一种蝴蝶兰开花基因PhalLFY及其应用的制作方法
本发明属于分子生物学、基因工程技术领域,具体涉及一种在蝴蝶兰中表达的花分生组织决定基因—PhalLFY基因、含有该基因的重组植物表达载体,以及PhalLFY基因在促进植物多花枝发育和促进植物提前开花中的应用
背景技术:
:
在植物个体发育中,花的分化是生殖生长的开始。首先,营养分生组织转变为花序分生组织,然后花序分生组织转变为花分生组织,接下来,花分生组织产生花器官原基,最后产生各种花器官。已被确定的花分生组织决定基因有LEAFY(LFY),APETALA1(AP1),CAULIFL OWER(CAL),APETALA2(AP2),and UNUSUAL FLORAL ORGANS(UFO)(Weigel et al.,1992;Yanofsky,1995),其中,LFY起着中心调控作用,在植物花发育中发挥着重要作用(Weigel et al.,1992;Pidkowich et al.,1999)。
已经有一些FLO/LFY同源基因从不同的物种中被克隆出来,LFY同源基因编码的氨基酸在C-末端区域内高度保守,而N-末端不保守(Southerton et al.,1998;Mouradov et al.,1998;Wada et al.,2002)。它们的结构也有所不同,氨基酸序列中大部分含有5’-N端脯氨酸富集区、中央酸性区和亮氨酸拉链结构(XEL…XTL…XEL…)特征,拟南芥中还含有锌指结构,这些结构具有转录因子的特征。所有已知的花分生组织决定基因所编码的蛋白质都参与调节下游基因的表达。但是LFY编码的产物和其它任何已知的转录因子没有相似性,LFY蛋白质被显示优先定位于核内,能够在活体内结合DNA(Weigel,1995;Weigel and Meyerowitz,1993)。当LFY融合一个不同的激活域时,它能在酵母中调节转录活性(Parcy et al.,1998),因此,LFY有转录因子的功能。
所有的花分生组织决定基因都在幼嫩的花原基中表达。然而,LFY不仅在花原基中表达,它也在营养器官中表达,在营养阶段,LFY的表达在幼嫩的叶原基中被发现,它的表达在芽分生组织中增加。LFY在其它花分生组织决定基因之前表达,且随着花的诱导,表达会增加。在营养阶段,过量表达LFY导致早花。在ful/ap1/cal三突变体中,不能开花,而过量表达LFY可以修复这种突变体植物的不开花表型(Ferrandiz et al.,2000)。这个结果暗示LFY在开花转变中起着关键性作用。然而,LFY本身并不能立即充分地将SAM转变成花分生组织,没有其它基因的上游调控,LFY的表达不能到达能成功启动开花的域值。对LFY同源基因的时空表达研究发现,不同植物中其表达的时间和空间有所不同。金鱼草中的FLO基因最早是在花分生组织中表达,而后是萼片、花瓣和心皮原基,而在雄蕊原基中没有表达,FLO只在植物的开花期表达(Coen,1990)。但在拟南芥、烟草、豌豆、凤仙花、矮牵牛、葡萄、桉树、山杨和番茄等植物中,LFY同源基因除了在开花阶段有表达外,在叶原基中也有表达(Kelly et al.,1995;Blazquez et al.,1997;Hofer et al.,1997;Pouteau et al.,1997;Souer et al.,1998;Ca rmona et al.,2002;Southerton et al.,1998;Rottmann et al.,2002;Andrea et al.,2003;Do rnelas et al.,2004)。在豌豆的叶子中LFY同源基因(UNIFOLIATA)mRNA具有很强的表达(Hofer et al.,1997);苹果中的AFL2在根、叶及花分生组织中均有很强的表达(Wada et al.,2002);辐射松的2个同源基因中,NLY在营养生长时期表达,PRFLL在雄球果中表达(Mellerowicz et a1.,1998;Mouradov et al.,1998)。单子叶植物水稻中LFY的同源基因RFL的表达明显区别于双子叶植物,RFL基因在幼穗中表达,而在成熟的小花和叶子中没有表达(Kyozuka et al.,1998)。LFY同源基因这种表达特性上的差异很可能反映植物本身花结构的进化关系。
不同植物中LFY同源基因转基因植株的表达效果并不同。拟南芥LFY基因转入其本身后,转基因植株的侧芽全部转变为花芽,使花期提前(Blazquez et a1.,1997)。转LFY基因的杨树、烟草、菊花、水稻及柑橘等植物也具有同样的效果。把杨树中LFY同源基因导入拟南芥,转基因植株花期提前,但杨树自身的转基因植株开花时间变化不明显(Rottmann et a1.,2000),这可能与其幼年期较长有关。转入辐射松NLY基因的拟南芥对日长不敏感,均提前开花(Mour adov et a1.,1998)。但烟草NFL1基因在转入拟南芥后不能使花期提前(Ahearn et a1.,2001)。上述情况说明尽管LFY同源基因所编码的蛋白序列很保守,但在进化过程中,由于一些结构上的改变,导致其功能也发生了变化。
蝴蝶兰是兰科植物中栽培最广泛、最受欢迎的种类之一,花形似蝴蝶,别致美丽、色彩艳丽,花期长达数月之久,观赏价值极高,在热带兰中素有“兰花皇后”之美誉,国内外一直视其为花中珍品,倍受人们青睐。在我国,蝴蝶兰消费主要集中在春节、中秋、国庆、元旦等重大节日,生产上确保蝴蝶兰如期上市非常重要,花期调控是生产中的关键环节之一。影响蝴蝶兰生长发育与开花的因子有肥料、基质、温度、光照、激素等,生产上主要通过调控这几个因子来实现对蝴蝶兰的花期调控。开花受环境因子和内源因子共同调控,调节开花的环境信息和内源信息必须通过花分生组织决定基因发挥作用,花的命运最终被花分生组织决定基因所确定。LFY是最关键的花分生组织决定基因,它直接决定植物开花时间和开花状态。
技术实现要素:
:
本发明的第一个目的是提供一种在蝴蝶兰中表达的花分生组织决定基因—蝴蝶兰开花基因PhalLFY及其编码蛋白和含有该基因的重组植物表达载体。
本发明研究了一种从蝴蝶兰中克隆出的PhalLFY基因,将其转入拟南芥和烟草后可以促进侧芽或腋芽转化为花芽,导致多花枝发育,并促进提前开花。而在发明公布之前,尚未有任何公开报道过本专利申请中提及的蝴蝶兰开花基因PhalLFY在转基因植株中的功能应用。
本发明的第二个目的是提供了一种含有上述蝴蝶兰开花基因PhalLFY的重组植物表达载体。
优选,所述的重组植物表达载体中的载体为pBI系列载体、pBin系列载体、GatewayTW系列载体或pCAMBIA系列载体。
本发明的第三个目的是提供一种含有所述的重组植物表达载体的植物细胞系。
本发明的第四个目的是提供一种含有所述的重组植物表达载体的植物转化体。
优选,所述的植物转化体中的受体植物为烟草或拟南芥。
本发明的第五个目的是提供蝴蝶兰开花基因PhalLFY在促进侧芽或腋芽发育成花芽,促进花芽发育,缩短植物童期,促进植物提前开花中的应用,如促进拟南芥和烟草多花芽发育,缩短烟草和拟南芥童期,促进它们提前开花。
本发明以“蝴蝶兰(Phalaenopsis hybrida cv.Wedding Promenade)”的花芽为材料,提取其总RNA,再将mRNA逆转录成cDNA第一链,以该cDNA第一链模板,根据LFY的同源基因设计引物,利用RT-PCR和RACE的方法扩增拼接获得全长cDNA序列,该cDNA序列长度为1655bp,其序列如SEQ ID NO.1所示,其开放阅读框为自5’端第41位至第1462位碱基,共1422bp,将此开放阅读框命名为蝴蝶兰开花基因PhalLFY,其编码的蛋白的氨基酸序列具有473个氨基酸残基,其序列如SEQ ID NO.2所示,将该蛋白命名为PhalLFY蛋白。
将蝴蝶兰开花基因PhalLFY编码的蛋白质-PhalLFY蛋白的序列用ClustalW2分析其同源性,结果显示PhalLFY蛋白在N-和C-末端都有两个保守的结构域,它包含亮氨酸重复区域,酸性区域,碱性区域;氨基酸序列的同源性分析显示PhalLFY分别与倒距兰LFY的同源性为68%、与红门兰LFY的为67%、与风信子LFY-like的为61%,与水稻RFL为的60%、与百合LFY1的为59%、与金鱼草FLO的为59%、与拟南芥LFY的为54%。进化树显示所有的LFY/FLO同源基因被分成双子叶和单子叶两个分支,PhalLFY属于单子叶分支。
本发明人将上述PhalLFY基因(蝴蝶兰开花基因PhalLFY)与植物表达载体连接,通过农杆菌介导,将含有PhalLFY基因的重组植物表达载体转化到拟南芥中,经筛选培养获得含有PhalLFY基因的转基因拟南芥。通过实验发现,PhalLFY基因超表达促进拟南芥提前开花,促进腋芽分化成花芽,出现多花枝现象;该转基因拟南芥与野生型拟南芥相比,转基因拟南芥花期提前,腋芽分化成花芽,花枝增多,10个转化株的平均开花时间为15天,有3个花枝,而野生型的平均开花时间为26天,仅有1个花枝。
本发明人将上述PhalLFY基因与植物表达载体连接,利用农杆菌介导的转化技术,将含有PhalLFY基因的重组植物表达载体转化到烟草中,经筛选培养获得含有PhalLFY基因的转基因烟草,PhalLFY基因超表达改变了烟草的生长发育进程,促进了花枝发育,并促使烟草提前开花。
因此可以将本发明的蝴蝶兰开花基因PhalLFY应用到缩短植物童期,促进植物开花中。本发明的PhalLFY基因与植物表达载体连接,通过农杆菌介导进入植物细胞、组织或器官,通过组织培养成转基因的植物转化体,从而实现缩短植物童期生长期,促进植物开花。所述的植物表达载体可为任意一种可用于根瘤农杆菌或发根农杆菌转化植物的双元载体或可用于植物微弹轰击的载体等,如pBI系列载体、pBin系列载体、GatewayTW系列载体、pCAMBIA系列载体或其它衍生植物表达载体等,上述载体均为公开销售的商品。
本发明首次从蝴蝶兰中克隆到LFY同源基因—PhalLFY基因(蝴蝶兰开花基因PhalLFY),并且通过实验证明该基因能够影响植物生长发育进程,缩短了童期,促使植物提前开花,并促进腋芽/侧芽发育成花芽。因此,本发明提供的蝴蝶兰开花基因PhalLFY为基因工程改良植物的生长进程,对植物的花期进行调控提供了一种有效的技术手段,具有广泛的应用前景和极大的经济价值。
附图说明:
图1是蝴蝶兰PhalLFY蛋白的氨基酸序列与LFY/FLO同源基因氨基酸序列比对。OrcLFY(BAC 54955)、AnaLFY(BAC55081),DacLFY(BAC55083),LFY-like(AAS00458),RFL(BAA21547),LiLFY1(ABR13015),LFY(NP_200993),BraLFY(ABF06559)。具有一致性和相似性的氨基酸分别用黑色和灰色阴影表示。酸性水解域用黑色线在序列下注明,碱性域用灰色线在序列下注明。暗示重复的亮氨酸残基
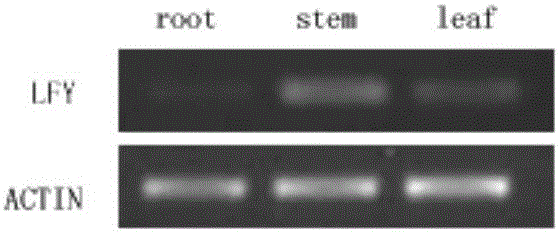
图2是RT-PCR检测在蝴蝶兰营养生长时期PhalLFY基因在植株根、茎、叶中的表达模式,其中root为根;stem为茎;leaf为叶;
图3是RT-PCR检测在蝴蝶兰生殖生长期当花梗长度达1cm时PhalLFY基因在植株根、茎、叶、花梗中的表达模式,其中root为根;stem为茎;scape为花梗;leaf为叶;
图4是转基因(PhalLFY基因)拟南芥的PCR鉴定。其中M为Marker 2000,WT为野生型拟南芥、-为转空载体质粒pBI121的拟南芥。1-6为筛选到的PhalLFY转基因拟南芥株系编号。
图5是转基因拟南芥(35S::PhalLFY)和野生型拟南芥的(WT)开花表型比较。转基因拟南芥明显比野生型拟南芥提前开花,且转基因拟南芥的腋芽多分化为花芽。
图6是转基因(PhalLFY)烟草的PCR鉴定。其中M为Marker 2000,WT为野生型烟草、-为转空载体质粒pBI121的烟草。1-6为筛选到的PhalLFY转基因烟草株系编号。
图7是转基因烟草(35S::PhalLFY)和野生型烟草的(WT)开花表型比较。右图的转基因烟草盛花后,左图的野生型烟草还没有萌发花芽。转基因烟草明显比野生型烟草提前开花,且转基因烟草的腋芽多已分化为花芽。
具体实施方式:
下面结合具体实施例进一步阐释本发明。应理解,这些实施例仅以用于说明本发明而不用于限制本发明的范围。下列实施例中未注明具体的实验方法,均可按照常规方法进行。如J.萨姆布鲁克等《分子克隆实验指南》、F.奥斯伯等《精编分子生物学实验指南》中所述条件,或按照所用产品生产厂商的使用说明。
实施例1:
一、蝴蝶兰PhalLFY基因的克隆及其同源性分析。
(1)以蝴蝶兰(Phalaenopsis hybrida cv.Wedding Promenade)的花芽为实验材料,材料在温室正常条件下生长(12h/12h;25-28℃)。
(2)RNA提取:用Trizol reagent(购自Invitrogen公司)进行试验材料的总RNA提取,整个操作过程严格按照Trizol试剂的RNA提取流程说明,然后再以该总RNA为模板逆转录得到cDNA第一链。
(3)基因的克隆:以逆转录的蝴蝶兰花芽cDNA第一链为模板,利用引物LFY-F和LFY-R进行PCR扩增,回收PCR产物,测序,获得600bp的核心片段。
LFY-F:5’-CGGAYATIAAYAARCCIAARATGMGICAYTA-3’
LFY-R:5’-CGACGTGICKIARIYKIGTIGGIACRTACCA-3’
利用该600bp的核心片段,根据RACE方法,设计扩增3’端序列的引物PR1和PR2,以及扩增5’端序列的引物PF1和PF2,引物为:
PF1:5'-TACAAGCCCCTCGTCGCCATCT-3'
PF2:5'-GACATCGACGCCGTCTTCA-3'
PR1:5'-GGCAGAGCTGACGGAGTTTGGTG-3'
PR2:5'-GGACGTAGTGTCGCATCTTAGGCTTGTT-3'.
PCR扩增获得5’端和3’端序列,最后与核心片段拼接获得全长cDNA序列,其序列如SEQ ID NO.1所示,该序列长度为1655bp,该序列的开放阅读框为SEQ ID NO.1的自5’端第41位至第1462位碱基,共1422bp,将该开放阅读框命名为PhalLFY基因(蝴蝶兰开花基因PhalLFY),该基因编码由473个氨基酸残基组成的多肽,命名为PhalLFY蛋白,其氨基酸序列如SEQ ID NO.2所示。将蝴蝶兰的PhalLFY基因编码的氨基酸序列用ClustalW2分析其同源性,结果显示PhalLFY蛋白的氨基酸结构非常保守,在N-和C-末端都有两个保守的结构域,它包含亮氨酸重复区域,酸性区域,碱性区域;氨基酸序列的同源性分析显示PhalLFY蛋白分别与倒距兰LFY的同源性为68%、与红门兰LFY的为67%、与风信子LFY-like的为61%,与水稻RFL为的60%、与百合LFY1的为59%、与金鱼草FLO的为59%、与拟南芥LFY的为54%(见图1)。进化树显示所有的LFY/FLO同源基因被分成双子叶和单子叶两个分支,PhalLFY属于单子叶分支。
实施例2
蝴蝶兰开花基因PhalLFY的基因表达模式分析
在蝴蝶兰营养生长期和生殖生长时期,取营养生长期的根、茎、叶以及生殖生长期的根、茎、叶、和花梗器官,用Trizol reagent(Invitrogen)分别提取这些器官的总RNA,测定OD260后定量取出2μgRNA,然后再使用oligo-dT引物和PrimeScriptTM Reverse Transcriptase(购自Takara公司)进行逆转录反应(方法参考说明书),以特异引物LF:5'-ATCCCCGCCTCTCAATCT-3'和LR:5'-TCTCATCCGCACCAGTTC-3'.进行PCR扩增,反应产物以1.0%琼脂糖凝胶电泳分离,电泳结果如图2和3所示,PhalLFY基因在蝴蝶兰各器官中均有表达,营养生长阶段,PhalLFY基因在各个器官中的表达都非常弱(图2);生殖生长阶段,在根中的表达很弱,但在茎中的表达水平明显提高(图3)。
实施例3
蝴蝶兰PhalLFY基因在拟南芥中的表达及转基因植株的表型鉴定。
(1)含目的基因PhalLFY表达载体的构建
设计引物PhLFYF 5’AATTCTAGAATGGACCCAAGCGACGCCTTCTCC3’
PhLFYR 5’CATGGATCCCTAAAGCATGGAAGGAGGGATTCC3’
以蝴蝶兰花芽的cDNA作为模板,以PhLFYF和PhLFYR作为引物扩增带酶切位点Xba I和BamH I的蝴蝶兰PhalLFY基因,连接到Takara的PMD18-T载体上,再根据J.萨姆布鲁克等《分子克隆实验指南》进行转化和鉴定。将鉴定的阳性质粒利用酶切位点Xba I和EcoR I从T载体上切下PhalLFY基因,与带有35s启动子和nos终止子的植物表达载体pBI121同样用Xba I和EcoR I酶切后连接,构建表达载体pBI121-35s-PhalLFY(经测序验证,确认如SEQ ID NO.1中的第41位至1462位碱基所示的PhalLFY基因插入了植物表达载体pBI121中)。
(2)将重组表达载体pBI121-35s-PhalLFY采用冻融法转化到根瘤农杆菌GV3101中。
从-70℃中取出农杆菌感受态细胞,冰上解冻,加入5μl重组表达载体pBI121-35s-PhalLFY,轻轻混匀,冰浴30min。
将此混合物转移置研钵中,液氮速冻1min,迅速转入37℃金属浴中,将其融化,约2-3min。
向混合物中加入1ml LB液体培养基,于28℃摇床200rpm培养2-4h。所述的LB液体培养基为本领域常用培养基,其配方参考J.萨姆布鲁克等《分子克隆实验指南》。
将转化产物涂布于LB+Kan(卡那霉素)50mg/L+Genta 25mg/L的平板上,吹干,封口,放于28℃培养箱培养48小时。
挑取单克隆进行菌落PCR鉴定,鉴定引物采用实施例2中的特异扩增引物LF和LR,筛选含有目的基因(PhalLFY)的根瘤农杆菌GV3101阳性克隆(即重组表达载体pBI121-35s-PhalLFY转化进入了根瘤农杆菌GV3101中)。
(3)利用花序浸染法转化拟南芥
挑取一个含有目的基因(PhalLFY)的根瘤农杆菌GV3101阳性克隆,接种于LB+Kan(卡那霉素)50mg/L+Genta 25mg/L的液体培养基中,28℃200rpm振荡培养24-36h,使OD600=0.8左右。
将菌液转入无菌的50ml离心管中,5000rpm,15min离心收集菌种,再用同等体积的渗透培养基(1/2MS+0.5g/L MES+5%Sucrose,pH5.7)重悬农杆菌,并加入表面活性剂Silwet,使其终浓度达到0.02%(200μl/L)。所述的MS培养基是本领域常用培养基,其配方参考(Murashige T and Skoog F,1962)。
将刚开花的拟南芥植株花序在此溶液中浸泡1min。斜置晾干多余的菌液。
将浸染菌液的拟南芥花序盖膜保湿,黑暗培养8小时左右,再揭膜置于光照下正常管理成熟收获种子。
(4)转基因植株的分子鉴定。
收集当代转基因植株种子(T0代),灭菌(10%NaClO 10min,灭菌水漂洗6次)后播种于含有Kan抗生素(50mg/L)的1/2MS固体培养基上筛选。将生长10天左右的绿色抗性植株转栽到盆钵里,基质为泥炭土与蛭石比为3:1(体积比)。收集T1代种子。将该种子种植于含有Kan抗生素(50mg/L)的1/2MS培养基上,统计分离比,选取符合3存活:1死亡分离比的转基因株系植株移栽至钵里进行栽培,收集T2代种子,将该种子种植于含有Kan抗生素(50mg/L)的1/2MS培养基上,种子在培养基上全部为绿苗,T2代种子为纯合系。以T2代转基因植株的基因组DNA为模板进行PCR鉴定检测,PCR引物为PhLFYF2:5’ATGGACCCAAGCGACGCCTTCTCC3’,PhLFYR2:5’CTAAAGCATGGAAGGAGGGATTCC3’,检测结果如图4所示(部分结果)。PCR结果表明携带有外源蝴蝶兰PhalLFY基因的表达质粒已经插入到拟南芥基因组中。由此而获得转入pBI121-35s-PhalLFY表达载体的转基因拟南芥。
(5)转基因植株进行表型鉴定。
将转基因拟南芥T2代植株(转入pBI121-35s-PhalLFY表达载体的转基因拟南芥)和野生型拟南芥同等条件下进行培养,与野生型拟南芥相比,转基因拟南芥(转入pBI121-35s-PhalLFY表达载体的转基因拟南芥)抽薹及开花时莲座叶仅有7片叶,转基因植株都比野生型拟南芥提前开花,同等培养条件下,野生型拟南芥需要26天开花,而转入PhalLFY基因的转基因拟南芥仅需要15天开花。根据结果表明,PhalLFY基因超表达促进拟南芥提前开花,同时促进腋芽分化成花芽,出现多花枝现象,试验结果表明,蝴蝶兰PhalLFY基因为有功能的基因,不仅可以影响植株的发育,具有促进生殖生长,缩短开花时间的作用,而且可以促进腋芽分化成花芽,产生多花枝现象。
实施例4
蝴蝶兰PhalLFY基因在烟草中的表达及转基因植株的表型鉴定
1.含目的基因PhalLFY表达载体的构建
设计引物PhLFYF 5’AATTCTAGAATGGACCCAAGCGACGCCTTCTCC3’
PhLFYR 5’CATGGATCCCTAAAGCATGGAAGGAGGGATTCC3’
以蝴蝶兰花芽的cDNA作为模板,以PhLFYF和PhLFYR作为引物扩增带酶切位点Xba I和BamH I的蝴蝶兰PhalLFY基因,连接到Takara的PMD18-T载体上,再根据J.萨姆布鲁克等《分子克隆实验指南》进行转化和鉴定。将鉴定的阳性质粒利用酶切位点Xba I和EcoR I从T载体上切下PhalLFY基因,与带有35s启动子和nos终止子的植物表达载体pBI121同样用Xba I和EcoR I酶切后连接,构建表达载体pBI121-35s-PhalLFY(经测序验证,确认如SEQ ID NO.1中的第41位至1462位碱基所示的PhalLFY基因插入了植物表达载体pBI121中)。
2.将表达载体pBI121-35s-PhalLFY采用电击法转化到根瘤农杆菌LBA4404中
1)取1μl质粒与刚制好的农杆菌LBA4404感受态细胞混匀,冰上放置10min。
2)将此混合物转移至已预冷的电极杯电击。
3)向电击杯中加入1000μl的YM液体培养基,重悬细胞后,转移到1.5ml的离心管中,28℃,220-250rpm复苏2-4小时。
4)将转化产物涂板,放于28℃培养箱培养48小时。
5)挑取单克隆进行菌落PCR鉴定筛选阳性克隆,鉴定引物采用实施例2中的特异扩增引物LF和LR,此阳性克隆即为转化进入了表达载体pBI121-35s-PhalLFY的根瘤农杆菌LBA4404。
3.利用叶盘法转化烟草
1)挑取阳性克隆(含有表达载体pBI121-35s-PhalLFY的根瘤农杆菌LBA4404),接种于YM+Kan 50mg/L+硫酸链霉素25mg/L的液体培养基中,28℃200rpm振荡培养24-36h,使OD600=0.5左右。
2)将菌液转入无菌的50ml离心管中,5000rpm,15min离心收集菌种,再用同等体积的MS液体培养基悬浮菌种,准备浸染烟草叶片。
3)于无菌条件下将烟草无菌苗的成熟叶片除去叶边缘,切成0.5×1.0cm的小块,浸泡在制备好的菌液中;
4)10min后取出叶片于灭菌的滤纸上吸去菌液,放在MS+3%蔗糖+0.6-0.7%琼脂的培养基中,置于24±1℃光照培养箱中,14h光照/10h黑暗条件下共培养48小时;
5)当看到叶片与培养基接触边缘长出白色农杆菌时,将叶片转移至不加激素的MS液体培养基中,于28±1℃恒温摇床上振荡漂洗45-60min,夹出叶片放于灭菌的滤纸上除去多余的液体培养基;
6)将叶片转入生芽筛选培养基(MS+500mg/L Carbincine+300mg/L Kanamicine+0.1mg/L NAA+1.0mg/L BA+0.59g/L+MES+3%蔗糖+0.6-0.7%琼脂、pH 5.7-5.8)上,置于24±1℃光照培养箱中,14h光照/10h黑暗条件下培养;
7)约20天后可见分化芽长出,待芽长大后,切下并转入生根培养基(MS+200mg/L Carbincine+100mg/L Kanamicine+1.5%蔗糖+0.6-0.7%琼脂、pH 5.7-5.8)中进行筛选,置于24±1℃光照培养箱中,14h光照/10h黑暗条件下培养筛选;
8)待根系发达后,将植株取出,用无菌水洗净附着的固体培养基,移入土壤中,温室中培养,得到转基因植株。
4.转基因植株的分子鉴定
DNA的提取采用Takara的“Universal Genomic DNA Extraction Kit”。以转基因植株的基因组DNA为模板进行PCR鉴定检测,PCR引物为PhLFYF2:5’ATGGACCCAAGCGACGCCTT CTCC3’,PhLFYR2:5’CTAAAGCATGGAAGGAGGGATTCC3’。检测结果如图6所示(部分结果)。PCR结果表明携带有外源蝴蝶兰PhalLFY基因的表达质粒已经插入到烟草基因组中,由此得到转有PhalLFY基因的转基因植株。
5.转基因植株进行表型鉴定
与野生型烟草相比,转基因植株(转有PhalLFY基因的转基因植株)都比野生型烟草提前开花,花枝增多(图7)。根据结果推测外源PhalLFY基因在宿主烟草中实现了超量表达,促进了转基因烟草的生殖生长和花芽发育,从而促进了开花时间。
- 还没有人留言评论。精彩留言会获得点赞!