ACGs的新应用的制作方法
本发明属于医学检测领域,具体的是涉及acgs的新应用。
背景技术:
1971年人们从红豆杉植物中提取得到抗癌药物紫杉醇后,人们开始更加关注天然抗癌化合物。1982年,jolad等在美洲植物uvariaaccuminata(番荔枝科)的根部提取物中分离得到天然化合物uvaricinin,发现它对小鼠白血病细胞有强烈杀伤作用,后来科学家发现它还具有强大细胞毒性、t细胞抑制活性、抗肿瘤、抗虫、抗寄生虫、杀菌等药理活性,由此掀起了植物化学家在各种番荔枝科植物中分离这类化合物的热潮以及科学家对acgs的这些药理活性进行了深层次、宽范围、多角度的研究,被科学家们预测为继紫杉醇之后天然化合物中的“明日抗癌之星”。天然番荔枝内酯中活性最强的有4个分别是bullatacin,asimicin,trilobacin和trilobin,由于种类太多导致还无法有效分离单独一种成分,目前主要是以番荔枝内酯化合物类化合物(annonaceousacetogenins,acgs)来进行研究。acgs的结构主要是一系列含35到39个碳原子构成的基本骨架,末端通常具有一个a,β,γ不饱和五元内酯环,有时异构成酮式五元内酯环,分子链中具有0到3个四氢呋喃环,并常有多个如羟基,乙酰基,酮基等含氧取代基团,分子中含有多个手性碳[yanghj,lix,zhangn,etal.twonewcytotoxicacetogeninsfromannonasquamosa[j].jasiannatprodres,2009.11(3):p.250-6.]
[kojiman,tanakat.medicinalchemistryofannonaceousacetogenins:
design,synthesis,andbiologicalevaluationofnovelanalogues[j].
molecules,2009.14(9):p.3621-61.]。在过去20多年,有更多的证据证明acgs在不同的肿瘤中都有抗癌作用。acgs中aa005能够导致大肠癌细胞生长抑制并诱导细胞凋亡,还可以导致结肠癌细胞的自噬。同样,acgs中desacetyluvaricin抑制结直肠癌细胞系sw480的增长。在异种移植小鼠试验中,acgs模拟药thiophene-3-carboxamide强烈抑制人肺癌细胞株nci-h23的生长。在番荔枝属graviola叶子提取的黄酮类化合物对前列腺癌具有保护作用]。从番荔枝属annonamuricata果实中提取的三种新的acgs对人的前列腺癌pc-3细胞具有抑制增殖作用,a.muricata在hela细胞中具有相同的作用。等等。
与makp,stat,pi3k等信号通路一样,notch信号通路是细胞乃至人体中关键的信号通路之一,参与了细胞增殖、凋亡和分化等决定细胞命运的关键环节。notch信号通路中包含5个配体(dll1、dll3、dll4、jagged1和jagged2)和4个受体(notch1/2/3/4),配受体的相互作用可以促使受体释放胞内段,胞内段进入细胞核内与相关蛋白结合进而促使下游基因hes1和hey1等表达。已有研究表明通路内相关蛋白编码基因的突变常常会导致各种肿瘤的发生和恶性发展。然而,在不同肿瘤中,notch信号通路所起的作用并不完全相同,比如,在结直肠癌、乳腺癌、肺癌、神经母细胞瘤中notch信号通路起致癌作用,而在皮肤鳞状细胞癌、宫颈癌、肝细胞癌、肺癌、胃肠道癌和神经内分泌肿瘤中notch表现出肿瘤抑制作用。所以,notch信号通路的多面性特征提示检测其在特定肿瘤类型中的特定激活模式和潜在的作用方式是必要的。
gc指发生在胃粘膜上皮细胞发生的恶性肿瘤,它是世界范围内最常见的恶性肿瘤之一。胃癌发病率在全球癌症发病中位居第五,死亡率位居第三,约占全球癌症7%的病例和9%的死亡数,其发病在发展中国家更为常见。由此可见,胃癌的情况依然不容乐观,而这很大程度上是由于胃癌在早期诊断中并无明显的特异症状,确诊状态下已经是晚期甚至出现转移。目前胃癌的主要治疗方法包括手术切除、化疗和放疗,但这些传统治疗方式效果并不理想。因此,人们正在积极探索新的治疗方法。
目前,已经有大量的研究探讨了notch信号通路和gc之间的联系,据一篇meta分析报道,相对于正常胃组织,gc细胞中notch1,notch2,dll4和hes1的表达均有所上升,而相关研究多数集中于notch1和notch2蛋白。notch1和notch2受体胞内域能促进人胃腺癌细胞sc-m1的增殖,在gc细胞mkn-45中下调notch2能增强肿瘤细胞侵袭能力。有研究发现在gc中过表达notch2蛋白引起的下游蛋白hes1核转移效果要比notch1蛋白显著,这表明notch2信号在gc致癌作用和发展更重要。还有研究发现notch1和notch2的高表达与gc早期阶段有着亲密的联系,高表达notch1的早期gc生存率显著增加,而高表达notch2的早期肠胃型肿瘤能够更好的生存。notch2信号通路还有可能通过抑制pi3k-akt信号通路来负调控gc细胞侵袭,抑制notch2信号通路使得gc细胞侵袭力增强。很多文献都表明notch2与gc有着紧密联系,但其作用机制却还未阐明。
直到现在,对acgs的抗肿瘤作用机制尚不十分清楚,acgs是否可以用于防治胃癌,目前更是未知。
技术实现要素:
本发明的目的之一在于提供一种acgs在制药中的新应用。
实现上述目的的技术方案如下。
acgs在制备防治胃癌的药物中的应用。
本发明的另一目的是提供一种治疗胃癌的药物。
具体技术方案如下。
一种治疗胃癌的药物,其活性成份包括有acgs。
我们的研究表明,acgs抑制gc细胞增殖,促进细胞凋亡,阻滞细胞周期停留在g0-g1期。随着acgs浓度升高对gc的增殖抑制,促进凋亡和阻滞细胞周期停留在g0-g1期效果越明显,这说明acgs对gc的作用具有剂量依赖性。我们首次证明了acgs通过作notch2信号通路发挥作用,acgs提高gc中notch2的表达,过表达notch2促进acgs诱导的细胞毒作用,notch2-sirna干扰则降低了acgs诱导的细胞毒作用。通过上述研究发现,acgs可用于防治胃癌。
附图说明
图1为q-pcr检测gc细胞中notch2的表达。
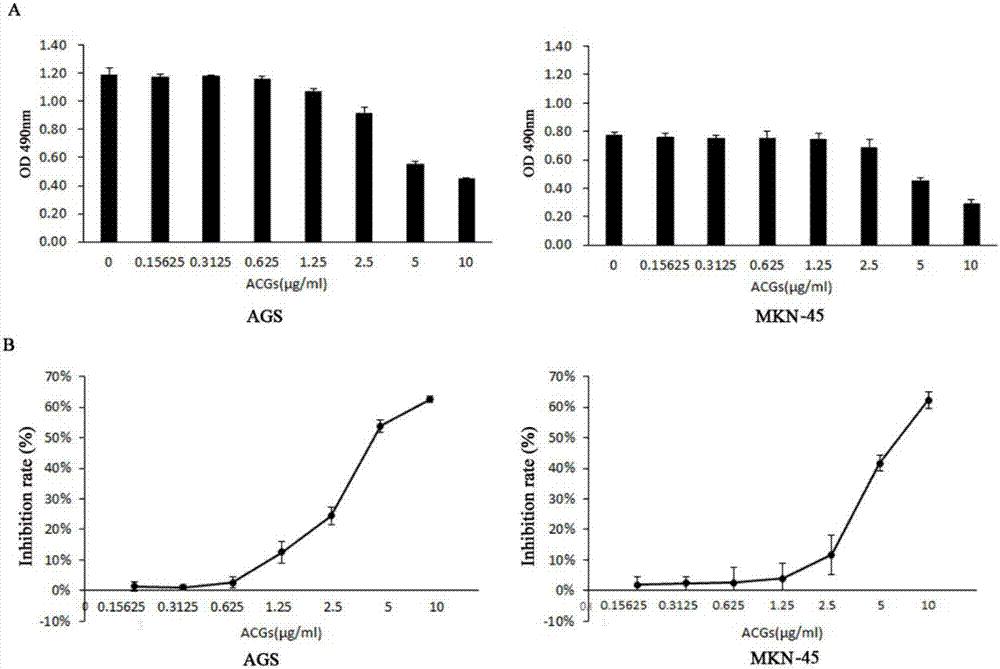
图2mts检测acgs对gc细胞增殖的抑制作用,其中,a:acgs作用ags、mkn-4524h后,mts检测细胞增殖;b:由mts结果换算出的抑制率结果。
图3acgs对gc细胞的增殖抑制能力呈时间和剂量依赖性,其中,a:2.5、5、10ug/mlacgs刺激ags细胞12、24、36hmts检测细胞增殖;b:2.5、5、10ug/mlacgs刺激mkn-45细胞12、24、36hmts检测细胞增殖。
图4显微镜下观察acgs抑制gc细胞的增殖,其中,2.5、5、10ug/mlacgs刺激ags、mkn-45细胞36h显微镜下细胞状态结果。
图5acgs对gc细胞凋亡的影响,其中,a:5ug/mlacgs分别作用ags、mkn-45细胞12、24、36h后流式细胞技术检测凋亡;b:2.5、5、10ug/mlacgs分别作用ags、mkn-45细胞36h后流式细胞技术检测凋亡。
图6acgs对gc细胞细胞周期的影响,其中,a:2.5、5、10ug/mlacgs作用ags细胞36h后流式细胞技术检测细胞周期;b:2.5、5、10ug/mlacgs作用mkn-45细胞36h后流式细胞技术检测细胞周期。
图7westernblot检测acgs刺激gc细胞后notch2蛋白表达,其中,通过westernblot以β-actin为内控对gc细胞notch2蛋白进行分析,直方图表示每组细胞notch2蛋白相对量。
图8westernblot检测acgs刺激gc细胞后hes1的表达,其中,通过westernblot以β-actin为内控对gc细胞hes1蛋白进行分析,直方图表示每组细胞notch2蛋白相对量。
图9acgs对转染notch2-cdna的gc细胞的影响,其中,a:westernblot检测notch2的表达;b:过表达notch2对gc细胞增殖影响。
图10acgs对转染notch2-sirna的gc细胞的影响,其中,a:westernblot检测notch2的表达;b:notch2-sirna对gc细胞增殖影。
具体实施方式
为了便于理解本发明,下面将参照相关附图对本发明进行更全面的描述。附图中给出了本发明的较佳实施例。但是,本发明可以以许多不同的形式来实现,并不限于本文所描述的实施例。相反地,提供这些实施例的目的是使对本发明的公开内容的理解更加透彻全面。
除非另有定义,本文所使用的所有的技术和科学术语与属于本发明的技术领域的技术人员通常理解的含义相同。本文中在本发明的说明书中所使用的术语只是为了描述具体的实施例的目的,不是旨在于限制本发明。本文所使用的术语“和/或”包括一个或多个相关的所列项目的任意的和所有的组合。
实施例
1、实验材料
1.1细胞株
胃正常上皮细胞ges-1,gc细胞株ags、sgc-7901、mgc-803、mkn-28、mkn-45。(所有细胞株均为本科室存种)
3.1.2主要试剂盒仪器
1.3主要试剂的配制
1.3.1acgs母液的配制
秤取32mgacgs固体溶于3.2mldmso中配成浓度为10mg/ml的母液,过滤后分装-20℃保存。
1.3.20.2medta.na2溶液配制
秤取74.448mgedta.na2用去离子水溶解定容至1ml,过滤后4℃保存。
1.3.32mg/mlpi溶液配制
将10mgpi用去离子水溶解定容至5ml,-20℃保存。
1.3.410%tritonx-100溶液配制
将1mltritonx-100溶于9ml去离子水中,4℃保存。
1.3.550mg/mlpi工作液配制
1.3.55mg/mlmts溶液的配制
秤取5mgmts粉末,用pbs溶解定容至1ml,过滤-20℃避光保存。
2、实验方法
1)q-pcr检测notch2在gc细胞中的表达
为了评估notch2在gc中的作用,我们筛选5株gc细胞系通过q-pcr技术检测gc细胞中notch2的mrna水平。用10%fbs的高糖dmem培养基培养6株细胞,待细胞长满10cm培养皿后,pbs洗3遍加0.25%胰蛋白酶消化,然后用完全培养基终止胰酶消化,吹打下来细胞,1000rcf/min、5min离心,去除上清,再用pbs清洗3遍,用rnaisoplus试剂按照说明书步骤提取细胞总rna。
第一链cdna合成按照primescriptrtreagentkitwithgdnaeraser试剂盒说明书完成及real-timequantitativepcr按照primescriptⅱreversetranscriptase试剂操作进行,以β-actin为内参(nm_001101.3),q-pcr引物见(表2):
表2real-timequantitativepcr特异引物
混匀,42℃,2分钟,4℃储存。
混匀,37℃、15min,85℃、5s,4℃保存。
q-pcr反应,20ul体系配制方法如下:
混匀,反应程序为95℃30s,95℃5s、60℃34s40个循环,95℃15s,60℃1min,95℃15s。定量方法参照,以2-δδct值表示目的基因mrna的相对扩增量。
2)mts检测gc细胞增殖
为了探索acgs是否抑制gc细胞的生长,用mts实验检测细胞的增殖。培养gc细胞,待细胞长满后消化铺96孔板,每个空3000个gc细胞,37℃培养过夜,将acgs用高糖dmem培养基稀释为0、0.15625、0.3125、0.625、1.25、2.5、5、10μg/ml8个浓度,每个浓度组设有96孔板中5个孔gc细胞,10个浓度组acgs同时作用24小时后加20ulmts试剂37℃孵育30min,酶标仪490nm读值。同时,通过mts实验数据计算出细胞增殖的抑制率,由抑制率检测acgs对gc细胞抑制作用效果(抑制率=1-实验组/对照组)。另外,以2.5、5、10μg/ml低、中、高3个浓度的acgs(用10%fbs的高糖dmem培养基稀释)刺激gc细胞12、24、36h后,用mts检测acgs作用于gc细胞是否具有时间依赖性,并镜下观察细胞生长状态。
3)流式细胞技术检测gc细胞凋亡
为了探索acgs抑制gc细胞生长是否伴随着诱导细胞凋亡,以5μg/ml的acgs刺激gc细胞12、24、36h,用不含edta的胰蛋白酶消化细胞后annexinⅴ/pi染色流式细胞术分析。此外,以2.5、5、10μg/ml的acgs刺激gc细胞36h,进行上述分析。流式细胞技术检测gc细胞周期
为了探索acgs对gc细胞的生长抑制是否伴随着细胞周期阻滞,以2.5、5、10μg/ml的acgs刺激gc细胞36h,用不含edta的胰蛋白酶消化细胞后pi染色流式细胞术分析。
4)westernblot检测acgs作用gc细胞后notch2的表达
为了确定acgs对gc细胞的作用是否通过notch2信号通路,以2.5、5、10μg/ml的acgs刺激gc细胞36h,消化收集细胞后用ripa裂解液裂解细胞,westernblot检测gc细胞中notch2蛋白水平。
5)westernblot检测下游蛋白hes1的表达
为了确定acgs对gc细胞的作用是否影响notch2信号通路的下游蛋白hes1对3.2.6中获取的细胞蛋白通过westernblot检测gc细胞中hes1蛋白水平。
6)过表达n2icd对gc细胞增殖的影响
构建n2icd/pcmv-tag4质粒:notch2蛋白胞内区基因的扩增根据ncbi数据库中的notch2的基因序列(af308601.1),设计扩增蛋白胞内区基因n2icd的引物(表1),下划线为酶切位点;以hela细胞的总rna为模板,rt-pcr扩增n2icd,反应为50℃30min,94℃2min,94℃30s、57℃30s、72℃2.5min,35个循环,72℃5min。rt-pcr产物经1%琼脂糖凝胶电泳鉴定并切胶回收。
n2icd引物
n2icd与pmd-18t载体连接与鉴定将切胶回收的n2icd与克隆载体pmd-18t在16℃下连接3h,连接产物转化到大肠杆菌dh5α中,从含有青霉素、iptg、x-gal的lb固体培养板挑取白色单菌落过夜培养和提取质粒,经pcr、双酶切及测序鉴定。
真核表达重组体n2icd/pcmv-tag4的构建和鉴定用xhoⅰ和notⅰ双酶切n2icd/pmd-18t和真核载体pcmv-tag4,将酶切过后的pcmv-tag4和n2icd分别进行切胶回收。将纯化的pcmv-tag4和n2icd在t4连接酶的作用下16℃连接过夜,产物转化大肠杆菌dh5α,从含有卡那霉素的lb固体培养板挑取单菌落过夜培养和提取质粒,经双酶切及测序鉴定无误。
为了进一步证实notch2在acgs中的作用,n2icd/pcmv-tag4质粒转染gc细胞24h后,空载体pcmv-tag4转染gc细胞作为对照,一部分细胞加5μg/mlacgs刺激36h,收集细胞westernblot检测gc细胞中notch2的蛋白水平,另一部分消化下来铺96孔板,10%fbs高糖dmem培养基培养过夜,待细胞贴壁后加5μg/mlacgs刺激12、24、36h后,通过mts实验获取od490nm值统计分析数据。
7)notch2-sirna干扰对gc细胞增殖的影响
通过notch2-sirna下调gc细胞中notch2的表达,以notch2-sirna和control-sirna转染gc细胞24h后,一部分细胞加5μg/mlacgs刺激36h,收集细胞westernblot检测gc细胞中notch2的蛋白水平,另一部分消化下来铺96孔板,10%fbs高糖dmem培养基培养过夜,待细胞贴壁后加5μg/mlacgs刺激12、24、36h后,通过mts实验获取od490nm值统计分析数据。
8)统计学分析
数据采用spss20.0软件进行统计分析,两组间数据的比较采用独立样本t检验的方法,三组或三组以上数据的比较,采用one-wayanova分析。
3.2实验结果
1)在基因水平上,notch2在gc细胞中呈高或低的表达
q-pcr结果显示,与正常胃粘膜上皮细胞ges-1相比,notch2在gc细胞株ags、sgc-7901中的表达高,而在gc细胞株mgc-803、mkn-28、mkn-45中的表达低。因此,我们从表达高低组中分别选出一株代表性细胞ags、mkn-45做后续实验研究。参见图1。
2)acgs抑制gc细胞的增殖
利用不同浓度的acgs分别作用ags和mnk-45后,通过mts实验检测acgs对gc细胞增殖的影响。由图2a可知,随着acgs浓度的升高,其对ags和mkn-45的生长抑制作用越强。从图2b可知acgs对ags和mkn-45的抑制率折线图趋势大体一致,经计算得知acgs对ags的ic50=5.02μg/ml对mkn-45的ic50=6.25μg/ml。
3)acgs对gc细胞增殖的抑制作用呈时间和剂量依赖性
以2.5、5、10μg/ml的acgs分别作用于ags和mkn-45胃癌细胞株12h、24h、36h后,通过mts实验检测不同浓度和不同时间作用下acgs对胃癌细胞的增殖抑制作用。从图3可知acgs对ags、mkn-45的生长抑制作用具有时间和剂量依赖性,随着时间的增加和acgs浓度的升高对细胞生长的抑制越明显。显微镜下观察发现(图4)随着acgs浓度的升高,细胞数及密度逐渐降低。
4)acgs诱导gc细胞的凋亡
以2.5、5、10μg/ml的acgs分别刺激ags、mkn-4536h和以5μg/ml的acgs分别刺激ags、mkn-4512、24、36h后,流式细胞检测细胞凋亡。从图5可知acgs刺激ags、mkn-45后使凋亡细胞比例增加。实验结果是acgs可以导致ags、mkn-45细胞凋亡。
5)acgs对gc细胞周期影响
以2.5、5、10μg/mlacgs刺激gc细胞36h后,利用流式细胞技术检测细胞周期。从图6可知,细胞周期阻滞在g0-g1期,从而阻止细胞有丝分裂影响细胞增殖。
6)acgs上调notch2的表达
以2.5、5、10μg/mlacgs刺激gc细胞36h后,用westernblot检测notch2蛋白的表达水平,从图7可知,acgs作用后notch2蛋白表达升高,且随着acgs浓度升高而升高。
7)acgs作用于gc细胞对下游蛋白hes1的影响
通过westernblot结果发现(图8)acgs刺激ags、mkn-45后对notch2下游蛋白hes1的表达与notch2的表达趋势并不十分一致,acgs刺激ags和mkn-45后(图7、图8)其notch2表达升高,而notch2下游蛋白hes1表达降低。这种偏差可能由于notch基因的表达是一个复杂的细胞调控过程。
8)过表达n2icd促进acgs对gc细胞生长的抑制作用
westernblot结果(图9)可知,与空载体转染对照组相比,转染n2icd/pcmv-tag4后的gc细胞notch2的表达升高,并且转染n2icd/pcmv-tag4组与同时加acgs组相比,后者notch2的表达比前者高。mts实验表明,与空载体转染对照组相比,转染n2icd/pcmv-tag4后的gc细胞生长受到抑制,并且转染n2icd/pcmv-tag4组与同时加acgs组相比,后者gc细胞的生长比前者受到的抑制作用强。结果表明,过表达notch2和acgs联合应用对gc细胞的生长抑制作用更明显。
9)notch2-sirna下调n2icd降低acgs对gc细胞的生长抑制作用
westernblot结果(图10)可知,与对照组相比,转染notch2-sirna后的gc细胞notch2的表达有明显降低,并且转染notch2-sirna组与同时加acgs组相比,后者notch2的表达比前者高。mts实验表明,与对照组相比,转染notch2-sirna后的gc细胞增殖能力升高,并且转染notch2-sirna组与同时加acgs组相比,后者gc细胞的增殖比前者受到的抑制作用强。结果表明,notch2-sirna干扰抑制了acgs诱导的对gc细胞的生长抑制作用。
在本发明中,与正常胃粘膜细胞相比,notch2在ags、sgc-7901中表达高,在mgc-803、mkn-28、mkn-45中表达低,我们选择ags和mkn-45做后续实验。我们首次证明了acgs能够上调gc细胞中notch2且存在剂量依赖性,还能够诱导细胞凋亡、阻滞细胞周期在g0-g1期,说明notch2在gc细胞中是抑癌基因,并且过表达notch2和notch2-sirna干扰实验也证明了这一点。与单独应用acgs相比,过表达notch2与acgs联合应用更能抑制gc细胞的增殖。相反,notch2-sirna干扰与acgs联合应用则降低了acgs诱导的细胞毒性作用。综上所述,acgs诱导notch2上调与gc细胞生长抑制和凋亡存在一定联系。然而,在本实验中发现hes1与notch2没有关联性,这正表明基因表达的调节是个复杂的过程。
总之,在体外实验中,acgs抑制gc细胞的增殖,诱导凋亡、阻滞细胞周期在g0-g1期和上调notch2的表达,且具有剂量依赖性。因此,acgs可能是通过notch2直接或间接发挥作用。据目前所知,本发明是第一个研究acgs通过notch2信号通路发挥抑制gc细胞的增殖作用。正如结果所示,acgs通过上调notch2发挥抑癌作用,抑制gc细胞的增殖、促进细胞凋亡和阻滞细胞周期具有剂量依赖性。另外,联合应用acgs和过表达notch2会产生协同作用,会比单独高剂量acgs用药效果好,这对减轻药物副作用和研制新的治疗gc的方案提供了可能性。这为研制新的治疗gc方法提供了理论依据和实验基础。
以上所述实施例的各技术特征可以进行任意的组合,为使描述简洁,未对上述实施例中的各个技术特征所有可能的组合都进行描述,然而,只要这些技术特征的组合不存在矛盾,都应当认为是本说明书记载的范围。
以上所述实施例仅表达了本发明的几种实施方式,其描述较为具体和详细,但并不能因此而理解为对发明专利范围的限制。应当指出的是,对于本领域的普通技术人员来说,在不脱离本发明构思的前提下,还可以做出若干变形和改进,这些都属于本发明的保护范围。因此,本发明专利的保护范围应以所附权利要求为准。
sequencelisting
<110>南方医科大学、珠海南医大生物医药公共服务平台有限公司
<120>acgs的新应用
<160>6
<170>patentinversion3.3
<210>1
<211>21
<212>dna
<213>人工序列
<400>1
aaaaatggggccaaccgagac21
<210>2
<211>20
<212>dna
<213>人工序列
<400>2
ttcatccagaaggcgcacaa20
<210>3
<211>19
<212>dna
<213>人工序列
<400>3
tggcacccagcacaatgaa19
<210>4
<211>25
<212>dna
<213>人工序列
<400>4
ctaagtcatagtccgcctagaagca25
<210>5
<211>31
<212>dna
<213>人工序列
<400>5
atttgcggccgcgccatgcgaaagcgtaagc31
<210>6
<211>33
<212>dna
<213>人工序列
<400>6
ccgctcgagcgcataaacctgcatgttgttgtg33
- 还没有人留言评论。精彩留言会获得点赞!