一种通过降低叶绿素A二级激发态提高光合效率的方法与流程
一种通过降低叶绿素a二级激发态提高光合效率的方法
技术领域
1.本技术涉及中药种植领域,尤其是适用于通过光调节叶绿素激发提高中药种植效率的领域。
背景技术:
2.光照射到植物叶片上,其能量将由叶绿素吸收,吸收的能量根据光量子的能级(波长越长,能级越低)分别引起叶绿素分子轨道的电子跃迁到一级激发态还是二级激发态(都是单线态)(图1)。植物进行光合作用的能量来自于太阳能,光合色素吸收太阳能,产生激发能,再经过类囊体膜的电子传递链上进行一系列氧化还原反应,最终进行能量转化,产生生物能(atp)和还原势(nadph2),提供叶绿体固定二氧化碳,合成碳水化合物的所需的能量和化学势。
3.虽然太阳能提供了光合作用过程所需的基本能量,但是,过强太阳能会使叶绿体产生过剩的激发能,导致活性氧等有害物质的产生,最终引起细胞死亡。为了应对多变的环境因子,植物产生了一系列的保护机制,包括非光化学淬灭和光破坏机制,就是说,叶绿体中过剩的激发能并不能转变为化学能,而是以热的形式(非光化学淬灭),或者部分的光合器官解体的形式消耗掉(光破坏),这部分能量消耗无疑降低了植物光合效率。因此,如何有效地设计光能输入是关系到植物光能利用效率的关键因素。植物的叶绿体中与吸收太阳能有关的光合色素有两大类分子,即:叶绿素(chla和 chlb)和各种类胡萝卜素分子。不同的光合色素由于其分子轨道分布不同而具有不同的跃迁能级,因而吸收具有不同能量的光子,由上所述,叶绿体吸收光能,产生的激发能具有两个去向,一个是光化学淬灭,这部分能量转化为生物质;而另一部分能量为非光化学淬灭,这部分能量作为维持细胞活动,过剩能量的消耗和光破坏。为了能够尽可能增加农作物的光合产量,必须尽量减少非光化学淬灭,增加光化学淬灭的成分。
4.叶绿素不同激发态的命运:1、二级激发态(蓝光区,或者400nm周围的光)弛豫到一级激发态(红光区,或者说是650nm周围的能量),一级激发态在弛豫到基态的过程中,可以引起一系列物理化学反应,最后将能量转换为化学能,体现为光合作用碳固定、产生生物量。但是,研究表明,由于太阳能很强,植物经常会面临能量过剩的状态,叶绿素吸收的能量并不能100%地转换为化学能,而是有相当部分要作为过剩能量消耗掉,所以,一级激发态的能量可以有以下不同的出路(图2):1、光化学淬灭(光合作用、或者生物量积累),这部分能量是经过一系列氧化还原反应,最终转换成化学能(产生生物量);2、经过斯托克斯位移(stokes shift)放出一部分能量,发出荧光(基础荧光,过剩能量消耗,光保护);3、直接将能量传递给类胡萝卜素,以热的形式消耗过剩能量(过剩能量消耗,光保护);4、叶绿素单线态激发态发生电子自旋,产生三线态激发态(长寿命激发态),而叶绿素的三线态激发态会与氧气作用产生活性氧自由基(ros),ros会引起光合色素氧化(光漂白)与光合作用色素蛋白复合体解体(光破坏),最后引起光合效率降低或者细胞死亡。所以,长寿命三线态的产生是破坏光合作用的。
5.不同叶绿素分子结构的差异导致其激发态的寿命不同,chlb的激发能寿命非常短,目前科学的认知认为,chlb能够非常迅速地将其激发能传递给chla,而chla的激发能寿命相对比较长,并且会根据其所在的介质不同而改变,当chla的能量不能及时地转化成化学能,则会引起激发电子的自旋,产生长寿命的三线态激发态,进而导致活性氧的产生,引起光破坏。如果能通过调控减少chla二级激发态的量,可以大大地降低叶绿素三线态的危害,达到合理地利用和分配类囊体膜的激发态分布,达到最佳的光合效率,增加作物产量。
6.现有技术中,对于入射光的不同波长光产生的一级激发态的去向以及对于光合作用效率的影响已经进行了大量的研究,但是,关于二级激发态对于不同一级激发态的影响、以及最终对于光合作用效率的影响的问题和技术,并没有解决。
技术实现要素:
7.为了解决上述通过降低叶绿素a二级激发态提高光合效率的方法调节问题,本发明提供一种通过降低叶绿素a二级激发态提高光合效率的方法,公开了通过降低入射光中能够引起叶绿素a(chla)二级激发态的光质的权重而提高作物产量的方法,通过改变入射光谱中“长/短波”比例,达到降低光破坏、进而提高植物光合作用效率。
8.叶绿素的吸收能级根据其分子结构及所处的环境不同而不同,而叶绿体则也根据入射光的能级(波长)引起不同叶绿素的激发态跃迁,而按照本技术对植物叶绿素进行不同光质条件下的激发,并利用以下公式计算激发能不同的弛豫方式:1chl
* = fn+pq+npq
ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ
(1)其中:1chl
*
:叶绿素分子吸收太阳能产生的单线态激发能;fn:基础荧光(维持植物本身的热量平衡,分子振动等等);pq:光化学淬灭(经过一系列电子传递过程,进行能量转换,产生生物能atp,还原势nadph2);npq:非光化学淬灭,此部分的能量消耗是光合作用中非常重要的过程,即通过以不同形式消耗掉过剩能量,避免过剩能量对于光合器官的伤害,所谓光保护过程。光合作用的npq有不同形式,根据能量淬灭过程的机理分为以下三个部分:npq=qe + qt + qi
ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ
(2)其中:qe: 快相激发能淬灭:这一过程直接和高能淬灭有关,由类囊体膜跨膜质子梯度调控,与叶黄素循环和玉米黄素的产生有关,是一个非常迅速的能量淬灭过程,持续时间为飞秒到一分钟内,这部分淬灭过程能够有效地消耗掉过剩能量,保护光合组织; qt:中间相激发能淬灭:这个过程与状态转换有关,通过光合膜蛋白的磷酸化/去磷酸化过程,产生蛋白质的构象变化,进而在光合膜不同光系统之间迁移,通过这种形式在不同光系统之间分配激发能,这种形式的能量淬灭持续时间比qe长,能够持续10分钟左右;qi:慢相激发能淬灭:也称光破坏/光抑制。是一个与光合膜色素蛋白复合体解体有关的过程,这部分淬灭方式持续的时间很长,达到几个小时以上,甚至引起细胞死亡。
9.由此确定本技术的技术方案:通过降低叶绿素a二级激发态提高光合效率的方法。
10.具体来说,本技术的技术方案包括:一种通过降低叶绿素a二级激发态提高光合效率的方法,照射植物的光线中波长
大于440nm光权重高的光作为长波光,波长小于或者等于440nm光的权重高的光作为短波光,使得长波光照射的植物产生的qe与短波光照射植物的qe之比:qe
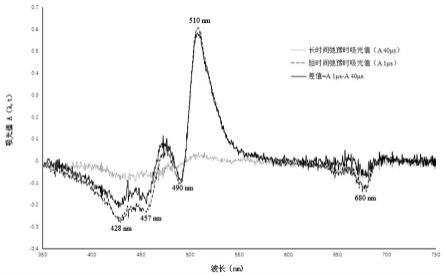
长波
/qe
短波
》1.1,长波光中波长大于440nm光权重高是指∑(380nm-440nm)/∑(441nm-550nm)《1;短波光中的小于或者等于440nm光的权重高是指∑(380nm-440nm)/∑(441nm-550nm)》1;∑为光谱图中相应区域的积分面积。
11.上述光线可以采用包括自然光的光源或者包括led的光源。
12.进一步的,当其中照射植物的光线采用包括自然光的光线时,qe
长波
/qe
短波
》3.0;或者照射植物的光线采用白色led光源,其中蓝光峰值为470nm的长波led入射光作为长蓝光,相比较蓝光峰值为440nm的短波led入射光作为短蓝光,使得长蓝光照射植物产生的qe与短蓝光照射植物产生的qe之比:qe
长蓝光
/qe
短蓝光
》1.9。
13.进一步的,qe
长波
/qe
短波
》3.1,或者 qe
长蓝光
/qe
短蓝光
》1.95;进一步的,qi
长波
/qi
短波
《0.80, qt
长波
/qt
短波
《0.80;进一步的,qi
长蓝光
/qi
短蓝光
《0.8,qt
长蓝光
/qt
短蓝光
《0.90;进一步的,npq
长波
/npq
短波
》1.2,或者 npq
长蓝光
/npq
短蓝光
》1.1;本技术可以达到增加光合作用中npq量,降低光破坏,提高光合产量目的。
14.实现:生物量w
长蓝光
/w
短蓝光
》1.1,或者净光合效率pn
长波
/pn
短波
》1.3;进一步的,生物量w
长蓝光
/w
短蓝光
》1.3,净光合效率pn
长波
/pn
短波
》1.5。
15.此外,本技术提供一种根据上述方法通过降低叶绿素a二级激发态提高光合效率的用途,这种方法能够降低叶绿素的光漂白,有效地提高植物的光能利用效率,提高植物的光合作用效率。所述植物包括穿心莲和拟南芥。根据这一发明的理论应用于植物的光合作用测量,发现如果在自然光条件下, 430nm的光处理,增加光破坏量30%,降低净光合作用44%。反之,则能够相应地降低30%光破坏,提高净光合作用效率,提高植物生物量30-40%。
附图说明
16.图1:分子吸收光谱和能级跃迁图。
17.图2:叶绿体中叶绿素一级激发态的能量去向图。
18.图3:光合作用天线分子的典型的瞬态吸收光谱动力学变化图。
19.图4:植物在不同光照条件下光合作用非光化学淬灭调查对比图。
20.图5:不同白色led灯光谱分布图。
21.图6:不同led灯条件下生长4周后的穿心莲照片图。
具体实施方式
22.植物进行光合作用的能量来自于太阳能,植物吸收太阳能主要的光合色素是叶绿素chla和 chlb。利用超快激光泵浦-探测技术检验了光合天线(lhcii)的瞬态吸收光谱,分析了在不同激发光条件下lhcii的激发能的能量分布。根据不同光合色素分子(chla、chlb)的跃迁矩不同,分别用437nm和475nm的光激发lhcii系统中的chla或者chlb分子,通过观察光合天线所结合的光合色素chla或者chlb分子的电子吸收泵浦光跃迁到激发态后的弛豫过程,分析和判断lhcii色素电子激发后产生的单线态和三线态状态、以及三线态的传递、分布、以及淬灭过程。图3表示出用不同波长的泵浦激光(437nm和475nm)分别激发光合作用
捕光天线中的chla和chlb之后的三线态分布情况,反应出系统中产生单线态能量传递给三线态而淬灭的过程。
23.从图3可以看出,捕光天线的色素激发,经过短时间弛豫(1μs)后,其瞬态吸收光谱呈现出一个510nm正峰(类胡萝卜素三线态峰)、在soret区有三个负峰:428nm、457nm、和490nm峰,这是单线态叶黄素和叶绿素交互作用,一个680nm的负峰(三线态叶黄素和叶绿素交互作用),激发态经过长时间弛豫(40μs)后,从其瞬态吸收光谱中,可以观察到一个680nm的负峰,反映出不能将能量传递给类胡萝卜素三线态的叶绿素激发态(叶绿素的峰:自由叶绿素),随着510nm峰的消失,单线态叶黄素和叶绿素交互作用的峰消失。soret区的负峰大幅度降低,从不同时间(t)弛豫过程的差谱(1μs-40μs)可以看出,在这个条件下,lhcii的单线态激发态基本上可以将能量传递给类胡萝卜素,产生类胡萝卜素三线态,并通过类胡萝卜素的三线态淬灭,回到基态过程。但是,从40μs的瞬态吸收光谱也可以看到,lhcii中的叶绿素激发态还有一小部分能量以叶绿素三线态的形式残留(680nm),这部分能量并不能传递到类胡萝卜素、并通过类胡萝卜素淬灭,而叶绿素三线态的寿命比较长,可以将能量传递给细胞中的氧分子,产生活性氧自由基(ros),造成细胞氧化损伤,引起光合色素的光漂白,光合作用色素蛋白复合体的解体(光破坏),甚至引起细胞死亡。而光合作用色素蛋白复合体的解体的必然结果是光合能力的大大降低。因此,需要提高光合作用效率,增加作物产量,需要尽量降低长寿命的叶绿素三线态激发态,减少光破坏。
24.从两次瞬态吸收光谱的差谱可以由以下计算推导出不同色素产生的三线态激发态的量:根据朗伯-比耳定律,物质的吸光度与浓度与液层厚度的关系:其中,a为吸收程度,ε为摩尔消光系数,c 为溶液浓度,l为比色皿路径,为1。
25.因此,类胡萝卜素三线态吸收计算公式由510nm处的吸收差值决定(4),同理,chla和chlb的残留三线态量由以下计算过程(chla的吸收峰为 674nm,chlb为 650nm)(5)和(6)推导出,将chla和chlb的三线态的摩尔消光系数与类胡萝卜素的相关联(7):
其中:
+
指的是三线态,ε指的是摩尔消光系数。δa
car
:两次瞬态吸收光谱差谱中类胡萝卜素在510nm的值;δa
chla
、δa
chlb
:分别表示两次瞬态吸收光谱差谱中chla在674nm处和chlb在650nm处的值;类胡萝卜素三线态的摩尔消光系数。ε
chlb
、ε
chla
分别为chlb或chla的摩尔消光系数。、:分别表示chla或chlb的残留三线态量。y:相对三线态产率。
26.根据上述计算过程,从图3计算出不同波长泵浦光激发不同光合色素(chla和chlb)引起的激发态在经过40微秒(t=40μs)弛豫后,光合天线经过不同色素的三线态能量淬灭的比例,其结果如表1所示。从表1中,我们发现,激发chlb,天线的能量能有75%能量通过类胡萝卜素三线态转变成热量淬灭,而激发chla,只有59%的能量能够通过类胡萝卜素淬灭,这个结果表明了,激发chla二级激发态的光大大降低了lhcii类胡萝卜素的三线态淬灭比例(cc),提高了有害的chla三线态的产生,而chla三线态是破坏植物光合器官的关键因素。所以,降低入射光中小于440 nm波长的权重,可以在保证尽可能大量吸收光量子的情况下,尽量降低产生chla的三线态。避免光破坏的成分,增加光合产量。
27.表1:天然光合作用捕光天线中不同色素的三线态状态**:ca:通过叶绿素a淬灭的三线态,cb:通过叶绿素b淬灭三线态,cc:通过类胡萝卜素淬灭的三线态,cc/(ca+cb+cc) 通过类胡萝卜素淬灭的三线态在总三线态中的比例,y:相对三线态产率。
28.表1的结果展示了激发光合作用捕光天线中(lhcii)不同叶绿素分子所产生就激发能的淬灭规律,以下的实验结果证明了,lhcii的结果在植物叶片(叶绿体)中也能得到证明:光合作用通过吸收光能,产生激发能,并将激发能转化成化学能(生物能atp 和还原势 nadph2),进而达到固定二氧化碳、合成碳水化合物的目的。叶绿体中进行固定二氧化碳,合
成碳水化合物的主要能量是通过叶绿素(chla、chlb)吸收、传递和转化的。在蓝光区,chla的吸收峰在437nm左右,而chlb的吸收峰在475nm左右,本发明主要提出了通过降低入射光中能够产生chla二级激发态的光量子的权重,可以减少光系统中的叶绿素三线态激发态,降低叶绿体中由于光破坏的引起的能量消耗,从而达到增加光合作用效率目的的方法。
29.具体来说,一种通过降低叶绿素a二级激发态提高光合效率的方法,照射植物的光线中波长大于440nm光权重高的光作为长波光,波长小于或者等于440nm光的权重高的光作为短波光,使得长波光照射的植物产生的qe与短波光照射植物的qe之比:qe
长波
/qe
短波
》1.1,长波光中波长大于440nm光权重高是指∑(380nm-440nm)/∑(441nm-550nm)《1;短波光中的小于或者等于440nm光的权重高是指∑(380nm-440nm)/∑(441nm-550nm)》1;∑为光谱图中相应区域的积分面积。
30.上述光线可以采用包括自然光的光源或者包括led的光源。
31.进一步的,当其中照射植物的光线采用包括自然光的光线时,qe
长波
/qe
短波
》3.0;或者照射植物的光线采用白色led光源,其中蓝光峰值为470nm的长波led入射光作为长蓝光,相比较蓝光峰值为440nm的短波led入射光作为短蓝光,使得长蓝光照射植物产生的qe与短蓝光照射植物产生的qe之比:qe
长蓝光
/qe
短蓝光
》1.9。
32.进一步的,qe
长波
/qe
短波
》3.1,或者 qe
长蓝光
/qe
短蓝光
》1.95;进一步的,qi
长波
/qi
短波
《0.80, qt
长波
/qt
短波
《0.80;进一步的,qi
长蓝光
/qi
短蓝光
《0.8,qt
长蓝光
/qt
短蓝光
《0.90;进一步的,npq
长波
/npq
短波
》1.2,或者 npq
长蓝光
/npq
短蓝光
》1.1;本技术可以达到增加光合作用中npq量,降低光破坏,提高光合产量目的。
33.实现:生物量w
长蓝光
/w
短蓝光
》1.1,或者净光合效率pn
长波
/pn
短波
》1.3;进一步的,生物量w
长蓝光
/w
短蓝光
》1.3,净光合效率pn
长波
/pn
短波
》1.5。
34.下面我们针对中药穿心莲和植物拟南芥应用本技术的方法进行下述实验:实施例1:1、叶绿素光合作用能量平衡:在自然光条件下,选择两盆规格相同的穿心莲植物,一盆用430nm蓝光(自然光+430nm光)处理。另一盆不处理(自然光)。两个处理的光通量相同,处理时间18周,利用光合作用测定仪6400测定净光合效率(单位时间固定的二氧化碳的量),通过荧光仪测定叶片光系统ii叶绿素荧光,分析叶片接受太阳能后产生的类囊体膜激发能淬灭的组分,分析其中非光化学淬灭形式,进而分析光合作用器官中的能量转换效率。
35.叶绿体中光合作用叶绿素分子吸收太阳能,产生单线态激发能1chl
*
,这部分能量将传递到反应中心,经过一系列氧化还原反应,最终转化为生物能(atp)和还原势(nadph2),提供固定co2所需的能量和电子,合成碳水化合物(c6h
12
o6)。但是,由于根据环境条件的变化,叶绿体从吸收太阳光子、产生激发能,到固定co2,产生碳水化合物的不同环节会产生能量不匹配的现象,所以,吸收光子产生的单线态激发能并不能全部用来进行固定co2的化学反应,而是如图2所示,以光化学淬灭和非光化学淬灭(npq)等形式消耗掉。如前所述,npq可以概括为三种形式,1)物理淬灭,这部分能量淬灭发生的很快,是个高效地淬灭过剩能量的途径:2)状态转换,这部分能量用来合理分配类囊体膜上的能量分配;3)光破坏,或者光抑制。这部分能量淬灭不利于光合积累,有可能引起细胞凋亡。
36.npq(图4)的测量方式:为了测量植物增加了430nm蓝光处理后在类囊体膜上的激发能的淬灭过程,用便携式调制叶绿素荧光仪pam-2500对于用430nm光处理过的穿心莲叶片进行慢速荧光诱导曲线测量,植物进行30分钟暗适应,开始测量,测量光是非常弱的红光(650 nm,0.15 μmol m-2 s-1
),此时的荧光为初始荧光(f0),打一次饱和脉冲光(4500 μmol m-2 s-1
,0.8s),测得暗适应条件下的最大荧光(fm),当荧光降低接近 f
0 点后,打开活化光(100 μmol m-2 s-1
),接着打一个饱和脉冲光,测得光适应条件下的最大荧光(fm′
),之后,用活化光照射一次,然后,每隔40s打一次饱和脉冲光,测定fm′
,根据公式:npq=( fm‑ꢀfm
′
)/ fm′ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ
(8)计算npq(无量纲),10分钟后,进行暗弛豫30分钟,期间每隔1分钟测量一次,测定npq发生和弛豫过程(参见图4),根据暗弛豫过程的半对数曲线,对npq的成分(qe、qt、qi)进行分解。为了比较不同形式npq的变化对于光合作用效率的影响测定的净光合作用一并表示在npq组分分析的表2中。
37.表2:自然光加上短波光的穿心莲净光合效率和非光化学淬灭从表2可以看出,附加了430nm光源后,光破坏(qi)分量从0.0479增加到了0.0626,增加了30%。更重要的是净光合作用效率pn(即单位时间的co2固定量)从31.84μmolm-2
s-1
降低到17.57μmolm-2
s-1
,这一实验证明了chla的二级激发态不利于迅速去除类囊体膜的过剩能量,容易产生对光系统的伤害,降低光合作用。
38.总结上述结果,可以看出,与光合作用天线分子的典型的瞬态吸收光谱动力学变化的结论一致,降低chla二级激发态主要虽然增加了非光化学淬灭,但是,碳固定量反而增加,这个的机理是增加蓝光造成了类囊体膜上快相激发能淬灭(qe)的降低和光抑制,即慢相激发能淬灭(qi)的增加,从表2中可以清楚地看到这个结果。
39.实施例2(实际应用案列:穿心莲):已经证明了chla的二级激发态引起过剩能量积累,引起光破坏,降低净光合效率,我们通过改变led芯片的方式,将培养间培养穿心莲的led光谱在蓝光区峰值红移了30nm,几乎消除了小于440nm的光在整个光谱中的权重(图5)。
40.穿心莲种植条件:在植物工厂环境下,培养基质经过120℃高温消毒,营养液为hogland培养液,穿心莲在不同光质条件生长4周(图6)。收获洗净根上沾的基质,吸去水分,秤鲜重,分别为0.22克和0.16克。计算不同实验光条件下生长植株的鲜重变化,降低chla二级激发态可以增加生物量37%。
41.表3:不同led光条件下穿心莲生物量积累及非光化学淬灭测量
实施例3(实际应用案列:拟南芥):拟南芥种植环境的光源为led冷光灯,红光区光谱一致,蓝光区呈现一个峰,处理1的峰值在470nm,处理2的峰值在440nm,总的光通量调节为140 μmol m-2
s-1
。培养基质经过120℃高温消毒,营养液为hogland培养液,生长室温度为22℃,在不同光质条件生长4周。收获后洗净根上沾的基质,吸去水分,秤鲜重,计算不同实验光条件下生长植株的鲜重变化来测定生物量,降低chla二级激发态可以增加生物量15%。
42.表4:不同led光条件下拟南芥生物量积累及非光化学淬灭测量由此可见,在本技术的长波光或者长蓝光条件下,有效降低入射光的短波成分,可以有效地降低chla二级激发态,进而增加类囊体膜上快相激发能淬灭(qe),使得光合器官迅速地去除有害激发状态,降低光抑制(qi),保护光系统结构和类囊体膜,增加植物光合作用效率。经验证,上述方法可以在中药种植领域得到应用,并通过光调节叶绿素激发提高中药种植效率。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1