用于检测靶RNA的方法和组合物与流程
本申请要求2016年6月16日提交的美国临时专利申请号62/351,172、2016年8月22日提交的美国临时专利申请号62/378,156和2017年3月23日提交的美国专利申请号15/467,922的权益,这些申请以引用方式整体并入本文。
关于联邦政府资助的研究或开发的声明
本发明是在国家科学基金会(nationalsciencefoundation)授予的1244557号在政府支持下完成的。政府对本发明享有一定权利。通过引用并入作为文本文件提供的序列表
序列表在此提供为在2017年3月21日创建且大小为67kb的文本文件“berk-337wo_seqlist_st25.txt”。该文本文件的内容以引用方式整体并入本文。
导言
细菌适应性免疫系统采用crispr(成簇规律间隔短回文重复序列)和crispr相关(cas)蛋白进行rna指导的核酸切割。虽然iii型和vi型crispr系统通常靶向dna底物,但是指导针对单链rna(ssrna)底物的干扰复合物。在vi型crispr系统中,单亚基c2c2蛋白作为rna指导的rna内切核酸酶起作用。
crispr-cas系统通过rna指导的核酸干扰而赋予细菌和古细菌中的适应性免疫。在多种crispr类型中,仅相对罕见的vi型crispr系统被认为仅靶向单链rna底物,这是由大效应蛋白c2c2赋予的活性。vi型操纵子共享其它crispr-cas基因组基因座的共同特征,包括充当短病毒dna区段的储库的crispr序列阵列。为了提供抗病毒免疫,将经处理的crispr阵列转录物(crrna)与含cas蛋白的监督复合物组装,所述监督复合物识别与crrna的病毒来源区段(称为间隔区)具有序列互补性的核酸。
免疫监督的第一步需要将由侧接病毒间隔区序列的重复序列组成的前体crrna(pre-crrna)加工成单独功能性crrna,每个所述crrna含有单个病毒来源的序列片段。crispr系统采用多种机制产生成熟的crrna,包括使用专用的内切核酸酶(例如,i型和iii型系统中的cas6或cas5d)、宿主内切核酸酶(例如,rna酶iii)与反式激活crrna(tracrrna,ii型系统)的偶联、或效应器酶(例如,来自v型系统的cpf1)本身内源的核糖核酸酶活性。
技术实现要素:
本公开提供了用于检测单链靶rna的方法。本公开提供了将前体c2c2指导rna阵列切割成两个或更多个c2c2指导rna的方法。本公开提供了用于检测样品中的靶rna的试剂盒。
提供了用于检测单链靶rna的组合物和方法,其中所述方法包括(i)使具有多种rna的样品与(a)c2c2指导rna和(b)c2c2蛋白接触,所述c2c2指导rna与单链靶rna杂交,所述c2c2蛋白切割样品rna;以及(ii)测量由切割产生的可检测信号。一旦主题c2c2蛋白由c2c2指导rna激活,c2c2蛋白就变成切割样品rna的内切核糖核酸酶,所述主题c2c2蛋白的激活是当样品包括指导rna杂交的单链靶rna(即,样品包括所靶向的单链靶rna)时发生的。因此,当所靶向的单链靶rna存在于样品中(例如,在一些情况下高于某一阈值量)时,结果是样品中rna的切割,该切割可以使用任何合宜的检测方法检测(例如,使用经标记的检测rna)。
在一些情况下,可以通过使用前体c2c2指导rna阵列来提供两个或更多个c2c2指导rna,所述前体c2c2指导rna阵列可以由c2c2蛋白切割成单独的指导rna,并且这与c2c2蛋白是否具有完整的hepn1结构域和/或hepn2结构域无关。因此,还提供了将前体c2c2指导rna阵列切割成两个或更多个c2c2指导rna的方法。在一些情况下,c2c2蛋白缺乏催化活性的hepn1结构域和/或缺乏催化活性的hepn2结构域。
附图说明
图1a至图1e示出了与内源c2c2基因座相关的示意图和证明重组c2c2蛋白的异源表达和纯化的实验。
图2描绘了使用c2c2蛋白执行的切割测定的结果。
图3描绘了显示c2c2强烈切割单链rna的结果。
图4a至图4b示出了使用c2c2蛋白的各种切割测定的图和结果。
图5描绘了使用经标记的检测rna(使用猝灭剂/荧光剂对)执行的检测由c2c2蛋白进行的rna切割的实验的结果。
图6描绘了证明由c2c2蛋白进行pre-crrna加工的实验的结果。
图7a至图7b描绘了示例性c2c2指导rna的示意图和序列。
图8a至图8f提供了各种c2c2多肽的氨基酸序列。
图9a至图9c描绘了对前体crrna转录物进行c2c2家族加工以产生成熟的crrna。
图10a至图10c描绘了crispr重复序列的结构和序列对lbuc2c2介导的crrna生物发生的影响。
图11a至图11c描绘了lbuc2c2对顺式和反式靶的指导依赖性ssrna降解。
图12a至图12d描绘了lbuc2c2的两种不同的核糖核酸酶活性。
图13a至图13d描绘了c2c2介导的对复杂混合物中转录物的灵敏目视检测。
图14提供了表2:在实施例8-12中描述且在图12a至图12d中示出的研究中使用的各种dna底物。
图15提供了表3:实施例8-12中描述的研究中使用的各种rna底物。
图16a至图16f描绘了数据,所述数据显示由c2c2进行的pre-crrna加工是不依赖于间隔区序列的,可以在串联crrna阵列上发生,受pre-crna的5'和/或3'侧翼区域中的突变影响,并且是不依赖于金属的。
图17提供了pre-crrna双突变对pre-crrna加工活性的影响的总结。
图18a至图18b描绘了lbuc2c2ssrna靶切割位点映射。
图19a至图19c描绘了crrna间隔区长度、反应温度和crrna的5'末端序列对靶rna切割效率的依赖性。
图20a至图20c描绘了lbuc2c2与成熟crrna和靶ssrna的结合数据。
图21描绘了rna酶检测测定λ2-ssrna的时间进程。
图22a至图22b描绘了c2c2家族和c2c2比对的系统发生树。
图23a至图23d描绘了c2c2的纯化和生产。
图24a至图24c描绘了数据,所述数据显示c2c2蛋白加工前体crrna转录物以产生成熟crrna。a.c2c2蛋白的最大似然系统发生树。本研究中使用的同系物以黄色突出显示。b.本研究中使用的三种vi型crispr基因座的图。黑色矩形表示重复元件,黄色菱形表示间隔区序列。cas1和cas2仅存在于lshc2c2附近的基因组中。c.c2c2介导的对来源于lbuc2c2、lsec2c2和lshc2c2crispr重复序列基因座的pre-crrna的切割。oh:碱水解梯带;t1:rna酶t1水解梯带;用100nm的c2c2和小于1nm的pre-crrna执行加工切割反应。右侧描绘了切割的示意图,并且下方示出了所预测的pre-crrna二级结构,其中箭头指示了所映射(mapped)的c2c2切割位点。
图25a至图25c描绘了数据,所述数据显示lbuc2c2介导的crrna生物发生取决于crispr重复序列的结构和序列。a.由lbuc2c2进行的对含有发夹的茎区和环区内的结构突变的pre-crrna的代表性切割测定。下面列出的加工百分比是在60分钟处定量的(平均值±标准差,n=3)。b.条形图显示pre-crrna加工对crispr重复序列的依赖性。野生型重复序列如下方所示,其中单独的条代表以红色标示的串联核苷酸突变。切割位点用卡通剪刀指示。在60分钟后测量加工百分比(平均值±标准差,n=3)。所描绘的测试突变体的发夹可以在扩展数据图3至4中找到。c.通过加入10-50mm的edta和egta以实现标准反应条件来测试crrna加工反应的二价金属离子依赖性。
图26a至图26d描绘了数据,所述数据显示lbuc2c2含有两种不同的核糖核酸酶活性。a.在37℃下由lbuc2c2执行的顺式ssrna靶(黑色)和pre-crrna(青色)切割的定量时间过程数据。指数拟合示出为实线(n=3),并且所计算的伪一级速率常数(pseudo-first-orderrateconstant)(kobs)(平均值±标准差)对于顺式ssrna靶切割和pre-crrna切割分别为9.74±1.15分钟-1和0.12±0.02分钟-1。b.lbuc2c2架构描绘了hepn基序和加工缺陷型点突变体的位置。c、d.lbuc2c2突变体针对c中的pre-crrna加工和d中的ssrna靶向的核糖核酸酶活性,并且扩展数据为图6d。
图27a至图27e示出,c2c2提供了对复杂混合物中转录物的灵敏检测。a.使用猝灭的荧光rna报告物进行的lbuc2c2rna检测方法的示意图。b.在存在人总rna的情况下下,由lbuc2c2针对不同浓度的靶rna在30分钟后产生的荧光信号的定量。rna酶a示出为阳性rna降解对照。(平均值±标准差,n=3)。c.对于不同量的人总rna或细菌总rna,由负载有β-肌动蛋白靶向crrna的lbuc2c2在3h后产生的荧光信号的定量(作为β-肌动蛋白空阴性对照)。(平均值±标准差,n=3)。d.串联pre-crrna加工也使得能够进行rna检测。(平均值±标准差,n=3)。e.vi型crispr途径的模型突出显示了c2c2的核糖核酸酶活性。
图28a至图28b描绘了c2c2家族和c2c2比对的完整系统发生树。a.c2c2蛋白的最大似然系统发生重建。叶包括gi蛋白数和来源生物;100个重新取样中的自展支持值被呈现用于内部分割。标尺是每个位点的取代。b.所分析的三种c2c2同系物的多重序列比对;坐标基于lbuc2c2。
图29a至图29d描绘了与c2c2的纯化和生产有关的数据。所有c2c2同系物在大肠杆菌(e.coli)中表达为his-mbp融合物,并通过亲和性、离子交换和尺寸排阻色谱的组合进行纯化。通过与tev蛋白酶一起孵育去除ni+亲和标签。色谱级分的代表性sds-page凝胶示出在(a、b)中。c.来自superdex200(16/60)柱的色谱图,表明c2c2作为单峰洗脱,缺乏核酸。d.对本稿图中使用的纯化蛋白的sdspage分析。
图30a至图30i描绘了在体外和体内通过c2c2进行的pre-crrna加工的映射。a.单个同源pre-crrna阵列的lsec2c2和lshcc2c2切割的切割位点映射。oh:碱水解梯带;t1:t1rna酶水解梯带。用100nm的c2c2和小于1nm的pre-crrna进行切割反应。b-i.对来自shmakov等人10的lshc2c2(b-f)和lsec2c2(g-i)crispr阵列rna测序实验的再分析(分别为图s7和图5)。使用bowtie2(参见方法)将所有读段(b、g)和经过滤的读段(55nt或更少;根据原始shmakov等人的分析;c、h)与每个crispr阵列进行严格比对。示出了lsh(d-f)和lse(i)的单独crispr重复序列间隔区的详细视图。5'末端pre-crrna加工的差异由每个序列下方的箭头指示。可以使用分析的bam比对文件。该映射清楚地表明,从lshpre-crrna产生的小rna测序读段的5'末端映射到距所预测的发夹的碱基2nt的位置,这与体外加工数据(a)一致。这种模式适用于从在沙氏纤毛菌(l.shahii)中的天然表达和在大肠杆菌中的异源表达两者检测到的所有成熟crrna。不幸的是,lsec2c2crrna测序数据(用于g-i中)由于较低的读段深度而具有较少信息,并且每个比对的crrna表现出略微不同的5'末端,几乎没有明显的均一性。经加工的重复序列中的一个重复序列(重复序列间隔区2;i)的映射与数据一致,但由于读段深度不足而仅具有低置信度。
图31a至图31d描绘了由c2c2进行的pre-crrna加工是与间隔区序列无关的,可以在串联crrna阵列上发生,受pre-crna的5'侧翼区域中的突变影响并且产生3'磷酸酯产物。a.串联pre-crrna阵列的lbucc2c2切割的切割位点映射。oh:碱水解梯带;t1:t1rna酶水解梯带。用100nm的lbuc2c2和小于1nm的pre-crrna进行切割反应。右侧描绘了切割产物的示意图,其中箭头指示了映射的c2c2切割产物。b.lbuc2c24聚突变体的pre-crrna加工数据,表明5'单链侧翼区域对于有效的pre-crrna加工的重要性。在60分钟后测量pre-crrna加工的百分比(平均值±标准差,n=3)。c.代表性的lbuc2c2pre-crrna切割时间过程,表明相似的pre-crrna加工速率与crrna间隔区序列无关地发生,间隔区a和间隔区λ2的伪一级速率常数(kobs)(平均值±标准差)分别为0.07±0.04分钟-1和0.08±0.04分钟-1。d.通过t4多核苷酸激酶(pnk)处理对经切割的rna进行的端基分析。使用标准加工测定条件产生切割产物,然后将所述切割产物与pnk一起孵育1小时以去除任何2',3'-环状磷酸酯/3'-单磷酸酯。条带的延迟迁移指示从放射性标记的5'产物的3'末端去除了带电荷的单磷酸酯。
图32a至图32c示出lbuc2c2催化顺式和反式靶的指导依赖性ssrna降解。a.两种c2c2指导依赖性ssrna降解模式的示意图。b.由lbuc2c2切割两种不同的放射性标记的ssrna底物a和b。在37℃预形成100nm的c2c2与50nm的crrna的复合物,并在25℃下加入小于1nm的5'-标记的靶rna时开始反应。反式切割反应包含等摩尔(小于1nm)浓度的放射性标记的非指导互补底物和未标记的在靶ssrna。对于多种ssrna底物,观察到lbuc2c2仅在与互补crrna结合时催化有效切割,表明lbuc2c2:crrna以rna指导的方式切割ssrna。此活性在下文中称为在靶切割或顺式靶切割。lbuc2c2介导的顺式切割导致多种产物的梯化,其中切割优先在尿嘧啶残基之前发生,类似于lshc2c29。在存在未标记的在靶(与crrna互补的)ssrna的情况下重复非靶切割反应。与顺式执行的非靶切割实验不同,观察到了非靶rna的反式快速降解。对于顺式在靶切割和反式非靶切割观察到相似的rna切割速率和几乎相同的切割产物暗示两种活性中的相同核酸酶中心。c.在存在25mmedta的情况下,测试负载有crrna靶向间隔区a的lbuc2c2在顺式(靶a经标记)和反式(在存在未标记的靶a的情况下,靶b经标记)切割条件下的切割活性。
图33a至图33b示出了lbuc2c2ssrna靶切割位点映射。a.按照方法进行的ssrna靶切割测定,证明lbuc2c2介导对具有相同的间隔区互补序列但不同的5'侧翼序列(具有不同长度和核苷酸组成)的若干个放射性标记的ssrna底物的‘顺式’切割。ssrna底物的序列示出在右侧,其中crrna-a的间隔区互补序列以黄色突出显示。箭头指示检测到的切割位点。为了清楚起见而裁剪了凝胶。应当注意的是,在不同底物上产生的切割产物的模式(例如a.1对比a.2对比a.3)表明,切割位点选择主要由尿嘧啶偏好性驱动,并且表现为在crrna互补靶序列内明显缺乏排他性切割机制,这与关于其它ii类crispr单效应复合物(诸如cas9和cpf1)11,21观察到的相反。有趣的是,针对底物a.0观察到的切割模式提示了对polyg序列的次要偏好性。b.按照方法,使用一系列覆盖ssrna靶的长度的crrna进行的lbuc2c2ssrna靶切割测定。在该实验中使用的ssrna底物的序列示出在凝胶下方,其中每个crrna的间隔区互补序列以黄色突出显示。箭头指示所预测的切割位点。在每组泳道上方的小图指示间隔区序列沿着靶(黄色框)的位置以及观察到(红色箭头)或不存在(黑色箭头)切割产物。同样地,应当注意的是,对于每种crrna,切割产物长度分布是非常相似的,再次表明在crrna结合序列内明显缺乏排他性切割。在反应的子集中不存在几种切割产物可能是由于ssrna靶上存在结合的c2c2:crrna,这可以通过任何顺式(分子内)或反式(分子间)lbuc2c2活性位点在空间上阻断对尿嘧啶的接取。虽然对lbuc2c2的原型间隔区侧翼位点(pfs)偏好性的适当分析超出了本研究的范围,但观察到了3'侧翼核苷酸的最小影响。预期的pfs碱基在所测试的每个指导序列旁边的图表中以红色标示。
图34a至图34d描绘了rna靶向对crrna变异、温度和点突变的依赖性。a.按照方法进行的lbuc2c2ssrna靶切割测定,其中crrna具有16nt、20nt或24nt的间隔区。b.按照方法在25℃和37℃处进行的lbuc2c2ssrna靶切割时间过程。c.按照方法进行的lbuc2c2ssrna靶切割时间过程,其中crrna具有不同的5'端侧翼核苷酸突变。突变以红色突出显示。1-2个核苷酸的5'延伸可忽略不计地影响切割效率。相比之下,将侧翼区域缩短至3nt会减慢切割速率。d.点突变对c2c2在用于ssrna靶向的hepn基序内的保守残基突变体中的核糖核酸酶活性的影响。
图35a至图35d描绘了lbuc2c2与成熟crrna和靶ssrna的结合数据。a.如在方法中所描述地进行过滤结合测定以确定成熟crrna-a_gg与lbuc2c2-wt、lbuc2c2-dhepn1、lbuc2c2-dhepn2或lbuc2c2-dhepn1/dhepn2的结合亲和力。将定量的数据与标准结合等温线拟合。误差条代表三次独立实验的标准偏差。从三个独立实验测量的解离常数(平均值±标准偏差)为27.1±7.5nm(lbuc2c2-wt)、15.2±3.2nm(lbuc2c2-dhepn1)、11.5±2.5nm(lbuc2c2-dhepn2)和43.3±11.5nm(lbuc2c2-dhepn1/dhepn2)。b.如所示,关于lbuc2c2-dhepn1/dhepn2:crrna-a_gg与‘在靶’assrna或‘脱靶’bssrna之间的结合反应的代表性电泳迁移率变动测定。如方法中所述进行三次独立实验。为了清楚起见而裁剪了凝胶。c.将来自(b)的定量结合数据与标准结合同种型拟合。误差条代表三次独立实验的标准偏差。从三个独立实验测量的解离常数(平均值±标准偏差)对于ssrnaa为1.62±0.43nm并且对于ssrnab为未检测到的(>>10nm)。d.如在方法中所描述地进行过滤结合测定以确定成熟crrna-a_ga与lbuc2c2-wt和lbuc2c2-r1079a的结合亲和力。将定量的数据与标准结合等温线拟合。误差条代表三次独立实验的标准偏差。从三个独立实验测量的解离常数(平均值±标准偏差)为4.65±0.6nm(lbuc2c2-wt)和2.52±0.5nm(lbuc2c2-r1079a)。应当注意的是,这些结合亲和力与图a不同。这种差异被解释为在指导序列的5'端序列方面略有不同,其中图a中的指导序列以5`-ggcca……开始并且图d中的指导序列以5`-gacca开始。虽然天然指导序列(5`-gacca)与lbuc2c2更紧密地结合,但是在这些指导序列变体的rna靶向效率方面没有看到差异(扩展数据图6c)。
图36a至图36b描绘了λ2-ssrna的rna酶检测测定的时间过程。a.在存在不断增加量的λ2ssrna(0-1nm)的情况下,将lbuc2c2:crrna-λ2与rna酶-alert底物(thermo-fisher))和100nghela总rna在37℃孵育120分钟。每5分钟进行荧光测量。1nm的λ2ssrna反应在可以测量第一时间点之前达到饱和。误差条代表三次独立实验的标准偏差。b.在存在或不存在在靶激活λ4ssrna的情况下,将lbuc2c2:crrna-λ4或apolbuc2c2在hela总rna中孵育2小时。按照方法在bioanalyzer2100中的小rna芯片上解析背景小rna的降解。在apolbuc2c2和lbuc2c2:crrna-λ4之间的片段谱中观察到了小差异。相比之下,在向反应中加入在靶ssrna时,trna峰的急剧变宽和移位揭示了在激活lbuc2c2反式活性时反应中存在的其它结构化和非结构化rna的广泛降解。
图37描绘了切割实验,所述切割实验证明当蛋白在选自r1079(例如,r1079a)、r1072(例如,r1072a)和k1082(例如,k1082a)的任何氨基酸位置处包括突变时,lbuc2c2对前体指导rna的切割(指导rna加工)严重减少。
图38a至图38c描绘了cas13a家族内pre-crrna加工的保守性。
图39a至图39c描绘了cas13a同系物的crispr基因座和crrna重复序列架构。
图40a至图40f描绘了对由lbucas13a进行的pre-crrna切割重要的残基。
图41a至图41b描绘了cas13a家族成员的螺旋1结构域和hepn结构域的比对。
图42a至图42d描绘了cas13a家族成员对ssrna的效率。
图43a至图43d描绘了由cas13a同系物进行的反式ssrna切割。
图44a至图44f描绘了cas13a家族内的crrna可交换性。
图45a至图45c描绘了用于rna检测的正交cas13a亚家族的功能验证。
图46a至图46d描绘了野生型(wt)lbucas13a和lbucas13ar1079a/k1080a双重突变体进行的crrna阵列加工。
图47a至图47c描绘了在pre-crrna加工所涉及的区域中由lbucas13a点突变体进行的反式切割。
图48a至图48b描绘了lbucas13ar1079a/k1080a双重突变体相对于野生型lbucas13a的特征。
图49提供表4。
图50提供表5。
图51提供表6。
图52提供表7。
图53提供表8。
图54提供表9。
图55a至图55b呈现vi-a型crispr系统功能的模型。
图56a至图56k提供各种cas13a多肽的氨基酸序列。
图57提供各种cas13a多肽的氨基酸序列的比对。
定义
本文可互换使用的术语“多核苷酸”和“核酸”是指任何长度的核苷酸的聚合形式,为核糖核苷酸或脱氧核糖核苷酸。因此,术语“多核苷酸”和“核酸”涵盖单链dna;双链dna;多链dna;单链rna;双链rna;多链rna;基因组dna;cdna;dna-rna杂交体;以及这样的聚合物,所述聚合物包含嘌呤碱基和嘧啶碱基或其它天然、经化学或生物化学修饰的、非天然的或衍生的核苷酸碱基。
术语“寡核苷酸”是指3与100个核苷酸之间的单链或双链核酸(例如,dna、rna或经修饰的核酸)的多核苷酸。然而,出于本公开的目的,寡核苷酸的长度没有上限。寡核苷酸也称为“寡聚体”或“寡聚物”,并且可以从基因中分离、(体外和/或体内地)转录,或化学合成。术语“多核苷酸”和“核酸”应理解为包括适用于所描述的实施方案的单链(诸如有义或反义)和双链多核苷酸。
“可杂交的”或“互补的”或“基本上互补的”是指核酸(例如rna、dna)包含这样的核苷酸序列,所述核苷酸序列使所述核酸能够在适当的体外和/或体内温度和溶液离子强度条件下,与另一核酸以序列特异性、反向平行方式(即,核酸特异性结合互补核酸)非共价结合(即,形成watson-crick碱基对和/或g/u碱基对)、“退火”或“杂交”。标准watson-crick碱基配对包括:腺嘌呤/腺苷)(a)与胸腺嘧啶/胸苷(t)配对,a与尿嘧啶/尿苷(u)配对,以及鸟嘌呤/鸟苷(g)与胞嘧啶/胞苷(c)配对。此外,对于两个rna分子(例如,dsrna)之间的杂交,以及对于dna分子与rna分子的杂交(例如,当dna靶核酸碱基与c2c2指导rna配对时,等):g也可以与u碱基配对。例如,在trna反密码子与mrna中的密码子进行碱基配对的情境下,g/u碱基配对部分地负责遗传密码的简并性(即,冗余)。因此,在本公开的上下文中,g(例如,c2c2指导rna分子的蛋白结合区段(dsrna双链体)的g;与c2c2指导rna碱基配对的靶核酸的g)被认为是与u和c两者互补。例如,当可以在c2c2指导rna分子的蛋白结合区段(例如,dsrna双链体)的给定核苷酸位置处制备g/u碱基对时,该位置不被认为是不互补的,而是替代地被认为是互补的。
杂交和洗涤条件是众所周知的,并在sambrook,j.,fritsch,e.f.和maniatis,t.molecularcloning:alaboratorymanual,第二版,coldspringharborlaboratorypress,coldspringharbor(1989)中,特别是该参考文献中的第11章和表11.1中;和sambrook,j.和russell,w.,molecularcloning:alaboratorymanual,第三版,coldspringharborlaboratorypress,coldspringharbor(2001)中例示。温度和离子强度的条件决定了杂交的“严谨性”。
杂交需要两个核酸含有互补序列,但是碱基之间的错配是可能的。适合于两个核酸之间杂交的条件取决于核酸的长度和互补程度,核酸的长度和互补程度是本领域中众所周知的变量。两个核苷酸序列之间的互补程度越大,则具有这些序列的核酸的杂交体的解链温度(tm)值越大。对于具有短链互补性(例如35个或更少个、30个或更少个、25个或更少个、22个或更少个、20个或更少个或18个或更少个核苷酸的互补性)的核酸之间的杂交,错配位置可能变得重要(参见sambrook等人,出处同上,11.7-11.8)。通常,可杂交核酸的长度为8个核苷酸或更多个(例如,10个核苷酸或更多个、12个核苷酸或更多个、15个核苷酸或更多个、20个核苷酸或更多个、22个核苷酸或更多个、25个核苷酸或更多个或30个核苷酸或更多个)。可以根据诸如互补区域的长度和互补程度之类的因素,而根据需要调节温度和洗涤溶液盐浓度。
应当理解,多核苷酸的序列不需要与其可特异性杂交或可杂交的靶核酸的序列100%互补。此外,多核苷酸可以在一个或多个区段上杂交,使得插入或相邻区段不参与杂交事件(例如,环结构或发夹结构)。多核苷酸可包含与其将杂交的靶核酸序列内的靶区域60%或更多、65%或更多、70%或更多、75%或更多、80%或更多、85%或更多、90%或更多、95%或更多、98%或更多、99%或更多、99.5%或更多或100%的序列互补性。例如,这样的反义核酸将代表百分之90的互补性,在所述反义核酸中,反义化合物的20个核苷酸中的18个与靶区域互补并因此与靶区域特异性杂交。在该实例中,剩余的非互补核苷酸可以与互补核苷酸成簇或散布,并且不需要彼此邻接或与互补核苷酸邻接。可以使用任何合宜的方法确定核酸内的特定核酸序列段之间的互补百分比。示例性方法包括blast程序(基本局部比对搜索工具)和powerblast程序(altschul等人,j.mol.biol.,1990,215,403-410;zhang和madden,genomeres.,1997,7,649-656),或通过使用使用默认设置的gap程序(wisconsin序列分析包,用于unix的第8版,geneticscomputergroup,universityresearchpark,madisonwis.),所述程序使用smith和waterman算法(adv.appl.math.,1981,2,482-489)。
术语“肽”、“多肽”和“蛋白”在本文中可互换使用,并且是指任何长度的氨基酸聚合形式,其可包括编码的和非编码的氨基酸、经化学修饰或生物化学修饰的或衍生化的氨基酸,以及具有经修饰的肽骨架的多肽。
如本文所用的“结合”(例如,参照多肽的rna结合结构域、与靶核酸结合等)是指大分子之间的非共价相互作用(例如,蛋白与核酸之间的非共价相互作用;在c2c2指导rna复合物和靶核酸之间的非共价相互作用;等等)。当处于非共价相互作用状态时,大分子被称为“缔合的”或“相互作用”或“结合”(例如,当分子x被称为与分子y相互作用时,则意味着分子x以非共价方式与分子y结合)。并非结合相互作用的所有组分都需要是序列特异性的(例如,与dna骨架中的磷酸残基接触),但结合相互作用的一些部分可以是序列特异性的。结合相互作用的特征通常在于解离常数(kd)小于10-6m、小于10-7m、小于10-8m、小于10-9m、小于10-10m、小于10-11m、小于10-12m、小于10-13m、小于10-14m,或小于10-15m。“亲和力”是指结合强度,增加的结合亲和力与较低的kd相关。
“结合结构域”是指能够与另一分子非共价结合的蛋白结构域。结合结构域可以与例如rna分子(rna结合结构域)和/或蛋白分子(蛋白结合结构域)结合。在具有蛋白结合结构域的蛋白的情况下,所述蛋白在一些情况下可以与其自身结合(以形成同源二聚体、同源三聚体等)和/或所述蛋白可以与一个或多个不同蛋白的一个或多个区域结合。
术语“保守氨基酸取代”是指蛋白中具有相似侧链的氨基酸残基的可互换性。例如,一组具有脂肪族侧链的氨基酸由甘氨酸、丙氨酸、缬氨酸、亮氨酸和异亮氨酸组成;一组具有脂肪族羟基侧链的氨基酸由丝氨酸和苏氨酸组成;一组具有含酰胺侧链的氨基酸由天冬酰胺和谷氨酰胺组成;一组具有芳香族侧链的氨基酸由苯丙氨酸、酪氨酸和色氨酸组成;一组具有碱性侧链的氨基酸由赖氨酸、精氨酸和组氨酸组成;一组具有酸性侧链的氨基酸由谷氨酸和天冬氨酸组成;并且一组具有含硫侧链的氨基酸由半胱氨酸和甲硫氨酸组成。示例性保守氨基酸取代基是:缬氨酸-亮氨酸-异亮氨酸、苯丙氨酸-酪氨酸、赖氨酸-精氨酸、丙氨酸-缬氨酸-甘氨酸,以及天冬酰胺-谷氨酰胺。
多核苷酸或多肽与另一多核苷酸或多肽具有一定百分比的“序列同一性”,意味着当比对时,在比较两个序列时,碱基或氨基酸的百分比是相同的,并且在相同的相对位置中。序列同一性可以以多种不同方式确定。为了确定序列同一性,可以使用各种方法和计算机程序(例如blast、t-coffee、muscle、mafft、phyre2等)比对序列,所述各种方法和计算机程序可通过万维网在包括ncbi.nlm.nili.gov/blast、ebi.ac.uk/tools/msa/tcoffee/、ebi.ac.uk/tools/msa/muscle/、mafft.cbrc.jp/alignment/software/、http://www.sbg.bio.ic.ac.uk/~phyre2/的网站处获得。参见例如altschul等人,(1990),j.mol.bioi.215:403-10。
“编码”特定rna的dna序列是转录成rna的dna核酸序列。dna多核苷酸可编码转化为蛋白的rna(mrna),或者dna多核苷酸可编码未翻译成蛋白的rna(例如trna、rrna、微rna(mirna)、“非编码”rna(ncrna)、c2c2指导rna等)。
本文可互换使用的术语“dna调控序列”、“控制元件”和“调控元件”是指转录和翻译控制序列,诸如启动子、增强子、多腺苷酸化信号、终止子、蛋白降解信号等,所述转录和翻译控制序列提供和/或调控非编码序列(例如,cas9指导rna)或编码序列(例如,cas9蛋白)的转录和/或调控编码多肽的翻译。
如本文所用,“启动子序列”是能够结合rna聚合酶并启动下游(3'方向)编码或非编码序列的转录的dna调控区。出于本公开的目的,启动子序列在其3'末端处由转录起始位点结合并向上游(5'方向)延伸,以包含以高于背景的可检测水平起始转录所必需的最少数量的碱基或元件。在启动子序列内将发现转录起始位点,以及负责结合rna聚合酶的蛋白结合结构域。真核启动子通常将(但不总是)包含“tata”盒和“cat”盒。包括诱导型启动子在内的各种启动子可用于驱动本公开的各种载体。
如本文所用的应用于核酸、多肽、细胞或生物体的术语“天然存在的”或“未修饰的”或“野生型”是指在自然中存在的核酸、多肽、细胞或生物体。例如,存在于生物体(包括病毒)中、可以从自然中的来源分离并且尚未由人在实验室中有意修饰的多肽或多核苷酸序列是野生型(并且是天然存在的)。
如本文所用的“重组”是指特定核酸(dna或rna)是克隆、限制、聚合酶链式反应(pcr)和/或连接步骤的各种组合的产物,导致具有结构编码或非编码序列的构建体可与自然系统中存在的内源核酸区分开来。编码多肽的dna序列可以由cdna片段或由一系列合成的寡核苷酸组装而成,以提供能够从包含在细胞中或无细胞转录和翻译系统中的重组转录单元表达的合成核酸。包含相关序列的基因组dna也可用于形成重组基因或转录单位。非翻译dna的序列可以存在于开放阅读框的5'端或3'端,其中此类序列不干扰编码区的操纵或表达,并且确实可以作用以通过各种机制调节所需产物的产生(参见下文的“dna调控序列”)。或者,编码未翻译的rna(例如,c2c2指导rna)的dna序列也可以被认为是重组的。因此,例如,术语“重组”核酸是指这样的核酸,所述核酸是非天然存在的,例如,是通过两个经由人工干预以其它方式分开的序列区段的人工组合而制备的。这种人工组合往往通过化学合成手段或通过人工操纵分离的核酸区段(例如通过基因工程技术)来实现。通常这样做是为了用编码相同氨基酸、保守氨基酸或非保守氨基酸的密码子替换密码子。或者,执行将所需功能的核酸区段连接在一起以产生所需的功能组合。这种人工组合往往通过化学合成手段或通过人工操纵分离的核酸区段(例如通过基因工程技术)来实现。当重组多核苷酸编码多肽时,所编码的多肽的序列可以是天然存在的(“野生型”)或可以是天然存在序列的变体(例如,突变体)。因此,术语“重组”多肽不一定是指序列不是天然存在的多肽。相反,“重组”多肽由重组dna序列编码,但是所述多肽的序列可以是天然存在的(“野生型”)或非天然存在的(例如,变体、突变体等)。因此,“重组”多肽是人工干预的结果,但可以是天然存在的氨基酸序列。
“载体”或“表达载体”是复制子,诸如质粒、噬菌体、病毒或粘粒,另一dna区段(即“插入物”)可以附接到所述复制子以便引起附接片段在细胞中的复制。
“表达盒”包含与启动子可操作连接的dna编码序列。“可操作地连接”是指并置,其中如此描述的组分处于允许它们以它们预期的方式起作用的关系中。例如,如果启动子影响编码序列的转录或表达,则所述启动子与所述编码序列可操作地连接。
术语“重组表达载体”或“dna构建体”在本文中可互换地使用,以指代包含载体和一个插入物的dna分子。通常产生重组表达载体以用于表达和/或增殖插入物的目的,或用于构建其它重组核苷酸序列的目的。插入物可以或可以不与启动子序列可操作地连接,并且可以或可以不与dna调控序列可操作地连接。
任何给定的部件或部件组合可以是未标记的,或者可以用标记部分可检测地标记。在一些情况下,当标记两种或更多种组分时,它们可以用彼此可区分的标记部分标记。
一般的分子和细胞生物化学方法可以在标准教科书中找到,所述标准教科书为诸如molecularcloning:alaboratorymanual,第3版(sambrook等人,harborlaboratorypress2001);shortprotocolsinmolecularbiology,第4版(ausubel等人编辑,johnwiley&sons1999);proteinmethods(bollag人,johnwiley&sons1996);nonviralvectorsforgenetherapy(wagner等人编辑,academicpress1999);viralvectors(kaplift和loewy编辑,academicpress1995);immunologymethodsmanual(i.lefkovits编辑,academicpress1997);以及cellandtissueculture:laboratoryproceduresinbiotechnology(doyle和griffiths,johnwiley&sons1998),这些参考文献的公开内容以引用方式并入本文中。
在进一步描述本发明之前,应当理解本发明不限于所描述的特定实施方案,因此当然可以改变。还应当理解,本文所使用的术语仅用于描述特定实施方案的目的,而非旨在为限制性的,因为本发明的范围将仅受所附权利要求书的限制。
在提供值范围的情况下,应理解,除非上下文另有明确规定,每个居间值至该下限单位的十分之一在该范围的上限和下限之间,并且任何其它所述范围或该所述范围中的居间值被涵盖在本发明内。这些较小范围的上限和下限可以独立地包括在该等较小范围内,并且也被涵盖在本发明内,经受所述范围中任何特别排除的极限。在所述范围包括上限和下限中的一个或两个的情况下,排除了这些所包括的极限中的一者或两者的范围也被包括在本发明中。
除非另有定义,本文所使用的所有技术和科学术语具有与本发明所属领域的普通技术人员通常理解的相同的含义。尽管与本文描述的那些方法和材料类似或等同的任何方法和材料也可用于本发明的实践或测试中,但是现在描述优选的方法和材料。本文提及的所有出版物均以引用方式并入本文,以公开和描述与所引用的出版物相关的方法和/或材料。
必须注意,如本文和随附权利要求书中所使用,单数形式“一”、“一个”和“该”包括复数指示物,除非上下文另有明确规定。因此,例如,提及“一蛋白”包括多个此类蛋白,并且提及“该指导rna”包括提及一个或多个此类指导rna及本领域的技术人员已知的其等同物,等等。还应当注意,权利要求书可以撰写成排除任何任选要素。因此,本声明旨在充当使用与权利要求要素的叙述相关的诸如“单独地”、“仅”等排他性术语或使用“否定”限制的先行基础。
应当理解,为了清楚起见而在单独实施方案的上下文中描述的本发明的某些特征也可以在单个实施方案中组合提供。相反,为简洁起见而在单个实施方案的上下文中描述的本发明的各种特征也可以单独地或以任何合适的子组合提供。与本发明有关的实施方案的所有组合由本发明具体包括,并且在本文中公开为如同每个和各个组合被单独和明确地公开一样。此外,各种实施例及其要素的所有子组合也由本发明具体包含,并且在本文中公开为如同每个此类子组合在此单独和明确地公开一样。
本文所论述的出版物仅仅是出于它们在本申请的申请日之前的公开内容而提供的。本文中的任何内容都不应被解释为承认本发明不能凭借优先权发明而在这些出版物之前授权。此外,所提供的出版物的日期可能与实际出版日期不同,这可能需要独立确认。
具体实施方式
本公开提供了用于检测单链靶rna的方法。本公开提供了将前体c2c2指导rna阵列切割成两个或更多个c2c2指导rna的方法。本公开提供了用于检测样品中的靶rna的试剂盒。术语“c2c2指导rna”在本文中与“cas13a指导rna”可互换地使用,并且在一些情况下指导rna被称为crrna(例如,“cas13acrrna”);术语“c2c2蛋白”(或“c2c2多肽”)在本文中与“cas13a蛋白”(或“cas13a多肽”)可互换地使用。
检测单链rna的方法
提供了用于检测单链靶rna的组合物和方法,其中所述方法包括(i)使具有多种rna的样品与(a)c2c2指导rna和(b)c2c2蛋白接触,所述c2c2指导rna与单链靶rna杂交,所述c2c2蛋白切割样品中存在的rna;以及(ii)测量由切割产生的可检测信号。一旦主题c2c2蛋白被c2c2指导rna激活,c2c2蛋白就被激活并作为非特异性地切割样品中存在的rna(包括非靶rna)的内切核糖核酸酶起作用,所述主题c2c2蛋白的激活是当样品包括指导rna杂交的单链靶rna(即,样品包含所靶向的单链靶rna)时发生的。因此,当所靶向的单链靶rna存在于样品中(例如,在一些情况下高于某一阈值量)时,结果是样品中rna(包括非靶rna)的切割,该切割可以使用任何合宜的检测方法检测(例如,使用经标记的检测rna)。接触步骤通常在包含二价金属离子的组合物中进行。接触步骤可以在无细胞环境中进行,例如在细胞外进行。接触步骤可以在细胞内进行。接触步骤可以在体外细胞中进行。接触步骤可以在离体细胞中进行。接触步骤可以在体内细胞中进行。在一些情况下,c2c2指导rna作为rna被提供;并且c2c2蛋白本身作为蛋白被提供。在一些情况下,c2c2指导rna作为编码指导rna的dna被提供;并且c2c2蛋白本身作为蛋白被提供。在一些情况下,c2c2指导rna作为rna被提供;并且c2c2蛋白作为编码c2c2蛋白的rna被提供。在一些情况下,c2c2指导rna作为编码指导rna的dna被提供;并且c2c2蛋白作为编码c2c2蛋白的rna被提供。在一些情况下,c2c2指导rna作为rna被提供;并且c2c2蛋白作为包含编码c2c2蛋白的核苷酸序列的dna被提供。在一些情况下,c2c2指导rna作为编码指导rna的dna被提供;并且c2c2蛋白作为包含编码c2c2蛋白的核苷酸序列的dna被提供。在一些情况下,本公开的方法提供了对样品中两种不同靶rna(第一单链靶rna和第二单链靶rna)的基本上同时检测。
在一些情况下,可以通过使用前体c2c2指导rna阵列来提供两个或更多个(例如,3个或更多个、4个或更多个、5个或更多个、或6个或更多个)c2c2指导rna,所述前体c2c2指导rna阵列可以由c2c2蛋白切割成单独的(“成熟”)指导rna;前体c2c2指导rna的切割与c2c2蛋白是否具有完整的hepn1结构域和/或hepn2结构域无关。因此,还提供了将前体c2c2指导rna阵列切割成两个或更多个c2c2指导rna的方法。因此,c2c2指导rna阵列可包括多于一个指导序列。在一些情况下,主题c2c2指导rna可以包括来自前体crrna的柄,但不一定必须包括多个指导序列。在一些情况下,c2c2蛋白缺乏催化活性的hepn1结构域和/或缺乏催化活性的hepn2结构域。接触步骤可以在无细胞环境中进行,例如在细胞外进行。接触步骤可以在细胞内进行。接触步骤可以在体外细胞中进行。接触步骤可以在离体细胞中进行。接触步骤可以在体内细胞中进行。
在一些情况下(例如,当与c2c2指导rna和c2c2蛋白接触时,当与前体c2c2指导rna阵列和c2c2蛋白接触时,等等),在测量步骤之前使样品接触2小时或更短时间(例如,1.5小时或更短时间、1小时或更短时间、40分钟或更短时间、30分钟或更短时间、20分钟或更短时间、10分钟或更短时间、或5分钟或更短时间、或1分钟或更短时间)。例如,在一些情况下,在测量步骤之前使样品接触40分钟或更短时间。在一些情况下,在测量步骤之前使样品接触20分钟或更短时间。在一些情况下,在测量步骤之前使样品接触10分钟或更短时间。在一些情况下,在测量步骤之前使样品接触5分钟或更短时间。在一些情况下,在测量步骤之前使样品接触1分钟或更短时间。在一些情况下,在测量步骤之前使样品接触50秒至60秒。在一些情况下,在测量步骤之前使样品接触40秒至50秒。在一些情况下,在测量步骤之前使样品接触30秒至40秒。在一些情况下,在测量步骤之前使样品接触20秒至30秒。在一些情况下,在测量步骤之前使样品接触10秒至20秒。
本公开提供了检测包含多种rna(例如,包含靶rna和多种非靶rna)的样品中的单链rna的方法。在一些情况下,所述方法包括:a)使样品与以下接触:(i)与单链靶rna杂交的c2c2指导rna,和(ii)切割样品中存在的rna的c2c2蛋白;以及b)测量由c2c2蛋白介导的rna切割产生的可检测信号。在一些情况下,所述方法包括:a)使样品与以下接触:i)前体c2c2指导rna阵列,所述前体c2c2指导rna阵列包含两个或更多个c2c2指导rna,每个所述c2c2指导rna具有不同的指导序列;和(ii)c2c2蛋白,所述c2c2蛋白将前体c2c2指导rna阵列切割成单独的c2c2指导rna,并且还切割样品的rna;以及b)测量由c2c2蛋白介导的rna切割产生的可检测信号。在一些情况下,本公开的方法提供了对样品中两种不同靶rna(第一单链靶rna和第二单链靶rna)的基本上同时检测。
用于检测包含多种rna(包括单链靶rna和多种非靶rna)的样品中的单链rna(单链靶rna)的本公开的方法可以以高灵敏度检测单链靶rna。在一些情况下,本公开的方法可用于检测包含多种rna(包括单链靶rna和多种非靶rna)的样品中存在的靶单链rna,其中靶单链rna以每107个非靶rna一个或多个拷贝(例如,每106个非靶rna一个或多个拷贝、每105个非靶rna一个或多个拷贝、每104个非靶rna一个或多个拷贝、每103个非靶rna一个或多个拷贝、每102个非靶rna一个或多个拷贝、每50个非靶rna一个或多个拷贝、每20个非靶rna一个或多个拷贝、每10个非靶rna一个或多个拷贝、或每5个非靶rna一个或多个拷贝)存在。
在一些情况下,本公开的方法可以检测在包含多种rna(包括单链靶rna和多种非靶rna)的样品中存在的靶单链rna,其中所述靶单链rna以每107个非靶rna一个拷贝至每10个非靶rna一个拷贝(例如,从每107个非靶rna1个拷贝至每102个非靶rna1个拷贝、从每107个非靶rna1个拷贝至每103个非靶rna1个拷贝、从每107个非靶rna1个拷贝至每104个非靶rna1个拷贝、从每107个非靶rna1个拷贝至每105个非靶rna1个拷贝、从每107个非靶rna1个拷贝至每106个非靶rna1个拷贝,从每106个非靶rna1个拷贝至每10个非靶rna1个拷贝、从每106个非靶rna1个拷贝至每102个非靶rna1个拷贝、从每106个非靶rna1个拷贝至每103个非靶rna1个拷贝、从每106个非靶rna1个拷贝至每104个非靶rna1个拷贝、从每106个非靶rna1个拷贝至每105个非靶rna1个拷贝、从每105个非靶rna1个拷贝至每10个非靶rna1个拷贝、从每105个非靶rna1个拷贝至每102个非靶rna1个拷贝、从每105个非靶rna1个拷贝至每103个非靶rna1个拷贝或从每105个非靶rna1个拷贝至每104个非靶rna1个拷贝)存在。
在一些情况下,本公开的方法可以检测在包含多种rna(包括单链靶rna和多种非靶rna)的样品中存在的靶单链rna,其中所述靶单链rna以每107个非靶rna一个拷贝至每100个非靶rna一个拷贝(例如,从每107个非靶rna1个拷贝至每102个非靶rna1个拷贝、从每107个非靶rna1个拷贝至每103个非靶rna1个拷贝、从每107个非靶rna1个拷贝至每104个非靶rna1个拷贝、从每107个非靶rna1个拷贝至每105个非靶rna1个拷贝、从每107个非靶rna1个拷贝至每106个非靶rna1个拷贝,从每106个非靶rna1个拷贝至每100个非靶rna1个拷贝、从每106个非靶rna1个拷贝至每102个非靶rna1个拷贝、从每106个非靶rna1个拷贝至每103个非靶rna1个拷贝、从每106个非靶rna1个拷贝至每104个非靶rna1个拷贝、从每106个非靶rna1个拷贝至每105个非靶rna1个拷贝、从每105个非靶rna1个拷贝至每100个非靶rna1个拷贝、从每105个非靶rna1个拷贝至每102个非靶rna1个拷贝、从每105个非靶rna1个拷贝至每103个非靶rna1个拷贝或从每105个非靶rna1个拷贝至每104个非靶rna1个拷贝)存在。
在一些情况下,对于检测样品中单链靶rna的本方法,检测阈值为10nm或更低。术语“检测阈值”在本文中用于描述为了进行检测样品中必须存在的靶rna的最小量。因此,作为说明性实例,当检测阈值为10nm时,则当靶rna以10nm或更高的浓度存在于样品中时,可以检测到信号。在一些情况下,本公开的方法具有5nm或更低的检测阈值。在一些情况下,本公开的方法具有1nm或更低的检测阈值。在一些情况下,本公开的方法具有0.5nm或更低的检测阈值。在一些情况下,本公开的方法具有0.1nm或更低的检测阈值。在一些情况下,本公开的方法具有0.05nm或更低的检测阈值。在一些情况下,本公开的方法具有0.01nm或更低的检测阈值。在一些情况下,本公开的方法具有0.005nm或更低的检测阈值。在一些情况下,本公开的方法具有0.001nm或更低的检测阈值。在一些情况下,本公开的方法具有0.0005nm或更低的检测阈值。在一些情况下,本公开的方法具有0.0001nm或更低的检测阈值。在一些情况下,本公开的方法具有0.00005nm或更低的检测阈值。在一些情况下,本公开的方法具有0.00001nm或更低的检测阈值。在一些情况下,本公开的方法具有10pm或更低的检测阈值。在一些情况下,本公开的方法具有1pm或更低的检测阈值。在一些情况下,本公开的方法具有500fm或更低的检测阈值。在一些情况下,本公开的方法具有250fm或更低的检测阈值。在一些情况下,本公开的方法具有100fm或更低的检测阈值。在一些情况下,本公开的方法具有50fm或更低的检测阈值。
在一些情况下,检测阈值(在本方法中用于检测单链靶rna)在500fm至1nm(例如,500fm至500pm、500fm至200pm、500fm至100pm、500fm至10pm、500fm至1pm、800fm至1nm、800fm至500pm、800fm至200pm、800fm至100pm、800fm至10pm、800fm至1pm、1pm至1nm、1pm至500pm、1pm至200pm、1pm至100pm或1pm至10pm)的范围内(其中浓度是指可以检测至靶rna的阈值靶rna浓度)。在一些情况下,本公开的方法具有在800fm至100pm的范围内的检测阈值。在一些情况下,本公开的方法具有在1pm至10pm的范围内的检测阈值。在一些情况下,本公开的方法具有在10fm至500fm,例如10fm至50fm、50fm至100fm、100fm至250fm或250fm至500fm的范围内的检测阈值。
在一些情况下,可以在样品中检测到单链靶rna的最小浓度在500fm至1nm(例如,500fm至500pm、500fm至200pm、500fm至100pm、500fm至10pm、500fm至1pm、800fm至1nm、800fm至500pm、800fm至200pm、800fm至100pm、800fm至10pm、800fm至1pm、1pm至1nm、1pm至500pm、1pm至200pm、1pm至100pm或1pm至10pm)的范围内。在一些情况下,可以在样品中检测到单链靶rna的最小浓度在800fm至100pm的范围内。在一些情况下,可以在样品中检测到单链靶rna的最小浓度在1pm至10pm的范围内。
在一些情况下,本公开的方法可以检测包含多种rna(包括单链靶rna和多种非靶rna)的样品中存在的靶单链rna,其中所述靶单链rna以低至500fm(例如,低至800fm、低至1pm、低至10pm或低至100pm)的浓度存在。在一些情况下,本公开的方法可以检测包含多种rna(包括单链靶rna和多种非靶rna)的样品中存在的靶单链rna,其中所述靶单链rna以低至1pm的浓度存在。
在一些情况下,本公开的方法可以检测包含多种rna(包括单链靶rna和多种非靶rna)的样品中存在的靶单链rna,其中所述靶单链rna以低至500fm(例如,低至800fm、低至1pm、低至10pm或低至100pm)的浓度存在,并且其中在测量步骤之前使样品接触60分钟或更短时间(例如,在一些情况下为40分钟或更短时间)。在一些情况下,本公开的方法可以检测包含多种rna(包括单链靶rna和多种非靶rna)的样品中存在的靶单链rna,其中所述靶单链rna以低至1pm的浓度存在,并且其中在测量步骤之前使样品接触60分钟或更短时间(例如,在一些情况下为40分钟或更短时间)。
例如,在一些情况下,本公开的方法提供用于检测以500fm或更高(例如,800fm或更高、1pm或更高、5pm或更高、10pm或更高)的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以1pm或更高(例如,2pm或更高、5pm或更高、或8pm或更高)的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以500fm或更高(例如,1pm或更高、5pm或更高、10pm或更高)的浓度存在于样品中的靶rna,其中在测量步骤之前使样品接触60分钟或更短时间(例如,在一些情况下为40分钟或更短时间)。在一些情况下,本公开的方法提供用于检测以1pm或更高(例如,2pm或更高、5pm或更高、或8pm或更高)的浓度存在于样品中的靶rna,其中在测量步骤之前使样品接触60分钟或更短时间(例如,在一些情况下为40分钟或更短时间)。
在一些情况下,本公开的方法提供用于检测以10nm或更低的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以5nm或更低的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以1nm或更低的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以0.5nm或更低的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以0.1nm或更低的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以0.05nm或更低的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以0.01nm或更低的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以0.005nm或更低的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以0.001nm或更低的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以0.0005nm或更低的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供了以0.0001nm或更低的浓度检测样品中存在的靶rna。在一些情况下,本公开的方法提供用于检测以0.00005nm或更低的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以0.00001nm或更低的浓度存在于样品中的靶rna。
在一些情况下,本公开的方法提供用于检测以10-6nm至1nm,例如10-6nm至5×10-6nm、5×10-6nm至10-5nm、10-5nm至5×10-5nm、5×10-5nm至10-4nm、10-4nm至5×10-4nm、5×10-4nm至10-3nm、10-3nm至5×10-3nm、5×10-3nm至10-2nm、10-2nm至5×10-2nm、5×10-2nm至0.1nm、0.1nm至0.5nm、0.5nm至1nm、1nm至5nm或5nm至10nm的浓度存在于样品中的靶rna。
在一些情况下,本公开的方法提供用于检测以低于10nm的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以低于5nm的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以低于1nm的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以低于0.5nm的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以低于0.1nm的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以低于0.05nm的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以低于0.01nm的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以低于0.005nm的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以低于0.001nm的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以低于0.0005nm的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以低于0.0001nm的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以低于0.00005nm的浓度存在于样品中的靶rna。在一些情况下,本公开的方法提供用于检测以低于0.00001nm的浓度存在于样品中的靶rna。
在一些情况下,本公开的方法可用于确定样品(例如,包含靶rna和多种非靶rna的样品)中靶rna的量。确定样品中靶rna的量可以包括将从测试样品产生的可检测信号的量与从参考样品产生的可检测信号的量进行比较。确定样品中靶rna的量可包括:测量可检测信号以产生测试测量值;测量由参考样品产生的可检测信号以产生参考测量值;以及将测试测量值与参考测量值进行比较以确定样品中存在的靶rna的量。
例如,在一些情况下,用于确定样品中靶rna的量的本公开的方法包括:a)使样品(例如,包含靶rna和多种非靶rna的样品)与以下接触:(i)与单链靶rna杂交的c2c2指导rna,和(ii)切割样品中存在的rna的c2c2蛋白;b)测量由c2c2蛋白介导的rna切割产生的可检测信号,产生测试测量值;c)测量由参考样品产生的可检测信号以产生参考测量值;以及d)将测试测量值与参考测量值进行比较,以确定样品中存在的靶rna的量。
作为另一实例,在一些情况下,用于确定样品中靶rna的量的本公开的方法包括:a)使样品(例如,包含靶rna和多种非靶rna的样品)与以下接触:i)前体c2c2指导rna阵列,所述前体c2c2指导rna阵列包含两个或更多个c2c2指导rna,每个所述c2c2指导rna具有不同的指导序列;和(ii)c2c2蛋白,所述c2c2蛋白将前体c2c2指导rna阵列切割成单独的c2c2指导rna,并且还切割样品的rna;b)测量由c2c2蛋白介导的rna切割产生的可检测信号,产生测试测量值;c)测量由两个或更多个参考样品中的每一个产生的可检测信号,以产生两个或更多个参考测量值;以及d)将测试测量值与参考测量值进行比较,以确定样品中存在的靶rna的量。
样品
主题样品包括多种靶rna。术语“多个”在本文中用于表示两个或更多个。因此,在一些情况下,样品包括两种或更多种(例如,3种或更多种、5种或更多种、10种或更多种、20种或更多种、50种或更多种、100种或更多种、500种或更多种、1,000种或更多种、或5,000种或更多种)rna。本方法可以用作检测复杂的rna混合物中存在的单链靶rna的非常灵敏的方法。因此,在一些情况下,样品包括5种或更多种(例如,10种或更多种、20种或更多种、50种或更多种、100种或更多种、500种或更多种、1,000种或更多种、或5,000种或更多种rna)序列彼此不同的rna。在一些情况下,样品包括10种或更多种、20种或更多种、50种或更多种、100种或更多种、500种或更多种、103种或更多种、5×103种或更多种、104种或更多种、5×104种或更多种、105种或更多种、5×105种或更多种、106种或更多种、5×106种或更多种、或107种或更多种序列彼此不同的rna。在一些情况下,样品包含10种至20种、20种至50种、50种至100种、100种至500种、500种至103种、103种至5×103种、5×103种至104种、104种至5×104种、5×104种至105种、105种至5×105种、5×105种至106种、106种至5×106种、或5×106种至107种、或多于107种序列彼此不同的rna。在一些情况下,样品包含5种至107种序列彼此不同的rna(例如,5种至106种、5种至105种、5种至50,000种、5种至30,000种、10种至106种、10种至105种、10种至50,000种、10种至30,000种、20种至106种、20种至105种、20种至50,000种、或20种至30,000种序列彼此不同的rna)。在一些情况下,样品包含5种至50,000种序列彼此不同的rna(例如,5种至30,000种、10种至50,000种、或10种至30,000种序列彼此不同的rna)。在一些情况下,样品包括20种或更多种序列彼此不同的rna。在一些情况下,样品包括来自细胞裂解物(例如,真核细胞裂解物、哺乳动物细胞裂解物、人细胞裂解物、原核细胞裂解物、植物细胞裂解物等)的rna。例如,在一些情况下,样品包括从细胞诸如真核细胞(例如,哺乳动物细胞,诸如人细胞)表达的rna。
术语“样品”在本文中用于表示包括单链rna的任何样品。样品可以来源于任何来源,例如,样品可以是纯化rna的合成组合;样品可以是细胞裂解物、富含rna的细胞裂解物,或从细胞裂解物中分离和/或纯化的rna。样品可以来自患者(例如,用于诊断目的)。样品可来自经透化的细胞。样品可以来自交联细胞。样品可以在组织切片中。样品可以来自通过交联之后进行脱脂和调节以产生均匀的折射率而制备的组织。在例如shah等人,development(2016)143,2862-2867doi:10.1242/dev.138560中已经描述了通过交联之后进行脱脂和调节以产生均匀的折射率而制备的组织的实例。
“样品”可包括单链靶rna和多种非靶rna。在一些情况下,靶单链rna以每10个非靶rna一个拷贝、每20个非靶rna一个拷贝、每25个非靶rna一个拷贝、每50个非靶rna一个拷贝、每100个非靶rna一个拷贝、每500个非靶rna一个拷贝、每103个非靶rna一个拷贝、每5×103个非靶rna一个拷贝、每104个非靶rna一个拷贝、每5×104个非靶rna一个拷贝、每105个非靶rna一个拷贝、每5×105个非靶rna一个拷贝、每106个非靶rna一个拷贝、或每106个非靶rna少于一个拷贝存在于样品中。在一些情况下,靶单链rna以每10个非靶rna一个拷贝至每20个非靶rna1个拷贝、每20个非靶rna1个拷贝至每50个非拷贝1个拷贝、每50个非靶rna1个拷贝至每100个非靶rna1个拷贝、每100个非靶rna1个拷贝至每500个非靶rna1个拷贝、每500个非靶rna1个拷贝至每103个非靶rna1个拷贝、每103个非靶rna1个拷贝至每5×103个非靶rna1个拷贝、每5×103个非靶rna1个拷贝至每104个非靶rna1个拷贝、每104个非靶rna1个拷贝至每105个非靶rna1个拷贝、每105个非靶rna1个拷贝至每106个非靶rna1个拷贝,或每106个非靶rna1个拷贝至每107个非靶rna1个拷贝存在于样品中。
合适的样品包括但不限于血液样品、血清样品、血浆样品、尿液样品、抽吸物样品和活组织检查样品。因此,关于患者的术语“样品”涵盖血液和其它生物来源的液体样品、实体组织样品(诸如活检样本或组织培养物或来源于其的细胞及其后代)。该定义还包括在获得后已经以任何方式操纵的样品,诸如通过用试剂处理;洗涤;或富集某些细胞群,诸如癌细胞。该定义还包括已经富集了特定类型的分子(例如,rna)的样品。术语“样品”涵盖生物样品,诸如临床样品,诸如血液、血浆、血清、抽吸物、脑脊髓液(csf),并且还包括通过手术切除获得的组织、通过活组织检查获得的组织、培养中的细胞、细胞上清液、细胞裂解物、组织样品、器官、骨髓等。“生物样品”包括来源于其的生物流体(例如,癌细胞、感染细胞等),例如包含从此类细胞获得的rna的样品(例如,细胞裂解物或其它包含rna的细胞提取物)。
样品可包括或可从多种细胞、组织、器官或无细胞流体中的任何一种获得。合适的样品来源包括真核细胞、细菌细胞和古细菌细胞。合适的样品来源包括单细胞生物和多细胞生物。合适的样品来源包括单细胞真核生物;植物或植物细胞;藻类细胞,例如布朗葡萄藻(botryococcusbraunii)、莱茵衣藻(chlamydomonasreinhardtii)、迦得拟微绿球藻(nannochloropsisgaditana)、蛋白核小球藻(chlorellapyrenoidosa)、展枝马尾藻(sargassumpatens)、羽藻(c.agardh)等等;真菌细胞(例如,酵母细胞);动物细胞、组织或器官;来自无脊椎动物的细胞、组织或器官(例如,果蝇、刺胞动物、棘皮动物、线虫、昆虫、蜘蛛类动物,等);来自脊椎动物(例如,鱼类、两栖动物、爬行动物、鸟类、哺乳动物)的细胞、组织、流体或器官;来自哺乳动物(例如,人;非人灵长类动物;有蹄类动物;猫科动物;牛;绵羊;山羊;等)的细胞、组织、流体或器官。合适的样品来源包括线虫、原生动物等。合适的样品来源包括寄生虫,诸如蠕虫、疟疾寄生虫等。
合适的样品来源包括例如以下六个界中任何一个界的细胞、组织或生物:细菌界(例如,真细菌界);古细菌界;原生生物界;真菌界;植物界;和动物界。合适的样品来源包括原生生物界的植物样成员,包括但不限于藻类(例如,绿藻、红藻、灰胞藻、蓝细菌);原生生物界的真菌样成员,例如粘液菌、水霉菌等;原生生物界的动物样成员,例如鞭毛虫类(例如,眼虫藻)、变形虫类(amoeboids)(例如,变形虫)、孢子虫类(例如,顶复门(apicomplexa)、粘体动物门(myxozoa)、微孢子虫纲(microsporidia))和纤毛虫类(例如,草履虫)。合适的样品来源包括真菌界的成员,包括但不限于以下门中的任何门的成员:担子菌门(担子菌类;例如,伞菌属、鹅膏菌属、牛肝菌属、鸡油菌属等成员);子囊菌门(子囊菌类,包括例如酵母菌);菌藻门(地衣);接合菌门(接合真菌);以及不完全菌门。合适的样品来源包括植物界的成员,包括但不限于以下门中的任何门的成员:苔藓植物门(例如,藓类)、角苔植物门(例如,角苔类)、苔类植物门(hepaticophyta)(例如,苔类)、石松植物门(例如,石松类)、楔叶植物门(例如,木贼类)、裸蕨植物门(例如,松叶蕨类)、瓶尔小草门、蕨门(例如,蕨类)、苏铁门、银杏门、松柏门、买麻藤门和木兰门(例如,显花植物)。合适的样品来源包括动物界的成员,包括但不限于以下门中的任何门的成员:多孔动物门(海绵动物);扁盘动物门;直泳虫门(海洋无脊椎动物的寄生物);菱形虫门;刺胞动物门(珊瑚、海葵、海蜇、海笔、海肾、立方水母);栉水母门(栉水母类);扁虫动物门(扁虫类);纽形动物门(纽虫类);颚胃动物门(ngathostomulida)(有颚蠕虫);腹毛动物门;轮虫动物门;曳鳃动物门;动吻动物门;铠甲动物门;棘头动物门;内肛亚门;线虫动物门;线形动物门;环口动物门;软体动物门(软体动物);星虫动物门(方格星虫(peanutworms));环节动物门(环节蠕虫);缓步动物门(缓步动物);有爪动物门(栉蚕);节肢动物门(包括以下亚门:有螯肢亚门、多足亚门、六足亚门和甲壳亚门,其中有螯肢亚门包括例如蛛形纲、肢口纲和海蜘蛛纲,多足亚门包括例如唇足纲(唇足类)、倍足纲(多足类)、少足纲(paropoda)和综合纲,六足亚门包括昆虫纲,甲壳亚门包括虾、磷虾、藤壶等;帚虫动物门;外肛动物门(苔藓动物);腕足动物门;棘皮动物门(例如,海星、海雏菊、毛头星、海胆、海参、海蛇尾、脆篮(brittlebaskets)等);毛颚动物门(箭虫);半索动物门(玉钩虫);和脊索动物门。合适的脊索动物门成员包括以下亚门的任何成员:尾索动物亚门(海鞘纲;包括海鞘目、樽海鞘目和幼形目);头索亚门(文昌鱼);盲鳗纲(盲鳗);和脊椎动物亚门,其中脊椎动物亚门成员包括例如以下纲的成员:鳃鳗纲(七鳃类),软骨鱼纲(软骨鱼),辐鳍鱼纲(辐鳍鱼),腔棘焦纲(腔棘鱼),肺鱼纲(肺鱼),爬行纲(爬行动物,例如蛇、短吻鳄、鳄鱼、蜥蜴等),鸟纲(鸟);和哺乳纲(哺乳动物)。合适的植物包括任何单子叶植物和任何双子叶植物。
合适的样品来源包括取自生物;从生物体分离的特定细胞或细胞群的细胞、流体、组织或器官等等。例如,在生物是植物的情况下,合适的来源包括木质部、韧皮部、形成层、叶、根等。在生物是动物的情况下,合适的来源包括特定组织(例如,肺、肝、心脏、肾、脑、脾、皮肤、胎儿组织等)、或特定细胞类型(例如,神经元细胞、上皮细胞、内皮细胞、星形胶质细胞、巨噬细胞、神经胶质细胞、胰岛细胞、t淋巴细胞、b淋巴细胞等)。
在一些情况下,样品的来源是患病的细胞、流体、组织或器官。在一些情况下,样品的来源是正常(非患病的)细胞、流体、组织或器官。在一些情况下,样品来源是病原体感染的细胞、组织或器官。病原体包括病毒、真菌、蠕虫、原生动物、疟疾寄生虫、疟原虫寄生虫、弓形虫寄生虫,血吸虫寄生虫等。“蠕虫”包括蛔虫、犬恶丝虫和植食性线虫(线虫纲)、吸虫(吸虫纲)、棘头虫、和绦虫(绦虫纲)。原生动物感染包括来自贾第虫属,毛滴虫属、非洲锥虫病、阿米巴痢疾、巴贝虫病、小袋虫性痢疾、查加斯病(chaga'sdisease)、球虫病、疟疾和弓形体病的感染。病原体(诸如寄生/原生动物病原体)的实例包括但不限于:恶性疟原虫(plasmodiumfalciparum)、间日疟原虫(plasmodiumvivax)、克氏锥虫(trypanosomacruzi)和刚地弓形虫(toxoplasmagondii)。真菌病原体包括但不限于:新型隐球菌(cryptococcusneoformans)、荚膜组织胞浆菌(histoplasmacapsulatum)、粗球孢菌(coccidioidesimmitis)、皮炎芽生菌(blastomycesdermatitidis)、沙眼衣原体(chlamydiatrachomatis)和白色念珠菌(candidaalbicans)。病原性病毒包括例如免疫缺陷病毒(例如,hiv);流感病毒;登革热病毒;西尼罗河病毒;疱疹病毒;黄热病毒;肝炎病毒c;肝炎病毒a;肝炎病毒b;乳头瘤病毒;等等。病原体包括例如hiv病毒、结核分枝杆菌(mycobacteriumtuberculosis)、无乳链球菌(streptococcusagalactiae)、耐甲氧西林金黄色葡萄球菌(methicillin-resistantstaphylococcusaureus)、嗜肺军团菌(legionellapneumophila)、酿脓链球菌(streptococcuspyogenes)、大肠杆菌(escherichiacoli)、淋病奈瑟氏菌(neisseriagonorrhoeae)、脑膜炎奈瑟氏菌(neisseriameningitidis),肺炎球菌(pneumococcus)、新型隐球菌(cryptococcusneoformans)、荚膜组织胞浆菌(histoplasmacapsulatum)、流感嗜血杆菌b(hemophilusinfluenzaeb)、梅毒密螺旋体(treponemapallidum)、莱姆病螺旋体(lymediseasespirochetes)、铜绿假单胞菌(pseudomonasaeruginosa)、麻风分枝杆菌(mycobacteriumleprae)、流产布鲁氏菌(brucellaabortus)、狂犬病病毒(rabiesvirus)、流感病毒(influenzavirus)、巨细胞病毒(cytomegalovirus)、单纯疱疹病毒i(herpessimplexvirusi)、单纯疱疹病毒ii(herpessimplexvirusii)、人血清细小样病毒(humanserumparvo-likevirus)、呼吸道合胞体病毒(respiratorysyncytialvirus)、水痘-带状疱疹病毒(varicella-zostervirus)、乙型肝炎病毒(hepatitisbvirus)、丙型肝炎病毒(hepatitiscvirus)、麻疹病毒(measlesvirus)、腺病毒(adenovirus)、人t细胞白血病病毒(humant-cellleukemiaviruses)、艾普斯登-巴尔病毒(epstein-barrvirus)、鼠白血病病毒(murineleukemiavirus)、腮腺炎病毒(mumpsvirus)、水疱性口炎病毒(vesicularstomatitisvirus)、辛德毕斯病毒(sindbisvirus)、淋巴细胞脉络丛脑膜炎病毒(lymphocyticchoriomeningitisvirus)、疣病毒(wartvirus)、蓝舌病毒(bluetonguevirus)、仙台病毒(sendaivirus)、猫白血病病毒(felineleukemiavirus)、呼肠孤病毒(reovirus)、脊髓灰质炎病毒(poliovirus)、猿猴病毒40(simianvirus40)、小鼠乳腺肿瘤病毒(mousemammarytumorvirus)、登革热病毒(denguevirus)、风疹病毒(rubellavirus)、西尼罗河病毒(westnilevirus)、恶性疟原虫(plasmodiumfalciparum)、间日疟原虫(plasmodiumvivax)、刚地弓形虫(toxoplasmagondii)、蓝氏锥虫(trypanosomarangeli)、克氏锥虫(trypanosomacruzi)、罗氏锥虫(trypanosomarhodesiense)、布氏锥虫(trypanosomabrucei)、曼氏血吸虫(schistosomamansoni)、日本血吸虫(schistosomajaponicum)、巴贝斯虫(babesiabovis)、柔嫩艾美球虫(eimeriatenella)、盘尾丝虫(onchocercavolvulus)、利什曼原虫(leishmaniatropica)、结核分枝杆菌(mycobacteriumtuberculosis)、旋毛虫(trichinellaspiralis)、泰勒原虫(theileriaparva)、胞状绦虫(taeniahydatigena)、羊绦虫(taeniaovis)、牛肉绦虫(taeniasaginata)、细粒棘球绦虫(echinococcusgranulosus)、柯氏中殖孔绦虫(mesocestoidescorti)、关节炎支原体(mycoplasmaarthritidis)、猪鼻支原体(m.hyorhinis)、口腔支原体(m.orale)、精氨酸支原体(m.arginini)、莱氏无胆甾原体(acholeplasmalaidlawii)、唾窦支原体(m.salivarium)和肺炎支原体(m.pneumoniae)。
靶rna
靶rna可以是任何单链rna(ssrna)。实例包括但不限于mrna、rrna、trna、非编码rna(ncrna)、长非编码rna(lncrna)和微rna(mirna)。在一些情况下,靶ssrna是mrna。在一些情况下,单链靶核酸是来自病毒(例如,寨卡病毒、人免疫缺陷病毒、流感病毒等)的ssrna。在一些情况下,单链靶核酸是寄生虫的ssrna。在一些情况下,单链靶核酸是细菌(例如,病原细菌)的ssrna。如上所述,靶rna的来源可以与rna样品的来源相同。
测量可检测信号
在一些情况下,本方法包括测量步骤(例如,测量由c2c2蛋白介导的rna切割产生的可检测信号)。因为c2c2蛋白一旦被激活就切割非靶向rna(所述激活当在存在c2c2蛋白的情况下c2c2指导rna与靶rna杂交时发生),所以可检测信号可以是当rna被切割时产生的任何信号。例如,在一些情况下,测量步骤可以包括以下中的一者或多者:基于金纳米颗粒的检测(例如参见xu等人,angewchemintedengl.2007;46(19):3468-70;和xia等人,procnatlacadsciusa.2010年6月15日;107(24):10837-41)、荧光偏振、胶体相变/分散(例如,baksh等人,nature.2004年1月8日;427(6970):139-41)、电化学检测、基于半导体的感测(例如,rothberg等人,nature.2011年7月20日;475(7356):348-52;例如,可以在rna切割反应后使用磷酸酶以通过打开2'-3'环状磷酸酯并通过将无机磷酸盐释放到溶液中而产生ph变化),以及检测经标记的检测rna(更多细节参见下文)。这种检测方法的读出可以是任何合宜的读出。可能的读出的实例包括但不限于:所测量的可检测荧光信号的量;对凝胶上条带(例如,表示切割产物对比未切割底物的条带)的视觉分析、基于视觉或传感器的对颜色存在与否的检测(即,颜色检测方法),以及是否存在电信号(或电信号的特定量)。
在一些情况下,例如在检测到的信号量可以用于确定样品中存在的靶rna的量的意义上,测量可以是定量的。在一些情况下,例如在可检测信号的存在或不存在可以指示靶rna的存在或不存在的意义上,测量可以是定性的。在一些情况下,除非靶rna以高于特定阈值浓度存在(例如,参见图5),否则可检测信号将不存在(例如,高于给定阈值水平)。在一些情况下,可以通过修改c2c2蛋白、指导rna、样品体积和/或检测rna(如果使用的话)的量来滴定检测阈值。因此,例如,如本领域的普通技术人员应当理解的,如果需要的话,可以使用许多对照以建立一个或多个反应,每个反应设置用于检测靶rna的不同阈值水平,并且因此可以使用这样的一系列反应来确定样品中存在的靶rna的量(例如,可以使用这样的一系列反应来确定靶rna‘以至少x的浓度’存在于样品中)。
经标记的检测rna
在一些情况下,本方法包括使样品(例如,包含靶rna和多种非靶rna的样品)与以下接触:i)经标记的检测rna;ii)c2c2蛋白;以及iii)c2c2指导rna(或前体c2c2指导rna阵列)。例如,在一些情况下,本方法包括使样品与经标记的检测rna接触,所述经标记的检测rna包含荧光发射染料对;c2c2蛋白在被激活(通过在指导rna与靶rna杂交的背景下与c2c2指导rna结合)后切割所述经标记的检测rna;并且所测量到的可检测信号是由荧光发射染料对产生的。例如,在一些情况下,本方法包括使样品与经标记的检测rna接触,所述经标记的检测rna包含荧光共振能量转移(fret)对或猝灭剂/荧光剂对或两者。在一些情况下,本方法包括使样品与包含fret对的经标记的检测rna接触。在一些情况下,本方法包括使样品与包含荧光剂/猝灭剂对的经标记的检测rna接触。荧光发射染料对包含fret对或猝灭剂/荧光剂对。在fret对和猝灭剂/荧光剂对的两种情况下,染料中的一种染料的发射光谱与所述对中的另一种染料的吸收光谱的某一区域重叠。如本文所用的,术语“荧光发射染料对”是用于包括“荧光共振能量转移(fret)对”和“猝灭剂/荧光剂对”两者的通用术语,这两个术语将在下面更详细地讨论。术语“荧光发射染料对”与短语“fret对和/或猝灭剂/荧光剂对”可互换地使用。
在一些情况下(例如,当检测rna包括fret对时),经标记的检测rna在被切割之前产生一定量的可检测信号,并且当所述经标记的检测rna被切割时所测量到的可检测信号的量减少。在一些情况下,经标记的检测rna在被切割之前产生第一可检测信号(例如,来自fret对),并且当所述经标记的检测rna被切割时产生第二可检测信号(例如,来自猝灭剂/荧光剂对)。因此,在一些情况下,经标记的检测rna包含fret对和猝灭剂/荧光剂对。
在一些情况下,经标记的检测rna包括fret对。fret是这样的过程,通过所述过程,能量的无辐射转移发生为从激发态荧光团到紧邻的第二发色团。能量转移可以发生的范围限制在约10纳米(100埃),并且转移效率对荧光团之间的分开距离非常敏感。因此,如本文所使用的,术语“fret”(“荧光共振能量转移”;也被称为“萤光共振能量转移”)是指这样的物理现象,所述物理现象涉及供体荧光团和匹配的受体荧光团被选择为使得供体的发射光谱与受体的激发光谱重叠,并且被进一步选择为使得当供体和受体彼此非常接近(通常为10nm或更短距离)时,供体的激发将引起来自受体的激发和发射,这是因为一些能量经由量子耦合效应从供体传递到受体。因此,fret信号用作供体和受体的接近度尺度;只有当它们彼此非常接近时才会产生信号。fret供体部分(例如,供体荧光团)和fret受体部分(例如,受体荧光团)在本文中统称为“fret对”。
供体-受体对(fret供体部分和fret受体部分)在本文中被称为“fret对”或“信号fret对”。因此,在一些情况下,主题经标记的检测rna包括两个信号配偶体(信号对),当一个信号配偶体是fret供体部分时,另一个信号配偶体是fret受体部分。因此,当信号配偶体非常接近时(例如,在相同的rna分子上时),包括这种fret对(fret供体部分和fret受体部分)的主题经标记的检测rna将显示可检测信号(fret信号),但是当所述配偶体分开时(例如,在用c2c2蛋白切割rna分子后),信号将减少(或不存在)。
fret供体和受体部分(fret对)将是本领域的普通技术人员已知的,并且可以使用任何合宜的fret对(例如,任何合宜的供体和受体部分对)。合适的fret对的实例包括但不限于表1中所示的那些。还参见:bajar等人,sensors(basel).2016年9月14日;16(9);和abraham等人,plosone.2015年8月3日;10(8):e0134436。
表1.fret对的实例(供体和受体fret部分)
(1)5-(2-碘乙酰基氨基乙基)氨基萘-1-磺酸
(2)n-(4-二甲基氨基-3,5-二硝基苯基)马来酰亚胺
(3)羧基荧光素琥珀酰亚胺酯
(4)4,4-二氟-4-硼杂-3a,4a-二氮杂-s-吲哒生
在一些情况下,可检测信号在经标记的检测rna被切割时产生(例如,在一些情况下,经标记的检测rna包含猝灭剂/荧光剂对。信号猝灭对的一个信号配偶体产生可检测信号,而另一个信号配偶体是猝灭剂部分,所述猝灭剂部分猝灭第一信号配偶体的可检测信号(即,猝灭剂部分猝灭信号部分的信号,使得当信号配偶体彼此接近时(例如,当信号对的信号配偶体非常接近时),来自信号部分的信号减少(被猝灭)。
例如,在一些情况下,当经标记的检测rna被切割时,可检测信号的量增加。例如,在一些情况下,由一个信号配偶体(信号部分)表现的信号被另一个信号配偶体(猝灭剂信号部分)猝灭,例如当在由c2c2蛋白进行切割之前两者存在于相同的rna分子上时。这种信号对在本文中称为“猝灭剂/荧光剂对”、“猝灭对”或“信号猝灭对”。例如,在一些情况下,一个信号配偶体(例如,第一信号配偶体)是产生可检测信号的信号部分,所述可检测信号由第二信号配偶体(例如,猝灭剂部分)猝灭。因此,当配偶体被分开时(例如,在由c2c2蛋白切割检测rna之后),这种猝灭剂/荧光剂对的信号配偶体将产生可检测信号,但是当配偶体非常接近(例如,在由c2c2蛋白切割检测rna之前)时,所述信号将被猝灭。
猝灭剂部分可以将来自信号部分的信号(例如,在由c2c2蛋白切割检测rna之前)猝灭至不同程度。在一些情况下,猝灭剂部分猝灭来自信号部分的信号,其中在存在猝灭剂部分的情况下(当信号配偶体彼此接近时)检测到的信号是在不存在猝灭剂部分(当信号配偶体分开时)时检测到的信号的95%或更少。例如,在一些情况下,在存在猝灭剂部分的情况下检测到的信号可以是在不存在猝灭剂部分的情况下检测到的信号的90%或更少、80%或更少、70%或更少、60%或更少、50%或更少、40%或更少、30%或更少、20%或更少、15%或更少、10%或更少、或5%或更少。在一些情况下,在存在猝灭剂部分的情况下未检测到信号(例如,高于背景)。
在一些情况下,在不存在猝灭剂部分的情况下(当信号配偶体分开时)检测到的信号是在存在猝灭剂部分的情况下(当信号配偶体彼此接近时)检测到的信号的至少1.2倍(例如,至少1.3倍、至少1.5倍、至少1.7倍、至少2倍、至少2.5倍、至少3倍、至少3.5倍、至少4倍、至少5倍、至少7倍、至少10倍、至少20倍或至少50倍)。
在一些情况下,信号部分是荧光标记。在一些此类情况下,猝灭剂部分猝灭来自荧光标记的信号(光信号)(例如,通过吸收标记的发射光谱中的能量)。因此,当猝灭剂部分不与信号部分接近时,来自荧光标记的发射(信号)是可检测的,因为信号不被猝灭剂部分吸收。可以使用任何合宜的供体受体对(信号部分/猝灭剂部分对),并且许多合适的对是本领域中已知的。
在一些情况下,猝灭剂部分从信号部分(在本文中也称为“可检测标记”)吸收能量,然后发射信号(例如,不同波长的光)。因此,在一些情况下,猝灭剂部分本身是信号部分(例如,信号部分可以是6-羧基荧光素,而猝灭剂部分可以是6-羧基-四甲基罗丹明),并且在一些此类情况下,所述对也可以是fret对。在一些情况下,猝灭剂部分是暗猝灭剂。暗猝灭剂可以吸收激发能量并以不同方式耗散能量(例如,作为热量)。因此,暗猝灭剂本身具有极小荧光至没有荧光(不发射荧光)。暗猝灭剂的实例进一步描述于美国专利号8,822,673和8,586,718;美国专利公开20140378330、20140349295和20140194611;以及国际专利申请:wo200142505和wo200186001中,所有这些专利都据此以引用方式整体并入本文。
荧光标记的实例包括但不限于:alexa
在一些情况下,可检测标记是选自以下的荧光标记:alexa
在一些情况下,可检测标记是选自以下的荧光标记:alexa
atto染料的实例包括但不限于:atto390、atto425、atto465、atto488、atto495、atto514、atto520、atto532、attorho6g、atto542、atto550、atto565、attorho3b、attorho11、attorho12、attothio12、attorho101、atto590、atto594、attorho13、atto610、atto620、attorho14、atto633、atto647、atto647n、atto655、attooxa12、atto665、atto680、atto700、atto725、以及atto740。
alexafluor染料的实例包括但不限于:alexa
猝灭剂部分的实例包括但不限于:暗猝灭剂、blackhole
在一些情况下,猝灭剂部分选自:暗猝灭剂、blackhole
atto猝灭剂的实例包括但不限于:atto540q、atto580q和atto612q。blackhole
对于一些可检测标记(例如,荧光染料)和/或猝灭剂部分的实例,参见例如bao等人,annurevbiomedeng.2009;11:25-47;以及美国专利号8,822,673和8,586,718;美国专利公开20140378330、20140349295、20140194611、20130323851、20130224871、20110223677、20110190486、20110172420、20060179585和20030003486;以及国际专利申请:wo200142505和wo200186001,所有这些参考文献据此以引用方式整体并入。
在一些情况下,可以通过测量比色读出来检测对经标记的检测rna的切割。例如,荧光团的释放(例如,从fret对释放、从猝灭剂/荧光剂对释放等)可导致可检测信号的波长偏移(并因此色移)。因此,在一些情况下,可以通过色移检测主题经标记的检测rna的切割。这种偏移可以表示为一种颜色(波长)信号的量损失、另一种颜色的量的增益、一种颜色与另一种颜色的比率的变化等。
核酸修饰
在一些情况下,经标记的检测rna包含一种或多种修饰,例如碱基修饰、骨架修饰、糖修饰等,以为核酸提供新的或增强的特征(例如,改善的稳定性)。如本领域中已知的,核苷是碱基-核糖组合。核苷的碱基部分通常是杂环碱基。此类杂环碱基的两种最常见的类别是嘌呤和嘧啶。核苷酸是这样的核苷,所述核苷还包括与核苷的糖部分共价连接的磷酸基团。对于那些包括戊呋喃糖基糖的核苷,磷酸基团可以与所述糖的2'、3'或5'羟基部分连接。在形成寡核苷酸时,磷酸基团将相邻的核苷彼此共价连接以形成直链聚合化合物。继而,该直链聚合化合物的各个末端可以进一步连接以形成环状化合物,然而,直链化合物通常是合适的。此外,线性化合物可以具有内部核苷酸碱基互补性,并且因此可以以产生完全或部分双链化合物的方式折叠。在寡核苷酸内,磷酸基团通常被称为形成寡核苷酸的核苷间骨架。rna和dna的正常键或骨架是3'至5'磷酸二酯键。
经修饰的骨架和经修饰的核苷间键
合适的修饰的实例包括经修饰的核酸骨架和非天然的核苷间键。具有经修饰的骨架的核酸包括在骨架中保留磷原子的核酸和在骨架中不具有磷原子的核酸。
其中含有磷原子的合适的经修饰寡核苷酸骨架包含例如硫代磷酸酯、手性硫代磷酸酯、二硫代磷酸酯、磷酸三酯、氨基烷基磷酸三酯、甲基和其它烷基膦酸酯(包括3'-亚烷基膦酸酯、5'-亚烷基膦酸酯和手性膦酸酯)、次膦酸酯、氨基磷酸酯(包括3'-氨基氨基磷酸酯和氨基烷基氨基磷酸酯)、二氨基磷酸酯、硫代氨基磷酸酯、硫代烷基膦酸酯、硫代烷基磷酸三酯、具有正常3'-5'键的硒代磷酸酯和硼代磷酸酯、硒代磷酸酯和硼代磷酸酯的2'-5'连接的类似物,以及具有反向极性的硒代磷酸酯和硼代磷酸酯,其中一个或多个核苷酸间键是3'至3'键、5'至5'键或2'至2'键。合适的具有反向极性的寡核苷酸包含在最3'核苷酸间键处的单个3'至3'键,即单个反向核苷残基,所述单个反向核苷残基可以是碱性的(核碱基丢失或具有羟基代替其)。还包括各种盐(诸如,钾盐或钠盐)、混合盐和游离酸形式。
在一些情况下,经标记的检测rna包含一个或多个硫代磷酸酯和/或杂原子核苷间键,特别是-ch2-nh-o-ch2-、-ch2-n(ch3)-o-ch2-(称为亚甲基(甲基亚氨基)或mmi骨架)、-ch2-o-n(ch3)-ch2-、-ch2-n(ch3)-n(ch3)-ch2-和-o-n(ch3)-ch2-ch2-(其中天然磷酸二酯核苷酸间键表示为-o-p(=o)(oh)-o-ch2-)。mmi型核苷间键在上面引用的美国专利号5,489,677中公开。合适的酰胺核苷间键在美国专利号5,602,240中公开。
同样合适的是如在例如美国专利号5,034,506中所述的具有吗啉代骨架结构的核酸。例如,在一些情况下,经标记的检测rna包含6元吗啉代环代替核糖环。在一些情况下,二氨基磷酸酯或其它非磷酸二酯核苷间键取代磷酸二酯键。
其中不含磷原子的合适的经修饰多核苷酸骨架具有由短链烷基或环烷基核苷间键、混合杂原子和烷基或环烷基核苷间键、或一个或多个短链杂原子或杂环核苷间键形成的骨架。这些骨架包括具有吗啉代键(部分地由核苷的糖部分形成)的那些骨架;硅氧烷骨架;硫化物、亚砜和砜骨架;甲酰基和硫代甲酰基骨架;亚甲基甲酰基和硫代甲酰基骨架;核糖乙酰骨架;含有烯基的骨架;氨基磺酸酯骨架;亚甲基亚氨基和亚甲基肼基骨架;磺酸酯和磺酰胺骨架;酰胺骨架;以及其它具有混合的n、o、s和ch2组成部分的骨架。
模拟物
经标记的检测rna可以是核酸模拟物。应用于多核苷酸的术语“模拟物”旨在包括这样的多核苷酸,在所述多核苷酸中仅呋喃糖环或呋喃糖环和核苷酸间键两者被非呋喃糖基团替代,仅替代呋喃糖环也在本领域中被称为糖替代物。维持杂环碱基部分或经修饰的杂环碱基部分以与合适的靶核酸杂交。一种此类核酸,已显示具有优异杂交特性的多核苷酸模拟物,被称为肽核酸(pna)。在pna中,多核苷酸的糖骨架被含酰胺的骨架,特别是氨基乙基甘氨酸骨架取代。核苷酸被保留并直接或间接地结合骨架的酰胺部分的氮杂氮原子。
一种据报道具有优异杂交特性的多核苷酸模拟物是肽核酸(pna)。pna化合物中的骨架是两个或更多个连接的氨基乙基甘氨酸单元,所述氨基乙基甘氨酸单元赋予pna含酰胺的骨架。杂环碱基部分直接或间接地与骨架的酰胺部分的氮杂氮原子结合。描述pna化合物制备的代表性美国专利包括但不限于:美国专利号5,539,082;5,714,331;和5,719,262。
已经研究的另一类多核苷酸模拟物基于连接的吗啉代单元(吗啉代核酸),所述连接的吗啉代单元具有附接到吗啉代环的杂环碱基。已经报道了许多连接基团,所述连接基团连接吗啉代核酸中的吗啉代单体单元。一类连接基团已经被选择用于提供非离子寡聚化合物。非离子的基于吗啉代的寡聚化合物不太可能与细胞蛋白发生不希望的相互作用。基于吗啉代的多核苷酸是寡核苷酸的非离子模拟物,其不太可能与细胞蛋白形成不希望的相互作用(dwainea.braasch和davidr.corey,biochemistry,2002,41(14),4503-4510)。基于吗啉代的多核苷酸在美国专利号5,034,506中公开。已经制备了吗啉代类多核苷酸内的多种化合物,所述化合物具有各种连接单体亚基的不同连接基团。
另一类多核苷酸模拟物被称为环己烯基核酸(cena)。通常存在于dna/rna分子中的呋喃糖环被环己烯基环替代。cenadmt保护的亚磷酰胺单体已被制备并用于按照经典的亚磷酰胺化学进行寡聚化合物合成。已经制备并研究了完全修饰的cena寡聚化合物和具有用cena修饰的特定位置的寡核苷酸(参见wang等人,j.am.chem.soc.,2000,122,8595-8602)。通常,将cena单体掺入dna链中增加了其dna/rna杂交体的稳定性。cena寡腺苷酸以与天然复合物相似的稳定性而与rna和dna互补物形成复合物。通过nmr和圆二色性显示将cena结构掺入天然核酸结构中的研究,以进行简单的构象适应。
另一修饰包括锁定核酸(lna),其中2'-羟基与糖环的4'碳原子连接,从而形成2'-c,4'-c-氧亚甲基键,从而形成双环糖部分。键可以是亚甲基(-ch2-)、桥接2'氧原子与4'碳原子的基团,其中n是1或2(singh等人,chem.commun.,1998,4,455-456)。lna和lna类似物显示出非常高的互补dna和rna的双链热稳定性(tm=+3至+10℃),朝向3'-核酸外切降解的稳定性和良好的溶解性。已经描述了含有lna的有效和无毒的反义寡核苷酸(wahlestedt等人,proc.natl.acad.sci.u.s.a.,2000,97,5633-5638)。
已经描述了lna单体腺嘌呤、胞嘧啶、鸟嘌呤、5-甲基胞嘧啶、胸腺嘧啶和尿嘧啶的合成和制备,以及它们的寡聚化和核酸识别性质(koshkin等人,tetrahedron,1998,54,3607-3630)。lna及其制备还描述于wo98/39352和wo99/14226中。
经修饰的糖部分
经标记的检测rna还可包括一个或多个经取代的糖部分。合适的多核苷酸包含选自以下的糖取代基:oh;f;o-烷基、s-烷基或n-烷基;o-烯基、s-烯基或n-烯基;o-炔基、s-炔基或n-炔基;或o-烷基-o-烷基,其中所述烷基、烯基和炔基可以是取代或未取代的c1至c10烷基或c2至c10烯基和炔基。特别合适的是o((ch2)no)mch3、o(ch2)noch3、o(ch2)nnh2、o(ch2)nch3、o(ch2)nonh2以及o(ch2)non((ch2)nch3)2,其中n和m为1至约10。其它合适的多核苷酸包含选自以下的糖取代基:c1至c10低级烷基,取代的低级烷基、烯基、炔基,烷芳基,芳烷基,o-烷芳基或o-芳烷基,sh,sch3,ocn,cl,br,cn,cf3,ocf3,soch3,so2ch3,ono2,no2,n3,nh2,杂环烷基,杂环烷芳基,氨基烷基氨基,聚烷基氨基,取代的甲硅烷基,rna切割基团,报告基团,嵌入剂,用于改善寡核苷酸的药代动力学性质的基团,或用于改善寡核苷酸的药效学性质的基团,以及其它具有相似性质的取代基。合适的修饰包括2'-甲氧基乙氧基(2'-o-ch2ch2och3,也称为2'-o-(2-甲氧基乙基)或2'-moe)(martin等人,helv.chim.acta,1995,78,486-504),即烷氧基烷氧基基团。另一合适的修饰包括2'-二甲基氨氧乙氧基,即o(ch2)2on(ch3)2基团,也称为2'-dmaoe,如下文实施例中所述;以及2'-二甲氨基乙氧基乙氧基(在本领域中也称为2'-o-二甲基-氨基-乙氧基-乙基或2'-dmaeoe),即2'-o-ch2-o-ch2-n(ch3)2。
其它合适的糖取代基包括甲氧基(-o-ch3)、氨基丙氧基(--och2ch2ch2nh2)、烯丙基(-ch2-ch=ch2)、-o-烯丙基(--o--ch2—ch=ch2)和氟(f)。2'-糖取代基可以处于阿拉伯糖(上)位置或核糖(下)位置。合适的2'-阿拉伯糖修饰是2'-f。也可以在寡聚化合物上的其它位置,特别是3'末端核苷上或2'-5'连接的寡核苷酸的糖的3'位置处,以及5'末端核苷酸的5'位置处进行类似的修饰。寡聚化合物也可以具有糖模拟物,诸如环丁基部分代替戊呋喃糖基糖。
碱基修饰和取代
经标记的检测rna还可以包括核碱基(在本领域中通常简称为“碱基”)修饰或取代。如本文所用,“未修饰的”或“天然的”核碱基包括嘌呤碱基腺嘌呤(a)和鸟嘌呤(g),以及嘧啶碱基胸腺嘧啶(t)、胞嘧啶(c)和尿嘧啶(u)。经修饰的核碱基包括其它合成和天然核碱基,诸如5-甲基胞嘧啶(5-me-c)、5-羟甲基胞嘧啶、黄嘌呤、次黄嘌呤、2-氨基腺嘌呤、6-甲基腺嘌呤和鸟嘌呤以及腺嘌呤和鸟嘌呤的其它烷基衍生物、2-丙基腺嘌呤和鸟嘌呤以及腺嘌呤和鸟嘌呤的其它烷基衍生物、2-硫尿嘧啶、2-硫代胸腺嘧啶和2-硫代胞嘧啶、5-氟尿嘧啶和胞嘧啶、5-丙炔基(-c=c-ch3)尿嘧啶和胞嘧啶以及嘧啶碱基的其它炔基衍生物、6-偶氮尿嘧啶、胞嘧啶和胸腺嘧啶、5-尿嘧啶(假尿嘧啶)、4-硫尿嘧啶、8-卤代腺嘌呤和鸟嘌呤、8-氨基腺嘌呤和鸟嘌呤、8-巯基腺嘌呤和鸟嘌呤、8-硫代烷基腺嘌呤和鸟嘌呤、8-羟基腺嘌呤和鸟嘌呤以及其它8-取代的腺嘌呤和鸟嘌呤、5-卤代(特别是5-溴代)尿嘧啶和胞嘧啶、5-三氟甲基尿嘧啶和胞嘧啶以及其它5-取代的尿嘧啶和胞嘧啶、7-甲基鸟嘌呤和7-甲基腺嘌呤、2-f-腺嘌呤、2-氨基腺嘌呤、8-氮杂鸟嘌呤和8-氮杂腺嘌呤、7-脱氮杂鸟嘌呤和7-脱氮杂腺嘌呤,以及3-脱氮杂鸟嘌呤和3-脱氮杂腺嘌呤。其它经修饰的核碱基包括三环嘧啶,诸如吩噁嗪胞苷(1h-嘧啶并(5,4-b)(1,4)苯并噁嗪-2(3h)-酮)、吩噻嗪胞苷(1h-嘧啶(5,4-b)(1,4)苯并噻嗪-2(3h)-酮)、g-钳如取代的吩噁嗪胞苷(例如9-(2-氨基乙氧基)-h-嘧啶并(5,4-(b)(1,4)苯并噁嗪-2(3h)-酮)、咔唑胞苷(2h-嘧啶并(4,5-b)吲哚-2-酮)、吡啶并吲哚胞苷(h-吡啶并(3',2':4,5)吡咯并(2,3-d)嘧啶-2-酮)。
杂环碱基部分还可包括嘌呤碱基或嘧啶碱基被其它杂环取代的那些杂环碱基部分,例如7-脱氮杂腺嘌呤、7-脱氮杂鸟苷、2-氨基吡啶和2-吡啶酮。其它核碱基包括在美国专利号3,687,808中公开的那些核碱基;在theconciseencyclopediaofpolymerscienceandengineering,第858-859页,kroschwitz,j.i.编辑,johnwiley&sons,1990中公开的那些核碱基;由englisch等人,angewandtechemie,internationaledition,1991,30,613公开的那些核碱基;以及由sanghvi,y.s.,第15章,antisenseresearchandapplications,第289-302页,crooke,s.t.和lebleu,b.编辑,crcpress,1993公开的那些核碱基。这些核碱基中的某些可用于增加寡聚化合物的结合亲和力。这些核碱基包括5-取代的嘧啶、6-氮杂嘧啶,以及n-2、n-6和o-6取代的嘌呤,包括2-氨基丙基腺嘌呤、5-丙炔基尿嘧啶和5-丙炔基胞嘧啶。5-甲基胞嘧啶取代已显示为使核酸双链体稳定性增加0.6-1.2℃(sanghvi等人编辑,antisenseresearchandapplications,crcpress,bocaraton,1993,第276-278页),并且是合适的碱基取代,例如当与2'-o-甲氧基乙基糖修饰组合时。
检测两种不同的靶rna
如上所述,在一些情况下,本公开的方法提供了对样品中两种不同靶rna(第一单链靶rna和第二单链靶rna)的基本上同时检测。在一些情况下,所述方法包括:a)使样品(例如,包含两种不同靶rna和多种非靶rna的样品)与以下接触:(i)第一c2c2蛋白,其切割样品中存在的腺嘌呤+rna(即,包括a的rna,而不是缺乏a的rna,诸如聚urna);(ⅱ)第二c2c2蛋白,其切割尿嘧啶+rna(即,包括u的rna,而不是缺乏u的rna,如聚arna);(iii)第一c2c2指导rna,其包含与第一单链靶rna杂交的第一核苷酸序列和与第一c2c2蛋白结合的第二核苷酸序列;以及(iv)第二c2c2指导rna,其包含与第二单链靶rna杂交的第一核苷酸序列和与第二c2c2蛋白结合的第二核苷酸序列;以及b)测量由第一和第二c2c2蛋白介导的rna切割产生的可检测信号,其中第一可检测信号由第一c2c2蛋白产生,并且第二可检测信号由第二c2c2蛋白产生,其中所述第一可检测信号和所述第二可检测信号可彼此区分。在一些情况下,第一c2c2蛋白不被第二c2c2指导rna激活,并且第一c2c2蛋白切割包括a的ssrna(例如,不切割缺乏a的ssrna);并且第二c2c2蛋白不被第一c2c2指导rna激活,并且第二c2c2蛋白切割包括u的ssrna(例如,不切割缺乏u的ssrna)。在一些情况下,第一c2c2蛋白不被第二c2c2指导rna激活,并且第一c2c2蛋白切割包括u的ssrna(例如,不切割缺乏u的ssrna);并且第二c2c2蛋白不被第一c2c2指导rna激活,并且第二c2c2蛋白切割包括a的ssrna(例如,不切割缺乏a的ssrna)。
在一些情况下,所述方法还包括使样品与以下接触:i)第一经标记的检测rna,其包含第一fret对和/或第一猝灭剂/荧光剂对(上文描述了示例性fret对和猝灭剂/荧光剂对);以及ii)第二经标记的检测rna,其包含第二fret对和/或第二猝灭剂/荧光剂对(上文描述了示例性fret对和猝灭剂/荧光剂对)。在一些情况下,第一经标记的检测rna包含至少一个a并且不包含u;而第二经标记的检测rna包含至少一个u并且不包含a。在一些情况下,第一经标记的检测rna包含至少一个u并且不包含a;而第二经标记的检测rna包含至少一个a并且不包含u。第一c2c2蛋白切割第一经标记的检测rna,并且第一可检测信号由第一fret对和/或第一猝灭剂/荧光剂对产生,并且第二c2c2蛋白切割第二经标记的检测rna,并且第二可检测信号由第二fret对和/或第二猝灭剂/荧光剂对产生。检测到第一可检测信号表明样品中存在第一靶rna;并且检测到第二可检测信号表明样品中存在第二靶rna。在一些情况下,检测到的第一信号和第二信号的相对量指示样品中第一靶rna与第二靶rna的比率。
在一些情况下,第一经标记的检测rna包含可与第二经标记的检测rna的标记区分的标记。例如,第一经标记的检测rna可包含第一fret对和/或第一猝灭剂/荧光剂对;并且第二经标记的检测rna可包含第二fret对和/或第二猝灭剂/荧光剂对。作为一个非限制性实例,第一经标记的检测rna可包含含色氨酸的供体和含丹酰的受体;并且第二经标记的检测rna可包含含iaedans的供体和含ddpm的受体。作为另一个非限制性实例,第一经标记的检测rna包含含丹酰的供体和含fitc的受体;并且第二经标记的检测rna包含含cy3的供体和含cy5的受体。在一些情况下,第一经标记的检测rna包含5’fam(荧光素)-3’ibfq(iowa
在一些情况下,将第一和第二经标记的检测rna同时加入样品中(基本上同时接触)。在一些此类情况下,由第一和第二经标记的检测rna产生的信号同时被检测到(基本上同时接触),例如是因为在此类情况下,第一和第二经标记的检测rna可以被区别地标记。
“基本上同时”是指在约5分钟内、约3分钟内、约2分钟内、约1分钟内、约30秒内、约15秒内、约10秒内、约5秒内,或约1秒内。
然而,在一些情况下,由第一和第二经标记的检测rna产生的信号不是同时被检测到的,而是替代地被顺序检测到的(一个在另一个之前)。例如,在一些情况下,第一和第二经标记的检测rna不是同时加入到样品中的,而是替代地顺序加入(例如,可以在第一经标记的检测rna加入之后加入第二经标记的检测rna),并且在一些此类情况下,直到检测到由第一经标记的检测rna产生的信号之后才添加第二经标记的检测rna。因此,在一些情况下,不需要可区分地标记第一和第二经标记的检测rna(例如,它们可以在一些情况下产生相同的可检测信号,例如,可以在相同波长下发荧光),这是因为信号将被顺序地检测到。
作为说明性实例,在一些情况下:(i)没有可区别地标记第一和第二经标记的检测rna;(ii)使样品与一种经标记的检测rna接触,并且检测(例如,测量)由该经标记的检测rna产生的信号;以及(iii)然后使样品与另一种经标记的检测rna接触,并检测由所加入的第二经标记的检测rna产生的信号-因此,当样品中存在两种靶ssrna时,加入第二经标记的检测rna可以导致信号增强(例如,如果信号随着切割增加而增加,例如荧光剂/猝灭剂对)或者可以导致在加入第二经标记的检测rna之后可检测的信号减少(例如,如果信号随着切割增加而减少,例如fret对)。
关于c2c2指导rna结合,第一和第二c2c2蛋白可以彼此正交。在此类情况下,第一c2c2蛋白不与第二c2c2指导rna结合;并且第二c2c2蛋白不与第一指导rna结合。第一c2c2蛋白和第二c2c2蛋白还可以在它们的ssrna切割偏好性方面彼此不同,使得c2c2蛋白中的一种c2c2蛋白在a处切割ssrna而另一种c2c2蛋白在u处切割ssrna。
对c2c2蛋白的正交对的指导可以在图44e中找到。
适用于本公开的方法的c2c2蛋白的正交对的非限制性实例包括下表10中描述的那些。切割偏好性在cas13a蛋白名称后的括号中呈现。例如,“lba(a)”是指lbacas13a蛋白,其在a处切割ssrna;并且“lbu(u)”是指lbucas13a蛋白,其在u处切割ssrna。
表10
第一和第二经标记的检测rna可以各自独立地具有2至100个核糖核苷酸(例如,2至80个、2至60个、2至50个、2至40个、2至30个、2至20个、2至15个或2至10个核糖核苷酸)的长度。第一和第二经标记的检测rna可以各自独立地具有2个核糖核苷酸至100个核糖核苷酸的长度,例如2个核糖核苷酸至5个核糖核苷酸、5个核糖核苷酸至7个核糖核苷酸、7个核糖核苷酸至10个核糖核苷酸、10个核糖核苷酸至15个核糖核苷酸、15个核糖核苷酸至20个核糖核苷酸、20个核糖核苷酸至25个核糖核苷酸、25个核糖核苷酸至30个核糖核苷酸、30个核糖核苷酸至35个核糖核苷酸、35个核糖核苷酸至40个核糖核苷酸、40个核糖核苷酸至45个核糖核苷酸,或45个核糖核苷酸至50个核糖核苷酸的长度。
在一些情况下,第一经标记的检测rna包含至少一个a(例如,至少2个、至少3个或至少4个a)并且缺乏u;并且第二经标记的检测rna包含至少一个u(例如,至少2个、至少3个或至少4个u)并且缺乏a。
在一些情况下,第一经标记的检测rna缺乏u并且包括2至15个连续的a(例如,2至12个、2至10个、2至8个、2至6个、2至4个、3至15个、3至12个、3至10个、3至8个、3至6个、3至5个、4至15个、4至12个、4至10个、4至8个或4至6个连续的a)的段。在一些情况下,第一经标记的检测rna缺乏u并且包括至少2个连续的a(例如,至少3个、至少4个或至少5个连续的a)的段。在一些情况下,第二经标记的检测rna缺乏a并且包括2至15个连续的u(例如,2至12个、2至10个、2至8个、2至6个、2至4个、3至15个、3至12个、3至10个、3至8个、3至6个、3至5个、4至15个、4至12个、4至10个、4至8个或4至6个连续的u)的段。在一些情况下,第二经标记的检测rna缺乏a并且包括至少2个连续的u(例如,至少3个、至少4个或至少5个连续的u)的段。
在一些情况下,第一经标记的检测rna缺乏a并且包括2至15个连续的u(例如,2至12个、2至10个、2至8个、2至6个、2至4个、3至15个、3至12个、3至10个、3至8个、3至6个、3至5个、4至15个、4至12个、4至10个、4至8个或4至6个连续的u)的段。在一些情况下,第一经标记的检测rna缺乏a并且包括至少2个连续的u(例如,至少3个、至少4个或至少5个连续的u)的段。在一些情况下,第二经标记的检测rna缺乏u并且包括2至15个连续的a(例如,2至12个、2至10个、2至8个、2至6个、2至4个、3至15个、3至12个、3至10个、3至8个、3至6个、3至5个、4至15个、4至12个、4至10个、4至8个或4至6个连续的a)的段。在一些情况下,第二经标记的检测rna缺乏u并且包括至少2个连续的a(例如,至少3个、至少4个或至少5个连续的a)的段。
在一些情况下,第一经标记的检测rna包含至少一个u并且缺乏a;并且第二经标记的检测rna包含至少一个a并且缺乏u。
在一些情况下,第一经标记的检测rna包含至少一个a并且缺乏u。例如,在一些情况下,第一经标记的检测rna是同聚腺苷聚合物(聚arna)。作为另一实例,第一经标记的检测rna:i)包含至少一个a;ii)缺乏u;并且iii)包含一个或多个c和/或g。在一些情况下,第二经标记的检测rna包含至少一个u并且缺乏a。例如,在一些情况下,第二经标记的检测rna是同聚尿苷聚合物(聚urna)。作为另一实例,第二经标记的检测rna:i)包含至少一个u;ii)缺乏a;并且iii)包含一个或多个c和/或g。
在一些情况下,第一经标记的检测rna包含至少一个u并且缺乏a。例如,在一些情况下,第一经标记的检测rna是同聚尿苷聚合物(聚urna)。作为另一实例,第二经标记的检测rna:i)包含至少一个u;ii)缺乏a;并且iii)包含一个或多个c和/或g。在一些情况下,第二经标记的检测rna包含至少一个a并且缺乏u。例如,在一些情况下,第二经标记的检测rna是同聚腺苷聚合物(聚arna)。作为另一实例,第二经标记的检测rna:i)包含至少一个a;ii)缺乏u;并且iii)包含一个或多个c和/或g。
如上所述,本公开的方法可以包括使样品与以下接触:第一c2c2蛋白;第二c2c2蛋白;第一c2c2指导rna,其包含与第一单链靶rna杂交的第一核苷酸序列和在本文中也称为‘恒定区’或‘柄’的第二核苷酸序列,所述第二核苷酸序列与第一c2c2蛋白结合;以及第二c2c2指导rna,其包含与第二单链靶rna杂交的第一核苷酸序列和与第二c2c2蛋白结合的第二核苷酸序列(柄)。
例如,在一些情况下,第一c2c2蛋白是cas13a多肽,其包含与图56f中所示的lbacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第一c2c2指导rna包含恒定区(‘柄’-与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列agauagcccaagaaagagggcaauaac(seqidno:16)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt,其中crrna的长度为约25nt、26nt、27nt、28nt、29nt或30nt。在一些情况下,crrna具有核苷酸序列agauagcccaagaaagagggcaauaac(seqidno:16);并且长度为27nt。在一些情况下,第二c2c2蛋白包含与图56k中所示的hhecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列guaacaauccccguagacaggggaacugcaac(seqidno:17)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt。在一些情况下,第二c2c2蛋白包含与图56g中所示的rcacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列caucaccgccaagacgacggcggacugaacc(seqidno:18)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt。在一些情况下,第二c2c2蛋白包含与图56b中所示的pprcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列aauuaucccaaaauugaagggaacuacaac(seqidno:19)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt;其中所述柄的长度为约28nt、29nt、30nt、31nt或32nt。在一些情况下,第二c2c2指导rna包含柄,所述柄包含核苷酸序列aauuaucccaaaauugaagggaacuacaac(seqidno:19);并且所述柄的长度为30nt。在一些情况下,第二c2c2蛋白包含与图56i中所示的lnecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列gaguaccucaaaacaaaagaggacuaaaac(seqidno:20)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt(例如,包含与核苷酸序列gaguaccucaaaacaaaagaggacuaaaac(seqidno:20)仅差异1nt、2nt、3nt、4nt或5nt的核苷酸序列);其中所述柄的长度为约28nt、29nt、30nt、31nt或32nt。在一些情况下,第二指导rna包含柄,所述柄包含核苷酸序列gaguaccucaaaacaaaagaggacuaaaac(seqidno:20);其中所述柄的长度为30nt。在一些情况下,第二c2c2蛋白包含与图56c中所示的lbucas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列gaccaccccaaaaaugaaggggacuaaaaca(seqidno:9)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt(例如,包含与核苷酸序列gaccaccccaaaaaugaaggggacuaaaaca(seqidno:9)仅差异1nt、2nt、3nt、4nt或5nt的核苷酸序列);其中所述柄的长度为约28nt、29nt、30nt、31nt或32nt。在一些情况下,第二指导rna包含柄,所述柄包含核苷酸序列gaccaccccaaaaaugaaggggacuaaaaca(seqidno:9);其中所述柄的长度为31nt。在一些情况下,第二c2c2蛋白包含与图56e中所示的lwacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列gaccaccccaauaucgaaggggacuaaaacuu(seqidno:21)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt(例如,包含与核苷酸序列gaccaccccaauaucgaaggggacuaaaacuu(seqidno:21)仅差异1nt、2nt、3nt、4nt或5nt的核苷酸序列);其中所述柄的长度为约28nt、29nt、30nt、31nt或32nt。在一些情况下,第二指导rna包含柄,所述柄包含核苷酸序列gaccaccccaauaucgaaggggacuaaaacuu(seqidno:21);其中所述柄的长度为32nt。在一些情况下,第二c2c2蛋白包含与图56d中所示的lshcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列caccccaauaucgaaggggacuaaaac(seqidno:22)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt(例如,包含与核苷酸序列caccccaauaucgaaggggacuaaaac(seqidno:22)仅差异1nt、2nt、3nt、4nt或5nt的核苷酸序列);其中所述柄的长度为25nt、26nt、27nt、28nt或29nt。在一些情况下,第二指导rna包含柄,所述柄包含核苷酸序列caccccaauaucgaaggggacuaaaac(seqidno:22);其中所述柄的长度为27nt。
作为另一实例,在一些情况下,第一c2c2蛋白是cas13a多肽,其包含与图56j中所示的erecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第一c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列aaguagcccgauauagagggcaauaac(seqidno:23)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt,其中所述柄的长度为约25nt、26nt、27nt、28nt、29nt或30nt。在一些情况下,所述柄具有核苷酸序列aaguagcccgauauagagggcaauaac(seqidno:23);并且长度为27nt。在一些情况下,第一c2c2蛋白是cas13a多肽,其包含与图56j中所示的erecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第一c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列auacagcucgauauagugagcaauaag(seqidno:24)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt,其中所述柄的长度为约25nt、26nt、27nt、28nt、29nt或30nt。在一些情况下,所述柄具有核苷酸序列auacagcucgauauagugagcaauaag(seqidno:24);并且长度为27nt。在一些情况下,第二c2c2蛋白包含与图56k中所示的hhecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列guaacaauccccguagacaggggaacugcaac(seqidno:17)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt;其中所述柄的长度为约30nt、31nt、32nt、33nt、34nt或35nt。在一些情况下,第二c2c2指导rna包含柄,所述柄包含核苷酸序列guaacaauccccguagacaggggaacugcaac(seqidno:17);并且所述柄的长度为32nt。在一些情况下,第二c2c2蛋白包含与图56g中所示的rcacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列ucacaucaccgccaagacgacggcggacugaacc(seqidno:25)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt;其中所述柄的长度为约32nt、33nt、34nt、35nt、36nt或37nt。在一些情况下,第二c2c2指导rna包含柄,所述柄包含核苷酸序列ucacaucaccgccaagacgacggcggacugaacc(seqidno:25);并且所述柄的长度为34nt。在一些情况下,第二c2c2蛋白包含与图56b中所示的pprcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列aauuaucccaaaauugaagggaacuacaac(seqidno:19)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt;其中所述柄的长度为约28nt、29nt、30nt、31nt或32nt。在一些情况下,第二c2c2指导rna包含柄,所述柄包含核苷酸序列aauuaucccaaaauugaagggaacuacaac(seqidno:19);并且所述柄的长度为30nt。在一些情况下,第二c2c2蛋白包含与图56i中所示的lnecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列gaguaccucaaaacaaaagaggacuaaaac(seqidno:20)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt(例如,包含与核苷酸序列gaguaccucaaaacaaaagaggacuaaaac(seqidno:20)仅差异1nt、2nt、3nt、4nt或5nt的核苷酸序列);其中所述柄的长度为约28nt、29nt、30nt、31nt或32nt。在一些情况下,第二指导rna包含柄,所述柄包含核苷酸序列gaguaccucaaaacaaaagaggacuaaaac(seqidno:20);其中所述柄的长度为30nt。在一些情况下,第二c2c2蛋白包含与图56c中所示的lbucas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列gaccaccccaaaaaugaaggggacuaaaaca(seqidno:9)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt(例如,包含与核苷酸序列gaccaccccaaaaaugaaggggacuaaaaca(seqidno:9)仅差异1nt、2nt、3nt、4nt或5nt的核苷酸序列);其中所述柄的长度为约28nt、29nt、30nt、31nt或32nt。在一些情况下,第二指导rna包含柄,所述柄包含核苷酸序列gaccaccccaaaaaugaaggggacuaaaaca(seqidno:9);其中所述柄的长度为31nt。在一些情况下,第二c2c2蛋白包含与图56e中所示的lwacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列gaccaccccaauaucgaaggggacuaaaacuu(seqidno:21)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt(例如,包含与核苷酸序列gaccaccccaauaucgaaggggacuaaaacuu(seqidno:21)仅差异1nt、2nt、3nt、4nt或5nt的核苷酸序列);其中所述柄的长度为约28nt、29nt、30nt、31nt或32nt。在一些情况下,第二指导rna包含柄,所述柄包含核苷酸序列gaccaccccaauaucgaaggggacuaaaacuu(seqidno:21);其中所述柄的长度为32nt。在一些情况下,第二c2c2蛋白包含与图56d中所示的lshcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列caccccaauaucgaaggggacuaaaac(seqidno:22)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt(例如,包含与核苷酸序列caccccaauaucgaaggggacuaaaac(seqidno:22)仅差异1nt、2nt、3nt、4nt或5nt的核苷酸序列);其中所述柄的长度为25nt、26nt、27nt、28nt或29nt。在一些情况下,第二指导rna包含柄,所述柄包含核苷酸序列caccccaauaucgaaggggacuaaaac(seqidno:22);其中所述柄的长度为27nt。在一些情况下,第二c2c2蛋白包含与图56a中所示的lsecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列gacuaccucuauaugaaagaggacuaaaac(seqidno:7)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt;其中所述柄的长度为约28nt、29nt、30nt、31nt或32nt。在一些情况下,第二c2c2指导rna包含柄,所述柄包含核苷酸序列gacuaccucuauaugaaagaggacuaaaac(seqidno:7);并且所述柄的长度为30nt。
作为另一实例,在一些情况下,第一c2c2蛋白是cas13a多肽,其包含与图56h中所示的camcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第一c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列gaacagcccgauauagagggcaauagac(seqidno:26)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt,其中所述柄的长度为约26nt、27nt、28nt、29nt或30nt。在一些情况下,所述柄具有核苷酸序列gaacagcccgauauagagggcaauagac(seqidno:26);并且长度为28nt。在一些情况下,第二c2c2蛋白包含与图56k中所示的hhecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列guaacaauccccguagacaggggaacugcaac(seqidno:17)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt;其中所述柄的长度为约30nt、31nt、32nt、33nt、34nt,或35nt。在一些情况下,第二c2c2指导rna包含柄,所述柄包含核苷酸序列guaacaauccccguagacaggggaacugcaac(seqidno:17);并且所述柄的长度为32nt。在一些情况下,第二c2c2蛋白包含与图56g中所示的rcacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列ucacaucaccgccaagacgacggcggacugaacc(seqidno:25)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt;其中所述柄的长度为约32nt、33nt、34nt、35nt、36nt或37nt。在一些情况下,第二c2c2指导rna包含柄,所述柄包含核苷酸序列ucacaucaccgccaagacgacggcggacugaacc(seqidno:25);并且所述柄的长度为34nt。在一些情况下,第二c2c2蛋白包含与图56b中所示的pprcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列aauuaucccaaaauugaagggaacuacaac(seqidno:19)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt;其中所述柄的长度为约28nt、29nt、30nt、31nt或32nt。在一些情况下,第二c2c2指导rna包含柄,所述柄包含核苷酸序列aauuaucccaaaauugaagggaacuacaac(seqidno:19);并且所述柄的长度为30nt。在一些情况下,第二c2c2蛋白包含与图56i中所示的lnecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列gaguaccucaaaacaaaagaggacuaaaac(seqidno:20)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt(例如,包含与核苷酸序列gaguaccucaaaacaaaagaggacuaaaac(seqidno:20)仅差异1nt、2nt、3nt、4nt或5nt的核苷酸序列);其中所述柄的长度为约28nt、29nt、30nt、31nt或32nt。在一些情况下,第二指导rna包含柄,所述柄包含核苷酸序列gaguaccucaaaacaaaagaggacuaaaac(seqidno:20);其中所述柄的长度为30nt。在一些情况下,第二c2c2蛋白包含与图56c中所示的lbucas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列gaccaccccaaaaaugaaggggacuaaaaca(seqidno:9)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt(例如,包含与核苷酸序列gaccaccccaaaaaugaaggggacuaaaaca(seqidno:9)仅差异1nt、2nt、3nt、4nt或5nt的核苷酸序列);其中所述柄的长度为约28nt、29nt、30nt、31nt或32nt。在一些情况下,第二指导rna包含柄,所述柄包含核苷酸序列gaccaccccaaaaaugaaggggacuaaaaca(seqidno:9);其中所述柄的长度为31nt。在一些情况下,第二c2c2蛋白包含与图56e中所示的lwacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列gaccaccccaauaucgaaggggacuaaaacuu(seqidno:21)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt(例如,包含与核苷酸序列gaccaccccaauaucgaaggggacuaaaacuu(seqidno:21)仅差异1nt、2nt、3nt、4nt或5nt的核苷酸序列);其中所述柄的长度为约28nt、29nt、30nt、31nt或32nt。在一些情况下,第二指导rna包含柄,所述柄包含核苷酸序列gaccaccccaauaucgaaggggacuaaaacuu(seqidno:21);其中所述柄的长度为32nt。在一些情况下,第二c2c2蛋白包含与图56d中所示的lshcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列caccccaauaucgaaggggacuaaaac(seqidno:22)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt(例如,包含与核苷酸序列caccccaauaucgaaggggacuaaaac(seqidno:22)仅差异1nt、2nt、3nt、4nt或5nt的核苷酸序列);其中所述柄的长度为25nt、26nt、27nt、28nt或29nt。在一些情况下,第二指导rna包含柄,所述柄包含核苷酸序列caccccaauaucgaaggggacuaaaac(seqidno:22);其中所述柄的长度为27nt。在一些情况下,第二c2c2蛋白包含与图56a中所示的lsecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2指导rna包含柄(与cas13a多肽结合的一段核苷酸),其包含这样的核苷酸序列,所述核苷酸序列与核苷酸序列gacuaccucuauaugaaagaggacuaaaac(seqidno:7)的差异不超过1个核苷酸(nt)、不超过2nt、不超过3nt、不超过4nt或不超过5nt;其中所述柄的长度为约28nt、29nt、30nt、31nt或32nt。在一些情况下,第二c2c2指导rna包含柄,所述柄包含核苷酸序列gacuaccucuauaugaaagaggacuaaaac(seqidno:7);并且所述柄的长度为30nt。
混合检测
如上所述,在一些情况下,本公开的方法包括:a)使样品(例如,包含靶rna和多种非靶rna的样品)与以下接触:i)前体c2c2指导rna阵列,其包含两个或更多个c2c2指导rna,每个所述c2c2指导rna具有不同的指导序列;和(ii)c2c2蛋白,其将前体c2c2指导rna阵列切割成单独的c2c2指导rna,并且还切割样品的rna;以及b)测量由c2c2蛋白介导的rna切割产生的可检测信号。
在一些情况下,阵列(前体c2c2指导rna阵列)上可以存在两个或更多个c2c2指导rna。c2c2蛋白可以将前体c2c2指导rna阵列切割成单独的c2c2指导rna(例如,参见图4和图6)。
在一些情况下,主题c2c2指导rna阵列包括2个或更多个c2c2指导rna(例如,3个或更多个、4个或更多个、5个或更多个、6个或更多个、或7个或更多个c2c2指导rna)。给定阵列的c2c2指导rna可以靶向相同靶rna的不同靶位点(即,可以包括与所述不同靶位点杂交的指导序列)(例如,这可以增加检测的灵敏度)和/或可以靶向不同的靶rna分子(例如,例如基于诸如单核苷酸多态性、单核苷酸多态性(snp)等等之类的变异的转录物家族等可用于例如检测多种病毒株,诸如流感病毒变体、寨卡病毒变体、hiv变体等)。
c2c2蛋白
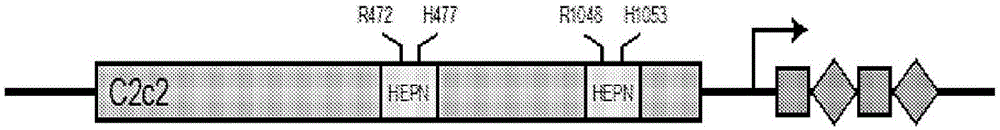
c2c2蛋白与c2c2指导rna结合,由指导rna(其与靶rna杂交)引导到单链靶rna,从而被“激活”。如果c2c2蛋白的hepn1结构域和hepn2结构域是完整的,则c2c2蛋白一旦被激活就切割靶rna,但是也切割非靶rna。
示例性的天然存在c2c2蛋白描绘于图8中,并且如seqidno:1至seqidno:6所示。在一些情况下,主题c2c2蛋白包括与seqidno:1至seqidno:6中任一项所示的氨基酸序列具有80%或更多(例如,85%或更多、90%或更多、95%或更多、98%或更多、99%或更多、99.5%或更多、或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,合适的c2c2多肽包含与seqidno:1所示的斯氏李斯特菌(listeriaseeligeri)c2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,合适的c2c2多肽包含与seqidno:2所示的口腔纤毛菌(leptotrichiabuccalis)c2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,合适的c2c2多肽包含与seqidno:4所示的荚膜红细菌(rhodobactercapsulatus)c2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,合适的c2c2多肽包含与seqidno:5所示的鸡肉杆菌(carnobacteriumgallinarum)c2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,合适的c2c2多肽包含与seqidno:6所示的herbinixhemicellulosilyticac2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,c2c2蛋白包括与seqidno:2所示的口腔纤毛菌(lbu)c2c2氨基酸序列具有80%或更多的氨基酸序列同一性的氨基酸序列。在一些情况下,c2c2蛋白是口腔纤毛菌(lbu)c2c2蛋白(例如,参见seqidno:2)。在一些情况下,c2c2蛋白包括seqidno:1至seqidno:2和seqidno:4至seqidno:6中任一项所示的氨基酸序列。
在一些情况下,在本公开的方法中使用的c2c2蛋白不是沙氏纤毛菌(leptotrichiashahii)(lsh)c2c2蛋白。在一些情况下,在本公开的方法中使用的c2c2蛋白不是与seqidno:3所示的lshc2c2多肽具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的c2c2多肽。
在一些情况下,所述c2c2蛋白比沙氏纤毛菌(lsh)c2c2蛋白在切割未由所述方法的c2c2指导rna靶向的rna方面更有效1.2倍或更多倍。在一些情况下,所述c2c2蛋白比沙氏纤毛菌(lsh)c2c2蛋白在切割未由所述方法的c2c2指导rna靶向的rna方面更有效1.5倍或更多倍。在一些情况下,在本公开的方法中使用的c2c2多肽当被激活时,比lshc2c2至少1.2倍、至少1.5倍、至少2倍、至少2.5倍、至少3倍、至少4倍、至少5倍、至少6倍、至少7倍、至少8倍、至少9倍、至少10倍、至少15倍、至少20倍、至少30倍或大于30倍有效地切割非靶rna。
在一些情况下,c2c2蛋白在所述接触的1小时内表现出至少50%的rna切割效率(例如,55%或更多、60%或更多、65%或更多、70%或更多、或75%或更多的切割效率)。在一些情况下,c2c2蛋白在所述接触的40分钟内表现出至少50%的rna切割效率(例如,55%或更多、60%或更多、65%或更多、70%或更多、或75%或更多的切割效率)。在一些情况下,c2c2蛋白在所述接触的30分钟内表现出至少50%的rna切割效率(例如,55%或更多、60%或更多、65%或更多、70%或更多、或75%或更多的切割效率)。
在一些情况下,适用于本公开的方法的c2c2蛋白在30秒至60分钟(例如,1分钟至60分钟、30秒至5分钟、1分钟至5分钟、1分钟至10分钟、5分钟至10分钟、10分钟至15分钟、15分钟至20分钟、20分钟至25分钟、25分钟至30分钟、30分钟至35分钟、35分钟至40分钟、40分钟至45分钟、45分钟至50分钟、50分钟至55分钟或55分钟至60分钟)的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适用于本公开的方法的c2c2蛋白在30秒至5分钟的时间段内(例如,1分钟至5分钟,例如在1分钟、2分钟、3分钟、4分钟或5分钟的时间段内)切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适用于本公开的方法的c2c2蛋白在5分钟至10分钟的时间段内(例如,在5分钟、6分钟、7分钟、8分钟、9分钟或10分钟的时间段内)切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适用于本公开的方法的c2c2蛋白在10分钟至15分钟(例如,10分钟、11分钟、12分钟、13分钟、14分钟或15分钟)的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适用于本公开的方法的c2c2蛋白在15分钟至20分钟的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适用于本公开的方法的c2c2蛋白在20分钟至25分钟的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适用于本公开的方法的c2c2蛋白在25分钟至30分钟的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适用于本公开的方法的c2c2蛋白在30分钟至35分钟的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适用于本公开的方法的c2c2蛋白在35分钟至40分钟的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适用于本公开的方法的c2c2蛋白在40分钟至45分钟的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适用于本公开的方法的c2c2蛋白在45分钟至50分钟的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适用于本公开的方法的c2c2蛋白在50分钟至55分钟的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适用于本公开的方法的c2c2蛋白在55分钟至60分钟的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适用于本公开的方法的c2c2蛋白在小于1分钟的时间段内(例如,在50秒至59秒、40秒至49秒、30秒至39秒或20秒至29秒的时间段内)切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,切割在生理条件下发生。在一些情况下,切割在15℃至20℃、20℃至25℃、25℃至30℃、30℃至35℃、或35℃至40℃的温度下发生。在一些情况下,切割在约37℃发生。在一些情况下,切割在约37℃发生并且反应条件包括二价金属离子。在一些情况下,二价金属离子是mg2+。在一些情况下,二价金属离子是mn2+。在一些情况下,反应条件的ph在ph5与ph6之间。在一些情况下,反应条件的ph在ph6与ph7之间。在一些情况下,反应条件的ph在ph6.5与ph7.5之间。在某些情况下,反应条件的ph高于ph7.5。
术语“切割效率”在本文中用于指c2c2蛋白的以下能力:一旦c2c2蛋白已经通过适当的c2c2指导rna/靶rna杂交而被激活,就快速切割样品中的rna。“切割效率”是指蛋白在给定时间段内可以切割的rna的量。例如,50%的切割效率应表明50%的给定rna在指定时间段内被切割。例如,如果样品中的rna以100μm的起始浓度存在,则当已经切割了50μm的rna时,已经实现了50%的切割。作为另一实例,如果样品中存在多个rna分子,则当已经切割了50%的rna分子时,已经实现了50%的切割;效率是切割一定百分比的总rna所需的时间量的表示。这可以通过任何合宜的方法测量,并且许多此类方法对于本领域的普通技术人员来说将是已知的。例如,可以使用经标记的检测rna。在一些情况下,rna种类(切割的对比未切割的)可以在凝胶上分离,并且可以将切割的rna的量与未切割的rna的量进行比较,例如参见图3。
当使用短语“其中c2c2蛋白切割样品中存在的rna的至少x%”(例如,在指定时间段内)时,意指样品中存在的‘信号产生’rna的x%在指定时间段内被切割。哪种rna是‘信号产生’rna可以取决于所用的检测方法。例如,当使用经标记的检测rna时,所述经标记的检测rna可能是唯一的‘信号产生rna'。然而,经标记的检测rna用于代表样品的rna,并且因此关于经标记的检测rna所观察到的情况被假定为代表样品的非靶rna发生的情况。因此,当50%的经标记的检测rna被切割时,这通常将被假设为表示当‘样品中存在的rna'的50%被切割时。在一些情况下,通常测量rna切割,因此样品的所有可切割rna都是“信号产生rna”。因此,当提及样品中存在的rna的被切割百分比时,可以使用任何合宜的方法测量该值,并且无论使用何种方法,该值通常在本文中意指当酶已经切割样品中的可切割靶的一半。
在一些情况下,c2c2蛋白不是沙氏纤毛菌(lsh)c2c2蛋白。在一些情况下,c2c2蛋白比沙氏纤毛菌(lsh)c2c2蛋白更有效(例如,在切割非靶rna方面)1.2倍或更高(例如,1.5倍或更高、1.7倍或更高、或2倍或更高)。因此,在一些情况下,主题c2c2蛋白比沙氏纤毛菌(lsh)c2c2蛋白在切割未由所述方法的c2c2指导rna靶向的rna方面更有效1.2倍或更多倍(例如,1.5倍或更多倍、1.7倍或更多倍、或2倍或更多倍)。在一些情况下,在本公开的方法中使用的c2c2多肽当被激活时,比lshc2c2至少1.2倍、至少1.5倍、至少2倍、至少2.5倍、至少3倍、至少4倍、至少5倍、至少6倍、至少7倍、至少8倍、至少9倍、至少10倍、至少15倍、至少20倍、至少30倍或大于30倍有效地切割非靶rna。
变体c2c2多肽
变体c2c2多肽包括seqidno:1、seqidno:2和seqidno:4至seqidno:6中任一者的变体,其中变体c2c2多肽表现出降低的(或不可检测的)核酸酶活性。例如,在一些情况下,变体c2c2蛋白缺乏催化活性的hepn1结构域。作为另一个实例,变体c2c2蛋白缺乏催化活性的hepn2结构域。在一些情况下,变体c2c2蛋白缺乏催化活性的hepn1结构域并且缺乏催化活性的hepn2结构域。
在一些情况下,变体c2c2多肽包含对seqidno:2(口腔纤毛菌c2c2)所示的氨基酸序列的氨基酸r472、h477、r1048和h1053中的1个、2个、3个或4个氨基酸的氨基酸取代,或seqidno:1、seqidno:4、seqidno:5或seqidno:6中所示的c2c2氨基酸序列的相应氨基酸的氨基酸取代。seqidno:1、seqidno:4、seqidno:5和seqidno:6中的相应氨基酸容易地被鉴别;参见例如图22b。例如,seqidno:1(斯氏李斯特菌c2c2)中对应于seqidno:2的r472、h477、r1048和h1053的氨基位置分别是r445、h450、r1016和h1021。作为另一实例,seqidno:4(荚膜红细菌c2c2)中对应于seqidno:2的r472、h477、r1048和h1053的氨基酸位置分别是r464、h469、r1052和h1057。作为另一实例,seqidno:5(鸡肉杆菌c2c2)中对应于seqidno:2的r472、h477、r1048和h1053的氨基酸位置分别是r467、h472、r1069和h1074。作为另一实例,seqidno:6(herbinixhemicellulosilyticac2c2)中对应于seqidno:2的r472、h477、r1048和h1053的氨基酸位置分别是r472、h477、r1044和h1049。
在一些情况下,变体c2c2多肽包含对seqidno:2所示的氨基酸序列的氨基酸r472和h477的氨基酸取代,或对seqidno:1、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含对seqidno:2所示的氨基酸序列的氨基酸r1048和h1053的氨基酸取代,或对seqidno:1、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含对seqidno:2所示的氨基酸序列的氨基酸r472、h477、r1048和h1053的氨基酸取代,或对seqidno:1、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列的相应氨基酸的氨基酸取代。
在一些情况下,变体c2c2多肽包含与seqidno:2所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472和h477的取代。在一些情况下,位置472处的氨基酸是除了arg以外的任何氨基酸;并且位置477处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r472a和h477a。在一些情况下,变体c2c2多肽包含与seqidno:2所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1048和h1053的取代。在一些情况下,位置1048处的氨基酸是除了arg以外的任何氨基酸;并且位置1053处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1048a和h1053a。在一些情况下,变体c2c2多肽包含与seqidno:2所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472、h477、r1048和h1053的取代。在一些情况下,位置472和1048处的氨基酸是除了arg以外的任何氨基酸;并且位置477和1053处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r472a、h477a、r1048a和h1053a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
在一些情况下,变体c2c2多肽包含与seqidno:1所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r445和h450的取代。在一些情况下,位置445处的氨基酸是除了arg以外的任何氨基酸;并且位置450处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r445a和h450a。在一些情况下,变体c2c2多肽包含与seqidno:1所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1016和h1021的取代。在一些情况下,位置1016处的氨基酸是除了arg以外的任何氨基酸;并且位置1021处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1016a和h1021a。在一些情况下,变体c2c2多肽包含与seqidno:1所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r445、h450、r1016和h1021的取代。在一些情况下,位置445和1016处的氨基酸是除了arg以外的任何氨基酸;并且位置450和1016处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r445a、h450a、r1016a和h1021a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
在一些情况下,变体c2c2多肽包含与seqidno:4(荚膜红细菌c2c2)所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r464和h469的取代。在一些情况下,位置464处的氨基酸是除了arg以外的任何氨基酸;并且位置469处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r464a和h469a。在一些情况下,变体c2c2多肽包含与seqidno:4所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1052和h1057的取代。在一些情况下,位置1052处的氨基酸是除了arg以外的任何氨基酸;并且位置1057处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1052a和h1057a。在一些情况下,变体c2c2多肽包含与seqidno:4所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r464、h469、r1052和h1057的取代。在一些情况下,位置464和1052处的氨基酸是除了arg以外的任何氨基酸;并且位置469和1057处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r464a、h469a、r1052a和h1057a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
在一些情况下,变体c2c2多肽包含与seqidno:5(鸡肉杆菌c2c2)所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r467和h472的取代。在一些情况下,位置467处的氨基酸是除了arg以外的任何氨基酸;并且位置472处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r469a和h472a。在一些情况下,变体c2c2多肽包含与seqidno:5所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1069和h1074的取代。在一些情况下,位置1069处的氨基酸是除了arg以外的任何氨基酸;并且位置1074处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1069a和h1074a。在一些情况下,变体c2c2多肽包含与seqidno:5所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r467、h472、r1069和h1074的取代。在一些情况下,位置467和1069处的氨基酸是除了arg以外的任何氨基酸;并且位置472和1074处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r469a、h472a、r1069a和h1074a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
在一些情况下,变体c2c2多肽包含与seqidno:6(herbinixhemicellulosilyticac2c2)所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472和h477的取代。在一些情况下,位置472处的氨基酸是除了arg以外的任何氨基酸;并且位置477处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r472a和h477a。在一些情况下,变体c2c2多肽包含与seqidno:6所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1044和h1049的取代。在一些情况下,位置1044处的氨基酸是除了arg以外的任何氨基酸;并且位置1049处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1044a和h1049a。在一些情况下,变体c2c2多肽包含与seqidno:6所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472、h477、r1044和h1049的取代。在一些情况下,位置472和1044处的氨基酸是除了arg以外的任何氨基酸;并且位置477和1049处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r472a、h477a、r1044a和h1049a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
在一些情况下,变体c2c2多肽包含与seqidno:2所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472、h477、r1048和h1053中的1个、2个、3个或4个氨基酸的取代,使得变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并且保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。例如,在一些情况下,变体c2c2多肽表现出由具有seqidno:2所示氨基酸序列的c2c2多肽表现出的rna指导的非靶rna切割的小于50%、小于40%、小于30%、小于20%、小于10%、小于5%、小于1%或小于0.1%。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
上述变体c2c2多肽中的任一者还可以包括突变(例如,在位置r1079、r1072和k1082中的任一者处,如下文进一步详细描述的),所述突变导致降低的切割前体c2c2指导rna的能力(例如,能力丧失)。
在一些情况下,变体c2c2多肽具有降低的切割前体c2c2指导rna的能力(例如,参见下面的实施例和相关的图26c至图26d、图35d和图37)。例如,在一些情况下,变体c2c2多肽包含对seqidno:2(口腔纤毛菌c2c2)所示的氨基酸序列的氨基酸r1079、r1072和k1082中的1个、2个或3个氨基酸的氨基酸取代,或者对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。seqidno:1、seqidno:4、seqidno:5和seqidno:6中的相应氨基酸容易被鉴别。例如,seqidno:1(斯氏李斯特菌c2c2)中对应于seqidno:2的r1079、r1072和k1082和k1082的氨基位置分别是r1048、r1041和k1051。作为另一实例,seqidno:4(荚膜红细菌c2c2)中对应于seqidno:2的r1079、r1072和k1082的氨基酸位置分别是r1085、r1078和k1088。作为另一实例,seqidno:5(鸡肉杆菌c2c2)中对应于seqidno:2的r1079、r1072和k1082的氨基酸位置分别是r1099、r1092和k1102。作为另一实例,seqidno:6(herbinixhemicellulosilyticac2c2)中对应于seqidno:2的r1079和r1072的氨基酸位置分别是r1172和r1165。
在一些情况下,变体c2c2多肽包含对seqidno:2所示的氨基酸序列的氨基酸r1079(例如,r1079a)的氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含对seqidno:2所示的氨基酸序列的氨基酸r1072(例如,r1072a)的氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含对seqidno:2所示的氨基酸序列的氨基酸k1082(例如,k1082a)的氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含在seqidno:2所示的氨基酸序列的选自r1079(例如,r1079a)、r1072(例如,r1072a)和k1082(例如,k1082a)的位置处的一个或多个(例如,两个或更多个、或全部三个)氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。
在一些情况下,变体c2c2多肽包含与seqidno:2所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对seqidno:2所示的氨基酸序列的氨基酸r1079(例如,r1079a)的氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含与seqidno:2所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对seqidno:2所示的氨基酸序列的氨基酸r1072(例如,r1072a)的氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含与seqidno:2所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对seqidno:2所示的氨基酸序列的氨基酸k1082(例如,k1082a)的氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含与seqidno:2所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含在seqidno:2所示的氨基酸序列的选自r1079(例如,r1079a)、r1072(例如,r1072a)和k1082(例如,k1082a)的位置处的一个或多个(例如,两个或更多个、或全部三个)氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。
在一些情况下,变体c2c2多肽包含与seqidno:1所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对seqidno:1所示的氨基酸序列的氨基酸r1041(例如,r1041a)的氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:2、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含与seqidno:1所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对seqidno:1所示的氨基酸序列的氨基酸r1048(例如,r1048a)的氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:2、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含与seqidno:1所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对seqidno:1所示的氨基酸序列的氨基酸k1051(例如,k1051a)的氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:2、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含与seqidno:1所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含在seqidno:1所示的氨基酸序列的选自r1048(例如,r1048a)、r1041(例如,r1041a)和k1051(例如,k1051a)的位置处的一个或多个(例如,两个或更多个、或全部三个)氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:2、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。
在一些情况下,变体c2c2多肽包含与seqidno:4所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对seqidno:4所示的氨基酸序列的氨基酸r1085(例如,r1085a)的氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:2、seqidno:5或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含与seqidno:4所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对seqidno:4所示的氨基酸序列的氨基酸r1078(例如,r1078a)的氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:2、seqidno:5或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含与seqidno:4所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对seqidno:4所示的氨基酸序列的氨基酸k1088(例如,k1088a)的氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:2、seqidno:5或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含与seqidno:4所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含在seqidno:4所示的氨基酸序列的选自r1085(例如,r1085a)、r1078(例如,r1078a)和k1088(例如,k1088a)的位置处的一个或多个(例如,两个或更多个、或全部三个)氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:2、seqidno:5或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。
在一些情况下,变体c2c2多肽包含与seqidno:5所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对seqidno:5所示的氨基酸序列的氨基酸r1099(例如,r1099a)的氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:4、seqidno:2或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含与seqidno:5所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对seqidno:5所示的氨基酸序列的氨基酸r1092(例如,r1092a)的氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:4、seqidno:2或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含与seqidno:5所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对seqidno:5所示的氨基酸序列的氨基酸k1102(例如,k1102a)的氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:4、seqidno:2或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含与seqidno:5所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含在seqidno:5所示的氨基酸序列的选自r1099(例如,r1099a)、r1092(例如,r1092a)和k1102(例如,k1102a)的位置处的一个或多个(例如,两个或更多个、或全部三个)氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:4、seqidno:2或seqidno:6所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。
在一些情况下,变体c2c2多肽包含与seqidno:6所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对seqidno:6所示的氨基酸序列的氨基酸r1172(例如,r1172a)的氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:4、seqidno:5或seqidno:2所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含与seqidno:6所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对seqidno:6所示的氨基酸序列的氨基酸r1165(例如,r1165a)的氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:4、seqidno:5或seqidno:2所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含与seqidno:6所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含在seqidno:6所示的氨基酸序列的选自r1172(例如,r1172a)和r1165(例如,r1165a)的位置处的一个或多个(例如,两个)氨基酸取代,或对任何c2c2氨基酸序列(例如,seqidno:1、seqidno:4、seqidno:5或seqidno:2所示的c2c2氨基酸序列)的相应氨基酸的氨基酸取代。
c2c2指导rna
主题c2c2指导rna(例如,c2c2crrna)包括指导序列和恒定区(例如,指导序列的5'端的区域)。指导序列的5'端的区域与c2c2蛋白结合(并且可以被认为是蛋白结合区),而指导序列与靶rna的靶序列杂交。
指导序列
指导序列与单链靶rna的靶序列具有互补性(与单链靶rna的靶序列杂交)。在一些情况下,直接位于靶序列(原型间隔区)3'端的靶rna的碱基不是g。在一些情况下,指导序列的长度为16-28个核苷酸(nt)(例如,长度为16-26nt、16-24nt、16-22nt、16-20nt、16-18nt、17-26nt、17-24nt、17-22nt、17-20nt、17-18nt、18-26nt、18-24或18-22nt)。在一些情况下,指导序列的长度为18-24个核苷酸(nt)。在一些情况下,指导序列的长度为至少16nt(例如,长度为至少18nt、20nt或22nt)。在一些情况下,指导序列的长度为至少17nt。在一些情况下,指导序列的长度为至少18nt。在一些情况下,指导序列的长度为至少20nt。
在一些情况下,指导序列与单链靶rna的靶序列具有80%或更多(例如,85%或更多、90%或更多、95%或更多、或100%)的互补性。在一些情况下,指导序列与单链靶rna的靶序列100%互补。
恒定区
以下3个序列各自是天然存在的c2c2指导rna的恒定区的实例(例如,在指导序列的5'端的区域):
gacuaccucuauaugaaagaggacuaaaac(seqidno:7)
(斯氏李斯特菌)(“lse”)
ccaccccaauaucgaaggggacuaaaaca(seqidno:8)
(沙氏纤毛菌)(“lsh”)
gaccaccccaaaaaugaaggggacuaaaaca(seqidno:9)
(口腔纤毛菌)(“lbu”)
在一些实施方案中,主题c2c2指导rna包括与seqidno:7至seqidno:9中任一项所示的序列具有70%或更多同一性(例如,80%或更多、85%或更多、90%或更多、95%或更多、98%或更多、99%或更多、或100%同一性)的核苷酸序列。在一些实施方案中,主题c2c2指导rna包括与seqidno:7至seqidno:9中任一项所示的序列具有90%或更多同一性(例如,95%或更多、98%或更多、99%或更多、或100%同一性)的核苷酸序列。在一些实施方案中,主题c2c2指导rna包含seqidno:7至seqidno:9中任一者所示的核苷酸序列。
在一些实施方案中,主题c2c2指导rna包括与seqidno:9所示的序列具有70%或更多同一性(例如,80%或更多、85%或更多、90%或更多、95%或更多、98%或更多、99%或更多、或100%同一性)的核苷酸序列。在一些实施方案中,主题c2c2指导rna包括与seqidno:9中所示的序列具有90%或更多同一性(例如,95%或更多、98%或更多、99%或更多、或100%同一性)的核苷酸序列。在一些实施方案中,主题c2c2指导rna包括seqidno:9所示的核苷酸序列。
在一些实施方案中,主题c2c2指导rna不包括沙氏纤毛菌(lsh)c2c2指导rna的核苷酸序列。例如,在一些情况下,所使用的c2c2蛋白不是来自沙氏纤毛菌的c2c2(例如,不是lshc2c2蛋白),并且在一些此类情况下,所使用的c2c2指导rna也不来自沙氏纤毛菌(例如,所使用的指导rna不包括lshc2c2指导rna的恒定区)。因此,在一些情况下,主题c2c2指导rna不包括seqidno:8所示的序列。
在一些情况下,c2c2指导rna包括双链rna双链体(dsrna双链体)。例如,参见图7a,其示出了来自lbu的c2c2指导rna与单链靶rna杂交,其中所述c2c2指导rna包括长度为4个碱基对(bp)的dsrna双链体。在一些情况下,c2c2指导rna包括长度为2至12bp(例如,2至10bp、2至8bp、2至6bp、2至5bp、2至4bp、3至12bp、3至10bp、3至8bp、3至6bp、3至5bp、3至4bp、4至12bp、4至10bp、4至8bp、4至6bp或4至5bp)的dsrna双链体。在一些情况下,c2c2指导rna包括长度为2bp或更长(例如,长度为3bp或更长、4bp或更长、5bp或更长、6bp或更长、或7bp或更长)的dsrna双链体。在一些情况下,c2c2指导rna包括比相应的野生型c2c2指导rna的dsrna双链体更长的dsrna双链体。例如,参见图7a,其示出了来自lbu的c2c2指导rna与单链靶rna杂交,其中所述c2c2指导rna包括长度为4个碱基对(bp)的dsrna双链体。因此,c2c2指导rna可以在一些情况下包括长度为5bp或更长(例如,长度为6bp或更长、7bp或更长、或8bp或更长)的dsrna双链体。在一些情况下,c2c2指导rna包括比相应的野生型c2c2指导rna的dsrna双链体更短的dsrna双链体。因此在一些情况下,c2c2指导rna包括长度小于4bp的dsrna双链体。在一些情况下,c2c2指导rna包括长度为2或3bp的dsrna双链体。
在一些情况下,c2c2指导rna的在指导序列的5'端的区域的长度为15个或更多个核苷酸(nt)(例如,长度为18nt或更长、20nt或更长、21nt或更长、22nt或更长、23nt或更长、24nt或更长、25nt或更长、26nt或更长、27nt或更长、28nt或更长、29nt或更长、30nt或更长、31nt或更长、32nt或更长、33nt或更长、34nt或更长、或35nt或更长)。在一些情况下,c2c2指导rna的在指导序列的5'端的区域的长度为29nt或更长。
在一些情况下,c2c2指导rna的在指导序列的5'端的区域的长度在12至100nt(例如,12至90nt、12至80nt、12至70nt、12至60nt、12至50nt、12至40nt、15至100nt、15至90nt、15至80nt、15至70nt、15至60nt、15至50nt、15至40nt、20至100nt、20至90nt、20至80nt、20至70nt、20至60nt、20至50nt、20至40nt、25至100nt、25至90nt、25至80nt、25至70nt、25至60nt、25至50nt、25至40nt、28至100nt、28至90nt、28至80nt、28至70nt、28至60nt、28至50nt、28至40nt、29至100nt、29至90nt、29至80nt、29至70nt、29至60nt、29至50nt或29至40nt)的范围内。在一些情况下,c2c2指导rna的在指导序列的5'端的区域的长度在28至100nt的范围内。在一些情况下,c2c2指导rna的在指导序列的5'端的区域的长度在28至40nt的范围内。
在一些情况下,c2c2指导rna的在指导序列的5'端的区域相对于(短于)相应野生型c2c2指导rna的相应区域被截短。例如,成熟的lsec2c2指导rna包括在指导序列的5'端的长度为30个核苷酸(nt)的区域,并且因此主题截短c2c2指导rna(相对于lsec2c2指导rna)可以具有在指导序列的5'端的长度小于30nt(例如,长度小于29nt、28nt、27nt、26nt、25nt、22nt或20nt)的区域。在一些情况下,截短c2c2指导rna包括在指导序列的5'端的长度范围为12至29nt(例如,12至28nt、12至27nt、12至26nt、12至25nt、12至22nt、12至20nt、12至18nt)的区域。在一些情况下,截短c2c2指导rna例如相对于相应的野生型c2c2指导物被截短一个或多个nt(例如,2nt或更长、3nt或更长、4nt或更长、5nt或更长、或10nt或更长))。
在一些情况下,c2c2指导rna的在指导序列的5'端的区域相对于(长于)相应野生型c2c2指导rna的相应区域被延长。例如,成熟的lsec2c2指导rna包括在指导序列的5'端的长度为30个核苷酸(nt)的区域,并且因此延长的c2c2指导rna(相对于lsec2c2指导rna)可以具有在指导序列的5'端的长于30nt(例如,长于31nt、长于32nt、长于33nt、长于34nt或长于35nt)的区域。在一些情况下,延长的c2c2指导rna包括在指导序列的5'端的长度范围为30至100nt(例如,30至90nt、30至80nt、30至70nt、30至60nt、30至50nt或30至40nt)的区域。在一些情况下,延伸的c2c2指导rna包括在指导序列的5'端的(例如,相对于相应野生型c2c2指导rna的相应区域)延伸一个或多个nt(例如,2nt或更长、3nt或更长、4nt或更长、5nt或更长、或10nt或更长)的区域。
在一些情况下,主题c2c2指导rna的长度为30个或更多个核苷酸(nt)(例如,长度为34nt或更长、40nt或更长、45nt或更长、50nt或更长、55nt或更长、60nt或更长、65nt或更长、70nt或更长、或80nt或更长)。在一些情况下,c2c2指导rna的长度为35nt或更长。
在一些情况下,主题c2c2指导rna的长度在30至120nt(例如,30至110nt、30至100nt、30至90nt、30至80nt、30至70nt、30至60nt、35至120nt、35至110nt、35至100nt、35至90nt、35至80nt、35至70nt、35至60nt、40至120nt、40至110nt、40至100nt、40至90nt、40至80nt、40至70nt、40至60nt、50至120nt、50至110nt、50至100nt、50至90nt、50至80nt或50至70nt)的范围内。在一些情况下,c2c2指导rna的长度在33至80nt的范围内。在一些情况下,c2c2指导rna的长度在35至60nt的范围内。
在一些情况下,主题c2c2指导rna相对于(短于)相应野生型c2c2指导rna被截短。例如,成熟的lsec2c2指导rna的长度可以是50个核苷酸(nt),并且因此截短的c2c2指导rna(相对于lsec2c2指导rna)的长度在一些情况下可以小于50nt(例如,长度为小于49nt、48nt、47nt、46nt、45nt、42nt或40nt)。在一些情况下,截短的c2c2指导rna的长度在30至49nt(例如,30至48nt、30至47nt、30至46nt、30至45nt、30至42nt、30至40nt、35至49nt、35至48nt、35至47nt、35至46nt、35至45nt、35至42nt、或35至40nt)的范围内。在一些情况下,截短的c2c2指导rna例如相对于相应的野生型c2c2指导物被截短一个或多个nt(例如,2nt或更长、3nt或更长、4nt或更长、5nt或更长、或10nt或更长))。
在一些情况下,主题c2c2指导rna相对于(长于)相应的野生型c2c2指导rna延长。例如,成熟的lsec2c2指导rna的长度可以为50个核苷酸(nt),并且因此延长的c2c2指导rna(相对于lsec2c2指导rna)在一些情况下可以长于50nt(例如,长于51nt、长于52nt、长于53nt、长于54nt或长于55nt)。在一些情况下,延长的c2c2指导rna的长度在51至100nt(例如,51至90nt、51至80nt、51至70nt、51至60nt、53至100nt、53至90nt、53至80nt、53至70nt、53至60nt、55至100nt、55至90nt、55至80nt、55至70nt或55至60nt)的范围内。在一些情况下,延长的c2c2指导rna(例如,相对于相应野生型c2c2指导rna)延长一个或多个nt(例如,2nt或更长、3nt或更长、4nt或更长、5nt或更长、或10nt或更长)。
切割前体c2c2指导rna阵列的方法
本公开提供了将前体c2c2指导rna阵列切割成两个或更多个c2c2指导rna的方法。该方法包括使前体c2c2指导rna阵列与c2c2蛋白接触。前体c2c2指导rna阵列包含两个或更多个(例如,2个、3个、4个、5个或更多个)c2c2指导rna,每个所述c2c2指导rna可以具有不同的指导序列。c2c2蛋白将前体c2c2指导rna阵列切割成单独的c2c2指导rna。在一些情况下,c2c2指导rna的恒定区(也称为‘柄’)包括来自前体指导rna的核苷酸序列(例如,在切割指导rna阵列之前通常存在的序列)。换句话说,在一些情况下,主题c2c2指导rna的恒定区包括前体crrna柄。
在一些情况下,接触步骤不在细胞内(例如在活细胞内)发生。在一些情况下,接触步骤在细胞(例如,体外细胞(培养中)、离体细胞、体内细胞)内发生。任何细胞都是合适的。其中可以发生接触的细胞的实例包括但不限于:真核细胞;原核细胞(例如细菌细胞、古细菌细胞);单细胞真核生物;植物细胞;藻类细胞,例如布朗葡萄藻、莱茵衣藻、迦得拟微绿球藻、蛋白核小球藻、展枝马尾藻、羽藻等等;真菌细胞(例如,酵母细胞);动物细胞;无脊椎动物细胞(例如,果蝇、刺胞动物、棘皮动物、线虫、昆虫、蜘蛛类动物,等);脊椎动物细胞(例如,鱼类、两栖动物、爬行动物、鸟类、哺乳动物);哺乳动物细胞(例如,人类;非人灵长类动物;有蹄类动物;猫科动物;牛;绵羊;山羊;大鼠;小鼠;啮齿动物;猪;绵羊;牛;等等);寄生虫细胞(例如,蠕虫、疟疾寄生虫等)。
c2c2蛋白
当c2c2蛋白具有完整的hepn结构域时,其可以在被‘激活’后切割rna(靶rna以及非靶rna)。然而,c2c2蛋白也可以以不依赖hepn的方式将前体c2c2指导rna切割成成熟c2c2指导rna。例如,当c2c2蛋白缺乏催化活性的hepn1结构域并且也缺乏催化活性的hepn2结构域时,其仍然可以将前体指导rna切割成成熟的指导rna。因此,当在包括前体c2c2指导rna和/或前体c2c2指导rna阵列的方法中使用时,c2c2蛋白可以(并且在一些情况下将)缺乏催化活性的hepn1结构域和/或催化活性的hepn2结构域。在一些情况下,c2c2蛋白缺乏催化活性的hepn1结构域并且缺乏催化活性的hepn2结构域。
缺乏催化活性hepn1结构域并且缺乏催化活性hepn2结构域的c2c2蛋白在一些情况下可用于结合方法(例如,成像方法)中。例如,在一些情况下,结合(和/或成像)方法包括使样品与前体c2c2指导rna阵列和缺乏催化活性hepn1结构域且缺乏催化活性hepn2结构域的c2c2蛋白接触。在此类情况下,可以可检测地标记c2c2蛋白(例如,融合表位标签、与荧光团融合、与荧光蛋白(诸如绿色荧光蛋白)融合等)。
适用于在本公开的方法中的用于切割前体c2c2指导rna阵列的c2c2蛋白可具有完整的hepn1和hepn2结构域。然而,在一些情况下,c2c2蛋白缺乏催化活性的hepn1结构域和/或缺乏催化活性的hepn2结构域。
在一些情况下,适用于在本公开的方法中的用于切割前体c2c2指导rna阵列的c2c2蛋白包括与seqidno:1至seqidno:6中任一项所示的氨基酸序列具有80%或更多(例如,85%或更多、90%或更多、95%或更多、98%或更多、99%或更多、99.5%或更多、或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,适用于在本公开的方法中的用于切割前体c2c2指导rna阵列的c2c2蛋白包含与seqidno:1所示的斯氏李斯特菌c2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,适用于在本公开的方法中的用于切割前体c2c2指导rna阵列的c2c2蛋白包含与seqidno:2所示的口腔纤毛菌c2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,适用于在本公开的方法中的用于切割前体c2c2指导rna阵列的c2c2蛋白包含与seqidno:4所示的荚膜红细菌c2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,适用于在本公开的方法中的用于切割前体c2c2指导rna阵列的c2c2蛋白包含与seqidno:5所示的鸡肉杆菌c2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,适用于在本公开的方法中的用于切割前体c2c2指导rna阵列的c2c2蛋白包含与seqidno:6所示的herbinixhemicellulosilyticac2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,适用于在本公开的方法中的用于切割前体c2c2指导rna阵列的c2c2蛋白包含与seqidno:2所示的口腔纤毛菌(lbu)c2c2氨基酸序列具有80%或更多的氨基酸序列同一性的氨基酸序列。在一些情况下,适用于包括在本公开的试剂盒中的c2c2蛋白是口腔纤毛菌(lbu)c2c2蛋白(例如,参见seqidno:2)。在一些情况下,适用于在本公开的方法中的用于切割前体c2c2指导rna阵列的c2c2蛋白包括seqidno:1至seqidno:2和seqidno:4至seqidno:6中任一项所示的氨基酸序列。
在一些情况下,本公开的方法中所使用的用于切割前体c2c2指导rna阵列的c2c2蛋白不是沙氏纤毛菌(lsh)c2c2蛋白。在一些情况下,本公开的方法中所使用的用于切割前体c2c2指导rna阵列的c2c2蛋白不是与seqidno:3所示的lshc2c2多肽具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的c2c2多肽。
在一些情况下,适用于在本公开的方法中的用于切割前体c2c2指导rna阵列的c2c2多肽是变体c2c2多肽。适用于在本公开的方法中的用于切割前体c2c2指导rna阵列的变体c2c2多肽包括seqidno:1、seqidno:2和seqidno:4至seqidno:6中任一者的变体,其中变体c2c2多肽表现出降低的(或不可检测的)核酸酶活性。例如,在一些情况下,变体c2c2蛋白缺乏催化活性的hepn1结构域。作为另一个实例,变体c2c2蛋白缺乏催化活性的hepn2结构域。在一些情况下,变体c2c2蛋白缺乏催化活性的hepn1结构域并且缺乏催化活性的hepn2结构域。
在一些情况下,变体c2c2多肽包含对seqidno:2(口腔纤毛菌c2c2)所示的氨基酸序列的氨基酸r472、h477、r1048和h1053中的1个、2个、3个或4个氨基酸的氨基酸取代,或seqidno:1、seqidno:4、seqidno:5或seqidno:6中所示的c2c2氨基酸序列的相应氨基酸的氨基酸取代。seqidno:1、seqidno:4、seqidno:5和seqidno:6中的相应氨基酸容易被鉴别;参见例如图22b。例如,seqidno:1(斯氏李斯特菌c2c2)中对应于seqidno:2的r472、h477、r1048和h1053的氨基位置分别是r445、h450、r1016和h1021。
在一些情况下,变体c2c2多肽包含对seqidno:2所示的氨基酸序列的氨基酸r472和h477的氨基酸取代,或对seqidno:1、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含对seqidno:2所示的氨基酸序列的氨基酸r1048和h1053的氨基酸取代,或对seqidno:1、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含对seqidno:2所示的氨基酸序列的氨基酸r472、h477、r1048和h1053的氨基酸取代,或对seqidno:1、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列的相应氨基酸的氨基酸取代。
在一些情况下,变体c2c2多肽包含与seqidno:2所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472和h477的取代。在一些情况下,位置472处的氨基酸是除了arg以外的任何氨基酸;并且位置477处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r472a和h477a。在一些情况下,变体c2c2多肽包含与seqidno:2所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1048和h1053的取代。在一些情况下,位置1048处的氨基酸是除了arg以外的任何氨基酸;并且位置1053处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1048a和h1053a。在一些情况下,变体c2c2多肽包含与seqidno:2所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472、h477、r1048和h1053的取代。在一些情况下,位置472和1048处的氨基酸是除了arg以外的任何氨基酸;并且位置477和1053处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r472a、h477a、r1048a和h1053a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
在一些情况下,变体c2c2多肽包含与seqidno:1所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r445和h450的取代。在一些情况下,位置445处的氨基酸是除了arg以外的任何氨基酸;并且位置450处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r445a和h450a。在一些情况下,变体c2c2多肽包含与seqidno:1所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1016和h1021的取代。在一些情况下,位置1016处的氨基酸是除了arg以外的任何氨基酸;并且位置1021处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1016a和h1021a。在一些情况下,变体c2c2多肽包含与seqidno:1所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r445、h450、r1016和h1021的取代。在一些情况下,位置445和1016处的氨基酸是除了arg以外的任何氨基酸;并且位置450和1016处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r445a、h450a、r1016a和h1021a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
在一些情况下,变体c2c2多肽包含与seqidno:4(荚膜红细菌c2c2)所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r464和h469的取代。在一些情况下,位置464处的氨基酸是除了arg以外的任何氨基酸;并且位置469处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r464a和h469a。在一些情况下,变体c2c2多肽包含与seqidno:4所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1052和h1057的取代。在一些情况下,位置1052处的氨基酸是除了arg以外的任何氨基酸;并且位置1057处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1052a和h1057a。在一些情况下,变体c2c2多肽包含与seqidno:4所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r464、h469、r1052和h1057的取代。在一些情况下,位置464和1052处的氨基酸是除了arg以外的任何氨基酸;并且位置469和1057处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r464a、h469a、r1052a和h1057a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
在一些情况下,变体c2c2多肽包含与seqidno:5(鸡肉杆菌c2c2)所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r467和h472的取代。在一些情况下,位置467处的氨基酸是除了arg以外的任何氨基酸;并且位置472处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r469a和h472a。在一些情况下,变体c2c2多肽包含与seqidno:5所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1069和h1074的取代。在一些情况下,位置1069处的氨基酸是除了arg以外的任何氨基酸;并且位置1074处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1069a和h1074a。在一些情况下,变体c2c2多肽包含与seqidno:5所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r467、h472、r1069和h1074的取代。在一些情况下,位置467和1069处的氨基酸是除了arg以外的任何氨基酸;并且位置472和1074处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r469a、h472a、r1069a和h1074a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
在一些情况下,变体c2c2多肽包含与seqidno:6(herbinixhemicellulosilyticac2c2)所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472和h477的取代。在一些情况下,位置472处的氨基酸是除了arg以外的任何氨基酸;并且位置477处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r472a和h477a。在一些情况下,变体c2c2多肽包含与seqidno:6所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1044和h1049的取代。在一些情况下,位置1044处的氨基酸是除了arg以外的任何氨基酸;并且位置1049处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1044a和h1049a。在一些情况下,变体c2c2多肽包含与seqidno:6所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472、h477、r1044和h1049的取代。在一些情况下,位置472和1044处的氨基酸是除了arg以外的任何氨基酸;并且位置477和1049处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r472a、h477a、r1044a和h1049a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
前体c2c2指导rna阵列
如下文工作实施例中所证明的,c2c2蛋白可以将前体c2c2指导rna切割成成熟的指导rna,例如通过对前体的内切核糖核酸酶切割。同样如以下工作实施例中所证明的,c2c2蛋白可以将前体c2c2指导rna阵列(其包括串联排列的多于一个c2c2指导rna)切割成两个或更多个单独的c2c2指导rna。因此,在一些情况下,前体c2c2指导rna阵列包含两个或更多个(例如,3个或更多个、4个或更多个、5个或更多个、2个、3个、4个或5个)c2c2指导rna(例如,作为前体分子串联排列)。在一些情况下,前体c2c2指导rna阵列的每个指导rna具有不同的指导序列。在一些情况下,前体c2c2指导rna阵列的两个或更多个指导rna具有相同的指导序列。
在一些情况下,前体c2c2指导rna阵列包含两个或更多个c2c2指导rna,所述两个或更多个c2c2指导rna靶向相同靶rna分子内的不同靶位点。例如,在一些情况下,当所述c2c2指导rna中的任一者与靶rna分子杂交时,这种情形可以通过激活c2c2蛋白而增加检测灵敏度。
在一些情况下,前体c2c2指导rna阵列包含两个或更多个c2c2指导rna,所述两个或更多个c2c2指导rna靶向不同的靶rna分子。例如,当存在潜在靶rna家族中的任何一者时,这种情况可以导致阳性信号。这种阵列可用于例如基于诸如单核苷酸多态性(snp)之类的变异而靶向转录物家族(例如,用于诊断目的)。这也可用于检测是否存在许多不同病毒株中的任何一种(例如,流感病毒变体、寨卡病毒变体、hiv变体等)。这也可用于检测是否存在许多不同物种、菌株、分离株或细菌变体中的任何一者(例如,分枝杆菌(mycobacterium)的不同物种、菌株、分离株或变体,奈瑟氏菌(neisseria)的不同物种、菌株、分离株、或变体,金黄色葡萄球菌(staphylococcusaureus)的不同物种、菌株、分离株、或变体;大肠杆菌的不同物种、菌株、分离株、或变体;等等)。
变体c2c2多肽
本公开提供了变体c2c2多肽,以及包含编码所述变体c2c2多肽的核苷酸序列的核酸(例如,重组表达载体)。
在一些情况下,变体c2c2多肽包含与seqidno:2所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472和h477的取代。在一些情况下,位置472处的氨基酸是除了arg以外的任何氨基酸;并且位置477处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r472a和h477a。在一些情况下,变体c2c2多肽包含与seqidno:2所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1048和h1053的取代。在一些情况下,位置1048处的氨基酸是除了arg以外的任何氨基酸;并且位置1053处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1048a和h1053a。在一些情况下,变体c2c2多肽包含与seqidno:2所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472、h477、r1048和h1053的取代。在一些情况下,位置472和1048处的氨基酸是除了arg以外的任何氨基酸;并且位置477和1053处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r472a、h477a、r1048a和h1053a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
在一些情况下,变体c2c2多肽包含与seqidno:1所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r445和h450的取代。在一些情况下,位置445处的氨基酸是除了arg以外的任何氨基酸;并且位置450处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r445a和h450a。在一些情况下,变体c2c2多肽包含与seqidno:1所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1016和h1021的取代。在一些情况下,位置1016处的氨基酸是除了arg以外的任何氨基酸;并且位置1021处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1016a和h1021a。在一些情况下,变体c2c2多肽包含与seqidno:1所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r445、h450、r1016和h1021的取代。在一些情况下,位置445和1016处的氨基酸是除了arg以外的任何氨基酸;并且位置450和1016处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r445a、h450a、r1016a和h1021a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
在一些情况下,变体c2c2多肽包含与seqidno:4(荚膜红细菌c2c2)所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r464和h469的取代。在一些情况下,位置464处的氨基酸是除了arg以外的任何氨基酸;并且位置469处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r464a和h469a。在一些情况下,变体c2c2多肽包含与seqidno:4所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1052和h1057的取代。在一些情况下,位置1052处的氨基酸是除了arg以外的任何氨基酸;并且位置1057处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1052a和h1057a。在一些情况下,变体c2c2多肽包含与seqidno:4所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r464、h469、r1052和h1057的取代。在一些情况下,位置464和1052处的氨基酸是除了arg以外的任何氨基酸;并且位置469和1057处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r464a、h469a、r1052a和h1057a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
在一些情况下,变体c2c2多肽包含与seqidno:5(鸡肉杆菌c2c2)所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r467和h472的取代。在一些情况下,位置467处的氨基酸是除了arg以外的任何氨基酸;并且位置472处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r469a和h472a。在一些情况下,变体c2c2多肽包含与seqidno:5所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1069和h1074的取代。在一些情况下,位置1069处的氨基酸是除了arg以外的任何氨基酸;并且位置1074处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1069a和h1074a。在一些情况下,变体c2c2多肽包含与seqidno:5所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r467、h472、r1069和h1074的取代。在一些情况下,位置467和1069处的氨基酸是除了arg以外的任何氨基酸;并且位置472和1074处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r469a、h472a、r1069a和h1074a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
在一些情况下,变体c2c2多肽包含与seqidno:6(herbinixhemicellulosilyticac2c2)所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472和h477的取代。在一些情况下,位置472处的氨基酸是除了arg以外的任何氨基酸;并且位置477处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r472a和h477a。在一些情况下,变体c2c2多肽包含与seqidno:6所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1044和h1049的取代。在一些情况下,位置1044处的氨基酸是除了arg以外的任何氨基酸;并且位置1049处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1044a和h1049a。在一些情况下,变体c2c2多肽包含与seqidno:6所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472、h477、r1044和h1049的取代。在一些情况下,位置472和1044处的氨基酸是除了arg以外的任何氨基酸;并且位置477和1049处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r472a、h477a、r1044a和h1049a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
在一些情况下,变体c2c2多肽包含与seqidno:2所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472、h477、r1048和h1053中的1个、2个、3个或4个氨基酸的取代,使得变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并且保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。例如,在一些情况下,变体c2c2多肽表现出由具有seqidno:2所示氨基酸序列的c2c2多肽表现出的rna指导的非靶rna切割的小于50%、小于40%、小于30%、小于20%、小于10%、小于5%、小于1%或小于0.1%。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
本公开提供了核酸(例如,分离的核酸),所述核酸包含编码本公开的变体c2c2多肽的核苷酸序列。在一些情况下,核苷酸序列与转录控制元件(例如,启动子)可操作地连接。在一些情况下,启动子是组成型启动子。在一些情况下,启动子是可调控的启动子。在一些情况下,启动子是诱导型启动子。在一些情况下,启动子是在真核细胞中起作用的。在一些情况下,启动子是在原核细胞中起作用的。
本公开提供了重组表达载体,所述重组表达载体包含本公开的核酸,例如包含编码本公开的变体c2c2多肽的核苷酸序列的核酸。
本公开提供了用本公开的核酸(例如,包含编码本公开的变体c2c2多肽的核苷酸序列的核酸)遗传修饰的宿主细胞。本公开提供了用包含本公开的核酸(例如,包含编码本公开的变体c2c2多肽的核苷酸序列的核酸)的重组表达载体遗传修饰的宿主细胞。在一些情况下,宿主细胞是原核细胞。在一些情况下,宿主细胞是真核细胞。在一些情况下,宿主细胞是体外的。在一些情况下,宿主细胞是离体的。在一些情况下,宿主细胞是体内的。在一些情况下,宿主细胞是细菌细胞。在一些情况下,宿主细胞是酵母细胞。在一些情况下,宿主细胞是植物细胞。在一些情况下,宿主细胞是哺乳动物细胞。在一些情况下,宿主细胞是人细胞。在一些情况下,宿主细胞是非人哺乳动物细胞。在一些情况下,宿主细胞是昆虫细胞。在一些情况下,宿主细胞是节肢动物细胞。在一些情况下,宿主细胞是真菌细胞。在一些情况下,宿主细胞是藻类细胞。
试剂盒
本公开提供了用于检测包含多种rna的样品中的靶rna的试剂盒。在一些情况下,所述试剂盒包含:(a)前体c2c2指导rna阵列,其包含两个或更多个c2c2指导rna,每个所述c2c2指导rna具有不同的指导序列;以及(b)c2c2蛋白和/或编码所述c2c2蛋白的核酸。在一些情况下,此类试剂盒还包括经标记的检测rna(例如,包含荧光发射染料对(即,fret对和/或猝灭剂/荧光剂对)的经标记的检测rna)。在一些情况下,给定前体c2c2指导rna阵列的两个或更多个c2c2指导rna(例如,在一些情况下,每个c2c2指导rna)包括相同的指导序列。
在一些情况下,主题试剂盒包含:(a)经标记的检测rna,其包含荧光发射染料对,即fret对和/或猝灭剂/荧光剂对;以及(b)c2c2蛋白和/或编码所述c2c2蛋白的核酸。在一些情况下,此类试剂盒还包括(c)c2c2指导rna(和/或编码c2c2指导rna的核酸),和/或(d)前体c2c2指导rna(和/或编码前体c2c2指导rna的核酸),和/或(e)前体c2c2指导rna阵列(和/或编码前体c2c2指导rna阵列的核酸,例如编码包括供用户插入指导序列的插入位点的前体c2c2指导rna阵列的核酸)。
1)包含前体c2c2指导rna阵列和c2c2蛋白的试剂盒
在一些情况下,所述试剂盒包含:(a)前体c2c2指导rna阵列,其包含两个或更多个c2c2指导rna,每个所述c2c2指导rna具有不同的指导序列;以及(b)c2c2蛋白和/或编码所述c2c2蛋白的核酸。如上所述,在一些情况下,此类试剂盒还包括经标记的检测rna(例如,包含荧光发射染料对(即,fret对和/或猝灭剂/荧光剂对)的经标记的检测rna)。
c2c2蛋白
适合于包括在本公开的试剂盒中的c2c2蛋白与c2c2指导rna结合,由指导rna(其与靶rna杂交)引导到单链靶rna,从而被“激活”。如果c2c2蛋白的hepn1结构域和hepn2结构域是完整的,则c2c2蛋白一旦被激活就切割靶rna,但是也切割非靶rna。
在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白包括与seqidno:1至seqidno:6中任一项所示的氨基酸序列具有80%或更多(例如,85%或更多、90%或更多、95%或更多、98%或更多、99%或更多、99.5%或更多、或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白包含与seqidno:1所示的斯氏李斯特菌c2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白包含与seqidno:2所示的口腔纤毛菌c2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白包含与seqidno:4所示的荚膜红细菌c2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白包含与seqidno:5所示的鸡肉杆菌c2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白包含与seqidno:6所示的herbinixhemicellulosilyticac2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白包括与seqidno:2所示的口腔纤毛菌(lbu)c2c2氨基酸序列具有80%或更多的氨基酸序列同一性的氨基酸序列。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白是口腔纤毛菌(lbu)c2c2蛋白(例如,参见seqidno:2)。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白包括seqidno:1至seqidno:2和seqidno:4至seqidno:6中任一项所示的氨基酸序列。
在一些情况下,包括在本公开的试剂盒中的c2c2蛋白不是沙氏纤毛菌(lsh)c2c2蛋白。在一些情况下,包括在本公开的试剂盒中的c2c2蛋白不是与seqidno:3所示的lshc2c2多肽具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的c2c2多肽。
在一些情况下,包括在本公开的试剂盒中的c2c2多肽是变体c2c2多肽。适合于包括在本公开的试剂盒中的变体c2c2多肽包括seqidno:1、seqidno:2和seqidno:4至seqidno:6中任一者的变体,其中变体c2c2多肽表现出降低的(或不可检测的)核酸酶活性。例如,在一些情况下,变体c2c2蛋白缺乏催化活性的hepn1结构域。作为另一个实例,变体c2c2蛋白缺乏催化活性的hepn2结构域。在一些情况下,变体c2c2蛋白缺乏催化活性的hepn1结构域并且缺乏催化活性的hepn2结构域。
在一些情况下,变体c2c2多肽包含对seqidno:2(口腔纤毛菌c2c2)所示的氨基酸序列的氨基酸r472、h477、r1048和h1053中的1个、2个、3个或4个氨基酸的氨基酸取代,或seqidno:1、seqidno:4、seqidno:5或seqidno:6中所示的c2c2氨基酸序列的相应氨基酸的氨基酸取代。seqidno:1、seqidno:4、seqidno:5和seqidno:6中的相应氨基酸容易被鉴别;参见例如图22b。例如,seqidno:1(斯氏李斯特菌c2c2)中对应于seqidno:2的r472、h477、r1048和h1053的氨基位置分别是r445、h450、r1016和h1021。
在一些情况下,变体c2c2多肽包含对seqidno:2所示的氨基酸序列的氨基酸r472和h477的氨基酸取代,或对seqidno:1、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含对seqidno:2所示的氨基酸序列的氨基酸r1048和h1053的氨基酸取代,或对seqidno:1、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列的相应氨基酸的氨基酸取代。在一些情况下,变体c2c2多肽包含对seqidno:2所示的氨基酸序列的氨基酸r472、h477、r1048和h1053的氨基酸取代,或对seqidno:1、seqidno:4、seqidno:5或seqidno:6所示的c2c2氨基酸序列的相应氨基酸的氨基酸取代。
在一些情况下,变体c2c2多肽包含与seqidno:2所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472和h477的取代。在一些情况下,位置472处的氨基酸是除了arg以外的任何氨基酸;并且位置477处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r472a和h477a。在一些情况下,变体c2c2多肽包含与seqidno:2所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1048和h1053的取代。在一些情况下,位置1048处的氨基酸是除了arg以外的任何氨基酸;并且位置1053处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1048a和h1053a。在一些情况下,变体c2c2多肽包含与seqidno:2所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472、h477、r1048和h1053的取代。在一些情况下,位置472和1048处的氨基酸是除了arg以外的任何氨基酸;并且位置477和1053处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r472a、h477a、r1048a和h1053a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
在一些情况下,变体c2c2多肽包含与seqidno:1所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r445和h450的取代。在一些情况下,位置445处的氨基酸是除了arg以外的任何氨基酸;并且位置450处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r445a和h450a。在一些情况下,变体c2c2多肽包含与seqidno:1所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1016和h1021的取代。在一些情况下,位置1016处的氨基酸是除了arg以外的任何氨基酸;并且位置1021处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1016a和h1021a。在一些情况下,变体c2c2多肽包含与seqidno:1所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r445、h450、r1016和h1021的取代。在一些情况下,位置445和1016处的氨基酸是除了arg以外的任何氨基酸;并且位置450和1016处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r445a、h450a、r1016a和h1021a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
在一些情况下,变体c2c2多肽包含与seqidno:4(荚膜红细菌c2c2)所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r464和h469的取代。在一些情况下,位置464处的氨基酸是除了arg以外的任何氨基酸;并且位置469处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r464a和h469a。在一些情况下,变体c2c2多肽包含与seqidno:4所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1052和h1057的取代。在一些情况下,位置1052处的氨基酸是除了arg以外的任何氨基酸;并且位置1057处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1052a和h1057a。在一些情况下,变体c2c2多肽包含与seqidno:4所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r464、h469、r1052和h1057的取代。在一些情况下,位置464和1052处的氨基酸是除了arg以外的任何氨基酸;并且位置469和1057处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r464a、h469a、r1052a和h1057a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
在一些情况下,变体c2c2多肽包含与seqidno:5(鸡肉杆菌c2c2)所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r467和h472的取代。在一些情况下,位置467处的氨基酸是除了arg以外的任何氨基酸;并且位置472处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r469a和h472a。在一些情况下,变体c2c2多肽包含与seqidno:5所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1069和h1074的取代。在一些情况下,位置1069处的氨基酸是除了arg以外的任何氨基酸;并且位置1074处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1069a和h1074a。在一些情况下,变体c2c2多肽包含与seqidno:5所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r467、h472、r1069和h1074的取代。在一些情况下,位置467和1069处的氨基酸是除了arg以外的任何氨基酸;并且位置472和1074处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r469a、h472a、r1069a和h1074a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
在一些情况下,变体c2c2多肽包含与seqidno:6(herbinixhemicellulosilyticac2c2)所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472和h477的取代。在一些情况下,位置472处的氨基酸是除了arg以外的任何氨基酸;并且位置477处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r472a和h477a。在一些情况下,变体c2c2多肽包含与seqidno:6所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1044和h1049的取代。在一些情况下,位置1044处的氨基酸是除了arg以外的任何氨基酸;并且位置1049处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r1044a和h1049a。在一些情况下,变体c2c2多肽包含与seqidno:6所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472、h477、r1044和h1049的取代。在一些情况下,位置472和1044处的氨基酸是除了arg以外的任何氨基酸;并且位置477和1049处的氨基酸是除了his以外的任何氨基酸。在一些情况下,取代是r472a、h477a、r1044a和h1049a。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),并保留结合c2c2指导rna和ssrna的能力。在一些情况下,变体c2c2多肽保留切割前体c2c2指导rna的能力。在一些情况下,变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
2)包含经标记的检测rna和c2c2蛋白的试剂盒
在一些情况下,所述试剂盒包含:(a)经标记的检测rna,其包含荧光发射染料对,即fret对和/或猝灭剂/荧光剂对;以及(b)c2c2蛋白和/或编码所述c2c2蛋白的核酸。在一些情况下,试剂盒还包括c2c2指导rna、前体c2c2指导rna阵列和/或编码c2c2指导rna的恒定区的核酸。如上所述,在一些情况下,此类试剂盒还包括(c)c2c2指导rna(和/或编码c2c2指导rna的核酸),和/或(d)前体c2c2指导rna(和/或编码前体c2c2指导rna的核酸),和/或(e)前体c2c2指导rna阵列(和/或编码前体c2c2指导rna阵列的核酸,例如编码包括供用户插入指导序列的插入位点的前体c2c2指导rna阵列的核酸)。
经标记的检测rna
在一些情况下,本公开的试剂盒包含经标记的检测rna,所述经标记的检测rna包含荧光发射染料对,即fret对和/或猝灭剂/荧光剂对。经标记的检测rna在被切割之前产生一定量的可检测信号,并且当所述经标记的检测rna被切割时所测量到的可检测信号的量减少。在一些情况下,经标记的检测rna在被切割之前产生第一可检测信号(例如,来自fret对),并且当所述经标记的检测rna被切割时产生第二可检测信号(例如,来自猝灭剂/荧光剂对)。因此,在一些情况下,经标记的检测rna包含fret对和猝灭剂/荧光剂对。
在一些情况下,经标记的检测rna包含fret对。fret是这样的过程,通过所述过程,能量的无辐射转移发生为从激发态荧光团到紧邻的第二发色团。能量转移可以发生的范围限制在约10纳米(100埃),并且转移效率对荧光团之间的分开距离非常敏感。供体-受体对(fret供体部分和fret受体部分)在本文中被称为“fret对”或“信号fret对”。因此,在一些情况下,主题经标记的检测rna包括两个信号配偶体(信号对),当一个信号配偶体是fret供体部分时,另一个信号配偶体是fret受体部分。因此,当信号配偶体非常接近时(例如,在相同的rna分子上时),包括这种fret对(fret供体部分和fret受体部分)的主题经标记的检测rna将显示可检测信号(fret信号),但是当所述配偶体分开时(例如,在用c2c2蛋白切割rna分子后),信号将减少(或不存在)。
fret供体和受体部分(fret对)将是本领域的普通技术人员已知的,并且可以使用任何合宜的fret对(例如,任何合宜的供体和受体部分对)。合适的fret对的实例包括但不限于上表1中所示的那些。
在一些情况下,可检测信号在经标记的检测rna被切割时产生(例如,在一些情况下,经标记的检测rna包含猝灭剂/荧光剂对。信号猝灭对的一个信号配偶体产生可检测信号,而另一个信号配偶体是猝灭剂部分,所述猝灭剂部分猝灭第一信号配偶体的可检测信号(即,猝灭剂部分猝灭信号部分的信号,使得当信号配偶体彼此接近时(例如,当信号对的信号配偶体非常接近时),来自信号部分的信号减少(被猝灭)。
例如,在一些情况下,当经标记的检测rna被切割时,可检测信号的量增加。例如,在一些情况下,由一个信号配偶体(信号部分)表现的信号被另一个信号配偶体(猝灭剂信号部分)猝灭,例如当在由c2c2蛋白进行切割之前两者存在于相同的rna分子上时。这种信号对在本文中称为“猝灭剂/荧光剂对”、“猝灭对”或“信号猝灭对”。例如,在一些情况下,一个信号配偶体(例如,第一信号配偶体)是产生可检测信号的信号部分,所述可检测信号由第二信号配偶体(例如,猝灭剂部分)猝灭。因此,当配偶体被分开时(例如,在由c2c2蛋白切割检测rna之后),这种猝灭剂/荧光剂对的信号配偶体将产生可检测信号,但是当配偶体非常接近(例如,在由c2c2蛋白切割检测rna之前)时,所述信号将被猝灭。
猝灭剂部分可以将来自信号部分的信号(例如,在由c2c2蛋白切割检测rna之前)猝灭至不同程度。在一些情况下,猝灭剂部分猝灭来自信号部分的信号,其中在存在猝灭剂部分的情况下(当信号配偶体彼此接近时)检测到的信号是在不存在猝灭剂部分(当信号配偶体分开时)时检测到的信号的95%或更少。例如,在一些情况下,在存在猝灭剂部分的情况下检测到的信号可以是在不存在猝灭剂部分的情况下检测到的信号的90%或更少、80%或更少、70%或更少、60%或更少、50%或更少、40%或更少、30%或更少、20%或更少、15%或更少、10%或更少、或5%或更少。在一些情况下,在存在猝灭剂部分的情况下未检测到信号(例如,高于背景)。
在一些情况下,在不存在猝灭剂部分的情况下(当信号配偶体分开时)检测到的信号是在存在猝灭剂部分的情况下(当信号配偶体彼此接近时)检测到的信号的至少1.2倍(例如,至少1.3倍、至少1.5倍、至少1.7倍、至少2倍、至少2.5倍、至少3倍、至少3.5倍、至少4倍、至少5倍、至少7倍、至少10倍、至少20倍或至少50倍。
在一些情况下,信号部分是荧光标记。在一些此类情况下,猝灭剂部分猝灭来自荧光标记的信号(光信号)(例如,通过吸收标记的发射光谱中的能量)。因此,当猝灭剂部分不与信号部分接近时,来自荧光标记的发射(信号)是可检测的,因为信号不被猝灭剂部分吸收。可以使用任何合宜的供体受体对(信号部分/猝灭剂部分对),并且许多合适的对是本领域中已知的。
在一些情况下,猝灭剂部分从信号部分(在本文中也称为“可检测标记”)吸收能量,然后发射信号(例如,不同波长的光)。因此,在一些情况下,猝灭剂部分本身是信号部分(例如,信号部分可以是6-羧基荧光素,而猝灭剂部分可以是6-羧基-四甲基罗丹明),并且在一些此类情况下,所述对也可以是fret对。在一些情况下,猝灭剂部分是暗猝灭剂。暗猝灭剂可以吸收激发能量并以不同方式耗散能量(例如,作为热量)。因此,暗猝灭剂本身具有极小荧光至没有荧光(不发射荧光)。暗猝灭剂的实例进一步描述于美国专利号8,822,673和8,586,718;美国专利公开20140378330、20140349295和20140194611;以及国际专利申请:wo200142505和wo200186001中,所有这些专利都据此以引用方式整体并入本文。
荧光标记的实例包括但不限于:alexa
在一些情况下,可检测标记是选自以下的荧光标记:alexa
在一些情况下,可检测标记是选自以下的荧光标记:alexa
atto染料的实例包括但不限于:atto390、atto425、atto465、atto488、atto495、atto514、atto520、atto532、attorho6g、atto542、atto550、atto565、attorho3b、attorho11、attorho12、attothio12、attorho101、atto590、atto594、attorho13、atto610、atto620、attorho14、atto633、atto647、atto647n、atto655、attooxa12、atto665、atto680、atto700、atto725以及atto740。
alexafluor染料的实例包括但不限于:alexa
猝灭剂部分的实例包括但不限于:暗猝灭剂、blackhole
在一些情况下,猝灭剂部分选自:暗猝灭剂、blackhole
atto猝灭剂的实例包括但不限于:atto540q、atto580q和atto612q。blackhole
对于一些可检测标记(例如,荧光染料)和/或猝灭剂部分的实例,参见例如bao等人,annurevbiomedeng.2009;11:25-47;以及美国专利号8,822,673和8,586,718;美国专利公开20140378330、20140349295、20140194611、20130323851、20130224871、20110223677、20110190486、20110172420、20060179585和20030003486;以及国际专利申请:wo200142505和wo200186001,所有这些参考文献据此以引用方式整体并入。
核酸修饰
在一些情况下,经标记的检测rna包含一种或多种修饰,例如碱基修饰、骨架修饰、糖修饰等,以为核酸提供新的或增强的特征(例如,改善的稳定性)。如本领域中已知的,核苷是碱基-核糖组合。核苷的碱基部分通常是杂环碱基。此类杂环碱基的两种最常见的类别是嘌呤和嘧啶。核苷酸是这样的核苷,所述核苷还包括与核苷的糖部分共价连接的磷酸基团。对于那些包括戊呋喃糖基糖的核苷,磷酸基团可以与所述糖的2'、3'或5'羟基部分连接。在形成寡核苷酸时,磷酸基团将相邻的核苷彼此共价连接以形成直链聚合化合物。继而,该直链聚合化合物的各个末端可以进一步连接以形成环状化合物;然而,直链化合物通常是合适的。此外,线性化合物可以具有内部核苷酸碱基互补性,并且因此可以以产生完全或部分双链化合物的方式折叠。在寡核苷酸内,磷酸基团通常被称为形成寡核苷酸的核苷间骨架。rna和dna的正常键或骨架是3'至5'磷酸二酯键。
经修饰的骨架和经修饰的核苷间键
合适的修饰的实例包括经修饰的核酸骨架和非天然的核苷间键。(具有经修饰的骨架的核酸包括在骨架中保留磷原子的核酸和在骨架中不具有磷原子的核酸。
其中含有磷原子的合适的经修饰寡核苷酸骨架包括例如硫代磷酸酯、手性硫代磷酸酯、二硫代磷酸酯、磷酸三酯、氨基烷基磷酸三酯、甲基和其它烷基膦酸酯(包括3'-亚烷基膦酸酯、5'-亚烷基膦酸酯和手性膦酸酯)、次膦酸酯、氨基磷酸酯(包括3'-氨基氨基磷酸酯和氨基烷基氨基磷酸酯)、二氨基磷酸酯、硫代氨基磷酸酯、硫代烷基膦酸酯、硫代烷基磷酸三酯、具有正常3'-5'键的硒代磷酸酯和硼代磷酸酯、硒代磷酸酯和硼代磷酸酯的2'-5'连接的类似物,以及具有反向极性的硒代磷酸酯和硼代磷酸酯,其中一个或多个核苷酸间键是3'至3'键、5'至5'键或2'至2'键。合适的具有反向极性的寡核苷酸包含在最3'核苷酸间键处的单个3'至3'键,即单个反向核苷残基,所述单个反向核苷残基可以是碱性的(核碱基丢失或具有羟基代替其)。还包括各种盐(诸如,钾盐或钠盐)、混合盐和游离酸形式。
在一些情况下,经标记的检测rna包含一个或多个硫代磷酸酯和/或杂原子核苷间键,特别是-ch2-nh-o-ch2-、-ch2-n(ch3)-o-ch2-(称为亚甲基(甲基亚氨基)或mmi骨架)、-ch2-o-n(ch3)-ch2-、-ch2-n(ch3)-n(ch3)-ch2-和-o-n(ch3)-ch2-ch2-(其中天然磷酸二酯核苷酸间键表示为-o-p(=o)(oh)-o-ch2-)。mmi型核苷间键在上面引用的美国专利号5,489,677中公开。合适的酰胺核苷间键在美国专利号5,602,240中公开。
同样合适的是如在例如美国专利号5,034,506中所述的具有吗啉代骨架结构的核酸。例如,在一些情况下,经标记的检测rna包含6元吗啉代环代替核糖环。在一些情况下,二氨基磷酸酯或其它非磷酸二酯核苷间键取代磷酸二酯键。
其中不含磷原子的合适的经修饰多核苷酸骨架具有由短链烷基或环烷基核苷间键、混合杂原子和烷基或环烷基核苷间键、或一个或多个短链杂原子或杂环核苷间键形成的骨架。这些骨架包括具有吗啉代键(部分地由核苷的糖部分形成)的那些骨架;硅氧烷骨架;硫化物、亚砜和砜骨架;甲酰基和硫代甲酰基骨架;亚甲基甲酰基和硫代甲酰基骨架;核糖乙酰骨架;含有烯基的骨架;氨基磺酸酯骨架;亚甲基亚氨基和亚甲基肼基骨架;磺酸酯和磺酰胺骨架;酰胺骨架;以及其它具有混合的n、o、s和ch2组成部分的骨架。
模拟物
经标记的检测rna可以是核酸模拟物。应用于多核苷酸的术语“模拟物”旨在包括这样的多核苷酸,在所述多核苷酸中仅呋喃糖环或呋喃糖环和核苷酸间键两者被非呋喃糖基团替代,仅替代呋喃糖环也在本领域中被称为糖替代物。维持杂环碱基部分或经修饰的杂环碱基部分以与合适的靶核酸杂交。一种此类核酸,已显示具有优异杂交特性的多核苷酸模拟物,被称为肽核酸(pna)。在pna中,多核苷酸的糖骨架被含酰胺的骨架,特别是氨基乙基甘氨酸骨架取代。核苷酸被保留并直接或间接地结合骨架的酰胺部分的氮杂氮原子。
一种据报道具有优异杂交特性的多核苷酸模拟物是肽核酸(pna)。pna化合物中的骨架是两个或更多个连接的氨基乙基甘氨酸单元,所述氨基乙基甘氨酸单元赋予pna含酰胺的骨架。杂环碱基部分直接或间接地与骨架的酰胺部分的氮杂氮原子结合。描述pna化合物制备的代表性美国专利包括但不限于:美国专利号5,539,082;5,714,331;和5,719,262。
已经研究的另一类多核苷酸模拟物基于连接的吗啉代单元(吗啉代核酸),所述连接的吗啉代单元具有附接到吗啉代环的杂环碱基。已经报道了许多连接基团,所述连接基团连接吗啉代核酸中的吗啉代单体单元。一类连接基团已经被选择用于提供非离子寡聚化合物。非离子的基于吗啉代的寡聚化合物不太可能与细胞蛋白发生不希望的相互作用。基于吗啉代的多核苷酸是寡核苷酸的非离子模拟物,其不太可能与细胞蛋白形成不希望的相互作用(dwainea.braasch和davidr.corey,biochemistry,2002,41(14),4503-4510)。基于吗啉代的多核苷酸在美国专利号5,034,506中公开。已经制备了吗啉代类多核苷酸内的多种化合物,所述化合物具有各种连接单体亚基的不同连接基团。
另一类多核苷酸模拟物被称为环己烯基核酸(cena)。通常存在于dna/rna分子中的呋喃糖环被环己烯基环替代。cenadmt保护的亚磷酰胺单体已被制备并用于按照经典的亚磷酰胺化学进行寡聚化合物合成。已经制备并研究了完全修饰的cena寡聚化合物和具有用cena修饰的特定位置的寡核苷酸(参见wang等人,j.am.chem.soc.,2000,122,8595-8602)。通常,将cena单体掺入dna链中增加了其dna/rna杂交体的稳定性。cena寡腺苷酸以与天然复合物相似的稳定性而与rna和dna互补物形成复合物。通过nmr和圆二色性显示将cena结构掺入天然核酸结构中的研究,以进行简单的构象适应。
另一修饰包括锁定核酸(lna),其中2'-羟基与糖环的4'碳原子连接,从而形成2'-c,4'-c-氧亚甲基键,从而形成双环糖部分。键可以是亚甲基(-ch2-)、桥接2'氧原子与4'碳原子的基团,其中n是1或2(singh等人,chem.commun.,1998,4,455-456)。lna和lna类似物显示出非常高的互补dna和rna的双链热稳定性(tm=+3至+10℃),朝向3'-核酸外切降解的稳定性和良好的溶解性。已经描述了含有lna的有效和无毒的反义寡核苷酸(wahlestedt等人,proc.natl.acad.sci.u.s.a.,2000,97,5633-5638)。
已经描述了lna单体腺嘌呤、胞嘧啶、鸟嘌呤、5-甲基胞嘧啶、胸腺嘧啶和尿嘧啶的合成和制备,以及它们的寡聚化和核酸识别性质(koshkin等人,tetrahedron,1998,54,3607-3630)。lna及其制备还描述于wo98/39352和wo99/14226中。
经修饰的糖部分
经标记的检测rna还可包括一个或多个经取代的糖部分。合适的多核苷酸包含选自以下的糖取代基:oh;f;o-烷基、s-烷基或n-烷基;o-烯基、s-烯基或n-烯基;o-炔基、s-炔基或n-炔基;或o-烷基-o-烷基,其中所述烷基、烯基和炔基可以是取代或未取代的c1至c10烷基或c2至c10烯基和炔基。特别合适的是o((ch2)no)mch3、o(ch2)noch3、o(ch2)nnh2、o(ch2)nch3、o(ch2)nonh2、以及o(ch2)non((ch2)nch3)2,其中n和m为1至约10。其它合适的多核苷酸包含选自以下的糖取代基:c1至c10低级烷基,取代的低级烷基、烯基、炔基,烷芳基,芳烷基,o-烷芳基或o-芳烷基,sh,sch3,ocn,cl,br,cn,cf3,ocf3,soch3,so2ch3,ono2,no2,n3,nh2,杂环烷基,杂环烷芳基,氨基烷基氨基,聚烷基氨基,取代的甲硅烷基,rna切割基团,报告基团,嵌入剂,用于改善寡核苷酸的药代动力学性质的基团,或用于改善寡核苷酸的药效学性质的基团,以及其它具有相似性质的取代基。合适的修饰包括2'-甲氧基乙氧基(2'-o-ch2ch2och3,也称为2'-o-(2-甲氧基乙基)或2'-moe)(martin等人,helv.chim.acta,1995,78,486-504),即烷氧基烷氧基基团。另一合适的修饰包括2'-二甲基氨氧乙氧基,即o(ch2)2on(ch3)2基团,也称为2'-dmaoe,如下文实施例中所述;以及2'-二甲氨基乙氧基乙氧基(在本领域中也称为2'-o-二甲基-氨基-乙氧基-乙基或2'-dmaeoe),即2'-o-ch2-o-ch2-n(ch3)2。
其它合适的糖取代基包括甲氧基(-o-ch3)、氨基丙氧基(--och2ch2ch2nh2)、烯丙基(-ch2-ch=ch2)、-o-烯丙基(--o--ch2—ch=ch2)和氟(f)。2'-糖取代基可以处于阿拉伯糖(上)位置或核糖(下)位置。合适的2'-阿拉伯糖修饰是2'-f。也可以在寡聚化合物上的其它位置,特别是3'末端核苷上或2'-5'连接的寡核苷酸的糖的3'位置处,以及5'末端核苷酸的5'位置处进行类似的修饰。寡聚化合物也可以具有糖模拟物,诸如环丁基部分代替戊呋喃糖基糖。
碱基修饰和取代
经标记的检测rna还可以包含核碱基(在本领域中通常简称为“碱基”)修饰或取代。如本文所用,“未修饰的”或“天然的”核碱基包括嘌呤碱基腺嘌呤(a)和鸟嘌呤(g),以及嘧啶碱基胸腺嘧啶(t)、胞嘧啶(c)和尿嘧啶(u)。经修饰的核碱基包括其它合成和天然核碱基,诸如5-甲基胞嘧啶(5-me-c)、5-羟甲基胞嘧啶、黄嘌呤、次黄嘌呤、2-氨基腺嘌呤、6-甲基腺嘌呤和鸟嘌呤以及腺嘌呤和鸟嘌呤的其它烷基衍生物、2-丙基腺嘌呤和鸟嘌呤以及腺嘌呤和鸟嘌呤的其它烷基衍生物、2-硫尿嘧啶、2-硫代胸腺嘧啶和2-硫代胞嘧啶、5-氟尿嘧啶和胞嘧啶、5-丙炔基(-c=c-ch3)尿嘧啶和胞嘧啶以及嘧啶碱基的其它炔基衍生物、6-偶氮尿嘧啶、胞嘧啶和胸腺嘧啶、5-尿嘧啶(假尿嘧啶)、4-硫尿嘧啶、8-卤代腺嘌呤和鸟嘌呤、8-氨基腺嘌呤和鸟嘌呤、8-巯基腺嘌呤和鸟嘌呤、8-硫代烷基腺嘌呤和鸟嘌呤、8-羟基腺嘌呤和鸟嘌呤以及其它8-取代的腺嘌呤和鸟嘌呤、5-卤代(特别是5-溴代)尿嘧啶和胞嘧啶、5-三氟甲基尿嘧啶和胞嘧啶以及其它5-取代的尿嘧啶和胞嘧啶、7-甲基鸟嘌呤和7-甲基腺嘌呤、2-f-腺嘌呤、2-氨基腺嘌呤、8-氮杂鸟嘌呤和8-氮杂腺嘌呤、7-脱氮杂鸟嘌呤和7-脱氮杂腺嘌呤,以及3-脱氮杂鸟嘌呤和3-脱氮杂腺嘌呤。其它经修饰的核碱基包括三环嘧啶,诸如吩噁嗪胞苷(1h-嘧啶并(5,4-b)(1,4)苯并噁嗪-2(3h)-酮)、吩噻嗪胞苷(1h-嘧啶(5,4-b)(1,4)苯并噻嗪-2(3h)-酮)、g-钳如取代的吩噁嗪胞苷(例如9-(2-氨基乙氧基)-h-嘧啶并(5,4-(b)(1,4)苯并噁嗪-2(3h)-酮)、咔唑胞苷(2h-嘧啶并(4,5-b)吲哚-2-酮)、吡啶并吲哚胞苷(h-吡啶并(3',2':4,5)吡咯并(2,3-d)嘧啶-2-酮)。
杂环碱基部分还可包括嘌呤碱基或嘧啶碱基被其它杂环取代的那些杂环碱基部分,例如7-脱氮杂腺嘌呤、7-脱氮杂鸟苷、2-氨基吡啶和2-吡啶酮。其它核碱基包括在美国专利号3,687,808中公开的那些核碱基;在theconciseencyclopediaofpolymerscienceandengineering,第858-859页,kroschwitz,j.i.编辑,johnwiley&sons,1990中公开的那些核碱基;由englisch等人,angewandtechemie,internationaledition,1991,30,613公开的那些核碱基;以及由sanghvi,y.s.,第15章,antisenseresearchandapplications,第289-302页,crooke,s.t.和lebleu,b.编辑,crcpress,1993公开的那些核碱基。这些核碱基中的某些可用于增加寡聚化合物的结合亲和力。这些核碱基包括5-取代的嘧啶、6-氮杂嘧啶,以及n-2、n-6和o-6取代的嘌呤,包括2-氨基丙基腺嘌呤、5-丙炔基尿嘧啶和5-丙炔基胞嘧啶。5-甲基胞嘧啶取代已显示为使核酸双链体稳定性增加0.6-1.2℃。(sanghvi等人编辑,antisenseresearchandapplications,crcpress,bocaraton,1993,第276-278页),并且是合适的碱基取代,例如当与2'-o-甲氧基乙基糖修饰组合时。
3)包含两种不同c2c2蛋白和两种不同经标记的检测rna的试剂盒
在一些情况下,主题试剂盒包含:(a)第一经标记的检测rna,其缺乏u但包含至少一个a(例如,至少2个、至少3个、或至少4个a)并且包含第一fret对和/或第一猝灭剂/荧光剂对;(b)第二经标记的检测rna,其缺乏a但包含至少一个u(例如,至少2个、至少3个或至少4个u)并且包含第二fret对和/或第二猝灭剂/荧光剂对;(c)第一c2c2蛋白和/或编码所述第一c2c2蛋白的核酸,其中所述第一c2c2蛋白在被激活时切割腺嘌呤+rna(包括a的rna)而不切割缺乏a的rna(例如,聚urna)[例如,第一c2c2蛋白可以切割第一经标记的检测rna而不切割第二经标记的检测rna];以及(d)第二c2c2蛋白和/或编码所述第二c2c2蛋白的核酸,其中所述第二c2c2蛋白在被激活时切割尿嘧啶+rna(包括u的rna)而不切割缺乏u的rna(例如,聚arna)(例如,第二c2c2蛋白可以切割第二经标记的检测rna而不切割第一经标记的检测rna)。
在一些情况下,第一经标记的检测rna缺乏u并且包括2至15个连续的a(例如,2至12个、2至10个、2至8个、2至6个、2至4个、3至15个、3至12个、3至10个、3至8个、3至6个、3至5个、4至15个、4至12个、4至10个、4至8个或4至6个连续的a)的段。在一些情况下,第一经标记的检测rna缺乏u并且包括至少2个连续的a(例如,至少3个、至少4个或至少5个连续的a)的段。在一些情况下,第二经标记的检测rna缺乏a并且包括2至15个连续的u(例如,2至12个、2至10个、2至8个、2至6个、2至4个、3至15个、3至12个、3至10个、3至8个、3至6个、3至5个、4至15个、4至12个、4至10个、4至8个或4至6个连续的u)的段。在一些情况下,第二经标记的检测rna缺乏a并且包括至少2个连续的u(例如,至少3个、至少4个或至少5个连续的u)的段。
在一些情况下,此类试剂盒还包括:(e)第一c2c2指导rna(和/或编码所述第一c2c2指导rna的核酸),例如包含编码所述第一c2c2指导rna的核苷酸序列的核酸,其中所述核酸包括供用户插入指导序列(例如,与靶rna杂交的核苷酸序列)的序列插入位点);以及(f)第二c2c2指导rna(和/或编码所述第二c2c2指导rna的核酸),例如包含编码所述第二c2c2指导rna的核苷酸序列的核酸,其中所述核酸包括供用户插入指导序列(例如,与靶rna杂交的核苷酸序列)的序列插入位点)。第一c2c2指导rna包含与第一单链靶rna杂交的第一核苷酸序列和与第一c2c2蛋白结合的第二核苷酸序列。第二c2c2指导rna包含与第二单链靶rna杂交的第一核苷酸序列和与第二c2c2蛋白结合的第二核苷酸序列。第一c2c2蛋白不被第二c2c2指导rna激活,并且第一c2c2蛋白切割包括至少一个a的ssrna(例如,不切割缺乏a的ssrna)。第二个c2c2蛋白不被第一c2c2指导rna激活,并且第二c2c2蛋白切割包括至少一个u的ssrna(例如,不切割缺乏u的ssrna)。
以下是适合于包括在本公开的试剂盒中的第一和第二c2c2蛋白的非限制性实例(下文列出为a)至w)):
a)第一c2c2蛋白包含与图56f所示的lbacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56k所示的hhecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
b)第一c2c2蛋白包含与图56f所示的lbacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56g所示的rcacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
c)第一c2c2蛋白包含与图56f所示的lbacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56b所示的pprcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
d)第一c2c2蛋白包含与图56f所示的lbacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56i所示的lnecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
e)第一c2c2蛋白包含与图56f所示的lbacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56c所示的lbucas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
f)第一c2c2蛋白包含与图56f所示的lbacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56e所示的lwacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
g)第一c2c2蛋白包含与图56f所示的lbacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56d所示的lshcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
h)第一c2c2蛋白包含与图56j所示的erecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56k所示的hhecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
i)第一c2c2蛋白包含与图56j所示的erecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56g所示的rcacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
j)第一c2c2蛋白包含与图56j所示的erecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56b所示的pprcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
k)第一c2c2蛋白包含与图56j所示的erecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56i所示的lnecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
l)第一c2c2蛋白包含与图56j所示的erecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56c所示的lbucas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
m)第一c2c2蛋白包含与图56j所示的erecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56e所示的lwacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
n)第一c2c2蛋白包含与图56j所示的erecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56d所示的lshcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
o)第一c2c2蛋白包含与图56j所示的erecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56a所示的lsecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
p)第一c2c2蛋白包含与图56h所示的camcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56k所示的hhecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
q)第一c2c2蛋白包含与图56h所示的camcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56g所示的rcacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
r)第一c2c2蛋白包含与图56h所示的camcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56b所示的pprcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
s)第一c2c2蛋白包含与图56h所示的camcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56i所示的lnecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
t)第一c2c2蛋白包含与图56h所示的camcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56c所示的lbucas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
u)第一c2c2蛋白包含与图56h所示的camcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56e所示的lwacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
v)第一c2c2蛋白包含与图56h所示的camcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56f所示的lshcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;或者
w)第一c2c2蛋白包含与图56h所示的camcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且第二c2c2蛋白包含与图56a所示的lsecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。
c2c2蛋白
适合于包括在本公开的试剂盒中的c2c2蛋白与c2c2指导rna结合,由指导rna(其与靶rna杂交)引导到单链靶rna,从而被“激活”。如果c2c2蛋白的hepn1结构域和hepn2结构域是完整的,则c2c2蛋白一旦被激活就切割靶rna,但是也切割非靶rna。
在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白包括与seqidno:1至seqidno:6中任一项所示的氨基酸序列具有80%或更多(例如,85%或更多、90%或更多、95%或更多、98%或更多、99%或更多、99.5%或更多、或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白包含与seqidno:1所示的斯氏李斯特菌c2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白包含与seqidno:2所示的口腔纤毛菌c2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白包含与seqidno:4所示的荚膜红细菌c2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白包含与seqidno:5所示的鸡肉杆菌c2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白包含与seqidno:6所示的herbinixhemicellulosilyticac2c2氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白包括与seqidno:2所示的口腔纤毛菌(lbu)c2c2氨基酸序列具有80%或更多的氨基酸序列同一性的氨基酸序列。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白是口腔纤毛菌(lbu)c2c2蛋白(例如,参见seqidno:2)。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白包括seqidno:1至seqidno:2和seqidno:4至seqidno:6中任一项所示的氨基酸序列。
在一些情况下,包括在本公开的试剂盒中的c2c2蛋白不是沙氏纤毛菌(lsh)c2c2蛋白。在一些情况下,包括在本公开的试剂盒中的c2c2蛋白不是与seqidno:3所示的lshc2c2多肽具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的c2c2多肽。
在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白比沙氏纤毛菌(lsh)c2c2蛋白在切割未由所述方法的c2c2指导rna靶向的rna方面更有效1.2倍或更多倍。在一些情况下,所述c2c2蛋白比沙氏纤毛菌(lsh)c2c2蛋白在切割未由所述方法的c2c2指导rna靶向的rna方面更有效1.5倍或更多倍。在一些情况下,在本公开的方法中使用的c2c2多肽当被激活时,比lshc2c2至少1.2倍、至少1.5倍、至少2倍、至少2.5倍、至少3倍、至少4倍、至少5倍、至少6倍、至少7倍、至少8倍、至少9倍、至少10倍、至少15倍、至少20倍、至少30倍、或大于30倍有效地切割非靶rna。
在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白在所述接触的1小时内表现出至少50%的rna切割效率(例如,55%或更多、60%或更多、65%或更多、70%或更多、或75%或更多的切割效率)。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白在所述接触的40分钟内表现出至少50%的rna切割效率(例如,55%或更多、60%或更多、65%或更多、70%或更多、或75%或更多的切割效率)。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白在所述接触的30分钟内表现出至少50%的rna切割效率(例如,55%或更多、60%或更多、65%或更多、70%或更多、或75%或更多的切割效率)。
在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白在30秒至60分钟(例如,1分钟至60分钟、30秒至5分钟、1分钟至5分钟、1分钟至10分钟、5分钟至10分钟、10分钟至15分钟、15分钟至20分钟、20分钟至25分钟、25分钟至30分钟、30分钟至35分钟、35分钟至40分钟、40分钟至45分钟、45分钟至50分钟、50分钟至55分钟或55分钟至60分钟)的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白在30秒至5分钟的时间段内(例如,1分钟至5分钟,例如在1分钟、2分钟、3分钟、4分钟或5分钟的时间段内)切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白在5分钟至10分钟的时间段内(例如,在5分钟、6分钟、7分钟、8分钟、9分钟或10分钟的时间段内)切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白在10分钟至15分钟(例如,10分钟、11分钟、12分钟、13分钟、14分钟或15分钟)的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白在15分钟至20分钟的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白在20分钟至25分钟的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白在25分钟至30分钟的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白在30分钟至35分钟的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白在35分钟至40分钟的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白在40分钟至45分钟的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白在45分钟至50分钟的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白在50分钟至55分钟的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白在55分钟至60分钟的时间段内切割样品中存在的rna的至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、至少98%、至少99%或大于99%。
在一些情况下,包括在本公开的试剂盒中的c2c2蛋白不是沙氏纤毛菌(lsh)c2c2蛋白。在一些情况下,c2c2蛋白比沙氏纤毛菌(lsh)c2c2蛋白更有效(例如,在切割非靶rna方面)1.2倍或更高(例如,1.5倍或更高、1.7倍或更高、或2倍或更高)。因此,在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白比沙氏纤毛菌(lsh)c2c2蛋白在切割未由所述方法的c2c2指导rna靶向的rna方面更有效1.2倍或更多倍(例如,1.5倍或更多倍、1.7倍或更多倍、或2倍或更多倍)。在一些情况下,适合于包括在本公开的试剂盒中的c2c2蛋白当被激活时,比lshc2c2至少1.2倍、至少1.5倍、至少2倍、至少2.5倍、至少3倍、至少4倍、至少5倍、至少6倍、至少7倍、至少8倍、至少9倍、至少10倍、至少15倍、至少20倍、至少30倍或大于30倍有效地切割非靶rna。
阳性对照
包含经标记的检测rna和c2c2多肽的本公开的试剂盒还可以包括阳性对照靶rna。在一些情况下,试剂盒还包括阳性对照指导rna,所述阳性对照指导rna包含与对照靶rna杂交的核苷酸序列。在一些情况下,阳性对照靶rna以不同的量在分开的容器中提供。在一些情况下,阳性对照靶rna以各种已知浓度在分开的容器中与对照非靶rna一起提供。
编码c2c2指导rna和/或前体c2c2指导rna阵列和/或c2c2蛋白的核酸
虽然本公开的rna(例如,c2c2指导rna和前体c2c2指导rna阵列)可以使用任何合宜的方法合成(例如,使用rna聚合酶(例如,t7聚合酶、t3聚合酶、sp6聚合酶)在体外化学合成,等等),但是也设想了编码2c2指导rna和/或前体c2c2指导rna阵列的核酸。另外,虽然可以以蛋白形式提供本公开的c2c2蛋白(例如,作为试剂盒的部分),但是也可以提供编码所述c2c2蛋白的核酸(诸如mrna和/或dna)。
例如,在一些实施方案中,本公开的试剂盒包含核酸(例如,dna,例如重组表达载体),所述核酸包含编码c2c2指导rna的核苷酸序列。在一些情况下,核苷酸序列编码没有指导序列的c2c2指导rna。例如,在一些情况下,核酸包含编码c2c2指导rna(没有指导序列的c2c2指导rna)的恒定区的核苷酸序列,并且包含编码指导序列的核酸的插入位点。在一些实施方案中,本公开的试剂盒包含核酸(例如,mrna、dna,例如重组表达载体),所述核酸包含编码c2c2蛋白的核苷酸序列。
在一些实施方案中,本公开的试剂盒包含核酸(例如,dna,例如,重组表达载体),所述核酸包含编码前体c2c2指导rna阵列的核苷酸序列(例如,在一些情况下,其中阵列的每个指导rna具有不同的指导序列)。在一些情况下,阵列的经编码的指导rna中的一者或多者不具有指导序列,例如,核酸可以包括所述阵列的一个或多个指导rna的指导序列的插入位点。在一些情况下,主题c2c2指导rna可以包括来自前体crrna的柄,但不一定必须包括多个指导序列。
在一些情况下,编码c2c2指导rna的核苷酸序列(和/或编码前体c2c2指导rna阵列的核苷酸序列)与启动子可操作地连接,所述启动子为例如在原核细胞中起作用的启动子、在真核细胞中起作用的启动子、在哺乳动物细胞中起作用的启动子、在人细胞中起作用的启动子等。在一些情况下,编码c2c2蛋白的核苷酸序列与启动子可操作地连接,所述启动子为例如在原核细胞中起作用的启动子、在真核细胞中起作用的启动子、在哺乳动物细胞中起作用的启动子、在人细胞中起作用的启动子、细胞型特异性启动子、可调控的启动子、组织特异性启动子等。
本公开的非限制性方面的实例
上述本主题的各方面(包括实施方案)可以单独或与一个或多个其它方面或实施方案一起为有益的。在不限制前述描述的情况下,下面提供了编号1-90的本公开的某些非限制性方面。对于本领域的技术人员在阅读本公开时将显而易见的是,每个单独编号的方面可以使用或与前面或后面的单独编号的方面中的任一者一起组合。这旨在为各方面的所有此类组合提供支持,并且不限于以下明确提供的各方面的组合:
方面1.一种检测包含多种rna的样品中的单链靶rna的方法,所述方法包括:
a)使所述样品与以下接触:(i)c2c2指导rna,其与所述单链靶rna杂交;和(ii)c2c2蛋白,其切割所述样品中存在的rna;以及
b)测量由c2c2蛋白介导的rna切割产生的可检测信号。
方面2.一种检测包含多种rna的样品中的单链靶rna的方法,所述方法包括:
(a)使样品与以下接触:i)前体c2c2指导rna阵列,其包含两个或更多个c2c2指导rna,每个所述c2c2指导rna具有不同的指导序列;和(ii)c2c2蛋白,其将所述前体c2c2指导rna阵列切割成单独的c2c2指导rna,并且还切割所述样品的rna;以及
(b)测量由c2c2蛋白介导的rna切割产生的可检测信号。
方面3.根据方面1或方面2所述的方法,其中所述c2c2蛋白在所述接触1小时内切割所述样品中存在的所述rna的至少50%。
方面4.根据方面3所述的方法,其中所述c2c2蛋白在所述接触40分钟内切割所述样品中存在的所述rna的至少50%。
方面5.根据方面4所述的方法,其中所述c2c2蛋白在所述接触5分钟内切割所述样品中存在的所述rna的至少50%。
方面6.根据方面5所述的方法,其中所述c2c2蛋白在所述接触1分钟内切割所述样品中存在的所述rna的至少50%。
方面7.根据方面1所述的方法,其中所述c2c2蛋白在所述接触1分钟内切割所述样品中存在的所述rna的50%至多于90%。
方面8.根据方面1-7中任一方面所述的方法,其中可以检测到所述单链靶rna的最小浓度在500fm至1nm的范围内。
方面9.根据方面1-7中任一方面所述的方法,其中可以在低至800fm的浓度下检测到所述单链靶rna。
方面10.根据方面1-9中任一方面所述的方法,其中所述c2c2蛋白不是包含与seqidno:3所示的氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列的沙氏纤毛菌(lsh)c2c2蛋白。
方面11.根据方面10所述的方法,其中所述c2c2蛋白比包含与seqidno:3所示的氨基酸序列至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的沙氏纤毛菌(lsh)c2c2蛋白至少1.2倍更有效地切割非靶rna。
方面12.根据方面11所述的方法,其中所述c2c2蛋白比包含seqidno:3所示的氨基酸序列的沙氏纤毛菌(lsh)c2c2蛋白至少1.5倍更有效地切割非靶rna。
方面13.根据方面1-9中任一方面所述的方法,其中所述c2c2蛋白包含与seqidno:1、seqidno:2或seqidno:4至seqidno:6中任一者所示的氨基酸序列具有80%或更多氨基酸序列同一性的氨基酸序列。
方面14.根据方面1-9中任一方面所述的方法,其中所述c2c2蛋白包含与seqidno:2所示的口腔纤毛菌(lbu)c2c2氨基酸序列具有80%或更多氨基酸序列同一性的氨基酸序列。
方面15.根据方面1-9中任一方面所述的方法,其中所述c2c2蛋白包含与seqidno:1所示的斯氏李斯特菌c2c2氨基酸序列具有80%或更多氨基酸序列同一性的氨基酸序列。
方面16.根据方面1-9中任一方面所述的方法,其中所述c2c2蛋白包含seqidno:1至seqidno:2和seqidno:4至seqidno:6中任一者所示的氨基酸序列。
方面17.根据方面1所述的方法,其中所述c2c2蛋白包含与seqidno:2所示的c2c2氨基酸序列具有至少80%氨基酸序列同一性的氨基酸序列,并且包含对r472、h477、r1048和h1053中的一者或多者的取代。
方面18.根据方面1所述的方法,其中所述c2c2蛋白包含与seqidno:2所示的c2c2氨基酸序列具有至少80%氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472和h477的取代。
方面19.根据方面1所述的方法,其中所述c2c2蛋白包含与seqidno:2所示的c2c2氨基酸序列具有至少80%氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r1048和h1053的取代。
方面20.根据方面1所述的方法,其中所述c2c2蛋白包含与seqidno:2所示的c2c2氨基酸序列具有至少80%氨基酸序列同一性的氨基酸序列,并且包含对氨基酸r472、h477、r1048和h1053的取代。
方面21.根据方面1-20中任一方面所述的方法,其中在所述测量之前使所述样品接触2小时或更短时间。
方面22.根据方面21所述的方法,其中在所述测量之前使所述样品接触60分钟或更短时间。
方面23.根据方面22所述的方法,其中在所述测量之前使所述样品接触30分钟或更短时间。
方面24.根据方面23所述的方法,其中在所述测量之前使所述样品接触10分钟或更短时间。
方面25.根据方面24所述的方法,其中在所述测量之前使所述样品接触1分钟或更短时间。
方面26.根据方面1-25中任一方面所述的方法,其包括确定在所述样品中存在的靶rna的量。
方面27.根据方面26所述的方法,其中所述确定包括:
测量所述可检测信号以产生测试测量值;
测量由参考样品产生的可检测信号以产生参考测量值;以及
将所述测试测量值与所述参考测量值进行比较以确定所述样品中存在的靶rna的量。
方面28.根据方面26所述的方法,其包括:
测量所述可检测信号以产生测试测量值,
测量由两个或更多个参考样品中的每一个产生的可检测信号,其中所述两个或更多个参考样品各自包括不同量的阳性对照rna,以产生两个或更多个参考测量值,以及
将所述测试测量值与所述两个或更多个参考测量值进行比较以确定所述样品中存在的靶rna的量。
方面29.根据方面1-28中任一方面所述的方法,其中所述样品包含5至107个序列彼此不同的rna。
方面30.根据方面1-28中任一方面所述的方法,其中所述样品包含10至106个序列彼此不同的rna。
方面31.根据方面1-30中任一方面所述的方法,其中所述样品包含来自细胞裂解物的rna。
方面32.根据方面1-31中任一方面所述的方法,其中测量可检测信号包括以下中的一者或多者:基于金纳米颗粒的检测、荧光偏振、胶体相变/分散、电化学检测和基于半导体的感测。
方面33.根据方面1-32中任一方面所述的方法,其中(i)所述方法包括使所述样品与包含荧光发射染料对(即,荧光共振能量转移(fret)对和/或猝灭剂/荧光剂对)的经标记的检测rna接触,(ii)所述c2c2蛋白切割所述经标记的检测rna,以及(iii)所述可检测信号由所述fret对和/或所述猝灭剂/荧光剂对产生。
方面34.根据方面33所述的方法,其中所述经标记的检测rna在被切割之前产生一定量的可检测信号,并且当所述经标记的检测rna被切割时可检测信号的量减少。
方面35.根据方面33所述的方法,其中所述经标记的检测rna在被切割之前产生第一可检测信号,并且当所述经标记的检测rna被切割时产生第二可检测信号。
方面36.根据方面35所述的方法,其中所述经标记的检测rna包含fret对和猝灭剂/荧光剂对。
方面37.根据方面33-36中任一方面所述的方法,其中所述经标记的检测rna包含fret对。
方面38.根据方面33所述的方法,其中在所述经标记的检测rna被切割时产生可检测信号。
方面39.根据方面33所述的方法,其中当所述经标记的检测rna被切割时,可检测信号的量增加。
方面40.根据方面38或方面39所述的方法,其中所述经标记的检测rna包含猝灭剂/荧光剂对。
方面41.根据方面33-40中任一方面所述的方法,其中所述经标记的检测rna包含经修饰的核碱基、经修饰的糖部分和/或经修饰的核酸键。
方面42.根据方面1-41中任一方面所述的方法,其中所述接触在无细胞样品中进行。
方面43.根据方面1-41中任一方面所述的方法,其中所述接触在体外、离体或体外细胞中进行。
方面44.一种将前体c2c2指导rna阵列切割成两个或更多个c2c2指导rna的方法,所述方法包括:
使包含两个或更多个c2c2指导rna的前体c2c2指导rna阵列与c2c2蛋白接触,每个所述c2c2指导rna阵列具有不同的指导序列,其中所述c2c2蛋白将所述前体c2c2指导rna阵列切割成单独的c2c2指导rna。
方面45.根据方面44所述的方法,其中所述c2c2蛋白缺乏催化活性的hepn1结构域和/或缺乏催化活性的hepn2结构域。
方面46.根据方面44或方面45所述的方法,其中所述前体c2c2指导rna阵列包含两个或更多个c2c2指导rna,所述两个或更多个c2c2指导rna靶向相同靶rna分子内的不同靶序列。
方面47.根据方面44-46中任一方面所述的方法,其中所述前体c2c2指导rna阵列包含两个或更多个c2c2指导rna,所述两个或更多个c2c2指导rna靶向不同靶rna分子。
方面48.根据方面44-47中任一方面所述的方法,其中所述接触不在细胞内发生。
方面49.根据方面44-48中任一方面所述的方法,其中所述指导rna和/或所述前体c2c2指导rna阵列中的至少一者是可检测地标记的。
方面50.一种用于检测包含多种rna的样品中的靶rna的试剂盒,所述试剂盒包含:
(a)前体c2c2指导rna阵列,和/或编码所述前体c2c2指导rna阵列的核酸,其中所述前体c2c2指导rna阵列包含两个或更多个c2c2指导rna,每个所述c2c2指导rna具有不同的指导序列和/或所选择的指导序列的插入位点;以及
(b)c2c2蛋白。
方面51.根据方面50所述的试剂盒,其中所述c2c2蛋白缺乏催化活性的hepn1结构域和/或缺乏催化活性的hepn2结构域。
方面52.根据方面50或方面51所述的试剂盒,其中所述前体c2c2指导rna阵列包含两个或更多个c2c2指导rna,所述两个或更多个c2c2指导rna靶向相同靶rna分子内的不同靶序列。
方面53.根据方面50-52中任一方面所述的试剂盒,其中所述前体c2c2指导rna阵列包含两个或更多个c2c2指导rna,所述两个或更多个c2c2指导rna靶向不同靶rna分子。
方面54.根据方面50-53中任一方面所述的试剂盒,其中所述指导rna和/或所述前体c2c2指导rna阵列中的至少一者是可检测地标记的。
方面55.根据方面50-54中任一方面所述的试剂盒,其还包含经标记的检测rna,所述经标记的检测rna包含荧光发射染料对(即,fret对和/或猝灭剂/荧光剂对)。
方面56.一种用于检测包含多种rna的样品中的靶rna的试剂盒,所述试剂盒包含:
(a)经标记的检测rna,其包含荧光发射染料对(即,fret对和/或猝灭剂/荧光剂对);以及
(b)c2c2蛋白。
方面57.根据方面56所述的试剂盒,其包含阳性对照靶rna。
方面58.根据方面57所述的试剂盒,其中所述阳性对照靶rna以不同的量在两个或更多个容器中的每一个中存在。
方面59.根据方面56-58中任一方面所述的试剂盒,其包含以下中的至少一者:
(c)c2c2指导rna和/或编码所述c2c2指导rna的核酸;
(d)前体c2c2指导rna和/或编码所述前体c2c2指导rna的核酸;以及
(e)前体c2c2指导rna阵列,和/或编码所述前体c2c2指导rna阵列的核酸,其中所述前体c2c2指导rna阵列包含两个或更多个c2c2指导rna,每个所述c2c2指导rna具有不同的指导序列和/或所选择的指导序列的插入位点。
方面60.根据方面56-59中任一方面所述的试剂盒,其包含dna,所述dna包含编码具有或不具有指导序列的c2c2指导rna的核苷酸序列。
方面61.根据方面60所述的试剂盒,其中所述dna包含用于插入指导序列的插入序列。
方面62.根据方面60或方面61所述的试剂盒,其中所述dna是表达载体,并且所述c2c2指导rna与启动子可操作地连接。
方面63.根据方面62所述的试剂盒,其中所述启动子是t7启动子。
方面64.根据方面56-63中任一方面所述的试剂盒,其包含缺乏核酸酶活性的c2c2内切核糖核酸酶变体。
方面65.根据方面56-64中任一方面所述的试剂盒,其中所述经标记的检测rna包含fret对。
方面66.根据方面56-65中任一方面所述的试剂盒,其中所述经标记的检测rna包含猝灭剂/荧光剂对。
方面67.根据方面56-66中任一方面所述的试剂盒,其中所述经标记的检测rna包含产生第一可检测信号的fret对和产生第二可检测信号的猝灭剂/荧光剂对。
方面68.一种变体c2c2多肽,其包含:
a)与seqidno:2所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对以下的取代:i)氨基酸r472和h477;ii)氨基酸r1048和h1053;或iii)氨基酸r472、h477、r1048和h1053;
b)与seqidno:1所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对以下的取代:i)氨基酸r445和h450;ii)氨基酸r1016和h1021;或iii)氨基酸r445、h450、r1016和h1021;
c)与seqidno:4所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对以下的取代:i)氨基酸r464和h469;ii)氨基酸r1052和h1057;或iii)氨基酸r464、h469、r1052和h1057;
d)与seqidno:5所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对以下的取代:i)氨基酸r467和h472;ii)氨基酸r1069和h1074;或iii)氨基酸r467、h472、r1069和h1074;或者
e)与seqidno:6所示氨基酸序列具有至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的氨基酸序列同一性的氨基酸序列,并且包含对以下的取代:i)氨基酸r472和h477;ii)氨基酸r1044和h1049;iii)或氨基酸r472、h477、r1044和h1049。
方面69.根据68所述的变体c2c2多肽,其中所述变体c2c2多肽具有减少的或不可检测的ssrna切割(例如,rna指导的切割活性),但保留了结合c2c2指导rna和ssrna的能力,并且保留了切割前体c2c2指导rna的能力。
方面70.一种核酸,其包含编码根据方面68或方面69所述的变体c2c2多肽的核苷酸序列。
方面71.根据方面70所述的核酸,其中所述核苷酸序列与组成型启动子或可调控的启动子可操作地连接。
方面72.一种重组表达载体,其包含根据方面70或方面71所述的核酸。
方面73.一种宿主细胞,用根据方面70或方面71所述的核酸或用根据方面72所述的重组表达载体遗传修饰。
方面74.根据方面73所述的宿主细胞,其中所述宿主细胞是真核细胞。
方面75.根据方面73所述的宿主细胞,其中所述宿主细胞是原核细胞。
方面76.根据方面73-75中任一方面所述的宿主细胞,其中所述宿主细胞是体外、离体或体内的。
方面77.一种检测包含多种rna的样品中的至少两种不同单链靶rna的方法,所述方法包括:
a)使所述样品与以下接触:
(i)第一c2c2蛋白,其切割包括至少一个a的单链rna(ssrna);
(ii)第二c2c2蛋白,其切割包括至少一个u的ssrna;
(iii)第一c2c2指导rna,其包含与所述第一单链靶rna杂交的第一核苷酸序列和与所述第一c2c2蛋白结合的第二核苷酸序列;以及
(iv)第二c2c2指导rna,其包含与所述第二单链靶rna杂交的第一核苷酸序列和与所述第二c2c2蛋白结合的第二核苷酸序列;
其中所述第一c2c2蛋白不被所述第二c2c2指导rna激活,并且其中所述第一c2c2蛋白切割包括至少一个a的ssrna,并且
其中所述第二c2c2蛋白不被所述第一c2c2指导rna激活,并且其中所述第二c2c2蛋白切割包括至少一个u的ssrna;以及
b)测量由所述第一c2c2蛋白和所述第二c2c2蛋白介导的rna切割产生的可检测信号,其中第一可检测信号在所述第一c2c2蛋白被激活时产生,并且第二可检测信号在所述第二c2c2蛋白被激活时产生,其中检测到所述第一信号表明所述第一靶ssrna存在于所述样品中,并且其中检测到所述第二信号表明所述第二靶ssrna存在于所述样品中。
方面78.根据方面77所述的方法,其中:
a)所述第一c2c2蛋白包含与图56f所示的lbacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56k所示的hhecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
b)所述第一c2c2蛋白包含与图56f所示的lbacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56g所示的rcacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
c)所述第一c2c2蛋白包含与图56f所示的lbacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56b所示的pprcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
d)所述第一c2c2蛋白包含与图56f所示的lbacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56i所示的lnecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
e)所述第一c2c2蛋白包含与图56f所示的lbacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56c所示的lbucas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
f)所述第一c2c2蛋白包含与图56f所示的lbacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56e所示的lwacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
g)所述第一c2c2蛋白包含与图56f所示的lbacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56d所示的lshcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
h)所述第一c2c2蛋白包含与图56j所示的erecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56k所示的hhecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
i)所述第一c2c2蛋白包含与图56j所示的erecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56g所示的rcacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
j)所述第一c2c2蛋白包含与图56j所示的erecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56b所示的pprcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
k)所述第一c2c2蛋白包含与图56j所示的erecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56i所示的lnecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
l)所述第一c2c2蛋白包含与图56j所示的erecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56c所示的lbucas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
m)所述第一c2c2蛋白包含与图56j所示的erecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56e所示的lwacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
n)所述第一c2c2蛋白包含与图56j所示的erecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56d所示的lshcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
o)所述第一c2c2蛋白包含与图56j所示的erecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56a所示的lsecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
p)所述第一c2c2蛋白包含与图56h所示的camcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56k所示的hhecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
q)所述第一c2c2蛋白包含与图56h所示的camcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56g所示的rcacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
r)所述第一c2c2蛋白包含与图56h所示的camcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56b所示的pprcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
s)所述第一c2c2蛋白包含与图56h所示的camcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56i所示的lnecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
t)所述第一c2c2蛋白包含与图56h所示的camcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56c所示的lbucas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
u)所述第一c2c2蛋白包含与图56h所示的camcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56e所示的lwacas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;
v)所述第一c2c2蛋白包含与图56h所示的camcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56f所示的lshcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;或者
w)所述第一c2c2蛋白包含与图56h所示的camcas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列;并且所述第二c2c2蛋白包含与图56a所示的lsecas13a氨基酸序列具有至少75%、至少80%、至少85%、至少90%、至少95%、至少98%、至少99%或100%的氨基酸序列同一性的氨基酸序列。
方面79.根据方面77或方面78所述的方法,其中所述方法包括使所述样品与以下接触:
i)第一经标记的检测rna,其包含第一荧光共振能量转移(fret)对和/或第一猝灭剂/荧光剂对,其中所述第一经标记的检测rna包含至少一个a并且不包含u;和
ii)第二经标记的检测rna,其包含第二fret对和/或第二猝灭剂/荧光剂对,其中所述第二经标记的检测rna包含至少一个u并且不包含a,
其中所述第一c2c2蛋白切割所述第一经标记的检测rna,并且所述第一可检测信号由所述第一fret对和/或所述第一猝灭剂/荧光剂对产生,并且
其中所述第一c2c2蛋白切割所述第二经标记的检测rna,并且所述第二可检测信号由所述第二fret对和/或所述第二猝灭剂/荧光剂对产生。
方面80.根据方面79所述的方法,其中所述第一经标记的检测rna包含2至15个连续的a的段和/或所述第二经标记的检测rna包含2至15个连续的u的段。
方面81.根据方面79所述的方法,其中所述第一经标记的检测rna包含4至15个连续的a的段和/或所述第二经标记的检测rna包含4至15个连续的u的段。
方面82.根据方面79所述的方法,其中所述第一经标记的检测rna包含至少3个连续的a的段和/或所述第二经标记的检测rna包含至少3个连续的u的段。
方面83.根据方面79所述的方法,其中所述第一经标记的检测rna包含至少4个连续的a的段和/或所述第二经标记的检测rna包含至少4个连续的u的段。
方面84.一种试剂盒,其包含:
(a)第一经标记的检测rna,其缺乏u并且包含至少一个a且包含第一荧光发射染料对;
(b)第二经标记的检测rna,其缺乏a并且包含至少一个u且包含第二荧光发射染料对;
(c)第一c2c2蛋白,和/或编码所述第一c2c2蛋白的核酸,其中所述第一c2c2蛋白可切割所述第一经标记的检测rna但不切割所述第二经标记的检测rna(例如,所述第一c2c2蛋白当被激活时切割腺嘌呤+rna并且不切割缺乏a的rna);以及
(d)第二c2c2蛋白,和/或编码所述第二c2c2蛋白的核酸,其中所述第二c2c2蛋白可切割所述第二经标记的检测rna但不切割所述第一经标记的检测rna(例如,所述第二c2c2蛋白当被激活时切割尿嘧啶+rna但不切割缺乏u的rna)。
方面85.根据方面84所述的试剂盒,其包含以下中的至少一者:
(e)第一c2c2指导rna和/或编码所述第一c2c2指导rna的核酸,其中所述第一c2c2指导rna包含与所述第一c2c2蛋白结合的恒定区序列;
(f)第二c2c2指导rna和/或编码所述第二c2c2指导rna的核酸,其中所述第二c2c2指导rna包含与所述第二c2c2蛋白结合的恒定区序列;
(g)核酸,其包含编码与所述第一c2c2蛋白结合的恒定区序列的核苷酸序列和所选择的指导序列的插入位点;
(h)核酸,其包含编码与所述第二c2c2蛋白结合的恒定区序列的核苷酸序列和所选择的指导序列的插入位点。
方面86.根据方面84所述的试剂盒,其包含核酸,所述核酸包含编码第一c2c2指导rna的核苷酸序列,其中所述第一c2c2指导rna包含与所述第一c2c2蛋白结合的恒定区序列。
方面87.根据方面84所述的试剂盒,其包含核酸,所述核酸包含编码第二c2c2指导rna的核苷酸序列,其中所述第二c2c2指导rna包含与所述第二c2c2蛋白结合的恒定区序列。
方面88.根据方面84所述的试剂盒,其包含核酸,所述核酸包含编码与所述第一c2c2蛋白结合的恒定区序列的核苷酸序列和所选择的指导序列的插入位点。
方面89.根据方面84所述的试剂盒,其包含核酸,所述核酸包含编码与所述第二c2c2蛋白结合的恒定区序列的核苷酸序列和所选择的指导序列的插入位点。
方面90.根据方面86-89中任一方面所述的试剂盒,其中所述核酸是表达载体,并且所述核苷酸序列与启动子可操作地连接。
实施例
提出以下实施例是为了向本领域的普通技术人员提供如何制备和使用本发明的完整公开和描述,而非旨在限制本发明人认为是其发明的范围,也非旨在表示以下实验是所执行的全部或唯一实验。已经努力确保关于所使用的数值(例如量、温度等)的准确性,但是应该考虑一些实验误差和偏差。除非另有说明,否则份数是重量份,分子量是重均分子量,温度以摄氏度计,并且压力处于大气压或接近大气压。可以使用标准缩写,例如bp表示碱基对;kb表示千碱基;pl表示皮升;s或sec表示秒;min表示分钟;h或hr表示小时;aa表示氨基酸;kb表示千碱基;bp表示碱基对;nt表示核苷酸;i.m.表示肌内(肌内地);i.p.表示腹膜内(腹膜内地);s.c.表示皮下(皮下地);等等。
本文描述的数据显示,c2c2具有负责crisprrna加工和ssrna降解的两种不同核糖核酸酶活性。前体crisprrna(pre-crrna)的成熟是相对缓慢的、高度特异性的催化事件。相比之下,当与具有与crrna的指导区段的序列互补性的靶rna结合时,c2c2被激活为强大的通用rna酶,所述rna酶通过引发尿嘧啶核苷酸处的链断裂而顺式和反式地切割rna。数据显示,这种反式切割活性可用于复杂混合物内的超灵敏rna检测。
实施例1:c2c2重组蛋白纯化
图1a至图1e.对内源c2c2基因座以及重组c2c2蛋白的异源表达和纯化的概述。图1a.口腔纤毛菌c2c2基因座的示意图。所预测的hepn活性位点以黄色指示,其中活性位点残基被标示。图1b.来自该vi型crispr系统的成熟crrna的示意图。图1c.对阳离子交换色谱级分的十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sds-page)分析。收获表达his-麦芽糖结合蛋白(mbp)-c2c2的大肠杆菌细胞并通过超声处理进行裂解。通过离心澄清裂解物,然后经由金属离子亲和色谱分离his-mbp-c2c2。将蛋白洗脱液与tev蛋白酶一起孵育以切除his-mbp标签。将切割的蛋白上样到hitrap肝素柱上,并以线性kcl梯度洗脱。将洗脱级分上样到10%sds-page凝胶上进行分析。图1d.将阳离子交换色谱级分汇集、浓缩,然后上样到s200尺寸排阻色谱柱上。经由sds-page分析来自尺寸交换色谱的级分。图1e.c2c2酶促活性的工作模型。
实施例2:c2c2是可编程的内切核糖核酸酶
纯化的c2c2以crrna指导的方式切割单链rna(ssrna),所述crrna指导的方式依赖于催化活性hepn结构域的存在(图2)
图2.在20mmtris-hclph6.8、50mmkcl、5mmmgcl2、5%甘油中执行切割测定。将100nmc2c2蛋白与crrna以2:1的比率在37℃下预孵育10分钟以促进复合物组装。约1nm的5’32p标记的ssrna的加入引发反应,并且通过用甲酰胺上样缓冲液猝灭来获取时间点。将切割产物形成通过15%变性尿素-page进行解析,并使用磷光成像仪可视化。灭活蛋白(由dc2c2表示法表示)是四点突变体(r472a、h477a、r1048a、h1053a),其在两个预测的活性位点中均从r-x4-6-hhepn基序消除精氨酸和组氨酸两者。在这种凝胶中,所关注的泳道用红色框突出显示。使用脱靶ssrna时或当hepn结构域失活时未检测到切割活性。
实施例3:c2c2切割活性对单链rna有特异性
c2c2表现为不切割双链rna或dna,其中与ssrna相比,以大大降低的速率有限地切割单链dna(图3)。
图3.c2c2强烈切割单链rna并且不切割双链底物。切割测定条件如图2所示。将切割产物在15%变性尿素-page凝胶上解析。在5分钟、10分钟、30分钟、75分钟时取样,其中所有对照泳道平行孵育75分钟。具有预期尺寸范围的切割产物可仅在ssrna反应中检测到,并且在单链dna条件下以更小程度检测到。对单链rna靶的3'末端标记表明,切割发生在靶-间隔区序列杂交区外部。图中顶部的标签指示是添加了蛋白(“lbuc2c2”)还是c2c2指导rna(“crgapdh1”)。对照包括(i)蛋白但没有指导rna,(ii)指导rna但没有蛋白,以及(iii)没有蛋白和没有指导rna。
实施例4:c2c2将前体crrna加工成成熟crrna
c2c2以不依赖于hepn结构域的方式将前体crrna加工成成熟crrna(图4a至图4b),表明存在另外的内切核酸酶结构域。
图4a.由c2c2催化的pre-crrna加工反应的图。图4b.在用甲酰胺上样缓冲液进行猝灭之前,将100nmlbuc2c2与约5nm5’32p标记的ssrna一起在20mm的hepesph6.8、50mmkcl、5mmmgcl2、5%甘油中孵育60分钟。将反应产物在15%变性尿素-page凝胶上解析,并使用磷光成像仪(phosphoimager)可视化。灭活蛋白(由dc2c2表示法表示)是四点突变体(r472a、h477a、r1048a、h1053a),其在两个预测的活性位点中均从r-x4-6-hhepn基序消除精氨酸和组氨酸两者。测试具有可变间隔区序列的三种不同pre-crrna的c2c2加工能力。
实施例5:复杂混合物中的转录物的灵敏检测
使用c2c2检测复杂混合物中的靶转录物(图5)。
图5.在存在不断增加量的λ2ssrna(0-10nm)的情况下,将50nm的靶向c2c2:crrna的λ2ssrna与185nm的rna酶-alert底物(小型荧光-猝灭剂rna寡核苷酸-用猝灭剂/荧光剂对标记的经标记的检测rna)和100ng的hek293t总rna一起在37c下孵育30分钟。当‘经激活的’c2c2(c2c2:crrna:λ2ssrna)切割rna酶-alert底物而从猝灭剂部分释放荧光染料时,观察到荧光增加。
实施例6:由c2c2蛋白进行前体crrna(pre-crrna)加工
c2c2切割(加工)前体crrna(pre-crrna),并且该切割不依赖于hepn结构域(图6)。
图6.将300nm的c2c2与1nm的5’-32p标记的pre-crrna一起在37c下孵育0-60分钟。将rna产物在15%尿素-page上分离,并使用磷光成像仪成像。通过较小的‘经切割的rna产物’的存在来确定pre-crrna的成功加工。
实施例7
材料和方法
使用以下材料和方法,并且以下材料和方法适用于实施例8-12。
c2c2系统发生和候选选择。使用具有protgammalg进化模型和100个自展抽样(bootstrapsampling)的raxml计算c2c2最大似然系统发生。通过mafft使用‘einsi’方法比对序列。niewoehner,o.和jinek,m.,structuralbasisfortheendoribonucleaseactivityofthetypeiii-acrispr-associatedproteincsm6.rna22,318–329(2016)。选择候选同系物来对蛋白家族的主要分支进行取样。
c2c2蛋白的生产和纯化。使用从integrateddnatechnologies订购的合成gblock组装用于蛋白纯化的表达载体。将密码子优化的c2c2基因组序列用his6-mbp-tev切割位点进行n末端标记,其中表达由t7启动子驱动(根据要求可用的映射(mapavailableuponrequest))。经由野生型c2c2构建体的圆角化(round-the-horn)、定点诱变来克隆突变蛋白。将表达载体转化到于37℃下在2xyt肉汤中生长的rosetta2大肠杆菌细胞中。将大肠杆菌细胞在对数期期间用0.5m的异丙基β-d-1-硫代半乳糖苷(itpg)诱导,并将温度降至16℃以过夜表达his-mbp-c2c2。随后收获细胞,重悬于裂解缓冲液(50mmtris-hclph7.0、500mmnacl、5%甘油、1mm三(2-羧乙基)膦(tcep)、0.5mm苯甲基磺酰氟(pmsf)以及不含乙二胺四乙酸(edta)的蛋白酶抑制剂(roche)中,并通过超声裂解,并且将裂解物通过离心澄清。通过金属离子亲和色谱分离出可溶性his-mbp-c2c2,将含蛋白的洗脱物与烟草蚀纹病毒(tev)蛋白酶一起在4℃下孵育过夜,同时透析到离子交换缓冲液(50mmtris-hclph7.0、250mmkcl、5%甘油、1mmtcep)中以切除his6-mbp标签。将切割的蛋白上样到hitrapsp柱上,并以线性kcl(0.25-1.5m)梯度洗脱。将阳离子交换色谱级分汇集,并用30kd的截留浓缩管(thermofisher)浓缩。将c2c2蛋白经由在s200柱上进行尺寸排阻色谱而进一步纯化,并储存在凝胶过滤缓冲液(20mmtris-hclph7.0、200mmkcl、5%甘油、1mmtcep)中以供后续酶测定。
生成ssrna。除了crrnaaes461是订购合成的(integrateddnatechnologies)之外,本研究中使用的所有rna都是在体外转录的[参见图15和图19]。如前所述进行体外转录反应,具有以下改变:t7聚合酶浓度降至10μg/ml,并且utp浓度降至2.5mm。将转录在37℃下孵育1-2小时以减少核苷酸的非模板加入。使用15%变性page凝胶纯化所有转录反应。将所有rna重悬于切割缓冲液(20mmhepesph6.8、50mmkcl、5mmmgcl2和5%甘油)中。对于放射性实验,在放射性标记之前用小牛肠磷酸盐(newenglandbiolabs)去除5'三磷酸酯,然后如前所述使用t4多核苷酸激酶(newenglandbiolabs)和[γ-32p]-atp(perkinelmer)对ssrna底物进行5'末端标记。sternberg,s.h.、haurwitz,r.e.和doudna,j.a.mechanismofsubstrateselectionbyahighlyspecificcrisprendoribonuclease.rna18,661–672(2012)。
在该研究中使用的dna底物列于图14的表2中。
在该研究中使用的rna底物列于图15的表3中。
pre-crrna加工测定。在37℃下在加工缓冲液(20mmhepesph6.8、50mmkcl、5mmmgcl2、10μg/ml牛血清白蛋白(bsa)、10μg/mltrna、0.05%igepalca-630和5%甘油)以及相对于5'-标记的pre-crrna100倍摩尔过量的c2c2(最终浓度分别为100nm和大于1nm)中执行pre-crrna切割测定。除非另有说明,否则在60分钟后用1.5xrna上样染料(100%甲酰胺、0.025w/v%溴酚蓝和200μgml-1肝素)猝灭反应。猝灭后,将反应在95℃变性5分钟,然后通过12%或15%变性page(0.5xtbe缓冲液)解析。通过向反应缓冲液中加入浓度范围为10-100mm的edta或egta来测试反应的金属依赖性。将条带通过磷光成像可视化,并用imagequant(gehealthcare)定量。切割百分比被确定为产物条带强度与产物条带和未切割的pre-crrna条带两者的总强度的比率,并使用imagequanttl软件(gehealthcare)针对各测量的底物内的背景进行归一化,并使用prism(graphpad)与单相指数关联进行拟合。
产物尺寸映射。通过使用得自ambion的rna酶t1试剂盒比较产物条带到碱水解梯带和rna酶t1消化梯带的凝胶迁移,而生物化学地确定切割产物的长度。对于水解梯带,将15nm的全长rna底物在95℃下于1x碱水解缓冲液(ambion)中孵育5分钟。用1.5xrna上样缓冲液猝灭反应,并冷却至-20℃以立即停止水解。对于rna酶t1梯带,将15nm的全长rna底物在65℃下在1xrna测序缓冲液(ambion)中解折叠。将反应冷却至环境温度,然后向反应中加入1μl的rna酶t1。15分钟后,通过苯酚-氯仿提取终止反应,并加入1.5xrna上样缓冲液进行储存。将水解条带与切割样品平行地在15%变性page上解析,并通过磷光成像可视化。
靶切割测定。在25℃和37℃下在切割缓冲液(20mmhepesph6.8、50mmkcl、5mmmgcl2和5%甘油)中执行靶切割测定。通过加热至65℃达5分钟,以及然后在切割缓冲液中缓慢冷却至环境温度而将crrna指导物预折叠。在加入5'末端标记的靶和/或其它非放射性标记的rna靶底物之前,通常以2:1的蛋白与crrna摩尔比在切割缓冲液中于37℃下执行rnp复合物形成达10分钟。除非另有说明,否则对于所有反应,蛋白、指导物和靶的最终浓度分别为100nm、50nm和大于1nm。用1.5xrna上样染料猝灭反应,并通过15%变性page(0.5xtbe缓冲液)解析。将条带通过磷光成像可视化,并用imagequant(gehealthcare)定量。切割百分比被确定为所有较短产物的总条带化强度相对于未切割条带的比率,并使用imagequanttl软件(gehealthcare)针对各测量的底物内的背景进行归一化,并使用prism(graphpad)与单相指数关联进行拟合。
crrna过滤结合测定。在rna加工缓冲液(20mmhepesph6.8、50mmkcl、5mmmgcl2、10μg/mlbsa、10μg/ml酵母trna、0.01%igepalca-630和5%甘油)中进行过滤结合测定。将lbuc2c2与放射性标记的crrna(小于0.1nm)一起在37℃下孵育1小时。将tufryn、protran和hybond-n+以上面列出的顺序组装到点印迹设备上。将膜用50μl的平衡缓冲液(20mmhepesph6.8、50mmkcl、5mmmgcl2和5%甘油)洗涤两次,然后将样品施加到所述膜上。将膜再次用50μl的平衡缓冲液洗涤,干燥并通过磷光成像可视化。将数据用imagequanttl软件(gehealthcare)定量,并使用prism(graphpadsoftware)与结合等温线拟合。所有实验一式三份地进行。图例中报告了解离常数和相关误差。
电泳迁移率变动测定。为了避免在ssrna结合实验期间解离低浓度的lbuc2c2-dhepn1/dhepn2:crrna复合物,结合反应含有恒定过量的lbuc22c2-dhepn1/dhepn2(200nm)以及不断增加浓度的crrna-a和小于0.1nm的靶ssrna。在c2c2电泳迁移率变动测定(emsa)缓冲液(20mmhepesph6.8、50mmkcl、10μg/mlbsa、100μg/ml酵母trna、0.01%igepalca-630和5%甘油)中进行测定。如上所述在37℃下预形成lbuc2c2-crrna-a复合物10分钟,然后加入5'-放射性标记的ssrna底物,并在37℃下进一步孵育45分钟。然后将样品通过4℃下的8%天然page(0.5xtbe缓冲液)进行解析。将凝胶通过磷光成像而成像,使用imagequanttl软件(gehealthcare)定量,并使用prism(graphpadsoftware)与结合等温线拟合。所有实验一式三份地进行。图例中报告了解离常数和相关误差。
荧光rna检测测定。通过将1μm的lbu-c2c2:c2c2与500nm的crrna在37℃下孵育而预组装lbuc2c2:crrna复合物。然后在存在185nm的rna酶-alert底物(thermo-fisher)、100ng的hela总rna和不断增加量的ssrna(0-1nm)的情况下,将这些复合物稀释至在rna加工缓冲液(20mmhepesph6.8、50mmkcl、5mmmgcl2、10μg/mlbsa、10μg/ml酵母trna、0.01%igepalca-630和5%甘油)中100nm的lbuc2c2:50nm的crrna-λ2。将这些反应在荧光平板读数器中于37℃下孵育长达120分钟,其中每5分钟进行一次荧光测量(λex:485nm;λem:535nm)。通过减去在不存在靶ssrna的情况下进行的反应获得的荧光值来获得经背景校正的荧光值。通过将50nm的rna酶a与185nm的rna酶-alert底物一起孵育来测量最大荧光。对于耦合的pre-crrna加工和rna检测测定,通过将1μm的lbu-c2c2:c2c2与500nm的pre-crrna-a-λ2一起在37℃下孵育20分钟而预组装lbucas9-sgrna复合物,并在存在不断增加量的ssrnaa和ssrnaλ2(各自0-1nm)的情况下如上所述地进行反应。在每种情况下,误差条代表三次独立实验的标准偏差。
实施例8:c2c2家族加工前体crrna转录物以产生成熟crrna
vi型crispr基因座缺乏明显的cas6或cas5d样内切核酸酶或tracrrna。问题是c2c2本身是否具有pre-crrna加工活性。为了测试此点,将来自口腔纤毛菌(lbu)、沙氏纤毛菌(lsh)和斯氏李斯特菌(lse)的重组c2c2同系物(所述重组c2c2同系物来源于c2c2蛋白家族的三个不同分支)在大肠杆菌中表达和纯化。图9a至图9b。
所有三种蛋白在60分钟内切割经5'末端放射性标记的pre-crrna底物,所述底物由全长共有重复序列和20个核苷酸(nt)的间隔区序列组成。图9c.使用t1rna酶梯带和氢氧化物水解梯带映射所有pre-crrna同系物的切割位点;结果表明,加工取决于同系物而发生在所预测的发夹结构上游2nt或5nt的位置处。图9c。
细胞因子可能进一步加工crrna,因为来自大肠杆菌中lsec2c2操纵子和lshc2c2操纵子的异源表达的测序数据发现是相似但不相同的加工事件。用lbuc2c2和含有串联发夹-重复序列阵列的pre-crrna进行切割测定产生了两种对应于两种连续切割事件的产物(图16a至图16f),这与c2c2在加工初级crrna转录物中的作用相符。这些结果表明,在vi型crispr系统中,c2c2同系物催化其相关crrna的成熟。
图9a至图9c.c2c2家族加工前体crrna转录物以产生成熟crrna。图9a.c2c2蛋白的最大似然系统发生树。图22a中提供了包括种质、生物名称和自展值的完整细节。本研究中使用的同系物以黄色突出显示。图9b,三种不同的vi型crispr基因座的图。黑色矩形表示重复元件,而黄色菱形表示间隔区序列。cas1和cas2仅存在于lshc2c2附近的基因组中。图9c,cc2c2介导的对来源于lbuc2c2、lsec2c2和lshc2c2的pre-crrna的切割。oh:碱水解梯带;t1:t1rna酶水解梯带;用100nm的c2c2和大于1nm的pre-crrna执行加工切割反应。右侧描绘了切割的示意图,并且下方示出了所预测的pre-crrna二级结构,其中箭头指示了所映射的c2c2切割位点。
图22a至图22b.c2c2家族的完整系统发生树以及c2c2比对。图22a,c2c2蛋白的最大似然系统发生重建。叶包括gi蛋白数和来源生物;100个重新取样中的自展支持值被呈现用于内部分割。标尺是每个位点的取代。图22b,三种所分析的c2c2的同系物的多重序列比对,坐标基于lbuc2c2。
图23a至图23d.c2c2的纯化和生产。所有c2c2同系物在大肠杆菌(e.coli)中表达为his-mbp融合物,并通过亲和性、离子交换和尺寸排阻色谱的组合进行纯化。通过与tev蛋白酶一起孵育去除ni+亲和标签。色谱级分的代表性sds-page凝胶如上文在(图23a至图23b)中所示。图23c,来自superdex200(16/60)柱的色谱图表明,c2c2作为单峰洗脱、缺乏核酸。图23d,本稿图中使用的所有纯化蛋白的sdspage分析。
图16a至图16f.由c2c2进行的pre-crrna加工是不依赖于间隔区序列的,可以在串联crrna阵列上发生,受pre-crrna的5'侧翼区域中的突变影响并且是不依赖于金属的。a,pre-crrna切割测定的代表性时间过程表明lbuc2c2、lshc2c2和lsec2c2进行的pre-crrna加工具有相似速率。b,单个同源pre-crrna阵列的lsec2c2切割和lshcc2c2切割的切割位点映射。oh:碱水解梯带;t1:t1rna酶水解梯带。用100nm的lbuc2c2和小于1nm的pre-crrna进行切割反应。描绘了切割产物的示意图,其中箭头指示所映射的c2c2切割产物。c,串联pre-crrna阵列的lbucc2c2切割的切割位点映射。oh:碱水解梯带;t1:t1rna酶水解梯带。用100nm的lbuc2c2和小于1nm的pre-crrna进行切割反应。右侧描绘了切割产物的示意图,其中箭头指示映射的c2c2切割产物。d,代表性的lbuc2c2pre-crrna切割时间过程表明,相似的pre-crrna加工速率不依赖于crrna间隔区序列地发生。e,lbuc2c24聚突变体pre-crrna加工数据表明了5'单链侧翼区域对于有效的pre-crrna加工的重要性。60分钟后测量pre-crrna加工的百分比(平均值±标准差,n=3)。f,变性凝胶示出了在一系列不同edta和egta浓度下的加工活性。高浓度的螯合剂改变产物的预期迁移模式
实施例9:lbuc2c2介导的crrna生物发生取决于crispr重复序列的结构和序列。
其它pre-crrna加工酶,诸如cas6和cas5d,以高度特异性的方式识别它们各自相应的crispr重复序列的发夹。确定lbuc2c2指导rna加工的序列和结构要求。用在共有重复序列的茎环或单链侧翼区域中具有突变的pre-crrna执行切割测定(图10a至图10c)。在改变重复区域中的茎长度后,pre-crrna切割活性显著减弱;而重复共有的4碱基对(bp)茎在60分钟后导致85±0.7%的切割,-1bp和+1bp的茎分别导致仅54±1.3%和6±3.8%的切割(图10a)。类似地,茎环的倒置或环长度的缩短降低了加工活性。对包括或靠近易断裂键的连续4nt突变的初步研究完全消除了lbuc2c2加工pre-crrna的能力(图16d)。对完整crrna重复序列的更广泛突变分析揭示发夹两侧的两个不同区域对碱基变化具有显著的敏感性(图10b;图17)。加工活性完全不受二价金属离子螯合剂edta或egta的存在的影响(图10c;图16e)。总之,这些数据表明,c2c2pre-crrna切割是一种不依赖于二价金属离子的过程,所述过程由对前体crispr转录物内的重复序列来源的发夹的结构特异性识别和序列特异性识别的组合的控制。
图10a至图10c.lbuc2c2介导的crrna生物发生取决于crispr重复序列的结构和序列。图10a,在发夹的茎区域和环区域内含有结构突变的pre-crrna的lbuc2c2加工的代表性凝胶。下面列出的加工百分比是在60分钟处针对每种条件定量的(平均值±标准差,n=3)。图10b,图形映射显示了pre-crrna加工对crispr重复序列的依赖性。野生型重复序列如下方所示,其中单独的条代表上文以红色标示的2个核苷酸突变。切割位点用卡通剪刀指示。在60分钟后测量加工百分比(平均值±标准差,n=3)。所测试的突变体的图示发夹可以在图16a至图16f和图17中找到。图2c,通过向标准加工反应条件中加入10-100mm的edta来激发加工反应的二价金属依赖性。示出了60分钟处的反应终点。
图17.pre-crrna双突变对pre-crrna加工活性的影响的详细总结。根据方法,在60分钟后测量pre-crrna加工百分比(平均值±标准差,n=3)。突变的核苷酸以黄色突出显示。关于图形表示,参见图10b。
实施例10:lbuc2c2催化顺式和反式靶上的指导依赖性ssrna降解
成熟后,crrna通常以高亲和力与cas效应蛋白结合,以产生能够进行序列特异性核酸识别的rna指导的监督复合物。为了测试经c2c2加工的crrna的功能,使用lbuc2c2蛋白,其在初始切割实验中表现出最强的活性。对于多种ssrna底物,观察到lbuc2c2仅在与互补crrna结合时有效地切割,表明lbuc2c2:crrna以rna指导的方式切割ssrna(图11b)。此活性在下文中称为在靶或顺式靶切割(图11a)。lbuc2c2介导的顺式切割导致多种产物的梯化,其中切割优先在尿嘧啶残基之前发生,类似于lshc2c2(图18a至图18b)9,这种活性不同于仅在由与crrna的碱基配对所指定的限定区域内催化靶核酸切割的其它ii类crispr效应子(诸如cas9和cpf1)。测试了lbuc2c2反式地充当经crrna激活的非特异性rna内切核酸酶(即,c2c2催化的对未由crrna靶向的rna分子的切割)的能力(图11a)。通过在存在未标记的在靶(与crrna互补的)ssrna的情况下重复非靶切割反应来研究该能力。观察到在这些反式切割条件下5'-标记的非靶rna快速降解(图11b)。该结果表明,识别与crrna的间隔区段互补的靶ssrna激活c2c2以进行快速和非特异性的rna反式降解。对于顺式在靶切割和反式非靶切割观察到相似的rna切割速率和几乎相同的切割产物暗示两种活性中的相同核酸酶中心。此外,这些结果表明,观察为“顺式”活性的情况仅是当只有一种底物可用时发生的反式活性(图18a至图18b)。
图11a-11c.lbuc2c2催化顺式和反式靶上的指导依赖性ssrna降解。图11a,c2c2指导依赖性ssrna降解的两种模式的示意图。图11b,由lbuc2c2切割两种不同的放射性标记的ssrna底物a和b。在37℃预形成100nmc2c2与50nmcrrna的复合物,并在25℃下加入大于1nm的5'-标记的靶rna时开始反应。反式切割反应含有等摩尔(大于1nm)浓度的放射性标记的非指导互补底物和未标记的在靶ssrna。图11c,在存在edta的情况下,激发负载有crrna以及间隔区a的lbuc2c2以获得其在顺式(靶a经标记)和反式(在存在未标记的靶a的情况下,靶b经标记)切割条件下的切割活性。
图18a至图18b.lbuc2c2ssrna靶切割位点映射a,按照方法进行的ssrna靶切割测定,证明lbuc2c2介导对具有相同的间隔区互补序列但不同的5'侧翼序列(具有不同长度和核苷酸组成)的若干个放射性标记的ssrna底物的‘顺式’切割。ssrna底物的序列示出在凝胶的右侧和底部,其中crrna-a和crrna-b的间隔互补序列分别以黄色和红色突出显示。箭头指示所预测的切割位点。为了清楚起见而裁剪了凝胶。应当注意的是,在不同底物上产生的切割产物的模式(例如a.1对比a.2对比a.3)表明,切割位点选择主要由尿嘧啶偏好性驱动,并且表现为明显缺乏‘基于crrna标尺的’切割机制,这与关于其它ii类crispr单效应复合物(诸如cas9和cpf1)<ref?>观察到的相反。有趣的是,针对底物a.0观察到的切割模式提示了对polyg序列的次要偏好性,所述次要偏好性可能可以设想地形成g-四链体或类似的二级结构。b,按照方法,使用一系列覆盖(tile)ssrna靶的长度的crrna进行的lbuc2c2ssrna靶切割测定。在该实验中使用的ssrna底物的序列示出在凝胶下方,其中每个crrna的间隔区互补序列以黄色突出显示。箭头指示所预测的切割位点。同样地,应当注意的是,对于每种crrna,切割产物长度分布是非常相似的,再次表明明显缺乏‘基于crrna标尺的’切割机制。在反应的子集中不存在几种切割产物可以通过ssrna靶上存在结合的c2c2:crrna,从而通过任何顺式(分子内)或反式(分子间)lbuc2c2活性位点在空间上阻断了对尿嘧啶的可及来解释。虽然对lbuc2c2的pfs偏好性的适当分析超出了本研究的范围,但观察到了3'侧翼序列的最小影响。预测的pfs碱基在所测试的每个指导序列旁边的图表中以红色标示。
实施例11:lbuc2c2含有两种不同的核糖核酸酶活性
pre-crrna加工活性以比crrna介导的对靶rna或非互补反式ssrna切割慢80倍的表观速率发生(图12a)。在饱和酶条件下,1h后pre-crrna加工仅达到约85%,而反式底物的rna指导切割的速率由于反应在37℃下于1分钟内完成而几乎是不可测量的(图19a至图19c)。还发现,与pre-crrna加工相对,在存在edta的情况下rna指导的切割被完全消除,表明该活性是二价金属离子依赖性的(图11c)。鉴于pre-crrna加工与rna指导的切割之间在动力学和金属离子依赖性方面的明显差异,有理由认为c2c2可能代表具有两种正交rna切割活性的新型crispr效应蛋白:一种用于crrna成熟,并且另一种用于crrna指导的非特异性rna降解。为了测试该假设,将lbuc2c2的保守hepn结构域内的几个残基(预测所述残基含有rna切割活性位点)进行系统突变;并且评定突变体的pre-crrna加工和rna指导的rna酶活性(图12b)。保守hepn残基(r472a、r477a、r1048a和r1053)的双重突变体和四重突变体保留了强pre-crrna切割活性(图12c)。相比之下,所有hepn突变体都消除了rna指导的切割活性,同时不影响与crrna或ssrna结合的能力(图20a至图20c)。切割速率差异和对点突变和edta的差别敏感性表明,pre-crrna加工和rna指导的rna切割是由c2c2蛋白内的不同催化中心介导的,前者是不依赖于金属的,而后者需要二价金属依赖性hepn结构域。
图12a至图12d.lbuc2c2含有两种不同的核糖核酸酶活性。图4a,由lbuc2c2在37℃下执行的对顺式ssrna靶(黑色)和pre-crrna(青色)切割的定量时间过程数据。指数拟合示出为实线(n=3),并且所计算的一级速率常数(kobs)对于顺式靶切割和pre-crrna切割分别为9.74±1.15和0.12±0.02。图12b,lbuc2c2的结构域架构描绘了hepn突变的位置。图12c和图12d,双重和四重hepn结构域突变体对图12c中的pre-crrna加工和图12d中的ssrna靶向的核糖核酸酶活性。对于两种活性,利用crrna靶向间隔物a,如图11c所示执行顺式和反式切割反应。
图19a至图19ccrrna间隔区长度、反应温度和crrna的5'末端序列对靶rna切割效率的依赖性。图19a,按照方法进行的lbuc2c2ssrna靶切割测定,其中crrna具有16nt、20nt或24nt的间隔区。图19b,按照方法在25℃或37℃下进行的lbuc2c2ssrna靶切割的时间过程。图19c,按照方法进行的lbuc2c2ssrna靶切割时间过程,其中crrna具有不同的5'-侧翼核苷酸突变。突变以红色突出显示。1-2个核苷酸的5'延伸可忽略不计地影响切割效率。相比之下,将侧翼区域缩短至3nt会减慢切割速率。关于代表性平行测定,指数拟合示出为实线。
图20a-20c.lbuc2c2与成熟crrna和靶ssrna的结合数据。图20a,如在方法中所描述地进行过滤结合测定以确定成熟crrna-a与lbuc2c2-wt、lbuc2c2-dhepn1、lbuc2c2-dhepn2或lbuc2c2-dhepn1/dhepn2的结合亲和力。将定量的数据与标准结合同种型拟合。误差条代表三次独立实验的标准偏差。来自三个独立实验的测量的解离常数(平均值±标准差)为27.1±7.5nm(lbuc2c2-wt)、15.2±3.2nm(lbuc2c2-dhepn1)、11.5±2.5nm(lbuc2c2-dhepn2),以及43.3±11.5nm(lbuc2c2-dhepn1/dhepn2)。图20b,如所示,关于lbuc2c2-dhepn1/dhepn2:crrna-a与‘在靶’assrna或‘脱靶’bssrna之间的结合反应的代表性电泳迁移率变动测定。如方法中所述进行三次独立实验。为了清楚起见而裁剪了凝胶。图20c,将来自(b)的定量结合数据与标准结合同种型拟合。误差条代表三次独立实验的标准偏差。从三个独立实验测量的解离常数(平均值±标准偏差)对于ssrnaa为1.62±0.43nm并且对于ssrnab为未检测到的(>>10nm)。
实施例12:c2c2提供对复杂混合物中转录物的灵敏视觉检测。
c2c2对反式底物的强rna刺激切割可用作检测转录物集合内的特定rna的手段。迄今为止,最灵敏的rna检测策略需要多轮基于聚合酶和/或逆转录酶的扩增。作为替代方案,测试了c2c2的rna指导的反式内切核酸酶活性是否可用于切割用荧光团-猝灭剂标记的报告rna底物,从而导致在靶rna触发的rna酶激活后荧光增加(图13a)。简而言之,使lbuc2c2负载有靶向细菌噬菌体λ的crrna,并测试其检测掺入hela细胞总rna的相应λssrna靶的能力。如果lbuc2c2成功检测到互补靶,则反式切割活性应被激活以切割报告物,从而从猝灭剂中释放荧光团。加入少至1-10pm的互补λ靶rna后,在30分钟后观察到显著的crrna特异性荧光增加(图13b和图21)。单独使用c2c2:crrna复合物或在存在crrna和非互补靶rna的情况下的对照实验导致相对于rna酶a阳性对照的可忽略不计的荧光增加(图13b和图21)。在10pm的λ靶rna浓度下,预测仅约0.02%的c2c2:crrna复合物处于活性状态,但观察到的荧光信号反映了取决于rna靶而对报告rna底物的约25-50%切割,表明了lbuc2c2具有强大的多转换酶促活性。因此,即使在极低的活性蛋白水平下,crrna指导的反式切割也是有效并且可检测的。
鉴于c2c2加工其自身的pre-crrna,测试了pre-crrna加工和rna检测是否可以在单一反应中组合。为了测试该想法,设计了含有与靶rnaa和λ2互补的间隔区的串联crrna重复序列,并测试了它们检测掺入hela总rna中的不断减少量的a和λ2rna的能力。观察到了显著的荧光增加,这在量值和灵敏度方面与使用成熟crrna的实验相似(图13b、图13c),表明串联的pre-crrna可以由c2c2成功加工并用于rna靶向。这些数据突出了在一个测定中使用单一串联pre-crrna转录物对单个和/或多个rna分子进行多重rna检测的可能性。
不受理论束缚,提出当经由与crrna的碱基配对而在宿主细胞内检测到侵入性转录物时,c2c2被激活以反式地进行混杂的rna切割(图13d)。
图13a至图13d.c2c2提供对复杂混合物中转录物的灵敏视觉检测。图13a,使用猝灭的荧光rna报告物通过c2c2进行的rna检测方法的示意图。在互补的靶rna结合后,c2c2催化报告rna的降解,从而导致荧光信号的积累。图13b,在存在100ng总rna的情况下,在30分钟后通过c2c2对不同浓度的靶rna的荧光信号进行的定量。rna酶a示出作为阳性rna降解对照。数据显示为n=3个独立实验的平均值±标准差。图13c,串联pre-crrna加工还使得能够进行rna检测。数据显示为n=3个独立实验的平均值±标准差。图13d,vi型crispr途径的模型突显了c2c2的两种核糖核酸酶活性。
图21.rna酶检测测定λ2-ssrna的时程。在存在不断增加量的λ2ssrna(0-1nm)的情况下,将lbuc2c2:crrna-λ2与rna酶-alert底物(thermo-fisher))和100nghela总rna在37℃孵育120分钟。每5分钟进行荧光测量。1nm的λ2ssrna反应在可以测量第一个时间点之前达到饱和。误差条代表三次独立实验的标准偏差。
实施例13
材料和方法
mec2c2系统发生和候选选择。使用具有protgammalg进化模型和100个自展抽样的raxml计算c2c2最大似然系统发生。通过mafft使用‘einsi’方法比对序列。
c2c2蛋白的生产和纯化.使用从integrateddnatechnologies订购的合成gblock组装用于蛋白纯化的表达载体。将密码子优化的c2c2基因组序列用his6-mbp-tev切割位点进行n末端标记,其中表达由t7启动子驱动。经由野生型c2c2构建体的定点诱变来克隆突变蛋白。将表达载体转化到于37℃下在2xyt肉汤中生长的rosetta2大肠杆菌细胞中。将大肠杆菌细胞在对数期期间用0.5m的itpg诱导,并将温度降至16℃以过夜表达his-mbp-c2c2。随后收获细胞,重悬于裂解缓冲液(50mmtris-hclph7.0、500mmnacl、5%甘油、1mmtcep、0.5mmpmsf、以及不含edta的蛋白酶抑制剂(roche))中,并通过超声裂解,并且将裂解物通过离心澄清。通过金属离子亲和色谱分离出可溶性his-mbp-c2c2,将含蛋白的洗脱物与tev蛋白酶一起在4℃下孵育过夜,同时透析到离子交换缓冲液(50mmtris-hclph7.0、250mmkcl、5%甘油、1mmtcep)中以切除his6-mbp标签。将切割的蛋白上样到hitrapsp柱上,并以线性kcl(0.25-1.5m)梯度洗脱。将阳离子交换色谱级分汇集,并用30kd的截留浓缩管(thermofisher)浓缩。将c2c2蛋白经由在s200柱上进行尺寸排阻色谱而进一步纯化,并储存在凝胶过滤缓冲液(20mmtris-hclph7.0、200mmkcl、5%甘油、1mmtcep)中以供后续酶测定。将表达质粒由addgene保藏。
产生rna。除了crrnaaes461是订购合成的(integrateddnatechnologies)之外,本研究中使用的所有rna都是在体外转录的[参见图33]。如前所述进行体外转录反应,具有以下改变:t7聚合酶浓度降至10μg/ml,并且utp浓度降至2.5mm。将转录在37℃下孵育1-2小时以减少核苷酸的非模板加入,并经由用dna酶i在37℃下处理0.5-1小时来进行猝灭。将转录反应通过15%变性聚丙烯酰胺凝胶电泳(page)纯化,并将所有rna重悬于切割缓冲液(20mmhepesph6.8、50mmkcl、5mmmgcl2和5%甘油)中。对于放射性实验,在放射性标记之前用小牛肠磷酸盐(newenglandbiolabs)去除5'三磷酸酯,然后如前所述使用t4多核苷酸激酶(newenglandbiolabs)和[γ-32p]-atp(perkinelmer)对ssrna底物进行5'末端标记。
pre-crrna加工测定。在37℃下在rna加工缓冲液(20mmhepesph6.8、50mmkcl、5mmmgcl2、10μg/mlbsa、10μg/mltrna、0.05%igepalca-630和5%甘油)以及相对于5'-标记的pre-crrna100倍摩尔过量的c2c2(最终浓度分别为100nm和小于1nm)中执行pre-crrna切割测定。除非另有说明,否则在60分钟后用1.5xrna上样染料(100%甲酰胺、0.025w/v%溴酚蓝和200μgml-1的肝素)猝灭反应。猝灭后,将反应在95℃变性5分钟,然后通过12%或15%变性page(0.5xtbe缓冲液)解析。通过向反应缓冲液中加入浓度范围为10-100mm的edta或egta来测试反应的金属依赖性。将条带通过磷光成像可视化,并用imagequant(gehealthcare)定量。切割百分比被确定为产物条带强度与产物条带和未切割的pre-crrna条带两者的总强度的比率,并使用imagequanttl软件(gehealthcare)针对各测量的底物内的背景进行归一化,并使用prism(graphpad)与单相指数关联进行拟合。
产物尺寸映射和3'末端部分鉴别。通过使用得自ambion的rna酶t1试剂盒比较产物条带到碱水解梯带和rna酶t1消化梯带的凝胶迁移,而生物化学地确定切割产物的长度。对于水解梯带,将15nm的全长rna底物在95℃下于1x碱水解缓冲液(ambion)中孵育5分钟。用1.5xrna上样缓冲液猝灭反应,并冷却至-20℃以立即停止水解。对于rna酶t1梯带,将15nm的全长rna底物在65℃下在1xrna测序缓冲液(ambion)中解折叠。将反应冷却至环境温度,然后向反应中加入1u的rna酶t1(ambion)。15分钟后,通过苯酚-氯仿提取终止反应,并加入1.5xrna上样缓冲液进行储存。将水解条带与切割样品平行地在15%变性page上解析,并通过磷光成像可视化。对于3'末端部分鉴别,将来自加工反应的产物与10u的t4多核苷酸激酶(newenglandbiolabs)一起在37℃下于加工缓冲液中孵育1小时。用1.5xrna上样缓冲液猝灭反应,在20%变性page上解析,并通过磷光成像可视化。
小rna测序分析。从sra运行srr3713697、srr3713948和srr3713950下载来自smakov等人的rna读段。使用具有以下选项的bowtie2将配对末端读段局部映射到参考序列:“--重排--非常快速地定位--定位”。然后使用具有“-m0-p两者”选项的mapped.py过滤映射以仅保留不包含错配的比对。所得映射的bam文件是可用的。使用geneious可视化读段覆盖,并使用prism(graphpad)绘图。
靶切割测定。在25℃或37℃下在切割缓冲液(20mmhepesph6.8、50mmkcl、5mmmgcl2和5%甘油)中执行靶切割测定。通过加热至65℃达5分钟,以及然后在切割缓冲液中缓慢冷却至环境温度而将crrna指导物预折叠。在加入5'末端标记的靶和/或其它非放射性标记的rna靶底物之前,通常以2:1的蛋白与crrna摩尔比在切割缓冲液中于37℃下执行c2c2:crrna复合物形成达10分钟。除非另有说明,否则对于所有反应,蛋白、指导物和靶的最终浓度分别为100nm、50nm和小于1nm。用1.5xrna上样染料猝灭反应,并通过15%变性page(0.5xtbe缓冲液)解析。将条带通过磷光成像可视化,并用imagequant(gehealthcare)定量。切割百分比被确定为所有较短产物的总条带化强度相对于未切割条带的比率,并使用imagequanttl软件(gehealthcare)针对各测量的底物内的背景进行归一化,并使用prism(graphpad)与单相指数关联进行拟合。
crrna过滤结合测定。在rna加工缓冲液(20mmhepesph6.8、50mmkcl、5mmmgcl2、10μg/mlbsa、10μg/ml酵母trna、0.01%igepalca-630和5%甘油)中进行过滤结合测定。将lbuc2c2与放射性标记的crrna(小于0.1nm)一起在37℃下孵育1小时。将tufryn、protran和hybond-n+以上面列出的顺序组装到点印迹设备上。将膜用50μl的平衡缓冲液(20mmhepesph6.8、50mmkcl、5mmmgcl2和5%甘油)洗涤两次,然后将样品施加到所述膜上。将膜再次用50μl的平衡缓冲液洗涤,干燥并通过磷光成像可视化。将数据用imagequanttl软件(gehealthcare)定量,并使用prism(graphpadsoftware)与结合等温线拟合。所有实验一式三份地进行。图例中报告了解离常数和相关误差。
电泳迁移率变动测定。为了避免在ssrna结合实验期间解离低浓度的lbuc2c2-dhepn1/dhepn2:crrna复合物,结合反应含有恒定过量的lbuc22c2-dhepn1/dhepn2(200nm),以及不断增加浓度的crrna-a和小于0.1nm的靶ssrna。在c2c2emsa缓冲液(20mmhepesph6.8、50mmkcl、10μg/mlbsa、100μg/ml酵母trna、0.01%igepalca-630和5%甘油)中进行测定。如上所述在37℃下预形成lbuc2c2-crrna-a复合物10分钟,然后加入5'-放射性标记的ssrna底物,并在37℃下进一步孵育45分钟。然后将样品通过4℃下的8%天然page(0.5xtbe缓冲液)进行解析。将凝胶通过磷光成像而成像,使用imagequanttl软件(gehealthcare)定量,并使用prism(graphpadsoftware)与结合等温线拟合。所有实验一式三份地进行。图例中报告了解离常数和相关误差。
荧光rna检测测定。通过将1μm的lbu-c2c2:c2c2与500nm的crrna在37℃下孵育10分钟而预组装lbuc2c2:crrna复合物。然后在存在185nm的rna酶-alert底物(thermo-fisher)、100ng的hela总rna和不断增加量的靶60ntssrna(0-1nm)的情况下,将这些复合物稀释至在rna加工缓冲液(20mmhepesph6.8、50mmkcl、5mmmgcl2、10μg/mlbsa、10μg/ml酵母trna、0.01%igepalca-630和5%甘油)中100nm的lbuc2c2:50nm的crrna-λ2。将这些反应在荧光平板读数器中于37℃下孵育长达120分钟,其中每5分钟进行一次荧光测量(λex:485nm;λem:535nm)。通过减去在不存在靶ssrna的情况下进行的反应获得的荧光值来获得经背景校正的荧光值。通过将50nm的rna酶a与185nm的rna酶-alert底物一起孵育来测量最大荧光。对于用人总rna中的β-肌动蛋白mrna测量crrna-actb介导的lbuc2c2激活,通过将1μm的lbuc2c2与500nm的crrna-actb一起在37℃下孵育10分钟而预组装lbucas9:crrna复合物,并在上述条件下在存在不断增加量(0-1μg)的hela细胞总rna或大肠杆菌总rna(作为阴性对照)的情况下进行反应。将这些反应在荧光平板读数器中于37℃下孵育长达180分钟,其中每5分钟进行一次荧光测量(λex:485nm;λem:535nm)。通过减去在不存在靶ssrna的情况下进行的反应获得的荧光值来获得经背景校正的荧光值。对于耦合的pre-crrna加工和rna检测测定,通过将1μm的lbuc2c2与500nm的pre-crrna-a-λ2一起在37℃下孵育20分钟而预组装lbucas9-crrna复合物,并在存在不断增加量的ssrnaa和ssrnaλ2(各自0-1nm)的情况下如上所述地进行反应。在每种情况下,误差条代表三次独立实验的标准偏差。
总rna中的背景切割。如先前关于荧光rna检测测定所述地组装lbuc2c2:crrnaλ4复合物。在存在含有和不含10nm的λ4ssrna靶的3μg总rna的情况下,将复合物在rna加工缓冲液中孵育。2小时后,通过trizol提取和乙醇沉淀分离rna。使用制造商的方案,使用小rna分析试剂盒(smallrnaanalysiskit)(agilent)在bioanalyzer2100(agilent)上解析重悬样品的rna片段尺寸分布。将荧光强度曲线在prism(graphpadsoftware)中针对曲线覆盖归一化。
结果
vi型crispr基因座缺乏明显的cas6或cas5d样内切核酸酶或tracrrna。通过使用来自c2c2蛋白家族的三个不同分支的纯化的重组c2c2蛋白同系物(图24a至图24b和图28和图29),在此呈现的数据显示所有三种c2c2酶切割经5'末端放射性标记的pre-crrna底物,所述底物由全长共有重复序列和20个核苷酸(nt)的间隔区序列组成(图24c)。映射每个pre-crrna:c2c2同源对的切割位点,揭示加工取决于c2c2同系物而发生在所预测的重复序列发夹结构上游2个或5个核苷酸的位置处(图24c、图30a)。令人惊讶的是,在生物化学上映射的5'-切割位点与之前报道的沙氏纤毛菌(lshc2c2)或斯氏李斯特菌(lsec2c2)pre-crrna的切割位点不一致。对shmakov等人的rna测序数据集表的重新分析表明体内切割位点与此处报道的体外位点一致(图30b至图30i)。此外,使用来自口腔纤毛菌(lbuc2c2)的c2c2和包含串联发夹-重复序列阵列的较大pre-crrna的切割测定产生了来源于两种单独的切割事件的两种产物(图31a),这与c2c2在加工从vi型crispr基因座产生的前体crrna转录物中的作用一致。
为了理解c2c2指导rna加工的底物要求和机制,产生在共有重复序列的茎环或单链侧翼区域中具有突变的pre-crrna,并测试其由lbuc2c2进行加工的能力(图25)。数据显示,在改变重复区域中的茎长度时,c2c2催化的切割减弱(图25a)。茎环的倒置或环长度的缩短也降低了c2c2的加工活性,而包括或靠近易断裂键的连续4nt突变完全消除了所述加工活性(图31b)。对全长crrna重复序列的更广泛突变分析揭示发夹任一侧的两个不同区域对碱基变化具有显著的敏感性(图25b)。相比之下,加工动力学不存在对间隔区序列的依赖性(图31c)。对发夹的两个侧翼区域的这种敏感性使人联想到许多cas6酶和cas5d酶所需的序列和结构基序。相比之下,cpf1不具有对3′发夹侧翼区域的任何依赖性,因为可变间隔区域邻接发夹茎。
lbuc2c2的机理研究揭示,加工活性不受二价金属离子螯合剂edta或egta存在的影响(图25c),表明了不依赖于金属离子的rna水解机制。不依赖于金属离子的rna水解的典型特征是分别在crrna切割产物的5'和3'半部上形成2',3'-环磷酸酯和5'-氢氧化物。为了确定c2c2加工的底物的末端基团化学特性,将5'侧翼产物与t4多核苷酸激酶一起进一步孵育,此举去除2',3'-环磷酸酯以留下3'-羟基。在激酶处理后观察到了5'侧翼产物的改变的变性凝胶迁移,这与3'磷酸基团的去除一致(图31d)。c2c2的pre-crrna加工活性的二价金属离子非依赖性与cpf1的二价金属离子依赖性形成鲜明对比,cpf1是唯一显示为执行指导加工的其它单蛋白crispr效应物。总的来说,这些数据表明,c2c2催化的pre-crrna切割是一种不依赖于二价金属离子的过程,该过程可能使用一般的酸碱催化机制。
成熟后,crrna通常以高亲和力与cas效应蛋白结合,以产生能够进行序列特异性核酸识别的rna指导的监督复合物。与先前使用lshc2c2的工作一致,lbuc2c2仅在此类底物可与crrna中的互补序列碱基配对时催化有效的靶rna切割(图32至图34)。鉴于观察到的c2c2混杂切割模式(图33),测试了lbuc2c2反式地充当经crrna激活的非特异性rna内切核酸酶的能力(图32b)。与顺式执行的非靶切割实验形成鲜明对比并且与先前对lshc2c2的观察一致,观察到了非靶rna的反式快速降解(图32b)。该结果显示,靶识别激活c2c2进行rna的一般非特异性降解。重要的是,对于相同rna底物的顺式在靶切割和反式非靶切割观察到的相似的rna切割速率和几乎相同的切割产物暗示两种活性中的相同核酸酶中心(图32b)。
值得注意的是,crrna介导的靶ssrna切割以比pre-crrna加工快约80倍的速率发生(图26a),并且与pre-crrna加工不同,在存在edta的情况下rna指导的靶切割严重减少,表明该活性是二价金属离子依赖性的(图26a,(图32c、图34)。鉴于这些明显的差异,有理由认为c2c2可能具有两种正交rna切割活性:一种用于crrna成熟,并且另一种用于crrna指导的非特异性rna降解。为了测试该假设,将lbuc2c2的保守hepn基序内的几个残基进行系统突变;并且评定突变体的pre-crrna加工和rna指导的rna酶活性(图26、图34d)。保守hepn残基(r472a、r477a、r1048a和r1053)的双重突变体和四重突变体保留了强pre-crrna切割活性(图26c)。相比之下,所有hepn突变体都消除了rna指导的切割活性,同时不影响与crrna或ssrna结合的能力(图34d、图35)。
接下来,寻找可以在不破坏靶rna切割的情况下消除pre-crrna加工活性的突变。鉴于不能预测超出hepn基序的任何其它潜在的rna酶基序,并且c2c2蛋白与cpf1没有同源性,所以整个lbuc2c2中的荷电残基被系统地突变。精氨酸残基(r1079a)被鉴别为在突变后导致严重减弱的pre-crrna加工活性(图26c)。该c2c2突变酶保留了crrna结合能力以及rna靶切割活性(图35d、图26d)。总的来说,结果显示c2c2蛋白内的不同活性位点催化pre-crrna加工和rna指导的rna切割。
接下来测试c2c2对反式底物的强rna刺激切割是否可用作检测转录物集合内的特定rna的手段。虽然已经开发了许多基于聚合酶的方法用于rna扩增和随后的检测,但是基本上没有方法能够直接检测靶rna而无需对每个新rna靶进行大量工程化或严格的设计约束。作为一种易于编程的替代方案,测试了c2c2的rna指导的反式内切核酸酶活性是否可用于切割用荧光团-猝灭剂标记的报告rna底物,从而导致在靶rna触发的rna酶激活后荧光增加(图27a)。使lbuc2c2负载有靶向细菌噬菌体λ的crrna,并测试其检测掺入hela细胞总rna的相应λssrna靶的能力。加入少至1-10pm的互补λ靶rna后,在30分钟内发生了显著的crrna特异性荧光增加(图27b和图36a)。单独使用c2c2:crrna复合物或在存在crrna和非互补靶rna的情况下的对照实验导致相对于rna酶a阳性对照的可忽略不计的荧光增加(图27b和图36a)。应当注意的是,在10pm的λ靶rna浓度下,预测仅约0.02%的c2c2:crrna复合物处于活性状态,但观察到的荧光信号反映了取决于rna靶而对报告rna底物的约25-50%切割。这些反应中背景rna的片段尺寸解析揭示了显著的降解,即使是对高度结构化的trna也是如此(图36b)。由于报告rna切割在存在大量过量的未标记rna的情况下发生,因此推断lbuc2c2是一种强多转换酶,其能够识别每靶rna至少104个转换。因此,与先前的观察结果不同,即使在极低的激活蛋白水平下,crrna指导的反式切割也是有效并且可检测的。
为了扩展该lbuc2c2rna检测系统,将crrna设计为靶向内源性β-肌动蛋白mrna。相对于大肠杆菌总rna,在存在人总rna的情况下观察到了可测量的荧光增加,证明了该方法的特异性(图27c)。此外,鉴于c2c2加工其自身的指导物,所以通过设计含有与靶rnaa和λ2互补的间接区的串联crrna重复序列而将pre-crrna加工和rna检测组合在单一反应中。在检测测定中将lbuc2c2与该未加工的串联指导rna一起孵育产生了显著的荧光增加,这在量值和灵敏度方面与使用成熟crrna的实验相似(图27b、图27d)。总的来说,这些数据突显了利用c2c2的两种不同rna酶活性进行一系列生物技术应用的令人兴奋的机会(图27e)。
在细菌中,c2c2可能作为病毒rna的前哨工作。不受理论束缚,提出当经由与crrna的碱基配对而在宿主细胞内检测到侵入性转录物时,c2c2被激活以反式地进行混杂的rna切割(图27e)。作为防御机制,这与真核生物中的rna酶l和半胱天冬酶系统具有惊人的相似性,由此细胞信号分别在宿主细胞内引发混杂的核糖核酸降解或蛋白水解性降解,从而导致细胞凋亡。虽然iii型crispr系统的rna靶向机制通常导致原型间隔区-指导双链体内的rna切割,但相关核酸酶csx1和csm6的最近实例提供了在vi型系统与多组分iii型推断复合物之间令人信服的相似性。
在此描述的数据显示,crispr-c2c2蛋白代表一类新的酶,其能够具有两种单独的rna识别和切割活性。有效的pre-crrna加工需要crispr重复序列内的序列和结构基序,这阻止了非内源性crrna负载并有助于降低该有效rna酶的潜在毒性。c2c2和v型crispr效应蛋白cpf1的完全不同的pre-crrna加工机制表明,每个蛋白家族已经会聚于独立的活性,所述独立的活性涵盖它们各自相应的crispr途径的加工和干扰功能。此外,c2c2的两种不同催化能力可以一起用于rna检测,因为激活c2c2以将每个检测到的靶rna切割成数千个反式rna使得能够实现有效的信号放大。c2c2从阵列加工其自身的指导rna的能力允许使用组织特异性polii启动子来指导表达,以及用于广泛应用的多靶点混合检测(targetmultiplexing)。c2c2酶在细菌适应性免疫中因其双rna酶活性而是独特的,并突显了利用crispr蛋白在细胞和无细胞系统中进行精确核酸操纵的实用性。
实施例14
在lbuc2c2中鉴别出了另外的氨基酸位置,所述另外的氨基酸位置在突变时导致严重减弱的pre-crrna加工活性(图37)。氨基酸位置包括r1079(例如,r1079a)、r1072(例如,r1072a)和k1082(例如,k1082a)。
实施例15
cas13a酶包含两种不同的官能团,这两种不同的官能团识别正交的crrna组并具有不同的ssrna切割特异性。cas13apre-crrna加工对于ssrna切割不是必需的,但是其增强了ssrna对在crispr阵列内部编码的crrna的靶向。定义了两种cas13a蛋白亚家族,这些亚家族可以并行操作,以用于在细菌中进行rna检测和破坏两者和用于诊断应用。
材料和方法
cas13a系统发生和重复保守分析。使用具有protgammalg进化模型和100个自展值的raxml(stamatakis,bioinformatics.2014年5月1日;30(9):1312-3)计算cas13a最大似然系统发生。蛋白进化枝α和β被定义为自展值大于90的分支点,表明在具有共同祖先方面的高置信度。其余的蛋白被标记为模糊的祖先,因为它们之间的系统发生关系是低置信度的,反映为自展值小于90。通过mafft使用‘einsi’方法(katoh和standley,molbiolevol.2013年4月;30(4):772-80)比对序列。选择候选者以代表cas13a蛋白树的每个主要分支。对所有非冗余同系物(图41)和候选蛋白进行比对。使用needleman-wunsch算法,经由embl-ebi上的needle工具计算逐对相似性得分而进行crispr-rna(crrna)重复序列的比较(mcwilliam等人,nucleicacidsres.2013年7月;41(webserverissue):w597-600)。使用相似性得分矩阵在r中进行crisprcrrna的分层聚类。
cas13a蛋白的表达和纯化。使用从integrateddnatechnologies(idt)订购的合成gblock组装用于蛋白纯化的表达载体。将密码子优化的cas13a基因组序列用his6-mbp-tev切割位点序列进行n末端标记,其中表达由t7启动子驱动。经由野生型cas13a构建体的定点诱变来克隆突变蛋白。如上文实施例7和13中所述地进行所有同系物的纯化。简而言之,将表达载体转化到于37℃下在2xyt肉汤中生长的rosetta2de3或bl21大肠杆菌细胞中,在对数中期用0.5mm的itpg诱导,然后将温度转变至16℃以进行过夜表达。将细胞沉淀物重悬于裂解缓冲液(50mmtris-clph7.0、500mmnacl、5%甘油、1mmtcep、0.5mmpmsf、以及不含edta的蛋白酶抑制剂(roche))中,通过超声裂解,并且通过以15,000g离心澄清。通过金属离子亲和色谱分离出可溶性his6-mbp-tev-cas13a,并且为了切除his6-mbp标签,将含蛋白的洗脱物与tev蛋白酶一起在4℃下孵育过夜,同时透析到离子交换缓冲液(50mmtris-clph7.0、250mmkcl、5%甘油、1mmtcep)中。将切割的蛋白上样到hitrapsp柱(gehealthcare)上,并以线性kcl(0.25-1.5m)梯度洗脱。将含有cas13a的级分汇集、浓缩,并在s200柱(gehealthcare)上于凝胶过滤缓冲液(20mmtris-clph7.0、200mmkcl、5%甘油、1mmtcep)中经由尺寸排阻色谱进一步纯化,随后在-80℃下储存。使用该方案纯化所有同系物,不同之处在于lwacas13a结合到hitrap肝素柱而不是sp柱,并且尺寸排阻色谱步骤由于样品在离子交换后的足够纯度而被省略。将所有表达质粒由addgene保藏。
体外rna转录。使用先前描述的方法(sternberg等人,rna.2012年4月;18(4):661-72)并且如在以上实施例中所述地在体外转录所有pre-crrna、成熟crrna和靶。简而言之,所有底物都从单链dna寡核苷酸模板(idt)转录,不同之处在于成熟crrna需要非gr的5'末端。对于这些成熟crrna,使用重叠pcr产生紧接在成熟crrna序列上游的含有锤头型核酶序列的t7聚合酶模板,然后纯化以用作t7转录的模板(关于序列,参见图50(表5))。lbu六聚crispr阵列体外转录模板由geneart(thermofisher)合成以作为质粒。将t7启动子-crispr阵列区域在用作t7转录的模板之前进行pcr扩增和纯化。使用15%的尿素-page纯化所有转录的rna,不同之处在于使用6%的尿素-page纯化阵列。随后用小牛碱性磷酸酶处理所有rna以去除5'磷酸酯。使用先前描述的方法(sternberg等人,rna.2012年4月;18(4):661-72)并且如在以上实施例中所述地执行放射性标记。由idt合成a、c、g和u均聚物,以及用于反式ssrna切割的荧光标记的rna报道基因。在放射性标记后使用25%的尿素-page纯化均聚物,以降低底物异质性。
放射性标记的ssrna核酸酶测定。在37℃下在rna加工缓冲液(20mmhepes-naph6.8、50mmkcl、5mmmgcl2、10μg/mlbsa、10μg/mltrna、0.05%igepalca-630和5%甘油)以及相对于5'-标记的pre-crrna100倍摩尔过量的cas13a(最终浓度分别为100nm和小于1nm)中执行pre-crrna加工测定。除非另有说明,否则在60分钟后用1.5xrna上样染料(100%甲酰胺、0.025%溴酚蓝和200μgml-1肝素)猝灭反应。猝灭后,将反应在95℃变性5分钟,然后通过15%变性page(0.5xtbe缓冲液)解析。在37℃下在切割缓冲液(20mmhepes-naph6.8、50mmkcl、5mmmgcl2和5%甘油)中执行靶切割测定。通常,在加入5'标记的靶和/或其它非放射性标记的rna靶底物之前,以2:1的蛋白与crrna摩尔比在切割缓冲液中于37℃下执行cas13a:crrna复合物形成达60分钟。除非另有说明,否则最终浓度为100nmcas13a、50nmcrrna或pre-crrna、50nm的与crrna互补的靶ssrna(此后称为“激活物”)和小于1nm的反式ssrna靶。将所有条带通过磷光成像(typhoon,gehealthcare)可视化,并用imagequant(gehealthcare)定量。对于pre-crrna加工,切割百分比被确定为产物条带强度与产物条带和未切割的pre-crrna条带两者的总强度的比率,并针对各测量的底物内的背景进行归一化。对于反式ssrna切割反应,切割百分比被确定为小于靶的所有片段与泳道内总强度的比率,并针对每个底物内的背景进行归一化。随后使用prism7(graphpad)将这些数据与单指数衰减拟合,并且在图例中报告切割率。
荧光ssrna核酸酶测定。如上所述,在切割缓冲液中组装cas13a:crrna复合物。加入150nm的rna酶alert报告物(idt)和各种最终浓度(0-1μm)的ssrna激活物以引发反应。值得注意的是,这些反应是在没有竞争trna或总rna的情况下,以更准确地测量反式切割活性。将这些反应在荧光平板读数器(tecaninfiniteprof2000)中于37℃下孵育长达120分钟,其中每5分钟进行一次荧光测量(λex:485nm;λem:535nm)。通过减去在不存在靶ssrna激活物的情况下进行的反应获得的荧光值来获得经背景校正的荧光值。为了确定同系物敏感度和阵列加工效果,使用prism7(graphpad)将经背景校正的值与单指数衰减进行拟合,并且将所计算的速率和它们与n=3的相关标准偏差一起进行绘图。为了比较非同源crrna指导的反式ssrna切割,替代地计算初始反应速率,这是由于整个数据集的荧光平台值差异。然后将速率相对于同源crrna进行缩放,以使速率针对同系物归一化。参见图52、图53和图54,分别给出了关于归一化值的表7、表8和表9。对于荧光均聚物ssrna报告物研究,使用标准条件在37℃下预孵育cas13a:crrna复合物60分钟。在将反应放入平板读数器之前,立即加入激活物ssrna和200nm的荧光ssrna报告物以引发反应。对于具有荧光均聚物ssrna报告物的含lbu-cas13a和lba-cas13a的样品,分别使用10pm和1nm的激活物。对于pre-crrna阵列实验,首先将300nm的cas13a与50nm的pre-crrna阵列在切割缓冲液中孵育1小时,以实现阵列的结合和加工。对于每种间隔区序列,以生物一式三份的方式将100pm的每种ssrna激活物与150nm的rna酶alert报告物(idt)一起加入以引发反应。使用单指数衰减、使用prism7(graphpad)来计算表观速率,并将所计算的速率与它们的相关标准偏差一起进行绘图。
crrna过滤结合测定。如上文实施例中所述地进行过滤结合测定。简而言之,将cas13a和放射性标记的crrna在37℃下在rna加工缓冲液(20mmhepes-naph6.8、50mmkcl、5mmmgcl2、10μg/mlbsa、10μg/ml酵母trna、0.01%igepalca-630和5%甘油)中孵育1小时。将tufryn、protran和hybond-n+以上面列出的顺序组装到点印迹设备上。将膜用50μl的平衡缓冲液(20mmhepes-naph6.8、50mm的kcl、5mmmgcl2和5%甘油)洗涤两次,然后将样品施加到所述膜上。将膜再次用50μl的平衡缓冲液洗涤,干燥并通过磷光成像可视化。将数据用imagequanttl软件(gehealthcare)定量,并使用prism(graphpadsoftware)与结合等温线拟合。所有实验一式三份地进行。图例中报告了解离常数和相关误差。
结果
大多数cas13a同系物具有pre-crrna加工活性
为了探索cas13a蛋白的功能多样性,对于它们从同源pre-crrna产生成熟crrna的能力,比较来自蛋白家族树的10种同系物的pre-crrna加工活性(图38a;图39)。与上面讨论的三种同系物类似,另外七种cas13a酶具有crrna成熟活性(图38b和图38c)。迄今为止测试的十一种cas13a蛋白中只有一种在广泛的测定条件下未表现出对其同源pre-crrna的可检测的切割(hhecas13a)(图38;图39a)。在加工其天然crrna的同系物中,除了lshcas13a之外全部都在保守crrna-重复序列发夹上游四个核苷酸的磷酸二酯键处进行切割(图38b和图38c)。这些结果显示了在vi-a型crispr系统中cas13a介导的crrna生物发生活性的强保守性。
大多数cas13a酶中的保守crrna成熟中心
先前的研究已经暗示cas13a的两个不同区域负责pre-crrna加工。由lshcas13a晶体结构建立的一般cas13a蛋白架构由n末端结构域和由两个螺旋结构域分开的两个hepn(高等真核生物和原核生物核苷酸结合)结构域组成(图40a)(liu等人,cell2017年1月12日;168(1-2):121-134.e12)。对于lbucas13a,hepn2结构域内单个残基(r1079)的突变足以显著降低pre-crrna加工活性。相比之下,位于螺旋1结构域中的两个位置r438和k441处的突变显示为减少由lshcas13a进行的pre-crrna切割(liu等人,cell2017年1月12日;168(1-2):121-134.e12)。虽然这两个区域都可能参与pre-crrna加工,但尚不清楚由lbucas13a进行的加工是否被螺旋1结构域突变抑制,以及哪个结构域主要负责跨cas13a蛋白家族的crrna成熟。
跨19个cas13a同系物检查了螺旋1结构域和hepn2结构域两者的保守性。先前报道的比对(liu等人,cell2017年1月12日;168(1-2):121-134.e12;shmakov等人,molcell.2015年11月5日;60(3):385-97)在螺旋1域区域中有冲突,表明了同系物之间关系在该结构域方面的高度模糊性。观察到了参与pre-crrna加工的螺旋1结构域的比对中的最小保守;相比之下,在hepn2结构域内存在一致的保守(图40b至图40c;图41)。唯一的pre-crrna加工缺陷型同系物hhecas13a,在两个结构域中保持大部分保守的荷电残基,表明蛋白或重复序列的其它部分可阻止pre-crrna切割。在该研究中使用的同系物中,lshcas13a在hepn2结构域中是最不同的,从而潜在地解释了该同系物的替代催化结构域和由该同系物进行的非典型切割位点选择。
测试了对lshcas13apre-crrna加工至关重要的螺旋1结构域残基对lbucas13a的pre-crrna加工活性的影响。测试了四种残基(e299、k310、r311和n314)在pre-crrna切割中的作用(图40b和图40c)(liu等人,cell2017年1月12日;168(1-2):121-134.e12;shmakov等人,molcell.2015年11月5日;60(3):385-97)。这些残基突变为丙氨酸揭示了对pre-crrna加工效率的以下一系列影响:n314a显著降低切割速率,r311a活性受损最小,并且e299a和k310对pre-crrna加工没有影响(图40d)。并行地执行lbucas13a的hepn2结构域内的诱变。r1072和k1082处的丙氨酸取代显著减少了pre-crrna切割,而同一区域中的其它突变(d1078a、k1080a和k1087a)具有对pre-crrna加工的最小影响(图40e)。这些结果表明,hepn2和(较小程度地)螺旋1结构域在lbucas13a的crrna生物发生中起重要作用,但是直接负责催化水解的残基仍然未知。lbucas13a和lshcas13a在参与pre-crrna加工的区域之间的差异(liu等人,cell2017年1月12日;168(1-2):121-134.e12)并不一定相互矛盾,因为crrna的5'末端保持在lshcas13a结构中的hepn2结构域与螺旋1结构域之间(liu等人,cell2017年1月12日;168(1-2):121-134.e12)。来自这两个结构域的残基可能通过使底物结合稳定化、促进适当的底物取向和/或催化水解而在pre-crrna加工中起关键作用。
lshcas13a的hepn2结构域内缺乏保守、其推定的pre-crrna加工活性位点以及该酶的非典型pre-crrna切割位点使我们假设,lshcas13a可利用螺旋1结构域内的不同区域来催化pre-crrna加工。在缺乏与pre-crrna结合的cas13a同系物的三维结构的情况下,通过将lshcas13a切割位点映射到非同源pre-crrna上来测试该假设(图40f)。lshcas13a能够加工来自lwacas13a和lbucas13a的pre-crrna,从而产生相对于所预测的发夹碱基移位一个核苷酸的切割位点。与该观察结果一致,lwacas13a和lbucas13a对lshpre-crrna的加工发生在距重复序列茎标准四核苷酸间隔处,不同于同源lshcas13a位点。这支持以下观察结果:不同的lshcas13a加工位点取决于蛋白架构而不是pre-crrna序列,并且lshcas13a是cas13a树内关于pre-crrna加工的异常蛋白。
cas13a酶在尿嘧啶或腺苷处引发ssrna切割
先前的研究证实,cas13a:crrna复合物识别并结合互补的ssrna靶,由此被称为ssrna激活物,以触发在暴露的尿苷残基处的一般rna酶活性(图42a)。测试同源组是否保留了由lbucas13a和lshcas13a所表现出的非特异性降解活性。还测试hepn活性位点内的尿苷偏好性是否在该家族中是普遍的。为了系统地测试一般rna酶活性,监测包含cas13a:crrna以及结合的ssrna激活物的三元复合物降解反式ssrna靶的能力(图42a)。在一小时的过程中检测到十种同系物中的八种具有反式ssrna切割活性(图42b;图43a)。最值得注意的是,具有反式ssrna切割活性的cas13a同系物是hhecas13a,其具有不可检测的pre-crrna加工活性,但是催化由其同源全长pre-crrna指导的底物的完全降解。虽然先前表征的同系物lshcas13a(abudayyeh等人,science.2016年8月5日;353(6299):aaf5573)和lbucas13a都表现出对易断裂键的5'端尿苷的偏好性,但是由其它同系物产生的不同长度的产物表明,该蛋白家族中可能存在不同的活性位点核苷酸偏好性(图43b)。
为了进一步探测cas13a同系物的反式ssrna切割核苷酸偏好性,使用a、c、g和u的5聚体均聚物作为底物来测量这些酶的反式切割能力。六种同系物能够切割这些短底物(图42c;图43c;图50(表5))。虽然同系物中的四种lbucas13a、lwacas13a、hhecas13a和pprcas13a表现出对同型尿苷底物的偏好性切割,但是观察到了具有最高活性的同系物的次要偏好性(图42c;图43d)。相比之下,lbacas13a和erecas13a偏好性同型腺苷,这与较长靶上的生物化学映射的切割位点一致(图43b)。camcas13a从这些长底物产生相同产物与cas13a家族树的该进化枝的腺苷偏好性一致(图42、图39;图43)。
这些酶之间的一个显著区别是在等摩尔的ssrna激活物和cas13a:crrna干扰复合物的测试条件下,反式ssrna切割达到饱和的速率。这些酶差异可能对cas13a的生物学作用具有巨大影响。定量cas13a同源家族内在反式ssrna切割方面的方差。开发考虑到ssrna激活物结合和反式ssrna切割两者(有助于总酶促输出的cas13a酶的两种核心属性),利用短荧光ssrna报告物进行的rna切割高通量筛选。为了查询每种cas13a同系物的灵敏度,加入不断减少量的互补ssrna激活物以引发反应,并且根据所得时间过程中的每一者计算荧光ssrna报告物切割的表观速率。虽然所计算的速率是所述酶中的每种酶的ssrna激活物结合亲和力和催化转换速率的卷积,但是它们给出了同系物中相当的切割活性的相对量度。
五种同系物(lbucas13a、lwacas13a、lbacas13a、hhecas13a和pprcas13a)在该测定中显示出足够用于可再现分析的可检测切割活性。在这五种同系物中,lbucas13a表现出最高的灵敏度,在存在仅10fm互补激活物的情况下具有可检测的报告物切割(图42d)。只有两种同系物lwacas13a和pprcas13a显示出足够用于在皮摩尔范围内检测激活物的活性,其中灵敏度分别为10pm和100pm。lbacas13a和hhecas13a的灵敏度要低得多,仅在纳摩尔水平的报告物时变得有活性,该纳摩尔水平相对于cas13a复合物接近等摩尔。由于该测定依赖于大量的反式切割事件以产生可检测的荧光,因此可以假设虽然具有相似的切割位点偏好性但不能产生高于背景的可检测信号的三种同系物(erecas13a、camcas13a和lshcas13a)具有灵敏性甚至更低的互补靶灵敏度。灵敏度范围非常广(约107倍)表明cas13a酶具有保护宿主生物免受外来rna影响的多样性能力。
crispr重复序列确定非同源pre-crrna加工
cas13a的双重活性提供了研究pre-crrna加工与不同vi-a型crispr操纵子之间靶向的相互依赖性的机会。为了确定这两种活性的底物需求,测试了不同同系物可以识别非同源crrna以进行指导加工和靶向的程度。最初,尝试经由对cas13a家族和crrna相似性的系统发生分析而生物信息学地预测可能的crrna可交换性。该分析表明存在两种进化枝的同系物,为了清楚起见称为α和β(stamatakis,bioinformatics.2014年5月1日;30(9):1312-3)(图44a)。所述纯化的同系物中的五种存在于这些进化枝之外,具有模糊的祖先关系;有疑问的是pre-crrna(crispr重复序列)序列是否可能决定功能正交性。由于crispr重复序列的短且结构化的内容物,所以使用逐对序列比对评分矩阵在crispr重复序列之间建立分层聚类关系以对整个家族的变异进行评分(burstein等人,natcommun.2016年2月3日;7:10613)。令人惊讶的是,该分析指出存在两种crrna聚类,该两种crrna聚类重叠但不同于由氨基酸序列确定的蛋白进化枝(图44b和图44c)。虽然聚类1crrna与α-进化枝蛋白的子集良好相关,并且所有β-进化枝相关的crrna在聚类2内,但是具有模糊系统发生关系的同系物分散在两个聚类中(图44c)。
不能单独使用生物信息学分析容易地预测cas13a:crrna正交性,所以测试了非同源crrna之间在由所述cas13a同系物中的每一种进行加工和反式ssrna靶切割两者方面功能可交换的程度(图44d和图44f)。对于pre-crrna加工,发现由逐对序列比较定义的crrna聚类预测了它们被它们的相关cas13a蛋白加工的能力(图44d)。例如,来自聚类1的pre-crrna仅由进化枝α的蛋白加工,并且对于聚类2和进化枝β反之亦然。相比之下,蛋白分类的预测性较差,因为大多数模糊分类的蛋白可加工来自重复聚类2的序列,而与它们的重复序列聚类的位置无关。
三种同系物(pprcas13a、hhecas13a和rca13a)是关于它们在crrna聚类内的位置方面异常的pre-crrna加工同系物。hhecas13a不能切割任何非同源的pre-crrna,并且反之,没有同系物(包括hhecas13a在内)加工hhepre-crrna。这表明,hhecas13a不能加工其同源pre-crrna不仅反映了不同的重复序列(图38;图39),还反映了该蛋白中pre-crrna加工活性的丧失。相比之下,pprcas13a和rcacas13a的偏离活性通过crrna重复序列差异来解释。pprcas13a和rcacas13a加工它们自己的crrna,但是两者也加工crrna重复聚类2非同源序列。pprcas13acrrna重复序列与其它聚类2crrna重复序列在5'侧翼区域上的切割位点方面不同,表明由pprcas13a进行的pre-crrna加工具有更大的底物灵活性。类似地,rcacas13acrrna的不同序列特征是相对于存在于该家族其余部分的crrna中的标准五碱基对茎环延伸的六碱基对茎环。值得注意的是,每个crrna重复序列内对cas13apre-crrna加工的位置取代耐受性与上文讨论的对lbucas13a:crrna复合物的突变研究,以及所获得的对lshcas13a:crrna复合物的最近结构见解一致(liu等人,cell2017年1月12日;168(1-2):121-134.e12)。总的来说,这些结果表明,vi-a型crispr重复序列的序列决定了其被cas13a家族进行pre-crrna加工的能力。然而,在存在不同重复序列的情况下进化的同系物(pprcas13a和rcacas13a)保留了加工其它聚类2序列的能力。
功能正交cas13a酶的两个亚家族
对在图44d中定义的pre-crrna加工可交换性聚类是否胜任于指导由非同源cas13a同系物进行的反式ssrna切割进行了研究。为了研究由非同源pre-crrna和成熟crrna指导的cas13a介导反式ssrna切割,对上述实施例中所描述的荧光测定进行修改,并且分析仅限于在ssrna切割实验中表现出显著切割活性的五种同系物(参见图42c)。概括地说,这些结果反映了pre-crrna加工结果,其中crrna重复聚类身份确定了功能组,但是其中与加工和靶向一致的一些鲜明对比是与酶促活性无关的(图44e和图44f)。例如,pprpre-crrna和成熟crrna可以指导由非同源蛋白lwacas13a和lbucas13a进行的ssrna切割,尽管它们不能加工这些pre-crrna。另一惊喜是hhecas13a(其由所有的聚类2的pre-crrna和成熟crrna指导进行反式ssrna切割)的混杂性,尽管其缺乏与对这些指导物中的任一者的pre-crrna加工活性。这表明,crrna成熟不是反式ssrna切割所需要的,这是与以下发现相符的观察结果:lbucas13apre-crrna加工缺陷型突变体具有不受影响的反式ssrna切割能力。
关于pre-crrna加工和反式ssrna切割的crrna可交换性比较限定了cas13a蛋白家族中的两种功能正交的亚家族。第一组(例如,lbucas13a)具有跨该蛋白家族的关于由crrna指导的pre-crrna加工和反式ssrna切割两方面的广泛混杂性。此多源组也显示出对反式ssrna切割活性位点内的尿苷的偏好性。由crrna和蛋白序列两者限定的第二组(例如,lbacas13a),具有不同的crrna可交换特征并且在反式ssrna靶切割期间优先在腺苷处进行切割。为了测试这些cas13a亚家族的正交反应性,验证lbucas13a和lbacas13a分别切割同型assrna报告物或同型ussrna报告物(图45a和图45b)。虽然不同的探针产生相似量的荧光信号,但应当注意的是,由于同系物的不同灵敏度(对于lbucas13a和lbacas13a分别为10pm和1nm),所以加入了实质上不同量的ssrna激活物。为了验证正交性,测试了一组具有在crrna、激活物和报告物之间的所有可能非同源组合的对照反应,其中除了同源组合外没有检测到实质信号(图45c)。总的来说,这些结果限定了可以在相同系统内并行起作用的不同cas13a同系物。
pre-crrna加工增强crispr阵列情境中的靶向效率
这项研究的一个令人费解的发现是,触发随后由cas13a进行的反式ssrna靶切割反应缺乏对成熟crrna的严格需要。此外,lbucas13a的加工缺陷型突变体保持相似的反式ssrna切割效率,甚至当由pre-crrna代替成熟crrna进行指导时也是如此(图46;图47)。这导致了以下假设:pre-crrna加工在vi型crispr基因座中的作用不一定是用于有效的ssrna靶向,而是替代地用于从长crispr阵列转录物的范围中释放每个crrna。有疑问的是pre-crrna加工是否可以缓解在crrna负载和/或ssrna靶向期间的rna折叠约束以及相邻cas13a:crrna-间隔区物质的潜在空间位阻。为了测试此点,比较使用野生型lbucas13a或无pre-crrna加工活性的突变体进行的crispr阵列指导的ssrna切割的效率。
由于lbucas13a的所有加工缺陷型单点突变体(图40)保留低水平的pre-crrna加工活性,所以建立了不具有可检测的加工活性,但保留了与野生型lbucas13a相似或更高的反式ssrna切割效率的双重突变体(r1079a/k1080a)(图46a和图46b)。在存在靶rna和由六个不同的重复间隔区单元组成的小crispr阵列转录物的情况下测试该双重突变体和野生型lbucas13a酶的ssrna切割(图46c和图46d)。对于该阵列内的所有间隔区序列,由无crrna加工活性的突变体进行的反式ssrna切割的速率显著下降(单侧t检验;对于所有对p<0.001)。与野生型lbucas13a相比活性降低对于该阵列内的各个连续间隔区更明显,其中最后一个间隔区以仅为由野生型酶催化的速率的15%的速率指导切割。该发现表明,虽然pre-crrna加工对于靶向不是必需的,但是其通过从crispr阵列释放crrna来增强活性,导致改进的vi型crispr系统模型(图55)。
图38a至图38c.pre-crrna加工在cas13a蛋白家族中是广泛保守的。(a)cas13a催化的crrna生物发生途径的示意图。由cas13a切割pre-crrna转录物以产生成熟crrna。下面是pre-crrna的示意图,突出显示了重要的功能特征。(b)来自所研究的vi型crispr系统的crispr重复序列的5'部分的比对,突出显示了pre-crrna切割位点。所映射的切割的切割位点示出为红色条。以黑色文本标示了与lbucrrna重复序列的偏差。小写g是转录目的所需要的,并且不是天然crrna重复序列的部分。完整的crispr重复序列示于图39b中。(c)与5'-放射性标记的pre-crrna底物一起孵育60分钟后,由9种cas13a同系物进行的cas13a介导pre-crrna切割的代表性凝胶。
图39a至图39c.关于本研究中使用的cas13a同系物的crispr基因座和crrna重复结构(a)cas13a蛋白的最大似然系统发生树,其中vi-a型基因座的图改编自(shmakov等人,molcell.2015年11月5日;60(3):385-97)。cas13aorf以青色示出。crispr阵列描绘为黑色框(重复序列)、黄色菱形(间隔区)和针对上面提到的较大阵列的间隔区阵列尺寸。基因座周围的所关注的orf以以下缩写标示:t-毒素、at-抗毒素,以及tp-转座酶。(b)对来自所用同系物的crispr重复序列的人工比对。pre-crrna加工切割位点以红线标示。以黑色文本标示了与lbucrrna重复序列的偏差。小写g是转录目的所需要的,并且不是天然crrna重复序列的部分。测试了两种单独的hhecrrna序列,第一种含有天然序列,而第二种具有四核苷酸延伸部以延长非典型短的天然重复序列。在任何研究条件下,hhecas13a都不切割crrna重复序列。(c)在可变的盐和ph条件下,使用hhecas13a对天然crrna重复序列进行的pre-crrna加工测定。没有观察到切割产物。
图40a至图40f.对于由lbucas13a进行的pre-crrna切割重要的残基的鉴别(a)lbucas13a结构域的组构示意图,其中结构域基于lshcas13a晶体结构注释(liu等人,cell2017年1月12日;168(1-2):121-134.e12)。(b)通过对lshcas13a的研究,对cas13a的螺旋1结构域中参与pre-crrna加工的局部区域,以及(c)通过对lbucas13a的研究,对cas13a的hepn2结构域内参与pre-crrna加工的区域进行多重序列氨基酸比对。关于这些区域的完整家族树比对参见图41,并且关于完整的蛋白比对参见图57。突变严重影响pre-crrna加工的残基以黄色菱形标记,并且突变最低限度地影响pre-crrna加工的残基以青色菱形标记。lbucas13a序列上方的符号对应于对lbucas13a进行的突变,并且lshcas13a序列下方的符号对应于由liu等人,cell2017年1月12日;168(1-2):121-134.e12对lshcas13a进行的突变。矩阵比对的着色表示使用clustalx方案的残基保守,其中较暗的色调指示保守的强度。测量突变体的(d)螺旋1结构域和(e)hepn2结构域中在单一转换条件下的pre-crrna加工。将定量数据与单指数衰减拟合,其中所计算的伪一级速率常数(kob)(平均值±标准差,n=3)如下:lbuwt为0.074±0.003分钟-1,e299a为0.071±0.005分钟-1,k310a为0.071±0.003分钟-1,r311a为0.054±0.007分钟-1,n314a为0.029±0.008分钟-1,r1079a为0.009±0.007分钟-1,d1078a为0.023±0.002分钟-1,k1080a为0.016±0.004分钟-1,并且k1087a为0.076±0.007分钟-1,而r1072a和k1082a无法进行拟合。(f)使用标准条件,通过lbucas13a、lshcas13a和lwacas13a蛋白对lshcas13a、lwacas13a和lbucas13apre-crrna进行pre-crrna加工的代表性凝胶。最右边泳道中的水解梯带允许相对尺寸比较,尽管三种pre-crrna之间的细微序列差异将改变这些小片段的迁移。
图41a至图41b.螺旋1结构域和hepn结构域的完全比对(a-b)跨(a)螺旋1结构域和(b)hepn2结构域对19种cas13a家族成员的多重序列比对。gi登录号列在每个物种名称之前。下面列出了基于比对上方列出的lshcas13a序列的坐标和lbucas13a序列的坐标。在该研究中测试的突变在比对上方以黄色和青色菱形标示,所述黄色和青色菱形分别对应于不利地影响pre-crrna加工反应的丙氨酸取代或对pre-crrna加工具有最小影响的丙氨酸取代。由(liu等人,cell2017年1月12日;168(1-2):121-134.e12)测试的突变描绘于lshcas13a序列下方。通过jalview,使用amas方法计算保守评分。
图42a至图42d.cas13a蛋白家族的成员以一系列效率切割ssrna(a)由cas13a进行的ssrna靶向的示意图。为简单起见,反式ssrna切割是研究的重点。(b)在孵育60分钟后,由所有十种同系物进行的cas13a介导反式ssrna切割的代表性凝胶。如方法中所述,使用成熟crrna产物形成cas13a:crrna复合物,其中最终rnp复合物浓度为50nm。在存在和不存在50nm未标记的与crrna互补的ssrna激活物的情况下,加入小于1nm的放射性标记的反式ssrna靶以引发反应。使用lshcas13a观察到了弱的反式ssrna切割活性,其中产物条带在右侧用双箭头指示。(c)热图,报告了对于六种不同的cas13a,各5聚体均聚物ssrna底物的cas13a催化的反式ssrna切割百分比。测定条件与部分(b)相同,不同的是lbucas13a和lwacas13a,它们孵育5分钟而不是60分钟。(n=3,具有相关误差的值在图51(表6)中示出。(d)在一系列ssrna激活物浓度下,五种同系物对荧光ssrna报告物的表观切割速率。将cas13a:crrna复合物分别以2:1的比率预孵育,其中最终活性复合物浓度为50nm。加入互补ssrna激活物和荧光ssrna切割报告物以引发反应。将归一化的报告物信号曲线的时间过程使用单指数衰减进行拟合,并且将表观速率绘图(n=3)。一些条件在第一测量时间点之前达到平台,因此它们的速率被最低限度地假设为0.5分钟-1,并在图表中用*标记。
图43a至图43d.由cas13a同系物进行的反式ssrna切割(a)对在存在和不存在ssrna激活物的情况下由十种不同的cas13a同系物进行的反式ssrna切割的时间过程分析。时间点取在1分钟、10分钟和60分钟处。标示了lbucas13a、lwacas13a、lbacas13a、erecas13a、hhecas13a、pprcas13a、camcas13a和lshcas13a的ssrna激活物特异性切割产物。虚线表示单独的page凝胶之间的边界。(b)针对四种cas13a同系物的多个切割反应映射的反式ssrna切割位点。不同的切割模式以红色箭头标示。似乎lbacas13a和erecas13a可具有腺苷偏好性,而lwacas13a似乎在核苷酸偏好性方面更加混杂。(c-d)由5种cas13a同系物进行的对均聚ssrna底物的代表性反式ssrna切割凝胶。
图44a至图44f.cas13a家族中的crrna可交换性(a)cas13a蛋白的最大似然系统发生树。本研究中使用的同系物以粗体表示,并且进化枝被突出显示。自展值位于图39中。(b)关于来自本研究中使用的同系物的crispr重复序列的对称相似性得分矩阵。行和列是按照crispr重复序列聚类排序的。(c)关于来自本研究中使用的同系物的crispr重复序列的不对称相似性得分矩阵。在(b)中呈现了相同的逐对评分,不同之处为行被重新排序以对应于cas13a系统发生树。(d-f)(d)由非同源蛋白进行的pre-crrna加工、(e)由pre-crrna指导的反式ssrna切割,以及(f)由成熟crrna指导的反式ssrna切割的功能活性矩阵。使用标准条件执行加工测定,并分析60分钟的反应终点。使用荧光ssrna报告物测定,以拟合的初始速率执行反式ssrna切割测定。ssrna激活物的浓度如下:lbucas13a:100pm、lwacas13a:100pm、pprcas13a:100nm、lbacas13a:10nm以及hhecas13a:100nm。对三次平行测定的初始速率进行拟合以计算荧光平台值的差异,并针对每种cas13a:crrna同源对进行归一化。关于数值和相关误差(n=3),参见图52、图53和图54(分别为表7、表8和表9)。
图45a至图45c.对用于rna检测的正交cas13a亚家族的功能验证(a)rna检测测定的示意图,所述rna检测测定经改进以使用荧光均聚ssrna报告底物来测定由lbucas13a或lbacas13a进行的反式ssrna切割激活。(b)由与lbucas13a:lbu-crrna:10pmssrna激活物或lbacas13a:lba-crrna:1nmssrna激活物一起孵育的均聚报告物产生的原始荧光测量值的时间过程(平均值±标准差,n=3)。(c)对于一组crrna、ssrna激活物和cas13a蛋白组合,由荧光均聚ssrna报告物产生的原始荧光测量值(平均值±标准差,n=3)。
图46a至图46d.对lbucas13a的crdna阵列加工作用的解释(a)相较于野生型lbucas13a的r1079a/k1080a突变体的pre-crrna加工测定的定量时间过程数据。将定量数据与单指数衰减拟合,并且lbucas13awt的伪一级速率常数(kob)(平均值±标准差,n=3)为0.074±0.003分钟-1,而r1079a/k1080a突变体不能以足够的置信度拟合产生速率常数。(b)由pre-crrna和成熟crrna指导的lbucas13a野生型和无r1079a/k1080a加工活性的突变体的荧光报告物表观比率。将cas13a:rna复合物以1:1的比率预孵育60分钟,然后加入10pm的激活物和150nm的报告物以引发反应。(平均值±标准差,n=3)(c)如由50nm的含有六个crrna重复间隔区的crispr阵列指导的,由300nm野生型lbucas13a或无r1079a/k1080apre-crrna加工活性的突变体进行的荧光ssrna报告物切割的表观速率。每个条组代表加入100pm的不同ssrna激活物序列,所述不同ssrna激活物序列与在每个条组下方指示的crispr阵列内的示意位置(schematizedposition)互补。将每个速率根据来自三次生物平行测定的数据进行拟合,并描绘速率的标准偏差。对于所有间隔区位置,突变蛋白的速率与野生型lbucas13a在统计学上不同(单侧t检验:p<0.001)。(d)来自(c)的数据被描述为野生型lbucas13a活性的百分比,证明了由无pre-crrna加工活性的突变体降低反式ssrna靶向效率的位置效应。
图47a至图47c.由lbucas13a点突变体在参与pre-crrna加工的区域中进行的反式切割。(a-c)由lbucas13a的各种‘参与pre-crrna加工的点突变体进行的反式ssrna切割。如上所述,在25℃或37℃用0.2nm或1nm的ssrna激活物使用50nm的cas13a:crrna执行切割反应。使用较低的温度和激活物浓度以减慢反应动力学,以进行更准确的测量。拟合的曲线是单指数衰减,其中所计算的伪一级速率常数(平均值±标准差,n=3)如下:25℃条件:lbuwt为1.76±0.24分钟-1,k299a为1.72±0.65分钟-1,k310a为3.19±0.27分钟-1,r1072a为2.95±0.53分钟-1,r1079a为0.93±0.28分钟-1,并且k1082a而2.64±0.63分钟-1,并且37℃条件:r311a为0.39±0.09分钟-1,并且n314a为0.54±0.11分钟-1,而野生型和无加工活性的突变体(r1079a/k1080a)过快地达到平台而无法进行准确的速率测量:wt为2.95±1.97分钟-1,并且r1079a/k1080a为4.86±4.99分钟-1。
图48a至图48b.无crrna加工活性的突变体r1079a/k1080a保留相似的crrna结合亲和力并且不加工pre-crna阵列(a)如方法中所述地进行过滤结合测定以确定成熟crrna与lbucas13awt和lbucas13ar1079a/k1080a的结合亲和力。将定量的数据与标准结合等温线拟合。从三个独立实验测量的解离常数(平均值±标准偏差)为1.21±0.57nm(lbucas13awt)和3.11±0.89nm(lbucas13ar1079a/k1080a)。(b)使用六聚体crispr阵列作为底物、以及lbucas13a和lbucas13ar1079a/k1080a突变体以及各种尺寸的标记物进行的pre-crna加工测定。产物标识描绘在凝胶的右侧。由于偶尔未加工的另外前导区,如所标示的偶尔会存在其它尺寸的产物。
图49(表4).crispr重复共有序列:针对在含有cas13a的操纵子(rcacas13a、lbacas13a和erecas13a)的10kb内具有多个crispr基因座的同系物,或具有重复变异(pprcas13a)的长阵列,列出了多种crrna重复序列,其中突变以红色文本突出显示。对于pprcas13a,选择阵列前导侧的第一crrna重复序列用于该研究。对于rcacas13a、lbacas13a和erecas13a,分析crrna重复序列的以下两个因素以选择用于该研究的代表性crrna(1):阵列的长度,以及(2)由同源cas13a蛋白指导反式ssrna切割的能力。以主要文本使用的序列在最后一列中注明。
图50(表5).*寡聚物编号-用于保持本研究中使用的rna底物的一致性的指数。**来源缩写:ss-单链dna寡核苷酸模板用于体外转录,hhpcr-体外转录模板是包含锤头型核酶模板序列的重叠寡核苷酸的pcr产物,idt-由idt合成的,pcr-从质粒扩增的体外转录模板。
图51表示表6。图52表示表7。图53表示表8。图54表示表9。
图55.vi-a型crispr系统功能的改进模型(a)本研究中的关键发现的图形概要。本研究中使用的同系物用粗体缩写表示,其中无反式ssrna切割活性的同系物用灰色绘示。彩色圆圈突出显示两个正交的cas13a酶组,所述两个正交的cas13a酶组如由它们的广义crrna可交换性和进行反式ssrna切割的底物核苷酸偏好性所限定。(b)描绘了vi-a型crispr系统功能的改进模型的示意图。
虽然已经参考本发明的特定实施方案描述了本发明,但是本领域的技术人员应当理解,在不脱离本发明的真实精神和范围的情况下,可以进行各种改变并且可以替换等同物。另外,可以进行许多修改以使具体情况、材料、物质组成、过程、一个或多个过程步骤适应本发明的目的、精神和范围。所有此类修改都在所附权利要求书的范围内。
序列表
<110>doudna,jennifera
o'connell,mitchell
seletsky,alexandrae
knight,spencerc
cate,jamie
<120>用于检测靶rna的方法和组合物
<130>berk-337
<150>us62/351,172
<151>2016-06-16
<150>us62/378,156
<151>2016-08-22
<160>26
<170>patentin3.5版
<210>1
<211>1120
<212>prt
<213>斯氏李斯特菌
<400>1
mettrpileserilelysthrleuilehishisleuglyvalleuphe
151015
phecysasptyrmettyrasnargargglulyslysileilegluval
202530
lysthrmetargilethrlysvalgluvalasparglyslysvalleu
354045
ileserargasplysasnglyglylysleuvaltyrgluasnglumet
505560
glnaspasnthrgluglnilemethishislyslysserserphetyr
65707580
lysservalvalasnlysthrilecysargprogluglnlysglnmet
859095
lyslysleuvalhisglyleuleuglngluasnserglnglulysile
100105110
lysvalseraspvalthrlysleuasnileserasnpheleuasnhis
115120125
argphelyslysserleutyrtyrpheprogluasnserproasplys
130135140
sergluglutyrargilegluileasnleuserglnleuleugluasp
145150155160
serleulyslysglnglnglythrpheilecystrpgluserpheser
165170175
lysaspmetgluleutyrileasntrpalagluasntyrileserser
180185190
lysthrlysleuilelyslysserileargasnasnargileglnser
195200205
thrgluserargserglyglnleumetaspargtyrmetlysaspile
210215220
leuasnlysasnlyspropheaspileglnservalserglulystyr
225230235240
glnleuglulysleuthrseralaleulysalathrphelysgluala
245250255
lyslysasnasplysgluileasntyrlysleulysserthrleugln
260265270
asnhisgluargglnileileglugluleulysgluasnsergluleu
275280285
asnglnpheasnilegluilearglyshisleugluthrtyrphepro
290295300
ilelyslysthrasnarglysvalglyaspileargasnleugluile
305310315320
glygluileglnlysilevalasnhisargleulysasnlysileval
325330335
glnargileleuglngluglylysleualasertyrgluilegluser
340345350
thrvalasnserasnserleuglnlysilelysilegluglualaphe
355360365
alaleulyspheileasnalacysleuphealaserasnasnleuarg
370375380
asnmetvaltyrprovalcyslyslysaspileleumetileglyglu
385390395400
phelysasnserphelysgluilelyshislyslyspheilearggln
405410415
trpserglnphepheserglngluilethrvalaspaspilegluleu
420425430
alasertrpglyleuargglyalailealaproileargasngluile
435440445
ilehisleulyslyshissertrplyslysphepheasnasnprothr
450455460
phelysvallyslysserlysileileasnglylysthrlysaspval
465470475480
thrserglupheleutyrlysgluthrleuphelysasptyrphetyr
485490495
sergluleuaspservalprogluleuileileasnlysmetgluser
500505510
serlysileleuasptyrtyrserseraspglnleuasnglnvalphe
515520525
thrileproasnphegluleuserleuleuthrseralavalprophe
530535540
alaproserphelysargvaltyrleulysglypheasptyrglnasn
545550555560
glnaspglualaglnproasptyrasnleulysleuasniletyrasn
565570575
glulysalapheasnserglualapheglnalaglntyrserleuphe
580585590
lysmetvaltyrtyrglnvalpheleuproglnphethrthrasnasn
595600605
aspleuphelysserservalasppheileleuthrleuasnlysglu
610615620
arglysglytyralalysalapheglnaspilearglysmetasnlys
625630635640
aspglulysproserglutyrmetsertyrileglnserglnleumet
645650655
leutyrglnlyslysglngluglulysglulysileasnhispheglu
660665670
lyspheileasnglnvalpheilelysglypheasnserpheileglu
675680685
lysasnargleuthrtyrilecyshisprothrlysasnthrvalpro
690695700
gluasnaspasnilegluileprophehisthraspmetaspaspser
705710715720
asnilealaphetrpleumetcyslysleuleuaspalalysglnleu
725730735
sergluleuargasnglumetilelysphesercysserleuglnser
740745750
thrglugluileserthrphethrlysalaarggluvalileglyleu
755760765
alaleuleuasnglyglulysglycysasnasptrplysgluleuphe
770775780
aspasplysglualatrplyslysasnmetserleutyrvalserglu
785790795800
gluleuleuglnserleuprotyrthrglngluaspglyglnthrpro
805810815
valileasnargserileaspleuvallyslystyrglythrgluthr
820825830
ileleuglulysleupheserserseraspasptyrlysvalserala
835840845
lysaspilealalysleuhisglutyraspvalthrglulysileala
850855860
glnglngluserleuhislysglntrpileglulysproglyleuala
865870875880
argaspseralatrpthrlyslystyrglnasnvalileasnaspile
885890895
serasntyrglntrpalalysthrlysvalgluleuthrglnvalarg
900905910
hisleuhisglnleuthrileaspleuleuserargleualaglytyr
915920925
metserilealaaspargasppheglnpheserserasntyrileleu
930935940
gluarggluasnserglutyrargvalthrsertrpileleuleuser
945950955960
gluasnlysasnlysasnlystyrasnasptyrgluleutyrasnleu
965970975
lysasnalaserilelysvalserserlysasnaspproglnleulys
980985990
valaspleulysglnleuargleuthrleuglutyrleugluleuphe
99510001005
aspasnargleulysglulysargasnasnileserhispheasn
101010151020
tyrleuasnglyglnleuglyasnserileleugluleupheasp
102510301035
aspalaargaspvalleusertyrasparglysleulysasnala
104010451050
valserlysserleulysgluileleuserserhisglymetglu
105510601065
valthrphelysproleutyrglnthrasnhishisleulysile
107010751080
asplysleuglnprolyslysilehishisleuglyglulysser
108510901095
thrvalserserasnglnvalserasnglutyrcysglnleuval
110011051110
argthrleuleuthrmetlys
11151120
<210>2
<211>1159
<212>prt
<213>口腔纤毛菌
<400>2
metlysvalthrlysvalglyglyileserhislyslystyrthrser
151015
gluglyargleuvallyssergluserglugluasnargthraspglu
202530
argleuseralaleuleuasnmetargleuaspmettyrilelysasn
354045
proserserthrgluthrlysgluasnglnlysargileglylysleu
505560
lyslysphepheserasnlysmetvaltyrleulysaspasnthrleu
65707580
serleulysasnglylyslysgluasnileaspargglutyrserglu
859095
thraspileleugluseraspvalargasplyslysasnphealaval
100105110
leulyslysiletyrleuasngluasnvalasnserglugluleuglu
115120125
valpheargasnaspilelyslyslysleuasnlysileasnserleu
130135140
lystyrserpheglulysasnlysalaasntyrglnlysileasnglu
145150155160
asnasnileglulysvalgluglylysserlysargasnileiletyr
165170175
asptyrtyrarggluseralalysargaspalatyrvalserasnval
180185190
lysglualapheasplysleutyrlysglugluaspilealalysleu
195200205
valleugluilegluasnleuthrlysleuglulystyrlysilearg
210215220
gluphetyrhisgluileileglyarglysasnasplysgluasnphe
225230235240
alalysileiletyrglugluileglnasnvalasnasnmetlysglu
245250255
leuileglulysvalproaspmetsergluleulyslysserglnval
260265270
phetyrlystyrtyrleuasplysglugluleuasnasplysasnile
275280285
lystyralaphecyshisphevalgluileglumetserglnleuleu
290295300
lysasntyrvaltyrlysargleuserasnileserasnasplysile
305310315320
lysargilepheglutyrglnasnleulyslysleuilegluasnlys
325330335
leuleuasnlysleuaspthrtyrvalargasncysglylystyrasn
340345350
tyrtyrleuglnaspglygluilealathrserasppheilealaarg
355360365
asnargglnasnglualapheleuargasnileileglyvalserser
370375380
valalatyrpheserleuargasnileleugluthrgluasngluasn
385390395400
aspilethrglyargmetargglylysthrvallysasnasnlysgly
405410415
gluglulystyrvalserglygluvalasplysiletyrasngluasn
420425430
lyslysasngluvallysgluasnleulysmetphetyrsertyrasp
435440445
pheasnmetaspasnlysasngluilegluaspphephealaasnile
450455460
aspglualaileserserilearghisglyilevalhispheasnleu
465470475480
gluleugluglylysaspilephealaphelysasnilealaproser
485490495
gluileserlyslysmetpheglnasngluileasnglulyslysleu
500505510
lysleulysilepheargglnleuasnseralaasnvalpheargtyr
515520525
leuglulystyrlysileleuasntyrleulysargthrargpheglu
530535540
phevalasnlysasnileprophevalproserphethrlysleutyr
545550555560
serargileaspaspleulysasnserleuglyiletyrtrplysthr
565570575
prolysthrasnaspaspasnlysthrlysgluileileaspalagln
580585590
iletyrleuleulysasniletyrtyrglyglupheleuasntyrphe
595600605
metserasnasnglyasnphephegluileserlysgluileileglu
610615620
leuasnlysasnasplysargasnleulysthrglyphetyrlysleu
625630635640
glnlysphegluaspileglnglulysileprolysglutyrleuala
645650655
asnileglnserleutyrmetileasnalaglyasnglnaspgluglu
660665670
glulysaspthrtyrileasppheileglnlysilepheleulysgly
675680685
phemetthrtyrleualaasnasnglyargleuserleuiletyrile
690695700
glyseraspglugluthrasnthrserleualaglulyslysglnglu
705710715720
pheasplyspheleulyslystyrgluglnasnasnasnilelysile
725730735
protyrgluileasnglupheleuarggluilelysleuglyasnile
740745750
leulystyrthrgluargleuasnmetphetyrleuileleulysleu
755760765
leuasnhislysgluleuthrasnleulysglyserleuglulystyr
770775780
glnseralaasnlysgluglualapheseraspglnleugluleuile
785790795800
asnleuleuasnleuaspasnasnargvalthrgluaspphegluleu
805810815
glualaaspgluileglylyspheleuasppheasnglyasnlysval
820825830
lysaspasnlysgluleulyslyspheaspthrasnlysiletyrphe
835840845
aspglygluasnileilelyshisargalaphetyrasnilelyslys
850855860
tyrglymetleuasnleuleuglulysilealaasplysalaglytyr
865870875880
lysileserileglugluleulyslystyrserasnlyslysasnglu
885890895
ileglulysasnhislysmetglngluasnleuhisarglystyrala
900905910
argproarglysaspglulysphethraspgluasptyrglusertyr
915920925
lysglnalailegluasnilegluglutyrthrhisleulysasnlys
930935940
valglupheasngluleuasnleuleuglnglyleuleuleuargile
945950955960
leuhisargleuvalglytyrthrseriletrpgluargaspleuarg
965970975
pheargleulysglyglupheprogluasnglntyrileglugluile
980985990
pheasnphegluasnlyslysasnvallystyrlysglyglyglnile
99510001005
valglulystyrilelysphetyrlysgluleuhisglnasnasp
101010151020
gluvallysileasnlystyrserseralaasnilelysvalleu
102510301035
lysglnglulyslysaspleutyrileargasntyrilealahis
104010451050
pheasntyrileprohisalagluileserleuleugluvalleu
105510601065
gluasnleuarglysleuleusertyrasparglysleulysasn
107010751080
alavalmetlysservalvalaspileleulysglutyrglyphe
108510901095
valalathrphelysileglyalaasplyslysileglyilegln
110011051110
thrleugluserglulysilevalhisleulysasnleulyslys
111511201125
lyslysleumetthraspargasnserglugluleucyslysleu
113011351140
vallysilemetpheglutyrlysmetgluglulyslysserglu
114511501155
asn
<210>3
<211>1389
<212>prt
<213>沙氏纤毛菌
<400>3
metglyasnleupheglyhislysargtrptyrgluvalargasplys
151015
lysaspphelysilelysarglysvallysvallysargasntyrasp
202530
glyasnlystyrileleuasnileasngluasnasnasnlysglulys
354045
ileaspasnasnlyspheilearglystyrileasntyrlyslysasn
505560
aspasnileleulysgluphethrarglysphehisalaglyasnile
65707580
leuphelysleulysglylysgluglyileileargilegluasnasn
859095
aspasppheleugluthrglugluvalvalleutyrileglualatyr
100105110
glylysserglulysleulysalaleuglyilethrlyslyslysile
115120125
ileaspglualaileargglnglyilethrlysaspasplyslysile
130135140
gluilelysargglngluasngluglugluilegluileaspilearg
145150155160
aspglutyrthrasnlysthrleuasnaspcysserileileleuarg
165170175
ileilegluasnaspgluleugluthrlyslysseriletyrgluile
180185190
phelysasnileasnmetserleutyrlysileileglulysileile
195200205
gluasngluthrglulysvalphegluasnargtyrtyrglugluhis
210215220
leuargglulysleuleulysaspasplysileaspvalileleuthr
225230235240
asnphemetgluileargglulysilelysserasnleugluileleu
245250255
glyphevallysphetyrleuasnvalglyglyasplyslyslysser
260265270
lysasnlyslysmetleuvalglulysileleuasnileasnvalasp
275280285
leuthrvalgluaspilealaaspphevalilelysgluleugluphe
290295300
trpasnilethrlysargileglulysvallyslysvalasnasnglu
305310315320
pheleuglulysargargasnargthrtyrilelyssertyrvalleu
325330335
leuasplyshisglulysphelysilegluarggluasnlyslysasp
340345350
lysilevallysphephevalgluasnilelysasnasnserilelys
355360365
glulysileglulysileleualagluphelysileaspgluleuile
370375380
lyslysleuglulysgluleulyslysglyasncysaspthrgluile
385390395400
pheglyilephelyslyshistyrlysvalasnpheaspserlyslys
405410415
pheserlyslysseraspgluglulysgluleutyrlysileiletyr
420425430
argtyrleulysglyargileglulysileleuvalasngluglnlys
435440445
valargleulyslysmetglulysilegluileglulysileleuasn
450455460
gluserileleuserglulysileleulysargvallysglntyrthr
465470475480
leugluhisilemettyrleuglylysleuarghisasnaspileasp
485490495
metthrthrvalasnthraspasppheserargleuhisalalysglu
500505510
gluleuaspleugluleuilethrphephealaserthrasnmetglu
515520525
leuasnlysilepheserarggluasnileasnasnaspgluasnile
530535540
aspphepheglyglyaspargglulysasntyrvalleuasplyslys
545550555560
ileleuasnserlysilelysileileargaspleuasppheileasp
565570575
asnlysasnasnilethrasnasnpheilearglysphethrlysile
580585590
glythrasngluargasnargileleuhisalaileserlysgluarg
595600605
aspleuglnglythrglnaspasptyrasnlysvalileasnileile
610615620
glnasnleulysileseraspglugluvalserlysalaleuasnleu
625630635640
aspvalvalphelysasplyslysasnileilethrlysileasnasp
645650655
ilelysileserglugluasnasnasnaspilelystyrleuproser
660665670
pheserlysvalleuprogluileleuasnleutyrargasnasnpro
675680685
lysasnglupropheaspthrilegluthrglulysilevalleuasn
690695700
alaleuiletyrvalasnlysgluleutyrlyslysleuileleuglu
705710715720
aspaspleuglugluasngluserlysasnilepheleuglngluleu
725730735
lyslysthrleuglyasnileaspgluileaspgluasnileileglu
740745750
asntyrtyrlysasnalaglnileseralaserlysglyasnasnlys
755760765
alailelyslystyrglnlyslysvalileglucystyrileglytyr
770775780
leuarglysasntyrglugluleupheasppheseraspphelysmet
785790795800
asnileglngluilelyslysglnilelysaspileasnaspasnlys
805810815
thrtyrgluargilethrvallysthrserasplysthrilevalile
820825830
asnaspasppheglutyrileileserilephealaleuleuasnser
835840845
asnalavalileasnlysileargasnargphephealathrserval
850855860
trpleuasnthrserglutyrglnasnileileaspileleuaspglu
865870875880
ilemetglnleuasnthrleuargasnglucysilethrgluasntrp
885890895
asnleuasnleugluglupheileglnlysmetlysgluileglulys
900905910
asppheaspaspphelysileglnthrlyslysgluilepheasnasn
915920925
tyrtyrgluaspilelysasnasnileleuthrgluphelysaspasp
930935940
ileasnglycysaspvalleuglulyslysleuglulysilevalile
945950955960
pheaspaspgluthrlysphegluileasplyslysserasnileleu
965970975
glnaspgluglnarglysleuserasnileasnlyslysaspleulys
980985990
lyslysvalaspglntyrilelysasplysaspglngluilelysser
99510001005
lysileleucysargileilepheasnserasppheleulyslys
101010151020
tyrlyslysgluileaspasnleuilegluaspmetgluserglu
102510301035
asngluasnlyspheglngluiletyrtyrprolysgluarglys
104010451050
asngluleutyriletyrlyslysasnleupheleuasnilegly
105510601065
asnproasnpheasplysiletyrglyleuileserasnaspile
107010751080
lysmetalaaspalalyspheleupheasnileaspglylysasn
108510901095
ilearglysasnlysilesergluileaspalaileleulysasn
110011051110
leuasnasplysleuasnglytyrserlysglutyrlysglulys
111511201125
tyrilelyslysleulysgluasnaspaspphephealalysasn
113011351140
ileglnasnlysasntyrlysserpheglulysasptyrasnarg
114511501155
valserglutyrlyslysileargaspleuvalglupheasntyr
116011651170
leuasnlysileglusertyrleuileaspileasntrplysleu
117511801185
alaileglnmetalaargphegluargaspmethistyrileval
119011951200
asnglyleuarggluleuglyileilelysleuserglytyrasn
120512101215
thrglyileserargalatyrprolysargasnglyseraspgly
122012251230
phetyrthrthrthralatyrtyrlysphepheaspglugluser
123512401245
tyrlyslyspheglulysilecystyrglypheglyileaspleu
125012551260
sergluasnsergluileasnlysprogluasngluserilearg
126512701275
asntyrileserhisphetyrilevalargasnprophealaasp
128012851290
tyrserilealagluglnileaspargvalserasnleuleuser
129513001305
tyrserthrargtyrasnasnserthrtyralaservalpheglu
131013151320
valphelyslysaspvalasnleuasptyraspgluleulyslys
132513301335
lysphelysleuileglyasnasnaspileleugluargleumet
134013451350
lysprolyslysvalservalleugluleuglusertyrasnser
135513601365
asptyrilelysasnleuileilegluleuleuthrlysileglu
137013751380
asnthrasnaspthrleu
1385
<210>4
<211>1285
<212>prt
<213>荚膜红细菌r121
<400>4
metglnileglylysvalglnglyargthrileserglupheglyasp
151015
proalaglyglyleulysarglysileserthraspglylysasnarg
202530
lysgluleuproalahisleuserseraspprolysalaleuilegly
354045
glntrpileserglyileasplysiletyrarglysproaspserarg
505560
lysseraspglylysalailehisserprothrproserlysmetgln
65707580
pheaspalaargaspaspleuglyglualaphetrplysleuvalser
859095
glualaglyleualaglnaspserasptyraspglnphelysargarg
100105110
leuhisprotyrglyasplyspheglnproalaaspserglyalalys
115120125
leulyspheglualaaspproprogluproglnalaphehisglyarg
130135140
trptyrglyalametserlysargglyasnaspalalysgluleuala
145150155160
alaalaleutyrgluhisleuhisvalaspglulysargileaspgly
165170175
glnprolysargasnprolysthrasplysphealaproglyleuval
180185190
valalaargalaleuglyilegluserservalleuproargglymet
195200205
alaargleualaargasntrpglygluglugluileglnthrtyrphe
210215220
valvalaspvalalaalaservallysgluvalalalysalaalaval
225230235240
seralaalaglnalapheaspproproargglnvalserglyargser
245250255
leuserprolysvalglyphealaleualagluhisleugluargval
260265270
thrglyserlysargcysserpheaspproalaalaglyproserval
275280285
leualaleuhisaspgluvallyslysthrtyrlysargleucysala
290295300
argglylysasnalaalaargalapheproalaasplysthrgluleu
305310315320
leualaleumetarghisthrhisgluasnargvalargasnglnmet
325330335
valargmetglyargvalserglutyrargglyglnglnalaglyasp
340345350
leualaglnserhistyrtrpthrseralaglyglnthrgluilelys
355360365
glusergluilephevalargleutrpvalglyalaphealaleuala
370375380
glyargsermetlysalatrpileaspprometglylysilevalasn
385390395400
thrglulysasnaspargaspleuthralaalavalasnilearggln
405410415
valileserasnlysglumetvalalaglualametalaargarggly
420425430
iletyrpheglygluthrprogluleuaspargleuglyalaglugly
435440445
asngluglyphevalphealaleuleuargtyrleuargglycysarg
450455460
asnglnthrphehisleuglyalaargalaglypheleulysgluile
465470475480
arglysgluleuglulysthrargtrpglylysalalysglualaglu
485490495
hisvalvalleuthrasplysthrvalalaalaileargalaileile
500505510
aspasnaspalalysalaleuglyalaargleuleualaaspleuser
515520525
glyalaphevalalahistyralaserlysgluhispheserthrleu
530535540
tyrsergluilevallysalavallysaspalaprogluvalserser
545550555560
glyleuproargleulysleuleuleulysargalaaspglyvalarg
565570575
glytyrvalhisglyleuargaspthrarglyshisalaphealathr
580585590
lysleuproproproproalaproarggluleuaspaspproalathr
595600605
lysalaargtyrilealaleuleuargleutyraspglyprophearg
610615620
alatyralaserglyilethrglythralaleualaglyproalaala
625630635640
argalalysglualaalathralaleualaglnservalasnvalthr
645650655
lysalatyrseraspvalmetgluglyargserserargleuargpro
660665670
proasnaspglygluthrleuargglutyrleuseralaleuthrgly
675680685
gluthralathrglupheargvalglnileglytyrgluseraspser
690695700
gluasnalaarglysglnalaglupheilegluasntyrargargasp
705710715720
metleualaphemetphegluasptyrileargalalysglypheasp
725730735
trpileleulysilegluproglyalathralametthrargalapro
740745750
valleuprogluproileaspthrargglyglntyrgluhistrpgln
755760765
alaalaleutyrleuvalmethisphevalproalaseraspvalser
770775780
asnleuleuhisglnleuarglystrpglualaleuglnglylystyr
785790795800
gluleuvalglnaspglyaspalathraspglnalaaspalaargarg
805810815
glualaleuaspleuvallysargpheargaspvalleuvalleuphe
820825830
leulysthrglyglualaargphegluglyargalaalapropheasp
835840845
leulyspropheargalaleuphealaasnproalathrpheasparg
850855860
leuphemetalathrprothrthralaargproalagluaspasppro
865870875880
gluglyaspglyalasergluprogluleuargvalalaargthrleu
885890895
argglyleuargglnilealaargtyrasnhismetalavalleuser
900905910
aspleuphealalyshislysvalargaspglugluvalalaargleu
915920925
alagluilegluaspgluthrglnglulysserglnilevalalaala
930935940
glngluleuargthraspleuhisasplysvalmetlyscyshispro
945950955960
lysthrileserproglugluargglnsertyralaalaalailelys
965970975
thrileglugluhisargpheleuvalglyargvaltyrleuglyasp
980985990
hisleuargleuhisargleumetmetaspvalileglyargleuile
99510001005
asptyralaglyalatyrgluargaspthrglythrpheleuile
101010151020
asnalaserlysglnleuglyalaglyalaasptrpalavalthr
102510301035
ilealaglyalaalaasnthraspalaargthrglnthrarglys
104010451050
aspleualahispheasnvalleuaspargalaaspglythrpro
105510601065
aspleuthralaleuvalasnargalaargglumetmetalatyr
107010751080
asparglysarglysasnalavalproargserileleuaspmet
108510901095
leualaargleuglyleuthrleulystrpglnmetlysasphis
110011051110
leuleuglnaspalathrilethrglnalaalailelyshisleu
111511201125
asplysvalargleuthrvalglyglyproalaalavalthrglu
113011351140
alaargpheserglnasptyrleuglnmetvalalaalavalphe
114511501155
asnglyservalglnasnprolysproargargargaspaspgly
116011651170
aspalatrphislysproprolysproalathralaglnsergln
117511801185
proaspglnlysproproasnlysalaproseralaglyserarg
119011951200
leuproproproglnvalglygluvaltyrgluglyvalvalval
120512101215
lysvalileaspthrglyserleuglypheleualavalglugly
122012251230
valalaglyasnileglyleuhisileserargleuargargile
123512401245
arggluaspalaileilevalglyargargtyrargpheargval
125012551260
gluiletyrvalproprolysserasnthrserlysleuasnala
126512701275
alaaspleuvalargileasp
12801285
<210>5
<211>1175
<212>prt
<213>鸡肉杆菌
<400>5
metargilethrlysvallysilelysleuaspasnlysleutyrgln
151015
valthrmetglnlysgluglulystyrglythrleulysleuasnglu
202530
gluserarglysserthralagluileleuargleulyslysalaser
354045
pheasnlysserphehisserlysthrileasnserglnlysgluasn
505560
lysasnalathrilelyslysasnglyasptyrileserglnilephe
65707580
glulysleuvalglyvalaspthrasnlysasnilearglysprolys
859095
metserleuthraspleulysaspleuprolyslysaspleualaleu
100105110
pheilelysarglysphelysasnaspaspilevalgluilelysasn
115120125
leuaspleuileserleuphetyrasnalaleuglnlysvalprogly
130135140
gluhisphethraspglusertrpalaaspphecysglnglumetmet
145150155160
protyrargglutyrlysasnlyspheilegluarglysileileleu
165170175
leualaasnserilegluglnasnlysglypheserileasnproglu
180185190
thrpheserlysarglysargvalleuhisglntrpalailegluval
195200205
glngluargglyasppheserileleuaspglulysleuserlysleu
210215220
alagluiletyrasnphelyslysmetcyslysargvalglnaspglu
225230235240
leuasnaspleuglulyssermetlyslysglylysasnproglulys
245250255
glulysglualatyrlyslysglnlysasnphelysilelysthrile
260265270
trplysasptyrprotyrlysthrhisileglyleuileglulysile
275280285
lysgluasnglugluleuasnglnpheasnilegluileglylystyr
290295300
phegluhistyrpheproilelyslysgluargcysthrgluaspglu
305310315320
protyrtyrleuasnsergluthrilealathrthrvalasntyrgln
325330335
leulysasnalaleuilesertyrleumetglnileglylystyrlys
340345350
glnpheglyleugluasnglnvalleuaspserlyslysleuglnglu
355360365
ileglyiletyrgluglypheglnthrlysphemetaspalacysval
370375380
phealathrserserleulysasnileilegluprometargsergly
385390395400
aspileleuglylysarggluphelysglualailealathrserser
405410415
phevalasntyrhishisphepheprotyrpheprophegluleulys
420425430
glymetlysaspargglusergluleuilepropheglygluglnthr
435440445
glualalysglnmetglnasniletrpalaleuargglyservalgln
450455460
glnileargasngluilephehisserpheasplysasnglnlysphe
465470475480
asnleuproglnleuasplysserasnpheglupheaspalaserglu
485490495
asnserthrglylysserglnsertyrilegluthrasptyrlysphe
500505510
leupheglualaglulysasnglnleugluglnphepheilegluarg
515520525
ilelysserserglyalaleuglutyrtyrproleulysserleuglu
530535540
lysleuphealalyslysglumetlyspheserleuglyserglnval
545550555560
valalaphealaprosertyrlyslysleuvallyslysglyhisser
565570575
tyrglnthralathrgluglythralaasntyrleuglyleusertyr
580585590
tyrasnargtyrgluleulysglugluserpheglnalaglntyrtyr
595600605
leuleulysleuiletyrglntyrvalpheleuproasnphesergln
610615620
glyasnserproalaphearggluthrvallysalaileleuargile
625630635640
asnlysaspglualaarglyslysmetlyslysasnlyslyspheleu
645650655
arglystyralaphegluglnvalargglumetgluphelysgluthr
660665670
proaspglntyrmetsertyrleuglnserglumetarggluglulys
675680685
valarglysalaglulysasnasplysglypheglulysasnilethr
690695700
metasnpheglulysleuleumetglnilephevallysglypheasp
705710715720
valpheleuthrthrphealaglylysgluleuleuleuserserglu
725730735
glulysvalilelysgluthrgluileserleuserlyslysileasn
740745750
gluargglulysthrleulysalaserileglnvalgluhisglnleu
755760765
valalathrasnseralailesertyrtrpleuphecyslysleuleu
770775780
aspserarghisleuasngluleuargasnglumetilelysphelys
785790795800
glnserargilelyspheasnhisthrglnhisalagluleuilegln
805810815
asnleuleuproilevalgluleuthrileleuserasnasptyrasp
820825830
glulysasnaspserglnasnvalaspvalseralatyrphegluasp
835840845
lysserleutyrgluthralaprotyrvalglnthraspaspargthr
850855860
argvalserpheargproileleulysleuglulystyrhisthrlys
865870875880
serleuileglualaleuleulysaspasnproglnpheargvalala
885890895
alathraspileglnglutrpmethislysargglugluileglyglu
900905910
leuvalglulysarglysasnleuhisthrglutrpalagluglygln
915920925
glnthrleuglyalaglulysarggluglutyrargasptyrcyslys
930935940
lysileaspargpheasntrplysalaasnlysvalthrleuthrtyr
945950955960
leuserglnleuhistyrleuilethraspleuleuglyargmetval
965970975
glypheseralaleuphegluargaspleuvaltyrpheserargser
980985990
phesergluleuglyglygluthrtyrhisileserasptyrlysasn
99510001005
leuserglyvalleuargleuasnalagluvallysproilelys
101010151020
ilelysasnilelysvalileaspasnglugluasnprotyrlys
102510301035
glyasngluprogluvallyspropheleuaspargleuhisala
104010451050
tyrleugluasnvalileglyilelysalavalhisglylysile
105510601065
argasnglnthralahisleuservalleuglnleugluleuser
107010751080
metileglusermetasnasnleuargaspleumetalatyrasp
108510901095
arglysleulysasnalavalthrlyssermetilelysileleu
110011051110
asplyshisglymetileleulysleulysileaspgluasnhis
111511201125
lysasnphegluilegluserleuileprolysgluileilehis
113011351140
leulysasplysalailelysthrasnglnvalsergluglutyr
114511501155
cysglnleuvalleualaleuleuthrthrasnproglyasngln
116011651170
leuasn
1175
<210>6
<211>1285
<212>prt
<213>herbinixhemicellulosilytica
<400>6
metlysleuthrargargargileserglyasnservalaspglnlys
151015
ilethralaalaphetyrargaspmetserglnglyleuleutyrtyr
202530
aspsergluaspasnaspcysthrasplysvalileglusermetasp
354045
phegluargsertrpargglyargileleulysasnglygluaspasp
505560
lysasnprophetyrmetphevallysglyleuvalglyserasnasp
65707580
lysilevalcysgluproileaspvalaspseraspproaspasnleu
859095
aspileleuileasnlysasnleuthrglypheglyargasnleulys
100105110
alaproaspserasnaspthrleugluasnleuilearglysilegln
115120125
alaglyileprogluglugluvalleuprogluleulyslysilelys
130135140
glumetileglnlysaspilevalasnarglysgluglnleuleulys
145150155160
serilelysasnasnargilepropheserleugluglyserlysleu
165170175
valproserthrlyslysmetlystrpleuphelysleuileaspval
180185190
proasnlysthrpheasnglulysmetleuglulystyrtrpgluile
195200205
tyrasptyrasplysleulysalaasnilethrasnargleuasplys
210215220
thrasplyslysalaargserileserargalavalserglugluleu
225230235240
argglutyrhislysasnleuargthrasntyrasnargphevalser
245250255
glyaspargproalaalaglyleuaspasnglyglyseralalystyr
260265270
asnproasplysgluglupheleuleupheleulysgluvalglugln
275280285
tyrphelyslystyrpheprovallysserlyshisserasnlysser
290295300
lysasplysserleuvalasplystyrlysasntyrcyssertyrlys
305310315320
valvallyslysgluvalasnargserileileasnglnleuvalala
325330335
glyleuileglnglnglylysleuleutyrtyrphetyrtyrasnasp
340345350
thrtrpglngluasppheleuasnsertyrglyleusertyrilegln
355360365
valgluglualaphelyslysservalmetthrserleusertrpgly
370375380
ileasnargleuthrserphepheileaspaspserasnthrvallys
385390395400
pheaspaspilethrthrlyslysalalysglualailegluserasn
405410415
tyrpheasnlysleuargthrcysserargmetglnasphisphelys
420425430
glulysleualaphephetyrprovaltyrvallysasplyslysasp
435440445
argproaspaspaspilegluasnleuilevalleuvallysasnala
450455460
ilegluservalsertyrleuargasnargthrphehisphelysglu
465470475480
serserleuleugluleuleulysgluleuaspasplysasnsergly
485490495
glnasnlysileasptyrservalalaalaglupheilelysargasp
500505510
ilegluasnleutyraspvalphearggluglnileargserleugly
515520525
ilealaglutyrtyrlysalaaspmetileseraspcysphelysthr
530535540
cysglyleugluphealaleutyrserprolysasnserleumetpro
545550555560
alaphelysasnvaltyrlysargglyalaasnleuasnlysalatyr
565570575
ileargasplysglyprolysgluthrglyaspglnglyglnasnser
580585590
tyrlysalaleugluglutyrarggluleuthrtrptyrilegluval
595600605
lysasnasnaspglnsertyrasnalatyrlysasnleuleuglnleu
610615620
iletyrtyrhisalapheleuprogluvalarggluasnglualaleu
625630635640
ilethrasppheileasnargthrlysglutrpasnarglysgluthr
645650655
glugluargleuasnthrlysasnasnlyslyshislysasnpheasp
660665670
gluasnaspaspilethrvalasnthrtyrargtyrgluserilepro
675680685
asptyrglnglygluserleuaspasptyrleulysvalleuglnarg
690695700
lysglnmetalaargalalysgluvalasnglulysglugluglyasn
705710715720
asnasntyrileglnpheileargaspvalvalvaltrpalaphegly
725730735
alatyrleugluasnlysleulysasntyrlysasngluleuglnpro
740745750
proleuserlysgluasnileglyleuasnaspthrleulysgluleu
755760765
pheprogluglulysvallysserpropheasnilelyscysargphe
770775780
serileserthrpheileaspasnlysglylysserthraspasnthr
785790795800
seralaglualavallysthraspglylysgluaspglulysasplys
805810815
lysasnilelysarglysaspleuleucysphetyrleupheleuarg
820825830
leuleuaspgluasngluilecyslysleuglnhisglnpheilelys
835840845
tyrargcysserleulysgluargargpheproglyasnargthrlys
850855860
leuglulysgluthrgluleuleualagluleuglugluleumetglu
865870875880
leuvalargphethrmetproserileprogluileseralalysala
885890895
gluserglytyraspthrmetilelyslystyrphelysasppheile
900905910
glulyslysvalphelysasnprolysthrserasnleutyrtyrhis
915920925
seraspserlysthrprovalthrarglystyrmetalaleuleumet
930935940
argseralaproleuhisleutyrlysaspilephelysglytyrtyr
945950955960
leuilethrlyslysglucysleuglutyrilelysleuserasnile
965970975
ilelysasptyrglnasnserleuasngluleuhisgluglnleuglu
980985990
argilelysleulysserglulysglnasnglylysaspserleutyr
99510001005
leuasplyslysaspphetyrlysvallysglutyrvalgluasn
101010151020
leugluglnvalalaargtyrlyshisleuglnhislysileasn
102510301035
phegluserleutyrargilepheargilehisvalaspileala
104010451050
alaargmetvalglytyrthrglnasptrpgluargaspmethis
105510601065
pheleuphelysalaleuvaltyrasnglyvalleuglugluarg
107010751080
argpheglualailepheasnasnasnaspaspasnasnaspgly
108510901095
argilevallyslysileglnasnasnleuasnasnlysasnarg
110011051110
gluleuvalsermetleucystrpasnlyslysleuasnlysasn
111511201125
glupheglyalaileiletrplysargasnproilealahisleu
113011351140
asnhisphethrglnthrgluglnasnserlysserserleuglu
114511501155
serleuileasnserleuargileleuleualatyrasparglys
116011651170
argglnasnalavalthrlysthrileasnaspleuleuleuasn
117511801185
asptyrhisileargilelystrpgluglyargvalaspglugly
119011951200
glniletyrpheasnilelysglulysgluaspilegluasnglu
120512101215
proileilehisleulyshisleuhislyslysaspcystyrile
122012251230
tyrlysasnsertyrmetpheasplysglnlysglutrpilecys
123512401245
asnglyilelysglugluvaltyrasplysserileleulyscys
125012551260
ileglyasnleuphelyspheasptyrgluasplysasnlysser
126512701275
seralaasnprolyshisthr
12801285
<210>7
<211>30
<212>rna
<213>斯氏李斯特菌
<400>7
gacuaccucuauaugaaagaggacuaaaac30
<210>8
<211>29
<212>rna
<213>沙氏纤毛菌
<400>8
ccaccccaauaucgaaggggacuaaaaca29
<210>9
<211>31
<212>rna
<213>口腔纤毛菌
<400>9
gaccaccccaaaaaugaaggggacuaaaaca31
<210>10
<211>50
<212>rna
<213>斯氏李斯特菌
<400>10
gacuaccucuauaugaaagaggacuaaaaccaaacaugaucugggucauc50
<210>11
<211>55
<212>rna
<213>斯氏李斯特菌
<400>11
uaagagacuaccucuauaugaaagaggacuaaaaccaaacaugaucugggucauc55
<210>12
<211>49
<212>rna
<213>沙氏纤毛菌
<400>12
ccaccccaauaucgaaggggacuaaaacaggggcagagaugaugacccu49
<210>13
<211>58
<212>rna
<213>沙氏纤毛菌
<400>13
ggauuuagaccaccccaauaucgaaggggacuaaaacaggggcagagaugaugacccu58
<210>14
<211>51
<212>rna
<213>口腔纤毛菌
<400>14
gaccaccccaaaaaugaaggggacuaaaacaggggcagagaugaugacccu51
<210>15
<211>56
<212>rna
<213>口腔纤毛菌
<400>15
auuuagaccaccccaaaaaugaaggggacuaaaacaggggcagagaugaugacccu56
<210>16
<211>27
<212>rna
<213>人工序列
<220>
<223>合成序列
<400>16
agauagcccaagaaagagggcaauaac27
<210>17
<211>32
<212>rna
<213>人工序列
<220>
<223>合成序列
<400>17
guaacaauccccguagacaggggaacugcaac32
<210>18
<211>31
<212>rna
<213>人工序列
<220>
<223>合成序列
<400>18
caucaccgccaagacgacggcggacugaacc31
<210>19
<211>30
<212>rna
<213>人工序列
<220>
<223>合成序列
<400>19
aauuaucccaaaauugaagggaacuacaac30
<210>20
<211>30
<212>rna
<213>人工序列
<220>
<223>合成序列
<400>20
gaguaccucaaaacaaaagaggacuaaaac30
<210>21
<211>32
<212>rna
<213>人工序列
<220>
<223>合成序列
<400>21
gaccaccccaauaucgaaggggacuaaaacuu32
<210>22
<211>27
<212>rna
<213>人工序列
<220>
<223>合成序列
<400>22
caccccaauaucgaaggggacuaaaac27
<210>23
<211>27
<212>rna
<213>人工序列
<220>
<223>合成序列
<400>23
aaguagcccgauauagagggcaauaac27
<210>24
<211>27
<212>rna
<213>人工序列
<220>
<223>合成序列
<400>24
auacagcucgauauagugagcaauaag27
<210>25
<211>34
<212>rna
<213>人工序列
<220>
<223>合成序列
<400>25
ucacaucaccgccaagacgacggcggacugaacc34
<210>26
<211>28
<212>rna
<213>人工序列
<220>
<223>合成序列
<400>26
gaacagcccgauauagagggcaauagac28
- 还没有人留言评论。精彩留言会获得点赞!