一种甘蓝型油菜与拟南芥中NAC87转录因子基因及其应用的制作方法
本发明属于生物基因工程技术领域,具体地说,涉及一种甘蓝型油菜与拟南芥中nac87转录因子基因及其应用。
背景技术:
叶片衰老代表了叶片发育的后期阶段,关系到养分的转移与再利用,用于保障生殖生长的顺利进行,而且衰老有助于植物适应多变的环境条件。因此,研究调控叶片衰老的基因并用于改良作物的抗逆、抗病性、提高作物的产量与品质,意义非常重大。
国内外的多个报道表明,延缓植物的衰老是一个有效地提高植物抗旱性进而提高产量的方法。拟南芥是重要的模式植物,同属十字花科的甘蓝型油菜则是我国重要的油料作物,两者的基因的相似性较高,平均达80%以上,因此可以相互借鉴、相互促进。环境胁迫包括干旱以及冷害等严重影响菜籽的产量与品质。利用分子生物学与遗传学方法,挖掘油菜中调控抗逆的基因并阐明它们在增强油菜抗逆性上的应用,通过分子辅助的育种技术培育高产、抗逆的油菜新品种,是一条行之有效的途径。
在叶片衰老的信号转导通路中,转录因子发挥着核心调控作用。目前,已鉴定、发现了拟南芥中一些能显著促进或延缓叶片衰老的转录因子,比如wrky53、wrky75、nap(anac029)、ataf1(anac002)、jub1(anac042)、ore1(anac092)等。研究表明,这些基因在植物中的表达水平不仅决定叶片衰老的速度,而且也调控对于干旱、盐害等的耐受性,说明具有重要的应用价值。但是,油菜中有哪些基因调控叶片衰老,至今未见任何报道。
nac(nam,ataf,cuc)转录因子家族是植物中特有的,构成较大的基因家族,在植物的生长、发育、激素应答以及提高植物对干旱、盐害、冷害等逆境的耐受中发挥着重要作用。但是,有关油菜中调控叶片衰老与抗逆的nac基因的鉴定与研究,鲜见报道。而且,即使在模式植物拟南芥中,由于nac基因众多,仅有少数被研究过,还有很多nac基因的功能尚待解析。
nac转录因子基因是植物特有的基因家族,广泛存在于陆生植物中。最早被发现参与矮牵牛与水稻的胚胎发育、器官建成以及花发育。nac转录因子的一般特点是在氨基端有一个高度保守的约含160个氨基酸残基的能结合下游靶基因的启动子区顺式作用元件(nacrecognitionsequence,nacrs)的nac结构域,羧基端有一个高度可变的调控转录活性的区域。拟南芥与水稻基因组中分别有117、151个nac基因,划分为8个亚类,其中仅有部分nac的功能被研究报道过。该基因家族成员功能多样,有研究表明其功能主要与植物生长发育、激素调控以及植物对干旱、高盐、低温等非生物和病原菌侵染等生物胁迫的抗逆反应相关。例如,拟南芥的nap(anac029)调控依赖脱落酸(aba)的叶片与果实的衰老,并且jub1(anac042)、ore1(anac092)、ors1(anac059)、anac016以及anac019/055/072也被发现调控叶片衰老;而且ore1、anac016与anac019/055/072调控的叶片衰老与叶绿素的降解有关。此外,拟南芥anac016还被发现通过调控areb1/abf2转录因子基因的表达而影响植物的抗旱性,再一次暗示叶片的衰老进程与抗旱等性能紧密相关。但是,油菜以及拟南芥中有无其他的nac转录因子基因调控叶片衰老以及抗逆等,并不清楚,尚待进一步研究。
技术实现要素:
本发明的目的在于提供一种甘蓝型油菜与拟南芥中nac87转录因子基因及其应用。发明人在前期开展有关甘蓝型油菜中nac转录因子基因家族的研究的基础上,首先根据公共数据库中的表达序列标签(est)信息,设计基因特异性引物,从油菜cdna中克隆获得nac87转录因子全长cdna,并且在油菜以及烟草中进行功能验证。同时,发现拟南芥的同源基因anac087也具有类似地诱导活性氧积累与叶片衰老的功能。
其具体技术方案为:
一种甘蓝型油菜与拟南芥中nac87转录因子基因,油菜nac87转录因子基因的核苷酸如seqidno:1所示,氨基酸如seqidno:2所示,拟南芥nac87转录因子基因的核苷酸如seqidno:3所示,氨基酸如seqidno:4所示。
本发明所述甘蓝型油菜与拟南芥中nac87转录因子基因在调控叶片衰老以及抗逆过程中的应用。
进一步,利用本发明所述的nac87基因促进叶片衰老的方法,是将nac87构建到过表达载体pyjha,然后导入宿主植物的细胞、组织或植株个体,得到衰老速度加快的植株。利用本发明所述的nac87基因延缓叶片衰老的方法,是通过生物技术手段将nac87基因敲除或沉默,得到衰老速度延迟的植株。
进一步,所述宿主植物为拟南芥和油菜。
与现有技术相比,本发明的有益效果为:
本发明首次发现油菜与拟南芥的nac87同源基因均能正调控叶片衰老,并且与活性氧的累积相关,在调控作物的抗逆以及抗病方面也具有重要的应用价值。
附图说明
图1是油菜与拟南芥中nac87基因的rt-pcr扩增产物的电泳检测,其中,m,1kbdna分子量标准(fermentas);1,油菜nac87基因的扩增;2,拟南芥nac87基因的扩增。
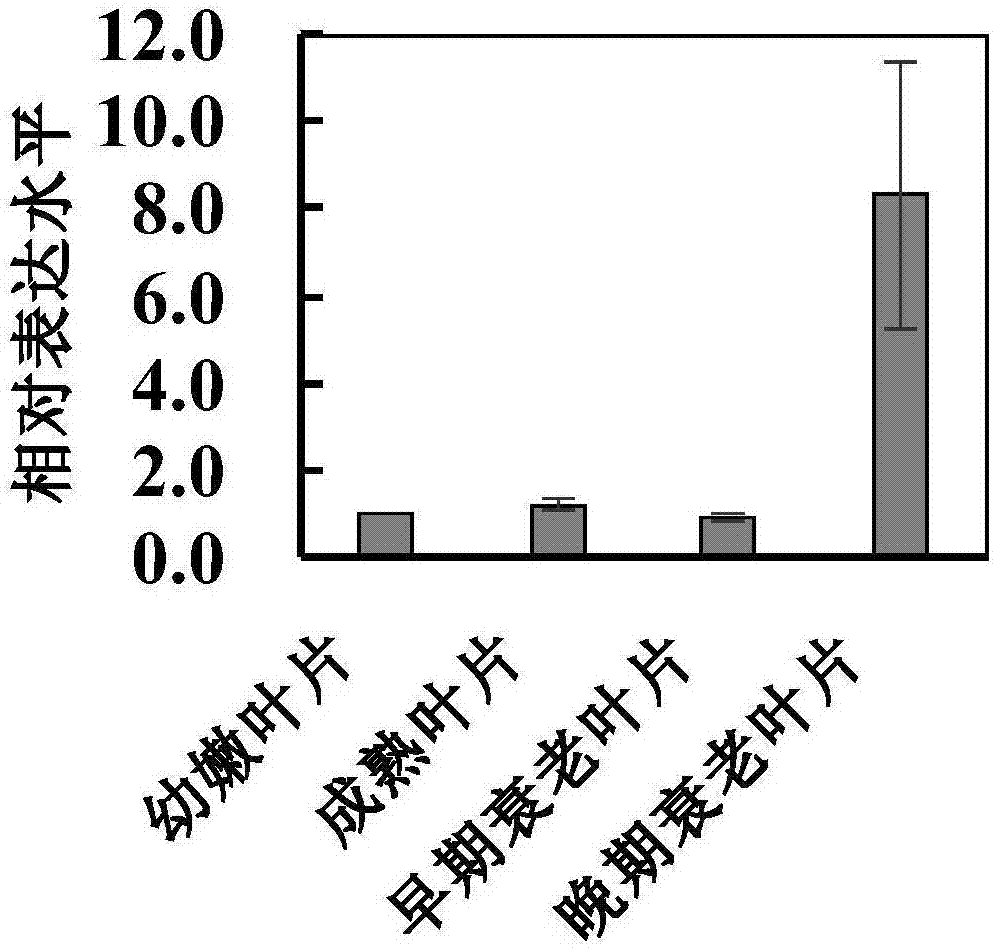
图2是油菜nac87在叶片发育进程中的表达含量的实时荧光定量rt-pcr分析,数值来自三个生物学重复±标准方差;
图3是拟南芥中anac087基因的t-dna插入突变体的pcr鉴定,m,1kbdna分子量标准(fermentas)。wt表示野生型col-0;1-13代表13个独立的株系。a是lp+rp引物组合的pcr扩增的电泳检测图;b是gabi8409+rp引物组合的pcr扩增的电泳检测图。
图4是拟南芥nac87基因的突变体与过表达株系的表型分析,其中图4a为野生型col-0,图4b为anac087突变体,图4c为anac087的过表达株系;
图5是nac87的表达诱导活性氧(ros)的累积与叶片提前衰老,其中图5a通过农杆菌介导的注射,瞬时表达bnanac87及gus对照基因,分别是2、3、4天时观察表型并进行dab染色。左侧为叶片正面,中间为背面,右侧为dab染色后的背面拍照,图5b是bnanac87及gus表达材料的相对电导率的测定分析,图5c是bnanac87及gus表达材料的双氧水含量的测定分析,统计数据是三个独立生物学重复的平均值±s.e,不同字母代表显著性差异(anova分析);
图6是nac87的表达诱导叶片衰老的生理生化指标的测定分析,其中图6a是bnanac87及gus表达材料的叶绿素含量的测定分析,图6b是bnanac87及gus表达材料的花青素含量的测定分析,图6c是bnanac87及gus表达材料的丙二醛含量的测定分析,相关数据是三个独立生物学重复的平均值±s.e,不同的字母表示显著性差异(anova分析)。
具体实施方式
下面结合附图和具体实施例对本发明的技术方案作进一步详细地说明。
本发明提供了一种甘蓝型油菜与拟南芥中nac87转录因子基因及其编码蛋白与应用。
本发明涉及的甘蓝型油菜与拟南芥的基因,名称分别为bnanac87与anac087,其基因座位号分别是bnac02g07990d、at5g18270,编码的蛋白分别含有337、335个氨基酸残基。
含有本发明基因的表达载体、转基因植株也在本发明的保护范围之内。
本发明的保护范围还包括基因nac87在调控叶片衰老以及抗逆中的应用,该植物为拟南芥和油菜。
利用该nac87基因促进叶片衰老的方法,是将nac87构建到过双元表达载体pyjha,通过农杆菌介导,导入宿主植物的细胞、组织或植株个体,得到衰老速度加快的植株。利用本发明所述的nac87基因延缓叶片衰老的方法,是通过生物技术手段(比如rna干涉、t-dna插入突变或crispr-cas9基因组编辑)将nac87基因敲除或沉默,得到衰老速度延迟的植株。
以上所说的植物宿主为拟南芥和油菜。
实施例i油菜nac87基因cdna序列的克隆与序列测定
根据ncbiest数据库以及tair公用数据库的序列信息,分别设计基因特异性的两对引物,分别以甘蓝型油菜与拟南芥col-0生态型的cdna为模板扩增基因的编码区,引物序列为(下划线表示限制性内切酶位点):
bnanac87-f:5'-ttaggtcatgatcgcggttgttgttgttgaaga-3'
bnanac87-r:5'-cgcgtcgacgtagtcccataagatccctcttaag-3'
anac087-f:5'-ggactagtgcggttgtggttgaagaa-3',
anac087-r:5’-cgcgtcgacgaagtcccacaagtcccccct-3’。
采用plantrnakit(omegabio-tek)提取油菜与拟南芥col-0幼苗的总rna,经过nanodrop1000定量与电泳检测后,各取2.5μg总rna,采用revertaidfirststrandcdnasynthesiskit(fermentas)进行逆转录反应,按说明书进行。
2.pcr反应体系(50μl)
5xprimestarbuffer10μl
cdna模板:1μl
dntps(10mmeach)1μl
primerf(20μμ)1μl
primerr(20μμ)1μl
primestardnapolymerase(takara)0.5μl
ddh2o加至终体积50μl
3.pcr反应程序为:94℃预变性2min;94℃变性30sec,50℃复性30sec,72℃延伸1min,循环35次;72℃延伸10min。
4.1%琼脂糖凝胶电泳
pcr扩增产物用1%琼脂糖凝胶电泳检测后发现在1kb处有一条目的条带如附图1所示。
5.扩增片段的回收、酶切以及与pjet1.2载体的链接
对扩增条带采用omegabio-tek公司的琼脂糖胶回收试剂盒,步骤按说明书进行。
对回收片段,通过平末端克隆的方法,克隆入pjet1.2载体(fermentas),按照clonejetpcrcloningkit(fermentas)的说明书进行。
本实验所用的pjet1.2载体的多克隆位点的上游有t7启动子、下游有pjet1.2-r引物结合的序列,所以可以用这两个引物序列对重组质粒进行双向测序。结果表明,我们获得了甘蓝型油菜与拟南芥的nac87基因的cdna序列。
nac87转录因子基因在发育过程中的表达分析
i.材料处理
甘蓝型油菜种子经过次氯酸钠消毒处理后,播种在1/2xms培养基上,于4℃层积处理两天后,置于萌发7天,然后移栽到泥炭土基质,放于温室正常生长,三周时取幼嫩叶片、4周时取成熟的叶片、5周时取早期衰老的叶片、6周时取晚期衰老的叶片,分别用液氮速冻后存于-80℃冰箱。
2.trizol(sangon)法提取油菜叶片的总rna,按照说明书进行。
用nanodrop1000检测rna的浓度与纯度,根据1od=40ug/ml计算浓度。根据在230nm、260nm和280nm处的吸光值,检测rna的纯度,纯rna的od260/od280应为1.9左右,od260/od230应大于2。
采用revertaidfirststrandcdnasynthesiskit(fermentas)进行逆转录反应,按说明书进行。
以10倍稀释的cdna为模板,进行定量pcr反应,采用sybrpremixextaqtm‖(perfectrealtime,takara)试剂盒,在cfx96(bio-rad)仪器上进行。引物如下:
qpcr体系
2xsybrgreenimix5μl
primerqf(20μμ)0.2μl
primerqr(20μμ)0.2μl
ddh2o2.6μl
cdna模板2μl
totalvolume10μl
pcr程序
95℃2min;94℃30s;60℃1min;35cycles。
根据内参基因bnaup1与bnaubc9,计算目的基因在各个组织材料中的表达倍数。结果表明油菜的nac87基因倾向于在正在衰老的叶片内表达(图2)。
植物过表达载体的构建
植物表达载体pyjha是含有camv35s启动子和hptii(潮霉素)植物选择标记基因的双元载体,在其多克隆位点上含有限制性内切酶ncoi、spei和sali等位点。用限制性内切酶ncoi或spei和sali双酶切载体pyjha;同时,用bsphi和sali双酶切目的基因片断。完全酶切的载体在1%琼脂糖凝胶上电泳分离后,经胶回收,然后与双酶切的目的基因片段连结,构建获得植物表达载体pyjha-bnanac87与pyjha-anac087。
(i)质粒pyjha空载体和目的基因片断双酶切体系如下:
ncoi(或者spei)0.5μl
sali0.5μl
双酶切过的pyjha载体或目的基因片段3μl
10xbuffer1.5μl
ddh2oto15μl
于37℃恒温水浴锅酶切4小时;
酶切产物的电泳与回收:双酶切完成后,以1xtae为电泳缓冲液,将酶切产物进行1%琼脂糖凝胶电泳。在紫外透射仪下用洁净刀片切下pyjha中载体的11kb大片段,琼脂糖凝胶回收试剂盒(omegabio-tek)回收目的条带;目的基因片段用pcr产物纯化试剂盒(omegabio-tek)回收。
连接:经酶切的pyjha载体片段和目的基因片段以摩尔比1:3的比例进行16℃连接2小时;
采用常规的cacl2介导的热激法转化入大肠杆菌dh5α感受态细胞之后,通过菌落pcr筛选阳性克隆,
本实验所用的pyjha载体,根据genbank中登陆过的pcsgfpbt载体改造而来,在基因的编码区的末尾增加了2xha亲和标签序列,设计了上下游测序引物,所以可以用这两个引物对插入片段进行扩增,对重组质粒进行鉴定;
pcr程序:94℃预变性3min;94℃变性1min,50℃复性1min,72℃延伸lmin,循环35次;72℃延伸5min;
pcr扩增产物用1%琼脂糖电泳检测,然后挑取两个阳性克隆摇菌,用质粒小量提取试剂盒(omegabio-tek)提取质粒。重组质粒送至生物公司采用abi3730仪器双向测序。
农杆菌感受态的制备与转化
采用cacl2法制备根癌农杆菌gv3101的感受态细胞,采用冻融法将上述pyjha-nac87重组质粒转入农杆菌细胞。涂于含34μg/ml利福平、25μg/ml庆大霉素、50μg/ml卡那霉素的yeb培养基平板上,28℃倒置暗培养2天。
菌落pcr鉴定验证重组质粒确已转入农杆菌。
菌体pcr所用引物以及扩增条件等同上述。
转基因功能验证一拟南芥转化、筛选及表型分析
拟南芥的遗传转化
首先小量培养转化后的农杆菌,然后转入大量(330ml)培养,直至od600达到1.2。离心收集菌体,等量重悬于含有5%蔗糖、1/2xms以及0.03%silwetl-77的溶液中,将约5周的处于盛花前期的野生型拟南芥col-0倒扣并使所有花序浸入悬浮菌液中,轻轻搅动蘸花30sec,取出花盆侧放于托盘中,盖盖,将托盘置暗处放置14h,然后取出营养钵并直立放置,恢复光照,继续培养植株至成熟并收获种子。
阳性植株的筛选:t1代种子用次氯酸钠溶液(包括0.03%tween-20)消毒后,点播在1/2xms选择培养板(含有30mg/l潮霉素)上,于4℃下层积2天,移入培养室中培养,7天后挑选潮霉素抗性植株(长出真叶1~2对,根伸长至培养基中)并移栽到营养钵中,培养直至种子成熟,采用同样的方法筛选t2代种子得到t3代植株,并在t2代植物中挑选抗性比为3:1的单插入独立株系,并获得纯合t3代株系进行转基因拟南芥的分子检测和表型鉴定;
转基因拟南芥的rt-pcr鉴定与高表达株系的筛选
采用plantrnakit(omegabio-tek)提取t2代植物与野生型col-0叶片的总rna,经过nanodrop1000定量后,各取2.5μg总rna,采用revertaidfirststrandcdnasynthesiskit(fermentas)进行逆转录反应,按说明书进行。
定量rt-pcr筛选高表达株系
以10倍稀释的cdna为模板,进行定量pcr反应,采用sybrpremixextaqtm‖(perfectrealtime,takara)试剂盒,在cfx96(bio-rad)仪器上进行。引物如下
atubq10-qf:5’-gttccaatctatgagggatacacgc-3’
atubq10-qr:5’-agaagttcgacttgtcattagaaagaaa-3’
anac087-qf:5’-ttggttctgcttcgggctctacg-3’
anac087-qr:5'-ccattcccatcatccccatttctt-3'
qpcr体系
2xsybrgreenimix5μl
primerqf(20μμ)0.2μl
primerqr(20μμ)0.2μl
ddh2o2.6μl
cdna模板2μl
totalvolume10μl
pcr程序
95℃2min;94℃30s;60℃1min;35cycles。
根据内参基因atubq10的扩增情况,与野生型比较,计算目的基因在各个转基因株系中的表达倍数。
突变体的鉴定
从英国nasc购得拟南芥anac087基因的t-dna插入突变体gabi_622h06种子,播种在营养土表面,4℃层积处理两天后放置于温室萌发,对四周龄的幼苗,取一片真叶,用常规的2%ctab法提取基因组dna,溶解于20ul1xte缓冲液中。利用taq聚合酶、两对引物(gabi-8409+rp、lp+rp),以野生型col-0与突变体的基因组dna作为模板,进行常规pcr扩增,然后进行1%琼脂糖凝胶电泳,根据带型筛选出纯合的突变体株系(图3)。
所用引物序列如下:
lp:5’-tcgttgggatgaagaaaacac-3’
rp:5’-tcccaaattatcacaatcctttc-3’
gabi-8409:5’-atattgaccatcatactcattgc-3’
转基因拟南芥的表型鉴定
将t3代纯合的拟南芥株系的种子与野生型col-0种子,分别用次氯酸钠溶液消毒2分钟,然后用无菌水漂洗4次,点播于1/2xms(含有1%蔗糖)培养基平板上,于4℃层积处理2天,然后放在温室中竖直生长7天,22℃培养,14h/10h光周期,光强100μmol·m-2·s-1。然后,将萌发7天的拟南芥幼苗(对照及转基因株系)用灭菌过的镊子小心移至盆钵(里面有营养土),继续在同一温室生长并观察表型,结果表明转anac087基因的拟南芥植株的叶片衰老进程明显快于野生型对照植株,而功能丧失的突变体则表现出一定程度的延缓衰老的表型(图4)。说明本发明涉及的拟南芥anac087基因的的确能调控植物叶片衰老的进程。
烟草叶片的瞬时表达
转化有pyjha-nac87重组质粒的农杆菌,在yeb液体培养基中过夜振荡培养过夜,同样的方法培养抑制基因沉默的p19菌株,次日分别离心收集菌体,将得到的菌体分别用新鲜配制的as溶液(10mmmes-koh,10mmmgcl2,0.15mm乙酰丁香酮)重悬,使其od600达到0.5后,按照1:1(v/v)等体积混匀,对照为转化有pyjha-gus的农杆菌,黑暗静置2-3h后共注射30d的本氏烟草叶片的下表皮。继续在温室中生长,光周期为14小时光照、10小时黑暗,光强为120μmolm-2sec-1,湿度为60-70%。观察、记录表型并取样检测以下生理指标。结果发现油菜nac87的瞬时表达能显著增加活性氧的累积与加快叶片的衰老(图5)。
二氨基联苯胺(3,3'-diaminobenzidine,dab)染色
将叶片剪下,放入12cm培养皿,加入50mldab染色液,确保叶片完全浸入到染色液中,用铝箔纸盖住培养皿,80-100rpm振荡4-5小时。移去dab,加入70ml脱色液(乙醇:乙酸:甘油3:1:1),放于沸水浴中脱色15min。移去脱色液,换上新的脱色液,保持30min即可,拍照记录结果。
电导率测定
用1cm直径的打孔器取直径为1cm的叶圆片,转入含5mlddh2o的离心管中,抽真空处理10min,在25℃摇床中150rpm培育30min,利用电导仪测定电导率并记录数据,记录为c1。将上述样品煮沸5min,冷却至室温后,再次测定样品电导率(c2)。同时检测ddh2o的电导率作为c0,计算相对导电率(c1-c0)/(c2-c0)*100%。
叶绿素含量测定
称取约100mg的叶圆片,置于盛有5ml无水乙醇的指形管中,室温黑暗旋转振荡过夜,次日利用分光光度计测定649及665nm波长处吸光值,计算叶绿素含量。叶绿素(μg/gfw)=(6.1*od665+20.04*od649)/样品鲜重(gfw)。
花青素含量的测定:取约100mg的叶圆片,记录鲜重,用液氮充分研磨,用甲醇(含1%hcl)4℃黑暗旋转混合过夜,次日检测530nm及657nm波长下的吸光值,计算花青素的含量。花青素/mgfw=(od530-od657)*1000/样品鲜重(mgfw)。
丙二醛含量的测定
取新鲜叶圆片,称重记录后用液氮充分研磨,加1ml10%的三氯乙酸(tca)重悬混匀,65℃水浴15min,转速10000g离心10min后吸取700μl上清,加入等量0.6%的硫代巴比妥酸(tba)混匀,90℃水浴15min后,冷却至室温,取上清检测在532nm和600nm波长处的吸光值并计算其含量。计算公式:mda(μmol/gfw)=[6.45*(od532-od600)-0.56*od450]*提取液总体积(ml)*10-3/样品鲜重(gfw)。
h2o2含量的测定
取新鲜的叶圆片,称重并记录后用液氮充分研磨,加入5%tca充分混匀,65℃水浴15min,冷却至室温后,加200μl氯仿:异戊醇(24:1,v/v)充分混匀,转速12000g离心10min。转移上清至新的离心管中,用1m氨水调ph至7.5。设置对照组和实验组,其中对照组加过氧化氢酶(cat,sigma)处理。取100μl样品至新的离心管,后各管加400μltris-hcl(ph8.5)。反应30min后,加入500μlpar溶液[30mmpotassiumtitaniumoxalate:30mm4-pyridylazoresorcinol=1:1(v/v)],室温稳定15min后,利用紫外分光光度计检测508nm处的吸光值,计算得到叶片中h2o2的含量。首先计算标准曲线:y=4913.5*od508-873.77,h2o2(总量)=(y实验组-y对照组)*(800+330)/100,h2o2(μm/mgfw)=h2o2(总量)/样品鲜重(mgfw)。
上述多个生理与生化指标的定量分析表明,油菜nac87基因的表达能引起叶片组织的坏死导致的电导率上升、过氧化氢含量的上升、叶绿素含量的降低、花青素含量的升高以及丙二醛含量的上升(图5与图6)。
以上所述,仅为本发明较佳的具体实施方式,本发明的保护范围不限于此,任何熟悉本技术领域的技术人员在本发明披露的技术范围内,可显而易见地得到技术方案的简单变化或等效替换均属于本发明的保护范围内。
sequencelisting
<110>西北农林科技大学
<120>一种甘蓝型油菜与拟南芥中nac87转录因子基因及其应用
<160>21
<170>patentinversion3.3
<210>1
<211>1014
<212>dna
<213>人工序列
<400>1
atggcggttgttgttgttgaagaaggcgtggtgttgaatcatggaggtgaagagcttgtg60
gatttaccacctggattcaggtttcatccaacagacgaggagatcataacattctacctc120
aaagaaaaggttttagacagccgattcacggctgtggccatgggtgaagcagatctcaac180
aagtgtgagccctgggatttgccgaagagagcaaagatgggggagaaagagttttacttc240
ttctgtcaaagggacaggaagtacccgaccgggatgaggacgaaccgtgcgaccgagtcc300
gggtattggaaagcgaccgggaaggataaggagattttcaaaggcaaaggttgtctcgtt360
gggatgaagaaaacacttgtgttttatagaggaagagctccacgaggtgaaaagactaat420
tgggtcatgcatgaatatcgtcttgatggcaaatattctcatcacaatctccccaaatcc480
gcaagggacgagtgggtcgtgtgtagggtttttcacaagaacaatccttccactacaacc540
caacaaatgacaagaatgcccattgaaaatctcacaagaatggattctctagagaacatt600
gatcatctcctagacttctcatctcttcctcctctcatggatccgagtttcacgggtcaa660
cacgaccatcacaacttcaaacctatcaaccctcctacctataacatctcgtcaccaatc720
caaccccatcacttcaactctaattaccaatcaatatttaaccaccagggttatggttct780
gcttcgggttccggttctacatacaacaacaataaggaaatgataaagatggagccgtcg840
cttgttagtgtatctcaagaaacctgtctgagctcagatgtgaacgcgaccacgaccatt900
gaggtatcttcgggtccggtaatgaagcaagagatgagtatgacgggaatggtgaatggt960
agcaagtcatatgaagatctatgtgacctgagggggatcttatgggactactga1014
<210>2
<211>337
<212>prt
<213>人工序列
<400>2
metalavalvalvalvalglugluglyvalvalleuasnhisglygly
151015
glugluleuvalaspleuproproglypheargphehisprothrasp
202530
glugluileilethrphetyrleulysglulysvalleuaspserarg
354045
phethralavalalametglyglualaaspleuasnlyscysglupro
505560
trpaspleuprolysargalalysmetglyglulysgluphetyrphe
65707580
phecysglnargasparglystyrprothrglymetargthrasnarg
859095
alathrgluserglytyrtrplysalathrglylysasplysgluile
100105110
phelysglylysglycysleuvalglymetlyslysthrleuvalphe
115120125
tyrargglyargalaproargglyglulysthrasntrpvalmethis
130135140
glutyrargleuaspglylystyrserhishisasnleuprolysser
145150155160
alaargaspglutrpvalvalcysargvalphehislysasnasnpro
165170175
serthrthrthrglnglnmetthrargmetproilegluasnleuthr
180185190
argmetaspserleugluasnileasphisleuleuasppheserser
195200205
leuproproleumetaspproserphethrglyglnhisasphishis
210215220
asnphelysproileasnproprothrtyrasnileserserproile
225230235240
glnprohishispheasnserasntyrglnserilepheasnhisgln
245250255
glytyrglyseralaserglyserglyserthrtyrasnasnasnlys
260265270
glumetilelysmetgluproserleuvalservalserglngluthr
275280285
cysleuserseraspvalasnalathrthrthrilegluvalserser
290295300
glyprovalmetlysglnglumetsermetthrglymetvalasngly
305310315320
serlyssertyrgluaspleucysaspleuargglyileleutrpasp
325330335
tyr
<210>3
<211>1008
<212>dna
<213>人工序列
<400>3
atggcggttgtggttgaagaaggtgtggtgttgaatcatggaggtgaagagcttgtggat60
ttgccacctggtttcaggtttcatccaacagacgaagagatcataacatgttaccttaag120
gagaaggttttaaacagccgattcacggctgtggccatgggagaagctgatctcaacaag180
tgtgagccttgggatttgccaaagagggcaaagatgggggagaaagagttctacttcttc240
tgtcaaagggacaggaagtatccgactgggatgaggacgaaccgtgcgacggagtcagga300
tactggaaagccaccgggaaggataaggagatcttcaaaggcaaaggttgtctcgttggg360
atgaagaaaacacttgtgttttatagaggaagagctccaaaaggtgaaaagactaattgg420
gtcatgcatgaatatcgtcttgaaggcaaatattcgtattacaatctcccaaaatctgca480
agggacgaatgggtcgtgtgtagggtttttcacaagaacaatccttctaccacaacccaa540
ccaatgacgagaatacccgttgaagatttcacaaggatggattctctagagaacattgat600
catctcctagacttctcatctcttcctcctctcatagacccgagtttcatgagtcaaacc660
gaacaaccaaacttcaaacccatcaaccctccaacttacgatatctcatcaccaatccaa720
ccccatcatttcaattcttaccaatcaatctttaaccaccaggtttttggttctgcttcg780
ggctctacgtacaacaacaacaacgagatgatcaagatggagcaatcacttgttagtgta840
tctcaagaaacatgcctaagctcagatgtgaacgcgaacatgactacaaccacggaggta900
tcttcgggtcctgtaatgaaacaagaaatggggatgatgggaatggtgaatggtagcaag960
tcgtatgaagatctatgtgacttgaggggggacttgtgggacttctaa1008
<210>4
<211>335
<212>prt
<213>人工序列
<400>4
metalavalvalvalglugluglyvalvalleuasnhisglyglyglu
151015
gluleuvalaspleuproproglypheargphehisprothraspglu
202530
gluileilethrcystyrleulysglulysvalleuasnserargphe
354045
thralavalalametglyglualaaspleuasnlyscysgluprotrp
505560
aspleuprolysargalalysmetglyglulysgluphetyrphephe
65707580
cysglnargasparglystyrprothrglymetargthrasnargala
859095
thrgluserglytyrtrplysalathrglylysasplysgluilephe
100105110
lysglylysglycysleuvalglymetlyslysthrleuvalphetyr
115120125
argglyargalaprolysglyglulysthrasntrpvalmethisglu
130135140
tyrargleugluglylystyrsertyrtyrasnleuprolysserala
145150155160
argaspglutrpvalvalcysargvalphehislysasnasnproser
165170175
thrthrthrglnprometthrargileprovalgluaspphethrarg
180185190
metaspserleugluasnileasphisleuleuasppheserserleu
195200205
proproleuileaspproserphemetserglnthrgluglnproasn
210215220
phelysproileasnproprothrtyraspileserserproilegln
225230235240
prohishispheasnsertyrglnserilepheasnhisglnvalphe
245250255
glyseralaserglyserthrtyrasnasnasnasnglumetilelys
260265270
metgluglnserleuvalservalserglngluthrcysleuserser
275280285
aspvalasnalaasnmetthrthrthrthrgluvalserserglypro
290295300
valmetlysglnglumetglymetmetglymetvalasnglyserlys
305310315320
sertyrgluaspleucysaspleuargglyaspleutrpaspphe
325330335
<210>5
<211>33
<212>dna
<213>人工序列
<400>5
ttaggtcatgatcgcggttgttgttgttgaaga33
<210>6
<211>34
<212>dna
<213>人工序列
<400>6
cgcgtcgacgtagtcccataagatccctcttaag34
<210>7
<211>26
<212>dna
<213>人工序列
<400>7
ggactagtgcggttgtggttgaagaa26
<210>8
<211>30
<212>dna
<213>人工序列
<400>8
cgcgtcgacgaagtcccacaagtcccccct30
<210>9
<211>25
<212>dna
<213>人工序列
<400>9
caatccaaccccatcacttcaactc25
<210>10
<211>24
<212>dna
<213>人工序列
<400>10
gcgacggctccatctttatcattt24
<210>11
<211>24
<212>dna
<213>人工序列
<400>11
agcctgaggagatattagcaggaa24
<210>12
<211>21
<212>dna
<213>人工序列
<400>12
atctcactgcagctccaccat21
<210>13
<211>21
<212>dna
<213>人工序列
<400>13
gcatctgcctcgacatcttga21
<210>14
<211>21
<212>dna
<213>人工序列
<400>14
gacagcagcaccttggaaatg21
<210>15
<211>25
<212>dna
<213>人工序列
<400>15
gttccaatctatgagggatacacgc25
<210>16
<211>28
<212>dna
<213>人工序列
<400>16
agaagttcgacttgtcattagaaagaaa28
<210>17
<211>23
<212>dna
<213>人工序列
<400>17
ttggttctgcttcgggctctacg23
<210>18
<211>24
<212>dna
<213>人工序列
<400>18
ccattcccatcatccccatttctt24
<210>19
<211>21
<212>dna
<213>人工序列
<400>19
tcgttgggatgaagaaaacac21
<210>20
<211>23
<212>dna
<213>人工序列
<400>20
tcccaaattatcacaatcctttc23
<210>21
<211>23
<212>dna
<213>人工序列
<400>21
atattgaccatcatactcattgc23
- 还没有人留言评论。精彩留言会获得点赞!