳动物包含含有异源κ轻链基因座的转基因和含有一个或多个异源重链基因座的转基因。
备选地,非人哺乳动物可包含含有异源λ轻链基因座的转基因和含有一个或多个异源重链基因座的转基因。
在进一步的备选中,非人哺乳动物可包含含有异源κ轻链基因座的转基因、含有λ轻链基因座的转基因和含有一个或多个异源重链基因座的转基因。
优选每个异源基因座整合了关联lcr。
每个异源基因座优选为人基因座。
然而,每个异源基因座可以是包含来源于多于一种物种的可变区和恒定区的杂交基因座,例如包含人可变区和大鼠或鼠恒定区的杂交基因座。
非人哺乳动物可包含含有不同群的不同异源重链基因座的转基因群,其中每个转基因群包含不同的显性选择标记基因。
备选地,非人哺乳动物可包含含有异源轻链基因座的转基因和含有异源重链基因座的转基因,其中含有异源轻链基因座的转基因和含有异源重链基因座的转基因各自包含不同的显性选择标记基因。
非人哺乳动物优选为啮齿动物,例如小鼠。
按照第二个方面,本发明提供产生抗原特异性异源单克隆抗体的方法,所述方法包括:
(a)用抗原对前述权利要求中任一项的非人转基因哺乳动物进行免疫;
(b)从免疫的转基因哺乳动物的b细胞制备分别产生单克隆抗体的杂交瘤或无限增殖化b细胞系;
(c)任选地,通过使用存在于包含异源免疫球蛋白轻链和重链基因座的转基因上的显性选择标记基因来选择至少一种表达异源抗体的杂交瘤或无限增殖化b细胞系;以及
(d)选择至少一种产生与抗原特异性结合的抗体的杂交瘤或无限增殖化b细胞系。
按照本发明又一个方面,提供由杂交抗体得到哺乳动物(优选人)的抗体的方法,所述方法包括:
(a)进行上述方法;
(b)选择至少一种产生特异性结合抗原并包含选定物种的vh和vl结合结构域的抗体的杂交瘤或无限增殖化b细胞系;
(c)对vh和vl结构域进行克隆和测序;
(d)使选出的包含vh和vl结合结构域编码序列的序列与来自同一物种的选定的恒定效应子结构域再克隆;和
(e)在选定的细胞类型中使用表达载体共表达再克隆的编码所需物种的重链和轻链多肽的序列。
按照本发明又一个方面,提供用于产生如上定义的非人哺乳动物的方法,所述方法包括使哺乳动物祖细胞中的内源重链基因座、内源κ轻链基因座和任选内源λ轻链基因座突变,并由所述祖细胞产生哺乳动物,其中突变使得:在哺乳动物中,每个基因座可以重排,使得在抗原攻击后在b细胞中形成并表达免疫球蛋白重链和轻链基因,但是形成包含由所述突变基因座产生的重排重链和轻链的功能性免疫球蛋白四聚体的能力已大大降低或消除。
优选祖细胞为非人胚胎干细胞。
非人哺乳动物优选为啮齿动物,例如小鼠。
本发明还提供使用如上定义的非人哺乳动物或方法得到的优选人的抗原特异性异源功能性免疫球蛋白四聚体作为药物、诊断剂或试剂的用途。
本发明的发明人通过开发用于产生非人哺乳动物(特别是小鼠)的简化方法,预料不到地克服了现有技术的局限,其中通过由优选在κ恒定区和/或λ恒定区中导致移码或mrna翻译过早终止的小插入事件引起恒定区非功能性,或者通过部分或全部剔除关联内源lcr,使内源κ和/或λ轻链基因的功能性表达大大减少。这与需要通过完全或部分缺失一些或所有轻链基因编码序列而使内源免疫球蛋白轻链基因功能性沉默的备选策略截然不同。
所述策略同样可很好地应用于免疫球蛋白重链基因座。因此,例如,lcr的完全或部分缺失将引起重链基因表达大大减少。优选在μc区的ch1结构域,但备选在所有免疫球蛋白同种型恒定区的ch1区中引入导致移码或mrna翻译过早终止的序列,将大大减少、消除或阻断与轻链形成免疫球蛋白四聚体。同样,如前所述,如果通过mrna翻译的过早终止体内阻断igm在前b细胞的细胞表面上的表达,则抑制进一步的发育,导致对抗原攻击的非生产性应答(kitamura等(1991)nature,350,423-426)。引入重链免疫球蛋白转基因拯救了b细胞膨胀,包含转基因编码的重链和内源鼠轻链的功能性免疫球蛋白四聚体在血浆中循环。
因此,内源轻链基因座和/或重链基因座产生相互作用并形成功能性免疫球蛋白四聚体的重链和轻链的能力在引入移码突变后可消除或大大降低,因为移码突变导致无关蛋白序列的合成,并一般伴有由存在异读框终止密码子引起的蛋白质合成过早终止。
这可通过在负责形成功能性免疫球蛋白四聚体的重链或轻链多肽序列中或上游插入外源dna或者少量缺失dna来实现。优选的方法是插入新的序列。所设计的在氨基酸编码序列中引起移码、导致编码mrna的翻译过早终止的有效插入事件,可限于单个核苷酸的引入。因此,编码区内插入一个或多个核苷酸(nucleocleotide)可导致移码。备选地,编码其它肽且包含终止密码子的序列的框内插入也将导致截短蛋白的合成(us5591669)。靶向插入事件也可包括引入选择标记基因和其它功能体,条件是保留所有内源序列,且所得融合蛋白破坏免疫球蛋白四聚体的形成,或者一个或多个框内终止密码子的存在引起mrna翻译的过早终止,导致不能形成功能性免疫球蛋白四聚体的截短蛋白的合成。
优选插入事件位于免疫球蛋白κ轻链恒定区。任选插入事件位于免疫球蛋白κ和/或λ轻链恒定区。实际上,插入可包含任何重组酶识别位点,例如lox位点。仅此就可导致移码。加入确保移码的其它核苷酸或者一个或多个终止密码子的密码子,也可提高插入序列在破坏重链和轻链通过重链ch1结构域与轻链κ和/或λ恒定区的相互作用二聚化方面的效果。插入可包含单个核苷酸,优选少于50个核苷酸。插入可以仅导致移码,但优选设计为使得一个或多个终止密码子引起mrna翻译过早终止。通过利用邻接插入位点的支臂的同源重组进行插入。优选在操作过程中,加入选择标记,然后接着切割,只留下识别位点和基因组中原位插入的任何其它序列,例如lox位点。
对于本领域技术人员显而易见的是,移码和截短蛋白的合成还可通过在编码序列中缺失一个或多个核苷酸以及加入包含终止密码子的最小添加序列来实现。优选插入或缺失事件发生在重链和轻链基因的恒定区,而不发生在内源基因座的多个v、d和j区。优选的选择是κ轻链恒定区。
因此,对于消除内源免疫球蛋白基因重排或mrna转录,不依赖大规模基因缺失的策略。相同的插入策略可用来通过重链同种型ch1区内的目标插入事件而抑制内源重链免疫球蛋白与轻链形成功能性四聚体复合物的能力。优选采用与kitamura等人(kitamura等(1991)nature,350,423-426)所述的类似策略,在前b细胞阶段阻断内源免疫球蛋白重链基因的表达,使得内源重链不在b细胞表面上表达,并且阻断由b细胞膨胀引起的生产性表达(productiveexpression),同时,轻链与内源igm的功能性ch1的缔合通过插入事件抑制,该插入事件导致翻译编码无法与免疫球蛋白重链进行功能性相互作用的轻链恒定区的κ和/或λ轻链mrna,从而防止功能性内源免疫球蛋白四聚体形成。
假若内源免疫球蛋白四聚体的功能性装配在功能上受损,则具有vh结构域的相关亲和力成熟的b细胞膨胀将受限于并依赖于外源免疫球蛋白重链和轻链转基因的存在和表达。免疫球蛋白转基因将以b细胞特异性方式参与所选择的非人哺乳动物宿主的等位基因排斥过程,导致对抗原攻击的生产性应答、b细胞膨胀和循环、转基因编码的抗原特异性免疫球蛋白四聚体。
本发明还提供非人哺乳动物,其中通过剔除部分或全部λ轻链lcr使内源λ轻链基因表达大大减少,通过剔除部分或全部κ轻链lcr使内源κ轻链基因表达大大减少。在本发明的一个实施方案中,通过剔除部分或全部相关lcr,仅使内源κ轻链表达大大减少,或者仅使λ轻链表达大大减少。
内源轻链基因座可保持功能性,因为它们可以重排,并被转录为功能性mrna,但是转录水平通过剔除一些或全部内源lcr功能性而大大降低(wo90/10077)。通过在小鼠es细胞中靶向核酸酶超敏位点的基因,或者在es细胞不存在时通过使用核移植(soulter(1998)nature,394,315-316)或来源于其它哺乳动物物种的ips细胞(参见gottweiss.和minger(2008)naturebiotechnology,26,271-272)进行克隆,除去lcr功能性。备选地,可通过靶向诱变,例如锌指核酸酶技术和dna修复(例如http://www.sigmaaldrich.com/life-science/functional-genomics-and-rnai/zinc-finger-nuclease-technology),实现重链或轻链的破坏。
通过剔除部分或全部κ轻链lcr,可使内源κ轻链基因表达大大减少,而在缺失或插入事件之后,可使λ轻链基因在功能上沉默。
通过剔除部分或全部λ轻链lcr,可使内源λ轻链基因表达大大减少,而在缺失或插入事件之后,可使κ轻链基因在功能上沉默。
在lcr剔除或插入事件之后,可使内源κ轻链基因在功能上沉默。
本发明还提供非人哺乳动物,其中通过剔除部分或全部κ轻链lcr和/或剔除部分或全部λ轻链lcr,使内源κ轻链基因表达和内源λ轻链基因表达的任一种或两种大大减少。通过剔除部分或全部λ轻链lcr,可使内源κ基因在功能上沉默且使内源λ基因活性大大降低。
通过剔除部分或全部κ轻链lcr,可使κ轻链基因表达大大减少,并且使λ基因表达在功能上沉默。
可通过剔除κ链lcr而大大减少或者通过缺失或插入事件而在功能上沉默仅内源κ轻链基因表达。
上述内源κ和/或λ轻链基因表达在功能上沉默或者内源κ和/或λ轻链基因表达大大减少的非人哺乳动物,其内源重链基因表达也可减少或在功能上沉默。按照一个实施方案,在本发明的非人哺乳动物中,内源重链基因表达在缺失一些或全部包含lcr的核酸酶超敏位点后减少,或者在缺失或插入事件之后在功能上沉默。优选在前b细胞阶段阻断内源重链基因的表达,使得内源重链不在b细胞表面上表达,并且采用与kitamura等人(kitamura等(1991)nature,350,423-426)所述类似的策略,阻断由b细胞膨胀引起的生产性表达。
上述非人哺乳动物另可包含一个或多个包含异源重链和轻链基因座及相关的b细胞特异性调节元件(优选包含关联lcr)的转基因。
在本发明的上下文中,术语“异源(的)”是指对于其所位于的哺乳动物而言不是内源的本文所述核苷酸序列或基因座。
因此,非人哺乳动物可包含含有异源κ轻链基因座和相关的b细胞特异性调节元件(优选包含lcr)的转基因和/或含有异源λ轻链基因座和相关的b细胞特异性调节元件(优选包含lcr)的转基因。
关联lcr的存在不是b细胞特异性表达所必需的。在基因座内包含关联lcr确保高水平转基因表达发生在每个整合位点,而不依赖于只有一些整合事件偶尔发生在b细胞中活跃转录的染色质区内的随机整合事件。关联lcr的使用显著减少筛选抗体表达所需要的转基因动物的数量,并允许插入多于一个的基因座,确保插入的所有基因座可以b细胞特异性方式按基本正常的水平表达。因此,lcr技术的应用,结合等位基因排斥机制可将内源免疫球蛋白基因与多个竞争转基因区分开来的出人意料的观察结果,开启了包含一个或多个免疫球蛋白重链或轻链基因座的转基因非人哺乳动物的装配途径,每个因座的v基因的复杂性比内源基因的低,而且与内源基因座(1-2mb)相比,包含相对易处理的dna断片(<300kb)以进行体外装配。例如,可将39个功能性人免疫球蛋白重链v基因区段克隆至两个或更多个免疫球蛋白重链基因座上。每一个可包含不同的v基因区段,但却具有共同的d和j基因区段及恒定(效应子)区。加入lcr确保了不论基因组内的整合位点,每一个都以相同的方式表达。因此,与存在于技术上难以操作的单个大基因片段上的单个更复杂基因相比,以这种方式加入两个或更多个小基因座提供相同的v基因复杂性。
每个异源轻链基因座可包含vl基因区段、j基因区段和轻链恒定区区段。优选vl和j基因区段及轻链恒定区区段来源于同一哺乳动物源,例如啮齿动物、猪、牛、山羊或绵羊。优选来源于人。
备选地,异源轻链基因座可以是包含哺乳动物源(优选人源)的可变结构域和来自不同哺乳动物的恒定(效应子)区的杂交基因座,该不同哺乳动物例如但不限于小鼠、大鼠、仓鼠、兔、猪、山羊和牛。如果宿主哺乳动物是小鼠,则优选恒定区来源于啮齿动物,更优选为小鼠或大鼠。这类异源轻链基因座包含优选仅来自一个物种的vl和j区段和来自另一个物种的轻链恒定区。
如果预期杂交κ轻链转基因,则vl和j基因区段优选来自提供重链v、d和j基因区段的相同物种,且优选来源于人。κ轻链恒定区和重链恒定区也同样优选来源于同一物种,但是该物种不同于提供可变结构域的物种,并且优选来源于啮齿动物,优选来源于大鼠或小鼠。
所预期的所有轻链转基因的特征是,在抗原攻击后,轻链以b细胞特异性方式重排,在转录和翻译后,所得轻链能够与在同一b细胞中产生的转基因来源的重链免疫球蛋白复合。免疫球蛋白四聚体的生产性表达引起b细胞膨胀和转基因编码的抗原特异性四价免疫球蛋白复合体在缺乏显著水平的内源免疫球蛋白四聚体时在血清中蓄积。
如果内源λ轻链表达在功能上未受抑制,则可检测到低水平的包含内源λ轻链的宿主抗体或杂交抗体。这些可在筛选杂交瘤上清液后弃去。
在人中,有36个功能性κvl基因区段、5个jl基因区段和一个κ轻链恒定区(http://imgt.cines.fr)。优选存在于本发明非人哺乳动物的转基因上的异源κ轻链基因座包含一个或多个人vl基因区段、所有人jl基因区段和一个人κ轻链恒定区。任选人κ轻链恒定区可置换为优选大鼠或小鼠源的备选哺乳动物κ轻链恒定区。
存在于本发明非人哺乳动物的转基因上的异源λ轻链基因座优选包含鼠λlcr及人λ轻链v1和v2基因区段、人λj1、j2、j3和j4基因区段及人λ轻链c1、c2、c3和c4恒定区区段(wo90/10077和wo2003/000737)。任选人λ轻链c1、c2、c3和c4恒定区区段可置换为优选大鼠或小鼠源的备选λ轻链恒定区。
存在于本发明非人哺乳动物的转基因上的异源重链基因座优选包含重链免疫球蛋白lcr(优选为鼠源)、一个或多个人v基因区段,一个或多个j基因区段和一个或多个d基因区段。优选存在10个或更多个不同的人v基因区段及所有的人j和d基因区段。
基因座还可包含一个或多个人恒定(效应子)区,优选μ和γ恒定区。任选人恒定效应子区可置换为来自其它非人哺乳动物的效应子区。如果非人哺乳动物宿主是小鼠或大鼠,则优选恒定(效应子)区来源于大鼠或小鼠。与人的相比,小鼠和大鼠b细胞受体复合体(bcr)的跨膜结构域是100%保守的。因此,在抗原攻击后,包含大鼠恒定(效应子)区基因的抗体基因座的小鼠转基因应发挥与包含小鼠恒定(效应子)区基因的基因座一样良好的作用,并且可优于包含人恒定(效应子)区基因的基因座(defranco等(1995)ann.nyacad.sci.,766,195-201)。转基因可包含优选小鼠或人源的重链、κ和λ轻链lcr。在所有哺乳动物物种中起作用的lcr是已知的,并且可替代转基因的人或小鼠lcr(li等(1999)trendsgenet.,10,403-8)。
如果预期产生完整人抗体,则克隆的来源于由杂交瘤表达的杂交抗体的人抗原特异性vh和vl结合结构域可与人恒定重链和轻链恒定区融合,因此得到任何种类的完整的人四聚体抗体。
作为进一步的改进,每个免疫球蛋白κ和/或λ轻链基因座还可包含显性选择标记基因。
掺入基因座的显性选择标记基因可具有相同或不同的作用机制。对于本发明的目的,可使用任何显性选择标记基因,条件是在选择性或毒性攻击存在下,该基因的表达为杂交瘤或来源于非人转基因哺乳动物的转化b细胞提供选择性益处。通常,显性选择标记基因是原核来源的,并选自:赋予有毒药物抗性的基因,例如嘌呤霉素(vara等(1986)nar,14,4617-4624)、潮霉素(santerre等(1984)gene,30,147-156)和g418(colbere-garapin等(1981)150,1-14),或者包括这样的基因:其消除对某些营养的需要,使得其表达将有毒物质转化成必需氨基酸,例如将吲哚转化为色氨酸或将组氨醇转化为组氨酸(hartmann和mulligan,(1988)pnas,85,8047-8051)。
本发明这个方面的必要条件是,将显性选择标记掺入免疫球蛋白轻链转基因座内,并且与所需的免疫球蛋白轻链等位基因共表达,以确保b细胞特异性表达。备选地,可采用同源重组结合es细胞或核移植方法,将药物抗性基因插入内源或外源(转基因)免疫球蛋白基因座(teriele等(1992),pnas,89,11,5128-5132)。
非人哺乳动物还可包含含有异源重链基因座和相关b细胞特异性lcr及调节元件的转基因。可存在多于一个的不同转基因重链基因座,各包含lcr和调节元件。
各包含一个或多个v基因区段、一个或多个d基因区段、一个或多个j基因区段和一个或多个恒定(效应子)区的重链基因座作为转基因引入,各基因座包含关联lcr。
各基因座包含驱动b细胞特异性表达所必需的5’和3’调节元件。在响应抗原攻击时,每个重链或轻链基因座以与内源基因座基本相同的方式表达,导致转基因编码的抗原特异性亲和力成熟的四聚体免疫球蛋白在小鼠血清中循环。
优选每个重链基因座包含一个或多个v基因区段,例如1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、45、50、60或更多个v基因区段,其可来源于任何脊椎动物物种,优选非人哺乳动物。优选不超过20个v基因区段存在于任何单个重链基因座上。
在一个实施方案中,每个基因座可只包含一个v基因区段。在该实施方案的一个备选实施方案中,存在多个v基因区段,每个v基因区段不同于所有其它v基因区段。在这个实施方案中,任一个基因座上的v基因区段可全部来源于同一物种的生物,例如所有的v基因区段可以来源于人。备选地,任一个基因座上的v基因区段可来源于不同物种的生物,例如一些v基因区段来自人,其它则来自绵羊、牛、兔、骆驼(camelid)或甚至鲨鱼。在第二个备选实施方案中,每个v基因区段与所有其它v基因区段相同。不论存在的v基因区段数目和性质,每个基因座上其余的d和j基因区段可与所有其它基因座上的区段相同或不同。
因此设想非人哺乳动物可含有多个拷贝的重链基因座。这具有使在b细胞中发生生产性重排的可能性最优化的优势,因此允许用于抗原识别的免疫球蛋白重链的最适产生。
在另一个实施方案中,每个基因座包含多个v基因区段。
优选v基因区段来源于人。
术语“v基因区段”涵盖任何来源于脊椎动物的天然存在的v基因区段,包括但不限于鲨鱼、啮齿动物、骆驼和人。v基因区段必须能够与d基因区段、j基因区段和编码重链恒定(效应子)区的基因区段重组,以当重排核酸在b细胞中表达时产生能够与κ或λ免疫球蛋白轻链复合的免疫球蛋白重链抗体。
v基因区段在其范围内还包括编码天然或工程改造的同源物、衍生物或蛋白质片段的任何基因序列,其能够与d基因区段、j基因区段和编码重链恒定区的基因区段重组,以当重排核酸在b细胞中表达时产生能够与κ或λ免疫球蛋白轻链复合的免疫球蛋白重链抗体。v基因区段可来源于例如t细胞受体基因座。
优选本发明的多个重链基因座包含39种功能性人v基因区段及其改造变体的任何数目或组合。它们可在任何数目的基因座上,例如4个包含8个v基因区段的基因座加1个包含7个v基因区段的基因座;7个包含4个v基因区段的基因座加1个包含3个v基因区段的基因座;或39个各包含1个v基因区段的基因座。
将人v基因分成7个家族vh1-vh7,并将各个基因归类到各自编号的家族内。每个基因使用的频率取决于特定免疫应答的不同需要。例如,与vh5家族的基因相比,在响应细菌抗原时,可优先使用vh3家族的基因。因此,在本发明的另一个优选实施方案中,将已证实可用于产生针对特异性抗原的抗体应答的v基因区段群归类为单独的基因座,各自包含不同的显性选择标记基因。v基因区段可按照家族归类,或者可按照各自的功能归类。例如,如果vh3家族的v基因显示可用于产生针对细菌抗原的免疫应答,则它们可用于产生特别可用于产生针对细菌抗原的仅有重链的抗体的基因座。备选地,如果表明来自vh3和vh5家族的若干个别基因可用于产生针对细菌抗原的免疫应答,则它们可归类在一起并用于产生特别可用于产生针对细菌抗原的抗体的基因座。
在本发明上下文中,“免疫球蛋白重链基因座”涉及最小的微基因座,其编码包含一个或多个v基因区段、一个或多个d基因区段和一个或多个j基因区段并与一个或多个编码重链恒定(效应子)区的基因区段有效连接的vh结构域。优选抗体库变化性的主要来源是通过选择v、d和j基因区段以及通过v-d和d-j连接形成的cdr3区。
本发明的优势是,在相同的转基因非人哺乳动物中通过利用等位基因排斥来使用多个免疫球蛋白重链基因座,可使在重排v、d和j基因区段中得到的抗体库和多样性最大化。随机选择基因座之一开始重组,如果第一重组是非生产性的,则接着下一个基因座等等,直到从基因座之一产生生产性重组,这种等位基因排斥的过程可确保实际上所有存在于合并基因座上的v基因区段都可成为整个重组过程的部分。
为了提高任何给定免疫球蛋白重链基因座上所有vh基因区段生产性参与vdj重排的概率,可将ctcf位点分散在vh基因区段群之间。
根据在b细胞中进行的距离测量以及在vh区方向上和遍及vh区的测量,免疫球蛋白基因座在其正常构象中似乎具有三维折叠结构(jhunjhunwala等(2008)cell,133,265-279)。这类折叠或成环的结构解释了不同的vh区即使以距免疫球蛋白重链基因座的d、j和恒定区完全不同的距离排列时可同等有效地使用的原因。
最近也越来越清晰的是,在许多基因座上通过成环形成的折叠结构经由称为ctcf的特定染色质结合蛋白介导。如诱变实验证实的一样,ctcf似乎直接参与染色质成环的形成(splinter等(2006)genesdev.,20,2349-2354)。更近期的研究表明,黏结蛋白这种将姐妹染色单体保持在一起的蛋白质复合体存在于ctcf结合位点上(wendt等(2008)nature,451,796-801)。免疫球蛋白vh区的许多ctcf位点的加入(kim等(2007)cell,128,1231-1245;denger,wong,jankevicius和feeney(2009)j.immunol.,182,44-48)提高了转基因免疫球蛋白重链基因座的vh区可正确折叠并允许有效使用存在于该基因座上的所有不同的v基因区段的概率。
包含异源重链基因座的每个转基因还可包含显性选择标记。优选显性选择标记不同于引入κ或λ轻链基因座内的显性选择标记。
对于本发明的目的,可使用任何显性选择标记基因,条件是在选择性或毒性攻击存在下,基因的表达为杂交瘤或来源于非人转基因哺乳动物的转化b细胞提供选择性益处。通常,显性选择标记基因可以是原核来源的,并选自:赋予有毒药物抗性的基因,例如嘌呤霉素(vara等(1986)nar,14,4617-4624)、潮霉素(santerre等(1984)gene,30,147-156)和g418(colbere-garapin等(1981)150,1-14),或者包括这样的基因:其消除对某些营养的需要,使得其表达将有毒物质转化成必需氨基酸,例如吲哚转化为色氨酸或组氨醇转化为组氨酸(参见hartmann和mulligan(1988)pnas,85,8047-8051)。
本发明的必要条件是,使显性选择标记(如使用的话)位于免疫球蛋白重链转基因座内,以确保b细胞特异性共表达。备选地,可采用同源重组结合es细胞或核移植方法,将药物抗性基因插入内源或外源(转基因)免疫球蛋白基因座(例如teriele,robanusmaandag和berns(1992),pnas,89,11,5128-5132)。
可将相同的显性选择标记基因掺入所有重链基因座内。备选地,不同的重链基因座或重链基因座群可包含不同的显性选择标记基因。
由于转基因轻链基因座内的功能性显性选择标记基因共表达,因此可以选择不含表达内源免疫球蛋白的细胞的来源于本发明表达四聚体抗体的转基因小鼠的杂交瘤或转化b细胞,优选转化的长寿浆细胞(slifka等(1998)immunity,8,363-372)。此外,与掺入轻链基因座中的显性选择标记相比,由于重链基因座内不同显性选择标记的存在和共表达,因此也可选择表达来源于转基因重链基因座上存在的特定v区段群的抗体的杂交瘤或转化b细胞系。例如,在κ轻链转基因座内加入嘌呤霉素抗性基因使得能够选择所有表达κ轻链转基因的细胞。备选地,在包含优选的v基因区段的重链转基因座中加入g418抗性基因使得能够选择所有表达优选的v基因区段的细胞。
具体地讲,本发明提供产生抗原特异性异源单克隆抗体的方法,所述方法包括:
(a)用抗原对上述非人转基因哺乳动物进行免疫;
(b)从已免疫的转基因哺乳动物的b细胞制备分别产生单克隆抗体杂交瘤或无限增殖化b细胞系;
(c)通过使用存在于包含异源免疫球蛋白轻链和重链基因座的转基因上的显性选择标记基因,选出至少一种表达异源抗体的杂交瘤或无限增殖化b细胞系;和
(d)选择至少一种产生与抗原特异性结合的抗体的杂交瘤或无限增殖化b细胞系。
下面参照下列附图的详细描述,仅通过实施例对本发明进行描述。
附图:
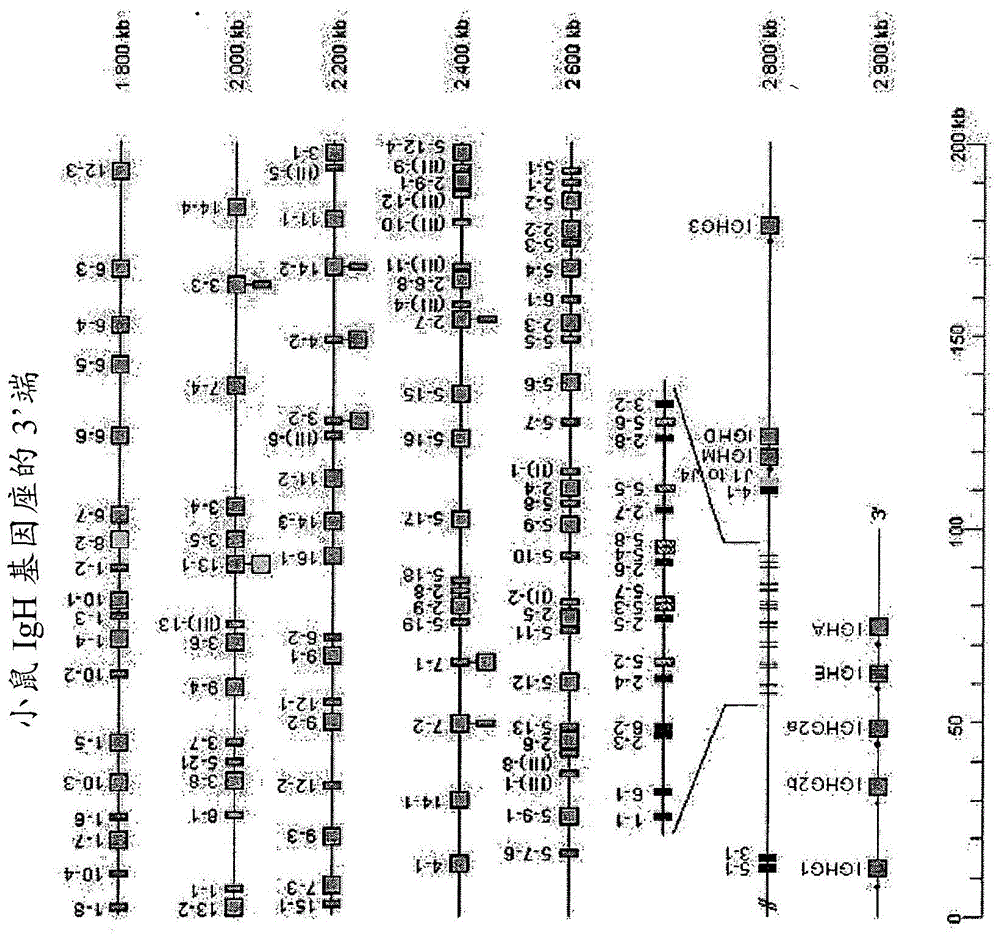
图1a:小鼠igh基因座的3’端。
该图谱复印自imgt数据库(http://imgt.cines.f)。标尺以千碱基(kb)为单位。绿色正方形为功能性vh区段;红色和黄色正方形为非功能性vh区段;橙色正方形为jh区段;蓝色正方形为恒定区。未标出基因座3’端上的内含igh增强子和lcr。
图1b:使igh失效的策略
首行显示具有包括编码膜形式的igm的2个外显子在内的不同外显子的小鼠cμ区。往左为基因座的j、d和vh区,往右为从cδ开始的其它恒定区。尾行显示重组后正常m1外显子的部分氨基酸序列。dna序列表示整合序列。终止密码子用红色表示,spei位点用红色和蓝色表示。
图1c:es细胞中使cμ失效的重组
该图显示通过覆盖重组插入片段3’侧的pcr分析而得到的es细胞的两个重组阳性克隆。较大的片段相当于在图1b标出的位置上插入m1外显子中的neo选择标记。
图1d:cμ敲除小鼠的facs分析
在m1外显子被终止密码子和neo基因中断后,将es细胞引入胚泡得到嵌合体。将这些嵌合体培育成纯合性,并分析血液中b细胞的存在情况。上两图表示使用b细胞标记b220和cd19对正常野生型小鼠和杂合中断的m1外显子小鼠进行的facs分析。下两图表示两个纯合小鼠,其未显示b220+、cd19+细胞,即无功能性b细胞。
图1e:培育成重组小鼠后neo的缺失
使图1d的小鼠与表达重组酶的小鼠杂交以缺失neo基因。右边两条泳道显示高于亲本动物的neo基因的长片段pcr产物,左侧紧邻的两条泳道表示携带neo缺失和野生型等位基因的两个杂合小鼠。左侧相邻的4条泳道为野生型小鼠,而最左一条泳道表示携带纯合缺失的neo基因和失活的m1外显子的小鼠。
图2:小鼠igκ基因座的图谱
该图谱复印自imgt数据库(http://imgt.cines.f)。标尺以千碱基(kb)为单位。绿色正方形为vκ区段;橙色正方形为jκ区段;蓝色正方形为恒定区;黑色圆形为κ-增强子和红色圆形为κ-lcr序列。
图3:小鼠vκ敲减
通过lcr缺失使小鼠igκ基因座在功能上失活的方案
通过同源重组将lox新霉素抗性基因盒插入到es细胞中置换3’κ-lcr(尾行)。用cre重组酶(+cre)处理可除去lox位点之间的所有序列,在κ基因座上留下单个lox位点(首行)。
图4:插入小鼠cκ使κ基因座失活。
基因座(首行)与图3的相同。尾行表示cκ外显子5’端上的序列(首行的蓝色),在碱基上方标出氨基酸编码。开始的gg碱基对紧邻剪接后的编码氨基酸r的剪接受体位点。中间行表示插入34个碱基对lox位点插入片段(蓝色和红色反向重复序列),这使恒定区的密码子使用是异读框的,并产生下游终止密码子(例如粗体加下划线的tga)。黑色圆形为κ-增强子和红色圆形为κ-lcr序列。
图5:小鼠cκ插入
基因座(首行)与图3的相同。尾行表示cκ外显子的5’端序列(第一行中的蓝色),碱基上标出氨基酸编码。开始的gg碱基对紧邻剪接后的编码氨基酸r的剪接受体位点。中间行表示插入的含有lox序列(蓝色和红色反向重复序列)和4个终止密码子的46个碱基对插入片段,这也使恒定区的密码子使用是异读框的,并产生下游终止密码子(例如粗体加下划线的tga)。黑色圆形为κ-增强子和红色圆形为κ-lcr序列。
图6a:小鼠cκ恒定区终止密码子和移码插入
基因座(首行)与图3的相同。尾行表示cκ外显子部分序列(黑色),碱基上标出氨基酸编码。中间行表示cκ编码区部分序列。其上一行表示插入含有lox序列(蓝色和红色反向重复序列)、3个终止密码子的44个碱基对的插入片段,这也使恒定区的密码子使用是异读框的,并产生下游终止密码子(taa)。黑色圆形为κ-增强子,红色圆形为κ-lcr序列,用于插入的hpai位点用红色表示。
图6b:es细胞中使cκ失效的重组
凝胶显示了图13a所示es细胞中对cκ重组的多个克隆插入位点进行pcr扩增的结果。克隆351和623为阳性,将其注射入胚泡中以产生igκ阴性小鼠。
图7:人igκ基因座。
该图谱复印自imgt数据库(http://imgt.cines.f)。标尺以千碱基(kb)为单位。绿色正方形为功能性vk区段;红色和黄色正方形为非功能性vκ区段;橙色正方形为jκ区段;蓝色正方形为恒定区;黑色圆形为κ-增强子,红色圆形为κ-lcr序列。
图8:用于转基因作用的杂交人/大鼠igκ基因座的产生
基因座的3’端获自含有小鼠κ3’增强子(黄色)的小鼠(黄色)。小鼠恒定编码序列用大鼠的恒定编码序列置换,包括其通过长片段pcr获自大鼠基因组dna(红色)的5’增强子。从vκ4-1下游直到人jκ序列的人区段获自小鼠(黄色)以保持v区与j区的适当间隔。人jκ区段获自覆盖人基因座该部分的pac(绿色)。绿色正方形为分别添加或作为区组(block)添加的vκ区段(参见正文)。存在于pac载体上的嘌呤霉素抗性基因用红色表示,黑色圆形为κ-增强子,红色圆形为κ-lcr序列。
图9:vh4重链基因座的图谱
在5’端含有新霉素选择标记的该基因座用作构建人/小鼠杂交基因座的起始原料。按照wo2008/035216中所述构建该基因座。标尺以千碱基为单位。基因座含有4个vh区(1-46、3-53、3-23、3-11)、全部的人d区段、全部的人j区段和igg恒定区以及3’人ighlcr。
图10:4个vh人/大鼠恒定区igh基因座的产生
存在于vh4人基因座的ceui位点(图6)用于通过添加经pcr由大鼠基因组dna扩增的大鼠恒定编码区和转换区来产生人/大鼠基因座。同样,小鼠lcr区作为几个片段由小鼠基因组dna扩增,这些片段先克隆在一起产生完整的小鼠ighlcr。人vh4基因座的5’端含有4个vh区段、全部的人d区段和全部人j区段(图6)。所有人序列用蓝色表示,所有小鼠序列用淡绿色表示,大鼠序列用红色表示,新霉素抗性基因用紫色表示。
图11:包含λlcr的igλ增强子的缺失
使用潮霉素抗性基因和杀稻瘟菌素s(blasticins)抗性基因,采用标准置换型载体通过同源重组剔除λ基因座增强子,所述潮霉素抗性基因两侧是与邻接增强子2-4的区同源的序列,所述杀稻瘟菌素s抗性基因两侧是与邻接4-10增强子的区同源的区段。置换产生丢失增强子且表达减少的λ基因座。潮霉素抗性基因用红色表示,杀稻瘟菌素s抗性基因用橙色表示。小鼠vλ区段用绿色表示,j区段用黄色表示,恒定区用蓝色表示。图谱复印自igmt数据库。
图12:用于转基因作用的人大鼠vκ基因座
基因座与图8的相同,但已将其它人vκ区段添加入该基因座。所得基因座含有所有频繁和中等频繁使用的人vκ区段。绿色圆形和绿色线为鼠κ-增强子和内含子;红色正方形和圆形为大鼠κ-恒定区和增强子;蓝色正方形为人vh区段;深蓝色为嘌呤霉素选择标记。
图13a:人大鼠杂交基因座的产生
通过将vh区连接在一起形成在scei位点之间克隆的17个连续人vh区的多联体,来产生该基因座。将小鼠间隔区添加入含有所有人dh和jh区段的人40kb片段中以在dh与vh区段之间保持适当距离。接着添加含有scei位点的vh6-1区段。然后将多联体添加至vh6-1上。最后,将各种大鼠恒定区和小鼠lcr添加至3’侧。所得基因座含有全部频繁和中等频繁使用的人vh区段。
图13b:人/大鼠igh基因座的起始构建体
凝胶表示在添加vh6-1后人大鼠igh基因座构建第一步的结果。右边的泳道表示noti消化的两个克隆质粒。泳道(notixmlui)表示泳道1的质粒在载体中具有正确朝向,而泳道2的质粒具有错误朝向。泳道1的质粒用于产生基因座的下一步骤中。
图13c:vh区段的多联体
凝胶表示xhoi/sali消化的17个不同vh区的多联体。标记泳道含有用bsteii消化的λ噬菌体dna。
图14:转基因人大鼠重链免疫球蛋白基因座
引入同一动物中的两个重链基因座的实例。彼此的对立选择通过等位基因排斥进行。可将其它vh区段添加至各基因座。备选地,可采用更多的重链基因座引入其它vh区段。显然,可采用相同策略增加κ或λ轻链基因座的多样性。
图15:人/大鼠λ转基因轻链基因座
图显示人/大鼠基因座的实例。其3’端获自含有小鼠λ3’lcr(黄色)的小鼠(黄色)。小鼠恒定编码序列用通过长片段pcr从大鼠基因组dna获得的大鼠恒定编码序列(红色)替换。vλ2-14下游直到人jκ序列的人区段获自小鼠(黄色),以保持v区和j区之间的适当间隔。通过覆盖人基因座该部分(蓝色)的人pac的长片段pcr获得人jλ区段。蓝色正方形为分别添加或作为区组添加的vλ区段。存在于pac载体上的潮霉素抗性基因用紫色表示。
实施例
在下面的实施例中,产生转基因小鼠,其表达作为转基因通过显微注射引入受精卵(常规转基因过程)的杂交人/大鼠重链和轻链基因座。对供卵小鼠进行修饰以不表达或非常低地表达内源小鼠重链基因和小鼠轻链基因。小鼠中有κ链和λ链两种轻链基因座,其中λ仅用于约2%的小鼠h2l2抗体。因此,实施例在具有igh基因座且仅内源κ基因座失活的小鼠中,或者在具有igh和失活的κ基因座且剔除λ基因座的调节序列以更进一步地降低λ基因座的表达的小鼠中。
用于构建重链和轻链基因座,在抗原攻击后形成和筛选转基因小鼠的方法基本上如前所述(janssens等(2006)pnas,10,103(41),15130-5;wo2006/008548、wo2007/096779、gb0805281.3和2008年6月11日申请的要求gb0805281.3的优先权的pct申请),在所有重链基因座上保留ch1结构域的除外。用于获得表达功能性异源免疫球蛋白基因座和/或具有工程改造的内源基因座的脊椎动物(包括小鼠以外的哺乳动物)的通用方法参见wo2006/047367。在下面的实施例中,在es细胞中使用重组,并使用修饰的es细胞产生具有所需性质的小鼠。然而,相同方法可在诱导性多能干细胞(ips细胞)中进行,其然后用来产生小鼠(例如boland,hazen,nazor,rodriguez,gifford,martin,kupriyanov和baldwin(2009),461,7260,91-4及其中的参考文献)。备选地,在体细胞或体干细胞进行修饰,随后改编(reprogram)到ips细胞中产生修饰小鼠。同样,可将修饰的造血干细胞移植到缺乏b细胞的受体小鼠中以产生人抗体或人杂交抗体。
实施例1
在该实施例中,通过类似于kitamura和rajewsky发表的策略,使igh基因座(图1a)失活,其差别在于将终止密码子在kitamura等人(kitamura等(1991)nature,350,423-426)所述位置前一个氨基酸的位置上引入cμ区。es(或ips)细胞用构建体转染,该构建体将小鼠igm基因第一膜外显子的第二密码子变为终止密码子。这涉及包括用于转染的neo选择的常规步骤。将spei位点加入重组序列中,以能够监测成功重组(图1b)。随后通过dna印迹法筛选es细胞以证实成功的重组克隆。这产生10个正确的重组体(例如图1c)。这些当中,将3个注射到小鼠胚泡中获得嵌合体,随后培育嵌合体得到对于igh突变为纯合的小鼠。这类小鼠b细胞的facs分析(b220对cd19)表明,外周血中不存在b细胞(图1d)。随后将小鼠与表达重组酶的小鼠杂交除去neo基因(图1e)。同样,在es细胞中通过重组使小鼠igκ基因座(图2)失活或减少(图2、3、4-6)。所得κ失活小鼠与重链ko小鼠杂交。通过将3’κ基因lcr替换为两侧是lox位点的neo抗性标记,实现活性敲减(图3)。任选用重组酶处理除去neo基因。
备选地,可在ighko细胞中直接敲除κ等位基因。可采用若干不同的策略达到κ失活。可通过使用es细胞或ips细胞中的同源重组将两侧是lox位点的neo基因插入cκ外显子的5’端(图4),或者通过插入两侧是lox位点的neo基因和编码终止密码子的其它序列(图5和图6a),来实现κ基因活性的阻遏。重组的es细胞用cre处理可使该序列为异读框的(图4),或另含有新的终止密码子(图5-6a)。图6b表示图6a所示构建体转染后在es细胞中的重组结果,产生具有失活的一个cκ等位基因的2个es细胞克隆(共获得5个这样的克隆)。用肌动蛋白驱动的重组酶质粒通过标准瞬时转染用重组酶处理细胞,以剔除两个lox位点间的区。随后通过常规方法使用该细胞产生小鼠,培育子代得到纯合小鼠。这类小鼠包含b细胞,其中包含κ轻链的免疫球蛋白四聚体的装配基本被破坏或完全阻断。
接下来,按照之前对于人vh区段所述,将最常用的人igκ基因座的vκ基因(利用ig数据库评价;http://imgt.cines.fr/;参见图7、8)通过标准pcr扩增,并亚克隆至xhoi/sali位点之间。这使得vκ区能够多聚化,并将多聚体保持在xhoi和sali位点之间。
同样,将包括3’κ增强子的小鼠κ基因座的3’端与大鼠恒定(cκ)区和大鼠5’增强子克隆在一起(图8)。接着,克隆入人jκ区与得自小鼠vκ和jκ之间的区域(17kb)(图2)以保持vκ区和jκ区之间的正常间隔。最后,通过常规方法(例如janssens等(2006),同上),将人vκ插入含有嘌呤霉素选择标记的pac(图8)。
在所示实施例中,将最常用的vκ区段(4-1、3-11、3-15、3-20和1-39)多聚化,并连接到含有人j区和小鼠增强子及大鼠cκ区的pac载体中。这产生由puro抗性标记基因、人vκ区段和大鼠恒定(cκ)区组成的人-大鼠杂交基因座(图8)。
同样,构建杂交人/大鼠igh基因座。此外,就起始原料而言有许多可能性。在该实施例中,起始原料为含有4个人vh区、全部的人dh和jh区段、2个人恒定区及人lcr的人pac(图9和英国专利申请号0905023.8)。
后面的pac在j区和恒定区之间具有独特的ceui大范围核酸酶位点。为使得能够容易地构建杂交基因座,使用该ceui位点除去人3’端序列,并将这些替换为大鼠恒定区和转换区(cμ、cγ3、cγ1和cγ2)。这些通过标准长片段pcr扩增自大鼠基因组dna。最后,添加小鼠重链lcr。该调节序列以3个部分由小鼠基因组dna扩增,并亚克隆到一起以恢复完整的lcr,并加至大鼠恒定区3’侧(图10)。因此,所得杂交igh基因座含有neo选择标记、人v、d和j区及大鼠恒定区与小鼠调节序列。
随后杂交基因座插入片段作为大的dna片段从pac中分离,并注射入来源于igh/igκ杂合或纯合失效小鼠的小鼠受精卵,产生对于人/大鼠杂交igh和igκ基因座是转基因的小鼠。所有这些都通过常规方法进行(例如janssens等(2006),同上)。
随后培育杂交igh和杂交igκ转基因小鼠,得到对于内源小鼠igh和igκ表达而言是纯合失效且对于人/大鼠杂交igh和igκ表达而言为阳性的小鼠。随后通过常规方法,对这些小鼠进行免疫产生抗原特异性杂交人/大鼠h2l2抗体。如果通过杂交瘤产生单克隆的人/大鼠ig(抗体),则嘌呤霉素和新霉素的双重选择确保只选出既含igh又含igκ基因座的骨髓瘤融合体。
本领域技术人员应理解的是,可对该方法作各种变化以产生杂交转基因小鼠,例如使用不同的载体、不同的选择标记、不同的重组位置以使小鼠基因失活,或可在杂交基因座的实际(常规)克隆策略中作出各种变化。可采用同一方法,使用来源于任何单一哺乳动物物种的免疫球蛋白dna产生任何正常基因座或杂交基因座,或者使用来自两个或更多个物种的dna得到杂交基因座。
实施例2
本实施例基本上与实施例1相同,除了通过降低内源小鼠igλ基因座的表达频率更进一步提高获得igh/igκh2l2抗体的高频率。这可通过用选择标记置换两个igλ的调节区来实现(图11),在该情况下为潮霉素抗性基因和tk-bsd基因。
后者使得能够阳性选择杀稻瘟菌素s抗性(karreman,(1998)nar,26,(10),2508-2510)。标记的这种组合当置换调节区时,使得能够在2种es细胞重组中进行阳性选择。重组在实施例1产生的es细胞中进行,或者备选地,在正常es细胞中平行进行,并且培育成上面实施例1中所述小鼠。
所得转基因小鼠含有杂交人大鼠igh和igκ基因座,对于内源小鼠igh和igκ而言为阴性(或以非常低的水平表达cκ),并以非常低的水平表达igλ。在通过常规方法免疫和产生杂交瘤后,通过neo(图10)和puro(图8)选择,选出只表达人大鼠杂交h2l2抗体的杂交瘤用于表达转基因杂交基因座。
实施例3
实施例3类似于上述实施例,但是杂交igκ基因座通过添加较不常使用的vκ区段而延伸(图12;vκ1-9、1-33、2-30、2-28、1-27、1-5)。备选地,可另外添加突变/修饰的vκ区段或vλ区段。可通过采用上述相同的xhoi/sali克隆策略进行其它区段的添加。对在该实施例中产生的小鼠进行免疫,允许在响应用抗原免疫时更大的复杂度。可通过添加其它vκ区段或使用所有上述vl区段的组合,使vl区的数目进一步改变。
实施例4
实施例4类似于上述实施例中所描述的方法,但在这里通过使用18个vh区段和5个大鼠恒定区产生杂交人/大鼠igh基因座(图13a;人vh6-1、1-2、4-4、2-5、3-07、1-8、4-39、3-15、1-18、3-23、3-30、3-33、3-48、4-34、3-49、3-53、4-59、1-69)。首先人基因座的中心70kbdj区在5’端用来自小鼠igh内含子的8kb延伸,以保持vh区段与d区之间的适当距离。接下来,将10kb具有人工scei大范围核酸酶位点的第一vh区(6-1)克隆在小鼠内含子序列的5’端(图13b)。在单独的质粒中,所有剩余的vh区通过加插如上所述的xhoi/salivh区段而克隆在一起(图13c)。还可以将更多vh区段或已经修饰/突变的vh区段加入这些基因座中。还可包括ctcf位点。在所示实施例中,使用了3个这类位点。它们通过包括上游ctcf位点的vh区的长片段pcr获得。此外,与图10中的基因座相比,已加入了大鼠cα。在该实施例中,免疫允许响应用抗原进行免疫时更大的复杂度,特别与实施例3组合时。将vh多聚体克隆到vh6-1dj质粒中,之后加入大鼠恒定区完成基因座。
在所有这些实施例中,通过加入v区段作为存在于同一小鼠中的其它重链或轻链转基因座的一部分,将进一步提高应答的复杂度。由于所有基因座经历等位基因排斥(参见wo2007/096779),因此在抗原攻击后,体内仅选出优选的重排,导致b细胞膨胀及抗体在血清积累。图14表示可引入同一动物的2个重链基因座的实例,每一个可通过等位基因排斥而被利用。显然,可加入更多的vh区段(包括修饰的vh区段),或者可引入甚至更多的基因座以增加转基因免疫库的复杂度。还可利用对其它物种有特异性的v区段将该方法应用于这些物种。
实施例5
在该实施例中,通过培育将人/大鼠igλ基因座加入携带上述实施例中所述的人/大鼠igh和/或igκ基因座的小鼠更进一步地增加人/大鼠杂交抗体的多样性。人/大鼠杂交λ基因座的产生与前述实施例中对于人/大鼠igκ基因座所述非常相似。区别是由jλ和cλ区成对出现所引起的,因此将2个大鼠cλ区克隆至2个人jλ区(图15)。通过克隆出现在该位置的正常小鼠序列,来保持vλ和jλ区段之间的间隔(参见图11)。在该实施例中,2个人jλ和大鼠cλ区段与4个人vλ区段一起使用。综合起来,这些包括超过80%的人igλ应答。调节序列(lcr,图15)来源于小鼠以确保最佳表达,选择标记被加至基因座的5’端。如上所述,将基因座作为限制片段分离,并注射到受精卵中以产生携带转基因λ基因座的小鼠。
在所有这些实施例中,可使用来自不同物种的vh、vl、d、j和恒定区来产生其它单一物种抗体或杂交物种抗体。对本领域技术人员还显而易见的是,一旦鉴定出抗原特异性抗体,则可通过完全常规的方法,将vhdj和vlj区克隆至来自同一物种或来自不同物种的备选恒定区。
- 还没有人留言评论。精彩留言会获得点赞!