拟南芥SAL基因在调节开花时间和莲座叶数量中的应用的制作方法
本发明属于生物技术领域,具体涉及拟南芥SAL基因在调节开花时间和莲座叶数量中的应用。
背景技术:
在开花植物中,营养生长到生殖生长的转变,开花,是植物发育的最关键阶段。在农业中,开花的时间很大程度决定了种子和果实的数量和质量。植物的开花受到一系列的环境和内源因素的影响,如日照长度、生物钟和赤霉素等,并通过体内复杂的基因调控网络调节开花的时间。对植物开花的遗传学研究大多数以模式植物拟南芥为对象,而后将相应的研究结果应用于其他经济作物中,如水稻,大豆和玉米中,极大改善了作物的经济性状以及对不利环境的适应性。
对拟南芥开花相关基因的研究已经有几十年的历史,期间发现了许多重要的开花调节因子,如SOC1、FT、AGL24和CO等,响应不同的环境信号,通过激活分生组织中的LFY、AP1以及SEP3等基因,控制营养生长到生殖生长的转变。值得注意的是,开花的早晚通常与拟南芥莲座叶的数量呈负相关,即开花早的植物莲座叶数量较小。一直以来,植物被认为在营养和生殖生长上的分配是此消彼长,也因此作物的收成被认为是有上限的,因为开花早的作物基本营养生长较欠缺(叶片数量少,光合作用产物量低)导致产量低。那么是否营养和生殖生长在遗传学层面不能独立被控制呢。SAL突变体和过表达植株的表型显示这是有可能的,该基因功能的研究结果对突破作物产量增长的瓶颈有极其重要的意义。
技术实现要素:
本发明的目的在研究拟南芥SAL基因的在营养和生殖生长转变过程(开花)中的功能,提供拟南芥SAL基因在调节开花时间和莲座叶数量中的应用。
为实现上述目的,本发明采用如下技术方案:
拟南芥SAL1 /2抑制开花时间和莲座叶数量,所述的SAL1 /2分别为拟南芥基因AT1G13360和AT3G25870为同源基因。其氨基酸序列分别如SEQ ID No.1-2所示。
所述的拟南芥SAL1 /2延迟了开花时间并抑制了莲座叶的生成。
本发明的优点在于:本发明通过对SAL1/2的功能缺失型突变体纯合植株的表型进行分析得知:单突sal1, sal2以及双突sal1sal2与野生型Col-0相比,均出现开花时间缩短7天左右,但开花时莲座叶数量增加2-3片,开花后茎秆的生长速度较快。同时,通过构建SAL2的过表达植株后发现:与野生型相比,过表达植株出现明显的矮化,并且开花时间延长了一个月左右。SAL1/2的突变体的开花表型与正常的早花表型出现明显差异,即莲座叶的数量增加,尚无该类表型的报道;通过SAL2突变体和过表达植株的表型可以推测其功能为限制植物过快发育,该基因功能的研究结果对突破作物产量增长的瓶颈有指导意义。
附图说明
图1 SAL1/2 功能缺失型突变体纯合子的鉴定。其中a/b/c/d/e分别为纯合子鉴定所用引物名称,a为SALK_052626-LP+SALK_052626-RP,b为SALK_052626-RP+LBa1.3,c为SALK_027501-LP+SALK_027501-RP,d为SALK_027501-RP+ LBa1.3,e为SALK_027501-LP+ LBa1.3,序列详见“实施例1”。
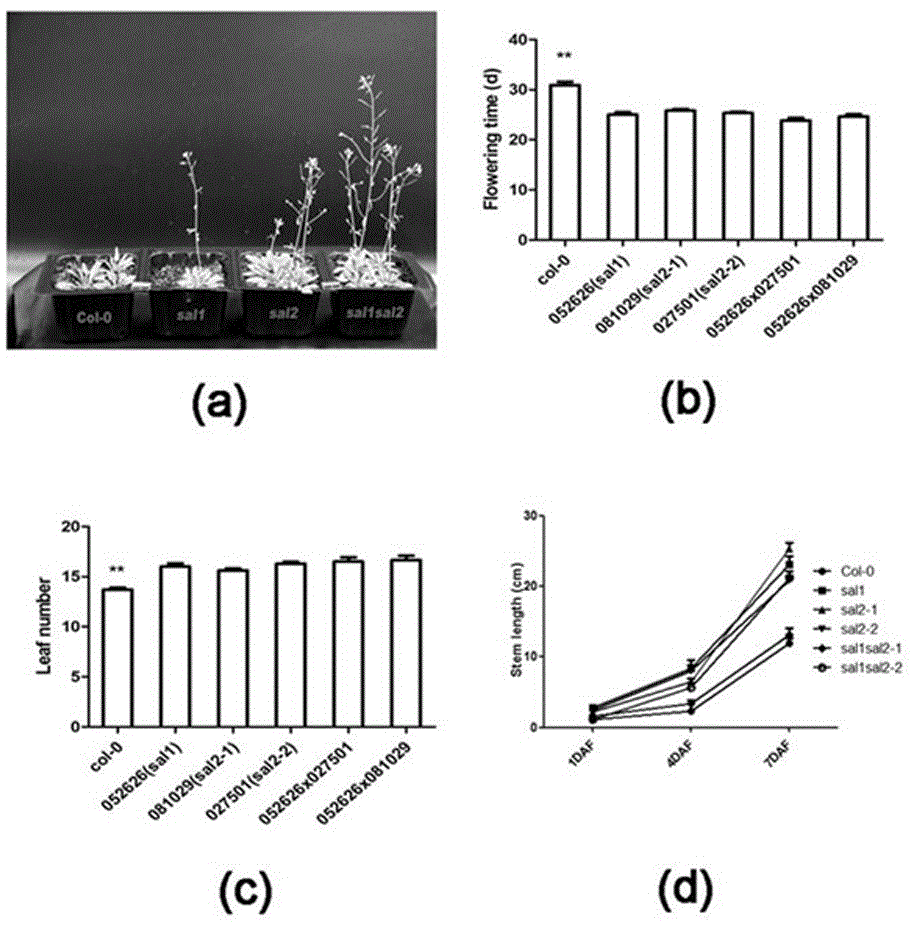
图2 SAL1/2功能缺失型突变体的开花相关表型。其中a为4周大的突变体形态;b为突变体植株的开花时间;c为突变体植株的莲座叶数量;d为突变体植株开花后茎秆的生长速率。
图3 过表达植株中SAL2的相对表达量。
图4 SAL2 过表达的开花时间。
具体实施方式
实施例1
1.突变体的获得
所用SAL1/2突变体均订购自拟南芥资源中心(ABRC, The Ohio State University Rightmire Hall 1060 Carmack Road Columbus, OH 43210 USA)。其中sal1编号为SALK_052626,sal2-1编号为SALK_018029c,sal2-2编号为SALK_027501,所有突变体构建方法均为T-DNA插入。
双突变体sal1sal2-1和 sal1sal2-2的构建,采用单突变体相互杂交的方法获得,具体操作方法为:寻找还是紧闭的花蕾,但柱头开始从花顶部突出。用已灭菌的镊子,去掉一株中不去雄的任何花蕾。用已灭菌的镊子去掉除雌 蕊以外的花的所有部分,立即授粉。选择已盛开的花,用灭菌的镊子取下雌蕊。用花粉轻轻碰已去雄的花柱头。如柱头伸长,子房变大,则表明杂交成功。
2.突变体的纯合子鉴定
纯合子的鉴定采用PCR法,以植物叶片的DNA为模板,T-DNA中间引物选用LBa1.3(5’到3’, ATTTTGCCGATTTCGGAAC),基因引物分别为:
引物名称 序列(5'到3')
SALK_027501-LP CAAAACTCAACTTGTTTGGGTG
SALK_027501-RP AGGAGAGGCTAACTCGGACTG
SALK_052626-LP TGTTTCAGGTCCAAGTCGAAC
SALK_052626-RP GGAGCTTTTGATTTAGAGCGG
其中,由于SALK_018029c和SALK_027501插入位点十分接近,所以共用鉴定引物SALK_027501-LP和SALK_027501-RP。
3.过表达载体的构建
过表达载体的构建采用Gateway系统,SAL1/2首先利用BP反应被克隆入入门载体pDONR207,而后用LR反应克隆入表达载体pEarleyGate104(N端YFP),获得35S::YFP-SAL1和35S::YFP-SAL2。
4.植物的转化
采用农杆菌介导的浸花法进行植物转化,首先将构建好的35S::YFP-SAL1和35S::YFP-SAL2通过冻融法转入农杆菌GV3101中,通过浸花法侵染野生型拟南芥rdr6-11(该基因型植株可以有效防止转化基因在子代RNA水平的沉默),共侵染两次,间隔一周。收获的种子以除草剂Basta为筛选标记,得到转化子植株,并通过SAL1/2的表达量(qPCR)作为过表达植株的评判标准。
5.表型的观察
5.1开花时间的测定:以种子发芽为起始时间,看到第一个花芽为终止时间;
5.2 莲座叶数量的统计:计算植株开花时的莲座叶数量;
5.3 茎秆生长速率的测定:在开花后的第1,4,7天分别测量茎秆的高度。
实验结果:
SAL突变体的开花相关表型
借助PCR的手段,鉴定出了SAL1/2的单突sal1, sal2以及双突sal1sal2的纯合子植株(图1)。通过对开花时间、莲座叶数量以及生长速率的分析显示:与野生型Col-0相比,单突sal1, sal2以及双突sal1sal2均出现开花时间缩短7天左右(图2b),但开花时莲座叶数量比野生型多2-3片(图2c),开花后一周内茎秆的生长速度明显较野生型快(图2d)。
突变体的开花时间表型
通过构建SAL1/2的过表达植株后发现:无法获得rdr6-11为背景的35S::YFP-SAL1转化植株,35S::YFP-SAL2的转化植株的表达量变化仅为野生型的2倍左右(图3)。与野生型相比,SAL2过表达植株出现明显的矮化,并且开花时间较野生型rdr6-11延长了一个月左右(图4)。
以上所述仅为本发明的较佳实施例,凡依本发明申请专利范围所做的均等变化与修饰,皆应属本发明的涵盖范围。
SEQUENCE LISTING
<110> 福建农林大学
<120> 拟南芥SAL基因在调节开花时间和莲座叶数量中的应用
<130> 7
<160> 7
<170> PatentIn version 3.3
<210> 1
<211> 194
<212> PRT
<213> SAL1
<400> 1
Met Glu Lys Lys Leu Leu Asp Ile Thr Arg Thr Asp Ser Ala Glu Lys
1 5 10 15
Lys Arg Val Arg Asp Glu Ser Phe Asp Glu Ala Val Leu Asp Ser Pro
20 25 30
Glu Val Lys Arg Leu Arg Asp Asp Leu Phe Asp Val Leu Asp Asp Ser
35 40 45
Asp Pro Glu Pro Val Ser Gln Asp Leu Asp Ser Val Met Lys Ser Phe
50 55 60
Glu Asp Glu Leu Ser Thr Val Thr Thr Thr Thr Ala Gln Gly Ser Ser
65 70 75 80
Thr Ala Gly Glu Thr Gln Pro Asp Leu Gly Tyr Leu Leu Glu Ala Ser
85 90 95
Asp Asp Glu Leu Gly Leu Pro Pro Pro Pro Ser Ile Ser Pro Val Pro
100 105 110
Val Ala Lys Glu Glu Val Thr Thr Glu Thr Val Thr Asp Leu Val Arg
115 120 125
Ala Ser Ser Asp Ser Ser Gly Ile Asp Glu Ile Trp Gly Phe Glu Asp
130 135 140
His Val Ser Asn Tyr Gly Gly Leu Asp Phe Gly Ser Gly Val Gly Asp
145 150 155 160
Gly Gly Asp Tyr Val Ala Val Glu Gly Leu Phe Glu Phe Ser Asp Asp
165 170 175
Cys Phe Asp Ser Gly Asp Leu Phe Ser Trp Arg Ser Glu Ser Leu Pro
180 185 190
Ala Glu
<210> 2
<211> 170
<212> PRT
<213> SAL2
<400> 2
Met Glu Asp Leu Leu Glu Glu Arg Leu Thr Arg Thr Asp Ser Val Gly
1 5 10 15
Asn Lys Arg Val Arg Asp Gly Leu Asp Leu Asp Ser Pro Asp Val Lys
20 25 30
Arg Leu Arg Asp Asp Leu Phe Asp Asp Ser Gly Leu Asp Pro Val Ser
35 40 45
Gln Asp Leu Asp Ser Val Met Lys Ser Phe Glu Asn Glu Leu Ser Thr
50 55 60
Thr Thr Ala Ala Leu Ser Ser Gly Glu Thr Gln Pro Asp Leu Gly Tyr
65 70 75 80
Leu Phe Glu Ala Ser Asp Asp Glu Leu Gly Leu Pro Pro Pro Leu Thr
85 90 95
Pro Pro Gln Thr Leu Leu Pro Pro Ser Cys Glu Glu Thr Val Thr Glu
100 105 110
Leu Val Arg Ala Ser Ser Asp Ser Ser Glu Val Gly Glu Leu Cys Gly
115 120 125
Phe Glu Asp His Val Thr Glu Phe Gly Pro Cys Asp Leu Gly Asp Asp
130 135 140
Gly Leu Phe Glu Tyr Phe Asp Gly Cys Leu Asp Ser Gly Asp Leu Phe
145 150 155 160
Ser Trp Arg Pro Glu Phe Leu Pro Ala Glu
165 170
<210> 3
<211> 19
<212> DNA
<213> 人工序列
<400> 3
attttgccga tttcggaac 19
<210> 4
<211> 22
<212> DNA
<213> 人工序列
<400> 4
caaaactcaa cttgtttggg tg 22
<210> 5
<211> 21
<212> DNA
<213> 人工序列
<400> 5
aggagaggct aactcggact g 21
<210> 6
<211> 21
<212> DNA
<213> 人工序列
<400> 6
tgtttcaggt ccaagtcgaa c 21
<210> 7
<211> 21
<212> DNA
<213> 人工序列
<400> 7
ggagcttttg atttagagcg g 21
- 还没有人留言评论。精彩留言会获得点赞!