用于调制α-酮戊二酸(2OG)依赖型加氧酶并作为其功能替代的化合物的制作方法
本发明涉及的是作为2og依赖型加氧酶(dios)中的α-酮戊二酸(也称2-氧戊二酸酯或2og)共基质的功能等价的化合物,特别是用于在存在dios的竞争性抑制剂,如2-羟基戊二酸(2hg),的情况下来实现其功能建立。
背景技术:
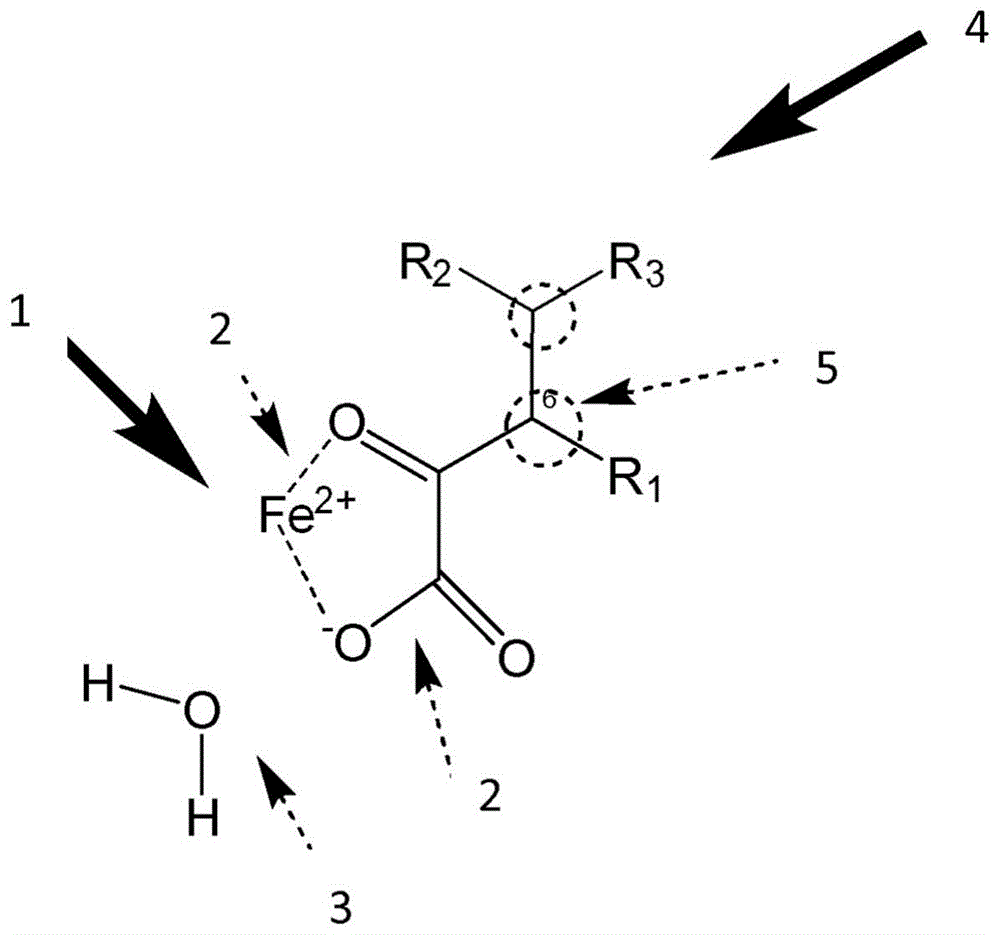
:α-酮戊二酸-(2-氧戊二酸酯-2og)依赖型二氧酶(dios也被称为2og加氧酶)是氧依赖型酶,其将α-酮戊二酸(2-氧戊二酸酯2og,2kg)用作共基质,将非血红素fe(ii)用作辅助因子。其催化多种氧化反应。这些氧化反应包括(但不限于)羟基化反应、去甲基化、环膨胀、闭环和引入双键。[1][2、3]。2og依赖型二氧酶参与大量生物过程和功能。[4、5]。在细菌等微生物中,2og依赖型二氧酶参与生物合成的基本功能[6-8]。在植物中,2og依赖型二氧酶参与大量不同的植物代谢反应[9]。其中包括(但不限于)黄酮类生物合成和乙烯生物合成[10]。就哺乳动物和人类而言,2og依赖型二氧酶在生物合成(如胶原蛋白的生物合成[11]和l-半胱氨酸的生物合成[12])、翻译后修饰(如蛋白质羟化[13])、表观遗传调控(如组蛋白和dna去甲基化[14])以及能量代谢的传感器等方面均具有功能作用。[15]。2og依赖型二氧酶通过将单个氧原子嵌入基质来催化氧化反应。这一点总是与共基质2og氧化成琥珀酸酯和二氧化碳相关[16]。许多2og依赖型二氧酶的催化活性取决于能够将辅助因子铁保持其二价形式或将其还原为这种氧化态的还原剂[1][17-20]。就维生素c(抗坏血酸)也讨论过这种机制。但并非所有功能均能得到解释[21]。2og依赖型二氧酶的特征在于共同的催化机制。在第一步骤中,将2og和基质结合至活性结合位点中[22-24]。2og在活性中心与铁(ii)(niii;mnii)直接配位,而基质以紧邻但非直接配位的方式与金属结合。第二步骤是分子氧的结合,该分子氧在fe(ii)(niii;mnii)中心占据第三位点。从而以形成琥珀酸、二氧化碳和活性金属(iv)氧化物中间体的方式实现氧化脱羧反应,其随后氧化该基质[25-31]。铁的替代和作用是研究的主题[59]。2004年讨论过细菌2og依赖型二氧酶,脱乙酰氧基头孢菌素-c合成酶(daocs)的替代机制。所提出的“乒乓”机制不同于共识机制(见上文),2og和氧首先在没有基质的情况下与酶活性位点的fe(ii)中心结合。然后发生2og的非偶联氧化,以产生反应性的fe(iv)-氧化物质,接着释放琥珀酸和二氧化碳并与被氧化的基质结合。但后来在2014年的研究表明,daocs也很可能遵循2og依赖型加氧酶的一般共识机制[32]。所有的2og依赖型二氧酶均含有一个保守的双链β-螺旋(dsbh),其与两个β-片形成裂隙[33、34]。活性位点含有一个高度保守的2-his-1-羧酸(hxd/e...h)-氨基酸残基三联体基序,其中催化必需金属由两个组氨酸残基和一个天冬氨酸或谷氨酸残基固定[35]。但它们在活性中心的氨基酸排列方式不同。从x射线晶体学、分子动力学计算(md)和核磁共振光谱发现,某些2og依赖型二氧酶通过诱导适应机制结合其基质。例如曾观察到人脯氨酰羟化酶同工型2(phd2)的基质结合中的蛋白质结构变化[36、37],[38]参与氧稳态的2og依赖型二氧酶[39],和异青霉素-n合酶(ipns),一种微生物2og依赖型二氧酶[40]。在许多疾病中,dios的活性会发生改变,通常会降低。鉴于2og依赖型二氧酶所发挥的重要生物学作用,它们是治疗疾病的重要靶点。使用小分子作为2og的功能替代作为共基质的观点迄今为止在此功能方面尚不为人所知,并且是通过影响dios来治疗疾病的全新药理方法。从而使得缺乏2-og或者肿瘤代谢物(如2hg)在酶中的竞争性抑制所引起的病证变得可以治疗。α-酮戊二酸(2og)依赖型加氧酶(dios)催化大量显著的氧化反应。在人类和动物中是通过羟基化反应发生的羟基化和n-去甲基化;在植物和微生物中,它们催化环形成、重排、去饱和和卤化等反应。dios的催化灵活性反映在它们的生物学功能上。dios除了被发现在胶原生物合成中的作用以外,它们在动植物的发育、转录调控、核酸(dna,rna)的修饰/修复、脂肪酸代谢、干细胞(psips)的形成和稳定化以及次生代谢产物的生物合成中,包括医学上重要的抗生素,均发挥重要作用。发明目的本发明的目的是提供用于为2og依赖型加氧酶中的共基质2og承担功能的化合物,及其在治疗疾病方面的应用,其与2og依赖型加氧酶相关(dios)。技术实现要素:本发明提供化合物,其中所述化合物选自通式(i)或通式(ii)中的化合物其中as为来自酶的结合口袋的氨基酸;me为来自催化中心的金属;r1和r2为氧(羟基)或羧基、卤素,特别是氟、氯或碘,单重至多重卤化的甲基,特别是ch2f至cf3;以及cn表示一个碳原子、一个杂原子或通向一个杂环的桥。其特征在于,其可以占据与催化中心的反应距离,从而形成相应的过渡态(ts),以实现作为共基质的必要功能。此外就所述化合物而言,r1为氢或ch2r3基团,其中r3为氢或氧(羟基、羰基)或较短的c链(c1至c4)。在所述化合物的另一方面中,所述化合物是一个环系统的组成部分,其中环的大小在3至5个原子之间且包含至少一个杂原子。在所述化合物的另一实施方式中,c7可以由具有最多5个原子的碳链构成且含有双键。所述化合物还可以包括作为羧酸的f2。对于通式(ii)中的化合物,r2可以表示氢原子、甲基、具有最多6个碳原子的烷基,其可以是支链饱和的或不饱和的或者本身也含有杂原子。此外对于通式(ii)中的化合物,在r1和r2之间布置有具有单键或双键的架桥环状结构,并且含有杂原子。根据本发明,也可以设有通式(i)和通式(ii)中的化合物的混合物。在另一方面中,单独或者以前述混合比例组合地使用相应化合物的制药学上可接受的盐和互变异构体。本发明的另一主题是前述化合物中的一个作为药物的应用。本发明的另一主题是前述化合物作为药物中与其他有效成分的功能共基质的应用,其中所述其他有效成分可以选自但不限于包括以下的群组:化疗药物、细胞抑制剂(烷化剂、抗代谢物、拓扑异构酶抑制剂、有丝分裂抑制剂、抗生素、抗体、激酶抑制剂、蛋白质组抑制剂和支持性肿瘤治疗药物,如干扰素、细胞因子、肿瘤坏死因子和idh抑制剂)。本发明还包括前述化合物作为用于预防、治疗或后续护理癌症、神经退行性疾病以及先天性或后天性代谢紊乱的药物,的应用。本发明的另一主题是通式(i)或通式(ii)中的化合物在酶中显示ts的应用,其用于制造用于预防、治疗或后续护理癌症、神经退行性疾病以及先天性或后天性代谢紊乱的药物。最后,本发明还包括一种药物,其包括通式(i)或通式(ii)中的化合物。附图说明下面结合附图和实施例对本发明进行描述和展示。对本案中的本领域技术人员而言显而易见的是,本发明并非仅限于附图和实施例的内容。图中:图1为酶中作为共基质的功能所需的结合比例图2为tet2酶,tet抑制剂nga(n-草酰甘氨酸)的结合图3、4为tet酶中2og的结合比例图5a、5b为2hg和2og竞争导致的tet酶功能丧失图6为dka对共基质2og的功能替代图7为已知疗法以及本发明的新型药理学理念图8为dka毒性图9为2og毒性图10为idh抑制剂与本发明的共基质的组合图11为idh1-mthct116中2-羟基戊二酸水平与5-hmdc水平的相关性图12为在存在竞争性抑制剂2hg的情况下通过dka恢复tet酶的功能图13-18为可以用作tet和hif上共基质2og的功能替代的选定化合物图19-27为作为hf酶的功能替代的选定化合物图28为借助dka来恢复功能性图29为mtt细胞毒性测试图30、31为采用2,3-二酮古洛糖酸的细胞凋亡测试图32、33为采用2,3-二酮古洛糖酸的hmdc测量图34为hct116细胞的蛋白质印迹具体实施方式本发明涉及一种酮戊二酸依赖型二氧酶的替代共基质([1][6、7]表1-2;表4),用于其功能建立和调控,目的是实现对癌症、神经退行性疾病和年龄相关疾病的治疗效果。由失调引起,特别是由柠檬酸循环中的代谢脱轨引起的表观遗传疾病,也是该疗法的重点。本发明的说明书中的术语共基质指的是,在酶促反应中使得真正意义上的基质发生反应所需的低分子量化合物。因此,共基质用作与基质一起发生反应的某种形式的“辅助分子”,但自身不具有催化作用。一般而言,治疗疾病时通过抑制靶结构(竞争性、非竞争性、全能性、共价性)来影响药物受体。涉及dios时同样通过抑制idhs或α-hif来寻求这个目标,其结果和毒性事件部分地存在矛盾[8-13]。本发明涉及一种针对疾病治疗的酶影响新途径,即用一种我们的物质来对天然共基质进行功能替代。通过相应的共基质替代(调制因子)来调控dios可对其加以优化,其中如此地继续实施这一点,以便对表观遗传调控、细胞代谢的恢复和调控以及对相关的更多遗传调控进行针对性影响。还可以用来治疗dio依赖型孤儿病,如2-羟基-戊二酸尿症。以及应用于对传统治疗的肿瘤进行后续治疗。本发明提供了用于替代和调制或者调控α-酮戊二酸(2og)依赖型加氧酶的化合物。本发明的化合物能够用来治疗与α-酮戊二酸二氧酶(dios)的功能(活性)相关的疾病。这些疾病特别是包括癌症、阿尔茨海默病、帕金森病、老年性疾病。本发明所提供的化合物的在本文中也称为共基质的变体,能够在适应征中调控和适应相应的靶结构。本发明提供通式(i)或通式(ii)中的以下化合物:通式(i)中的化合物可以采用以下方案:其中as为来自酶的结合口袋的氨基酸,以及me为来自催化中心的金属;r1和r2可以是氧(羟基)或羧基、卤素,特别是氟、氯或碘,单重至多重卤化的甲基,特别是ch2f至cf3;r1可以是单重氢或ch2r3基团,其中r3为氢或氧(羟基、羰基)或较短的c链(c1至c4)。下面示出本发明的化合物的一个例子:通式(i)中的化合物可以是一个环系统的组成部分,其中环的大小在3至5个原子之间且包含一或多个杂原子。c7可以由具有最多5个原子的碳链构成且含有下例所示的双键:这个链也可以按以下方式键合在一个环系统中cn在通式(i)中是至少一个碳原子、一个杂原子或通向一个杂环的桥,其含有一或多个杂原子。r2表示一个羧酸,cn可以表示例如2-氧代己二酸中的一个、两个或多个碳原子:或者氧代丁二酸酯中r2和cn可以是包含或不包含杂原子的不饱和或饱和环。2-氧代-4-苯基丁酸3-环丙基-2-氧代丙酸酯2-(2-氨基噻唑-4-基)-2-氧代乙酸通式(ii)中的化合物可以采用以下方案:其中as为来自酶的结合口袋的氨基酸,以及me为来自催化中心的金属;r1和r2可以是氧(羟基)或羧基、卤素,特别是氟、氯或碘,单重至多重卤化的甲基,特别是ch2f至cf3;r1可以是单重结合的氢或ch2r3基团,其中r3为氢或氧(羟基、羰基)或较短的c链(c1至c4)。通式(ii)中的化合物可以是一个环系统的组成部分,其中环的大小在3至5个原子之间且包含一或多个杂原子。c7可以由具有最多5个原子的碳链构成且含有以下示例性示出的双键:5-氧代己-2-烯二酸4-甲基-5-氧代己-2-烯二酸这个链也可以键合在一个环系统中:cn在通式(ii)中是一个碳原子或一个杂原子。r2表示一个羧酸,cn可以表示例如2-氧代己二酸(参见上文)中或者氧代丁二酸酯(参见上文)中的一个、两个或多个碳原子。cn可以是含有一或多个杂原子的杂环的组成部分或者是通向该杂环的桥,例如:在通式(ii)中,r1可以氢原子、具有最多5个碳原子的甲基或烷基,其可以是支链饱和的或不饱和的或者也含有杂原子。r2同样可以是氢原子、甲基、具有最多6个碳原子的烷基,其可以是支链饱和的或不饱和的或者也含有杂原子,参阅下例。在通式(ii)中,在r1和r2之间可以布置有架桥环状结构,其可以不饱和或饱和或者含有杂原子。该所形成的循环可以是3环、4环、5环或6环。该环可以含有一或多个双键,也可以另外地或单独含有一或多个杂原子,参见下例。一个双键,与杂原子结合一或多个双键,与杂原子结合一或多个双键,与杂原子结合一或多个双键,与杂原子结合一或多个双键,与杂原子结合可以延长碳原子的链并且按下例的方式使用取代基。所述化合物可以用作物质、用作盐以及以缓冲形式使用。也可以以酯化的方式使用羧酸,其中也可以用高链醇(最多c12原子)进行酯化。在制药技术层面上实施加工。制剂,如霜剂、软膏、凝胶、纳米制剂、输液、片剂、胶囊。维生素c可以用作传统的药物前体,其在体内可以代谢成通式(i)或通式(ii)中的化合物以及代谢成通式(ii)中的下文所示结构。羧基的酯化同样产生吸收和细胞摄取有所改善的药物前体。药物前体结构的更多选项参阅[48]。药物前体与共基质替代(调制因子)的组合是可行的。可以通过口服、局部或输注进行给药。可以将传统肿瘤治疗药物与调制因子相混合或者组合式给药以提高治疗效率并实现治疗优化。针对本文所主张的结构要素,多种已知结构(cas)可供选择,它们可以根据具体dios而用作调制因子从而对dios产生影响。用作药物前体的结构同样可供选择。最简单的例子是维生素c及其衍生物,如2-o-α-d-吡喃葡萄糖-1-抗坏血酸,其可以代谢为2,3-二酮古洛糖酸(dka),也可以代谢为3,4,5-三羟基-2-氧代戊酸(iii;2kgl)并在dios中用作共基质的无毒功能替代。维生素c在各种dios和相关过程中的作用机制尚未阐明,可以通过其作为dkg和2kgl的药物前体功能来解释。这样就除了氧化还原作用以外,还导致维生素c在不同靶结构上具有非常广泛的活性。[17,49-61]。通过使用调制因子,就能重新激活dios并且竞争性地将肿瘤代谢物(羟基戊二酸hg)从活性中心挤出。从而恢复并调整该功能。表1和表2中总结了hgs的影响和共基质调控目标的实例。表1表2使用本发明的化合物也包括与相应化合物的制药学上可用的盐、互变异构体和立体异构体(以及上述的混合物)一起使用。所述化合物是这些基本式的进一步研发和修改的基础。在本发明的范围内,通式(i)的制药学上相容或可用的盐、药物前体、对映异构体、非对映异构体、外消旋混合物、结晶形式、非晶形式、无定形形式、未溶解形式和溶解物均如公开内容。本文所用的表达式“制药学上可用的盐”将通式(i)和(ii)中的化合物的盐包括在内,其用相对无毒(即制药学上可接受的)酸或碱根据在本发明的化合物上发现的特定取代基而制成。例如在本发明的化合物含有酸性官能团的情况下,可以通过将这类化合物的中性形式与足够量的期望碱进行接触而获得碱加成盐(要么以纯形式,要么以适宜的惰性溶剂)。制药学上可接受的碱加成盐的非限制性实例包括钠盐、钾盐、钙盐、铵盐、有机氨基盐或镁盐或类似盐。例如在本发明的化合物含有碱性官能团的情况下,可以通过将这类化合物的中性形式与足够量的期望酸进行接触而获得酸加成盐(要么以纯形式,要么以适宜的惰性溶剂)。制药学上相容的酸加成盐的非限制性实例包括衍生自无机酸(如盐酸、氢溴酸、硝酸、碳酸、磷酸、部分中和的磷酸、硫酸、部分中和的硫酸、氢碘酸或亚磷酸等诸如此类)的酸加成盐,以及衍生自相对无毒的有机酸(如乙酸、丙酸、异丁酸、马来酸)的盐。丙二酸、苯甲酸、琥珀酸、亚麻酸、富马酸、扁桃酸、邻苯二甲酸、苯磺酸、对甲苯磺酸、柠檬酸、酒石酸、甲磺酸等。同样包括在内的还有氨基酸的盐,如精氨酸等,以及有机酸的盐,如葡萄糖醛酸或银河酸等。本发明的某些特定化合物可以既包括碱性官能团又包括酸性官能团,使得这些化合物可转化为碱性或酸性加成盐。盐与碱的接触可使本发明的化合物的中性形式或酸再生,并以传统方式分离母体化合物。所述化合物的起始形式与各种盐形式在某些物理特性上有所不同,如在极性溶剂中的溶解度方面,但就本发明的目的而言,这些盐在其他方面与所述化合物的起始形式等效。本发明的化合物可以具有手性或不对称的碳原子(光学中心)和/或双键。本发明也将外消旋体、非对映异构体、几何异构体和各个光学异构体包括在内。本发明的化合物也可以以非溶解形式和溶解形式(包括超螺旋形式)存在。一般而言,溶解形式与非溶解形式等效并且也被本发明包括在内。本发明的化合物也可以以多晶或无定形形式存在。本发明的化合物还可以在所谓的药物前体形式中存在。本发明的化合物的药物前体指的是易于在生理条件下发生化学变化以提供本发明的化合物的化合物。此外,本发明化合物中的药物前体可以在离体环境中通过化学或生物工艺而发生转化。举例而言,在将药物前体与适宜的酶或化学试剂一起例如置于透皮贴剂储存器中的情况下,其能够缓慢地转化成本发明的化合物。本文所描述的本发明化合物可以合适的剂量施用于哺乳动物,如人或家畜。家畜的非限制性实例为猪、牛、水牛、绵羊、山羊、兔子、马、驴、鸡、鸭、猫、狗、真猪或仓鼠。最佳供人服用。优选服用方式取决于本发明的化合物(通式(i))的形式。如前所述,通式(i)中的化合物的形式为制药学上相容的盐、药物前体、对映异构体、非对映异构体、外消旋混合物、结晶形式、非晶形式、无定形形式、未溶解形式或溶解物。本发明的化合物可以口服、肠外服用,如皮下、静脉注射、肌肉注射、腹膜内、鞘内、眼内、透皮、透粘膜、硬膜下、局部或局部通过离子导入、舌下、吸入喷雾的方式服用。还可以采用气雾剂或直肠等类似方式,视情况还包括传统的制药学上可接受的赋形剂。结合本发明所使用的本发明化合物可以使用一或多个生理载体或赋形剂来调配成医药组合物。实施口服时,本发明的医药组合物例如可以采取片剂或胶囊的形式,其按传统方式用制药学上可接受的辅剂制成,这些辅剂例如为粘合剂(如预胶化玉米淀粉、聚乙烯吡咯烷酮、羟丙基甲基纤维素)、填料(如乳糖、微晶纤维素、磷酸氢钙)、润滑剂(如硬脂酸镁、滑石粉、二氧化硅)、崩解剂(如钾淀粉、淀粉糖酸钠)或润湿剂(如十二烷基硫酸钠)。所述医药组合物可以用生理相容的载体给患者服用。在一种特殊实施方式中,“制药学上相容”这一表述的意思是其经监管机构或其他公认的药典授权应用于动物,特别是人类。“载体”这一表述指的是用来服用治疗药物的稀释剂、佐剂、赋形剂或介质。这类制药学上的载体可以是无菌液体,如水和油,包括源于石油、动物、植物或合成的水和油,如花生油、大豆油、矿物油、芝麻油等诸如此类。静脉注射医药组合物时,水是优选载体。盐溶液和水性葡萄糖与甘油溶液也可用作液态载体,特别是用于注射溶液。适宜的制药学辅剂包括淀粉、葡萄糖、乳糖、蔗糖、明胶、麦芽、大米、面粉、白垩、硅胶、硬脂酸钠、甘油单硬脂酸酯、滑石粉、钠离子、干脱脂乳、甘油、丙二醇、乙二醇、水、乙醇等诸如此类。所述组合物视需要也可以含有少量润湿剂或乳化剂或ph缓冲剂。这些组合物可以以软膏、溶液、悬浮剂、乳剂、片剂、丸剂、胶囊、粉剂、缓释调配物等诸如此类的形式存在。优选形式为软膏。所述组合物可以调配成具有传统粘合剂的栓剂和载体,如甘油三酸酯。所述口服制剂可以含有具备制药学品质的标准辅料,如甘露醇、乳糖、淀粉、硬脂酸镁、糖精钠、纤维素、碳酸镁等等。e.w.martin在《remington'spharmaceuticalsciences》一文中描述过适宜的制药学上的载体。这类组合物含有治疗有效量的上述化合物,优选以纯化的形式,连同适量的载体,以便为患者提供正确服用形式。该制剂应符合服用方式。用于口服的液体制剂例如可以是溶液、糖浆或悬浮液的形式,或者可以作为干品呈现,以便在使用前与水或其他合适的介质一起使用。这种液态制剂可以用常规方式制成,使用的是制药学上相容的添加剂,例如悬浮剂(如山梨醇、糖浆、纤维素衍生物、氢化食用脂肪)、乳化剂(如卵磷脂、洋槐胶)、非水性介质(如杏仁油、油、油性酯、乙醇、分馏植物油)、防腐剂(如甲基或丙基对羟基碳酸盐、山梨酸)。这些制剂视情况也可以含有缓冲盐,调味剂,着色剂和甜味剂。可以对口服制剂进行相应调配以实现本发明的医药组合物的可控释放。为了进行吸入式服用,使用适宜的推进剂(如二氯二氟甲烷、三氯氟甲烷、二氯四氟乙烷、二氧化碳或其他适宜的气体),方便地从加压包或雾化器中以气溶胶喷雾呈现的形式来释放本发明的医药组合物。在涉及预分散的气雾剂的情况下,可以通过提供用于释放适当量的阀来确定剂量单位。可以对例如用于吸入器或注气器的明胶所构成的胶囊和套筒进行调配,其含有本发明的医药组合物的粉末混合物和适宜的粉末基质,如乳糖或淀粉。为了通过注射进行肠外服用,例如可以通过推注注射或连续输注来对本发明的医药组合物进行调配。注射部位是静脉内、腹膜内或皮下。注射用制剂可以以单位剂型(如安瓿瓶、多剂量容器)和附加防腐剂呈现。本发明的医药组合物可以采取在油性或水性介质中的悬浮剂、溶液或乳剂等形式,并可以含有调配剂,如悬浮剂、稳定剂或分散剂。替代地,所述制剂可以在使用前以用于与适宜的介质(如无菌无热原水)混合使用的粉末形式存在。通常情况下,所述用于静脉服用的组合物是在无菌等渗水性缓冲液中的溶液。所述组合物视需要也可以含有增溶剂和局部麻醉剂,如利多卡因,以减轻注射部位的疼痛。一般而言,将这些成分分离或以单位剂型混合在一起,例如作为干冻干粉剂或无水浓缩物装在密封的容器中,如安瓿瓶或标明活性剂的量的小袋中。通过输注来服用所述组合物时,无需使用装有无菌水或医药级食盐的输液瓶。通过注射来服用所述组合物时,可以提供装有注射用无菌水或食盐溶液的安瓿瓶,以便在服用前将这些成分混合在一起。对本领域普通技术人员而言显而易见的是,本发明也将缓释剂型包括在内,其设计为以预定速率释放药物,以在特定时间内保持恒定的药物浓度且副作用最小。这一点可以通过多种制剂或装置来实现,包括微球、纳米颗粒、脂质体和其他聚合物基质,如药物-聚合物共轭物(如水凝胶)或生物降解材料,如包封有效成分的羟基乙酸共聚物(plga)。优选根据治疗特定疾病的特殊需要来对释放进行调整,例如在治疗糖尿病时持续释放注射液。缓释的定义更像是“可控释放”或“库房用药”而非“持续”。视需要也可以在包装或分配器中提供本发明的医药组合物,其可以具有包含该制剂的一或多个单位剂型。所述包装例如可以包括金属或塑料薄膜,如透明塑料包装。该包装或分配设备可以配有服用说明。本发明的医药组合物可以作为单独的有效成分服用或者与其他有效成分一起服用。这类附加的活性剂主要应选自与治疗同一疾病相关的有效成分。治疗肥胖症时,应从抗肥胖药物的群组中选择附加有效成分。以此类推,抗糖尿病药物和抗nafld/nash(非酒精性脂肪肝)药物以及抗血脂异常药物可以作为附加的有效成分。此外还应从具有增重副作用的抗精神病治疗的有效成分中选择这种附加的有效成分。也可以将本发明的化合物或本发明的组合物用作用于预防、治疗和后续治疗肿瘤疾病的共基质。优选地,肿瘤疾病选自由以下构成的群组:耳鼻喉区域的肿瘤,包括内鼻、鼻旁窦、鼻咽、唇、口腔、口咽、喉、下咽、耳、唾液腺的肿瘤和副神经瘤、肺部肿瘤,包括非副支气管癌、副支气管癌、纵隔肿瘤、胃肠道肿瘤,包括食管、胃、胰腺、肝脏、胆囊、胆管肿瘤、小肠、结肠的肿瘤和肠癌及肛门癌、泌尿生殖系统肿瘤,包括肾脏、输尿管、膀胱、前列腺、尿道、阴茎和睾丸的肿瘤、妇科肿瘤,包括宫颈、阴道、外阴的肿瘤、子宫癌、恶性滋养细胞病、卵巢癌、子宫输卵管(tubafaloppii)、腹腔肿瘤、乳腺癌、内分泌器官肿瘤,包括甲状腺、甲状旁腺、肾上腺皮质的肿瘤、胰腺内分泌肿瘤、类癌和类癌综合征、多发性内分泌肿瘤、骨和软组织肉瘤、间皮瘤、皮肤肿瘤、皮肤和眼内黑色素瘤、中枢神经系统肿瘤、婴儿肿瘤,包括视网膜母细胞瘤、威尔姆斯肿瘤、神经纤维瘤、神经母细胞瘤、尤文肉瘤肿瘤家族、横纹肌肉瘤,包括非霍奇金淋巴瘤的淋巴瘤、皮肤t细胞淋巴瘤、原发性中枢神经系统淋巴瘤、霍奇金病、包括急性白血病的白血病、慢性髓细胞和淋巴细胞白血病、浆细胞瘤、骨髓增生异常综合征、副肿瘤综合征、原发肿瘤不明的转移(cup综合征)、转移性肿瘤,包括脑转移瘤、肺转移瘤、肝转移瘤、骨转移瘤、胸膜和心包转移瘤、以及恶性腹水、腹膜癌、免疫系统相关的恶性肿瘤,包括艾滋病相关的恶性肿瘤,如卡波西肉瘤、艾滋病相关的淋巴瘤、艾滋病相关的中枢神经系统淋巴瘤、艾滋病相关的霍奇金病和艾滋病相关的肛门肿瘤、移植相关的恶性肿瘤。治疗肿瘤疾病时,将本发明的化合物与化疗药物相结合,其可以选自包括以下的群组:抗体、烷基化剂、铂类似物、嵌入剂、抗生素、有丝分裂抑制剂、紫杉烷。本发明的更多方面、特征和优点可从以下详述中轻易得出,该详述中简单地示出优选实施方式和实施方案。本发明也可以以其他不同的实施方式实现且其各种细节可以在各种显而易见的方面中加以变更,而不脱离本发明的原理和范围。相应地,附图和说明书起展示作用,不具有限制性。本发明的其他目的和优点部分地在下文中加以说明并且部分地从说明书中是显而易见的或者可从本发明的阐述中得出。图1为酶中作为共基质的功能所需的结合比例。为此,在酶的反应中心存在二价铁原子1,其不与共基质共价相互作用。共基质-铁的所需距离为(箭头2)。水分子具有的距离(箭头3)并且对进行中的反应同样其主要作用。综上所述,此处展示了对于共基质功能替代的酶中的反应的过渡态(transistionstate;ts)。此外,图1还示出了dios的活性中心的周围环境和共基质/辅助因子络合物4的嵌入和非共价固定(参阅2-his-1-羧基面部三联体[41][42])。图中还示出了具有相同电子特性的一个碳原子5。通式(i)和(ii)中的本发明的化合物可以主动或被动地通过转运体(如dka、2-okg)或经过相应的衍生化后进入靶细胞。其中as为来自酶的结合口袋的氨基酸;me为来自催化中心的金属;r1和r2可以是氧(羟基)或羧基、卤素,特别是氟、氯或碘,单重至多重卤化的甲基,特别是ch2f至cf3;以及cn表示一个碳原子、一个杂原子或通向一个杂环的桥。这种在dios中使用生理共基质2og的功能等价的做法尚未被披露,总体上代表了人类代谢酶的功能影响和恢复以治疗疾病的范式切换。因此,这种做法与其他旨在影响酶的催化金属(辅助因子)或减少肿瘤代谢物(如2hg)形成的方案(所谓idh抑制剂)不同。从而明确了这种范式切换。通过这种新方法和所展示的效果,迄今为止没有得到解答的dios功能,如十-十一易位(tet)酶和hif脯氨酸羟化酶的功能问题变得可以解释[21]。本发明建立在发现某些化合物的基础上,其在存在肿瘤代谢物(hg)的情况下恢复dios功能,从而提供广泛的疾病治疗选项。迄今为止未曾描述过能够模拟2og作为共基质的生理功能的化合物。本发明一方面是影响活细胞中酶的全新做法,另一方面,本发明能够首次全面回答关于dios的未解问题,并将其转化为dio依赖型疾病的新治疗方案。本发明还基于对接实验和分子动力学研究并采用homann法和算法,从而使得dio酶的活性位点(结合位点或相互作用位点)的实际条件得以显示,并在无细胞和细胞系统中通过相应的实验加以证明。举例而言,根据对tet2酶的x射线结构研究,可了解tet抑制剂nga(n-草酰甘氨酸)的结合(参阅图2)。2og在tet酶中的结合比例尚不清楚,假设与图2类似。图3和图4中,首次对这些结合比例(ts中)进行展示。结合图1和图2就能看出,保持了酶反应所需的结合距离。如图1所阐述的那样,水是酶络合物中的必要组分,此处一并显示。为了解释肿瘤代谢物2hg导致的dios功能丧失(这里以tet为例),必须明确其在酶中的空间结构(参阅图4)。再次可以看出tet中的2hg和hif酶进行相互作用,并且由于羟基与催化金属(fe2+)的相互作用而无法作为共基质进入反应。同时,基于2hg在酶中较强的亲和力而竞争性地将2og挤出。使得酶失去功能(图5a和图5b)。基于前述研究和结果而得出以下认识:需要承担2og功能性的化合物必须在类似的电子条件下以相同方式存在于酶中(图1)。根据这些初步结果而得出以下结论:为了实现2og的功能替代和dio功能的恢复,只有采用在dio酶中的结合力强于2og,但同时具有并能承担作为共基质的功能的化合物。单凭较强的结合力会对dios造成抑制,从而与2hg的抑制类似地导致竞争性抑制[43]。这一点是与作为功能等价的期望功能相悖的。但如果dio酶中较强的结合力与进入相应反应的功能等价并存,则会存在某些化合物,其可以将2hg挤出并通过共基质功能恢复dios(如5-甲基胞嘧啶5mc的去甲基化的tet功能)。这些化合物本身能够取代2og。图6为dka对共基质2og的功能替代,其显示了在tet酶中作为功能辅助因子的dka的功能的无细胞测定的结果。为此,在无细胞tet测试中进行研究(在含有50mmnacl8μmfe(nh4)2(so4)25mmatp、3mmdtt、0.25μgtet酶和鲱鱼精子dna的50mmhepes缓冲液中)。培养8小时后,借助于质谱进行评价。用功能性辅酶dka取代天然2og。测试表明,dka可以承担天然2og的功能并可以用作天然共基质的替代。编码异柠檬酸盐脱氢酶(idh)的基因变异促进将2og还原为肿瘤代谢物d-2-羟基戊二酸(2hg),此举导致(tet)抑制dna中的5-甲基胞嘧啶(5mc)脱甲基化为5-羟甲基胞嘧啶(5hmc),以及抑制jumonjic-domain脱甲基酶(jmjc)甲基化为组蛋白赖氨酸(hk)残基(hkme),和fto抑制n6-甲基腺苷(m6a)[44、45]。2hg是一种能抑制多种去甲基化酶的肿瘤代谢物,这一点导致基因组和转录组甲基化谱的变化,以及基因表达和基因组拓扑结构的变化[46]。从而导致相应的病证,包括癌症,考虑到基因组的总甲基化时还导致衰老过程。癌症治疗的基石,包括后来表观遗传治疗的成功,均采用了有效合理的药物组合。核心在于将表观遗传活性药物(包括我们的2og功能替代共基质)与其他疗法相结合,并进行优化。有鉴于此,tet酶由于其核心的表观遗传作用,而成为替代天然辅酶2og的专门目标。下面结合tet酶对本发明的化合物的应用进行阐述。迄今为止已知的科学文献在考虑将tet酶作为治疗的靶点时,以图7所示以下关系为出发点。图7示出已知疗法以及本发明的新型药理学理念作为补充。缩写的含义如下:hdac组蛋白去乙酰化酶;ezh2zeste同源物2的增强子是一种组蛋白赖氨酸n-甲基转移酶;dot1l端粒沉默1-like的破坏者;bet溴结构域和末端外基序。通过外部输入的化合物来实现共基质的功能替代和功能承担,这一点迄今为止从未被考虑过,因而构成范式切换。本发明的化合物,特别是dka和二酮古洛糖酸(2og),因其无毒作用而也可以以相应的药理浓度(使用的最高浓度为1mm)使用,参阅图8和图9,其显示毒性研究结果。目前在临床试验中正在寻求各种组合策略,包括表观遗传治疗与化疗的组合,以及靶向治疗与免疫治疗的组合。2og在tet中的功能等价在迄今为止的组合治疗中尚未被考虑过,因其仍属未知,只有用本发明才得以受到考虑。癌细胞可能由于转录适应而逃避选择性压力,这一证据为利用表观遗传学疗法来阻断或逆转耐药性提供了分子依据。当前正在进行首次临床研究的idh抑制剂中,也显示了这一点[47]。通过将idh抑制剂与本发明的共基质组合在一起,就能在细胞水平上显示idh抑制剂的有效性增强(参见图10),从而令人印象深刻地展示了本发明的新化合物在新的酶影响的背景下的潜力。同时还会促进继发性细胞凋亡,此举在分子水平上恢复细胞功能(参阅图11)。天然的2og重新进入正常的细胞代谢,从而再次抑制细胞的恶性脱轨。(由谷氨酸形成2og的次生途径消失)。用显著降低2-hg水平的强idh抑制剂ml309进行培养,不能激活idh1r132h/+细胞中被抑制的tet酶。通过添加功能性共基质dka来激活tet酶。证明取代天然共基质2og的本发明化合物的优点的另一个事实是,由此便能研发出新的化合物,其在突变的dios上也具有活性,并且在动力学和结合亲和力上与天然共基质不同。众所周知,idh1变异与有所变化的idh1酶功能相关,其可诱导新变态代谢物2-羟基戊二酸的过量生产。为了验证hct116idh1r132h/+细胞中与hct116idh1+/+细胞相比,2-hg的频率增加,借助于lc-ms/ms分析来分析2-hg含量(图1)。idh1r132h/+细胞中的细胞内2-hg比idh1野生型细胞中的细胞内2-hg高56倍(分别为8.5nmol/mg蛋白或0.15nmol/mg蛋白;高56倍;p<0.0001)。此外,用idh1抑制剂ml309(10μm)进行治疗可使hct116idh1r132h/+细胞中的肿瘤代谢物2-hg显著降低至(0.3nmol/mg蛋白),idh1+/+的水平降低(0.04nmol/mg蛋白)(附图)。相比之下,dka在突变体细胞中仅产生极小的2-hg浓度变化(5.9nmol/mg蛋白)。有趣的是,用10μmml309和1mmdka的组合治疗导致idh1r132h/+中2-hg的最大下降,并达到了与idh1野生型细胞相似的浓度(0.12nmol/mg蛋白),参阅图10。图11为idh1-mthct116中2-羟基戊二酸水平与5-hmdc水平的相关性。图12为在存在竞争性抑制剂2hg的情况下单凭将dka用作功能共基质来恢复tet酶的功能。图13-18列出可以用作tet和hif上共基质2og的功能替代的更多选定化合物在酶中的构象。为此,下文列出相应的细胞实验。图13为tet酶中的dka,图13中显示氨基酸as与哪个原子配位。图14为dka的空间位置,图15为dka在酶中的空间配位。图15中的箭头指向fe2+与dka的距离如图所示,dka在酶中处于ts并且在此也达到了规定的条件(图1),从而实现共基质2og的功能替代。图16示出二酮古洛糖酸(2okg)并且示出,2-okg如何在酶中处于ts并且在此也达到了规定的条件(图1),从而实现共基质2og的功能替代。图16b在测试中表明,2-ogk的效果与剂量相关。图17示出具有fe2+的4-甲基-5-氧代己-2-烯二酸,显示出进行相互作用的氨基酸。图18a中示出具有fe2+和氨基酸的aof(2-(呋喃-2-基)-2-氧代乙酸,图18b为其效果,图18c中示出酶与aof的相互作用。图19-27为针对hf酶的化合物。图19a中示出结构,在图19b中,在2g19中,2-(((羟基(4-羟基-8-碘异喹啉-2-鎓-3-基)亚甲基)氨基)乙酸盐与缺氧诱导因子(hypoxia-induciblefactor脯氨酰羟化酶,phd2)拮抗剂结合。图20示出酶phd2中的2og(根据homann法)。酶中的方位依据的是作为共基质功能的必要原则。图21中,nga被显示为酶中的拮抗剂。图22示出酶中的竞争性拮抗剂2hg,其在酶中与2og进行竞争性相互作用,导致酶的功能丧失。图23示出酶中作为功能性辅酶的2-okg。这项功能的必要原则得以保持。图24示出酶中作为功能性辅酶的3-溴-2-氧代戊酸酯。图25示出酶中作为功能性辅酶的(e)-5-氧代己-2-烯二酸。图26示出酶中作为功能性辅酶的4-(s)-甲基-5-氧代己-2-烯二酸。图27示出酶中作为功能性辅酶的dkg。实施例tet酶(ten-eleventranslocation(tet)酶)。在实例1(图28)中能够展现功能性的恢复,并且将致癌的表观遗传变化消除。此外,能够将细胞的正常新陈代谢再生,并且将癌症代谢(cancermetabolismus)中断。在与其他化学疗法(联合疗法)相结合的情况下,或单独凭借此举措,能够实现细胞功能的正常化,并防止退化细胞中的凋亡。肿瘤抑制基因(cdkn1a(p21))同样因功能恢复的tet而重新激活。图28示出dka–实例1中的2,3-二酮古洛糖酸和2og(2kg-2-酮-l-古洛糖酸:细胞培养和处理从americantypeculturecollection(atcc;https://www.atcc.org/)购得hct116(atcc编号ccl-247),人类结肠癌细胞系。人类结肠癌细胞hct116idh1+/+和hct116idh1r132h/+来自horizondiscovery,并且为了试验目的而被提供给我们。在包含2mml-谷氨酰胺并且添加有10%胎牛血清(fbs)、45iu/ml青霉素和45iu/ml链霉素的dulbecco'smodifiedeagle'smedium(dmem)中对所述细胞进行培养。在使用前的六个月内,所述细胞系的支原体感染测试结果为阴性。细胞活力测试如前文所述,借助mtt还原测定法对经测试的物质的可能存在的细胞毒素效应进行研究。将hct116细胞种入96孔板(tpp,特拉萨丁根,瑞士)。在24小时后,为24、48和72小时添加给定的浓度。随后以100μl的mtt溶液(0.5mg/ml,pbs中)对所述细胞进行为时4小时的培养。在将上清液移除后,添加50μl的二甲基亚砜,以将甲盐溶解,并且借助酶标仪(tecan,克赖尔斯海姆,德国)对光密度(od)进行测量。将激励调节至540nm。以0.002%sds对阳性对照进行处理。<75%的细胞活力预示了细胞毒性效应。细胞凋亡测试在使用ebiosciencetmannexinvapoptosisdetectionkitapc(thermofisher,达姆施塔特,德国)的情况下,借助流式细胞仪确定凋亡和死亡细胞的水平。以每孔2x105hct116细胞的密度,将细胞种入6孔板(tpp,特拉萨丁根,瑞士)。在24小时后,借助给定浓度的物质对细胞进行为时72小时的培养。随后对细胞进行洗涤,并按照制造商的指示用annexinv和碘化丙啶对细胞进行染色。在facscantoii(bdbiosciences,海德堡,德国)上对细胞进行分析。使用软件flowjo(treestar,阿什兰,美国)来进行数据分析。rt-pcr根据rnahighpurernakit(roche,曼海姆,德国)的说明提取rna,并且借助revertaidreversetranskriptase(thermofisher,达姆施塔特,德国)根据研究方案将rna中的0.5-5μg(理想情形下为3μg)逆转录。借助maximasybrgreenqpcrmix(thermofisher,达姆施塔特,德国)在lightcycler480ii实时pcr系统(roche,曼海姆,德国)上实施qrt-pcr。借助δδct方法进行定量,并且将gapdh表达用作内参。熔解曲线分析证实,所有qrt-pcr产物均以双链dna的形式产生。使用的引物在表3中列出。表3:借助同位素稀释法、液相色谱法、串联质谱法(lc-ms/ms)确定全基因组甲基化和羟甲基化借助来源于金黃色葡萄球菌的微球菌核酸酶、牛脾磷酸二酯酶和小牛肠碱性磷酸酶(均来自sigma-aldrich,陶夫基兴,德国)将基因组dna的试样(20μg)水解成2'-脱氧核苷,如前文[62]借助应用改性所述。将10μl的50nm5-hmdc-d3(torontoresearchchemicals,多伦多,加拿大)作为内标添加至dna酶解混合物,且两阶段水解的培养时间分别为1小时。随后对dna水解产物进行离心(5分钟,16000xg),并将上清液中的10μl用于dc以及5-mdc稳定标记参考的定量。化合物[15n2,13cl]dc和5-mdc-d3(两者均来自torontoresearchchemicals,多伦多,加拿大)。在有所减小的压力下,借助savantspeedvac浓缩仪(thermofisherscientific,德赖艾希,德国)将dna水解物的残余(~310μl)蒸发至干燥。在将100μl甲醇添加至经干燥的残余物和短暂的涡旋后,在-20℃下将试样放置一夜。在下一日对试样进行为时10分钟的充分涡旋(1400rev/min),随后在16000xg条件下进行为时10分钟的离心。将上清液转移到新的试管中。通过再添加100μl甲醇以及为时5分钟的离心(1400rev/min),重复提取蛋白质颗粒的操作。在16000xg条件下进行为时10分钟的离心后,将两个甲醇馏分合并,并在有所减小的压力下蒸发至干燥。在包含0.0075%甲酸的50μl水中将经干燥的残余物重构,对其进行为时10分钟的超声处理,随后进行为时5分钟的离心(1400rev/min),并且在16000xg条件下进行为时5分钟的离心。借助agilent1260infinitylc系统,结合以agilent6490三重串联四极杆质谱仪(两者均来自瓦尔德布龙,德国)对上清液进行lc-ms/ms分析,该仪器与处于正离子模式(esi+)下的电喷雾离子源连接。色谱条件和esi源设置与结合dc以及5-mdc的定量所作的描述[62]相同。将agilentporoshell120ec-c18(2.7μm,3.0x150mm)用作分离柱,其注射容积为5μl。借助多反应监测(multiplereactionmonitoring,mrm)方案,相对在4.9分钟内从lc柱洗提的稳定的同位素标记标准物(dc和5-mdc的保留时间为4.7或6.0分钟)对5-hmdc进行定量。其中将以下质量转移(2'-脱氧核糖的损耗)用作定量(括号中为经优化的碰撞能量):5-hmdc:m/z258.1>142.0(8ev)和5-hmdc-d3:m/z261.1>145.0(8ev)。为了实现明确识别,将额外的质量转移记录。针对分析的四个质量转移中的每一个的停留时间均为50ms。对人类idh1(r132h/+)hct116细胞系进行的试验图29示出mtt细胞毒性测试的结果,此测试如下实施:·以浓度增大(100μm-10mm)的dka进行24小时、48小时和72小时的培养·结肠癌细胞系hct116,具有杂合突变idh1-(r132h/+)-mt和hct116-idh1-wt图30示出细胞凋亡测试的结果,此测试如下实施:·以浓度增大(100μm-10mm)的dka进行72小时的培养·结肠癌细胞系hct116,具有杂合突变idh1-(r132h/+)-mt和hct116-idh1-wt统计:双因素方差分析,dunnett修正-****=p<0.0001图31示出另一细胞凋亡测试的结果,此测试如下实施:·以浓度增大(100μm-10mm)的dka进行48小时的培养+24小时tnfα(10ng/ml)/chx(1μg/ml)·结肠癌细胞系hct116,具有杂合突变idh1-(r132h/+)-mt和hct116-idh1-wt统计:双因素方差分析,dunnett修正-****=p<0.0001;***=p<0.001图32示出hmdc测量的结果(不同的显示),所述测量如下实施:·以浓度增大(100μm-10mm)的dka进行72小时的培养·结肠癌细胞系hct116,具有杂合突变idh1-(r132h/+)-mt和hct116-idh1-wt统计:双因素方差分析,dunnett修正-****=p<0.0001;*=p<0.05图33示出hmdc测量的结果(不同的显示),所述测量如下实施:hmdc测量(未经处理的对照充当100%,其他显示采用相对此对照的%)·以浓度增大(10μm-1mm)的dka进行72小时的培养·结肠癌细胞系hct116,具有杂合突变idh1-(r132h/+)-mt和hct116-idh1-wt统计:双因素方差分析,dunnett修正-****=p<0.0001;*=p<0.05图34示出采用hct116细胞的蛋白质印迹法,其如下实施:·以浓度增大(100μm-1mm)的dka进行72小时的培养·结肠癌细胞系hct116,具有杂合突变idh1-(r132h/+)-mt和hct116-idh1-wt这些示例表明,用诸如dkg的无毒示例物质进行共基质替代和调制的请求是成功的。诸如继发性凋亡的效应表明细胞代谢的重构。通过增大或改变肿瘤细胞的能量需求,这些肿瘤细胞不再能够从正常的代谢循环生成这个能量需求,故通过治疗进入继发性凋亡。tet的活性产生导致胞嘧啶的去甲基化,以及导致与肿瘤抑制基因再生相关的表观遗传变化消除。所有这些都证明了共基质替代(调制因子)具备影响dios和与之相关的各种病理状态的能力。人类的dios,尤其是调控转录的那一些,是目前针对各种贫血症和癌症中的治疗靶点的研究方法的主题。[63-66]dios影响和调控着大量蛋白质,从本文开篇揭示的数个反应便能推知这一点。[67]genbankdna数据库中的已知和推定的2og依赖性二氧酶参见表4。根据本发明,考虑对这些进行调制。dna/rnajmjc域jmjc域脯氨酸/赖氨酸其他改性包含包含羟化酶羟化酶tet1kdm2akdm7aegln1asphtet2kdm2bkdmxegln2asphd1tet3kdm3ahregln3asphd2abh1kdm3bjarid2p4ha1bbox1abh2kdm4ajhdm1cp4ha2fih1abh3kdm4bjmjd1cp4ha3hspbap1abh4kdm4cjmjd4p4hbogfod1abh5kdm4djmjd6p4htmogfod2abh6kdm5ajmjd7plod1pahx-ftokdm5bjmjdxplod2phyhkdm5cminaplod3phyhd1kdm5dno66lepre1kdm6aphf2leprel1kdm6bphf8leprel2utybbox2表4参考1.flashman,e.和c.j.schofield,themostversatileofallreactiveintermediates?naturechemicalbiology,2007.3:第86页。2.hausinger,r.p.,fe(ⅱ)/α-ketoglutarate-dependenthydroxylasesandrelatedenzymes.criticalreviewsinbiochemistryandmolecularbiology,2004.39(1):第21-68页。3.perry,c.等人,rieskenon-hemeiron-dependentoxygenasescatalysediversereactionsinnaturalproductbiosynthesis.naturalproductreports,2018.35(7):第622-632页。4.loenarz,c.和c.j.schofield,physiologicalandbiochemicalaspectsofhydroxylationsanddemethylationscatalyzedbyhuman2-oxoglutarateoxygenases.trendsinbiochemicalsciences,2011.36(1):第7-18页。5.prescott,a.g.和m.d.lloyd,theiron(ⅱ)and2-oxoacid-dependentdioxygenasesandtheirroteinmetabolism.naturalproductreports,2000.17(4):第367-383页。6.scotti,j.s.等人,humanoxygensensingmayhaveoriginsinprokaryoticelongationfactortuprolyl-hydroxylation.proceedingsofthenationalacademyofsciences,2014.111(37):第13331-13336页。7.clifton,i.j.等人,crystalstructureofcarbapenemsynthase(carc).journalofbiologicalchemistry,2003.278(23):第20843-20850页。8.farrow,s.c.和p.j.facchini,functionaldiversityof2-oxoglutarate/fe(ⅱ)-dependentdioxygenasesinplantmetabolism.frontiersinplantscience,2014.5:第524页。9.cheng,a.x.等人,thefunctionandcatalysisof2-oxoglutarate-dependentoxygenasesinvolvedinplantflavonoidbiosynthesis.intjmolsci,2014.15(1):第1080-95页。10.zhang,z.等人,crystalstructureandmechanisticimplicationsof1-aminocyclopropane-l-carboxylicacidoxidase—theethylene-formingenzyme.chemistry&biology,2004.11(10):第1383-1394页。11.myllyharju,j.,prolyl4-hydroxyiases,thekeyenzymesofcollagenbiosynthesis.matrixbiology,2003.22(1):第15-24页。12.leung,i.k.h.等人,structuralandmechanisticstudiesonγ-butyrobetainehydroxylase.chemistry&biology,2010.17(12):第1316-1324页。13.markolovic,s.,s.e.wilkins,和c.j.schofield,proteinhydroxylationcatalyzedby2-oxoglutarate-dependentoxygenases.journalofbiologicalchemistry,2015.290(34):第20712-20722页。14.walport,l.j.,r.j.hopkinson,和c.j.schofield,mechanismsofhumanhistoneandnucleicaciddemethylases.currentopinioninchemicalbiology,2012.16(5):第525-534页。15.salminen,a.,a.kauppinen,和k.kaarniranta,2-oxoglutarate-dependentdioxygenasesaresensorsofenergymetabolism,oxygenavailability,andironhomeostasis:potentialroleintheregulationofagingprocess.cellularandmolecularlifesciences,2015.72(20):第3897-3914页。16.martinez,s.和r.p.hausinger,catalyticmechanismsoffe(ⅱ)-and2-oxoglutarate-dependentoxygenases.journalofbiologicalchemistry,2015.290(34):第20702-20711页。17.r.等人,ascorbateisconsumedstoichiometricallyintheuncoupledreactionscatalyzedbyprolyl4-hydroxylaseandlysylhydroxylase.journalofbiologicalchemistry,1984.259(9):第5403-5405页。18.flashman,e.等人,investigatingthedependenceofthehypoxia-induciblefactorhydroxylases(factorinhibitinghifandprolylhydroxylasedomain2)onascorbateandotherreducingagents.biochemj,2010.427(1):第135-42页。19.lappin,t.和n.masson,twoantioxidantsarebetterthanone.blood,2011.117(20):第5276页。20.mastrangelo,d.等人,mechanismsofanti-cancereffectsofascorbate:cytotoxicactivityandepigeneticmodulation.bloodcells,molecules,anddiseases,2018.69:第57-64页。21.shenoy,n.等人,ascorbicacidincancertreatment:letthephoenixfly.cancercell,2018。22.solomon,e.i.,a.decker,和n.lehnert,non-hemeironenzymes:contraststohemecatalysis.proceedingsofthenationalacademyofsciences,2003.100(7):第3589页。23.huang,c.w.等人,thedifferentcatalyticrolesofthemetal-bindingligandsinhuman4-hydroxyphenylpyruvatedioxygenase.biochemj,2016.473(9):第1179-89页。24.hewitson,k.s.等人,oxidationby2-oxoglutarateoxygenases:non-haemironsystemsincatalysisandsignalling.philosophicaltransactionsoftheroyalsocietya:mathematical,physicalandengineeringsciences,2005.363(1829):第807页。25.raili,m.,t.leena,和k.k.i.,mechanismoftheprolylhydroxylasereaction.europeanjournalofbiochemistry,1977.80(2):第349-357页。26.price,j.c.等人,thefirstdirectcharacterizationofahigh-valentironintermediateinthereactionofana-ketoglutarate-dependentdioxygenase:ahigh-spinfe(iv)complexintaurine/α-ketoglutaratedioxygenase(taud)fromescherichiacoli.biochemistry,2003.42(24):第7497-7508页。27.proshlyakov,d.a.等人,directdetectionofoxygenintermediatesinthenon-hemefeenzymetaurine/α-ketoglutaratedioxygenase.journaloftheamericanchemicalsociety,2004.126(4):第1022-1023页。28.welford,r.w.等人,incorporationofoxygenintothesuccinateco-productofiron(ⅱ)and2-oxoglutaratedependentoxygenasesfrombacteria,plantsandhumans.febslett,2005.579(23):第5170-4页。29.grzyska,p.k.等人,insightintothemechanismofanirondioxygenasebyresolutionofstepsfollowingthefelv=hospecies.procnatlacadsciusa,2010.107(9):第3982-7页。30.valegird,k.等人,thestructuralbasisofcephalosporinformationinamononuclearferrousenzyme.naturestructural&;molecularbiology,2003.11:第95页。31.wiek,c.r.等人,structuralinsightintotheprolylhydroxylasephd2:amoleculardynamicsanddftstudy.europeanjournalofinorganicchemistry,2012.2012(31):第4973-4985页。32.tarhonskaya,h.等人,studiesondeacetoxycephalosporincsynthasesupportaconsensusmechanismfor2-oxoglutaratedependentoxygenases.biochemistry,2014.53(15):第2483-2493页。33.mcdonough,m.a.等人,structuralstudiesonhuman2-oxoglutaratedependentoxygenases.currentopinioninstructuralbiology,2010.20(6):第659-672页。34.clifton,i.j.等人,structuralstudieson2-oxoglutarateoxygenasesandrelateddouble-strandedbeta-helixfoldproteins.jinorgbiochem,2006.100(4):第644-69页。35.l.,h.e.和j.l.que,the2-his-l-carboxylatefacialtriad—anemergingstructuralmotifinmononuclearnon-hemeiron(ⅱ)enzymes.europeanjournalofbiochemistry,1997.250(3):第625-629页。36.chowdhury,r.等人,structuralbasisforbindingofhypoxia-induciblefactortotheoxygen-sensingprolylhydroxylases.structure,2009.17(7):第981-9页。37.deng,g.等人,novelcomplexcrystalstructureofprolylhydroxylasedomain-containingprotein2(phd2):2,8-diazaspiro[4.5]decan-l-onesaspotent,orallybioavailablephd2inhibitors.bioorgmedchem,2013.21(21):第6349-58页。38.chowdhury,r.等人,structuralbasisforoxygendegradationdomainselectivityofthehifprolylhydroxylases.naturecommunications,2016.7:第12673页。39.willam,c.等人,theprolylhydroxylaseenzymesthatactasoxygensensorsregulatingdestructionofhypoxia-induciblefactoralpha.advenzymeregul,2004.44:第75-92页。40.rabe,p.等人,rolesof2-oxoglutarateoxygenasesandisopenicillinnsynthasein6-lactambiosynthesis.naturalproductreports,2018。41.hegg,e.l.和l.q.jr,the2-his-l-carboxylatefacialtriad—anemergingstructuralmotifinmononuclearnon-hemeiron(ⅱ)enzymes.europeanjournalofbiochemistry,1997.250(3):第625-629页。42.kal,s.和l.que,dioxygenactivationbynonhemeironenzymeswiththe2-his-l-carboxylatefacialtriadthatgeneratehigh-valentoxoironoxidants.jbicjournalofbiologicalinorganicchemistry,2017.22(2):第339-365页。43.xu,w.等人,oncometabolite2-hydroxyglutarateisacompetitiveinhibitorofα-ketoglutarate-dependentdioxygenases.cancercell,2011.19(1):第17-30页。44.xu,w.等人,oncometabolite2-hydroxygiutarateisacompetitiveinhibitorofalpha-ketoglutarate-dependentdioxygenases.cancercell,2011.19(1):第17-30页。45.elkashef,s.m.等人,idhmutation,competitiveinhibitionoffto,andrnamethylation.cancercell,2017.31(5):第619-620页。46.flavahan,w.a.等人,insulatordysfunctionandoncogeneactivationinidhmutantgliomas.nature,2016.529(7584):第110-4页。47.buege,m.j.,a.j.dipippo,和c.d.dinardo,evolvingtreatmentstrategiesforelderlyleukemiapatientswithidhmutations.cancers(basel),2018.10(6)。48.rautio,j.等人,theexpandingroleofprodrugsincontemporarydrugdesignanddevelopment.natrevdrugdiscov,2018.17(8):第559-587页。49.vangorkom,g.等人,influenceofvitaminconlymphocytes:anoverview.antioxidants,2018.7(3):第41页。50.cimmino,l.,b.g.neel,和i.aifantis,vitamincinstemcellreprogrammingandcancer.trendscellbiol,2018。51.li,x.等人,ten-eleventranslocation2demethylatesthemmp9promoter,anditsdown-regulationinpreeclampsiaimpairstrophoblastmigrationandinvasion.jbiolchem,2018。52.sant,d.w.等人,vitamincpromotesapoptosisinbreastcancercellsbyincreasingtrauexpression.scientificreports,2018.8(1):第5306页。53.park,s.等人,vitamincincancer:ametabolomicsperspective.frontiersinphysiology,2018.9(762)。54.monacelli,f.等人,vitaminc,agingandalzheimer'sdisease.nutrients,2017.9(7)。55.ebata,k.t.等人,vitamincinducesspecificdemethylationofh3k9me2inmouseembryonicstemcellsviakdm3a/b.epigenetics&chromatin,2017.10(1):第36页。56.d'aniello,c.等人,vitamincandi-prolineantagonisticeffectscapturealternativestatesinthepluripotencycontinuum.stemcellreports,2017.8(1):第1-10页。57.schonberger,k.和n.cabezas-wallscheid,vitaminc:c-inganewwaytofightleukemia.cellstemcell,2017.21(5):第561-563页。58.unlu,a.等人,high-dosevitamincandcancer.journalofoncologicalscience,2016.1:第10-12页。59.li,r.,vitaminc,amulti-taskingmolecule,findsamoleculartargetinkillingcancercells.reactoxygspecies(apex),2016.1(2):第141-156页。60.levine,m.和p.-c.violet,datatriumphatc.cancercell,2017.31(4):第467-469页。61.monfort,a.和a.wutz,breathing-inepigeneticchangewithvitaminc.emborep,2013.14(4):第337-46页。62.schumacher,f.等人,optimizedenzymatichydrolysisofdnaforlc-ms/msanalysesofadductsofl-methoxy-3-indolylmethylglucosinolateandmethyleugenol.analbiochem,2013.434(1):第4-11页。63.islam,m.s.等人,2-oxoglutarate-dependentoxygenases.annualreviewofbiochemistry,2018.87(1):第585-620页。64.lv,x.等人,theroleofhypoxia-induciblefactorsintumorangiogenesisandcellmetabolism.genes&diseases,2017.4(1):第19-24页。65.wigerup,c.,s.pahlman,和d.bexell,therapeutictargetingofhypoxiaandhypoxia-induciblefactorsincancer.pharmacolther,2016.164:第152-69页。66.xia,y.,h.k.choi,和k.lee,recentadvancesinhypoxia-induciblefactor(hif)-linhibitors.eurjmedchem,2012.49:第24-40页。67.semenza,g.l.,acompendiumofproteinsthatinternetwithhif-lalpha.expcellres,2017.356(2):第128-135页。序列表<110>托马斯·梅尔希奥·霍曼(homann,thomas,melchior)<120>用于调制α-酮戊二酸(2og)依赖型加氧酶并作为其官能替代的化合物<130>80405wo<150>lu100900<151>2018-08-10<160>20<170>patentinversion3.5<210>1<211>20<212>dna<213>人工序列<220><223>正向引物hhmbs<220><221>misc_feature<223>正向引物hhmbs<400>1accaaggagcttgaacatgc20<210>2<211>21<212>dna<213>人工序列<220><223>反向引物hhmbs<220><221>misc_feature<223>反向引物hhmbs<400>2gaaagacaacagcatcatgag21<210>3<211>20<212>dna<213>人工序列<220><223>正向引物hdnmt1<220><221>misc_feature<223>正向引物hdnmt1<400>3acctggctaaagtcaaatcc20<210>4<211>20<212>dna<213>人工序列<220><223>反向引物hdnmt1<220><221>misc_feature<223>反向引物hdnmt1<400>4attcacttcccggttgtaag20<210>5<211>20<212>dna<213>人工序列<220><223>正向引物hdnmt3a<220><221>misc_feature<223>正向引物hdnmt3a<400>5actacatcagcaagcgcaag20<210>6<211>20<212>dna<213>人工序列<220><223>反向引物hdnmt3a<220><221>misc_feature<223>反向引物hdnmt3a<400>6catccaccaagacacaatgc20<210>7<211>20<212>dna<213>人工序列<220><223>正向引物hdnmt3b<220><221>misc_feature<223>正向引物hdnmt3b<400>7ccagctcttaccttaccatc20<210>8<211>20<212>dna<213>人工序列<220><223>反向引物hdnmt3b<220><221>misc_feature<223>反向引物hdnmt3b<400>8cagacatagcctgtcgcttg20<210>9<211>20<212>dna<213>人工序列<220><223>正向引物htet1<220><221>misc_feature<223>反向引物htet1<400>9gctgctgtcagggaaatcat20<210>10<211>20<212>dna<213>人工序列<220><223>反向引物htet1<220><221>misc_feature<223>反向引物htet1<400>10accatcacagcagttggaca20<210>11<211>20<212>dna<213>人工序列<220><223>正向引物htet2<220><221>misc_feature<223>正向引物htet2<400>11ccaataggacatgatccagg20<210>12<211>20<212>dna<213>人工序列<220><223>反向引物htet2<220><221>misc_feature<223>反向引物htet2<400>12tctggatgagctctctcagg20<210>13<211>20<212>dna<213>人工序列<220><223>正向引物htet3<220><221>misc_feature<223>正向引物htet3<400>13tcggagacaccctctaccag20<210>14<211>20<212>dna<213>人工序列<220><223>反向引物htet3<220><221>misc_feature<223>反向引物htet3<400>14cttgcagccgttgaagtaca20<210>15<211>18<212>dna<213>人工序列<220><223>正向引物cdkn2a(p16)<220><221>misc_feature<223>正向引物cdkn2a(p16)<400>15gagcagcatggagccttc18<210>16<211>20<212>dna<213>人工序列<220><223>反向引物cdkn2a(p16)<220><221>misc_feature<223>反向引物cdkn2a(p16)<400>16cctccgaccgtaactattcg20<210>17<211>20<212>dna<213>人工序列<220><223>正向引物cdkn1a(p21)<220><221>misc_feature<223>正向引物cdkn1a(p21)<400>17agtggacagcgagcagctga20<210>18<211>23<212>dna<213>人工序列<220><223>反向引物cdkn1a(p21)<220><221>misc_feature<223>反向引物cdkn1a(p21)<400>18tagaaatctgtcatgctggtctg23<210>19<211>23<212>dna<213>人工序列<220><223>正向引物cdkn1b(p27)<220><221>misc_feature<223>正向引物cdkn1b(p27)<400>19aaacgtgcgagtgtctaacggga23<210>20<211>23<212>dna<213>人工序列<220><223>反向引物cdkn1b(p27)<220><221>misc_feature<223>反向引物cdkn1b(p27)<400>20cgcttccttattcctgcgcattg23当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1