分子验证系统的制作方法
本申请要求2016年9月20日提交的美国临时申请号62/396,932、2016年10月12日提交的美国临时申请号62/407,331和2017年5月5日提交的美国临时申请号62/502,522在u.s.c.§119(e)下的权益,它们中的每一篇通过引用整体并入本文。
联邦资助的研究
本发明在美国国防部海军研究办公室(u.s.departmentofdefense,officeofnavalresearch)授予的n00014-16-1-2410、n00014-14-1-0610和n00014-13-1-0593下在政府支持下完成。美国政府具有本发明的某些权利。
背景
越来越复杂的安全措施正在被用于限制对敏感信息、应用和管辖地的权限。例如,许多软件平台现在使用两步验证来确保仅仅适当的个体有权进入该系统。除了具有密码以外,一个人必须同时提供第二种形式的证据来证明他或她有权进入该系统,诸如通过输入被发送至他们的手机的一次性密码(otp)。
生物测定分析作为安全地鉴别个体的身份的方法也已经随着装置(诸如智能电话)中或在进入管辖地(诸如游乐园)的入口处的指纹扫描仪的出现来到最前线。其它类型的生物测定分析着重于个体的特征,诸如眼、手或脸。但是,这些方法中的许多可以被容易地欺骗。例如,手的高分辨率照片可以揭示模仿指纹所必需的特征并因而获得对安全信息的权限。这些当前方法中的一些也可能被视作侵袭性的,且容易受到攻击而暴露敏感的私人信息。
技术实现要素:
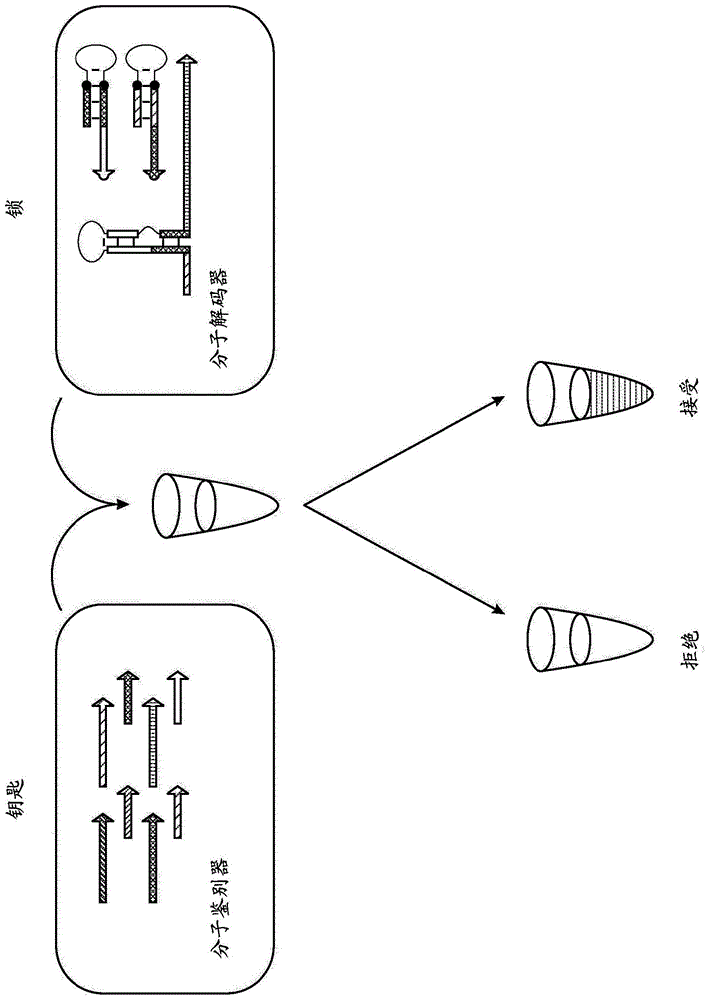
本文提供了分子验证(鉴定)系统、方法和组合物,其使用分子(dna/rna)电路来实现特异性的分子加密溶液。与分子解码器(“锁”)结合使用的分子标识符(“钥匙”)充当分子加密系统来独特地鉴别正确的钥匙溶液所拥有的特定实体(例如,人或产品)(参见,例如,图1)。不同于当前的验证系统,本公开内容的分子验证系统不需要使用私人生物测定特征,因而消除了生物测定分析和储存的敏感的和有时侵袭性的性质。本文提供的分子验证系统仍然维持稳健地和独特地检测特定分子特征的强大能力。
尽管当前的生物测定分析在它们的区分个体的统计学的能力方面可以是非常强大的,但是它们也具有几个功能缺点。一个缺点是,必须为每个个体定制生物测定分析,因而即使验证操作意图应用于大的人群(例如,商业的、政府的或学校的机构),它也必须经过改造以适配每个个体,且因此,难以有效地传播。另一个缺点是,生物测定分析通常需要数据库和服务器来验证个体生物统计学,因而提供了另一个安全漏洞。另一个缺点是,这些生物测定分析倾向于储存无限敏感的私人信息。本文提供的分子验证系统能够在没有储存的情况下完成身份验证并补救当前安全措施的漏洞。
因而,本公开内容提供了使用一组寡核苷酸和有关的分子验证溶液作为稳健且安全鉴别的钥匙和锁系统的系统和方法。数字硬件和软件是不必要的,它们会进一步减小系统的安全性受到攻击的可能。进一步地,在许多情况下,为了在执行验证之前方便的长期储存和运输,可以将反应物干燥。
本公开内容的验证/鉴定系统通常包括一种或多种钥匙组分和一种或多种锁组分。应当理解,在本文的实施方案中的任一个中,一种或多种钥匙组分可以施加于产品或与产品结合,而一种或多种锁组分维持在与所述产品组合或添加给所述产品的溶液中。同样地,在本文的实施方案中的任一个中,一种或多种锁组分可以施加于产品或与产品结合,而一种或多种钥匙组分维持在与所述产品组合或添加给所述产品的溶液中。
本公开内容的一些方面提供了试剂盒或组合物,其包含(a)含有一组催化分子的组合物,每个催化发夹分子包含(i)未配对的3'支点(toehold)结构域,(ii)通过所述分子的3'子结构域和所述分子的5'子结构域之间的分子内核苷酸碱基配对形成的配对茎结构域,和任选的(iii)环结构域,(b)包含引物钥匙的组合物,所述引物钥匙与所述催化分子之一的3'支点结构域互补且与其结合,(c)包含rna支点开关锁的组合物,所述rna支点开关锁包含(i)未配对的支点结构域,其包含与所述催化分子之一的3'子结构域互补的序列,(ii)包含配对茎结构域和环结构域的发夹,所述配对茎结构域包含位于未配对的支点结构域的3'的起始密码子和与所述催化分子之一的3'支点结构域互补的序列,所述环结构域包含核糖体结合位点,和(iii)编码报道蛋白的编码结构域。
本公开内容的其它方面提供了方法,所述方法包括:在包含具有链置换活性的聚合酶和脱氧核糖核苷酸三磷酸(dntp)的第一反应缓冲液中组合(a)一组催化分子,每个催化发夹分子包含(i)未配对的3'支点结构域,(ii)通过所述分子的3'子结构域和所述分子的5'子结构域之间的分子内核苷酸碱基配对形成的配对茎结构域,和(iii)环结构域,和(b)引物钥匙,其与所述催化分子之一的3'支点结构域互补且与其结合;在导致核酸聚合、链置换和退火的条件下温育第一反应混合物足以产生单链核酸触发物的时间;在第二反应缓冲液中组合(c)所述单链核酸触发物,和(d)rna支点开关锁,其包含(i)包含与所述催化分子之一的3'子结构域互补的序列的未配对的支点结构域,(ii)包含配对茎结构域和环结构域的发夹,所述配对茎结构域包含位于未配对的支点结构域的3'的起始密码子和与所述催化分子之一的3'支点结构域互补的序列,所述环结构域包含核糖体结合位点,和(iii)编码报道蛋白的编码结构域,由此形成第二反应混合物;和在导致核酸杂交、rna转录和蛋白翻译的条件下温育所述第二反应混合物足以表达所述报道蛋白的时间。
本公开内容的其它方面提供了试剂盒或组合物,其包含(a)包含下述核酸链的组合物:所述核酸链连接至荧光分子且具有5'结构域和3'结构域,和(b)包含下述核酸链的组合物:所述核酸链连接至猝灭分子且具有(i)与(a)的核酸链的3'结构域互补的5'结构域,和(ii)与(a)的核酸链的5'结构域互补的3'结构域。
本公开内容的其它方面提供了试剂盒或组合物,其包含(a)含有下述核酸发夹分子的组合物:所述核酸发夹分子具有通过5'末端结构域和3'末端结构域的配对而形成的环结构域和茎结构域,其中一个末端结构域包括荧光分子且另一个末端包括猝灭分子;和(b)含有下述核酸发夹分子的组合物:所述核酸发夹分子具有通过5'末端结构域和3'末端结构域的配对而形成的环结构域和茎结构域,其中一个末端结构域包含与(a)的发夹分子的5'末端结构域的序列互补的序列,且另一个末端结构域包含与(a)的发夹分子的3'末端结构域的序列互补的序列,且所述环结构域包含与(a)的环结构域的序列互补的序列。
本公开内容的一些方面提供了试剂盒或组合物,其包含(a)含有催化分子的组合物,所述催化分子具有(i)未配对的支点结构域,(ii)配对的茎结构域,其包含与猝灭分子相对的荧光分子,和任选的(iii)环结构域,和(b)包含核酸链的组合物,所述核酸链具有两个邻近结构域,一个结构域与(a)的催化分子的支点结构域互补且另一个结构域与(a)的催化分子的茎结构域互补。
在某些实施方案中,本文提供了方法,所述方法包括:在反应缓冲液中组合(a)的组合物和组合物(b),和在导致核酸杂交的条件下温育所述反应缓冲液。
本公开内容的另一个方面提供了试剂盒,其包含(a)含有5'结构域和3'结构域的第一核酸链;(b)含有5'结构域的第二核酸链,所述5'结构域与所述第一核酸链的3'结构域互补;(c)第三核酸链,其含有(i)与所述第一核酸链的3'结构域互补的5'结构域和(ii)与所述第一核酸链的5'结构域互补的3'结构域;(d)含有5'结构域和3'结构域的第四核酸链;(e)含有5'结构域的第五核酸链,所述5'结构域与所述第四核酸链的3'结构域互补;(f)第六核酸链,其含有(i)与所述第四核酸链的3'结构域互补的5'结构域和(ii)与所述第四核酸链的5'结构域互补的3'结构域;(g)猝灭剂分子;和(h)荧光分子,其中所述第一核酸链的3'结构域和所述第四核酸链的3'结构域具有彼此不同的核苷酸组成。
在某些实施方案中,所述猝灭剂分子中的至少一个连接至所述第一核酸链的3'结构域和/或连接至所述第四核酸链的3'结构域。在某些实施方案中,所述荧光分子连接至所述第二核酸链的5'结构域和/或连接至所述第五核酸链的5'结构域。在某些实施方案中,存在包含与第二核酸链结合的第一核酸链的第一溶液。在某些实施方案中,存在包含与第五核酸链结合的第四核酸链的第二溶液。
本公开内容的其它方面提供了溶液,其包含(a)第一核酸链,所述第一核酸链含有5'结构域和连接至猝灭剂分子的3'结构域;(b)含有5'结构域的第二核酸链,所述5'结构域连接至荧光分子且结合所述第一核酸链的3'结构域;(c)第三核酸链,其含有(i)与所述第一核酸链的3'结构域互补的5'结构域和(ii)与所述第一核酸链的5'结构域互补的3'结构域;(d)含有5'结构域和3'结构域的第四核酸链,所述3'结构域连接至猝灭剂分子;(e)含有5'结构域的第五核酸链,所述5'结构域连接至荧光分子且结合所述第四核酸链的3'结构域;和(f)第六核酸链,其含有(i)与所述第四核酸链的3'结构域互补的5'结构域和(ii)与所述第四核酸链的5'结构域互补的3'结构域;其中所述第一核酸链的3'结构域和所述第四核酸链的3'结构域具有彼此不同的核苷酸组成。
本公开内容的其它方面提供了基底,其包括(a)至少一个离散区域,该区域包含(i)第一核酸链,所述第一核酸链含有5'结构域和连接至猝灭剂分子的3'结构域,和(ii)含有5'结构域的第二核酸链,所述5'结构域连接至荧光分子且结合所述第一核酸链的3'结构域;和(b)至少一个离散区域,该区域包含(i)第三核酸链,其含有5'结构域和连接至猝灭剂分子的3'结构域,和(ii)含有5'结构域的第四核酸链,所述5'结构域连接至荧光分子且结合所述第三核酸链的3'结构域,其中所述第一核酸链的3'结构域和所述第三核酸链的3'结构域具有彼此不同的核苷酸组成。
本公开内容的另外方面提供了方法,所述方法包括:用图形设计给基底加图案;在所述图形设计上以预定图案沉积以下至少一种:(a)第一核酸链,所述第一核酸链含有5'结构域和连接至猝灭剂分子的3'结构域;和含有5'结构域的第二核酸链,所述5'结构域连接至荧光分子且结合所述第一核酸链的3'结构域;和(b)第三核酸链,其含有5'结构域和连接至猝灭剂分子的3'结构域;和含有5'结构域的第四核酸链,所述5'结构域连接至荧光分子且结合所述第三核酸链的3'结构域。
在某些实施方案中,所述方法进一步包括在所述图形设计上沉积以下至少一种:(c)第五核酸链,其含有(i)与所述第一核酸链的3'结构域互补的5'结构域和(ii)与所述第一核酸链的5'结构域互补的3'结构域;和(d)第六核酸链,其含有(i)与所述第三核酸链的3'结构域互补的5'结构域和(ii)与所述第三核酸链的5'结构域互补的3'结构域。在一个实施方案中,针对荧光来测定所述图形设计。在某些实施方案中,所述测定包括将所述图形设计暴露于透照仪,任选地透过滤波器单元。
附图说明
图1显示了分子验证系统的一个实例。核酸(例如,dna)引物“钥匙”触发分子级联以打开核酸(例如,rna)支点开关“锁”。在该实例中,所述锁编码仅在有正确钥匙存在下才表达的报道分子。因而,与正确钥匙结合的实体会激活颜色变化。
图2a-2b显示了多步引物交换反应(per)的多步的各个步骤(循环)。
图3a-3c显示了per途径的一个实例,其使用一种引物和五种催化发夹分子,从而导致包括6个结构域的触发钥匙的产生。
图4显示了支点开关锁的示意图。对应的tarna具有序列5'-b*-a*-3',其中结构域a*和b*分别是结构域a和b的反向互补物。支点开关锁是rna依赖性的翻译激活剂。在有它的相应核酸触发钥匙存在下,支点开关锁改变它的构象以实现下游可读框(其编码例如报道蛋白)的翻译。
图5a-5e显示了另一种分子验证系统的一个实例,该分子验证系统使用催化发夹系统作为锁来编码荧光分子的募集。
图6a-6e显示了使用引物交换反应实现逻辑计算的分子验证系统的实例。
图7表明dna触发钥匙的效力与质粒表达的rna触发钥匙相当。
图8显示的凝胶证实,仅具有datp、dttp和dctp的3-字母密码例如可以用于合成触发钥匙。
图9表明经由per合成的触发钥匙成功地激活支点开关锁。
图10a-10c显示了用于复制保护验证系统的嵌合的和经修饰的核酸(寡核苷酸)的实例。图10a显示了包含两种互补嵌合寡核苷酸的分子验证系统,一种连接至荧光分子(星号)且一种连接至猝灭分子。一种寡核苷酸与另一种寡核苷酸的杂交导致荧光信号的丧失。所述嵌合寡核苷酸各自包括抑制酶测序的l-dna结构域。图10b显示了包含两种互补催化发夹分子的分子验证系统,其中一种连接至荧光分子和猝灭分子,使得在不与它的互补催化发夹分子结合时荧光被猝灭。仅在所述催化发夹分子彼此杂交后才产生荧光信号。图10c显示了图10a的分子验证系统的一种变体,其中将停止分子(stoppermolecule)包括在嵌合寡核苷酸中以抑制复制和测序。
图11a-11d显示了可以用于使信号序列结构域到一起(例如,使引物钥匙和锁到一起)的机制的实例。图11a显示了通过互补结构域的结合到一起的多个序列,使得'a'和'e'结构域共定位以触发其它过程。图11b显示了通过环-环相互作用(b-b*)到一起的两个发夹环。图11c显示了通过环-环相互作用(b-b*)到一起的两个环形序列。图11d显示了拓扑连接的两个环形序列。
图12a-12d显示了基本支点交换锁/钥匙机制。在图12a中,‘x*a’链结合退火的复合物上的暴露的‘a*’支点,所述退火的复合物最初使溶液中的荧光团(f)与匹配猝灭剂(q)共定位。荧光信号在该构型中被阻断。当所述链通过‘x’结构域替换时,它从猝灭剂释放含有荧光团的链,从而产生荧光溶液。图12b显示了在实施例7中使用的钥匙/锁对。两种含有退火的含荧光团(f)和猝灭剂(q)的链的锁溶液用带圆圈的‘1’和‘2’表示(即,①和②)。图12c显示了从钥匙混合物测得的荧光,所述钥匙混合物含有与等体积的退火的锁溶液(含有在图12b中所示的荧光团/猝灭剂对1)混合的0%至70%的乙醇浓度。不含有钥匙链(浅灰色)或含有错误钥匙链(中灰色,对应于在图12b中所示的荧光团/猝灭剂对2的钥匙链)的钥匙溶液在所有乙醇条件下具有非常小的荧光。当钥匙溶液含有荧光团/猝灭剂对的正确钥匙链(黑色条)时,测量到大量荧光。图12d显示了具有正确钥匙的荧光除以没有钥匙链的荧光的图。在试验的乙醇条件的全范围下观察到成功的钥匙锁对的显著分化(>30倍on/off(开/关)比率)。
图13呈现了在实施例7中描述的实验之一的示意图,从而证实钥匙‘2’特异性地将锁‘2’解锁。
图14a-14b显示了在实施例7中描述的图形设计应用的示意图。图14a显示了在所述实验之一中使用的图形设计的示意图及其结果。图14b显示了用于解释结果的示意图。
图15显示了可以如本文所提供的那样使用的多组分钥匙的实例。在左图中的示意设计显示了1-、2-和3-组分钥匙,且在右图中的图显示了在三种不同的钥匙构型中相对于滴入(dropin)钥匙组分浓度的各个滴入组装状态。
图16显示了为实施例8中所述的成像仪装置使用的蓝色led发射谱(上部)、fam谱(中间)和长通滤波器(mcmaster琥珀色丙烯酸透射率)。
图17显示了实施例8中所述的成像仪装置的示意图和实例。
图18a-18b的示意图描绘了在实施例10中所述的固定放大的机制。图18a显示了额外荧光团和猝灭剂(在这里,各2个)的连接,这导致信号输出(x*)的倍增。图18b显示了一个实例,其中从相同猝灭剂链除去多个荧光团链,导致增加的荧光信号传导(x1*和x2*都用荧光团标记)。
图19a-19c的示意图描绘了在实施例10中所述的固定放大的其它机制。例如,图19a显示了一种初步设置,其中所有发夹作为系统的锁组分存在于溶液中,每个发夹含有荧光团和猝灭剂。图19b显示了它们的杂交链式反应(hcr),因为固定数目的荧光团从单个钥匙链激活。图19c显示了含有荧光团的终产物。
图20a-20b显示了在实施例11中描述的使用未修饰的钥匙链的锁和钥匙系统的实例。图20a显示了一系列独特的未修饰的钥匙链以及测试条,所述测试条用于产生独特条形码图案然后在生产商的网站上验证。图20b显示了一种系统,其中给定的未修饰的钥匙系统是时间敏感的;失效日期和序列号确保鉴定正确的产品。
具体实施方式
本文提供了可以用于鉴定例如一个人、一群人或组合物的身份的分子验证系统(例如,组合物、方法和试剂盒)。将独特的分子钥匙提供给需要身份验证的实体并呈送给拥有分子锁系统的实体。在某些实施方案中,分子验证系统使用四种主要组分:(1)引物钥匙和(2)催化分子,其用于合成(3)触发钥匙,所述触发钥匙可以打开(4)支点开关锁。如在图1中所示,引物钥匙可以被称作分子标识符,且催化分子和支点开关锁一起可以被称作分子解码器。
引物钥匙
“引物钥匙”通常是具有与位于核酸“锁”上的结构域互补的核苷酸序列(结构域)的核酸(例如,单链核酸,也被称作寡核苷酸)。在某些实施方案中,所述锁是催化分子。在其它实施方案中,所述锁是支点开关。在其它实施方案中,所述锁简单地是包含(例如,连接至)功能分子(诸如荧光分子或猝灭分子)的互补核酸。
在某些实施方案中,引物钥匙是具有与催化分子的支点结构域或支点开关锁的支点结构域互补的核苷酸序列(引物结构域)的核酸(例如,单链寡核苷酸)。在某些实施方案中,引物钥匙可以作为典型引物起作用,因为当结合另一种核酸(例如,催化分子或支点开关)时,是在有聚合酶存在下的聚合的起始点。因而,在某些实施方案中,引物钥匙结合催化分子以开始引物交换反应。如在下面更详细地讨论的,在引物交换反应结束时释放的延伸产物被称作‘触发钥匙’。触发钥匙能够打开(线性化)支点开关锁。
在其它实施方案中,引物钥匙简单地是具有至少一个与锁互补的结构域的单链核酸。
在某些实施方案中,引物钥匙比常规核酸引物更短。这有助于通过阻碍引物钥匙序列的‘解码’来防止安全隐患。难以扩增短核酸序列用于随后的测序分析。因而,在某些实施方案中,引物钥匙(或引物钥匙的引物结构域,其为结合催化分子或支点开关锁的支点结构域的核苷酸序列)具有小于18个核苷酸的长度。例如,引物钥匙可以具有4-17、4-16、4-15、4-14、4-13、4-12、4-11、4-10、4-9、4-8、5-17、5-16、5-15、5-14、5-13、5-12、5-11、5-10、5-9、5-8、6-17、6-16、6-15、6-14、6-13、6-12、6-11、6-10、6-9、6-8、7-17、7-16、7-15、7-14、7-13、7-12、7-11、7-10、7-9、7-8、8-17、8-16、8-15、8-14、8-13、8-12、8-11、8-10、8-9、9-17、9-18、9-15、9-14、9-13、9-12、9-11、9-10、10-17、10-16、10-15、10-14、10-13、10-12、10-11、10-10、10-9或10-8个核苷酸的长度。在某些实施方案中,引物钥匙具有4、5、6、7、8、9、10、11、12、13、14、15、16、17或18个核苷酸的长度。在某些实施方案中,引物钥匙具有小于15个核苷酸的长度。
在某些实施方案中,例如,在其中不使用引物交换反应的实施方案中,引物钥匙(或引物钥匙的引物结构域)具有大于15个核苷酸的长度。在某些实施方案中,引物钥匙(或引物钥匙的引物结构域)具有10-50个核苷酸的长度。例如,引物钥匙(或引物钥匙的引物结构域)可以具有10-45、10-40、10-35、10-30、10-25、10-20、10-15、15-50、15-45、15-40、15-35、15-30、15-25、15-20、20-50、20-45、20-40、20-35、20-30、20-25、25-50、25-45、25-40、25-35、25-30、30-50、30-45、30-40、30-35、35-50、35-45、35-40、40-50、40-45或45-50个核苷酸的长度。在某些实施方案中,引物钥匙(或引物钥匙的引物结构域)具有10、15、20、25、30、35、40、45或50个核苷酸的长度。在某些实施方案中,引物钥匙(或引物钥匙的引物结构域)具有15、16、17、18、19、20、21、22、23、24或25个核苷酸的长度。在某些实施方案中,引物钥匙(或引物钥匙的引物结构域)长于50个核苷酸,或短于10个核苷酸。应当理解,引物钥匙(或引物钥匙的引物结构域)的长度至少部分地取决于催化分子或支点开关锁的支点结构域的长度。
引物钥匙可以存在于水溶液中或以干燥(冻干的)形式存在。在某些实施方案中,引物钥匙存在于墨液中,例如,被需要身份鉴定的人携带在钢笔中。在某些实施方案中,引物钥匙存在于醇(例如,乙醇)溶液中,诸如含有某种百分比的醇的香味剂(例如,香料)。例如,醇(例如,乙醇)溶液可以包括至少5%、10%、20%、30%、40%、50%、60%、70%、80%、90%或100%的醇。在某些实施方案中,引物钥匙存在于70%醇(例如,乙醇)溶液中。在某些实施方案中,将在溶液(例如,水性缓冲液)中的引物钥匙施加于(例如,喷在上面)产品或其它组合物/实体并干燥用于长期储存和运输。以此方式,可以将产品(例如,市售产品)用独特身份‘编码’。在某些实施方案中,所述引物钥匙没有与产品(例如,香料)混合,且可以存在于标签(诸如纸)上,所述标签附着于含有所述产品的器皿(即,容器)。在这样的实施方案中,可以将液体形式的锁溶液施加于包含钥匙的标签,且可见的颜色变化指示锁-钥匙匹配。所述系统也可以反过来设计:钥匙可以以液体形式存在,且锁可以存在于标签上。在某些实施方案中,所述标签没有连接至器皿。
在某些实施方案中,将引物钥匙与额外分子组合以进一步掩饰引物钥匙序列的身份。这样的额外分子包括,例如,随机序列寡核苷酸、蛋白或肽。
在某些实施方案中,改变引物钥匙与额外分子的比率。
在某些实施方案中,改变引物钥匙的浓度。例如,可能需要引物钥匙的阈值浓度或确切浓度来成功地结合催化分子或支点开关锁(打开锁)。例如,可能需要刚好2pm特定引物钥匙来打开锁,且更高或更低的浓度不能打开锁。
在某些实施方案中,引物钥匙在引物钥匙的核苷酸序列内包括抑制复制和/或测序的分子。例如,在某些实施方案中,引物钥匙包括“停止子(stopper)”,诸如间隔物、合成的核苷酸和修饰的核苷酸。修饰的核苷酸或核苷酸结构域(超过一个连续核苷酸)的例子包括、但不限于xna、pna和lna。在本文别处描述了其它停止子(终止聚合的分子或修饰)。在某些实施方案中,所述引物钥匙可以包含未修饰的(即,天然存在的)核苷酸碱基。
在某些实施方案中,引物钥匙包括l-dna。d-dna是形成规范右螺旋的天然存在的dna形式,尽管可以化学地合成它的镜像l-dna。还可以合成嵌合分子,其部分地包含d-dna且部分地包含l-dna。l-dna结构域通常不会被聚合酶识别。因此,在某些实施方案中,它们可以充当酶测序方法的有效抑制剂。因而,本文提供了包含l-dna结构域和d-dna的嵌合引物钥匙。这些嵌合引物的一个例子显示在图10a-10c中。在本文中包括其它嵌合引物钥匙。
在某些情况下,经稀释的物质(例如,香料)在加密试验下可以是试验阳性的,因为有某种量的“真实”物质存在。为了与该溶液斗争,可以将“钥匙”分成通过弱杂交保持在一起的多个组分,如在图15中所示。不希望受理论的约束,这可能使得钥匙功能更依赖于组分浓度。对于单组分钥匙,10倍滴入钥匙浓度会在试验中产生1/10x浓度的钥匙。但是,在2-组分系统中,相同的10倍滴入产生例如100倍滴入功能性的(组装的)钥匙,从而减小了错误鉴别经稀释的伪品的机会。在某些实施方案中,相同的10倍滴入可能产生20倍至1000倍(例如,30倍,40倍,50倍,60倍,70倍,80倍,90倍,100倍,150倍,200倍,250倍,300倍,350倍,400倍,450倍,500倍,550倍,600倍,650倍,700倍,750倍,800倍,850倍,900倍,950倍)或更大的滴入功能性的(组装的)钥匙,这至少部分地取决于序列设计和反应条件。较多的组分会导致较大的效果,如在图15中的3-组分模型所证实的。通过改变dna序列和结构域长度,也可以在设计中任意调节使钥匙稳定的链相互作用。
因而,在某些实施方案中,钥匙是包含2、3个或更多个彼此弱结合的链的复合物。在某些实施方案中,钥匙包含(a)含有5'结构域和3'结构域的第一核酸链,和(b)含有5'结构域和3'结构域的第二核酸链,其中所述第二核酸链的3'结构域结合(例如,弱结合)所述第一核酸链的5'结构域,其中所述第一核酸链的3'结构域与连接至荧光分子的核酸链互补。在其它实施方案中,钥匙包含(a)含有5'结构域和3'结构域的第一核酸链,(b)含有5'结构域和3'结构域的第二核酸链,和(c)含有5'结构域和3'结构域的第三核酸链,其中所述第二核酸链的3'结构域结合(例如,弱结合)所述第一核酸链的5'结构域,其中所述第二核酸链的5'结构域结合(例如,弱结合)所述第三核酸链的3'结构域,且其中所述第一核酸链的3'结构域与连接至荧光分子的核酸链互补。
催化分子
在某些实施方案中,将催化分子用作锁系统的组分,以从起始引物钥匙合成触发钥匙。催化核酸分子(“催化分子”)通常包括未配对的(单链的)3'支点结构域和在3'支点结构域的5'侧(且,在某些实施方案中,直接地邻近)的配对的(双链的)结构域。“催化发夹分子”也包括环结构域。通过例如调整催化分子的长度、组成和浓度(例如,催化分子的一个或多个结构域),可以控制引物交换反应的动力学。
催化发夹(参见图2a作为一个示例性例子)包括与配对的发夹茎结构域(例如,通过子结构域“2”与子结构域“2'”的分子内结合而形成)连接的未配对的3'支点结构域(“1'”),所述配对的发夹茎结构域连接至发夹环结构域(环样结构)。术语“未配对的”在本文中用于表示没有结合互补核酸/序列的核酸/核苷酸序列,更具体地是核酸的结构域。在某些实施方案中,未配对的结构域可以简单地是在单链核酸上的区域。在其它实施方案中,未配对的结构域是在部分双链分子(包括配对的结构域和未配对的结构域的核酸分子)上的区域。部分双链分子的未配对的结构域的一个例子是支点结构域,例如,如在图2a中显示的(结构域1')。同样地,术语“配对的”在本文中用于表示结合互补核酸/序列的核酸/核苷酸序列,更具体地是核酸的结构域。部分双链分子的配对的结构域的一个例子是茎结构域,例如,如在图2a中显示的(与结构域2结合的结构域2')。
催化分子(例如,催化发夹分子)的长度可以变化。在某些实施方案中,催化分子具有25-300个核苷酸的长度。例如,催化分子可以具有25-250、25-200、25-150、25-100、25-50、50-300、50-250、50-200、50-150或50-100个核苷酸的长度。在某些实施方案中,催化分子具有30-50、40-60、50-70、60-80、70-90、80-100、100-125、100-150或100-200个核苷酸的长度。在某些实施方案中,催化分子具有35、36、37、38、39、40、41、42、43、44、45、46、47、48、49.50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99或100个核苷酸的长度。在某些实施方案中,催化分子长于300个核苷酸,或短于25个核苷酸。
“支点结构域”表示位于催化分子的3'末端处的未配对的核苷酸序列,且与引物钥匙(或引物钥匙的引物结构域)的核苷酸序列互补(和结合)。支点结构域的长度可以变化。在某些实施方案中,支点结构域具有4-17、4-16、4-15、4-14、4-13、4-12、4-11、4-10、4-9、4-8、5-17、5-16、5-15、5-14、5-13、5-12、5-11、5-10、5-9、5-8、6-17、6-16、6-15、6-14、6-13、6-12、6-11、6-10、6-9、6-8、7-17、7-16、7-15、7-14、7-13、7-12、7-11、7-10、7-9、7-8、8-17、8-16、8-15、8-14、8-13、8-12、8-11、8-10、8-9、9-17、9-18、9-15、9-14、9-13、9-12、9-11、9-10、10-17、10-16、10-15、10-14、10-13、10-12、10-11、10-10、10-9或10-8个核苷酸的长度。在某些实施方案中,支点结构域具有4、5、6、7、8、9、10、11、12、13、14、15、16、17或18个核苷酸的长度。在某些实施方案中,支点结构域具有5-40个核苷酸的长度。例如,支点结构域可以具有2-35、2-30、2-25、2-20、2-15、2-10、5-35、5-30、5-25、5-20、5-15、5-10、10-40、10-35、10-30、10-25、10-20、10-15、15-40、15-35、15-30、15-25、15-20、20-40、20-35、20-30、20-25、25-40、25-35、25-30、30-40、30-35或35-40个核苷酸的长度。在某些实施方案中,支点结构域具有5、10、15、20、25、30、35或40个核苷酸的长度。在某些实施方案中,支点结构域具有10、11、12、13、14、15、16、17、18、19、20、21、22、23、24或25个核苷酸的长度。在某些实施方案中,支点结构域长于40个核苷酸,或短于5个核苷酸。
在某些实施方案中,引物钥匙(或引物钥匙的引物结构域)结合催化分子的3'未配对的(单链的)支点结构域以起始引物交换反应。在该反应(参见图2a作为一个示例性例子)中,引物钥匙(“1”)结合催化分子的支点结构域(“1'”),并且存在于反应溶液中的链置换聚合酶对引物钥匙的延伸会通过分支迁移过程替换催化分子的茎结构域的子结构域之一(“2”)。总效果是发夹茎结构域的子结构域之一(“2”)被延伸的(新合成的)引物结构域替换。
催化分子的“配对的结构域”或“茎结构域”表示位于催化分子的未配对的支点结构域的5'侧(且,在某些实施方案中,直接邻近)的配对的核苷酸序列(例如,沃森-克里克核苷碱基配对)。催化发夹分子的配对茎结构域通过催化发夹分子的2个子结构域的分子内碱基配对(在同一分子内的核苷酸之间的碱基配对)而形成:例如,位于支点结构域的5'侧的内部/中央子结构域与位于催化发夹的5'末端处的子结构域结合(杂交)。配对的结构域的长度可以变化。在某些实施方案中,配对的结构域具有5-40个核苷酸的长度。例如,配对的结构域可以具有5-35、5-30、5-25、5-20、5-15、5-10、10-40、10-35、10-30、10-25、10-20、10-15、15-40、15-35、15-30、15-25、15-20、20-40、20-35、20-30、20-25、25-40、25-35、25-30、30-40、30-35或35-40个核苷酸的长度。在某些实施方案中,配对的结构域具有5、10、15、20、25、30、35或40个核苷酸的长度。在某些实施方案中,配对的结构域具有10、11、12、13、14、15、16、17、18、19、20、21、22、23、24或25个核苷酸的长度。在某些实施方案中,配对的结构域长于40个核苷酸,或短于5个核苷酸。
尽管配对的结构域通常通过催化分子(或支点开关,如下面讨论的)的2个子结构域的分子内碱基配对而形成,但是应当理解,配对的茎结构域可以含有至少一个错配对(例如,a与c或g的配对,或t与c或g的配对)。在某些实施方案中,所述茎结构域具有1-5个错配核苷酸碱基对。例如,配对的结构域可以具有1、2、3、4或5个错配核苷酸碱基对。
催化发夹的“环结构域”表示在茎结构域的末端处(邻近它)形成环样结构的基本上未配对的核苷酸序列。环结构域的长度可以变化。在某些实施方案中,环结构域具有3-200个核苷酸的长度。例如,环结构域可以具有3-175、3-150、3-125、3-100、3-75、3-50、3-25、4-175、4-150、4-125、4-100、4-75、4-50、4-25、5-175、5-150、5-125、5-100、5-75、5-50或5-25个核苷酸的长度。在某些实施方案中,环结构域具有3-10、3-15、32-10、3-25、3-30、3-35、3-40、3-35、3-40、3-45、3-50、4-10、4-15、4-10、4-25、4-30、4-35、4-40、4-35、4-40、4-45或4-50个核苷酸的长度。在某些实施方案中,环结构域具有3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、45、46、47、48、49或50个核苷酸的长度。在某些实施方案中,环结构域长于300个核苷酸。
在某些实施方案中,催化分子不含有发夹环结构域。例如,催化分子可以简单地是包含与配对的结构域邻近的3'未配对的支点结构域的双链体,类似于茎结构域(没有邻近环结构域)。不包括环结构域的催化分子可以在与3'支点结构域相对的末端处通过一段(例如,10个或更多个)核苷酸碱基对之间的交联或核苷酸碱基互补性而得以稳定。
“一组催化分子”指示触发钥匙的合成。因而,一组催化分子通常包括打开支点开关锁所需要的触发钥匙的所有结构域。在某些实施方案中,一组催化分子包括3-20个催化分子。例如,一组催化分子可以包括3、4、5、6、7、8、9、10、12、15或20个催化分子。在某些实施方案中,一组催化分子包括3-5、3-10、3-15、5-10、5-15或5-20个催化分子。一组催化分子通常彼此不同,因为每个茎结构域包括相对于该组的其它催化分子的茎结构域不同的序列。但是,每个催化分子的支点结构域包括与该组的仅一个其它催化分子的茎结构域互补的序列。该规则的例外是引物钥匙最初结合的催化分子。如在图3a中所示,引物‘a’结合催化分子‘a’的结构域a*。关于该组的其它催化分子,支点结构域序列与该组的一个其它催化分子的茎序列相同(或互补)。催化分子‘b’的支点结构域‘b*’与催化分子‘a’的结构域‘b’互补。催化分子‘c’的支点结构域‘c*’与催化分子‘b’的结构域‘c’互补。催化分子‘d’的支点结构域‘d*’与催化分子‘c’的结构域‘d’互补。催化分子‘e’的支点结构域‘c*’与催化分子‘d’的结构域‘e’互补。
在某些实施方案中,所述催化分子组可以以程序化的方式(即,杂交链式反应,hcr)用于放大荧光信号。如在图19a-19c中所示,发夹可以保持一个或多个荧光团和/或猝灭剂部分,然后经历上述的per,从而产生具有扩大的和程序化的荧光水平的双链产物。应当指出,发夹/无活性状态和活化的聚合物状态都是热力学平衡状态,从而提供在储存中的系统稳定性。
触发钥匙
触发钥匙能够打开支点开关锁(改变支点开关锁的发夹/茎结构域)。这些触发钥匙以反式工作,因而可以在本文中称作反式激活核酸(例如,反式激活rna或tarna)。
在某些实施方案中,使用包括一种引物钥匙和“一组催化分子”的引物交换反应合成触发钥匙。该组的每个催化分子包括与支点开关锁上的结构域对应的结构域,因而每个催化分子“指定”反向互补结构域,所述反向互补结构域与特定支点开关锁上的结构域互补且能够与其结合。
触发钥匙可以是dna或rna,或dna和rna二者的组合。在某些实施方案中,所述触发钥匙是mrna。在某些实施方案中,所述触发钥匙是rrna或任意其它非编码rna(例如,srna)。
普通技术人员将能够鉴别合适的触发钥匙序列。这可以例如如下完成:针对它们的特异性地和选择性地仅结合目标触发物的能力来筛选开关支点结构域序列。这样的筛选可以在特定条件下执行,所述特定条件模仿所述开关将结合它的相应触发物的条件。
触发钥匙的长度至少部分地取决于支点开关锁的长度。在某些实施方案中,触发钥匙具有20-100个核苷酸的长度。例如,触发钥匙可以具有20-90、20-80、20-70、20-60、20-50、20-40、20-30、30-100、30-90、30-80、30-70、30-60、30-50、30-40、40-100、40-90、40-80、40-70、40-60、40-50、50-100、50-90、50-80、50-70、50-60、60-100、60-90、60-80、60-70个核苷酸的长度。在某些实施方案中,触发钥匙具有20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95或100个核苷酸的长度。在某些实施方案中,触发钥匙具有长于100个核苷酸的长度。
所述触发物可以包含第一结构域和第二结构域,所述第一结构域与前述核糖调节剂(riboregulator)开关中的任一种的支点结构域杂交且不包含二级结构或包含最小二级结构,所述第二结构域与支点结构域的下游(3')序列(为茎结构域做出贡献的序列,任选地包括茎结构域的第一配对区域)杂交。第一结构域可以与支点结构域具有100%的互补性。第二结构域可以与支点结构域的下游序列具有小于100%的互补性。
所述触发物可以由超过一个核酸链组成,且这样的组合的多链提供第一和第二结构域用于与支点核糖调节剂开关杂交。在某些实施方案中,可以使用一种或多种其它核酸使多种触发物(或部分触发物序列)通过杂交紧密靠近以使它们能够与核糖调节剂开关有效地杂交。
在某些实施方案中,触发钥匙与支点开关锁的支点结构域和第一配对结构域杂交且不与未配对的凸起杂交。
触发钥匙可以包含二级结构,例如发夹结构,只要这样的发夹结构不会干扰触发钥匙与开关锁或彼此的杂交。
荧光分子和猝灭分子
在某些实施方案中,本公开内容的核酸包括(至少一种)荧光分子,诸如荧光团。荧光分子的非限制性例子包括5-fam、钙黄绿素、dio、荧光素、fluo-3、fluo-4、egfp、gfp、俄勒冈绿514、quantifluortmdsdna、quantifluortmssdna、quantifluortmrna、罗丹明绿、sybr金、sybr绿、syto9、
在某些实施方案中,本公开内容的核酸包括(至少一种)猝灭分子。猝灭表示减小给定物质的荧光强度的任何过程。多种过程可以导致猝灭,诸如激发态反应、能量转移、复合物形成和碰撞猝灭。分子氧、碘离子和丙烯酰胺是化学猝灭剂的非限制性例子。氯离子是众所周知的奎宁荧光猝灭剂。
当将荧光分子和猝灭剂分子用在本公开内容的钥匙/锁鉴定系统中时,在某些实施方案中,至少一种荧光分子定位在核酸钥匙链上,且至少一种分子定位在核酸锁链上,使得当钥匙链结合锁链时,荧光分子和猝灭剂分子彼此紧密靠近(例如,紧靠),从而导致否则由荧光分子发出的荧光信号的猝灭。在其它实施方案中,至少一种荧光分子定位在核酸锁链上,且至少一种分子定位在核酸钥匙链上,使得当然后钥匙链结合锁链时,荧光分子和猝灭剂分子彼此紧密靠近,从而导致荧光信号的猝灭。
在某些实施方案中,荧光分子和猝灭剂分子都位于相同锁链或相同钥匙链上,且将所述链构造成使得荧光分子和猝灭剂分子彼此紧密靠近,从而导致荧光信号的猝灭。例如,含有荧光分子和猝灭剂分子的链可以形成发夹(参见,例如,图10b)。
应当理解,在本文提供的任何实施方案(其中含有荧光分子的核酸链结合含有猝灭剂分子的核酸链)中,在核酸链(钥匙链或锁链)上的猝灭剂分子可以置换荧光分子,只要在互补链上的荧光分子用猝灭剂分子置换,使得两条链的结合导致荧光信号的猝灭。同样地,在核酸链(钥匙链或锁链)上的荧光分子可以置换猝灭剂分子,只要在互补链上的猝灭剂分子用荧光分子置换,使得两条链的结合导致荧光信号的猝灭。
在某些实施方案中,钥匙/锁系统(包括组合物、试剂盒和/或方法)包括(a)包含5'结构域和3'结构域的第一核酸链(例如,钥匙链),(b)第二核酸链(例如,锁链),其包含与第一链的3'结构域互补的5'结构域、与第一链的5'结构域互补的3'结构域,和彼此被第二链的3'结构域的x个核苷酸隔开的两个猝灭剂分子,和(c)第三核酸链(例如,另一个锁链),其包含与第二链的3'结构域互补的5'结构域,和彼此被第三链的5'结构域的x个核苷酸隔开的两个荧光分子,其中所述第三核酸链与所述第二核酸链的结合猝灭由两个荧光分子发出的荧光信号,且x大于0(参见,例如,图18a)。
在某些实施方案中,所述第二核酸链包含荧光分子,且所述第三核酸链包含猝灭剂分子。因而,在某些实施方案中,钥匙/锁系统(包括组合物、试剂盒和/或方法)包括(a)包含5'结构域和3'结构域的第一核酸链(例如,钥匙链),(b)第二核酸链(例如,锁链),其包含与第一链的3'结构域互补的5'结构域、与第一链的5'结构域互补的3'结构域,和彼此被第二链的3'结构域的x个核苷酸隔开的两个荧光分子,和(c)第三核酸链(例如,另一个锁链),其包含与第二链的3'结构域互补的5'结构域,和彼此被第三链的5'结构域的x个核苷酸隔开的两个猝灭剂分子,其中所述第三核酸链与所述第二核酸链的结合猝灭由两个荧光分子发出的荧光信号,且x大于0。
在某些实施方案中,x是1-50、2-50、3-50、4-50、5-50、1-20、2-20、3-20、4-20、5-20、1-10、2-10、3-10、4-10或5-10个核苷酸。
在某些实施方案中,钥匙/锁系统(包括组合物、试剂盒和/或方法)包括(a)包含第一5'子结构域、第二5'子结构域和3'结构域的第一核酸链,(b)第二核酸链,其包含与第一链的3'结构域互补的5’结构域、与第一链的第二5'子结构域互补的第一3'子结构域、与第一链的第一5'子结构域互补的第二3'子结构域和将第二链的第一3'子结构域与第二链的第二3'子结构域隔开的猝灭剂分子,(c)第三核酸链,其包含与第二链的第一3'子结构域互补的5'结构域和5'荧光分子,和(d)第四核酸链,其包含与第二链的第二3'子结构域互补的5'结构域和3'荧光分子,其中所述第三核酸链和所述第四核酸链与所述第二核酸链的结合猝灭由5'荧光分子和3'荧光分子发出的荧光信号(参见,例如,图18b)。
在某些实施方案中,钥匙/锁系统(包括组合物、试剂盒和/或方法)包括(a)包含第一5'子结构域、第二5'子结构域和3'结构域的第一核酸链,(b)第二核酸链,其包含与第一链的3'结构域互补的5’结构域、与第一链的第二5'子结构域互补的第一3'子结构域、与第一链的第一5'子结构域互补的第二3'子结构域和将第二链的第一3'子结构域与第二链的第二3'子结构域隔开的荧光分子,(c)第三核酸链,其包含与第二链的第一3'子结构域互补的5'结构域和5'猝灭剂分子,和(d)第四核酸链,其包含与第二链的第二3'子结构域互补的5'结构域和3'猝灭剂分子,其中所述第三核酸链和所述第四核酸链与所述第二核酸链的结合猝灭由荧光分子发出的荧光信号。
在某些实施方案中,钥匙/锁系统(包括组合物、试剂盒和/或方法)包括(a)第一核酸链,其包含5'结构域、3'结构域和5'荧光分子或5'猝灭剂分子,和(b)第二核酸链,其包含与第一链的3'结构域互补的5'结构域、与第一链的5'结构域互补的3'结构域和3'猝灭剂分子或3'荧光分子,其中所述第二核酸链与所述第一核酸链的结合猝灭由5'荧光分子或3'荧光分子发出的荧光信号(参见,例如,图10a)。在某些实施方案中,所述第一链包含5'荧光分子,且所述第二链包含3'猝灭剂分子,并且所述第一链与所述第二链的结合猝灭由5'荧光分子发出的荧光信号。在其它实施方案中,所述第一链包含5'猝灭剂分子,且所述第二链包含3'荧光分子,并且所述第一链与所述第二链的结合猝灭由3'荧光分子发出的荧光信号。
在某些实施方案中,钥匙/锁系统(包括组合物、试剂盒和/或方法)包括(a)第一核酸链,其包含5'结构域、环结构域和与所述5'结构域互补的3'结构域,和(b)第二核酸链,其包含5'末端、5'结构域、环结构域、与所述5'结构域互补的3'结构域和3'末端,其中一个末端包含荧光分子且另一个末端包含猝灭剂分子,其中第二链的5'结构域与第二链的3'结构域的结合猝灭由荧光分子发出的荧光信号(参见,例如,图10b(上部))。在某些实施方案中,钥匙/锁系统(包括组合物、试剂盒和/或方法)包括(a)包含5'结构域和3'结构域的第一核酸链,和(b)第二核酸链,其包含5’结构域、环结构域、与第二链的5'结构域互补且与第一链的3'结构域互补的第一3'子结构域和与第一链的5'结构域互补的第二3'子结构域,其中所述环结构域侧接荧光分子和猝灭剂分子,且第二链的5'结构域与第二链的第一3'子结构域的结合猝灭由荧光分子发出的荧光信号(参见,例如,图10b(中间))。
在某些实施方案中,钥匙/锁系统(包括组合物、试剂盒和/或方法)包括(a)包含5'结构域和3'结构域的第一核酸链,(b)第二核酸链,其包含含有猝灭剂分子的5'末端、与第一链的3'结构域互补的5'结构域和与第一链的5'结构域互补的3'结构域,和(c)第三核酸链,其包含与第二链的5'结构域互补的结构域和含有荧光分子的3'末端,其中第二链与第三链的结合猝灭由荧光分子发出的荧光信号(参见,例如,图10b(底部))。
引物交换反应
在引物交换反应(per)过程中,使用链置换聚合酶和催化地起作用的部分地配对的分子(例如,发夹分子),将离散的核苷酸序列(结构域)添加(合成)至“生长中的”核酸链(参见,例如,图2a-2b)。基本引物交换反应以三个一般步骤发生。首先,引物(例如,引物钥匙)(结构域1)可逆地结合促进延伸的催化分子(例如,发夹分子)。然后,链置换聚合酶延伸引物以复制催化分子中的茎序列(结构域2'),直到到达停止序列(或其它终止聚合的分子)。在延伸已经终止以后,所述分子的置换的茎区域可以与在所述分子上的它的相对链重新杂交以置换引物序列至它可以自发地从催化分子解离且自由地与溶液中的另一种相应催化分子相互作用的点。该引物交换反应能够以特异性的可编程的方式将序列附加到生长中的链上,以产生例如触发钥匙。这些反应都等温地进行,且由溶液中的dntp提供动力。通过用一个分子催化的反应的输出引物序列充当另一个反应的输入引物,可以容易地将引物交换反应连接到一起。引物交换反应描述在2016年2月17日提交的美国临时申请号62/296310和2016年2月24日提交的美国临时申请号62/299206中,它们中的每一篇通过引用整体并入本文。
在某些实施方案中,所述催化分子中终止聚合的分子或修饰的存在会终止置换聚合酶对引物钥匙的延伸。因而,在某些实施方案中,本公开内容的催化分子包含终止聚合的分子或修饰。终止聚合的分子或修饰(“停止子”)通常位于催化分子的配对的结构域(例如,茎结构域)中,使得聚合通过配对的结构域终止引物的延伸。对于以发夹形式排列的催化分子,终止聚合的分子或修饰可以位于配对茎结构域和环结构域之间。在某些实施方案中,终止聚合的分子是合成的非-dna接头,例如,三甘醇间隔物,诸如int间隔物9(isp9)或间隔物18(integrateddnatechnologies(idt))。应当理解,可以如本文中提供的使用终止聚合酶的聚合的任何非天然接头。这样的分子和修饰的其它非限制性例子包括三碳连接(/ispc3/)(idt)、acryditetm(idt)、腺苷酰化、叠氮化物、地高辛配基(nhs酯)、胆甾醇基-teg(idt)、i-linkertm(idt)和3-氰基乙烯基咔唑(cnvk)及其变体。通常,但并非总是这样,短接头(例如,isp9)导致较快的反应时间。
在某些实施方案中,终止聚合的分子是单个或配对的非天然的核苷酸序列,诸如异-dg和异-dc(idt),它们分别是胞嘧啶和鸟嘌呤的化学变体。异-dc将与异-dg进行碱基配对(氢键),但是不与dg进行碱基配对。类似地,异-dg将与异-dc进行碱基配对,但是不与dc进行碱基配对。通过在停止子位置处将这些核苷酸成对掺入发夹的相对侧,聚合酶将被停止,因为它不具有在溶液中的互补核苷酸来添加在该位置。
在某些实施方案中,通过将反应中的dntp浓度(例如,从200μm)降低至100μm、10μm、1μm或更低,提高“停止子”修饰的执行效率。
终止聚合的分子或修饰的包含经常会在催化分子的配对的结构域(例如,发夹结构的茎结构域)中建立“凸起”,因为所述分子或修饰没有配对。因而,在某些实施方案中,将催化分子设计成包括与所述分子或修饰相对的单个核苷酸(例如,胸腺嘧啶),至少两个相同的核苷酸(例如,胸腺嘧啶二聚体(tt)或三聚体(ttt))或非天然的修饰。
在图2a中描绘了引物交换反应的完整“步骤”。引物钥匙(“1”)结合催化分子的支点结构域(“1'”)以起始引物交换反应。在与含有聚合酶(例如,链置换聚合酶)和dntp的反应溶液中的催化分子结合后,所述引物钥匙通过配对的结构域延伸,从而置换配对的结构域的子结构域(“2”)。被置换的子结构域(“2”)然后与延伸的引物(“1+2”)竞争与它的互补子结构域(“2'”)的结合(重新退火),由此置换延伸的输出引物“1+2”。这完成引物交换反应的一个步骤。被置换的输出引物“1+2”然后可以作为输入引物在反应的下一步中继续起作用。
例如,如在图2b中所示,被置换的输出引物“1+2”在引物交换反应的另一个步骤中充当输入引物,通过它的引物结构域“2”结合另一种催化分子的支点结构域“2'”,由此起始引物交换反应中的另一个步骤。在与含有聚合酶和dntp的反应溶液中的催化分子结合后,输入引物“1+2”通过配对的结构域延伸,从而置换配对的结构域的子结构域(“3”)。被置换的子结构域(“3”)然后与延伸的引物(“1+2+3”)竞争与它的互补子结构域(“3'”)的结合(重新退火),由此置换延伸的输出引物“1+2+3”。这完成引物交换反应的另一个步骤。被置换的输出引物“1+2+3”然后可以作为输入引物在反应的下一步中继续起作用。
引物交换反应需要使用聚合酶。在某些实施方案中,所述聚合酶是dna聚合酶(dnap),诸如具有dna链置换活性的dna聚合酶(链置换聚合酶)。“链置换”描述了置换在合成过程中遇到的下游dna的能力。可以如本文所提供的使用的具有dna链置换活性的聚合酶的例子包括,但不限于,phi29dna聚合酶(例如,neb#m0269)、bstdna聚合酶大片段(例如,neb#m0275)或bsudna聚合酶大片段(例如,neb#m0330)。可以使用具有链置换活性的其它聚合酶。在某些实施方案中,所述聚合酶是rna聚合酶。
在某些实施方案中,所述聚合酶是phi29dna聚合酶。在这样的实施方案中,反应条件可以如下:补充了经纯化的牛血清白蛋白(bsa)的1x反应缓冲液(例如,50mmtris-hcl,10mmmgcl2,10mm(nh4)2so4,4mmdtt),ph7.5,在30℃温育。
在某些实施方案中,所述聚合酶是bstdna聚合酶大片段。在这样的实施方案中,反应条件可以如下:1x反应缓冲液(例如,20mmtris-hcl,10mm(nh4)2so4,10mmkcl,2mmmgso4,0.1%
在某些实施方案中,所述聚合酶是bsudna聚合酶。在这样的实施方案中,反应条件可以如下:1x反应缓冲液(例如,50mmnacl,10mmtris-hcl,10mmmgcl2,1mmdtt),ph7.9,在37℃温育。
在引物交换反应系统中的引物钥匙、催化分子和dntp的浓度可以随例如特定应用和该特定应用所需的动力学而变化。
在引物交换反应中的引物钥匙的浓度可以是,例如,10nm至1000nm。在某些实施方案中,在引物交换反应中的引物钥匙浓度是10-20、10-30、10-40、10-50、10-60、10-70、10-80、10-90、10-100、10-125、10-150、10-200、25-50、25-75、25-100、25-150、25-200、50-75、50-100、50-150或50-200nm。在某些实施方案中,在引物交换反应中的引物钥匙浓度是100-200、100-300、100-400、100-500、100-600、100-70、100-800、100-900或100-1000nm。在某些实施方案中,在引物交换反应中的引物钥匙浓度是10、15、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、100、105、110、115、120、125、130、135、140、145、150、155、160、165、170、175、180、185、190、195或200nm。在某些实施方案中,在引物交换反应中的引物钥匙浓度是100、200、300、400、500、600、700、800、900或1000nm。在引物交换反应中的引物钥匙浓度可以小于10nm或大于1000nm。
在引物交换反应中的催化分子(例如,催化发夹)的浓度可以是,例如,5nm至1000nm。在某些实施方案中,在引物交换反应中的催化分子浓度是5-10、5-20、5-30、5-40、5-50、5-60、5-70、5-80、5-90、5-100、5-125、5-150、5-200、10-50、10-75、10-100、10-150、10-200、25-75、25-100、25-125或25-200nm。在某些实施方案中,在引物交换反应中的催化分子浓度是10-200、10-300、10-400、10-500、10-600、10-70、10-800、10-900或10-100nm。在某些实施方案中,在引物交换反应中的催化分子浓度是10、15、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、100、105、110、115、120、125、130、135、140、145、150、155、160、165、170、175、180、185、190、195或200nm。在某些实施方案中,在引物交换反应中的催化分子浓度是10、20、30、40、50、60、70、80、90或100nm。在引物交换反应中的催化分子浓度可以小于5nm或大于1000nm。
在引物交换反应中的引物与催化分子的比率可以是2:1至100:1。在某些实施方案中,所述引物与催化分子的比率是2:1、3:1、4:1、5:1、6:1、:1、8:1、9:1、10:1、11:1、12:1、13:1、14:1、15:1、16:1、17:1、18:1、19:1或20:1。在某些实施方案中,所述引物与催化分子的比率是30:1、40:1、50:1、60:1、70:1、80:1或90:1。
在引物交换反应中的不同催化分子的数目是非限制性的。引物交换反应可以包含1-1010种不同的催化分子(例如,各自具有特定支点结构域序列)。在某些实施方案中,引物交换反应包含1-10、1-102、1-103、1-104、1-105、1-106、1-107、1-108、1-109、1-1010种或更多种不同的催化分子。在某些实施方案中,引物交换反应包含1-5、1-10、1-15、1-20、1-25、1-30、1-35、1-40、1-45、1-50、1-55、1-60、1-65、1-70、1-75、1-80、1-85、1-90、1-95、1-100、5-10、5-15、5-20、5-25、5-30、5-35、5-40、5-45、5-50、5-55、5-60、5-65、5-70、5-75、5-80、5-85、5-90、5-95、5-100、10-15、10-20、10-25、10-30、10-35、10-40、10-45、10-50、10-55、10-60、10-65、10-70、10-75、10-80、10-85、10-90、10-95或10-100种不同的催化分子。在某些实施方案中,引物交换反应包含1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、28、19、20、21、22、23、24或25种不同的催化分子。例如,如果它们的支点结构域彼此不同,则催化分子是彼此不同的。
通过例如改变温度、时间、缓冲液/盐条件和脱氧核糖核苷酸三磷酸(dntp)浓度,可以控制引物交换反应的动力学。象大多数酶一样,聚合酶对许多缓冲条件是敏感的,包括离子强度、ph和存在的金属离子的类型(例如,钠离子相对于镁离子)。因而,进行引物交换反应的温度可以例如在4℃至65℃变化(例如,4℃、25℃、37℃、42℃或65℃)。在某些实施方案中,进行引物交换反应的温度是4-25℃、4-30℃、4-35℃、4-40℃、4-45℃、4-50℃、4-55℃、4-60℃、10-25℃、10-30℃、10-35℃、10-40℃、10-45℃、10-50℃、10-55℃、10-60℃、25-30℃、25-35℃、25-40℃、25-45℃、25-50℃、25-55℃、25-60℃、25-65℃、35-40℃、35-45℃、35-50℃、35-55℃、35-60℃或35-65℃。在某些实施方案中,在室温进行引物交换反应,而在其它实施方案中,在37℃进行引物交换反应。
可以进行引物交换反应(温育)30分钟(min)至24小时(h)。在某些实施方案中,进行引物交换反应10min、35min、40min、45min、50min、55min、60min、1小时、2小时、3小时、4小时、5小时、6小时、7小时、8小时、9小时、10小时、11小时、12小时、18小时或24小时。
脱氧核糖核苷酸(dntp)是驱动引物交换反应的“燃料”。因而,在某些实施方案中,引物交换反应的动力学主要取决于反应中dntp的浓度。在引物交换反应中dntp的浓度可以是,例如,2-1000μm。在某些实施方案中,在引物交换反应中dntp浓度是2-10μm、2-15μm、2-20μm、2-25μm、2-30μm、2-35μm、2-40μm、2-45μm、2-50μm、2-55μm、2-60μm、2-65μm、2-70μm、2-75μm、2-80μm、2-85μm、2-90μm、2-95μm、2-100μm、2-110μm、2-120μm、2-130μm、2-140μm、2-150μm、2-160μm、2-170μm、2-180μm、2-190μm、2-200μm、2-250μm、2-300μm、2-350μm、2-400μm、2-450μm、2-500μm、2-600μm、2-700μm、2-800μm、2-900μm或2-1000μm。例如,在引物交换反应中dntp浓度可以是2μm、5μm、10μm、15μm、20μm、25μm、30μm、35μm、40μm、45μm、50μm、55μm、60μm、65μm、70μm、75μm、80μm、85μm、90μm、95μm、100μm、105μm、110μm、115μm、120μm、125μm、130μm、135μm、140μm、145μm、150μm、155μm、160μm、165μm、170μm、175μm、180μm、185μm、190μm、195μm或200μm。在某些实施方案中,在引物交换反应中dntp浓度是10-20μm、10-30μm、10-40μm、10-50μm、10-60μm、10-70μm、10-80μm、10-90μm或10-100μm。
在某些实施方案中,使用dntp变体。例如,引物交换反应可以使用热启动/cleanampdntp、硫代磷酸酯dntp或荧光dntp。可以使用其它dntp变体。因为一些经修饰的dntp不如正常的(未修饰的)dna-dna结合有利,所以发夹回复置换过程(hairpinbackdisplacementprocess)可能因它们的使用而增加。类似地,在某些实施方案中可以使用包含不同类型的核酸(例如,lna、rna或散布的经修饰的碱基诸如甲基dc或超tidt修饰)的发夹如下增加per的速度:通过形成就催化分子而言比合成的引物更强的键。
支点开关锁
核糖调节剂开关概述。支点开关锁是核糖调节剂开关,其通常是可以用于控制可读框的翻译并因而控制蛋白的生产的rna分子。所述开关通过互补碱基配对形成包含茎结构域和环结构域的发夹。所述发夹阻断核糖体对rna转录物的接近,由此阻止翻译。所述发夹通常将核糖体结合位点(rbs)隔离在它的茎结构域或它的环结构域中。
在触发核酸(例如,触发钥匙)与开关结合后发生的发夹结构域中的改变允许核糖体接近在起始密码子上游的转录物区域,由此将开关从它的被抑制状态释放并促进从转录物的蛋白翻译。所述开关通常是经工程改造的rna分子和/或由经工程改造的dna序列产生。所述触发物也可以是经工程改造的核酸。
关于核糖调节剂开关的其它一般教导参见公开的国际pct申请号wo2004/046321、wo2014/074648和wo2016/011089,它们中的每一篇的内容通过引用并入本文。
支点开关锁。在支点开关锁系统中,开关和触发物之间的相互作用通过位于发夹的5'末端的未配对的(单链的)核酸结构域介导。该未配对的核酸结构域(其被称作支点结构域)给触发物提供了足够结合亲和力以使它展开发夹的茎结构域。因而,本文还提供了包含rna的支点核糖调节剂开关(支点开关锁),所述rna包含(a)未配对的支点结构域,其包含与所述催化分子之一的3'子结构域互补的序列,(b)发夹,其包含(i)配对的茎结构域,其包含位于所述未配对的支点结构域的3'的起始密码子和与所述催化分子之一的3'支点结构域互补的序列,和(ii)包含核糖体结合位点的环结构域,和(c)编码报道蛋白的编码结构域。通常,所述编码结构域编码报道蛋白(例如,绿色荧光蛋白,gfp)。
触发钥匙和支点结构域之间的互补性程度可以变化。在某些实施方案中,它是至少60%、至少70%、至少80%、至少90%、至少95%或100%。为了最佳的核糖调节剂动力学,触发钥匙应当具有最小的二级结构和与支点开关锁的支点结构域的完全互补性(即,100%)。本文中使用的二级结构表示非线性结构,包括例如发夹结构、茎环结构等。因此,在某些实施方案中,所述触发物包括这样的序列(例如,由其组成):在它的使用条件下,所述序列具有极小形成二级结构的可能性至没有形成二级结构的可能性。本领域普通技术人员能够手工地或通过使用本领域中可得到的计算机程序确定这样的序列。
在某些实施方案中,所述支点开关锁包含超过一个编码结构域,且因而编码超过一种蛋白。所述编码结构域可以以直链方式排列在发夹结构域的下游(3')。
在某些实施方案中,未配对的支点结构域可以与催化核酸分子(例如,催化发夹分子)的引物钥匙或支点或茎结构域互补。
支点开关锁的支点结构域的长度可以变化。在某些实施方案中,支点结构域可以具有2-35、2-30、2-25、2-20、2-15、2-10、5-35、5-30、5-25、5-20、5-15、5-10、10-40、10-35、10-30、10-25、10-20、10-15、15-40、15-35、15-30、15-25、15-20、20-40、20-35、20-30、20-25、25-40、25-35、25-30、30-40、30-35或35-40个核苷酸的长度。在某些实施方案中,支点结构域具有5、10、15、20、25、30、35或40个核苷酸的长度。在某些实施方案中,支点结构域具有10、11、12、13、14、15、16、17、18、19、20、21、22、23、24或25个核苷酸的长度。在某些实施方案中,支点结构域长于40个核苷酸,或短于5个核苷酸。在某些实施方案中,未配对的支点结构域具有15、16、17、18个核苷酸或更长的长度。
在某些实施方案中,完全地或部分地配对的茎结构域是部分地配对的茎结构域,其中起始密码子位于将第一和第二配对的结构域隔开的未配对的凸起中。在某些实施方案中,所述第一配对的结构域邻近所述支点结构域。在某些实施方案中,所述环结构域邻近所述第二配对的结构域。
完全地或部分地配对的茎结构域可以包含一个或多个另外的未配对的凸起,且这样的凸起可以具有1、2或3个核苷酸的长度,尽管它们没有这样限制。
在某些实施方案中,所述起始密码子完全地或部分地存在于茎结构域中的未配对的凸起中。
在某些实施方案中,在起始密码子下游的序列不编码终止密码子。
在某些实施方案中,所述第一配对的结构域可以具有11-100、11-50、11-40、11-30或11-20个核苷酸碱基对的长度。在某些实施方案中,所述第一配对的结构域可以具有11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30个碱基对或更长的长度。在某些实施方案中,所述第一配对的结构域可以具有长于100个碱基对的长度,包括例如多达120、140、160、180或200个或更多个碱基对。
支点开关锁的配对的茎结构域的长度可以变化。在某些实施方案中,配对的结构域具有5-40个核苷酸的长度。例如,配对的结构域可以具有5-35、5-30、5-25、5-20、5-15、5-10、10-40、10-35、10-30、10-25、10-20、10-15、15-40、15-35、15-30、15-25、15-20、20-40、20-35、20-30、20-25、25-40、25-35、25-30、30-40、30-35或35-40个核苷酸的长度。在某些实施方案中,配对的结构域具有5、10、15、20、25、30、35或40个核苷酸的长度。在某些实施方案中,配对的结构域具有10、11、12、13、14、15、16、17、18、19、20、21、22、23、24或25个核苷酸的长度。在某些实施方案中,配对的结构域长于40个核苷酸,或短于5个核苷酸。
配对的茎结构域可以具有第一配对的结构域(子结构域)和第二配对的结构域(子结构域)。在某些实施方案中,所述第一配对的结构域具有11或12个碱基对的长度。在某些实施方案中,所述第一配对的结构域比所述第二配对的结构域长。在某些实施方案中,所述第二配对的结构域具有小于11个碱基对的长度。在某些实施方案中,所述第二配对的结构域具有10、9、8、7或6个碱基对的长度。在某些实施方案中,所述第二配对的结构域具有5或6个碱基对的长度。在某些实施方案中,所述第一配对的结构域具有11个碱基对的长度,且所述第二配对的结构域具有5个碱基对的长度。在某些实施方案中,所述第一配对的结构域具有12个碱基对的长度,且所述第二配对的结构域具有6个碱基对的长度。在某些实施方案中,所述环结构域具有12-14个核苷酸的长度,尽管它没有这样限制。
在某些实施方案中,所述支点开关锁进一步包含位于发夹的下游(3')且在茎结构域和/或第一配对的结构域和编码结构域之间的间隔物结构域。在某些实施方案中,所述间隔物结构域编码低分子量氨基酸。在某些实施方案中,所述间隔物结构域具有约9-33个核苷酸或约21个核苷酸的长度。
支点开关锁通常不会将rbs隔离在它们的茎结构域内。相反,将rbs被限制至通过抑制茎结构域而形成的环结构域。这允许在起始密码子前面(上游或5')和后面(下游或3')紧挨着的区域被隔离在茎结构域内,从而破坏翻译起始。可以在大程度上改变支点、茎和环结构域的各自长度,而不影响支点开关锁的性能,如将在下面详述的。另外,茎结构域可以保留它的抑制效率,即使它含有许多凸起或错配的碱基,这使不含有起始密码子aug序列的触发物能够触发支点开关锁。原则上,凸起的公差(tolerance)允许任意核酸序列(包括内源性rna)充当触发物进入支点开关锁中,尽管其它标准诸如高二级结构可以影响调节剂的应答。
支点开关锁的一个示例性的、非限制性的类别具有约12个核苷酸(nt)长的支点结构域和约11个核苷酸长的环结构域,且其含有任选地在它的3'末端的rbs序列agaggaga。与该环结构域紧邻的是包含6碱基对双链体间隔物区域和侧接起始密码子(aug)的9碱基对双链体区域的茎结构域。将在起始密码子下游(3')的9个核苷酸程序化以确保它们不会编码任何终止密码子,因为这会导致过早的翻译终止。如基于本公开内容会理解的,触发物负责展开茎结构域。另外,与起始密码子三联体相对的3-核苷酸区域是完全不配对的,从而产生具有3-核苷酸长凸起的茎结构域(该设计排除了触发物在程序化为与该凸起杂交的位置处具有aug序列)。为了减小9-核苷酸双链体区域编码影响目标蛋白的折叠的氨基酸的可能性,将含有许多低分子量残基的常见21-核苷酸(7-氨基酸)间隔物结构域插入在茎结构域和编码结构域(例如,编码目标蛋白的结构域)之间。因而,在某些情况下,支点开关锁将11个残基添加至编码的蛋白的n-端,所述蛋白包括茎的12-核苷酸翻译部分和紧随其后的常见21-核苷酸接头区域。
应当理解,该实施方案是非限制性的,并且本公开内容包括具有不同长度和功能的其它支点开关锁。因而,支点结构域、茎结构域、环结构域和接头结构域、以及在所述茎结构域内的双链体区域的长度可以在长度上不同于该实施方案。
应当理解,可以用对支点开关设计的一些修饰来避免前述施加于触发钥匙和效应物蛋白的条件。对触发钥匙的序列约束是为茎结构域和触发物-支点结构域复合物指定的碱基配对条件的副产物。但是,这些特定二级结构不是开关运转所严格需要的。高效开关可以具有在aug位置处的小于3-核苷酸凸起或在茎结构域根部的另外碱基对。向/从开关添加/减少碱基对的设计修饰仍然将允许支点开关调节蛋白翻译,同时提供足够的设计灵活性以消除对触发物序列的终止密码子-和aug-凸起相关的约束。
在某些情况下,支点开关包含一致的原核rbs。但是,多种替代性的天然存在的或经工程改造的序列中的任一种可以用作rbs。已经确定了大量细菌rbs的序列,且已知这些序列的重要特征。高水平翻译的优选rbs序列含有在相对于aug的位置-6至-11处的富含g的区域,且通常含有在位置-3处的a。示例性的rbs序列包括,但不限于agaggaga(或该序列的子序列,例如,至少6个核苷酸长度的子序列,诸如aggagg)。更短的序列也是可接受的,例如,agga、agggag、gaggag等。已经创建了众多合成的rbs,并已经试验了它们的翻译起始活性。使用任意合适的方法可以试验任何候选序列作为rbs起作用的活性。例如,如在公开的pct申请wo2004/046321的实施例1中所述,或如在该公开的pct申请的参考文献53中所述,可以测量表达,例如,通过测量由mrna编码的报道蛋白的活性,所述mrna含有适当地定位在aug上游的候选rbs。优选地,rbs序列支持的翻译水平为共有rbs支持的翻译水平的至少10%(例如,如通过报道蛋白的活性测量的)。例如,如果将候选rbs插入在对照质粒中替代一致rbs,测得的荧光将是使用共有rbs测得的荧光的至少10%。在某些实施方案中,可以使用这样的rbs:其支持的翻译水平为共有rbs支持的水平的至少1%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%或更多。在某些实施方案中,可以使用支持比共有rbs更高的翻译水平的rbs。
此外,还可以修饰支点开关锁以将输出蛋白的编码序列直接掺入开关茎结构域中。这类开关与对n-端修饰敏感的任何蛋白相容。支点介导的相互作用的特异性、凸起在茎结构域中的再分布和同义密码子的应用会为这些支点开关提供足够的序列空间以高动态范围和正交性运转。
在某些情况下,将具有至少5或6个核苷酸的长度的支点结构域用于最初的触发物结合。因此,在某些实施方案中,支点结构域是5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20个或更多个核苷酸。此外,还发现,为了允许编码的蛋白的翻译,触发钥匙仅需要展开茎结构域的三分之二(或茎结构域的第一双链区域的三分之二)。基于这些发现,为了适当抑制,茎结构域可以小至12个核苷酸碱基对。但是,茎结构域可以长于12个核苷酸碱基对,包括13、14、15、16、17、18、19、20个或更多个碱基对。此外,将环长度扩大至12-核苷酸和用具有规范shine-dalgarno序列的稍微更强的形式替换rbs,不减小开关的抑制程度。因此,环结构域的长度可以是11、12、13、14、15、16、17、18、19、20个或更多个核苷酸。
支点开关锁可以具有额外的特征。在某些情况下,茎结构域的顶部三个碱基可以是a-u碱基对。在某些情况下,茎结构域的底部三个碱基对可以包含两个强g-c碱基对和一个a-u碱基对。在某些情况下,开关支点的长度可以在约12至约15个核苷酸的范围内。该后一个特征在某些情况下可能强化触发物和它的相应支点结构域之间的最初结合。在某些情况下,环结构域的大小可以在约11至约15个核苷酸的范围内,以增强开关激活后的效应物蛋白翻译。在某些情况下,环大小是15个核苷酸。在其它情况下,可以使用展开茎结构域的18个碱基中的前15个的相应触发物。在某些情况下,可以同时使用这些特征中的一个或多个,包括全部。
在以下情况下,触发钥匙(其可以是rna)和支点开关锁是相应的:它们能够彼此结合并实现对支点开关锁的结构和功能改变,但是不能结合具有相同结构和功能效果的其它触发物和支点开关锁。在某些实施方案中,所述触发钥匙可以包含一个或多个结构域,其中至少一个结构域与支点开关锁的支点结构域具有100%互补性或能够在严谨条件下与支点开关锁的支点结构域杂交。因而,触发物和支点开关锁是对彼此特异性的,这意味着它们特异性地和选择性地彼此结合,但是不会结合其它不相应核酸。
应当理解,在本文提供的这个和其它实施方案的背景下,术语开关、支点开关、支点核糖调节剂、支点核糖调节剂开关、核糖调节剂开关、支点阻遏物、crrna、crrna核糖调节剂、crrna阻遏物等互换使用。类似地,在本文提供的这个和其它实施方案的背景下,术语输入和触发物/触发钥匙等表示这样的核酸:其完全地或部分地结合支点开关锁(通常在它的支点结构域),和/或其结合其它输入(例如,引物钥匙)或触发钥匙由此形成核酸复合物,所述核酸复合物结合支点开关锁的支点结构域并实现锁结构和/或功能的改变。后一类输入包括为and门(andgate)做贡献的那些。因而,在某些实施方案中,and门包括两个或更多个触发钥匙,它们必须彼此杂交以形成复合物,所述复合物本身能够结合支点核糖调节剂开关和造成核糖调节剂开关的结构和功能改变。一些、但并非所有的这样的and门触发物可以包含与核糖调节剂开关的支点结构域互补且能够与其杂交的核苷酸序列。
在某些实施方案中,支点开关锁与触发钥匙的比率小于1、小于0.5或小于0.1。
在某些实施方案中,所述支点开关锁被包含在第一核酸中或在第一核酸中编码。在某些实施方案中,所述第一核酸是第一质粒。在某些实施方案中,所述第一质粒包含中间拷贝复制起点。所述质粒可以是dna质粒或rna质粒。应该理解,在dna质粒的转录后,得到的rna种类将包括呈rna形式的支点开关锁。
进一步理解,任何给定的核酸构建体(不论在性质上是dna还是rna,诸如但不限于质粒或表达载体)可以包含或编码一个或多个支点开关锁。
编码的蛋白。所述开关可以编码一种或超过一种可检测的分子(例如,发出可检测信号诸如荧光或化学发光信号的分子),诸如报道蛋白(例如,荧光蛋白)。本文中使用的编码的蛋白包括蛋白和肽,尽管为了简洁将在本文中提及蛋白。应当理解,本公开内容同样包括蛋白和肽,除非另外说明。容易观察到报道物在细胞或生物体中的存在。
报道蛋白表示可以用于测量基因表达且通常产生可测量的信号(诸如荧光、发光或颜色)的蛋白。例如,当用特定波长的光激发时,荧光蛋白(例如,gfp)会造成溶液(或其它介质)发荧光,萤光素酶会造成产生光的催化反应,且酶诸如β-半乳糖苷酶会将底物转化成有色产物。
在某些实施方案中,报道蛋白是荧光蛋白。使用显微镜、平板读数器、流式细胞计或被装配成用适当波长的光激发荧光蛋白的其它装置(例如,手持装置)可以容易地量化荧光。可得到几种不同的荧光蛋白。根据本公开内容可以使用的编码荧光蛋白的基因的例子包括,但不限于,在美国专利申请号2012/0003630(参见表59)(通过引用并入本文)中提供的那些蛋白。
在某些实施方案中,报道蛋白是萤光素酶。使用平板读数器或发光计数器可以容易地量化发光。根据本公开内容可以使用的编码萤光素酶的基因的例子包括,但不限于,dmmyd88-接头-rluc、dmmyd88-接头-rluc-接头-pest191和萤火虫萤光素酶(来自北美萤火虫(photinuspyralis))。
逻辑解决方案
在某些实施方案中,使用逻辑运算来解锁支点开关锁。例如,支点开关锁可以在仅有引物钥匙a存在下打开,但是不会在有引物钥匙b存在下打开。在本文中包括其它更复杂的逻辑运算。例如,支点开关锁可以仅在有两种引物钥匙存在下打开,但是不会在有第三种引物钥匙存在下打开(参见,例如,图6a-6d)。
布尔逻辑函数是基于可以用于组装将实现任何数字组件的函数的组合的逻辑门。存在至少16种逻辑门:not、and、or、not、nor、nand、xor、xnor、aimplyb、bimplya、animplyb、bnimplya、a、b、false或true。
在药物鉴定操作中的应用
在某些应用(诸如处方药物鉴定和跟踪)中,可能优选的是,由于管理和安全性担忧,不修饰dna钥匙。为了保护免于钥匙读出和伪造,通过经常改变钥匙序列,可以使用未修饰的dna钥匙。
在图20a所示的一个实施方案中,每个产品含有独特的钥匙链。所述产品还包括测试条,其将产品钥匙翻译成可见代码。将产品施加于测试条会产生长的且独特的条形码图案。针对真实性在生产商的网站上检查所述图案,例如,用电话照相机、手工输入或其它方法。每个测试条结果仅可以检查一次,因为重复的试验或错误的钥匙将导致指示可能伪造的阴性结果。也会保护免于将真实产品稀释成几个复制产品。该应用也没有一定要求保护锁链免于读出。
在图20b所示的另一个实施方案中,将给定的钥匙序列用于短时间段,例如,当在液体或固体药物制剂内混合特定的未修饰的dna链时。从单独场合的单独来源(合法的药物生产商)接收的基于纸的试验允许鉴定特定的钥匙。测试条标有失效日期(此日期后不再有效)、以及所述试验相容的序列号或其它批号的范围。在某些实施方案中,所述锁链被保护免于通过共价修饰或其它方式读出。作为附加的保护水平,当测试条到期时,生产商改变掺入的钥匙链,使得从当前药物批次读出钥匙链仅在非常短的时期是有益的。
核酸
应当理解,本公开内容的核酸不存在于自然界中。因而,所述核酸可以被称作“经工程改造的核酸”。“经工程改造的核酸”是不存在于自然界中的核酸(例如,至少2个核苷酸共价地连接在一起,且在某些情况下,含有磷酸二酯键,被称作磷酸二酯“主链”)。经工程改造的核酸包括重组的核酸和合成的核酸。“重组的核酸”是通过连接核酸(例如,分离的核酸、合成的核酸或它们的组合)而构建的分子,且在某些实施方案中,可以在活细胞中复制。“合成的核酸”是扩增的或化学地或通过其它方式合成的分子。合成的核酸包括化学修饰的或以其它方式修饰的核酸,但是可以与天然存在的核酸分子进行碱基配对(也被称作“结合”,例如,暂时地或稳定地)。重组的核酸和合成的核酸也包括从前述任一种的复制产生的那些分子。
尽管经工程改造的核酸作为整体不是天然存在的,但是它可能包括野生型核苷酸序列。在某些实施方案中,经工程改造的核酸包含得自不同生物体(例如,得自不同物种)的核苷酸序列。例如,在某些实施方案中,经工程改造的核酸包括鼠核苷酸序列、细菌核苷酸序列、人核苷酸序列、病毒核苷酸序列或前述序列中的任意两种或更多种的组合。在某些实施方案中,经工程改造的核酸含有一个或多个随机碱基。
在某些实施方案中,本公开内容的经工程改造的核酸可以包含除磷酸二酯主链之外的主链。例如,在某些实施方案中,经工程改造的核酸可以包含磷酰胺、硫代磷酸酯、二硫代磷酸酯、o-甲基亚磷酰胺键、肽核酸或前述键中的任何两种或更多种的组合。如所指定的,经工程改造的核酸可以是单链的(ss)或双链的(ds),或者经工程改造的核酸可以含有单链和双链序列的部分。在某些实施方案中,经工程改造的核酸含有三链序列的部分或其它非沃森-克里克碱基配对,例如g-四联体,g-四链体和i-基序。经工程改造的核酸可以包含dna(例如基因组dna、cdna或基因组dna和cdna的组合)、rna或杂合分子,例如,其中核酸含有脱氧核糖核苷酸和核糖核苷酸(例如,人工的或天然的)的任何组合和两种或更多种碱基(包括尿嘧啶、腺嘌呤、胸腺嘧啶、胞嘧啶、鸟嘌呤、肌苷、黄嘌呤、次黄嘌呤、异胞嘧啶和异鸟嘌呤)的任何组合。
使用标准的分子生物学方法可以生产本公开内容的经工程改造的核酸(参见,例如,green和sambrook,molecularcloning,alaboratorymanual,2012,coldspringharborpress)。在某些实施方案中,使用gibson
读出分子验证系统
为了输出验证反应的结果,可以使用几个读出策略。例如,读出可以是在含有验证系统组分的溶液中的可见颜色变化。该颜色变化可以是由已经被打开的支点开关锁编码的蛋白表达的结果。这可以例如如下实现:激活支点开关,其产生造成颜色变化的蛋白。
作为另一个例子,读出可以是使用透照仪(诸如蓝光闪光灯或其它特定波长)检测到的荧光。例如,引物交换反应可以包括募集硫代黄素t(tht)的掺入的核苷酸序列,所述硫代黄素t当在蓝光下照射时发荧光(参见,例如,图4e)。还可以使用荧光嵌入染料(例如,
物理实现
所述分子验证系统可以以几种形式中的一种实现,所述形式包括微滴、油墨、毛细管条或喷雾剂。
微滴。可以将两种溶液(一种含有分子标识符且一种含有分子解码器)混合在一起以开始解码per和/或支点开关反应。以上述方式之一读出结果。
油墨。例如,可以在纸上运行支点开关锁,且可以将经干燥的dna稳定地储存在纸上长时间段,特别如果将它冻干的话。因此,这使得油墨(特别是经干燥的油墨)成为吸引人的分子鉴别和/或解码系统的储存模式。例如,一个人可以携带含有引物钥匙的钢笔。备选地,可以将油墨印或写和干燥在一张纸(或手或其它表面)上,所述油墨可以在需要验证时再水合并与适当的分子解码器(例如,催化分子和支点开关锁)混合。类似地,所述分子解码器可以是钢笔,其在经干燥的分子鉴别记录上书写以通过上述的分析和读出策略来验证身份。
毛细管条。可以使用毛细管作用将分子鉴别溶液上拉至经干燥的沉积的分子解码溶液用于验证,或反之亦然。条具有以下附加益处:能够容易地包括阳性和阴性对照泳道以进一步验证没有尝试欺骗,同时仍然要求所述条放在适当的水溶液中仅一次。
喷雾剂。可以在基底(诸如纸)上运行支点开关锁。在某些实施方案中,可以将支点锁沉积在确定区域或图形设计(其可以由疏水屏障诸如蜡限定)内。通过喷雾,将锁暴露于“钥匙”溶液,所述锁包含被猝灭剂阻止发荧光的荧光分子。当将匹配钥匙喷在锁上时,猝灭剂被从荧光分子上置换下来。然后可以针对荧光来测定图形设计。
额外的安全措施
可以使用分子验证系统的几个另外方面来给所述方案添加额外的安全层:
短引物钥匙。短核酸序列(例如,小于18个核苷酸)难以扩增和测序,这使得分子鉴别溶液特别难以复制(欺骗)。
经修饰的引物钥匙.还可以包括在引物钥匙上的修饰以抑制通过诸如聚合酶链式反应(pcr)的方法进行复制的可能。
引物钥匙的特定序列。可以将引物钥匙设计成具有故意地难以测序和复制的序列,诸如一系列重复的结构域。
相对和确切浓度。通过设计分子验证系统来区分具有正确的相对浓度和/或确切浓度的溶液中种类的多种溶液,实现额外的安全。即使某个人设法对存在于溶液中的种类测序,也非常难以鉴别它们的相对和/或确切浓度以有效地破坏代码。
模糊。通过包括额外的种类,如上所述,正确地确定引物钥匙和解码程序的鉴别特征变得明显更难。
温度或光敏感的种类。通过包括例如在升高的温度或在有光存在下被降解的物质,可以进一步保护免于某个人得到并复制溶液的可能。
时间敏感的种类。可以如下进一步保护反应混合物:将它们设计成在再水合以后在某个时间量以后降解,这会进一步限制某个人复制它们的可能。以及时方式读出这些溶液,且因为即将来临的降解,在沉积后仅可以读出一次。
对照。可以实现阳性和阴性对照机制来确定某个人是否在试图欺骗分子鉴别反应混合物。例如,反应混合物可以包括使得所有支点开关具有组成活性的化学剂。通过包括支点开关的阴性对照(其不会被激活,例如,在有相应钥匙存在下)并用其试验,消除了假权限的可能性。可以使用阳性对照来验证样品被适当地混合且材料没有过期。阳性对照特别容易添加至毛细管条(作为脱水的泳道)或钢笔,例如,在一个人可以用反应以及对照钢笔试验以验证钥匙的情况下。
经由核酸纳米结构自装配的鉴定
在某些实施方案中,通过使用例如原子力显微术(afm)或电子显微术(em)测定核酸纳米结构的存在,可以鉴定产品。在某些实施方案中,所述核酸纳米结构存在于(例如,附着于)产品。在其它实施方案中,所述核酸纳米结构存在于(例如,附着于)产品包装上。在某些实施方案中,所述产品是液体产品,诸如香料、油墨或其它基于油的或基于水的产品。在某些实施方案中,所述产品是固体。所述核酸纳米结构可以从dna、rna、或dna和rna的组合自装配。在某些实施方案中,经由已知的dnaorigami方法组装所述核酸纳米结构。在某些实施方案中,经由基于单链瓦片的(基于sst的)方法(参见,例如,weib.等人.nature485:626,2012和2014年5月15日公开的国际公开号wo2014/074597,2013年2月14日公开的国际公开号wo2013/022694,和2014年1月30日公开的国际公开号wo2014/018675,各自通过引用并入本文)组装所述核酸纳米结构。在某些实施方案中,所述核酸纳米结构包含可识别的形状,例如,字母,例如,如在us2015/0218204中所述。在某些实施方案中,经由核酸的一条单链的折叠,组装所述核酸纳米结构,例如,如在wo2016/144755(通过引用并入本文)中所述。在某些实施方案中,组装所述核酸纳米结构,然后附着于产品和/或包装。在其它实施方案中,在产品和/或包装的组装过程中组装所述核酸纳米结构。因而,在某些实施方案中,方法包括观察特定核酸纳米结构(例如,具有确定的二维或三维形状)在产品或包装的样品中的存在,和基于所述核酸纳米结构在样品中的存在或不存在而鉴定所述产品的身份。
另外的实施方案
1.一种试剂盒,其包括:
(a)含有一组催化分子的组合物,每个催化分子包含(i)未配对的3'支点结构域,(ii)通过所述分子的3'子结构域和所述分子的5'子结构域之间的分子内核苷酸碱基配对形成的配对的茎结构域,和任选的(iii)环结构域;
(b)包含引物钥匙的组合物,所述引物钥匙与所述催化分子之一的3'支点结构域互补且与其结合;
(c)包含rna支点开关锁的组合物,所述rna支点开关锁包含
(i)未配对的支点结构域,其包含与所述催化分子之一的3'子结构域互补的序列,
(ii)发夹,其包含
配对的茎结构域,其包含位于所述未配对的支点结构域的3'的起始密码子和与所述催化分子之一的3'支点结构域互补的序列,和
环结构域,其包含核糖体结合位点,和
(iii)编码报道蛋白的编码结构域。
2.段落1的试剂盒,其中所述钥匙引物具有小于18个核苷酸的长度。
3.段落2的试剂盒,其中所述钥匙引物具有4-15个核苷酸的长度。
4.段落1-3中的任一段的试剂盒,其中(c)的组合物进一步包含至少一种不结合所述催化发夹分子的分子。
5.段落1-4中的任一段的试剂盒,其中所述催化分子组包含3-20种催化分子。
6.段落1-5中的任一段的试剂盒,其中所述组的催化分子具有25-100个核苷酸的长度。
7.段落1-6中的任一段的试剂盒,其中所述组的催化分子的支点结构域具有4-17个核苷酸的长度。
8.段落1-7中的任一段的试剂盒,其中所述组的催化分子的茎结构域具有5-40个核苷酸的长度。
9.段落1-8中的任一段的试剂盒,其中所述组的催化分子的环结构域具有3-20个核苷酸的长度。
10.段落1-9中的任一段的试剂盒,其中所述rna支点开关锁的支点结构域具有5-20个核苷酸的长度。
11.段落1-10中的任一段的试剂盒,其中所述rna支点开关锁的茎结构域具有2-100个核苷酸的长度。
12.段落1-11中的任一段的试剂盒,其中所述rna支点开关锁的核糖体结合位点包含序列agaggaga。
13.段落1-12中的任一段的试剂盒,其中所述报道蛋白是荧光蛋白。
14.一种方法,其包括:
在包含具有链置换活性的聚合酶和脱氧核糖核苷酸三磷酸(dntp)的第一反应缓冲液中组合
(a)一组催化分子,每个催化发夹分子包含(i)未配对的3'支点结构域,(ii)通过所述分子的3'子结构域和所述分子的5'子结构域之间的分子内核苷酸碱基配对形成的配对茎结构域,和(iii)环结构域;和
(b)引物钥匙,其与所述催化分子之一的3'支点结构域互补且与其结合;
在导致核酸聚合、链置换和退火的条件下温育所述第一反应混合物足以产生单链核酸触发物的时间;
在第二反应缓冲液中组合
(c)所述单链核酸触发物,和
(d)rna支点开关锁,其包含
(i)未配对的支点结构域,其包含与所述催化分子之一的3'子结构域互补的序列,
(ii)发夹,其包含
配对的茎结构域,其包含位于所述未配对的支点结构域的3'的起始密码子和与所述催化分子之一的3'支点结构域互补的序列,和
环结构域,其包含核糖体结合位点,和
(iii)编码报道蛋白的编码结构域,由此形成第二反应混合物;和
在导致核酸杂交、rna转录和蛋白翻译的条件下温育所述第二反应混合物足以表达所述报道蛋白的时间。
15.一种试剂盒,其包括:
(a)包含核酸链的组合物,所述核酸链连接至荧光分子且具有5'结构域和3'结构域;和
(b)包含核酸链的组合物,所述核酸链连接至猝灭分子且具有(i)与(a)的核酸链的3'结构域互补的5'结构域和(ii)与(a)的核酸链的5'结构域互补的3'结构域。
16.段落15的试剂盒,其中所述(a)和/或(b)的核酸链包含至少一个停止子。
17.段落16的试剂盒,其中所述至少一个停止子选自间隔物、合成的核苷酸和修饰的核苷酸。
18.一种方法,其包括:
在反应缓冲液中组合段落15-17中的任一段的(a)的组合物和(b)的组合物;和
在导致核酸杂交的条件下温育所述反应缓冲液。
19.一种试剂盒,其包括:
(a)含有核酸发夹分子的组合物,所述核酸发夹分子具有通过所述核酸分子的5'末端结构域和3'末端结构域的配对而形成的环结构域和茎结构域,其中一个末端结构域包括荧光分子且另一个末端包括猝灭分子;和
(b)含有核酸发夹分子的组合物,所述核酸发夹分子具有通过所述核酸发夹分子的5'末端结构域和3'末端结构域的配对而形成的环结构域和茎结构域,其中一个末端结构域包含与(a)的发夹分子的5'末端结构域的序列互补的序列,且另一个末端结构域包含与(a)的发夹分子的3'末端结构域的序列互补的序列,且所述环结构域包含与(a)的环结构域的序列互补的序列。
20.段落19的试剂盒,其中所述(a)和/或(b)的核酸发夹分子包含至少一个停止子。
21.段落20的试剂盒,其中所述至少一个停止子选自间隔物、合成的核苷酸和修饰的核苷酸。
22.一种方法,其包括:
在反应缓冲液中组合段落19-21中的任一段的(a)的组合物和(b)的组合物;和
在导致核酸杂交的条件下温育所述反应缓冲液。
23.一种试剂盒,其包括:
(a)含有催化分子的组合物,所述催化分子具有(i)未配对的支点结构域,(ii)配对的茎结构域,其包含与猝灭分子相对的荧光分子,和任选的(iii)环结构域;和
(b)包含核酸链的组合物,所述核酸链具有两个邻近结构域,一个结构域与(a)的催化分子的支点结构域互补且另一个结构域与(a)的催化分子的茎结构域互补。
24.段落23的试剂盒,其中所述(a)的催化分子和/或(b)的核酸链包含至少一个停止子。
25.段落24的试剂盒,其中所述至少一个停止子选自间隔物、合成的核苷酸和修饰的核苷酸。
26.一种方法,其包括:
在反应缓冲液中组合段落23-25中的任一段的(a)的组合物和(b)的组合物;和
在导致核酸杂交的条件下温育所述反应缓冲液。
27.一种试剂盒,其包括:
(a)含有5'结构域和3'结构域的第一核酸链;
(b)含有5'结构域的第二核酸链,所述5'结构域与所述第一核酸链的3'结构域互补;
(c)第三核酸链,其含有(i)与所述第一核酸链的3'结构域互补的5'结构域和(ii)与所述第一核酸链的5'结构域互补的3'结构域;
(d)含有5'结构域和3'结构域的第四核酸链;
(e)含有5'结构域的第五核酸链,所述5'结构域与所述第四核酸链的3'结构域互补;
(f)第六核酸链,其含有(i)与所述第四核酸链的3'结构域互补的5'结构域和(ii)与所述第四核酸链的5'结构域互补的3'结构域;
(g)猝灭剂分子;和
(h)荧光分子,
其中所述第一核酸链的3'结构域和所述第四核酸链的3'结构域具有彼此不同的核苷酸组成。
28.段落27的试剂盒,其中所述猝灭剂分子中的至少一个连接至所述第一核酸链的3'结构域和/或连接至所述第四核酸链的3'结构域。
29.段落27或28的试剂盒,其中所述荧光分子连接至所述第二核酸链的5'结构域和/或连接至所述第五核酸链的5'结构域。
30.段落27-29中的任一段的试剂盒,其包含第一溶液,所述第一溶液包含与所述第二核酸链结合的第一核酸链。
31.段落27-30中的任一段的试剂盒,其包含第二溶液,所述第二溶液包含与所述第五核酸链结合的第四核酸链。
32.一种溶液,其包含:
(a)第一核酸链,所述第一核酸链含有5'结构域和连接至猝灭剂分子的3'结构域;
(b)含有5'结构域的第二核酸链,所述5'结构域连接至荧光分子且结合所述第一核酸链的3'结构域;
(c)第三核酸链,其含有(i)与所述第一核酸链的3'结构域互补的5'结构域和(ii)与所述第一核酸链的5'结构域互补的3'结构域;
(d)第四核酸链,所述第四核酸链含有5'结构域和连接至猝灭剂分子的3'结构域;
(e)含有5'结构域的第五核酸链,所述5'结构域连接至荧光分子且结合所述第四核酸链的3'结构域;和
(f)第六核酸链,其含有(i)与所述第四核酸链的3'结构域互补的5'结构域和(ii)与所述第四核酸链的5'结构域互补的3'结构域;
其中所述第一核酸链的3'结构域和所述第四核酸链的3'结构域具有彼此不同的核苷酸组成。
33.一种基底,其包括:
(a)至少一个离散区域,该区域包含(i)含有5'结构域和3'结构域的第一核酸链,所述3'结构域连接至猝灭剂分子,和(ii)含有5'结构域的第二核酸链,所述5'结构域连接至荧光分子且结合所述第一核酸链的3'结构域;和
(b)至少一个离散区域,该区域包含(i)第三核酸链,其含有5'结构域和连接至猝灭剂分子的3'结构域,和(ii)含有5'结构域的第四核酸链,所述5'结构域连接至荧光分子且结合所述第三核酸链的3'结构域,
其中所述第一核酸链的3'结构域和所述第三核酸链的3'结构域具有彼此不同的核苷酸组成。
34.一种方法,其包括:
用图形设计给基底加图案;
在所述图形设计上以预定图案沉积以下至少一种:
(a)含有5'结构域和3'结构域的第一核酸链,所述3'结构域连接至猝灭剂分子;和含有5'结构域的第二核酸链,所述5'结构域连接至荧光分子且结合所述第一核酸链的3'结构域;和
(b)第三核酸链,其含有5'结构域和连接至猝灭剂分子的3'结构域;和含有5'结构域的第四核酸链,所述5'结构域连接至荧光分子且结合所述第三核酸链的3'结构域。
35.段落34的方法,所述方法进一步包括在所述图形设计上沉积以下至少一种:
(c)第五核酸链,其含有(i)与所述第一核酸链的3'结构域互补的5'结构域和(ii)与所述第一核酸链的5'结构域互补的3'结构域;和
(d)第六核酸链,其含有(i)与所述第三核酸链的3'结构域互补的5'结构域和(ii)与所述第三核酸链的5'结构域互补的3'结构域。
36.段落35的方法,所述方法进一步包括测定所述图形设计的荧光。
37.段落36的方法,其中所述测定包括将所述图形设计暴露于透照仪,任选地透过滤波器单元。
38.段落34-37中的任一段的方法,其中所述基底是纸。
39.一种产品鉴定系统,其包括:
(a)第一核酸,其包含(i)5'结构域和(ii)3'结构域;
(b)第二核酸,其包含(i)与所述第一核酸的3'结构域互补的5'结构域和(ii)3'结构域;
(c)第三核酸,其包含(i)与所述第二核酸的3'结构域互补的5'结构域和(ii)3'结构域;和
(d)第四核酸,其包含(i)与所述第三核酸的3'结构域互补的5'结构域和(ii)3'结构域。
40.一种产品鉴定系统,其包含:
(a)第一核酸,其包含第一结构域、第二结构域、环结构域和与所述第二结构域互补且结合的第四结构域;和
(b)第二核酸,其包含第一结构域、第二结构域、环结构域和与所述第二核酸的第二结构域互补且结合的第四结构域,其中所述第一核酸的环结构域与所述第二核酸的环结构域互补。
41.一种产品鉴定系统,其包含:
(a)包含第一结构域和第二结构域的第一环形核酸;和
(b)包含第一结构域和第二结构域的第二环形核酸,其中所述第一环形核酸的第一结构域与所述第二环形核酸的第一结构域互补。
42.一种产品鉴定系统,其包含:
(a)包含5'结构域和3'结构域的第一核酸;
(b)第二核酸,其包含(i)与所述第一核酸的3'结构域互补且连接至猝灭剂分子的5'结构域,和(ii)与所述第一核酸的5'结构域互补的3'结构域;和
(c)第三核酸,其包含与所述第二核酸的5'结构域互补且连接至荧光分子的3'结构域。
43.一种产品鉴定系统,其包含:
(a)包含5'结构域和3'结构域的第一核酸;
(b)第二核酸,其包含(i)5'结构域和(ii)与所述第一核酸的5'结构域互补的3'结构域;
(c)第三核酸,其包含(i)与所述第一核酸的3'结构域互补且连接至猝灭剂分子的5'结构域和(ii)与所述第二核酸的5'结构域互补的3'结构域;和
(d)第四核酸,其包含与所述第三核酸的5'结构域互补且连接至荧光分子的3'结构域。
44.一种产品鉴定系统,其包含:
(a)包含5'结构域和3'结构域的第一核酸;
(b)第二核酸,其包含(i)5'结构域和(ii)与所述第一核酸的5'结构域互补的3'结构域;
(c)第三核酸,其包含(i)5'结构域和(ii)与所述第二核酸的5'结构域互补的3'结构域;
(d)第四核酸,其包含(i)与所述第一核酸的3'结构域互补且连接至猝灭剂分子的5'结构域和(ii)与所述第三核酸的5'结构域互补的3'结构域;和
(e)第五核酸,其包含与所述第四核酸的5'结构域互补且连接至荧光分子的3'结构域。
实施例
实施例1
图3a显示了为了引物序列的自动逐步生长而组合几个per发夹和其它试剂的示意图。图3b显示了五个延伸步骤的反应简图,用发夹a、b、c、d和e图示。图2c显示的变性凝胶证实了差别延伸,其中存在发夹种类的不同子集。在37℃温育反应物4小时,引物浓度为100nm,发夹为10nm,datp、dttp和dctp各为10μm。
实施例2
图5a显示了用单个per发夹实现合成端粒酶的示意图。图5b显示的page变性凝胶表明在不同的发夹浓度下的调聚反应(telomerization)。将引物与给定的发夹浓度一起在37℃温育4小时。图5c显示了无标记生物传感器的示意图,其中mirna靶标激活荧光端粒链的合成。图5d显示了生物传感器的系统组分和反应简图。设计门控的发夹(‘a’)和端粒酶发夹(‘b’)以反应来检测mirna信号和人端粒序列ttaggg的连环重复序列,在其中嵌入硫代黄素(tht)且变得发荧光。图5e显示的天然page凝胶表明在有10nmmirna信号存在下的条件调聚反应。透过琥珀色滤波器单元(vis)用蓝光透照仪可以直接观察靶标检测,且在fam通道(fam)下在typhoon扫描仪上也会观察反应的荧光。
实施例3
图6a显示了用rna输入评价逻辑表达的操作示意图。图6b显示了mir-19aortwt门反应组分和page变性凝胶,其描绘了响应于不同rna输入的转录物产生。如虚线指示的,通过寻找特定长度的转录物而读出真实输出。也显示了mir-19aandtwt(图6c)、notmir-21(图6d)以及(mir-19aortwt)and(notmir-21)(图6e)的反应设置和page变性凝胶结果。
实施例4
在该研究中,试验了两种不同的支点开关‘2_4'和‘2_10',之后接着在质粒上编码的gfp。还试验了在质粒上编码的几种dna触发物以及rna触发物。将在没有开关构建体的质粒上编码的gfp基因用作阳性对照。用不相应的开关、触发物对(‘sw2_4+trig2_10.35’和‘sw2_10+trig2_4.35’)观察到非常少的串扰(图7)。
在质粒上的有关序列如下。关于开关,所述序列开始于gcgc和t7启动子且终止于接头序列。关于触发物,所述序列开始于gcgc和t7启动子且终止于t7终止子的部分。
sw2_4
gcgctaatacgactcactatagggagtaagataatgaaggtaggtatgttaaactttagaacagaggagataaagatgaacatacctacgaacctggcggcagcgcaa(seqidno:1)
sw2_10
gcgctaatacgactcactatagggattgaatatgatagaagtttagtagtagacaatagaacagaggagatattgatgactactaaactaaacctggcggcagcgcaa(seqidno:2)
trig2_4
gcgctaatacgactcactatagggctcgatcactaatctgatcgagacgaacatacctaccttcattatcttacttgttagcataaccccttggggc(seqidno:3)
trig2_10
gcgctaatacgactcactatagggatacacatagaatcatgtgtataacactactaaacttctatcatattcaatcactagcataaccccttggggc(seqidno:4)
dna触发物序列:
2_04.32acgaacatacctaccttcattatcttacttgt(seqidno:5)
2_04.35acgaacatacctaccttcattatcttactccccac(seqidno:6)
2_04.38acatttaacatacctaccttcattatcttactccccac(seqidno:7)
2_10.32aacactactaaacttctatcatattcaatcac(seqidno:8)
2_10.35aacactactaaacttctatcatattcaatccccac(seqidno:9)
2_10.38aactctactactaaacttctatcatattcaatccccac(seqidno:10)
5μl溶液a
3.75μl溶液b(*向*a添加)
质粒dna
触发物(30μl储备物中的1.25μl)
h2o补至12.5μl
在37℃温育4小时以后,在biotek平板读数器上测量gfp荧光。
反应中的组分(开关,触发物):
2_4,2_10.35
2_4,2_4.32
2_4,2_4.35
2_4,2_4.38
2_4,2_4rna触发物
2_10,2_4.35
2_10,2_10.32
2_10,2_10.35
2_10,2_10.38
2_10,2_10rna触发物
gfp阳性对照
实施例5
该研究试验了通过per反应来延伸引物以产生支点开关的dna触发物序列。3-步延伸反应或1-步延伸反应工作良好(图8)。
per发夹的dna序列:
h1.1.2_04acctaccttcgggccttttggcccgaaggtaggtatgttcgt/3invdt/(seqidno:11)
h2.1.2_04attatcttgggccttttggcccaagataatgaaggtagg/3invdt/(seqidno:12)
h3.1.2_04actccccacgggccttttggcccgtggggagtaagataatga/3invdt/(seqidno:13)
h1.2.2_04acctaccttcgggccttttggcccgaaggtaggtatgttcg/3invdt/(seqidno:14)
h2.2.2_04attatcttgggccttttggcccaagataatgaaggtag/3invdt/(seqidno:15)
h3.2.2_04actccccacgggccttttggcccgtggggagtaagataatg/3invdt/(seqidno:16)
h.full.2_04acctaccttcattatcttactccccacgggccttttggcccgtggggagtaagataatgaaggtaggtatgttcgt/3invdt/(seqidno:17)
per(50μl反应):
12.5μlbstlf
12.5μlp.2_04至1μm终浓度
12.5μl发夹至100nm终浓度
12.5μldntp(仅a、t、c)至100μm终浓度
对照:h2.*.204+h3.*.204
3-步:h1.*.2_04+h2.*.2_04+h3.*.2_04
*={1,2}
反应:
a)无发夹
b)对照(无h.1.*.2_04发夹)(4个泳道)
c)3-步(8个泳道)
d)1-步(h.full.2_04)
实施例6
在该研究中,将得到的per反应的反应混合物加入具有支点开关的
实施例7
在该实施例中,使用在图12b中描绘的两个钥匙/锁对试验了6种不同图形设计的性能。描绘所述方法以及结果的图像的示意图显示在图13中。首先使用蜡印刷机将6种不同的图形设计印刷在滤纸上。然后将沉积的蜡在150℃加热2-3分钟以使蜡融化透过滤纸以建立离散疏水屏障。将锁‘1’(12μm)、锁‘2’(12μm)和游离荧光阳性对照链‘f’(5μm)中的每一个沉积在每个图形设计的离散疏水屏障内。将仅含有锁‘2’的钥匙的溶液(10μm:200μl钥匙,在2ml溶液总体积中)在6种设计中的每一种上喷2-3次,覆盖用锁或对照荧光链填充的每个离散疏水屏障。在蓝光透照仪下通过琥珀色滤波器单元的显现表明仅在含有锁‘2’和对照的疏水屏障内的荧光。
还将图12b所示的钥匙/锁对施加于图14a所示的另一个图形设计。将锁‘1’和锁‘2’溶液以预定方式沉积在图形‘x’设计内(在形成检查标志图案的离散区域中)。当施加于图形‘x’设计时,由荧光检查标志形状所描绘的,钥匙‘2’溶液仅将锁‘2’区域解锁。
实施例8
本实施例描述了用于读出基于纸的dna钥匙测定的成像仪。蓝光安全地且有效地激发荧光素(fam)或类似的荧光团。如在图16中所示,短通滤波器可以改良蓝色入射光,且长通滤波器(诸如mcmaster琥珀色丙烯酸透射率)可以相当好地透过fam发射,用于fam的最佳检测。在图17中所示的手持成像仪与上述的光路一起工作,在暗盒中利用蓝色led和短通和长通滤波器进行。
实施例9
本实施例描述了在含有醇的钥匙溶液中的锁/钥匙机制的试验。设计一个实验来在含有多达70%乙醇(etoh)的钥匙溶液中评价锁和钥匙对。
首先,通过在1xte+10mmmgcl2溶液中将含有荧光团的链(500nm)与过量猝灭剂(666.7nm)混合,使在图12b中描绘的荧光团/猝灭剂对1退火。然后将该溶液加热至80℃保持2分钟,然后在室温保持20分钟。随后,将10μl溶液分入384平板的24个孔中的每个。
接着,在室温,在所述孔中混合10μl的24种溶液中的一种与百分数在0%(无)至70%的范围内的乙醇(从100%乙醇溶液稀释),所述溶液不含有寡聚物或含有在图12b中描绘的两种钥匙触发链之一。试验了24种不同的条件:无钥匙,0%乙醇;无钥匙,10%乙醇;无钥匙,20%乙醇;无钥匙,30%乙醇;无钥匙,40%乙醇;无钥匙,50%乙醇;无钥匙,60%乙醇;无钥匙,70%乙醇;错误的钥匙(荧光团/猝灭剂对2的钥匙链),0%乙醇;错误的钥匙(荧光团/猝灭剂对2的钥匙链),10%乙醇;错误的钥匙(荧光团/猝灭剂对2的钥匙链),20%乙醇;错误的钥匙(荧光团/猝灭剂对2的钥匙链),30%乙醇;错误的钥匙(荧光团/猝灭剂对2的钥匙链),40%乙醇;错误的钥匙(荧光团/猝灭剂对2的钥匙链),50%乙醇;错误的钥匙(荧光团/猝灭剂对2的钥匙链),60%乙醇;错误的钥匙(荧光团/猝灭剂对2的钥匙链),70%乙醇;正确的钥匙(荧光团/猝灭剂对1的钥匙链),0%乙醇;正确的钥匙(荧光团/猝灭剂对1的钥匙链),10%乙醇;正确的钥匙(荧光团/猝灭剂对1的钥匙链),20%乙醇;正确的钥匙(荧光团/猝灭剂对1的钥匙链),30%乙醇;正确的钥匙(荧光团/猝灭剂对1的钥匙链),40%乙醇;正确的钥匙(荧光团/猝灭剂对1的钥匙链),50%乙醇;正确的钥匙(荧光团/猝灭剂对1的钥匙链),60%乙醇;和正确的钥匙(荧光团/猝灭剂对1的钥匙链),70%乙醇。
然后在33.8℃在平板读数器上测量这24种不同的钥匙/锁反应的荧光值。结果显示在图12c-12d中。图12c显示了以对数标度绘制的24个孔中的每个的原始荧光值。在所有情况下,正确的钥匙和锁对的荧光显著高于具有相同乙醇浓度的其它两种条件(“无钥匙”和“错误的钥匙”)的荧光。图12d显示了试验的8种乙醇条件中的每种的on/off比率,其通过将正确的钥匙和锁对溶液的荧光除以不具有钥匙的溶液的荧光来计算。在所有情况下,on/off比率远远超过30倍并自始至终保持大致恒定。这些数据证实了在图12b中描绘的设计所依赖的杂交和链置换反应可以在有醇存在下运行。
实施例10
本实施例描述了固定放大的几种机制。如在图18a中所示,所述锁或钥匙链包含超过一种荧光团和猝灭剂,从而导致2或3次荧光信号输出(“on”状态)。在图18b所示的一个备选实施方案中,单一猝灭剂会阻止来自2个荧光团链的荧光团信号传导(“off”状态;上部图)。当锁和钥匙相互作用时,2个荧光团链从猝灭剂链除去,从而导致放大的荧光信号(图18b,底部图)。
放大也可以由杂交链式反应导致,如在图19a-19c中所示。终产物(图19c)中的每个发夹是独特的,保持至少一个荧光团和/或猝灭剂部分。所有发夹存在于溶液中,从而形成锁机制(图19a)。发夹的整个级联被单个钥匙触发以相继次序打开(在图19b中的链‘ax’),从而从单个钥匙链激活固定数目的荧光团。所有溶液、发夹/无活性状态和活化的聚合物状态(图19c)都是热力学平衡,且因此,所述系统在储存中得以稳定。
实施例11
本实施例描述了未修饰的钥匙链的应用,其在用于鉴定处方药物或其它物质(在其中不希望引入经修饰的dna钥匙)的系统中可以是有益的。
图20a显示了一个实施例,其中每个产品含有独特的钥匙链和测试条。所述钥匙链可以是未修饰的。所述测试条将产品钥匙转换成可见代码。所述可见代码是长的且独特的条形码图案,其然后在生产商的网站上验证真实性。每个测试条仅可以使用一次,因为每个试验是独特的,从而保护免于一个真实产品被稀释成许多复制的产品。
图20b显示了未修饰的钥匙应用的另一个例子。在该情况下,为了额外的安全性单独运输产品包,所述产品包包括钥匙和试验包,所述试验包包括测试条和对照。给出的钥匙序列仅在短时间段中是有效的,诸如当将特定的未修饰的dna链混合在液体或固体药物制剂内时。所述测试条具有失效日期和所述试验相容的序列号的范围。另外,所述锁链被保护免于通过共价修饰或其它方式读出。因此,从当前药物批次读出钥匙链将仅在非常短的时间段中起作用,因为生产商会在失效日期以后改变掺入的钥匙链。
参考文献,其中每篇并入本文
[1]grosse,eric,和mayankupadhyay.“authenticationatscale.”ieeesecurity&privacy11.1(2013):15-22.
[2]corcoran,peter,和claudiacostache.“biometrictechnologyandsmartphones:aconsiderationofthepracticalitiesofabroadadoptionofbiometricsandthelikelyimpacts.”ieeeconsumerelectronicsmagazine5.2(2016):70-78.
[3]corcoran,peterm.“biometricsandconsumerelectronics:abravenewworldortheroadtodystopia?”ieeeconsumerelectronicsmagazine2.2(2013):22-33.
[4]fiebig,tobias,jankrissler,和ronny
[5]green,alexandera.,等人.“toeholdswitches:de-novo-designedregulatorsofgeneexpression.”cell159.4(2014):925-939.
[6]pardee,keith,等人.“paper-basedsyntheticgenenetworks.”cell159.4(2014):940-954.
[7]tribioli,carla,和thomaslufkin.“long-termroomtemperaturestorageofhigh-qualityembryonicstemcellgenomicdnaextractedwithasimpleandrapidprocedure.”journalofbiomoleculartechniques:jbt17.4(2006):249.
[8]anchordoquy,thomasj.,等人.“physicalstabilizationofdna-basedtherapeutics.”drugdiscoverytoday6.9(2001):463-470.
本文中公开的所有参考文献、专利和专利申请关于各自引用的主题通过引用并入,其在某些情况下可以包括所述文件的整体。
除非明确地相反指出,否则本文在说明书中和在权利要求中使用的不定冠词“一个/种(a)”和“一种/个(an)”应当被理解为是指“至少一个/种”。
还应当理解,除非明确地相反指出,否则在本文要求保护的包括超过一个步骤或动作的任何方法中,所述方法的步骤或动作的次序不一定限于列举所述方法的步骤或动作的次序。
在权利要求中以及在上述说明书中,所有过渡短语诸如“包含”、“包括”、“携带”、“具有”、“含有”、“涉及”、“容纳”、“由……组成”等应当理解为开放式的,即,意在包括但不限于。仅过渡短语“由……组成”和“基本上由……组成”应当分别是封闭式或半封闭式过渡短语,如在美国专利局专利审查程序手册(unitedstatespatentofficemanualofpatentexaminingprocedures)第2111.03部分中所述。
序列表
<110>presidentandfellowsofharvardcollege
<120>分子验证系统
<130>h0498.70624wo00
<140>notyetassigned
<141>2017-09-19
<150>us62/502,522
<151>2017-05-05
<150>us62/407,331
<151>2016-10-12
<150>us62/396,932
<151>2016-09-20
<160>17
<170>patentinversion3.5
<210>1
<211>108
<212>dna
<213>人工序列
<220>
<223>合成的多核苷酸
<400>1
gcgctaatacgactcactatagggagtaagataatgaaggtaggtatgttaaactttaga60
acagaggagataaagatgaacatacctacgaacctggcggcagcgcaa108
<210>2
<211>108
<212>dna
<213>人工序列
<220>
<223>合成的多核苷酸
<400>2
gcgctaatacgactcactatagggattgaatatgatagaagtttagtagtagacaataga60
acagaggagatattgatgactactaaactaaacctggcggcagcgcaa108
<210>3
<211>97
<212>dna
<213>人工序列
<220>
<223>合成的多核苷酸
<400>3
gcgctaatacgactcactatagggctcgatcactaatctgatcgagacgaacatacctac60
cttcattatcttacttgttagcataaccccttggggc97
<210>4
<211>97
<212>dna
<213>人工序列
<220>
<223>合成的多核苷酸
<400>4
gcgctaatacgactcactatagggatacacatagaatcatgtgtataacactactaaact60
tctatcatattcaatcactagcataaccccttggggc97
<210>5
<211>32
<212>dna
<213>人工序列
<220>
<223>合成的多核苷酸
<400>5
acgaacatacctaccttcattatcttacttgt32
<210>6
<211>35
<212>dna
<213>人工序列
<220>
<223>合成的多核苷酸
<400>6
acgaacatacctaccttcattatcttactccccac35
<210>7
<211>38
<212>dna
<213>人工序列
<220>
<223>合成的多核苷酸
<400>7
acatttaacatacctaccttcattatcttactccccac38
<210>8
<211>32
<212>dna
<213>人工序列
<220>
<223>合成的多核苷酸
<400>8
aacactactaaacttctatcatattcaatcac32
<210>9
<211>35
<212>dna
<213>人工序列
<220>
<223>合成的多核苷酸
<400>9
aacactactaaacttctatcatattcaatccccac35
<210>10
<211>38
<212>dna
<213>人工序列
<220>
<223>合成的多核苷酸
<400>10
aactctactactaaacttctatcatattcaatccccac38
<210>11
<211>43
<212>dna
<213>人工序列
<220>
<223>合成的多核苷酸
<220>
<221>misc_feature
<222>(43)..(43)
<223>3'反向的dt
<400>11
acctaccttcgggccttttggcccgaaggtaggtatgttcgtt43
<210>12
<211>40
<212>dna
<213>人工序列
<220>
<223>合成的多核苷酸
<220>
<221>misc_feature
<222>(40)..(40)
<223>3'反向的dt
<400>12
attatcttgggccttttggcccaagataatgaaggtaggt40
<210>13
<211>43
<212>dna
<213>人工序列
<220>
<223>合成的多核苷酸
<220>
<221>misc_feature
<222>(43)..(43)
<223>3'反向的dt
<400>13
actccccacgggccttttggcccgtggggagtaagataatgat43
<210>14
<211>42
<212>dna
<213>人工序列
<220>
<223>合成的多核苷酸
<220>
<221>misc_feature
<222>(42)..(42)
<223>3'反向的dt
<400>14
acctaccttcgggccttttggcccgaaggtaggtatgttcgt42
<210>15
<211>39
<212>dna
<213>人工序列
<220>
<223>合成的多核苷酸
<220>
<221>misc_feature
<222>(39)..(39)
<223>3'反向的dt
<400>15
attatcttgggccttttggcccaagataatgaaggtagt39
<210>16
<211>42
<212>dna
<213>人工序列
<220>
<223>合成的多核苷酸
<220>
<221>misc_feature
<222>(42)..(42)
<223>3'inverteddt
<400>16
actccccacgggccttttggcccgtggggagtaagataatgt42
<210>17
<211>77
<212>dna
<213>人工序列
<220>
<223>合成的多核苷酸
<220>
<221>misc_feature
<222>(77)..(77)
<223>3'反向的dt
<400>17
acctaccttcattatcttactccccacgggccttttggcccgtggggagtaagataatga60
aggtaggtatgttcgtt77
- 还没有人留言评论。精彩留言会获得点赞!