用于治疗肌病的用途的MTMR2-S多肽的制作方法
本公开内容涉及用于治疗与mtm1突变或缺乏相关的疾病或紊乱的用途的mtmr2-s多肽或者产生或编码所述mtmr2-s多肽的核酸序列。本发明提供用于在有需要的受试者中治疗肌病或者与mtm1突变或缺乏相关的疾病或紊乱的组合物和方法。本发明涉及将mtmr2-s多肽递送至需要改善肌肉功能的受试者(例如患有中央核性肌病的受试者)的方法。
背景技术:
中央核性肌病(cnm)是一类先天性肌病,其特征在于肌肉力弱,并在组织学上由纤维萎缩、i型纤维占优势以及核中央化增加而确认,并非是继发于肌肉再生。在cnm的三种主要特征形式中,x连锁型中央核性肌病(也称为xlcnm、肌小管肌病-xlmtm或omim310400)是cnm中最常见和最严重的形式,具有新生儿发作并且死亡通常发生在生命的头几年(jungbluth,h.等,orphanetjraredis,2008.3:p.26)。产后期的生存需要密集的支持,通常包括胃造口术喂养和机械通气。目前对于这种紊乱尚无治愈方法,也没有有效的治疗。
xlcnm是由磷酸肌醇磷酸酶肌微管素(mtm1)突变造成的(laporte,j.等,naturegenetics,1996,13(2):p.175-82)。目前为止已经报道了约450个家族中的超过200种不同的mtm1突变,其中大部分导致蛋白大量减少。之前已对mtm1敲除或敲入小鼠进行了表征,其重现了cnm表型,具有典型的组织学特征,包括细胞器定位异常、细胞核定位错误和肌肉萎缩,并伴有肌肉力量相应下降(buj-belloa,laugelv,messaddeqn,zahreddineh,laportej,pellissierjf,mandeljl.,thelipidphosphatasemyotubularinisessentialforskeletalmusclemaintenancebutnotformyogenesisinmice,procnatlacadsciusa.2002年11月12日;99(23):15060-5.epub2002年10月21日;piersoncr,dulin-smithan,durbanan,marshallml,marshalljt,snyderad,naiyern,gladmanjt,chandlerds,lawlormw,buj-belloa,dowlingjj,beggsah.,hummolgenet.2012年2月15日;21(4):811-25.doi:10.1093/hmg/ddr512.epub2011年11月7日;molcellbiol.2013年1月;33(1):98-110.doi:10.1128/mcb.01075-12.epub2012年10月29日.defectiveautophagyandmtorc1signalinginmyotubularinnullmice.fetalverokm,yuy,goetschkesm,liangg,valdezra,gouldt,triantafellowe,berglings,loureiroj,eashj,linv,porterja,finanpm,walshk,yangy,maox,murphylo)。已在具有不同形式cnm的数种动物模型和患者中检测到与异常兴奋-收缩偶联相关的三联体(triads)结构的缺陷,在全部cnm形式中识别出共同缺陷(toussaint,a.等,actaneuropathol,2011年2月;121(2):253-66)。这与所提出的mtm1在三联体肌质网部分的磷酸肌醇水平调节中的作用相一致。
肌微管素相关蛋白2中磷酸酶活性的丧失与charcot-marie-tooth病4b1型相关(charcot-marie-toothtype4biscausedbymutationsinthegeneencodingmyotubularin-relatedprotein-2.,bolinoa,mugliam,confortifl,leguerne,salihma,georgioudm,christodoulouk,hausmanowa-petrusewiczi,mandichp,schenonea,gambardellaa,bonof,quattronea,devotom,monacoap.charcot-marie-toothtype4biscausedbymutationsinthegeneencodingmyotubularin-relatedprotein-2-natgenet.2000年5月;25(1):17-9)。
肌微管素和肌微管素相关蛋白(mtm)定义了涉及不同神经肌肉疾病的一个保守蛋白家族(raess,m.a.,friant,s.,cowling,b.s.和laporte,j.(2016).wanted-deadoralive:myotubularins,alargedisease-associatedproteinfamily.advbiolregul.)。它们已被归类在磷酸酶超家族中。在人中,8种肌微管素共享双特异性磷酸酶和酪氨酸中发现的c(x)5r基序并显示酶活性,而其它6种肌微管素缺乏此基序并被称为非活性磷酸酶(dead-phosphatase)。出乎意料的是,发现具有酶活性的肌微管素并不作用于蛋白质,而是使磷酸肌醇(ppin)(集中在特定膜亚结构域中的脂质)脱磷酸(blondeau,f.,laporte,j.,bodin,s.,superti-furga,g.,payrastre,b.和mandel,j.l.(2000).myotubularin,aphosphatasedeficientinmyotubularmyopathy,actsonphosphatidylinositol3-kinaseandphosphatidylinositol3-phosphatepathway.hummolgenet9,2223-2229.;taylor,g.s.,maehama,t.和dixon,j.e.(2000).inauguralarticle:myotubularin,aproteintyrosinephosphatasemutatedinmyotubularmyopathy,dephosphorylatesthelipidsecondmessenger,phosphatidylinositol3-phosphate.procnatlacadsciusa97,8910-8915)。ppin是脂质第二信使,其涉及宽范围的细胞过程,包括信号传导和细胞内组织化(vicinanza,m.,d'angelo,g.,dicampli,a.和dematteis,m.a.(2008).functionanddysfunctionofthepisysteminmembranetrafficking.emboj27,2457-2470.)。肌微管素是使磷脂酰肌醇3-磷酸(ptdins3p)和磷脂酰肌醇3,5-二磷酸(ptdins(3,5)p2)脱磷酸的ppin3-磷酸酶,导致产生ptdins5p(berger,p.,bonneick,s.,willi,s.,wymann,m.和suter,u.(2002).inauguralarticle:myotubularin,aproteintyrosinephosphatasemutatedinmyotubularmyopathy,dephosphorylatesthelipidsecondmessenger,phosphatidylinositol3-phosphate.procnatlacadsciusa97,8910-8915;tronchere,h.,laporte,j.,pendaries,c.,chaussade,c.,liaubet,l.,pirola,l.,mandel,j.l.和payrastre,b.(2004).productionofphosphatidylinositol5-phosphatebythephosphoinositide3-phosphatasemyotubularininmammaliancells.(tronchèreh,laportej,pendariesc,chaussadec,liaubetl,pirolal,mandeljl,payrastreb.jbiolchem.2004年2月20日;279(8):7304-12.epub2003年12月1日)。ptdins5p涉及转录调控和生长因子信号传导,而ptdins3p和ptdins(3,5)p2调节膜运输和自噬。ptdins3p通过ptdins被ii类和iii类ptdins3-激酶磷酸化而产生,ptdins(3,5)p2主要由ptdins3p被pikfyve磷酸化而获得(jin,n.,lang,m.j.和weisman,l.s.(2016).phosphatidylinositol3,5-bisphosphate:regulationofcellulareventsinspaceandtime.biochemsoctrans44,177-184;schink,k.o.,raiborg,c.和stenmark,h.(2013).phosphatidylinositol3-phosphate,alipidthatregulatesmembranedynamics,proteinsortingandcellsignalling.bioessays35,900-912)。它们将蛋白质募集至特定的内体池(endosomalpools)或募集至内质网,分别使得内体能够成熟和相互转化或使得自噬泡能够形成。例如,eea1的fyve(fab1-yotb-vac1-eea1)结构域特异性结合集中在早期内体上的ptdins3p,以调节内体融合和货物递送(schink等,2013,同上)。非活性肌微管素与活性同源物寡聚并调节其酶活性和/或亚细胞定位(berger,p.,berger,i.,schaffitzel,c.,tersar,k.,volkmer,b.和suter,u.(2006).multi-levelregulationofmyotubularin-relatedprotein-2phosphataseactivitybymyotubularin-relatedprotein-13/set-bindingfactor-2.hummolgenet15,569-579.;kim等,2003,同上;nandurkar,h.h.,layton,m.,laporte,j.,selan,c.,corcoran,l.,caldwell,k.k.,mochizuki,y.,majerus,p.w.和mitchell,c.a.(2003).identificationofmyotubularinasthelipidphosphatasecatalyticsubunitassociatedwiththe3-phosphataseadapterprotein,3-pap.procnatlacadsciusa100,8660-8665)。除活性或非活性磷酸酶结构域外,肌微管素还共享结合至ppin或蛋白的ph-gram(普列克底物蛋白同源,葡萄糖基转移酶,rab样gtpase激活物和肌微管素)结构域以及牵涉其寡聚化的卷曲螺旋结构域(raess等,2016,同上)。
在人中有14种肌微管素,而在酵母中有1种活性肌微管素(酿酒酵母(saccharomycescerevisiae))(lecompte,o.,poch,o.和laporte,j.(2008).ptdins5pregulationthroughevolution:rolesinmembranetrafficking?trendsbiochemsci33,453-460.;raess等,2016,同上)。酵母肌微管素(ymr1p)调节液泡蛋白分选和片段化(parrish,w.r.,stefan,c.j.和emr,s.d.(2004).essentialroleforthemyotubularin-relatedphosphataseymr1pandthesynaptojanin-likephosphatasessjl2pandsjl3pinregulationofphosphatidylinositol3-phosphateinyeast.molbiolcell15,3567-3579.)。人肌微管素在酵母中的过表达因其磷酸酶活性而引起液泡增大(amoasii,l.,bertazzi,d.l.,tronchere,h.,hnia,k.,chicanne,g.,rinaldi,b.,cowling,b.s.,ferry,a.,klaholz,b.,payrastre,b.等(2012).phosphatase-deadmyotubularinamelioratesx-linkedcentronuclearmyopathyphenotypesinmice.plosgenet8,e1002965;blondeau等,2000,同上)。如前所述,在人中,肌微管素1(mtm1)中的功能丧失性突变引起称为xlcnm的严重先天性肌病,而活性肌微管素相关2基因蛋白(mtmr2)或非活性肌微管素相关蛋白mtmr13中的突变引起charcot-marie-tooth(cmt)周围神经病(分别为cmt4b1,omim601382和cmt4b2,omim604563)(azzedine,h.,bolino,a.,taieb,t.,birouk,n.,diduca,m.,bouhouche,a.,benamou,s.,mrabet,a.,hammadouche,t.,chkili,t.等(2003).mutationsinmtmr13,anewpseudophosphatasehomologueofmtmr2andsbf1,intwofamilieswithanautosomalrecessivedemyelinatingformofcharcot-marie-toothdiseaseassociatedwithearly-onsetglaucoma.amjhumgenet72,1141-1153.;bolino,a.,muglia,m.,conforti,f.l.,leguern,e.,salih,m.a.,georgiou,d.m.,christodoulou,k.,hausmanowa-petrusewicz,i.,mandich,p.,schenone,a.等(2000).charcot-marie-toothtype4biscausedbymutationsinthegeneencodingmyotubularin-relatedprotein-2.natgenet25,17-19;senderek,j.,bergmann,c.,weber,s.,ketelsen,u.p.,schorle,h.,rudnik-schoneborn,s.,buttner,r.,buchheim,e.和zerres,k.(2003).mutationofthesbf2gene,encodinganovelmemberofthemyotubularinfamily,incharcot-marie-toothneuropathytype4b2/11p15.hummolgenet12,349-356)。此外,mtmr5(sbf1)中的推定突变与cmt4b3(omim615284)和轴索神经病相关(alazami,a.m.,alzahrani,f.,bohlega,s.和alkuraya,f.s.(2014).setbindingfactor1(sbf1)mutationcausescharcot-marie-toothdiseasetype4b3.neurology82,1665-1666;manole,a.,horga,a.,gamez,j.,raguer,n.,salvado,m.,sanmillan,b.,navarro,c.,pittmann,a.,reilly,m.m.和houlden,h.(2016).sbf1mutationsassociatedwithautosomalrecessiveaxonalneuropathywithcranialnerveinvolvement.neurogenetics;nakhro等,2013)。
因此,尽管肌微管素普遍地表达,一种肌微管素的缺乏并不会被其同源物完全补偿。而且,相关疾病影响不同组织。值得注意的是,基于它们的序列,mtm1和mtmr2是同一进化亚组的一部分(lecompte等,2008,同上)。因此,这表明未表征的组织特异性功能潜在地反映不同的活性或不同的相互作用因子。
因此,对于合适的中央核性肌病治疗方法,特别是对于新的、更有效的治疗剂,具有显著需求。
本文中,在酵母和小鼠中比较了mtm1和mtmr2的体内功能,发现mtmr2的特定亚型(isoform)具有相当有效地对mtm1丧失进行补偿的能力。此类mtmr2形式可以挽救mtm1ko小鼠表现出的肌病,这使其成为治疗中央核性肌病、更具体而言是治疗xlcnm的有效试剂。
技术实现要素:
本公开内容提供了用于治疗中央核性肌病或者用于治疗与mtm1突变或缺乏相关的疾病或紊乱的方法和组合物。本发明提供了用于在有需要的受试者中治疗肌病或者与mtm1突变或缺乏相关的疾病或紊乱的组合物和方法。本发明涉及将特定的mtmr2多肽(在本文中称为mtmr2的短亚型)递送至需要改善肌肉功能的受试者的方法。本发明的组合物和方法在受试者中增加肌肉形成并改善肌肉功能。
在一个实施方式中,本发明可用于治疗患有肌病的个体。在另一实施方式中,本发明可用于治疗患有xlcnm的个体。本发明改善受困扰受试者的肌肉功能并延长其存活时间。然而,本发明不限于患有xlcnm的受试者。而是,本发明适用于在需要改善肌肉功能的任何受试者中改善肌肉功能或者适用于治疗与mtm1突变或缺乏相关的疾病或紊乱。
在一个具体的方面,本发明涉及一种组合物,所述组合物包含特定的mtmr2多肽(在本文中称为mtmr2的短亚型)或者产生或编码所述特定的mtmr2多肽的核酸序列。所述组合物可以用于治疗中央核性肌病或者用于治疗与mtm1突变或缺乏相关的疾病或紊乱。
在具体实施方式中,中央核性肌病选自于由如下疾病所组成的组:x连锁型cnm(xlcnm)、常染色体隐性cnm(arcnm)和常染色体显性cnm(adcnm)。在优选实施方式中,所述中央核性肌病是xlcnm。
本发明还提供了包含mtmr2多肽的短亚型的分离的多肽以及包含mtmr2多肽的短亚型与药物载体的组合的药物组合物。
本发明还公开了可用于产生此类多肽的构建体。另外,本发明涉及制备此类多肽或编码它们的构建体的方法。此外,本文公开了使用所述多肽例如用于治疗与mtm1突变或缺乏相关的疾病或紊乱的方法。
在详细描述本发明之后,本发明的这些和其它目的以及实施方式将变得更加显而易见。
附图说明
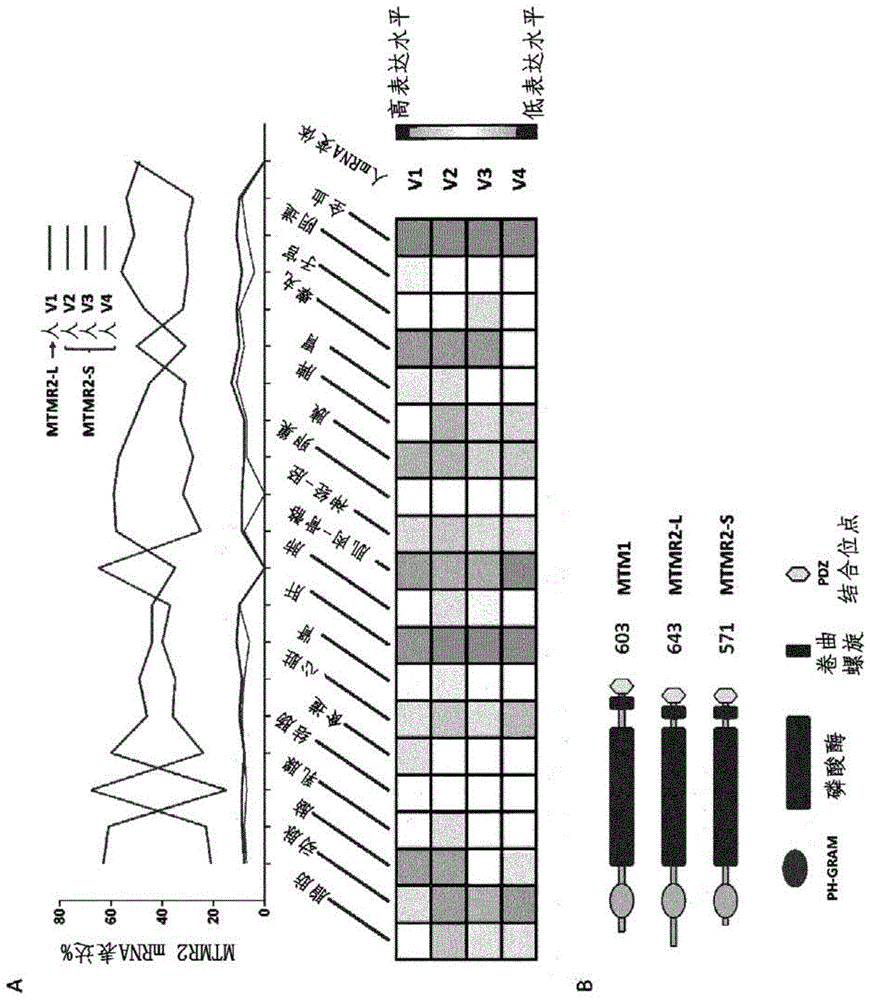
图1:mtmr2剪接亚型差异表达并编码长蛋白亚型和短蛋白亚型。(a)来自gtex数据库挖掘的20种人组织中mtmr2mrna亚型v1至v4的比较表达(上)。与v1相比,人mtmr2v2亚型包含额外的外显子1a和2a;v3包含外显子1a;v4包含外显子1a和2b。每种亚型的独立的组织表达(下)。(b)与mtm1相比,由v1mrna亚型编码的蛋白结构域mtmr2-l以及由其它亚型编码的mtmr2-s。
图2:短mtmr2亚型而非长mtmr2亚型展现出mtm1样活性。在ymr1δ酵母细胞中使用高拷贝数质粒2μ外源表达人mtm1以及mtmr2长亚型和短亚型。(a)在相同样品的两个独立印迹中,使用抗mtm1或抗mtmr2抗体通过western印迹对外源表达的人肌微管素进行检测。使用具有空载体的野生型(wt)和ymr1δ酵母菌株作为对照。使用pgk1p作为上样对照。该印迹代表至少3个独立实验。(b)过表达无标签肌微管素的酵母细胞中液泡形态的定量。每个构建体分析三个克隆;每个克隆计数的细胞数在上方标示。数据表示平均值±s.e.m.****p<0.0001,ns不显著(anova检验)。(c)带gfp标签的人肌微管素的定位。通过亲脂性染料fm4-64和nomarski微分相差来评价液泡形态。ymr1δ酵母细胞和表达mtmr2-l的细胞显示出碎片状液泡,而过表达mtm1和mtmr2-s的细胞具有大液泡。(d)在表达无标签肌微管素和与ptdins3p特异性结合的带dsred标签的fyve结构域的酵母克隆中的fyve点状定位(punctuatedlocalization)。(e)通过计数如(d)中所示的每个细胞的fyve阳性点的数量来进行ptdins3p定量。ptdins3p在mtm1和mtmr2-s表达时降低,但对于mtmr2-l不降低。数据表示平均值±s.e.m.*p<0.05,**p<0.01(anova检验)。(f)通过对来自过表达无标签肌微管素的酵母细胞的总脂质提取物的质量分析进行ptdins5p定量。每个构建体分析三个克隆。数据表示平均值±s.e.m.*p<0.05(anova检验)。
图3:mtmr2短亚型与mtm1类似地在mtm1ko肌病小鼠中挽救肌肉重量和力量。向来自2-3周龄mtm1ko小鼠的ta肌肉注射表达肌微管素的aav2/1,并在4周后进行分析。(a)使用抗mtm1或抗mtmr2抗体通过western印迹检测外源表达的人肌微管素;使用gapdh作为上样对照。非特异性条带以星号标示。该印迹代表每个构建体,每个构建体至少分析10份肌肉。(b)与注射空aav的对侧腿相比,表达人肌微管素的ta肌肉重量的比例。mtmr2-s与mtm1类似地改善了肌肉质量,而mtmr2-l没有效果。将注射空aav的mtm1ko小鼠设置为1的值。n>10。数据表示平均值±s.e.m.****p<0.0001,ns不显著(anova检验)。(c)ta肌肉的比最大力(specificmaximalforce)(绝对值)。两种mtmr2亚型均改善肌肉力量。n>7。数据表示平均值±s.e.m.**p<0.01,****p<0.0001,ns不显著(anova检验)。
图4:长mtmr2亚型和短mtmr2亚型均改善mtm1ko小鼠的组织学标志(hallmark)。向来自2-3周龄mtm1ko小鼠的ta肌肉注射表达肌微管素的aav2/1,并在4周后进行分析。(a)ta肌肉切片的苏木精-伊红染色。比例尺100μm。(b)ta肌肉切片的琥珀酸脱氢酶(sdh)染色。比例尺100μm。(c)纤维面积的定量。将纤维尺寸分为200μm2间隔的组,并表示为每组中总纤维的百分比。8只小鼠的n>1000。(d)超过800μm2的纤维的百分比。n>8。数据表示平均值±s.e.m.*p<0.05,***p<0.001,****p<0.0001(anova检验)。wt的值在统计学上不同于所有mtm1ko注射组。(e)在ta肌肉中的核定位。定位良好的外周核的百分比。n>6只动物。数据表示平均值±s.e.m.***p<0.001,****p<0.0001(anova检验)。wt的值在统计学上不同于所有mtm1ko注射组。
图5:mtmr2亚型挽救mtm1ko肌肉的肌肉超微结构和三联体形态。向来自mtm1ko小鼠的ta肌肉注射表达肌微管素的aav2/1。(a)显示肌节(sarcomere)、线粒体和三联体组织(triadorganization)的电子显微术照片。比例尺1μm。代表性三联体显示在缩放方块中。(b)对每个肌节中组织良好的三联体的数量的定量。对2只小鼠每只n>20张图像。所有表达肌微管素的肌肉的定量均不同于mtm1ko。数据表示平均值±s.e.m.*p<0.05,****p<0.0001(anova检验)。
图6:mtmr2-s短亚型在mtm1ko小鼠中减少,并且其过表达使ptdins3p水平正常化。(a)通过竞争性elisa在来自表达不同肌微管素的mtm1ko小鼠的ta肌肉和wt肌肉中对ptdins3p水平的定量。n>3只小鼠。数据表示平均值±s.e.m.*p<0.05,**p<0.01,***p<0.001(anova检验)。表达不同肌微管素的mtm1ko肌肉中的ptdins3p水平与wt对照无统计学差异。(b)通过qrt-pcr,相比于wt小鼠对mtm1ko小鼠的ta肌肉中的mtmr2亚型(v1至v4)进行定量。n>6。每种亚型以独立的比例表示,将wt小鼠中的表达设为1的值。数据表示平均值±s.d.**p<0.01,***p<0.001,****p<0.0001,ns不显著(学生t检验)。(c)通过qrt-pcr,相比于对照对mtm1患者肌肉中的mtmr2亚型(v1至v4)进行定量。n=3。每种亚型以独立的比例表示,将对照患者中的表达设为1的值。数据表示平均值±s.d.标示出每种亚型的p值(学生t检验)。
图7:人和小鼠中的mtmr2mrna和蛋白亚型。(a)小鼠mtmr2的基因组结构和mrna亚型。可变外显子1a或2a的任何组合的纳入带来提前终止密码子,并暴露(unmask)外显子3中的可变起始位点。鼠类mtmr2v1编码mtmr2-l,而亚型v2至v4编码mtmr2-s。(b)人和小鼠mtm1、mtmr2-l和mtmr2-s的n端区域的蛋白比对。ph-gram结构域在位置75处开始。(c)由来自肌肉的rt-pcr产物测序而来的小鼠可变外显子1a和2a的序列。(d)对来自从wt和mtm1ko小鼠分离的ta肌肉以及来自wt肝脏的cdna进行的mtmr2的外显子1和外显子3之间的pcr。检测到4种mrna变体。
图8:mtmr2亚型的表达不在wt小鼠中诱导肌肉肥厚。在3周龄时向wt小鼠的ta肌肉注射表达肌微管素的aav2/1,并在4周后进行分析。与注射空aav的对侧腿相比,表达人肌微管素的ta肌肉重量的比例。将wtta肌肉重量设为1的值。n>5。数据表示平均值±s.e.m.无显著差异(anova检验)。
图9:mtmr2-s亚型改善肌病小鼠的体重。对过表达不同肌微管素的mtm1ko或wt小鼠从3周龄到最大10周龄的体重进行测量。
图10:mtmr2-s亚型挽救mtm1ko小鼠的肌肉力量。通过从3周龄至10周龄每周进行悬挂测试(hangingtest)对过表达不同肌微管素的mtm1ko或wt小鼠的肌肉力量进行评价。
具体实施方式
除非另有定义,本文所使用的全部技术术语和科学术语具有与本发明所属领域中的普通技术人员所一般理解的含义相同的含义。
冠词“一个/一种(a/an)”在本文中用于指代一个/一种或指代多于一个/一种(即指代至少一个/一种)该冠词的语法对象。举例来说,“一个/一种要素(anelement)”是指一个/一种要素或多于一个/一种要素。
当涉及诸如量和持续时间(temporalduration)等可测量值时,本文所使用的“约/大约(about)”意在包括与规定值相差±20%或±10%、更优选±5%、甚至更优选±1%、并且还更优选±0.1%的变化,因为此类变化适合于实施所公开的方法或组合物。
范围:在整个本公开内容中,可以用范围格式来呈现本发明的各个方面。应当理解的是,范围格式的描述仅是为了方便和简洁,而不应被解释为对本发明范围的不灵活的限制。因此,应当认为范围的描述已具体公开了所有可能的子范围以及该范围内的各个数值。例如,应当认为例如从1到6的范围的描述已具体公开了例如从1到3、从1到4、从1到5、从2到4、从2到6、从3到6等子范围以及该范围内的各个数字(例如1、2、2.7、3、4、5、5.3和6)。无论范围的广度如何,这都适用。
根据本发明,术语“包括/包含/含有(comprise(s)/comprising)”(以及其它相当的术语,例如“containing”和“including”)是“开放式”的,并且通常可以解释为包括所有具体提到的特征以及任何任选的、额外的和未指定的特征。根据具体实施方式,除非另外说明,在包括指定特征以及不实质上影响所要求保护的发明的基本和新颖特征的任何任选的、额外的和未指定的特征的情况下,也可以解释为短语“基本上由……组成”;或在仅包括指定特征的情况下,也可以解释为短语“由……组成”。
术语“多肽”、“肽”和“蛋白”在本文中可互换使用,指通过肽键共价连接的氨基酸残基的聚合物。该术语适用于其中一个或多个氨基酸残基是相应的天然存在的氨基酸的人工化学模拟物的氨基酸聚合物以及适用于天然存在的氨基酸聚合物和非天然存在的氨基酸聚合物。“多肽”包括例如生物活性片段、基本上同源的多肽、寡肽、同二聚体、异二聚体、多肽的变体、修饰多肽、衍生物、类似物、融合蛋白等。多肽包括天然肽、重组肽、合成肽或它们的组合。
本文所使用的“治疗疾病或紊乱”是指降低患者经历疾病或紊乱的症状的频率。疾病和紊乱在本文中可互换使用。本文所使用的术语“治疗”疾病是指降低受试者经历的疾病或紊乱的至少一种迹象(sign)或症状的频率或严重性。在本发明的上下文中,术语治疗指治愈性(curative)、对症性(symptomatic)和预防性(preventive)处理。本文所使用的术语疾病的“治疗”是指任何旨在延长受试者(或患者)寿命的行为,如疾病进展的延迟和治疗。可将治疗设计为根除疾病、阻止疾病进展和/或促进疾病康复(regression)。术语疾病的“治疗”也指任何旨在减少与疾病相关的症状(如张力减退和肌肉力弱)的行为。更具体而言,根据本发明的治疗旨在推迟中央核性肌病表型或症状的出现、改善运动(motor)和/或肌肉行为和/或寿命。
如果疾病或紊乱的症状的严重性、患者经历此类症状的频率降低或这两者均降低,则疾病或紊乱“缓解(alleviated)”。“治疗性(therapeutic)”治疗是向表现出病理学迹象的受试者给予的治疗,其目的是减少或消除这些迹象中的至少一种或全部。
本文所使用的短语“治疗有效量”是指足以预防或治疗或有效地预防或治疗(延缓或预防其发作、阻止其进展、抑制、减少或逆转)疾病或紊乱的量,包括向受试者提供有益效果或减轻此类疾病的症状。
术语“患者”、“受试者”、“个体”等在本文中可互换使用,指适用于本文所述方法的任何动物或其细胞(无论是体外还是原位)。在某些非限制性实施方式中,患者、受试者或个体为人。优选地,受试者为人类患者,无论其年龄或性别。也包括新生儿、婴儿、儿童。
“编码”是指多核苷酸中核苷酸的特定序列(例如基因、cdna或mrna)在生物过程中充当合成其它聚合物和大分子(所述聚合物和大分子具有规定的核苷酸序列(即rrna、trna和mrna)或规定的氨基酸序列以及由此产生的生物学性质)的模板的固有性质。因此,如果对应于基因的mrna的转录和翻译在细胞或其它生物系统中产生蛋白,则该基因编码该蛋白。编码链(其核苷酸序列与mrna序列相同且通常在序列表中提供)和非编码链(用作基因或cdna转录的模板)均可以称为编码该基因或cdna的蛋白或其它产物。
“表达载体”是指包含重组多核苷酸的载体,所述重组多核苷酸包含可操作地连接至待表达的核苷酸序列的表达调控序列(在本文中称为构建体)。表达载体包含足够的用于表达的顺式作用元件;其它用于表达的元件可以由宿主细胞提供或在体外表达系统中提供。表达载体包括并入了所述重组多核苷酸的本领域已知的所有表达载体,如粘粒、质粒(例如裸露的或包含在脂质体中的)和病毒(例如慢病毒、逆转录病毒、腺病毒和腺相关病毒)。因此,术语“载体”包括自主复制质粒或病毒。该术语也应解释为包括有助于将核酸转移至细胞中的非质粒和非病毒化合物,例如聚赖氨酸化合物、脂质体等。病毒载体的实例包括但不限于腺病毒载体、腺相关病毒载体、逆转录病毒载体等。因此,将构建体并入表达载体中。
“同源的(homologous)”是指两个多肽之间或两个核酸分子之间的序列相似性或序列同一性。当两个被比较的序列中的位置均被相同的碱基或氨基酸单体亚单元占据时(例如,如果两个dna分子中的每一个的位置均被腺嘌呤占据),则分子在该位置是同源的。两个序列之间的同源性百分比是两个序列共享的匹配或同源位置数除以被比较的位置数×100的函数。例如,如果两个序列中10个位置中的6个是匹配或同源的,则该两个序列是60%同源的。举例而言,dna序列attgcc和tatggc具有50%的同源性。通常,当比对两个序列时,进行比较以给出最大同源性。
“分离的”是指由天然状态发生改变或去除。例如,天然存在于活动物中的核酸或肽不是“分离的”,而与其天然状态的共存物质部分或完全分离的相同核酸或肽是“分离的”。分离的核酸或蛋白质能够以基本纯化的形式存在,或者可以存在于非天然环境(例如宿主细胞)中。
在本发明的上下文中,对于常见的核酸碱基使用以下缩写。“a”是指腺嘌呤,“c”是指胞嘧啶,“g”是指鸟嘌呤,“t”是指胸腺嘧啶,并且“u”是指尿嘧啶。
除非另有说明,否则“编码氨基酸序列的核苷酸序列”包括彼此为简并形式并且编码相同氨基酸序列的所有核苷酸序列。在编码蛋白的核苷酸序列在某些版本中可能包含一个或多个内含子的程度上,短语编码蛋白或rna的核苷酸序列也可以包含内含子。
本文所使用的术语“多核苷酸”定义为核苷酸链。此外,核酸是核苷酸的聚合物。因此,本文所使用的核酸和多核苷酸是可互换的。本领域技术人员具有核酸是可被水解成单体“核苷酸”的多核苷酸的常识。单体核苷酸可以被水解为核苷。本文所使用的多核苷酸包括但不限于通过本领域可用的任何手段获得的所有核酸序列,所述手段包括但不限于重组手段(即从重组文库或细胞基因组克隆核酸序列,使用常规克隆技术和pcrtm等)以及通过合成手段。
本文所使用的术语“启动子”定义为起始多核苷酸序列的特异性转录所需的被细胞的合成机制或导入的合成机制识别的dna序列。
本文所使用的术语“启动子/调控序列”是指表达可操作地连接至启动子/调控序列的基因产物所需的核酸序列。在一些情况下,该序列可以是核心启动子序列;在其它情况下,该序列也可以包含增强子序列和表达基因产物所需的其它调控元件。例如,启动子/调控序列可以是以组织特异性方式表达基因产物的启动子/调控序列。
“组成型”启动子是这样的核苷酸序列:当与编码或指定基因产物的多核苷酸可操作地连接时,所述核苷酸序列在细胞的大多数或全部生理条件下引起该基因产物在细胞中产生。
“诱导型”启动子是这样的核苷酸序列:当与编码或指定基因产物的多核苷酸可操作地连接时,基本上仅当对应于该启动子的诱导物存在于细胞中时,所述核苷酸序列才引起该基因产物在细胞中产生。
“组织特异性”启动子是这样的核苷酸序列:当与编码或指定基因产物的多核苷酸可操作地连接时,基本上仅当细胞为对应于该启动子的组织类型的细胞时,所述核苷酸序列才引起该基因产物在细胞中产生。
本发明的mtmr2多肽(本文中也称为短亚型mtmr2或mtmr2-s)优选为人mtmr2的短剪接的天然存在的亚型,其长度为571个氨基酸。所述mtmr2多肽由seqidno:1表示。更具体而言,所述mtmr2多肽的短亚型不包括天然存在的长链人mtmr2多肽。
本文公开了由seqidno:1表示的所述mtmr2亚型具有相当有效地对mtm1丧失进行补偿的能力。此类mtmr2亚型能够挽救mtm1ko小鼠表现出的肌病,这使其成为用于治疗中央核性肌病(更具体地,用于治疗xlcnm)的有效试剂。这种方法可以引起肌肉力量、尺寸和功能的持续改善。
在一个方面,本文所使用的mtmr2-s包含与seqidno:1具有至少90%同一性(或同源性)的氨基酸序列或者其生物活性片段或变体。在一些实施方式中,mtmr2多肽包含与seqidno:1具有至少80%、85%、90%、95%、97%、98%、99%或100%同一性并且长度等于或小于571个氨基酸的氨基酸序列或者其生物活性片段或变体。
如本文所使用的,本文所使用的mtmr2-s包括人mtmr2的短剪接的天然存在的亚型(如上所述其长度为571个氨基酸并由seqidno:1表示)的各种剪接亚型、片段、变体、融合蛋白和修饰形式。天然存在的亚型mtmr2-s多肽的此类亚型、片段或变体、融合蛋白和修饰形式具有至少一部分与天然存在的亚型mtmr2-s多肽具有实质的序列同一性的氨基酸序列,并保留天然存在的mtmr2-s多肽的至少一种功能。在某些实施方式中,天然存在的亚型mtmr2-s多肽的生物活性片段、变体或融合蛋白包含与seqidno:1的天然存在的亚型mtmr2-s具有至少80%、85%并优选至少90%、95%、97%、98%、99%或100%同一性的氨基酸序列。本文所使用的“片段”理解为包括表现出如本文所述的“生物活性”的生物活性片段或生物活性变体。即,mtmr2-s的生物活性片段或变体表现出可以被测量和测试的生物活性。例如,生物活性片段或变体表现出与天然(即野生型或正常)mtm1蛋白相同或基本相同的生物活性,并且此类生物活性可以通过该片段或变体例如裂解或水解本领域已知的内源性磷酸肌醇底物或用于体外测定的人工磷酸肌醇底物的能力(即磷酸肌醇磷酸酶活性)进行评价。本文描述了评价任何这些标准的方法,并且必须更具体地参考以下实施例,所述实施例中在表达aav载体或aav肌微管素构建体的mtm1ko小鼠的肌肉提取物中借由elisa进行了ptdins3p定量,或者借由通过具有特异性ptdins3p结合能力的由串联fyve蛋白结构域组成的生物传感器进行ptdins3p检测。如下文在实施例的部分中所述(也参见图6a),当表达mtm1或天然存在的亚型mtmr2-s时,ptdins3p水平被正常化至wt水平。本文所使用的“基本上相同”是指为对照(例如实施例中的ko+mtm1或wt+空aav)(对该对照进行了参数测量)的至少70%的任何参数(例如活性)。在某些实施方式中,“基本上相同”也指为对照(对该对照进行了参数测量)的至少75%、80%、85%、90%、92%、95%、97%、98%、99%、100%、102%、105%或110%的任何参数(例如活性)。
在某些实施方式中,本文公开的任何前述或后述mtmr2-s多肽可用于嵌合多肽,所述嵌合多肽进一步包含增强体内稳定性、体内半衰期、摄取/给药和/或纯化中的一种或多种的一个或多个多肽部分。
本发明提供了增加mtmr2-s多肽或其生物活性片段或变体在肌肉中的表达的组合物。例如,在一个实施方式中,该组合物包含产生或编码mtmr2-s多肽或其生物学功能片段或变体的分离的核酸序列。如本文所述,递送包含此类核酸序列的组合物改善肌肉功能。此外,递送包含此类核酸序列的组合物延长具有mtm1功能丧失性突变的受试者的存活。
本发明还涉及药物组合物,该药物组合物包含如上定义的mtmr2-s多肽或用于产生此类多肽的构建体与药物载体的组合。还公开了所述组合物用于中央核性肌病的治疗或用于改善肌肉功能的治疗的用途。
本发明进一步涉及用于中央核性肌病的治疗或用于改善肌肉功能的治疗的方法,其中,该方法包括向需要此类治疗的受试者给予治疗有效量的如上定义的mtmr2-s多肽或提供该多肽的构建体的步骤。
最后,本发明涉及如上定义的mtmr2-s多肽或提供该多肽的构建体在制备用于与mtm1突变或缺乏相关的疾病或紊乱的治疗、用于中央核性肌病的治疗或用于改善肌肉功能的治疗的药物组合物中的用途。
在一个实施方式中,该组合物包含分离的核酸,所述核酸包含编码如上定义的mtmr2-s多肽或其生物学功能片段或变体的序列。在一个实施方式中,该核酸包括包含seqidno:2中至少一者的序列。在其它实施方式中,该核酸包含编码如上定义的mtmr2-s多肽或其生物学功能片段或变体的mrna序列。在具体实施方式中,该核酸包括包含seqidno:3、seqidno:4或seqidno:5中的至少一者的mrna序列,所述seqidno:3、seqidno:4或seqidno:5为编码mtmr2-s蛋白的3种亚型rna。如先前所述,该核酸编码如所定义的mtmr2多肽的所述短亚型,但不编码天然存在的人mtmr2多肽。可以使用本领域已知的多种重组方法中的任何方法来获得编码如上定义的mtmr2-s多肽或其生物学功能片段或变体的分离的核酸序列,例如,使用标准技术(例如pcr),通过从表达mtmr2基因的细胞中筛选文库、通过从已知包含该基因的载体中衍生该基因或通过直接从包含该基因的细胞和组织中分离。或者,感兴趣的基因可以通过合成的方式产生,而不是克隆。
本发明还包括其中插入了本发明的分离的核酸的载体。本领域有充足的可用于本发明的合适载体。它还指包含编码如上定义的mtmr2-s多肽的核酸序列的重组宿主细胞或核酸构建体;所述核酸序列可操作地连接至指导所述多肽产生的一个或多个调控序列。
总而言之,编码mtmr2-s的天然或合成核酸的表达通常通过将编码mtmr2-s或其部分的核酸可操作地连接至启动子、并将构建体并入表达载体中来实现。待使用的载体适于在真核细胞中复制并任选地适于在真核细胞中整合。典型的载体包含用于调控所需核酸序列表达的启动子、转录和翻译终止子以及起始序列。
使用标准的基因递送方案,本发明的载体也可用于基因疗法。基因递送方法是本领域已知的。参见例如美国专利no.5,399,346;5,580,859;或5,589,466。在另一实施方式中,本发明提供了基因疗法载体。
可以将本发明的分离的核酸克隆到多种类型的载体中。例如,可以将核酸克隆至载体中,所述载体包括但不限于质粒、噬菌粒、噬菌体衍生物、动物病毒和粘粒。特别感兴趣的载体包括表达载体、复制载体、探针生成载体(probegenerationvector)和测序载体。
此外,可以将载体以病毒载体的形式提供给细胞。病毒载体技术是本领域公知的,并在例如sambrook等(2001,molecularcloning:alaboratorymanual,冷泉港实验室,纽约)中以及其它病毒学和分子生物学手册中描述。可用作载体的病毒包括但不限于逆转录病毒、腺病毒、腺相关病毒、疱疹病毒和慢病毒。通常,合适的载体包含在至少一种有机体中起作用的复制起点、启动子序列、方便的限制性核酸内切酶位点和一种或多种选择标记(例如wo01/96584;wo01/29058;以及美国专利no.6,326,193)。
已经开发了多种基于病毒的系统,用于将基因转移至哺乳动物细胞中。例如,逆转录病毒为基因递送系统提供了便利的平台。可以使用本领域已知的技术将选定的基因插入载体并包装在逆转录病毒颗粒中。然后可以分离重组病毒并在体内或离体将其递送至受试者的细胞。多种逆转录病毒系统是本领域已知的。在一些实施方式中,使用腺病毒载体。多种腺病毒载体是本领域已知的。在一个实施方式中,使用慢病毒载体。
例如,由逆转录病毒(例如慢病毒)衍生而来的载体是实现长期基因转移的合适工具,因为它们能够使转基因长期稳定地整合并在子代细胞中传播。在优选实施方式中,组合物包含由腺相关病毒(aav)衍生而来的载体。腺相关病毒(aav)载体已成为治疗多种紊乱的强大基因递送工具。aav载体具有使其理想地适合基因疗法的多种特性,包括缺乏致病性、最小免疫原性以及以稳定有效的方式转导有丝分裂后细胞的能力。通过选择aav血清型、启动子和递送方法的适当组合,可以将aav载体中包含的特定基因的表达特异性地靶向至一种或多种类型的细胞。
在一个实施方式中,mtmr2-s编码序列包含在aav载体中。有超过30种天然存在的aav血清型可用。aav衣壳中存在许多天然变体,使得能够鉴别和使用具有特别适合骨骼肌的特性的aav。可以使用常规的分子生物学技术对aav病毒进行工程化,使得能够将这些颗粒优化,用于肌微管素核酸序列的细胞特异性递送、用于最小化免疫原性、用于调节稳定性和颗粒寿命、用于有效降解、用于准确递送至核等。
在从人或非人灵长类动物(nhp)分离并经良好表征的aav血清型中,人血清型2是第一个作为基因转移载体开发的aav;它已被广泛用于在不同靶组织和动物模型中进行有效的基因转移实验。基于aav2的载体在某些人类疾病模型中的实验性应用的临床试验正在进行中。其它有用的aav血清型包括aav1、aav3、aav4、aav5、aav6、aav7、aav8、aav9和aav10。
在一个实施方式中,在本文所述的组合物和方法中有用的载体至少包含编码选定的aav血清型衣壳(例如aav8衣壳)或其片段的序列。在另一实施方式中,有用的载体至少包含编码选定的aav血清型rep蛋白(例如aav8rep蛋白)或其片段的序列。任选地,此类载体可以包含aavcap蛋白和rep蛋白二者。
本发明的aav载体还包含小基因(minigene),其包含产生如上所述的mtmr2-s多肽的mtmr2-s核酸序列,侧翼为aav5'(反向末端重复)itr和aav3'itr。通过培养宿主细胞产生合适的重组腺相关病毒(aav),所述宿主细胞包含:编码如本文定义的腺相关病毒(aav)血清型衣壳蛋白或其片段的核酸序列;功能性rep基因;至少由aav反向末端重复(itr)和mtmr2-s核酸序列或其生物学功能片段组成的小基因;以及足够使小基因能够包装至aav衣壳蛋白中的辅助功能。可以将需在宿主细胞中培养以将aav小基因包装在aav衣壳中的组分以反式(intrans)提供给宿主细胞。或者,可以由稳定的宿主细胞提供任一种或多种所需组分(例如小基因、rep序列、cap序列和/或辅助功能),所述稳定的宿主细胞已使用本领域技术人员已知的方法工程化为包含一种或多种所需组分。
在具体实施方式中,此类稳定的宿主细胞将包含处于组成型启动子控制下的所需组分。在其它实施方式中,所需组分可以处于诱导型启动子控制下。合适的诱导型和组成型启动子的实例在本文其它地方提供,并且是本领域所公知的。在又一替代方式中,选定的稳定宿主细胞可以包含处于组成型启动子控制下的选定组分以及处于一个或多个诱导型启动子控制下的其它选定组分。例如,可以产生稳定的宿主细胞,其由293细胞衍生而来(其包含处于组成型启动子控制下的e1辅助功能),但包含处于诱导型启动子控制下的rep蛋白和/或cap蛋白。本领域技术人员还可以产生其它稳定的宿主细胞。
产生本发明的raav所需的小基因、rep序列、cap序列和辅助功能可以通过转移其上携带的序列的任何遗传元件的形式递送至包装宿主细胞。可以使用任何合适的方法来递送选定的遗传元件,包括本文所述的方法以及本领域中可获得的任何其它方法。用来构建本发明任何实施方式的方法是核酸操纵领域技术人员已知的,包括基因工程、重组工程和合成技术。类似地,生成raav病毒粒子的方法是公知的,并且对合适方法的选择不是对本发明的限制。
除非另有说明,否则本文所述的aavitr和其它选定的aav组分可以容易地选自任何aav血清型,包括但不限于aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9和aav10或者其它已知或尚未知的aav血清型。这些itr或其它aav组分可以使用本领域技术人员可利用的技术容易地从aav血清型中分离。可以从学术、商业或公共来源(例如美国模式培养物保藏中心,manassas,va.)分离或获得这种aav。或者,aav序列可以参考已公开的序列(例如文献中可得的公开序列或诸如genbank、pubmed等数据库中的公开序列)通过合成方式或其它合适的方式获得。
小基因至少由mtmr2-s编码核酸序列(转基因)及其调控序列以及5'和3'aav反向末端重复(itr)组成。在一个实施方式中,使用aav血清型2的itr。但是,也可以选择来自其它合适血清型的itr。正是这个小基因被包装到衣壳蛋白中并递送至选定的宿主细胞。编码mtmr2-s的核酸编码序列以允许转基因在宿主细胞中转录、翻译和/或表达的方式可操作地连接至调控组分。
除了上面确定的小基因的主要元件外,aav载体通常还包含常规调控元件,所述调控元件以允许转基因在本发明产生的被质粒载体转染或被病毒感染的细胞中转录、翻译和/或表达的方式可操作地连接至该转基因。本文所使用的“可操作地连接”的序列包括与感兴趣的基因相邻的表达调控序列和以反式或在一段距离处起作用以控制感兴趣的基因的表达调控序列这二者。表达调控序列包括适当的转录起始、终止、启动子和增强子序列;有效的rna加工信号(例如剪接和多聚腺苷酸化(polya)信号);稳定细胞质mrna的序列;增强翻译效率的序列(即kozak共有序列);增强蛋白稳定性的序列;以及当需要时增强编码产物分泌的序列。许多表达调控序列(包括天然、组成型、诱导型和/或组织特异性启动子)是本领域已知的并且可以被利用。其它启动子元件(例如增强子)调节转录起始的频率。通常,它们位于起始位点上游30-110bp的区域,不过最近已显示许多启动子也包含起始位点下游的功能元件。启动子元件之间的间隔通常是灵活的,从而当元件相对于彼此反转或移动时,启动子功能得以保留。取决于启动子,单个元件似乎可以协同地起作用或独立地起作用以激活转录。
为了评价mtmr2-s的表达,待导入细胞中的表达载体还可以包含选择标记基因或报告基因或这两者,以促进从试图通过病毒载体进行转染或感染的细胞群中鉴别和选择表达细胞。在其它方面,选择标记可以携带在单独的dna片段上并用于共转染程序。选择标记和报告基因均可以具有适当的调控序列作为侧翼,以使得能够在宿主细胞中表达。有用的选择标记包括例如抗生素抗性基因(例如neo等)。
报告基因用于鉴别潜在转染的细胞以及用于评价调控序列的功能性。通常,报告基因为这样的基因:该基因在受体生物体或组织中不存在或不表达并且编码多肽,所述多肽的表达通过一些易于检测的性质(例如酶活性)来显现。在将dna导入受体细胞后的适当时间时对报告基因的表达进行测定。合适的报告基因可以包括编码萤光素酶、β-半乳糖苷酶、氯霉素乙酰转移酶、分泌的碱性磷酸酶的基因或绿色荧光蛋白基因。合适的表达系统是公知的,并且可以使用已知技术制备或商购获得。通常,将显示最高水平的报告基因表达的具有最小5'侧翼区域的构建体鉴别为启动子。此类启动子区域可以连接至报告基因,并可用于评价试剂调节启动子驱动的转录的能力。
在一个实施方式中,该组合物包含编码mtmr2-s或其生物学功能片段的裸露的分离的核酸,其中,所述分离的核酸基本上不含促进转染的蛋白质、病毒颗粒、脂质体制剂等。使用裸露的分离的核酸结构(包括例如裸露的dna)能很好地诱导肌肉中的表达是本领域中所公知的。因此,本发明包括此类组合物用于局部递送至肌肉以及用于系统性给药的用途(wu等,2005,genether,12(6):477-486)。
将基因导入细胞并在细胞中表达的方法是本领域已知的。在表达载体的情况下,可以通过本领域的任何方法将载体容易地导入宿主细胞(例如哺乳动物、细菌、酵母或昆虫细胞)。例如,可以通过物理、化学或生物学手段将表达载体转移到宿主细胞中。
对于体内使用,可借助化学修饰(如磷酸骨架修饰(例如硫代磷酸酯键))对本发明的核苷酸进行稳定化。可以通过如下方式给予本发明的核苷酸:以游离(裸露)形式;或通过使用增强稳定性和/或靶向性的递送系统(例如脂质体);或纳入至其它载体(如水凝胶、环糊精、可生物降解的纳米胶囊、生物粘附性微球或蛋白性载体);或与阳离子肽组合。也可将其偶联至仿生细胞穿透肽。也可以通过其前体或编码dna的形式给药。
核苷酸的化学稳定化版本还包括“吗啉代(morpholinos)”(磷酰二胺吗啉代寡聚物-pmo)、2'-o-甲基寡聚物、带achn-(rxrrbr)2xb肽标签的pmo(r,精氨酸;x,6-氨基己酸;b,(r)-丙氨酸)(ppmo)、三环dna或小核(sn)rna。所有这些技术都是本领域公知的。这些核酸的版本还可以用于外显子跳跃,以促进内源mtmr2-s的表达。
在使用非病毒递送系统的情况下,示例性递送载体为脂质体。考虑了将脂质制剂用于将核酸导入宿主细胞(体外、离体或体内)。在另一方面,核酸可以与脂质缔合。可以将与脂质缔合的核酸封装在脂质体的水性内部中、散布在脂质体的脂质双层中、通过与脂质体和寡核苷酸均缔合的连接分子与脂质体连接、包埋在脂质体中、与脂质体复合、分散在含有脂质的溶液中、与脂质混合、与脂质结合、作为悬液包含在脂质中、包含在胶束中或与胶束复合、或以其它方式与脂质缔合。与脂质、脂质/dna或脂质/表达载体相关的组合物不限于溶液中的任何特定结构。
不管用于将外源核酸导入宿主细胞或以其它方式使细胞暴露于本发明的mtmr2-s的方法如何,为了确认宿主细胞中重组dna序列的存在,可以进行多种测定。此类测定包括例如本领域技术人员所公知的“分子生物学”测定,例如southern和northern印迹、rt-pcr和pcr;“生化”测定,例如检测特定肽存在或不存在,例如通过免疫学手段(elisa和western印迹)或通过本文所述的测定,来鉴别落入本发明范围内的试剂。
根据本发明,也可将基因组编辑用作工具。基因组编辑是一类基因工程,其中,使用人工工程化核酸酶或“分子剪刀”将dna插入基因组、从基因组中替换或从基因组中移除。核酸酶在基因组中的期望位置处制造特定的双链断裂(dsb),并利用细胞的内源机制借助同源重组(hr)和非同源末端连接(nhej)的天然过程来修复所诱导的断裂。目前使用的是四个家族的工程化核酸酶:锌指核酸酶(zfn)、转录激活因子样效应因子核酸酶(talen)、crispr/cas系统(更具体而言,如由p.mali等在naturemethods,第10卷第10期,2013年10月中描述的cas9系统)或工程化大范围核酸酶(meganuclease)再工程化的归巢(homing)核酸内切酶。可将所述核酸酶作为dna或mrna递送至细胞,将此类dna或mrna工程化以产生根据本发明的mtmr2多肽。
可以将根据本发明所使用的如上定义的核苷酸以dna前体或编码它们的分子的形式进行给药。
可以使用本领域已知的技术(例如常规固相化学)化学合成如上定义的mtmr2-s多肽,包括其片段或变体。可以产生(例如通过化学合成)片段或变体并对其进行测试以鉴别能够与天然mtm1蛋白同样好地或基本相似地发挥功能的片段或变体,例如通过测试它们的如下能力:切割或水解内源性磷酸肌醇底物或合成磷酸肌醇底物(即磷酸肌醇磷酸酶活性);募集和/或与其它蛋白(例如结蛋白(desmin)、pi3-激酶hvps34或hvps15)缔合(即适当的定位);或治疗中央核性肌病或者治疗与mtm1突变或缺乏相关的疾病或紊乱。
在某些实施方式中,本发明考虑出于诸如增强治疗或预防功效或稳定性(例如离体保存期限和对体内蛋白水解性降解的耐受力)的目的而修饰mtmr2-s多肽的结构。此类修饰mtmr2-s多肽具有与天然存在的(即天然或野生型)mtmr2-s多肽相同或基本相同的生物活性。修饰多肽可以通过例如在一个或多个位置处的氨基酸置换、删除或添加来产生。例如,可以合理地预期,例如,用异亮氨酸或缬氨酸单独置换亮氨酸、用谷氨酸单独置换天冬氨酸或用结构相关氨基酸对氨基酸进行类似置换(例如保守突变)将不会对所得分子的生物学活性产生大影响。保守置换是发生在其侧链具有相关性的氨基酸家族中的置换。
在具体实施方式中,根据本发明的待给予的治疗有效量是足以减轻与mtm1突变或改变相关的疾病或紊乱(包括中央核性肌病)的至少一种或全部迹象或者足以改善肌肉功能的量。可通过本领域普通技术人员公知的标准程序来确定待给予的mtmr2-s的量。必须对患者的生理数据(例如年龄、大小和重量)、给药途径和待治疗的疾病加以考虑来确定合适的剂量,任选地与不表现出中央核性肌病的受试者进行比较。本领域技术人员将认识到的是,待给予的mtmr2-s多肽的量或者包含或表达产生mtmr2-s的核酸的载体的量为足以治疗与mtm1突变相关的疾病或紊乱(包括中央核性肌病)的至少一种或全部迹象或者足以改善肌肉功能的量。该量可以改变,尤其是取决于诸如以下因素而改变:选定的mtmr2-s多肽或表达mtmr2-s多肽的载体;患者的性别、年龄、体重、整体身体状况等,并可基于个案来确定。该量也可根据治疗方案的其它组成因素(例如给予其它药物等)而改变。一般来说,当治疗试剂为核酸时,合适的剂量范围为约1mg/kg至约100mg/kg、更通常为约2mg/kg/天至约10mg/kg。如果选择基于病毒的核酸递送,合适的剂量将取决于不同因素,如所用病毒、递送途径(肌肉内、静脉内、动脉内或其它),但通常可为10-9病毒颗粒/kg至10-15病毒颗粒/kg。本领域技术人员将认识到的是,此类参数通常是在临床试验过程中解决。此外,本领域技术人员将认识到的是,通过本文所述的治疗,疾病症状可能完全缓解,然而并非必须如此。即便部分的或间歇性的症状缓解对于接受者也可以大有裨益。此外,对患者的治疗可为单个事件,或在多个时机给予患者mtmr2-s或编码mtmr2-s的核酸;取决于获得的结果,所述多个时机可相隔几天、相隔几个星期、或者相隔几个月、或者甚至相隔几年。
按照本领域技术人员知晓的标准制药实践(参见例如remington:thescienceandpracticeofpharmacy(第20版),a.r.gennaro,lippincottwilliams&wilkins编,2000;以及encyclopediaofpharmaceuticaltechnology,j.swarbrick和j.c.boylan编,1988-1999,marceldekker,newyork)来配制本发明的药物组合物。
可能的药物组合物包括那些适于经口、直肠、阴道内、粘膜、局部(包括经皮、口腔和舌下)或胃肠外(包括皮下(sc)、肌肉内(im)、静脉内(iv)、动脉内、皮内、胸骨内、腹膜内注射或输注技术)给药的药物组合物。对于这些制剂,可根据本领域技术人员公知的技术使用常规赋形剂。具体而言,优选肌肉内或系统性给药(例如腹膜内给药)。为了提供局部治疗效果,优选特异性肌肉给药途径或肌肉内给药途径。
可将根据本发明的药物组合物配制为在给药后实质上立即释放活性药物,或在给药后的任意预定时间或时间段释放活性药物。
出于说明的目的而并非限制的方式给出如下实施例。
实施例
申请文件中使用的缩写:
aa:氨基酸
aav:腺相关病毒
cmt:charcot-marie-tooth
cnm:中央核性肌病
fyve:fab1-yotb-vac1-eea1
he:苏木精-伊红
ko:敲除
mtm:肌微管素
mtmr:肌微管素相关
ph-gram:普列克底物蛋白同源,葡萄糖基转移酶,rab样gtpase激活物和肌微管素
ppin:磷酸肌醇
ptdins3p:磷脂酰肌醇3-磷酸
ptdins(3,5)p2:磷脂酰肌醇3,5-二磷酸
ta:胫骨前肌
wt:野生型
材料和方法
质粒和构建体
将人mtm1(1812bp,603aa)和mtmr2-l(1932bp,643aa)orf克隆到pdonr207质粒(invitrogen,carlsbad,ca)中来产生入门(entry)克隆(分别为psf108和psf98)。pdonr207-mtmr2-s(1716bp,571aa,psf101)已通过向psf98中在mtmr2-l上进行定点突变以删除对应于前72个氨基酸的前216个核苷酸而获得。使用gateway系统(invitrogen,carlsbad,ca)将不同肌微管素构建体克隆到从用于功能分析的欧洲酿酒酵母档案馆(europeansaccharomycescerevisiaearchiveforfunctionalanalysis,euroscarf)获得的酵母目的表达载体pag424gpd-ccdb-egfp(alberti,s.,gitler,a.d.和lindquist,s.(2007)asuiteofgatewaycloningvectorsforhigh-throughputgeneticanalysisinsaccharomycescerevisiae.yeast,24,913-919)和pvv200(vanmullem,v.,wery,m.,debolle,x.和vandenhaute,j.(2003)constructionofasetofsaccharomycescerevisiaevectorsdesignedforrecombinationalcloning.yeast,20,739-746)中或克隆到paav-mcs载体(cmv启动子)中。所有构建体均通过测序进行验证。pcs211dsred-fyve质粒先前已有描述(katzmann,d.j.,stefan,c.j.,babst,m.和emr,s.d.(2003)vps27recruitsescrtmachinerytoendosomesduringmvbsorting.j.cellbiol.,162,413-423)。
抗体
所用一抗是兔多克隆抗mtm1(2827)、小鼠单克隆抗mtmr2(4g3)、小鼠单克隆抗磷酸甘油酸激酶1(pgk1,invitrogen)和小鼠单克隆抗甘油醛-3-磷酸脱氢酶(抗gapdh,chemicon,merckmillipore,darmstadt,德国)。抗mtm1和抗mtmr2抗体在institutdegénétiqueetbiologiemoléculaireetcellulaire(igbmc)的抗体设施现场制备。针对全长人mtmr2产生抗mtmr2抗体,并在本研究中使用转染的cos-7细胞进行了验证。与辣根过氧化物酶(hrp)缀合的针对小鼠和兔igg的二抗从杰克逊免疫研究实验室(jacksonimmunoresearchlaboratories,westgrove,pa)获得。
体内模型
使酿酒酵母ymr1δ(matα,ura3-52,leu2-3,112,his3-δ200,trp1-δ901,lys2-801,suc2-δ9ymr1::his3)(14)和wt(matα,his3δ1,leu2δ0,lys2δ0,ura3δ0)菌株在30℃下在丰富培养基(ypd):1%酵母提取物、2%蛋白胨、2%葡萄糖中或合成营养缺陷型(drop-out)培养基(sc):0.67%无氨基酸酵母氮源基础、2%葡萄糖和适当的氨基酸混合物中生长以确保质粒维持。未使用来自酵母系统缺失集合的by4742背景下的ymr1δ(matα,his3δ1,leu2δ0,lys2δ0,ura3δ0,ymr1::kanmx),因为它不具有由scottdemr实验室描述的ymr1δ表型(parrish,w.r.,stefan,c.j.和emr,s.d.(2004)essentialroleforthemyotubularin-relatedphosphataseymr1pandthesynaptojanin-likephosphatasessjl2pandsjl3pinregulationofphosphatidylinositol3-phosphateinyeast.mol.biol.cell,15,3567-3579.)。
在该研究中,使用野生型和mtm1ko129pas小鼠。mtm1ko小鼠的特征在于从2-3周开始的进行性肌肉萎缩和无力,并到8周时导致死亡(30)。将动物圈养在温控房间内(19-22℃),光照/黑暗周期为12:12-h。
生物信息学分析
通过挖掘基因型-组织表达(gtex,www.gtexportal.org/home/)数据库获得mtmr2mrna亚型的表达水平,该数据库是使用来自数百个供体的51种不同组织的样本和培养中的2种转化细胞类型通过系统性rna测序而建立的。然后,将该数据用于计算20种最相关组织中mtmr2mrna亚型的相对表达,并创建强调特定亚型在何种组织中表达最多/最少的热图。
使用jalview(www.ialview.org/)对mtm1、mtmr2-l和mtmr2-s的n端部分进行比对,并通过clustalx颜色编码识别比对的氨基酸。
表达分析
根据制造商说明,使用trizol试剂(invitrogen,carlsbad,ca)从7周龄野生型和mtmlko小鼠的胫骨前肌(ta)肌肉和肝脏或从xlcnm患者和对照的肌肉活检中纯化总rna。使用superscriptii逆转录酶(invitrogen)和随机六聚体从500ng总rna合成cdna。
如先前所述,使用引物对来自ta肌肉和肝脏的1/10稀释的cdna进行pcr扩增,所述引物为来自mtmr2的5′-utr的正向引物:
seqidno6:5’-agcggcctccagtttctcgcgc-3’
和来自外显子3的反向引物:
seqidno7:5’-tctctcctggaagcagggctggttcc-3’,
所述pcr扩增在72℃下(65℃作为熔解温度)进行了35个扩增循环,并在72℃下进行30分钟的最终延伸(bolino,a.,marigo,v.,ferrera,f.,loader,j.,romio,l.,leoni,a.,diduca,m.,cinti,r.,cecchi,c.,feltri,m.l.等(2002)molecularcharacterizationandexpressionanalysisofmtmr2,mouse)。将产物在2%琼脂糖凝胶上进行分析,各条带使用nucleospingel和pcr纯化试剂盒(macherey-nagel,düren,德国)进行纯化,然后使用clonejetpcr克隆试剂盒(thermofisherscientific,waltham,ma)将产物克隆到pjet2.1载体中并通过sanger进行测序。
在light-cycler480ii仪器(roche,basel,swiss)上使用53℃作为熔解温度,对来自小鼠ta肌肉或人肌肉活检的1/10稀释的cdna进行定量pcr扩增。将特异性引物组用于各个小鼠mtmr2亚型:
对于小鼠v1,
seqidno8:正向5’-gactcactgtccagtgcttc-3’以及
seqidno9:反向5’-cctccctcaggaccctca-3’,
对于小鼠v2,
seqidno10:正向5’-gactcactgtccagtgcttc-3’以及
seqidno11:反向5’-cagctgggcactccctca-3’,
对于小鼠v3,
seqidno12:正向5’-aagataaaacatctcaaaaattataattgcttc-3’以及seqidno13:反向5’-cagctgggcactccctca-3’,
对于小鼠v4,
seqidno14:正向5’-aagataaaacatctcaaaaattataattgcttc-3’以及seqidno15:反向5’-gactcactgtccagtgcttc-3’。
将扩增所有小鼠亚型的共同序列的另一引物组(seqidno16:正向5’-tcctgtgtctaatggcttgc-3’以及seqidno17:反向5’-aaccaagagggcaggatatg-3’)用于对总小鼠mtmr2进行定量。将其它特异性引物组用于各个人mtmr2亚型:
对于人v1,
seqidno18:正向5’-actccttgtccagtgcctc-3’以及
seqidno19:反向5’-gactccctcaggaccctc-3’,
对于人v2,
seqidno20:正向5’-aagataaaacatctcaaaaattataattgcctc-3’以及seqidno21:反向5’-gactccctcaggaccctc-3’,
对于人v3,
seqidno22:正向5’-aagataaaacatctcaaaaattataattgcctc-3’以及seqidno23:反向5’-gagcgagactccctcctc-3’,
对于人v4,
seqidno24:正向5’-aagataaaacatctcaaaaattataattgcctc-3’以及seqidno25:反向5’-ctggactgcatgggcctc-3’。
将扩增所有人亚型的共同序列的另一引物组(seqidno26:正向5’-tttcctgtctctaataacctgcc-3’以及seqidno27:反向5’-ccaggagggcagggtatg-3’)用于对总人mtmr2进行定量。对于所有qpcr,由于hprt基因的表达在对照和xlcnm肌肉之间没有差异,因此将hprt基因表达用作对照。
western印迹
通过tca沉淀和naoh裂解从酵母细胞(od600nm=0.5-0.9,每个构建体最少3个克隆)中提取总蛋白(45),并使用组织匀浆器(omnith,kennesaw,ga)在ripa缓冲液中通过均质化从ta肌肉(每个构建体最少10份肌肉)中提取总蛋白。通过sds-page和硝酸纤维素膜上的western印迹对蛋白质裂解物进行分析。使用一抗(抗mtm11/500、抗mtmr21/1000、抗pgk11/1000和抗gapdh1/1000)检测蛋白,然后与偶联hrp的二抗孵育并彻底洗涤。通过ecl化学发光反应试剂盒(supersignalwestpicokit,thermofisherscientific,waltham,ma)显示膜。
酵母表型分析(phenotyping)
使用liac-peg方法(46),通过含有mtm1、mtmr2-l或mtmr2-scdna的酵母表达质粒pag424gpd-ccdb-egfp(2μ,c端处有gfp标签)或pvv200(2μ,无标签)对ymr1δ酵母细胞进行转化。使用通过空质粒进行转化的酵母细胞作为对照。
对于液泡染色,通过500×g离心1分钟收获1od600nm单位的细胞,并在含2μlfm4-64(200μm,invitrogen)的50μlypd培养基中在30℃下孵育15分钟,然后用900μlypd洗涤,接着在30℃下孵育10分钟,随后在sc完全培养基中进行第二次洗涤,通过荧光显微术观察染色的活酵母细胞。每个克隆计数了100至600个细胞(每个构建体3个不同克隆)并将其分为两类:大或中等单叶(unilobar)液泡以及小或碎片状液泡。
对于ptdins3p定量,通过pvv200质粒(空质粒或包含mtm1、mtmr2-l或mtmr2-scdna)和表达针对富含ptdins3p的膜结构的dsred-fyve报告基因的pcs211质粒对酵母细胞进行共转化(katzmann,d.j.,stefan,c.j.,babst,m.和emr,s.d.(2003)vps27recruitsescrtmachinerytoendosomesduringmvbsorting.j.cellbiol.,162,413-423)。在荧光显微术后,在每个克隆至少100个细胞上(每个构建体2个不同克隆)对每个细胞的点的数量进行定量。
对于ptdins5p定量,使产生不同mtm1和mtmr2构建体的酵母ymr1δ细胞生长到指数期。如hama等所述地对200od600nm单位的细胞进行脂质提取(hama,h.,takemoto,j.y.和dewald,d.b.(2000)analysisofphosphoinositidesinproteintrafficking.methods,20,465-473.)。如morrisj.b.等所述地确定ptdins5p细胞内水平。如morris等所述地进行ptdins(5)p水平的定量(morris,j.b.,hinchliffe,k.a.,ciruela,a.,letcher,a.j.和irvine,r.f.(2000)thrombinstimulationofplateletscausesanincreaseinphosphatidylinositol5-phosphaterevealedbymassassay.febslett.,475,57-60.),并基于总脂质浓度对结果进行归一化。
所有荧光显微术观察均在荧光axioobserverd1显微镜(zeiss)上使用gfp或dsred滤光片和dic光学元件借助100×/1.45油物镜(zeiss)完成。利用coolsnaphq2photometrix相机(roperscientific)捕获图像,并通过imagej进行处理(rasbandw.s.,imagej,美国国立卫生研究院,bethesda,maryland,usa,http://imagej.nih.gov/ij/)。
在肌肉提取物中通过elisa进行的ptdins3p定量
根据制造商的建议并使用ptdins3pmasselisa试剂盒(echelonbiosciences,saltlakecity,ut),对来自整个胫骨前肌(ta)肌肉的制品的脂质提取物进行ptdins3pmasselisa。对来自7周龄野生型或mtm1ko小鼠的ta肌肉进行称重,在液氮下使用研钵和研棒将其研磨成粉末,然后在冰冷的5%tca中孵育以提取脂质。将提取的脂质重悬于含3%蛋白稳定剂的pbs-t中,然后以2个重复点样在ptdins3pmasselisa板上。通过在读板仪上测量450nm处的吸光度来检测ptdins3p水平。通过将值与使用已知量的ptdins3p生成的标准曲线进行比较来确定具体量。
aav产生
通过利用paav2-插入物、pxr1和phelper对aav-293细胞系进行三重转染而生成raav2/1载体;paav2-插入物包含处于cmv启动子控制下的侧翼为血清型2反向末端重复的插入物;pxr1包含aav血清型1的rep和cap基因;phelper编码腺病毒辅助功能。将病毒载体纯化并借助实时pcr使用质粒标准paav-egfp进行定量。将滴度表示为每ml的病毒基因组(vg/ml),本文使用的raav滴度为5×1011-7×1011vg/ml。
野生型和mtm1ko小鼠胫骨前肌的aav转导
通过腹膜内注射5ml/g氯胺酮(20mg/ml;virbac,carros,法国)和甲苯噻嗪(0.4%,rompun;bayer,wuppertal,德国)麻醉2-3周龄的野生型或mtm1ko雄性129pas小鼠。向胫骨前肌(ta)肌肉注射20mlaav2/1制剂或无菌aav2/1空载体。四周后,麻醉小鼠,对ta肌肉进行功能分析(如下文所述);或将其直接解剖并冷冻在氮冷却的异戊烷中用于组织学检查;或将其固定用于电子显微术(如下文所述)。
野生型和mtm1ko小鼠的aav转导
对于系统性注射,在出生时或第1天对野生型或mtm1ko幼仔腹膜内注射1.5×1012单位的空aav病毒颗粒或者过表达人mtm1或mtmr2-s的aav。然后在注射后3周,每周通过两种不同的测试对体重和小鼠骨骼肌强度进行分析:抓握测试和悬挂测试(如下文所述)。
肌肉的功能分析
如先前所述(cowling,b.s.,chevremont,t.,prokic,i.,kretz,c.,ferry,a.,coirault,c.,koutsopoulos,o.,laugel,v.,romero,n.b.和laporte,j.(2014)reducingdynamin2expressionrescuesx-linkedcentronuclearmyopathy.jclininvest,124,1350-1363),通过测量响应于神经和肌肉刺激的原位肌肉收缩来对肌肉力量测量进行评价。通过腹膜内注射戊巴比妥钠(50mg/kg)麻醉动物。分离ta的远端肌腱(distaltendon),并用结扎丝线绑在等距传感器上(harvardbioscience,holliston,ma)。远端刺激坐骨神经,记录对强直刺激的响应(脉冲频率为50至143hz),并确定绝对最大力。收缩测量后,通过颈脱位法处死动物。为了确定比最大力,解剖ta肌肉并称重。
组织学
对于肌肉内注射,用苏木精和伊红(he)或琥珀酸脱氢酶(sdh)将小鼠ta骨骼肌的横向冰冻切片(9μm)染色,并用nanozoomer2.0ht玻片扫描仪(hamamatsu,hamamatsu市,日本)进行观察。使用imagej图像分析软件的roimanager插件在he切片上分析纤维面积。使用imagej图像分析软件的细胞计数器插件计算外周核的百分比。使用imagej插件将核定位与纤维尺寸相关联,并根据纤维尺寸对肌纤维进行颜色编码。
对于系统性注射,由石蜡包埋的器官制备5μm切片,将其固定并通过苏木精和伊红(h&e)进行染色。切片用nanozoomer2.0ht玻片扫描仪(hamamatsu)进行成像。
电子显微术
按照所描述的(cowling,b.s.,toussaint,a.,amoasii,l.,koebel,p.,ferry,a.,davignon,l.,nishino,i.,mandel,j.l.和laporte,j.(2011)increasedexpressionofwild-typeoracentronuclearmyopathymutantofdynamin2inskeletalmuscleofadultmiceleadstostructuraldefectsandmuscleweakness.am.j.pathol.,178,2224-2235),将麻醉小鼠的ta肌肉用0.1m磷酸盐缓冲液(ph7.2)中的4%pfa和2.5%戊二醛固定,并进行处理。通过将鉴别的三联体结构的数量除以切片上存在的肌节的总数来计算三联体/肌节的比例(每种基因型2只小鼠,每只小鼠分析最少10份纤维,每份纤维最少20个三联体)。
统计学分析
如图例中所注的,数据为平均值±s.e.m.或±sd。除表达分析(图6b-图6c)外(其中进行的是非配对双尾学生t检验),对于所有数据,使用单因素anova并继以tukey多重比较检验来进行统计学分析。将p值小于0.05认为具有显著性。
结果
mtmr2剪接变体差异表达并编码长蛋白亚型和短蛋白亚型
mtmr2基因突变导致charcot-marie-tooth神经病(cmt4b1),而mtm1突变则导致x连锁型中央核性肌病(xlcnm),表明这两种普遍表达的肌微管素具有不同功能。大多数组织包含多于一种亚型,因此它们的定位和表达程度可以帮助解释它们的不同功能。为了研究mtmr2功能,首先界定了其组织表达和亚型。在小鼠中,先前已在外周神经中报道了四种mtmr2mrna亚型(v1至v4),可能编码2种蛋白亚型(图7a-图7b)。变体v2至v4与变体v1的不同之处在于它们包含可变外显子1a和/或2a,导致提前终止密码子并暴露外显子3中的可变起始位点。变体v1编码643个氨基酸的蛋白,可以称为mtmr2-l(长);其它亚型则编码571aa蛋白,称为mtmr2-s(短),该蛋白先前已在多种细胞系中检测到(bolino,a.,marigo,v.,ferrera,f.,loader,j.,romio,l.,leoni,a.,diduca,m.,cinti,r.,cecchi,c.,feltri,m.l.等(2002)molecularcharacterizationandexpressionanalysisofmtmr2,mousehomologueofmtmr2,themyotubularin-related2gene,mutatedincmt4b.gene,283,17-26)。两种蛋白亚型仅在翻译起始位点不同;mtmr2-s紧邻ph-gram结构域之前开始,而mtmr2-l具有延伸的n端序列,其与任何蛋白结构域都不具有已知同源性并且在晶体结构中不可见(图1c;图7b)(begley,m.j.,taylor,g.s.,brock,m.a.,ghosh,p.,woods,v.l.和dixon,j.e.(2006)molecularbasisforsubstraterecognitionbymtmr2,amyotubularinfamilyphosphoinositidephosphatase.proc.natl.acad.sci.u.s.a.,103,927-932.;begley,m.j.,taylor,g.s.,kim,s.a.,veine,d.m.,dixon,j.e.和stuckey,j.a.(2003)crystalstructureofaphosphoinositidephosphatase,mtmr2:insightsintomyotubularmyopathyandcharcot-marie-toothsyndrome.mol.cell,12,1391-1402)。这些亚型的表达水平首先通过挖掘包含51种人组织的数据的gtex表达数据库(gtex_consortium.(2015)humangenomics.thegenotype-tissueexpression(gtex)pilotanalysis:multitissuegeneregulationinhumans.science,348,648-660)在人中进行研究。变体v1是脑、肝和脾中的主要mtmr2rna,而变体v2在其它组织中占主导。不同的变体在骨骼肌中仅较差地表达(图1a)。在小鼠中,rt-pcr和sanger测序证实了野生型(wt)和mtm1ko小鼠的胫骨前肌(ta)骨骼肌中以及肝中存在四种mtmr2mrna变体(v1至v4)(图7c-图1d),表明骨骼肌中存在mtmr2-l和mtmr2-s蛋白二者。
短mtmr2亚型而非长mtmr2亚型在酵母细胞中展现mtm1样活性
为了比较mtm1、mtmr2-l和mtmr2-s蛋白在体内的细胞功能,使用了这些人肌微管素在酵母细胞中的异源表达。酵母是研究磷酸肌醇依赖性膜运输的良好模型,因为磷酸肌醇依赖性膜运输从酵母到高等真核生物均为保守的(katzmann,d.j.,stefan,c.j.,babst,m.和emr,s.d.(2003)vps27recruitsescrtmachinerytoendosomesduringmvbsorting.j.cellbiol.,162,413-423)。在酵母细胞中,液泡体积、形态、酸度和膜电位受ptdins(3,5)p2控制,ptdins(3,5)p2通过fab1/pikfyve激酶使ptdins3p磷酸化而产生。在fab1δ突变细胞中,由于ptdins(3,5)p2的水平低,液泡相当大且是单叶的。相反,由于过量的ptdins(3,5)p2和/或ptdins3p,缺乏独特的酵母肌微管素的ymr1δ细胞具有碎片状液泡(14),并且该表型由诱导大液泡表型的人mtm1的表达补充(amoasii,l.,bertazzi,d.l.,tronchere,h.,hnia,k.,chicanne,g.,rinaldi,b.,cowling,b.s.,ferry,a.,klaholz,b.,payrastre,b.等(2012)plosgenet,8,e1002965)。为了确定mtm1、mtmr2-l和mtmr2-s的胞内定位,在ymr1δ细胞中过表达带gfp标签的融合物。mtm1-gfp和mtmr2-s-gfp蛋白被集中至与液泡相邻的膜点状结构(对fm4-64脂质染料也是阳性的),而mtmr2-l-gfp主要在细胞质中(图2c)。通过用亲脂性染料fm4-64将液泡膜染色,在ymr1δ细胞中对过表达带gfp标签或无标签的人肌微管素后的液泡形态进行了评价(图2b-图2c)。为了检测mtmr2亚型,针对重组全长人mtmr2-l生成小鼠单克隆抗体。该抗体在转化的酵母蛋白提取物中进行了验证,并特异性识别mtmr2-l和mtmr2-s(图2a)。在ymr1δ细胞中表达mtm1或mtmr2-s后,液泡显著增大;而对于mtmr2-l,液膜保持碎片状。由于这些磷酸酶的高表达水平(过表达质粒),mtm1和mtmr2-s诱导了模仿fab1δ表型的大液泡形态。这些结果表明,只有膜定位的肌微管素构建体挽救了ymr1δ细胞的液泡形态缺陷。由于液泡形态反映ptdins(3,5)p2水平,并且由于ptdins(3,5)p2丰度不足以在正常生长条件下检测到(dove,s.k.,cooke,f.t.,douglas,m.r.,sayers,l.g.,parker,p.j.和michell,r.h.(1997)osmoticstressactivatesphosphatidylinositol-3,5-bisphosphatesynthesis.nature,390,187-192),通过对ptdins5p(其是通过肌微管素磷酸酶活性由ptdins(3,5)p2产生的脂质)的水平进行质量测定对其进行定量(图2f)。在ymr1δ细胞中,mtm1和mtmr2-s过表达增加ptdins5p水平,而mtmr2-l没有作用。还通过对dsred-fyve(富含ptdins3p的膜的报告基因)阳性的点状结构进行计数对ptdins3p肌微管素底物水平进行了定量(katzmann,d.j.,stefan,c.j.,babst,m.和emr,s.d.(2003)vps27recruitsescrtmachinerytoendosomesduringmvbsorting.j.cellbiol.,162,413-423)(图2d-图2e)。mtm1和mtmr2-s的过表达显著降低ptdins3p水平,而mtmr2-l没有作用。然而,先前的数据显示mtmr2-l在体外具有强磷酸酶活性(berger,p.,bonneick,s.,willi,s.,wymann,m.和suter,u.(2002)lossofphosphataseactivityinmyotubularin-relatedprotein2isassociatedwithcharcot-marie-toothdiseasetype4b1.hum.mol.genet.,11,1569-1579),表明该亚型在酵母细胞中的胞质定位不允许ppin底物去磷酸化。总之,在酵母细胞中,只有mtmr2-s具有与mtm1相似的磷酸酶活性和定位,而mtmr2-l的表现不同。
类似于mtm1表达,mtm1ko小鼠中mtmr2短亚型的外源表达挽救肌肉重量和力量。
为了评估是否在哺乳动物中与mtmr2-l相比mtmr2-s在功能上也更接近mtm1,在mtm1ko小鼠中过表达mtm1、mtmr2-l和mtmr2-s,并对不同的肌病样表型进行分析。在cmv启动子的控制下由腺相关病毒aav2/1表达不同的肌微管素,并将重组病毒粒子注射至2-3周龄mtm1ko小鼠的ta肌肉中。mtm1ko小鼠在2-3周开始发展出进行性肌肉萎缩和无力,并到8周时导致死亡,ta肌肉是该模型中检测到的受影响最大的肌肉(buj-bello等,2002;cowling,b.s.,chevremont,t.,prokic,i.,kretz,c.,ferry,a.,coirault,c.,koutsopoulos,o.,laugel,v.,romero,n.b.和laporte,j.(2014)jclininvest,124,1350-1363)。先前已显示aav介导的mtm1在ta肌肉中表达4周纠正mtm1ko小鼠中的肌病表型(amoasii,l.,bertazzi,d.l.,tronchere,h.,hnia,k.,chicanne,g.,rinaldi,b.,cowling,b.s.,ferry,a.,klaholz,b.,payrastre,b.等(2012)plosgenet,8,e1002965)。因此,为了确定将mtmr2-l和mtmr2-s导入mtm1ko小鼠的影响,遵循了先前描述的aav注射方案(amoasii,l.,bertazzi,d.l.,tronchere,h.,hnia,k.,chicanne,g.,rinaldi,b.,cowling,b.s.,ferry,a.,klaholz,b.,payrastre,b.等(2012)plosgenet,8,e1002965),使用mtm1作为挽救的阳性对照,空aav2/1作为对侧肌肉中的疾病对照。如抗mtm1和抗mtmr2western印迹分析所示,mtm1、mtmr2-l和mtmr2-s人肌微管素在经注射的ta中表达(图3a)。在注射空aav的肌肉中未检测到内源性mtmr2蛋白,很可能归因于内源性表达水平低(图3a)。
aav注射后四周,与wt小鼠相比,mtm1ko小鼠的ta肌肉重量减少了2.5倍,二者均注射了空aav对照。与空aav对照相比,mtm1ko小鼠中的mtm1或mtmr2-s表达显著增加了肌肉质量(1.5倍),这与mtmr2-l相反(图3b)。为了解决人mtm1或mtmr2构建体在野生型(wt)小鼠中潜在的肥厚作用,对经注射的wt小鼠的ta肌肉重量进行定量(图8)。没有发现任何肌微管素增加肌肉质量,这表明在mtm1ko小鼠中观察到的改善不是由于肥厚作用,而是归因于功能挽救。
与wt小鼠相比,mtm1ko小鼠显示出相当弱的肌肉力量,并且包括mtmr2-l在内的所有肌微管素构建体均改善了ta比肌肉力(图3c)。值得注意的是,对于mtm1和mtmr2-s观察到了类似的挽救,显著高于对于注射mtmr2-l的肌肉观察到的挽救。这些结果表明,mtmr2-l和mtmr2-s亚型均改善因mtm1丧失引起的肌肉无力,并且mtmr2-s表达诱导了类似于通过mtm1基因置换所观察到的挽救。
mtmr2亚型挽救mtm1ko小鼠的组织病理学标志。
在mtm1ko小鼠中,带有mtm1、mtmr2-l或mtmr2-s的aav2/1的ta注射增加肌肉质量(mtmr2-l除外)和力量(图3)。为了在组织学水平上对挽救进行分析,确定了纤维尺寸和核定位(图4)。he(苏木精-伊红)染色显示,与用空aav或mtmr2-l处理的mtm1ko肌肉相比,aav-mtm1和aav-mtmr2-s的纤维尺寸增加(图4a),尽管我们观察到肌肉的空间异质性,有些区域仍显示出较小的萎缩纤维。形态计量分析显示,在测试的不同肌微管素中,与mtmr2构建体和空aav相比,mtm1诱导向较大纤维直径的明显转移(图4c)。在面积大于800μm2的肌肉纤维百分比方面,在aav-mtm1(平均58.4%)和aav-mtmr2-l(平均26.2%)之间观察到相当显著的差异(p<0.0001),而在mtm1和mtmr2-s(39.8%)之间差异较不显著(p=0.033)(图4d)。由于在mtm1ko小鼠中核在肌纤维内异常定位,对核分布进行了分析。与注射空aav的对侧对照肌肉相比,向mtm1ko的ta肌肉中注射mtm1、mtmr2-s或mtmr2-l显著增加定位良好的外周核的百分比(图4e)。琥珀酸脱氢酶(sdh)染色显示mtm1ko纤维中的外周和中央处的积聚(amoasii,l.,bertazzi,d.l.,tronchere,h.,hnia,k.,chicanne,g.,rinaldi,b.,cowling,b.s.,ferry,a.,klaholz,b.,payrastre,b.等(2012)plosgenet,8,e1002965),而这在表达不同肌微管素构建体后大为改善(图4b)。这些结果表明,两种mtmr2亚型均能够改善mtm1肌病的组织病理学标志,其中mtmr2-s更为有效。
mtmr2亚型挽救mtm1ko肌肉的组织混乱(disorganization)并使ptdins3p水平正常化。
患有肌小管肌病的患者和mtm1ko小鼠在超微结构水平上显示出其肌纤维的细胞内组织混乱(buj-bello,2002;spiro,a.j.,shy,g.m.和gonatas,n.k.(1966)myotubularmyopathy.persistenceoffetalmuscleinanadolescentboy.arch.neurol.,14,1-14)。为了确定收缩装置和三联体的组织化,通过电子显微术对不同注射的ta肌肉的超微结构进行评价。如先前所发表的,在注射空aav的mtm1ko肌肉中观察到z线和线粒体错位、肌节变细以及组织良好的三联体的缺乏(amoasii,l.,bertazzi,d.l.,tronchere,h.,hnia,k.,chicanne,g.,rinaldi,b.,cowling,b.s.,ferry,a.,klaholz,b.,payrastre,b.等(2012)plosgenet,8,e1002965)(图5a)。mtm1和两种mtmr2亚型的表达改善这些不同表型,在注射mtm1、mtmr2-l或mtmr2-s的肌肉中观察到组织良好的三联体,该三联体具有与中央横小管(t小管)相关联的两个肌质网池(图5a)。此外,aav介导的mtm1、mtmr2-l和mtmr2-s的表达使每个肌节的三联体数增加到几乎回到wt水平,mtmr2-s与mtmr2-l相比具有更好的效果(图5b)。
在酵母中,仅mtmr2-s而非mtmr2-l调节ptdins3p肌微管素底物水平以及ptdins(3,5)p2水平(如通过液泡形态评估的)(图2b)。为了确定mtmr2在小鼠中的挽救能力是否与其酶活性有关,对注射avv空、mtm1、mtmr2-l和mtmr2-s的mtm1ko小鼠ta肌肉中ptdins3p的细胞内水平进行定量(图6a)。注射空aav的mtm1ko肌肉中的ptdins3p水平比wt肌肉中高2.3倍,反映了mtm1丧失对其ptdins3p脂质底物的影响。mtm1表达后,ptdins3p水平降低至野生型水平,反映了mtm1的体内磷酸酶活性。当在mtm1ko小鼠中表达时,两种mtmr2亚型均诱导ptdins3p水平降低,然而只有短mtmr2-s亚型将ptdins3p正常化至野生型水平。这些结果表明mtmr2在肌肉中显示体内酶活性。此外,mtmr2催化活性与在mtm1ko肌病小鼠中外源表达所观察到的挽救相关。
综上所述,表达mtm1或mtmr2亚型的mtm1ko小鼠中的结果表明,与注射空aav的对照肌肉相比,与肌病相关的不同表型(包括肌肉力量降低、肌纤维萎缩、核错误定位、肌节和三联体组织混乱及ptdins3p水平增加)得到改善(表1)。值得注意的是,如在酵母研究中观察到的,较短的亚型mtmr2-s比mtmr2-l提供更好的挽救,并通常可与mtm1相当。
表1:mtm1和mtmr2亚型对肌小管肌病的若干标志的挽救作用
“+、++、+++”:递增的肌微管素挽救能力,范围从“-”(无挽救)到“+++”(wt表型)
mtm1ko小鼠肌肉中mtmr2短亚型的表达减少
基于gtex表达数据库,产生这两种mtmr2蛋白亚型的不同mtmr2mrna变体(v1至v4)在不同组织中表达,骨骼肌中的表达水平低(图1)。然而,尽管它们在mtm1ko小鼠的ta肌肉中过表达时具有强挽救特性(图3-图5、图6a;表1),但mtmr2变体的内源表达不能补偿肌病患者中mtm1功能的丧失。为了帮助理解在mtmr2-l和mtmr2-s亚型之间观察到的挽救差异,我们量化了与野生型(wt)小鼠相比mtm1kota肌肉中不同mtmr2变体(v1至v4)的mrna水平(图6b)。结果显示,mtm1ko肌肉中mtmr2mrna总水平降低了2倍。这主要归因于编码mtmr2-s亚型的v2和v3转录本强烈减少,而编码mtmr2-l的v1转录物水平在mtm1ko和wt小鼠之间在统计学上没有变化。注意到在图1b中未观察到这些下降,因为它呈现了不能定量的常规rt-pcr。在xlcnm患者肌肉中观察到编码mtmr2-s亚型的v2和v3转录本的类似下调(图6c)。这些数据表明由内源性mtmr2对mtm1丧失进行的补偿的缺乏与mtmr2的低表达水平(其与骨骼肌中mtmr2-s水平降低相关)有关。或者,这可能与肌肉中mtmr2蛋白水平低有关。
讨论
此处旨在确定属于同一蛋白家族但其突变导致影响不同组织的不同疾病(分别为肌病和神经病)的mtm1和mtmr2肌微管素的功能特异性和冗余性。还测试了它们作为潜在的新治疗策略相互进行补偿的能力。使用分子研究和这些人肌微管素在酵母细胞和mtm1ko肌病小鼠的骨骼肌中的过表达,表征了两种具有不同催化活性的mtmr2亚型,该催化活性与它们访问它们的ppin底物的能力有关。此外,表明了mtmr2的过表达挽救因mtm1丧失引起的肌病,并且与mtmr2-l相比,短mtmr2-s亚型在酵母和小鼠中表现出更好的ptdins3p磷酸酶活性,这与肌微管素耗竭的ymr1δ酵母细胞和mtm1ko小鼠中更好的挽救特性相关。尽管在酵母测定中表现不佳,mtmr2-l部分地改善了mtm1ko小鼠的表型这一事实可能归因于酵母异源系统中缺乏调节蛋白。
mtmr2亚型和功能
在人和小鼠中有四种天然存在的mtmr2mrna变体,它们编码两种蛋白亚型(mtmr2-l和mtmr2-s),差异为n端的72aa延伸。在酵母和小鼠中,mtmr2-s显示出比mtmr2-l更高的体内磷酸酶活性,表明n端对mtmr2功能的调节来说是重要的。该n端延伸中丝氨酸58的磷酸化被证明对mtmr2核内体膜定位和催化功能来说是重要的。事实上,与拟磷酸化(phosphomimetic)突变体mtmr2-s58e相反,mtmr2-s58a磷酸化缺陷型突变体定位至膜结构并且对ptdins3p具有活性。此处,显示当在酵母中表达时,缺少包含s58磷酸化残基的n端序列的mtmr2-s蛋白集中到膜上(图2b),并且在酵母中(图2d)和在鼠类肌肉中(图6a)对ptdins3p的活性与mtmr2-l相比更高。mtmr2的n端延伸未在晶体结构中得到解析,支持了它能够采用不同构象并能够调节mtmr2功能的假设(begley,m.j.,taylor,g.s.,brock,m.a.,ghosh,p.,woods,v.l.和dixon,j.e.(2006)molecularbasisforsubstraterecognitionbymtmr2,amyotubularinfamilyphosphoinositidephosphatase.proc.natl.acad.sci.u.s.a.,103,927-932;begley,m.j.,taylor,g.s.,kim,s.a.,veine,d.m.,dixon,j.e.和stuckey,j.a.(2003)crystalstructureofaphosphoinositidephosphatase,mtmr2:insightsintomyotubularmyopathyandcharcot-marie-toothsyndrome.mol.cell,12,1391-1402)。这些结果表明存在两种形式的mtmr2:mtmr2-s主要定位在膜并且具有高体内磷酸酶活性;mtmr2-l的膜定位取决于s58残基处的磷酸化。有趣的是,脑中表达偏向于(biasedtowards)编码mtmr2-l的mtmr2v1变体(图1)。s58磷酸化由erk2激酶介导,而erk2激酶在脑中的表达正好高于其它组织,与mtmr2-l的表达相关(gtex数据库)。
肌微管素中的功能冗余和补偿
存在大多数在人组织中普遍表达的14种肌微管素,但是mtm1丧失特别导致严重的先天性肌病。这表明当以内源水平表达时,mtm1同源物(尤其是较接近的mtmr2同源物)不能补偿骨骼肌中mtm1的缺乏。此处证明了mtmr2-s在肌病mtm1ko小鼠的骨骼肌中下调。此外,与大脑和其它组织相比,骨骼肌中mtmr2转录本的表达低。总之,这表明由于mtmr2在肌病小鼠模型和xlcnm患者肌肉中的下调而加剧的这种mtmr2在肌肉中的低表达是缺乏补偿的基础。实际上,mtmr2-s在肌病肌肉中比mtmr2-l更好地改善了功能性和结构性肌病表型二者,并且更显著地下调。这表明,mtm1和mtmr2之间的功能差异的分子基础在于ph-gram结构域上游的n端延伸,缺少该延伸的mtmr2-s在酵母和小鼠中表现出与mtm1类似的体内功能。在天然mtmr2-l亚型中移除该n端延伸将mtmr2活性转化为类似于mtm1的活性。
mtmr2-s作为肌小管肌病的新治疗靶标
mtmr2-s因此可以用作治疗靶标。将人mtmr2-s肌肉内aav转导至mtm1ko小鼠中大大改善了表型,支持了“挽救在肌肉中是细胞自主的”。虽然此实际方案旨在通过肌肉内注射研究由mtmr2进行的细胞自主补偿,但由于mtm1ko小鼠在2个月左右时死亡(最可能由于呼吸衰竭和进食缺陷),因此无法确定挽救的程度和mtmr2介导的挽救的长期潜力。这些数据支持了“mtmr2-s亚型比主要描述的mtmr2-l亚型具有更好的挽救能力并且是天然存在的变体,包括在肌肉中”。由于mtm1ko肌肉中mtmr2-s转录本减少,因此,潜在策略将是通过调节mtmr2可变剪接或外源表达来促进其表达。
通过系统性注射的mtmr2-s表达对小鼠整体的影响
如上文所示,肌肉内注射使得能够研究mtm1ko小鼠模型中mtm1和mtmr2的肌肉特异性功能和挽救能力。为了完成这项研究并观察mtmr2表达对小鼠整体的影响,进行了系统性注射。
在出生时或第1天对野生型或mtm1ko幼仔腹膜内注射1.5×1012单位的空aav病毒颗粒或过表达人mtm1或mtmr2-s的aav。然后在注射后3周,通过两种不同的测试来测量体重和小鼠骨骼肌的强度:抓握测试和悬挂测试。当注射空aav的mtm1ko影响的小鼠仍然存活时,从7周龄开始处死小鼠,从而可以将肌微管素过表达与空载体进行比较。
通过在胫骨前肌和膈肌骨骼肌上进行的western印迹来对mtm1和mtmr2-s亚型的过表达进行评价。在这两种情况下,如上文肌肉内注射所观察到的,都以预期大小很好地检测到肌微管素。这证实了在小鼠出生后第1天系统性递送aav后,所有肌微管素均在骨骼肌中良好表达。
分析了肌微管素的系统性表达对受注射小鼠的寿命和体重的影响。第一个主要观察结果是mtmr2-s延长了mtm1ko小鼠的寿命(3/4存活至7-10周)。虽然肌病小鼠通常在5-7周龄左右时死亡,但mtmr2-s过表达使得两只小鼠能够达到10周龄(测量的最老时间点)。已经发表了mtm1对寿命具有类似的挽救作用。该实验证实mtmr2-s亚型可以增加mtm1敲除小鼠的寿命。
肌病mtm1ko小鼠的主要临床表型是始自2-3周龄的与wt小鼠相比的较低体重以及在5周龄左右开始的体重逐渐减轻(cowling等,2014)。后者主要归因于肌肉质量丧失以及在疾病的最终阶段中难以获取食物。相反,wt小鼠在其生命的前10-12周内持续逐渐增加体重(图9)。过表达人mtm1的小鼠最初就比注射空aav的mtm1ko小鼠大,并且以与wt小鼠相同的速率理想地增加体重。相比之下,过表达mtmr2-s的小鼠从3-5周龄也能显示出对体重的良好挽救,但随后开始体重减少并达到mtm1ko水平(图9)。相关地,直到5周龄为止,mtmr2-s表达的积极作用在临床上是明确的(但未量化),然后小鼠开始逐渐发展出mtm1ko典型表型(肌肉重量和力量丧失、脊柱侧弯、呼吸和行走困难)。
这些结果表明,mtmr2短亚型改善了mtm1ko小鼠的寿命和体重。
肌小管肌病的另一明显临床特征是严重肌肉无力,这在mtm1ko小鼠模型中重现。悬挂测试测量全身力量。mtmr2-s亚型的过表达使mtm1ko小鼠能够逐渐悬挂更久(从30-40秒开始),直到它们达到wt水平,并能够在7周后悬挂3×60秒(图10)。在相同年龄,通常有半数mtm1ko小鼠死亡,并且接受测试的两只小鼠在5周后不能悬挂超过几秒钟。mtm1的效果甚至更好,使mtm1ko小鼠在4周后可以完美地悬挂60秒。对于始终能够悬挂60秒的wt小鼠,未观察到任何肌微管素的负面作用。
这些结果表明,mtmr2-s亚型挽救了mtm1ko小鼠的肌肉力量。值得注意的是,这些外观上患病(呼吸困难、脊柱侧弯)的注射了mtmr2亚型的小鼠与wt小鼠一样能悬挂60秒,表明全身力量的强烈改善(图10)。
结论
肌肉内注射确定了短mtmr2亚型和长mtmr2亚型均改善肌病表型,其中,并行比较时,短亚型(mtmr2-s)诱导更好的挽救效果。系统性注射证实mtmr2-s亚型表达能够延迟mtm1ko小鼠中的肌病表型发作,并显著挽救其肌肉力量。过表达mtmr2亚型的小鼠仍然受到影响,但显然比mtm1ko小鼠更具运动性。总而言之,这项系统研究显示了令人满意的初步数据,支持了“mtmr2-s的过表达能够改善mtm1敲除小鼠(肌小管肌病的小鼠模型)中的肌病表型”。
mtmr2-s序列
seqidno:1(人mtmr2-s蛋白)
人mtmr2-s蛋白序列(np_001230500.1、np_958438.1和np_958435.1)
seqidno:2(核苷酸人mtmr2-s,cdna)编码序列
人mtmr2-s编码序列(ccds:ccds8306.1-nm_201278.2:664..2379)编码相同蛋白mtmr2-s的3种亚型rna:
seqidno:3:对应于人mtmr2mrnav2的cdna(nm_201278.2)
seqidno:4:对应于人mtmr2mrnav3的cdna(nm_201281.2)
seqidno5:对应于人mtmr2mrnav4的cdna(nm_001243571.1)
序列表
<110>斯特拉斯堡大学等
<120>用于治疗肌病的组合物和方法
<130>b2537pc00
<160>29
<170>patentinversion3.5
<210>1
<211>571
<212>prt
<213>智人(homosapiens)
<400>1
metglugluproproleuleuproglygluasnilelysaspmetala
151015
lysaspvalthrtyrilecysprophethrglyalavalargglythr
202530
leuthrvalthrasntyrargleutyrphelyssermetgluargasp
354045
proprophevalleuaspalaserleuglyvalileasnargvalglu
505560
lysileglyglyalaserserargglygluasnsertyrglyleuglu
65707580
thrvalcyslysaspileargasnleuargphealahislysproglu
859095
glyargthrargargserilephegluasnleumetlystyralaphe
100105110
provalserasnasnleuproleuphealapheglutyrlysgluval
115120125
pheprogluasnglytrplysleutyraspproleuleuglutyrarg
130135140
argglnglyileproasnglusertrpargilethrlysileasnglu
145150155160
argtyrgluleucysaspthrtyrproalaleuleuvalvalproala
165170175
asnileproaspglugluleulysargvalalaserpheargserarg
180185190
glyargileprovalleusertrpilehisprogluserglnalathr
195200205
ilethrargcysserglnprometvalglyvalserglylysargser
210215220
lysgluaspglulystyrleuglnalailemetaspserasnalagln
225230235240
serhislysilepheilepheaspalaargproservalasnalaval
245250255
alaasnlysalalysglyglyglytyrglusergluaspalatyrgln
260265270
asnalagluleuvalpheleuaspilehisasnilehisvalmetarg
275280285
gluserleuarglysleulysgluilevaltyrproasnilegluglu
290295300
thrhistrpleuserasnleugluserthrhistrpleugluhisile
305310315320
lysleuileleualaglyalaleuargilealaasplysvalgluser
325330335
glylysthrservalvalvalhiscysseraspglytrpaspargthr
340345350
alaglnleuthrserleualametleumetleuaspglytyrtyrarg
355360365
thrileargglyphegluvalleuvalglulysglutrpleuserphe
370375380
glyhisargpheglnleuargvalglyhisglyasplysasnhisala
385390395400
aspalaaspargserprovalpheleuglnpheileaspcysvaltrp
405410415
glnmetthrargglnpheprothralapheglupheasnglutyrphe
420425430
leuilethrileleuasphisleutyrsercysleupheglythrphe
435440445
leucysasnsergluglnglnargglylysgluasnleuprolysarg
450455460
thrvalserleutrpsertyrileasnserglnleugluaspphethr
465470475480
asnproleutyrglysertyrserasnhisvalleutyrprovalala
485490495
sermetarghisleugluleutrpvalglytyrtyrileargtrpasn
500505510
proargmetlysproglngluproilehisasnargtyrlysgluleu
515520525
leualalysargalagluleuglnlyslysvalglugluleuglnarg
530535540
gluileserasnargserthrsersersergluargalaserserpro
545550555560
alaglncysvalthrprovalglnthrvalval
565570
<210>2
<211>1716
<212>dna
<213>智人(homosapiens)
<400>2
atggaagaaccacccttgcttccaggagaaaatattaaagacatggccaaagatgtaact60
tatatatgtccattcactggcgctgtacgaggaactctgactgtcacgaattataggtta120
tatttcaaaagcatggaacgggatcccccatttgttttagatgcttcccttggtgtgata180
aatagagtagaaaaaattggtggtgcttctagtcgaggtgaaaattcttatggactagaa240
actgtgtgtaaggatattaggaatttacgatttgctcataaacctgaggggcggacaaga300
agatccatatttgagaatctaatgaaatatgcatttcctgtctctaataacctgcctctt360
tttgcttttgaatacaaagaagtattccctgaaaatgggtggaagctatatgaccctctt420
ttagagtatagaaggcagggaattccaaatgaaagctggagaataacaaagataaatgaa480
cgatatgaactttgtgatacataccctgccctcctggttgtgccagcaaatattcctgat540
gaagaattaaagagagtggcatccttcagatcaagaggccgtatcccagttttatcatgg600
attcatcctgaaagtcaagccacaatcactcggtgtagccagcccatggttggagtgagt660
ggaaagcgaagcaaagaagatgaaaaataccttcaagctatcatggattccaatgcccag720
tctcacaaaatctttatatttgatgcccggccaagtgttaatgctgttgccaacaaggca780
aagggtggaggttatgaaagtgaagatgcctatcaaaatgctgaactagttttcctggat840
atccacaatattcatgttatgagagaatcattacgaaaacttaaggagattgtgtacccc900
aacattgaggaaacccactggttgtctaacttggaatctactcattggctagaacatatt960
aagcttattcttgcaggggctcttaggattgctgacaaggtagagtcagggaagacgtct1020
gtggtagtgcattgcagtgatggttgggatcgcacagctcagctcacttcccttgccatg1080
ctcatgttggatggatactatcgaaccatccgaggatttgaagtccttgtggagaaagaa1140
tggctaagttttggacatcgatttcaactaagagttggccatggagataagaaccatgca1200
gatgcagacagatcgcctgtttttcttcaatttattgactgtgtctggcagatgacaaga1260
cagtttcctaccgcatttgaattcaatgagtattttctcattaccattttggaccaccta1320
tacagctgcttattcggaacattcctctgtaatagtgaacaacagagaggaaaagagaat1380
cttcctaaaaggactgtgtcactgtggtcttacataaacagccagctggaagacttcact1440
aatcctctctatgggagctattccaatcatgtcctttatccagtagccagcatgcgccac1500
ctagagctctgggtgggatattacataaggtggaatccacggatgaaaccacaggaacct1560
attcacaacagatacaaagaacttcttgctaaacgagcagagcttcagaaaaaagtagag1620
gaactacagagagagatttctaaccgatcaacctcatcctcagagagagccagctctcct1680
gcacagtgtgtcactcctgtccaaactgttgtataa1716
<210>3
<211>4789
<212>dna
<213>智人(homosapiens)
<400>3
agatgtcgcgcggccggacacagccagcacggagagtcgatgccggcgtctgagctgcgc60
agtggggtcttcccgctgcccagcagcctacaggcgcggtgcactctgggggaacatggc120
cgcttccggtctccctcccgggccggcgctggcctgactgcggccccggtccgtagcact180
ccgccctccgcttctcccgccctgtagccgcgaagactgcttcagcctttccctgtgctg240
cccctgccgcgcgatggagaagagctcgagctgcgagagtcttggctcccagccggcggc300
ggctcggccgcccagcgtggactccttgtccagttaatgtgttaagagccattgacattt360
gaagatcatcagaagtgaagataaaacatctcaaaaattataattgcctccacttctcat420
tcagagaattcagtgcatacaaaatcagcttctgttgtatcatcagattccatttcaact480
tctgccgacaacttttctcctgatttgaggagggagtctcgctctatcccctaggctgga540
gtgcattggcgccatctcggctcatttgcaacctctgtctcccgggttcaagcgattctc600
ctgcctcagcttcccgaggagctgggattacaggtcctgagggagtctaacaagttagca660
gaaatggaagaaccacccttgcttccaggagaaaatattaaagacatggccaaagatgta720
acttatatatgtccattcactggcgctgtacgaggaactctgactgtcacgaattatagg780
ttatatttcaaaagcatggaacgggatcccccatttgttttagatgcttcccttggtgtg840
ataaatagagtagaaaaaattggtggtgcttctagtcgaggtgaaaattcttatggacta900
gaaactgtgtgtaaggatattaggaatttacgatttgctcataaacctgaggggcggaca960
agaagatccatatttgagaatctaatgaaatatgcatttcctgtctctaataacctgcct1020
ctttttgcttttgaatacaaagaagtattccctgaaaatgggtggaagctatatgaccct1080
cttttagagtatagaaggcagggaattccaaatgaaagctggagaataacaaagataaat1140
gaacgatatgaactttgtgatacataccctgccctcctggttgtgccagcaaatattcct1200
gatgaagaattaaagagagtggcatccttcagatcaagaggccgtatcccagttttatca1260
tggattcatcctgaaagtcaagccacaatcactcggtgtagccagcccatggttggagtg1320
agtggaaagcgaagcaaagaagatgaaaaataccttcaagctatcatggattccaatgcc1380
cagtctcacaaaatctttatatttgatgcccggccaagtgttaatgctgttgccaacaag1440
gcaaagggtggaggttatgaaagtgaagatgcctatcaaaatgctgaactagttttcctg1500
gatatccacaatattcatgttatgagagaatcattacgaaaacttaaggagattgtgtac1560
cccaacattgaggaaacccactggttgtctaacttggaatctactcattggctagaacat1620
attaagcttattcttgcaggggctcttaggattgctgacaaggtagagtcagggaagacg1680
tctgtggtagtgcattgcagtgatggttgggatcgcacagctcagctcacttcccttgcc1740
atgctcatgttggatggatactatcgaaccatccgaggatttgaagtccttgtggagaaa1800
gaatggctaagttttggacatcgatttcaactaagagttggccatggagataagaaccat1860
gcagatgcagacagatcgcctgtttttcttcaatttattgactgtgtctggcagatgaca1920
agacagtttcctaccgcatttgaattcaatgagtattttctcattaccattttggaccac1980
ctatacagctgcttattcggaacattcctctgtaatagtgaacaacagagaggaaaagag2040
aatcttcctaaaaggactgtgtcactgtggtcttacataaacagccagctggaagacttc2100
actaatcctctctatgggagctattccaatcatgtcctttatccagtagccagcatgcgc2160
cacctagagctctgggtgggatattacataaggtggaatccacggatgaaaccacaggaa2220
cctattcacaacagatacaaagaacttcttgctaaacgagcagagcttcagaaaaaagta2280
gaggaactacagagagagatttctaaccgatcaacctcatcctcagagagagccagctct2340
cctgcacagtgtgtcactcctgtccaaactgttgtataaaggactgtaagatcaggggca2400
tcattgctatacactcttgattacactggcagctctatgagtagaaagtcttcggaattt2460
agaacccatctatgagagaaagttcagtcactttatttattttaaatctctctaggatga2520
gtttagaactgtagcagtgcaggtggcttaagtgaagtaactccatatgtaattacatga2580
ttatgatactaatcttttaagtatccaaagaatattaaaatacttcaatcctggattcac2640
agtgggaacaagtttctattaaaaggcaaatgctgttacaaatttttggcatctggtaat2700
attaaaaccattttagaaatacactctgtgctcactgtgcagaggaacatcagttttcaa2760
accaacactgaaattctgtggcatcacatatattgggccttgatgtcatgacagatcaaa2820
atcatttgatatccctttctccattctaggtttttctttttttcagtaactgatttacct2880
tgatcacttttcaacttccatattcttcatatagtaaaaggcaaagtgttgaagatacta2940
cggtgtggtagtagttgaaaattattgccgtcattatttacatacttaagacatattagc3000
aagttgatccaaaatgggaggccttatagatgtgcttgggggaaaatgaaggggagaaag3060
tagccatacaggagttcaaagaattccatgcccttcagattagcccaattaccagaaaca3120
tcatgaaagatattttaaaaactaattatttactacagtgtatttcacttgtcttgtgtg3180
tctgaacacacagaagctaattagcaagtttttaagaagtatttaaaaatcttactagga3240
ttgacattttttctgaattctgtataaatagcttatagtgagaagtactgtgctcaaatt3300
ttacatttttttcctttgcaaattctgtaatttcactcaacgattaagtctaccaaagaa3360
cacactgcatgtaaaagatgtattacaatctcaaagccagtaaaagaaatcttgcttcac3420
tgttcacctgctacaagtaagagtttggtgctggtagaaacatttgactctgatgtctat3480
tttattctacataagagccatatgtaatgtactgtaacaaaggagcttcttgtccccttg3540
gtcttttaattaaaagaaattccaactgacttttaaactttgttcttgtccaaagttgcc3600
atttcttttttttccccagaaatatttggaaattattggaggaatatgcaccccagatga3660
aaatgttcagtttgtacccatttttccttaaccaacacccaaatcaaacaattaaaatat3720
acagtgtttttccactcactaattcactatacagagagtctgaaccttagcctccctctt3780
ggtcttgcagtgaggaagtttctattagtatatccaatttagcaaaattggtaccaaaat3840
gatttctttggtaattgtgtgaaatataagctttttaacagggcatttaagtggctagca3900
aatcagtaattaaaaattaagctttctactccaagtatttcacaaacgcatctgccattt3960
tcctcatttaaaccttggttatcttggcctgataccacataaaagaatgtagaatggctg4020
aagagatcaagaatttaaagcttctagtcttaacatacttgcatccacttcaaattcaaa4080
tcaaaagccagggaaatctaagtgcaaccctaccacttctctgctgagaaccttccagtg4140
gttcccctcaccttctgcagaagtctccaatatggagtacatgcacttgggcatttaata4200
tataccactggtgtgtgtgggagggagggaggaggaatactagccctttttatatattta4260
cacaagcaaaacttttaaatatttgaattgacagttacatgtttcataactttgtatgtc4320
tattggttgtgcaggtgtaattttttccctttttgattagggttacaaaatttagagacc4380
agtatgattaagttgaagctccttagcctccttcgacctagtctctgcatacctcaactt4440
ttacgtaccaatgctactctgctgttcacaattgcctcatgtaatctgcagattcctgcc4500
tccccactttggttcagtctgtcttgtgcacctggaacaactgttctcccttgtgactaa4560
ttcctattttctagagtttaggcatcatctcttcctttgggaagttatctgattcacgac4620
tgccttctctgacatccccaccttcctctgtgcccccatagcactgtgtataccgctact4680
accactgcaattcacattatattggaatgaacaattcacatgtctaccacaagtctgtaa4740
acataaccttatttgaaatgaattgcaataaagctctgttacaacgtaa4789
<210>4
<211>4666
<212>dna
<213>智人(homosapiens)
<400>4
agatgtcgcgcggccggacacagccagcacggagagtcgatgccggcgtctgagctgcgc60
agtggggtcttcccgctgcccagcagcctacaggcgcggtgcactctgggggaacatggc120
cgcttccggtctccctcccgggccggcgctggcctgactgcggccccggtccgtagcact180
ccgccctccgcttctcccgccctgtagccgcgaagactgcttcagcctttccctgtgctg240
cccctgccgcgcgatggagaagagctcgagctgcgagagtcttggctcccagccggcggc300
ggctcggccgcccagcgtggactccttgtccagttaatgtgttaagagccattgacattt360
gaagatcatcagaagtgaagataaaacatctcaaaaattataattgcctccacttctcat420
tcagagaattcagtgcatacaaaatcagcttctgttgtatcatcagattccatttcaact480
tctgccgacaacttttctcctgatttgagggtcctgagggagtctaacaagttagcagaa540
atggaagaaccacccttgcttccaggagaaaatattaaagacatggccaaagatgtaact600
tatatatgtccattcactggcgctgtacgaggaactctgactgtcacgaattataggtta660
tatttcaaaagcatggaacgggatcccccatttgttttagatgcttcccttggtgtgata720
aatagagtagaaaaaattggtggtgcttctagtcgaggtgaaaattcttatggactagaa780
actgtgtgtaaggatattaggaatttacgatttgctcataaacctgaggggcggacaaga840
agatccatatttgagaatctaatgaaatatgcatttcctgtctctaataacctgcctctt900
tttgcttttgaatacaaagaagtattccctgaaaatgggtggaagctatatgaccctctt960
ttagagtatagaaggcagggaattccaaatgaaagctggagaataacaaagataaatgaa1020
cgatatgaactttgtgatacataccctgccctcctggttgtgccagcaaatattcctgat1080
gaagaattaaagagagtggcatccttcagatcaagaggccgtatcccagttttatcatgg1140
attcatcctgaaagtcaagccacaatcactcggtgtagccagcccatggttggagtgagt1200
ggaaagcgaagcaaagaagatgaaaaataccttcaagctatcatggattccaatgcccag1260
tctcacaaaatctttatatttgatgcccggccaagtgttaatgctgttgccaacaaggca1320
aagggtggaggttatgaaagtgaagatgcctatcaaaatgctgaactagttttcctggat1380
atccacaatattcatgttatgagagaatcattacgaaaacttaaggagattgtgtacccc1440
aacattgaggaaacccactggttgtctaacttggaatctactcattggctagaacatatt1500
aagcttattcttgcaggggctcttaggattgctgacaaggtagagtcagggaagacgtct1560
gtggtagtgcattgcagtgatggttgggatcgcacagctcagctcacttcccttgccatg1620
ctcatgttggatggatactatcgaaccatccgaggatttgaagtccttgtggagaaagaa1680
tggctaagttttggacatcgatttcaactaagagttggccatggagataagaaccatgca1740
gatgcagacagatcgcctgtttttcttcaatttattgactgtgtctggcagatgacaaga1800
cagtttcctaccgcatttgaattcaatgagtattttctcattaccattttggaccaccta1860
tacagctgcttattcggaacattcctctgtaatagtgaacaacagagaggaaaagagaat1920
cttcctaaaaggactgtgtcactgtggtcttacataaacagccagctggaagacttcact1980
aatcctctctatgggagctattccaatcatgtcctttatccagtagccagcatgcgccac2040
ctagagctctgggtgggatattacataaggtggaatccacggatgaaaccacaggaacct2100
attcacaacagatacaaagaacttcttgctaaacgagcagagcttcagaaaaaagtagag2160
gaactacagagagagatttctaaccgatcaacctcatcctcagagagagccagctctcct2220
gcacagtgtgtcactcctgtccaaactgttgtataaaggactgtaagatcaggggcatca2280
ttgctatacactcttgattacactggcagctctatgagtagaaagtcttcggaatttaga2340
acccatctatgagagaaagttcagtcactttatttattttaaatctctctaggatgagtt2400
tagaactgtagcagtgcaggtggcttaagtgaagtaactccatatgtaattacatgatta2460
tgatactaatcttttaagtatccaaagaatattaaaatacttcaatcctggattcacagt2520
gggaacaagtttctattaaaaggcaaatgctgttacaaatttttggcatctggtaatatt2580
aaaaccattttagaaatacactctgtgctcactgtgcagaggaacatcagttttcaaacc2640
aacactgaaattctgtggcatcacatatattgggccttgatgtcatgacagatcaaaatc2700
atttgatatccctttctccattctaggtttttctttttttcagtaactgatttaccttga2760
tcacttttcaacttccatattcttcatatagtaaaaggcaaagtgttgaagatactacgg2820
tgtggtagtagttgaaaattattgccgtcattatttacatacttaagacatattagcaag2880
ttgatccaaaatgggaggccttatagatgtgcttgggggaaaatgaaggggagaaagtag2940
ccatacaggagttcaaagaattccatgcccttcagattagcccaattaccagaaacatca3000
tgaaagatattttaaaaactaattatttactacagtgtatttcacttgtcttgtgtgtct3060
gaacacacagaagctaattagcaagtttttaagaagtatttaaaaatcttactaggattg3120
acattttttctgaattctgtataaatagcttatagtgagaagtactgtgctcaaatttta3180
catttttttcctttgcaaattctgtaatttcactcaacgattaagtctaccaaagaacac3240
actgcatgtaaaagatgtattacaatctcaaagccagtaaaagaaatcttgcttcactgt3300
tcacctgctacaagtaagagtttggtgctggtagaaacatttgactctgatgtctatttt3360
attctacataagagccatatgtaatgtactgtaacaaaggagcttcttgtccccttggtc3420
ttttaattaaaagaaattccaactgacttttaaactttgttcttgtccaaagttgccatt3480
tcttttttttccccagaaatatttggaaattattggaggaatatgcaccccagatgaaaa3540
tgttcagtttgtacccatttttccttaaccaacacccaaatcaaacaattaaaatataca3600
gtgtttttccactcactaattcactatacagagagtctgaaccttagcctccctcttggt3660
cttgcagtgaggaagtttctattagtatatccaatttagcaaaattggtaccaaaatgat3720
ttctttggtaattgtgtgaaatataagctttttaacagggcatttaagtggctagcaaat3780
cagtaattaaaaattaagctttctactccaagtatttcacaaacgcatctgccattttcc3840
tcatttaaaccttggttatcttggcctgataccacataaaagaatgtagaatggctgaag3900
agatcaagaatttaaagcttctagtcttaacatacttgcatccacttcaaattcaaatca3960
aaagccagggaaatctaagtgcaaccctaccacttctctgctgagaaccttccagtggtt4020
cccctcaccttctgcagaagtctccaatatggagtacatgcacttgggcatttaatatat4080
accactggtgtgtgtgggagggagggaggaggaatactagccctttttatatatttacac4140
aagcaaaacttttaaatatttgaattgacagttacatgtttcataactttgtatgtctat4200
tggttgtgcaggtgtaattttttccctttttgattagggttacaaaatttagagaccagt4260
atgattaagttgaagctccttagcctccttcgacctagtctctgcatacctcaactttta4320
cgtaccaatgctactctgctgttcacaattgcctcatgtaatctgcagattcctgcctcc4380
ccactttggttcagtctgtcttgtgcacctggaacaactgttctcccttgtgactaattc4440
ctattttctagagtttaggcatcatctcttcctttgggaagttatctgattcacgactgc4500
cttctctgacatccccaccttcctctgtgcccccatagcactgtgtataccgctactacc4560
actgcaattcacattatattggaatgaacaattcacatgtctaccacaagtctgtaaaca4620
taaccttatttgaaatgaattgcaataaagctctgttacaacgtaa4666
<210>5
<211>4862
<212>dna
<213>智人(homosapiens)
<400>5
agatgtcgcgcggccggacacagccagcacggagagtcgatgccggcgtctgagctgcgc60
agtggggtcttcccgctgcccagcagcctacaggcgcggtgcactctgggggaacatggc120
cgcttccggtctccctcccgggccggcgctggcctgactgcggccccggtccgtagcact180
ccgccctccgcttctcccgccctgtagccgcgaagactgcttcagcctttccctgtgctg240
cccctgccgcgcgatggagaagagctcgagctgcgagagtcttggctcccagccggcggc300
ggctcggccgcccagcgtggactccttgtccagttaatgtgttaagagccattgacattt360
gaagatcatcagaagtgaagataaaacatctcaaaaattataattgcctccacttctcat420
tcagagaattcagtgcatacaaaatcagcttctgttgtatcatcagattccatttcaact480
tctgccgacaacttttctcctgatttgaggcccatgcagtccagttcgggagctaagtca540
cttaaacactgtgtgatagttcctacacatctcattatagtttagggagtctcgctctat600
cccctaggctggagtgcattggcgccatctcggctcatttgcaacctctgtctcccgggt660
tcaagcgattctcctgcctcagcttcccgaggagctgggattacaggtcctgagggagtc720
taacaagttagcagaaatggaagaaccacccttgcttccaggagaaaatattaaagacat780
ggccaaagatgtaacttatatatgtccattcactggcgctgtacgaggaactctgactgt840
cacgaattataggttatatttcaaaagcatggaacgggatcccccatttgttttagatgc900
ttcccttggtgtgataaatagagtagaaaaaattggtggtgcttctagtcgaggtgaaaa960
ttcttatggactagaaactgtgtgtaaggatattaggaatttacgatttgctcataaacc1020
tgaggggcggacaagaagatccatatttgagaatctaatgaaatatgcatttcctgtctc1080
taataacctgcctctttttgcttttgaatacaaagaagtattccctgaaaatgggtggaa1140
gctatatgaccctcttttagagtatagaaggcagggaattccaaatgaaagctggagaat1200
aacaaagataaatgaacgatatgaactttgtgatacataccctgccctcctggttgtgcc1260
agcaaatattcctgatgaagaattaaagagagtggcatccttcagatcaagaggccgtat1320
cccagttttatcatggattcatcctgaaagtcaagccacaatcactcggtgtagccagcc1380
catggttggagtgagtggaaagcgaagcaaagaagatgaaaaataccttcaagctatcat1440
ggattccaatgcccagtctcacaaaatctttatatttgatgcccggccaagtgttaatgc1500
tgttgccaacaaggcaaagggtggaggttatgaaagtgaagatgcctatcaaaatgctga1560
actagttttcctggatatccacaatattcatgttatgagagaatcattacgaaaacttaa1620
ggagattgtgtaccccaacattgaggaaacccactggttgtctaacttggaatctactca1680
ttggctagaacatattaagcttattcttgcaggggctcttaggattgctgacaaggtaga1740
gtcagggaagacgtctgtggtagtgcattgcagtgatggttgggatcgcacagctcagct1800
cacttcccttgccatgctcatgttggatggatactatcgaaccatccgaggatttgaagt1860
ccttgtggagaaagaatggctaagttttggacatcgatttcaactaagagttggccatgg1920
agataagaaccatgcagatgcagacagatcgcctgtttttcttcaatttattgactgtgt1980
ctggcagatgacaagacagtttcctaccgcatttgaattcaatgagtattttctcattac2040
cattttggaccacctatacagctgcttattcggaacattcctctgtaatagtgaacaaca2100
gagaggaaaagagaatcttcctaaaaggactgtgtcactgtggtcttacataaacagcca2160
gctggaagacttcactaatcctctctatgggagctattccaatcatgtcctttatccagt2220
agccagcatgcgccacctagagctctgggtgggatattacataaggtggaatccacggat2280
gaaaccacaggaacctattcacaacagatacaaagaacttcttgctaaacgagcagagct2340
tcagaaaaaagtagaggaactacagagagagatttctaaccgatcaacctcatcctcaga2400
gagagccagctctcctgcacagtgtgtcactcctgtccaaactgttgtataaaggactgt2460
aagatcaggggcatcattgctatacactcttgattacactggcagctctatgagtagaaa2520
gtcttcggaatttagaacccatctatgagagaaagttcagtcactttatttattttaaat2580
ctctctaggatgagtttagaactgtagcagtgcaggtggcttaagtgaagtaactccata2640
tgtaattacatgattatgatactaatcttttaagtatccaaagaatattaaaatacttca2700
atcctggattcacagtgggaacaagtttctattaaaaggcaaatgctgttacaaattttt2760
ggcatctggtaatattaaaaccattttagaaatacactctgtgctcactgtgcagaggaa2820
catcagttttcaaaccaacactgaaattctgtggcatcacatatattgggccttgatgtc2880
atgacagatcaaaatcatttgatatccctttctccattctaggtttttctttttttcagt2940
aactgatttaccttgatcacttttcaacttccatattcttcatatagtaaaaggcaaagt3000
gttgaagatactacggtgtggtagtagttgaaaattattgccgtcattatttacatactt3060
aagacatattagcaagttgatccaaaatgggaggccttatagatgtgcttgggggaaaat3120
gaaggggagaaagtagccatacaggagttcaaagaattccatgcccttcagattagccca3180
attaccagaaacatcatgaaagatattttaaaaactaattatttactacagtgtatttca3240
cttgtcttgtgtgtctgaacacacagaagctaattagcaagtttttaagaagtatttaaa3300
aatcttactaggattgacattttttctgaattctgtataaatagcttatagtgagaagta3360
ctgtgctcaaattttacatttttttcctttgcaaattctgtaatttcactcaacgattaa3420
gtctaccaaagaacacactgcatgtaaaagatgtattacaatctcaaagccagtaaaaga3480
aatcttgcttcactgttcacctgctacaagtaagagtttggtgctggtagaaacatttga3540
ctctgatgtctattttattctacataagagccatatgtaatgtactgtaacaaaggagct3600
tcttgtccccttggtcttttaattaaaagaaattccaactgacttttaaactttgttctt3660
gtccaaagttgccatttcttttttttccccagaaatatttggaaattattggaggaatat3720
gcaccccagatgaaaatgttcagtttgtacccatttttccttaaccaacacccaaatcaa3780
acaattaaaatatacagtgtttttccactcactaattcactatacagagagtctgaacct3840
tagcctccctcttggtcttgcagtgaggaagtttctattagtatatccaatttagcaaaa3900
ttggtaccaaaatgatttctttggtaattgtgtgaaatataagctttttaacagggcatt3960
taagtggctagcaaatcagtaattaaaaattaagctttctactccaagtatttcacaaac4020
gcatctgccattttcctcatttaaaccttggttatcttggcctgataccacataaaagaa4080
tgtagaatggctgaagagatcaagaatttaaagcttctagtcttaacatacttgcatcca4140
cttcaaattcaaatcaaaagccagggaaatctaagtgcaaccctaccacttctctgctga4200
gaaccttccagtggttcccctcaccttctgcagaagtctccaatatggagtacatgcact4260
tgggcatttaatatataccactggtgtgtgtgggagggagggaggaggaatactagccct4320
ttttatatatttacacaagcaaaacttttaaatatttgaattgacagttacatgtttcat4380
aactttgtatgtctattggttgtgcaggtgtaattttttccctttttgattagggttaca4440
aaatttagagaccagtatgattaagttgaagctccttagcctccttcgacctagtctctg4500
catacctcaacttttacgtaccaatgctactctgctgttcacaattgcctcatgtaatct4560
gcagattcctgcctccccactttggttcagtctgtcttgtgcacctggaacaactgttct4620
cccttgtgactaattcctattttctagagtttaggcatcatctcttcctttgggaagtta4680
tctgattcacgactgccttctctgacatccccaccttcctctgtgcccccatagcactgt4740
gtataccgctactaccactgcaattcacattatattggaatgaacaattcacatgtctac4800
cacaagtctgtaaacataaccttatttgaaatgaattgcaataaagctctgttacaacgt4860
aa4862
<210>6
<211>22
<212>dna
<213>人工序列
<220>
<223>正向引物5'utrmtmr2
<400>6
agcggcctccagtttctcgcgc22
<210>7
<211>26
<212>dna
<213>人工
<220>
<223>反向引物外显子3
<400>7
tctctcctggaagcagggctggttcc26
<210>8
<211>20
<212>dna
<213>人工序列
<220>
<223>正向引物小鼠mtmr2v1
<400>8
gactcactgtccagtgcttc20
<210>9
<211>18
<212>dna
<213>人工序列
<220>
<223>反向引物小鼠mtmr2v1
<400>9
cctccctcaggaccctca18
<210>10
<211>20
<212>dna
<213>人工序列
<220>
<223>正向引物小鼠mtmr2v2
<400>10
gactcactgtccagtgcttc20
<210>11
<211>18
<212>dna
<213>人工序列
<220>
<223>反向引物小鼠mtmr2v2
<400>11
cagctgggcactccctca18
<210>12
<211>33
<212>dna
<213>人工序列
<220>
<223>正向引物小鼠mtmr2v3
<400>12
aagataaaacatctcaaaaattataattgcttc33
<210>13
<211>18
<212>dna
<213>人工序列
<220>
<223>反向引物小鼠mtmr2v3
<400>13
cagctgggcactccctca18
<210>14
<211>33
<212>dna
<213>人工序列
<220>
<223>正向引物小鼠mtmr2v4
<400>14
aagataaaacatctcaaaaattataattgcttc33
<210>15
<211>20
<212>dna
<213>人工序列
<220>
<223>反向引物小鼠mtmr2v4
<400>15
gactcactgtccagtgcttc20
<210>16
<211>20
<212>dna
<213>人工序列
<220>
<223>正向引物总小鼠mtmr2
<400>16
tcctgtgtctaatggcttgc20
<210>17
<211>20
<212>dna
<213>人工序列
<220>
<223>反向引物总小鼠mtmr2
<400>17
aaccaagagggcaggatatg20
<210>18
<211>19
<212>dna
<213>人工序列
<220>
<223>正向引物人mtmr2v1
<400>18
actccttgtccagtgcctc19
<210>19
<211>18
<212>dna
<213>人工序列
<220>
<223>反向引物人mtmr2v1
<400>19
gactccctcaggaccctc18
<210>20
<211>33
<212>dna
<213>人工序列
<220>
<223>正向引物人mtmr2v2
<400>20
aagataaaacatctcaaaaattataattgcctc33
<210>21
<211>18
<212>dna
<213>人工序列
<220>
<223>反向引物人mtmr2v2
<400>21
gactccctcaggaccctc18
<210>22
<211>33
<212>dna
<213>人工序列
<220>
<223>正向引物人mtmr2v3
<400>22
aagataaaacatctcaaaaattataattgcctc33
<210>23
<211>18
<212>dna
<213>人工序列
<220>
<223>反向引物人mtmr2v3
<400>23
gagcgagactccctcctc18
<210>24
<211>33
<212>dna
<213>人工序列
<220>
<223>正向引物人mtmr2v4
<400>24
aagataaaacatctcaaaaattataattgcctc33
<210>25
<211>18
<212>dna
<213>人工序列
<220>
<223>反向引物人mtmr2v4
<400>25
ctggactgcatgggcctc18
<210>26
<211>23
<212>dna
<213>人工序列
<220>
<223>正向引物总人mtmr2
<400>26
tttcctgtctctaataacctgcc23
<210>27
<211>18
<212>dna
<213>人工序列
<220>
<223>反向引物总人mtmr2
<400>27
ccaggagggcagggtatg18
<210>28
<211>71
<212>dna
<213>智人(homosapiens)
<400>28
ttaatgtgttaagaaccattgacatttgaagatcatcagaagtgaagataaaacatctca60
aaaattataat71
<210>29
<211>105
<212>dna
<213>智人(homosapiens)
<400>29
gagtgcccagctgcttgggttttagttgatgccagacatagtcaagttgacagccaagat60
tagccatcacaccagtgttctatcctgttgaatccaattctatag105
- 还没有人留言评论。精彩留言会获得点赞!