用于检测和治疗胰岛素抵抗的组合物和方法与流程
相关申请
这是pct申请,要求35u.s.c.119的利益,其基于2018年7月13日提交的临时专利申请第62/697,935号和2017年8月25日提交的临时专利申请第62/550,233号的优先权,其各自通过引用整体上并入本文。
序列表
于2018年8月23日创建的计算机可读形式的序列表“sequencelisting.txt”(9782字节)通过引用并入本文。
本公开涉及与胰岛素抵抗有关的mirna及其拮抗剂作为治疗剂的用途。
背景技术:
下丘脑用作体内营养状况和能量稳态的关键调节剂。协调食物摄入和能量消耗的表达神经肽y(npy)/刺鼠调节肽(agrp)-和阿黑皮素原(pomc)的神经元对于维持能量稳态至关重要。定位于弓状核(arc)(gao和horvath,2008;varela和horvath,2012),位于第三脑室附近,并向正中隆起投射,共表达刺鼠相关肽(agrp)的这些npy神经元,和pomc神经元理想地定位于感知循环因子(诸如外周激素和饮食成分)的变化。该信息被传递到下丘脑的其他核以及其他大脑区域,以维持健康的能量平衡。npy/agrp或pomc神经元群的功能破坏,特别是那些改变其对周围信号(如胰岛素和瘦素)响应的那些功能破坏,与肥胖、胰岛素抵抗和ii型糖尿病(t2dm)的发展有关(konner和bruning,2012;roh等,2016)。还已经确定,通过低脂饮食减轻体重会导致肥胖小鼠中循环microrna的差异表达(hsieh等,2015)。
在外周组织中胰岛素抵抗是相当明确定义的,但是在脑中潜在的病理学特征较少(kleinridders等,2014)。在啮齿动物模型中进行脑胰岛素信号传导的研究在确定胰岛素信号传导在预防饮食诱导的肥胖,响应高脂肪摄入引起的肝葡萄糖生成失调以及维持正常的白色脂肪组织脂肪生成和脂肪分解(其在t2dm中受到干扰)中发挥着至关重要的作用(bruning等,2000;obici等,2002b;scherer等,2011)。进一步支持大脑在代谢失调发展中的重要性,clegg等研究发现,胰岛素抵抗在大脑中的发生先于外周组织,并在高脂饮食(hfd)下的肥胖发展之前(clegg等,2011)。实际上,仅arc的npy/agrp神经元中胰岛素受体(insr)水平的部分降低就足以诱发大鼠中的胰岛素抵抗和摄食过量(obici等,2002a)。尽管最近发现,npy神经元中的特异性胰岛素信号传导对于控制食物摄入和能量平衡至关重要(loh等,2017)。因此,中枢胰岛素信号传导的受损可能充当周围胰岛素抵抗发展的促发因素,然而,这种病理发生的机制仍未得到充分研究。
可以引发胰岛素抵抗的高胰岛素血症也可以独立于肥胖发生(rizza等,1985)。例如,已经证明,暴露于高水平胰岛素的npy/agrp神经元细胞模型在8小时至24小时之间变得胰岛素抵抗(mayer和belsham,2010a)。insr和insr底物1(irs-1)的蛋白水平降低,但其mrna水平却没有降低,这归因于观察到的细胞信号传导废止。延长胰岛素暴露后,蛋白体和溶酶体途径分别与irs-1和insr降解相关。类似地,也发现表达pomc的神经元细胞模型通过这种范例发展为胰岛素抵抗(nazarians-armavil等,2014)。这些实验阐明了在下丘脑细胞中高胰岛素暴露后,胰岛素信号传导途径中的关键蛋白可以被降解的方式。
技术实现要素:
根据本发明的一个方面,提供了一种mir-1983抑制剂,其包含与ctcacctggagcatgttttct(seqidno:1)的至少一部分互补的抗mir-1983寡核苷酸,所述部分至少包含ctcacctggagcatgttttct(seqidno:1)的核苷酸2至8。
在一个实施方式中,mir-1983抑制剂是抗mir-1983寡核苷酸。
根据本发明的另一方面,存在一种包含mir-1983抑制剂和稀释剂的组合物。
根据本发明的又一方面,存在一种检测细胞是否为胰岛素抵抗的方法,所述方法包括测量mir-1983的水平并与阈值或对照进行比较,其中,与对照相比,水平升高表明该细胞是胰岛素抵抗的。
根据另一方面,存在一种在受试者中检测糖尿病前期或发展糖尿病的可能性增加的方法,所述方法包括测量来自该受试者的生物样品中的mir-1983的水平,其中,与阈值或对照相比,水平升高表明所述受试者患有糖尿病前期或发展糖尿病的可能性增加。
根据另一方面,存在一种改善受试者的胰岛素敏感性的方法,所述方法包括向有需要的受试者施用mir-1983抑制剂。
根据以下具体实施方式,本公开的其他特征和优点将变得显而易见。然而,应当理解的是,在示出本公开的优选实施方式的同时,具体实施方式和具体实施例仅是举例说明的,这是因为由于在本公开的精神和范围内的各种变化和修改对于本领域技术人员而言从该具体实施方式是显而易见的。
附图描述
下面结合附图描述实施方式,其中:
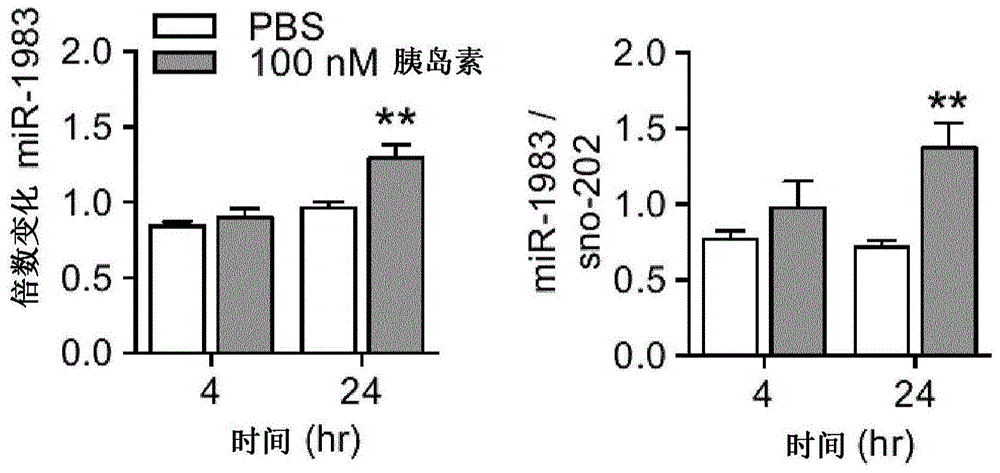
图1描绘了在诱导胰岛素抵抗后发现在mhypoe-46细胞系中改变的mirna的微阵列(n=3)和qpcr验证(n=4-5)。图1a描绘了mir-1983,图1b描绘了mir-20a-3p,图1c描绘了mir-296-5p,图1d描绘了let-7d-5p.*p<0.05,**p<0.01。
图2显示了mir-1983在成年细胞系中被调节,并且在胰岛素治疗后并未永久上调。在图2a的雌性mhypoa-npy/gfp(n=3-7)和图2b的雄性mhypoa-pomc/gfp-2(n=8-16)两个成体细胞系中,胰岛素抵抗诱导mir-1983的差异变化,但不引起mir-20a-3p的差异。图2c在48小时后,mhypoe-46细胞中没有mir-1983上调,而insrβ亚基蛋白水平的损失仍在持续(n=8)。显示的数据为平均值±s.e.m。*p<0.05,**p<0.01,****p<0.0001。
图3描绘了mir-1983的mrna靶标的计算机和体外分析。图3a显示了用于评估目的mir-1983基因靶标的计算机分析工作流示意图,图3b是小鼠中胰岛素受体(insr)3'非翻译区(3'utr)的靶标扫描分析(上图),以及mir-1983的insr3'utr序列靶标区域的保守性跨物种比对(下图)。图3c显示在将mir-1983mirna模拟物(25nm)转染到mhypoe-46细胞24小时后(n=4),insr的mrna水平未改变。图3d将mir-1983mirna模拟物(25nm)转染到mhypoe-46细胞24小时后(n=4),insrβ蛋白水平降低。图3e将mir-1983mirna模拟物(25nm)转染到mhypoe-46细胞48小时后(n=4),insr的mrna水平不变,而将mir-1983mirna模拟物(25nm)转染到mhypoe-46细胞48小时后(n=4),insrβ蛋白水平降低。图3f包含约3000bp3'insr-utr序列的pmirglo载体(1μg)与mir-1983模拟物(25nm)同时进行转染(n=3-4个独立实验/组,每个重复三次)。*显示的数据为平均值±s.e.m。p<0.05,**p<0.01。
图4显示尽管在神经肽mrna水平上表现出相似的变化,在喂食hfd7周至9周的雄性而非雌性的c57bl/6小鼠的下丘脑中mir-1983被上调。与7周至9周的蔗糖匹配的对照饮食(食物)相比,饲喂60%hfd的雄性和雌性c57bl/6小鼠体重增加(图4a,n=4-7/组),并且空腹血糖升高(图4b,n=3-7/组),并且显示了npy、agrp和insr(而不是pomc表达)的相似变化(图4c&d,n=7-8/组)。仅雄性c57bl/6表现出空腹胰岛素和下丘脑mir-1983水平升高(图4e&f,n=7-8/组),*p<0.05,**p<0.01,****p<0.0001。显示的数据为平均值±s.e.m。
图5显示在神经肽mrna水平完全改变之前,在饲喂hfd5周而非10周后,雄性cd-1小鼠的下丘脑中mir-1983被上调。与蔗糖匹配的对照饮食相比,饲喂60%hfd5周的小鼠的循环空腹血糖(图5a,n=4/组)和胰岛素水平(图5b,n=8/组)增加。在这些小鼠的下丘脑中mir-1983的表达增加(图5c,n=9-12),而npy、agrp、pomc或insrmrna水平无变化(图5d,n=8-14/组)。饲喂相同饮食10周的cd-1小鼠的下丘脑mir-1983水平没有升高(图5e,n=10-11/组),但mrna中的npy和agrp降低,而pomcmrna的升高。未检测到insrmrna丰度变化(图5f,n=8-11/组)。显示的数据为平均值±s.e.m,*p<0.05,**p<0.01,***p<0.001。
图6显示饲喂60%hfd5周的cd-1雌性小鼠的下丘脑中mir-1983没有升高,而雄性小鼠在其下丘脑和血清中表现出上调,而在肾脏中的水平降低。相对于同窝饲喂食物,对照组和60%hfd组的两种性别的体重增加(图6a&b,n=8-9/组)并且总体重增加(图6c,n=8-9/组)。饲喂两种食物(蔗糖匹配的对照饮食)或60%hfd的雄性和雌性cd-1小鼠4小时的空腹血糖(n=7-9/组)(图6d,n=7-9/组)。(图6e)与mkr(n=8)小鼠相比,饲喂食物(n=7-8),或60%hfd(n=8)的雄性cd-1小鼠的下丘脑mir-1983和空腹胰岛素水平。(图6f)与mkr(n=3)小鼠相比,饲喂食物(n=7-8),或60%hfd(n=8)的雄性和雌性cd-1小鼠的下丘脑mir-1983和空腹胰岛素水平。雄性(图6g,n=7-9/组)和雌性(图6h,n=8-9/组)小鼠表现出较低水平的npymrna,而agrp或pomcmrna的量没有变化;但是,insrmrna水平仅在雌性小鼠中增加。(图6i)与60%hfd相比,在饲喂食物的雄性cd-1小鼠中肌肉、肾脏和血清(n=5-8/组)中的mir-1983水平。显示的数据为平均值±s.e.m,*p<0.05,**p<0.01,****p<0.0001。
图7显示在60%hfd方案5周后,cd-1小鼠中体内鼻内递送mir-1983抑制剂或对照,诱导胰岛素敏感性、空腹血糖和下丘脑insrβ-亚基蛋白水平的变化。(图7a)向60%hfd的小鼠体内施用mir-1983抑制剂的实验范例(时间线)。与食物(与蔗糖匹配的对照饮食)相比,这些动物表现出体增重(图7b,n=5-6/组)和总体重(图7c,n=5-6/组)的增加。(图7d)腹膜内胰岛素耐受性测试(ipitt),(禁食4h加1单位胰岛素/kg体重)(n=5-6/组)。(图7e)在鼻内施用mir-1983抑制剂后两周后的空腹血糖水平(n=5-6/组)。(图7f)神经肽下丘脑npy、agrp、pomc和insrmrna水平在5周后在两个60%hfd组中相似地降低。(图7g)在5周研究结束时,在60%hfd的小鼠中而不是insrβ-亚基蛋白亚基降低,但在mir-1983抑制剂组中未降低(n=5-6/组)。显示的数据为平均值±s.e.m,*p<0.05,**p<0.01,***p<0.001,****p<0.0001。
图8描绘了在mhypoe-46细胞系中100nm胰岛素治疗后改变的60个mirna的热图。
图9描绘了在所有处理组中4小时和24小时改变的60个mirna的维恩图。载体pbs处理(veh)和100nm胰岛素处理(ins)。
图10描绘了验证数据,其表明在用100nm胰岛素处理4或24小时的mhypoe-46细胞中insr的mrna水平保持不变(图10a,n=5)。该细胞模型表现出胰岛素抵抗的insrβ亚基蛋白水平降低特性(图10b,n=10)。显示的数据为平均值±s.e.m。****p<0.0001.
图11显示,在雄性小鼠以及男性和女性人类受试者中,血清mir-1983水平与循环胰岛素水平相关。(图11a)用食物(蔗糖匹配的对照饮食)或60%hfd饲喂的雄性和雌性cd-1小鼠的总体重(n=8-9/组)。(图11b)在饲喂食物或60%hfd的雄性cd-1小鼠中,血清mir-1983水平与葡萄糖(左)和胰岛素(右)的相关性。(图11c)通过体重指数分开的人类中的mir-1983的血清水平为<25(n=2,sd显示为5名受试者没有可检测到的mir-1983水平),25-29.9(n=8),>30(n=9)。(图11d)血清mir-1983水平与各种血液参数的相关性分析(实心圆具有统计学意义,完整的分析细节参见表1)。(图11e)血清胰岛素(左)和稳态模型评估-胰岛素抵抗(homa-ir)(右)分数与人血清mir-1983水平的线性回归分析,除非另有说明,否则显示的数据为平均值±s.e.m。*p<0.05,**p<0.01,***p<0.001,****p<0.0001.
具体实施方式
除非另有定义,否则结合本公开使用的科学和技术术语应具有本领域普通技术人员通常理解的含义。此外,除非上下文另有要求,否则单数术语应包括复数,复数术语应包括单数。例如,术语“细胞”包括单个细胞以及多个细胞或细胞群。通常,本文描述的与细胞和组织培养、分子生物学以及蛋白质和寡核苷酸或多核苷酸化学和杂交相关的术语和技术是本领域众所周知的和常用的(参见,例如green和sambrook,2012)。
本文中通过端点对数值范围的引用包括该范围内所包含的所有数字和分数(例如1至5包括1、1.5、2、2.75、3、3.90、4和5)。还应理解的是,假定所有数字及其分数均被术语“约”修饰。此外,应理解的是,除非内容另有明确规定,否则“一个”,“一种”和“该”包括复数个指示物。术语“约”是指所参考数字的正负0.1%至20%,5%至20%或10%至20%,优选5%至15%,更优选5%或10%。
如本文所用,术语“包含”及其衍生词是开放式术语,其指定存在所述特征、元素、组件、组、整数和/或步骤,但不排除其他未述特征、元素、组件、组、整数和/或步骤的存在。前述内容也适用于具有类似含义的词,诸如术语“包括”、“具有”及其衍生词。最后,本文所用的程度术语,诸如“基本上”、“约”和“大约”,是指修饰术语的合理偏离量,使得最终结果不会显著改变。这些程度的术语应被解释为包括至少修饰词的±5%的偏离,如果该偏离不会否定其修饰词的含义。
在理解本公开的范围时,本文所用术语“由…….组成”及其衍生词是旨在指定所述特征、元素、组件、组、整数和/或步骤的存在的封闭式术语,并且还排除其他未述特征、元素、组件、组、整数和/或步骤的存在。
此外,如本领域技术人员所理解的,在特定部分中描述的定义和实施方式旨在适用于它们适合的本文所述的其他实施方式。例如,在以下段落中,更详细地定义了本公开的不同方面。如此定义的每个方面可以与任何一个或多个其他方面组合,除非明确指出相反。特别是,指示为优选或有利的任何特征可以与指示为优选或有利的任何其他一个或多个特征组合。
如本文所用的程度术语,诸如“约”、“基本上”和“大约”是指修饰术语的合理偏离量,使得最终结果不会显著改变。这些程度术语应被解释为包括修饰术语的至少±5%的偏离,如果该偏离不会否定其修饰词的含义。
如本文所用,术语“核酸”是指由天然存在的碱基、糖和糖间(主链)键组成的核苷或核苷酸单体的序列。该术语还包括包含非天然存在的单体或其部分的修饰或取代的序列。本公开的核酸可以是脱氧核糖核酸序列(dna)或核糖核酸序列(rna),并且可以包括天然存在的碱基,其包括腺嘌呤、鸟嘌呤、胞嘧啶、胸腺嘧啶和尿嘧啶。序列还可以包含修饰的碱基,例如提供以增加对核酸酶抗性。核酸可以是双链的或单链的,并且代表有义链或反义链。此外,术语“核酸”包括互补核酸序列以及密码子优化的或同义密码子等同物。如本文所用,术语“分离的核酸序列”是指当通过重组dna技术产生时基本上不含细胞材料或培养基的核酸,或当通过化学合成时产生的化学前体或其他化学物质。分离的核酸也基本上不含天然衍生于该核酸的核酸侧翼的序列(即位于该核酸的5'和3'末端的序列)。术语“核酸”还可以包括例如“反义核酸”、“反义寡核苷酸”、“mirna”、抗mirna等。
如本文所用,术语“寡核苷酸”是指短核酸,其包含由天然和非天然存在的碱基、糖和糖间(主链)键组成的核苷酸或核苷单体的序列,并且包括单链和双链分子,rna和dna。通常,寡核苷酸少于50个单体,包括非天然存在的单体,更通常为30个单体以下。术语“寡核苷酸”包括,例如“反义寡核苷酸”和“mirna”以及寡核苷酸类似物,如“吗啉代寡核苷酸”、“硫代磷酸寡核苷酸”。
本文所用的术语“化学修饰的寡核苷酸”是指包含至少一种非天然存在的单体(例如,修饰的单体)的寡核苷酸。此类化学修饰的寡核苷酸可具有与未修饰的寡核苷酸相似或比其更好的结合能力。在上下文中所用术语“相似”是指化学修饰的寡核苷酸以其缺乏特异性修饰的多核苷酸的20%以内的结合特异性结合其靶标。化学修饰可以是在锁核酸(lna)中发现的一种,也可以是2'-氟(2'-f)、2'-o-甲氧基乙基(2'-moe)或2'-o-甲基(2'-o-me),它们是以下2'位置的修饰:核糖部分,或其中六元吗啉环取代了糖部分的吗啉代单体,或其中硫取代了磷酸基团中的非桥接氧原子中的一个的硫代磷酸酯(ps)键。非天然存在的单体可以包含一种或多种化学修饰。例如,硫代磷酸酯键也可以掺入任何上述化学修饰的寡核苷酸中。由于诸如在核酸酶存在下增加的稳定性等性质,这种修饰或取代的核酸可能优于天然存在的形式。该术语还包括含有两个以上化学上不同区域的嵌合核酸。例如,嵌合核酸可包含至少一个赋予有益特性(例如,增加的核酸酶抗性,增加的细胞的摄取)的修饰的核苷酸区域,或两个修饰的核苷酸区域。
“反义核酸”或“反义寡核苷酸”包含与“有义”核酸互补(例如,与mir-1983互补)的核苷酸序列。因此,反义核酸可与有义核酸氢键结合。例如,核酸可以包含与靶dna或rna结合的dna、rna或化学类似物(例如,包含一种或多种修饰的单体)。可以使用天然存在的核苷酸或各种修饰的核苷酸化学合成反义核酸分子,所述修饰的核苷酸被设计成增加分子的生物稳定性或增加与靶rna或dna形成的双链体的物理稳定性,例如硫代磷酸酯衍生物和吖啶取代的核苷酸。反义核酸可以与整个靶链互补,或仅与其一部分互补。如本领域已知的,可以使用多种方法产生反义序列。可以使用以重组质粒、噬菌粒或减毒病毒的形式引入细胞中的表达载体,以生物学方式产生反义序列,其中在高效调节区的控制下产生反义序列,其活性可以由引入载体的细胞类型决定。此外,反义序列可从制造商处购买。
如本文所用,术语“抗mir-1983寡核苷酸”是指反义寡核苷酸,其长度为至少7个核苷酸并且与序列5’-3’,ctcacctggagcatgttttct(seqidno:1)的至少核苷酸2至8互补。抗mir-1983可以例如由dna、rna或lna单体或其他修饰的碱基或其任何组合组成。
术语“编码区”是指包含密码子的核苷酸序列的区域,该密码子被翻译成氨基酸残基。
术语基因或cdna的“非编码区”是指5'和3'序列,它们侧接在未翻译成氨基酸的编码区的侧翼(即也称为5'和3'非翻译区)。
术语“吗啉代单体”是指包含核酸碱基、6元吗啉环和非离子型二氨基磷酸酯亚基间键的亚基。吗啉代寡核苷酸是约25个吗啉代亚基的短链。每个吗啉代阻断核糖核酸(rna)碱基配对表面的小区域(约25个碱基)。
如本文所用,术语“细胞”包括多个细胞并且包括细胞系。
如本文所用,术语“细胞系”是指一组遗传上均匀的永生细胞,其在体外无限繁殖。细胞系可源自单个克隆(例如,单克隆细胞系)或来自多个克隆(例如,多克隆细胞系)。
如本文所用,术语“生物样品”是指源自受试者的流体样品或组织样品。流体样品的实例包括但不限于血液、血浆、血清、尿液、脊髓液、淋巴液、泪液、唾液、痰和乳汁。组织样品的实例包括脑组织样品或神经组织样品。获得此类生物样品的方法在本领域中是已知的,包括但不限于标准血液回收程序。
如本文所用,术语“方法”是指用于完成给定任务的方式、手段、技术和过程,其包括但不限于已知的方式、手段、技术和过程,或通过化学、药理、生物学、生化和医学领域的从业人员易于从已知的方式、手段、技术和过程发展的那些。
如本文所用,术语“治疗”包括废除、基本上抑制、减慢或逆转病症的进展,如减慢或逆转胰岛素抵抗的进展、基本上改善病症的临床或美学症状,如改善胰岛素敏感性或基本防止某种病症的临床或美学症状出现。
如本文所用,术语“施用”是指通过诸如鼻内施用途径等有效途径来提供或给予受试者一种试剂,诸如包含有效量的mir-1983抑制剂的组合物。
如本文所用,术语“有效量”是指足以产生期望的应答(如减轻或消除病症的征兆或症状)的试剂(如mir-1983抑制剂)的量。在某些实例中,“有效量”是治疗(包括预防)任何一种病症或疾病的一种或多种症状和/或潜在原因的量。有效量可以是治疗有效量,其包括防止特定疾病或病症的一种或多种征兆或症状发展的量,诸如与胰岛素抵抗或糖尿病前期相关的一种或多种征兆或症状。
如本文所用,术语“互补性”是指第一核酸与第二核酸之间的核碱基配对能力。
如本文所用,术语“胰岛素敏感性”是指细胞响应于激素胰岛素的正常作用的生理状况。这与胰岛素抵抗相反,后者是其中细胞无法对胰岛素的正常作用作出响应的生理状况。因此,细胞、组织或受试者中的“改善胰岛素敏感性”是指与参考水平(例如,采用所述方法和/或试剂之前的胰岛素敏感性)相比,细胞、组织或受试者中的胰岛素敏感性增加。
如本文所用,术语“锁核酸”是指双环rna类似物,其中核糖通过引入2'-o,4'-c亚甲基桥而被锁定在c3'-内构象中。所需的lna单体及其合成方法也公开在美国专利第6,043,060号、第6,268,490号、pct公开wo01/07455、wo01/00641、wo98/39352、wo00/56746、wo00/56748和wo00/66604,及以下论文中描述:morita等,bioorg.med.chem.lett.12(1):73-76,2002;hakansson等,bioorg.med.chem.lett.11(7):935-938,2001;koshkin等,j.org.chem.66(25):8504-8512,2001;kvaerno等,j.org.chem.66(16):5498-5503,2001;halkansson等,j.org.chem.65(17):5161-5166,2000;kvaerno等,j.org.chem.65(17):5167-5176,2000;pfundheller等,nucleosidesnucleotides18(9):2017-2030,1999;和kumar等,bioorg.med.chem.lett.8(16):2219-2222,1998。
术语“混聚体(mixmer)”是指包含天然和非天然存在的核苷酸的寡核苷酸。然而,不存在超过5个天然核苷酸的连续序列。
术语“a*c*a*t/u*g*c*t/u*c*c*a*g*g*t/u*g*a”(seqidno:2)是指所述序列的寡核苷酸。a*c*a*t/u*g*c*t/u*c*c*a*g*g*t/u*g*a”是mi-1983的互补链,其与mirna诱导的沉默复合体相关。
如本文所用,术语“错配”是指双链区域中的一个或多个错配的核苷酸。
如本文所用,术语“稀释剂”是指药学上可接受的载体,其不抑制待施用的活性化合物的生理活性或特性并且不刺激受试者并且不消除所施用的化合物的生物学活性和特性。稀释剂包括任何和所有溶剂、分散介质、涂料、表面活性剂、抗氧化剂、防腐剂盐、防腐剂、粘合剂、赋形剂、崩解剂、润滑剂,如本领域普通技术人员已知的材料及其组合(参见,例如,remington'spharmaceuticalsciences,第18版,mackprinting公司,1990年,第1289-1329页,通过引用并入本文)。除非任何常规载体与活性成分不相容,否则考虑将其用于药物组合物中。也可以使用允许或增强核酸跨细胞膜运输的稀释剂。
如本文所用,术语“糖尿病前期”是指血糖水平高于正常水平但不足以诊断糖尿病的病症。糖尿病前期为在开始口服葡萄糖耐受性测试(ogtt)2小时后,140mg/dl至199mg/dl的水平。
如本文所用,术语“鼻内制剂”是指配制用于鼻内递送的药物组合物,并且可以是粉末、水溶液、水性气雾剂、滴鼻剂和/或气雾剂的形式。
如本文所用,“药物组合物”是指本文所述的一种或多种活性成分(例如mir-1983抑制剂)与一种或多种其他化学成分(例如生理上合适的载体)的制剂。药物组合物的目的是促进化合物向生物体的施用。
如本文所用,术语“扩增”是指通常以模板依赖性方式复制靶多核苷酸、靶多核苷酸替代物或其组合的至少一部分的任何方式,其包括但不限于宽范围线性或指数扩增核酸序列的技术。进行扩增步骤的示例性程序包括聚合酶链式反应(pcr)。可以在本教导中使用的其他技术的描述可以在其他地方找到,sambrook等molecularcloning,第三版;ausbel等;pcrprimer:alaboratorymanual,diffenbach编,coldspringharborpress(1995);theelectronicprotocolbook,changbioscience(2002),msuih等,j.clin.micro.34:501-07(1996);thenucleicacidprotocolshandbook,r.rapley编,humanapress,totowa,n.j.(2002);abramson等,curropinbiotechnol.1993年2月;4(1):41-7,美国专利第6,027,998号;美国专利第6,605,451号,barany等,pct公开号wo97/31256;wenz等,pct公开号wo01/92579;day等,genomics,29(1):152-162(1995),ehrlich等,science252:1643-50(1991);innis等,pcrprotocols:aguidetomethodsandapplications,academicpress(1990);favis等,naturebiotechnology18:561-64(2000);和rabenau等,infection28:97-102(2000);belgrader,barany和lubin,developmentofamultiplexligationdetectionreactiondnatypingassay,sixthinternationalsymposiumonhumanidentification,1995(可从以下网址获得:promega.com/geneticidproc/ussymp6proc/blegrad.html);lcr试剂盒说明手册,cat.#200520,rev.#050002,stratagene,2002;barany,proc.natl.acad.sci.usa88:188-93(1991);bi和sambrook,nucl.acidsres.25:2924-2951(1997);zirvi等,nucl.acidres.27:e40i-viii(1999);dean等,procnatlacadsciusa99:5261-66(2002);barany和gelfand,gene109:1-11(1991);walker等,nucl.acidres.20:1691-96(1992);polstra等,bmcinf.dis.2:18-(2002);lage等,genomeres.2003年2月;13(2):294-307,和landegren等,science241:1077-80(1988),demidov,v.,expertrevmoldiagn.2002年11月;2(6):542-8.,cook等,jmicrobiolmethods..2003年5月;53(2):165-74,schweitzer等,curropinbiotechnol.2001年2月;12(1):21-7,美国专利第5,830,711号,美国专利第6,027,889号,美国专利第5,686,243号,公开的p.c.t.申请w00056927a3,和公开的p.c.t.申请wo9803673a1。
如本文所用,术语“mir-1983抑制剂”旨在指降低mir-1983的细胞内表达或活性的试剂。更详细地,该试剂直接作用于mir-1983或间接作用于mir-1983的前体,从而在转录水平上降低microrna-1983的表达,促进所表达的mir-1983的降解或破坏mir-1983的活性,从而降低了mir-1983的表达水平或活性。“mir-1983抑制剂”包括,例如抗mir-1983寡核苷酸及其修饰形式,其包含例如标记或标签,稳定部分和/或运输部分。
术语“改善胰岛素抵抗”通常是指在开始发展后改善胰岛素抵抗或病理状况的征兆或症状的治疗干预。“减轻”通常是指疾病征兆或症状的数量或严重程度的降低。
以上公开总体上描述了本申请。通过参考以下特定实施例,可以获得更完整的理解。仅出于说明的目的描述了这些实施例,而无意于限制本公开的范围。视情况提示或权宜之计,可以预期形式的改变和等同物的替换。尽管这里已经采用了特定术语,但是这些术语仅是描述性的,并非出于限制的目的。
组合物和方法
研究表明,mirna可以单独或与其他协同作用参与代谢性疾病(feng等,2016;hashimoto和tanaka,2016;vienberg等,2016)。某些涉及mirna调节介导下丘脑组织中的胰岛素敏感性。例如,ob/ob瘦素缺陷型小鼠下丘脑中的mir-200a的沉默足以通过增加瘦素受体和insr底物2(irs-2)表达水平来改善瘦素和胰岛素信号传导(crepin等,2014)。此外,dicer的下丘脑敲减是mirna生物发生的重要组分之一,可导致高食性肥胖,其在脑室内(icv)递送mir-103的模拟物后可能会逆转(vinnikov等,2014)。研究表明,中枢神经系统(cns)的mirna可能在调节全身能量稳态中起关键作用,而下丘脑可能表现出独特的特定代谢状态的mirna特征。这样的特征可以用作人类疾病治疗中的诊断工具或治疗靶标。考虑到胰岛素信号传导对代谢稳态的至关重要性,以及下丘脑神经元在实现这种平衡中的关键作用。本发明的发明人使用已建立的体外细胞来研究神经元是否在诱导胰岛素抵抗时表现出与hfd饲喂的小鼠一致的标志性的mirna表达谱。
发现mir-1983与胰岛素抵抗的诱导直接相关,并且下丘脑中mir-1983的升高与hfd小鼠血清中水平的升高一致,表明它可能是中枢胰岛素抵抗的生物标志物。另外,已经确定mir-1983也存在于超重和肥胖人类中,并且他们的血清mir-1983水平与循环胰岛素水平和胰岛素抵抗程度相关(胰岛素抵抗的稳态模型评估,homa-1r)。最后,确定了mir-1983抑制剂的鼻内转移可恢复小鼠体内的全身胰岛素信号传导和葡萄糖稳态。
下丘脑水平的胰岛素抵抗可以在葡萄糖稳态的失调之前发生。在本文中,在进食相关的下丘脑细胞系中,在体内胰岛素诱导的胰岛素抵抗的诱导过程中,micrornamir-1983显著变化,伴随着胰岛素受体(insr)蛋白的反向调节,这表明insr是mir-1983的下游靶标。人血清中mir-1983水平的检查显示与胰岛素抵抗性量度(包括homa-ir)呈正相关。鼻内暴露于mir-1983lna抑制剂恢复了hfd小鼠的周围胰岛素敏感性和空腹血糖水平,同时保持insr蛋白水平。总之,这些结果暗示mir-1983作为早期下丘脑胰岛素抵抗的可靶向预测生物标志物,具有阻止糖尿病进程的潜力。
第一方面是mir-1983抑制剂,其包含与ctcacctggagcatgttttct(seqidno:1)的至少一部分互补的抗mir-1983寡核苷酸,所述部分至少包含ctcacctggagcatgttttct(seqidno:1)的核苷酸2至8。在一个实施方式中,抗mir-1983寡核苷酸与seqidno:1的核苷酸1至21,1至20,1至19,1至18,1至17,1至16,1至15,1至14,1至13,1至12,1至11,1至10,1至9或1至8互补。在另一实施方式中,抗mir-1983寡核苷酸与seqidno:1的核苷酸2至21,2至20,2至19,2至18,2至17,2至16,2至15,2至14,2至13,2至12,2至11,2至10,2至9或2至8互补。
在一个实施方式中,抗mir-1983寡核苷酸与至少一部分互补,其中所述部分是或包含tcacctggagcatgt(seqidno:4)。在另一实施方式中,抗mir-1983寡核苷酸与至少一部分互补,其中所述部分是或者包含tcacctggagcatg(seqidno:5),tcacctggagcat(seqidno:6),tcacctggagca(seqidno:7),tcacctggagc(seqidno:8),tcacctggag(seqidno:9),tcacctgga(seqidno:10),tcacctgg(seqidno:11),或tcacctg(seqidno:12)。在另一实施方式中,抗mir-1983寡核苷酸与至少一部分互补,其中所述部分是或者包含ctcacctggagcatgt(seqidno:13),ctcacctggagcatg(seqidno:14),ctcacctggagcat(seqidno:15),ctcacctggagca(seqidno:16),ctcacctggagc(seqidno:17),ctcacctggag(seqidno:18),ctcacctgga(seqidno:19),ctcacctgg(seqidno:20),或ctcacctg(seqidno:21)。
在一个实施方式中,抗mir-1983寡核苷酸是或者包含seqidno:2的核苷酸9至15,8至15,7至15,6至15,5至15,4至15,3至15,2至15或1至15。抗mir1983寡核苷酸也可以更长,例如约或多达21个核苷酸,约或多达25个核苷酸,约或多达30个核苷酸,约或多达35个核苷酸,约或多达40个核苷酸,约或多达45个核苷酸,约或多达50个核苷酸,或者,更长,例如多达132个核苷酸,其包含例如dna和/或rna序列的稳定序列。
在另一实施方式中,抗mir-1983寡核苷酸包含rna单体。在一个实施方式中,抗mir-1983寡核苷酸包含dna单体、rna单体、lna单体或其组合。例如,抗mir-1983寡核苷酸包含仅dna单体、仅rna单体、dna和lna单体、rna和lna单体、或者仅lna单体。
在另一实施方式中,抗mir-1983寡核苷酸是化学修饰的。例如,抗mir-1983寡核苷酸可以包含一种或多种在核酸酶存在下增强稳定性的化学修饰。在另一实施方式中,抗mir-1983寡核苷酸是包含非化学修饰的和化学修饰的核苷酸的嵌合核酸。在另一实施方式中,通过将母体磷酸二酯键的主链修饰成硫代磷酸酯键也可以改善核酸酶抗性,其中硫原子取代磷酸基团中的一个非桥连氧原子,或者使用吗啉代低聚物,其中六元吗啉环取代糖部分。吗啉代例如是不带电荷的,并且固有地抗核酸酶降解。
在另一实施方式中,抗mir-1983寡核苷酸在2'位置包含化学修饰。例如,可以使用包括但不限于如专利us20130116419中所述的“点击化学”的化学修饰方法来合成在2'位置的化学修饰。化学修饰的核酸可以从不同的制造商处购买。在另一实施方式中,化学修饰可以是2'o甲基(2'o-me),2'-o-甲氧基乙基(2'o-moe),2'氟(2'f),锁核酸单体(lnam),炔基官能团或可用于靶向配体的其他官能团。
在另一实施方式中,抗mir-1983寡核苷酸包含化学修饰,其中,所述修饰选自2'o甲基(2'o-me),2'-o-甲氧基乙基(2'o-moe),2'氟(2'f)和锁核酸单体(lnam)。在另一实施方式中,抗mir-1983寡核苷酸包含一个或多个2’o-me化学修饰。在另一实施方式中,抗mir-1983寡核苷酸包含一个或多个2’o-moe化学修饰。在另一实施方式中,抗mir-1983寡核苷酸包含一个或多个(2'f)化学修饰。在另一实施方式中,抗mir-1983寡核苷酸包含一个或多个lnam化学修饰。在另一实施方式中,抗mir-1983寡核苷酸包含2’o-me、2’o-moe和2’f化学修饰的组合。在另一实施方式中,使用本领域已知的方法合成化学修饰的寡核苷酸。在另一实施方式中,修饰的寡核苷酸购自制造商。
在另一实施方式中,抗mir-1983寡核苷酸包含多个核苷酸单体的化学修饰。在一个实施方式中,抗mir-1983包含2’o-me、2’o-moe、2’f之一或其组合的多个化学修饰。在另一实施方式中,抗mir-1983仅包含多个lnam。在另一实施方式中,化学修饰的寡核苷酸购自制造商。
在另一实施方式中,抗mrna1983寡核苷酸包含多个锁核酸单体(lnam)。在一个实施方式中,寡核苷酸包含一个或多个lna,其是连续的或被一个或多个非lna核苷酸隔开。
在另一实施方式中,抗mir-1983寡核苷酸是锁核酸(lna)或lna/dna混聚体。在一个实施方式中,混聚体不包含超过5个连续的dna单体。在另一实施方式中,寡核苷酸包含lna和rna单体。在另一实施方式中,存在不超过5个连续单体。在另一实施方式中,寡核苷酸包含含有2’-o,4’-c-亚甲基核糖核苷(结构a)的锁核苷酸或lna,其中核糖糖部分处于“锁定”构象。在另一实施方式中,抗mir-1983寡核苷酸包含至少一个2′,4′-c桥连的2′脱氧核糖核苷(cdna,结构b)。参见,例如,美国专利第6,403,566号和wang等(1999)bioorganicandmedicinalchemistryletters,第9卷:1147-1150,将其全部内容通过引用以其整体并入本文。
在另一实施方式中,抗mir-1983寡核苷酸是锁核酸(lna)或lna/dna混聚体,其中,lna包含序列5’-3’,a*c*a*t/u*g*c*t/u*c*c*a*g*g*t/u*g*a(seqidno:2),或其部分,其中,所述部分至少是7个核苷酸长,并且包含seqidno:2的第9至15位核苷酸的核苷酸。例如,当抗mir-1983寡核苷酸为lna时,核苷酸间键(以星号表示)为硫代磷酸酯核苷酸间键。
锁核酸(lnatm)是一类高亲和力的rna类似物,其中核糖环被“锁定”在沃森-克里克结合的理想构象中。当与互补的dna或rna链杂交时,lnatm寡核苷酸显示出高的热稳定性。此外,lnatm寡核苷酸可以比传统的dna或rna寡核苷酸短,并且仍然保留高tm。lnatm寡核苷酸可以由lnatm和dna或rna的混合物组成。已显示将lnatm掺入寡核苷酸可提高许多基于杂交的技术(包括pcr、微阵列和原位杂交)的敏感性和特异性。
在另一实施方式中,抗mir-1983寡核苷酸仅包含dna和lna单体。
星号表示形成lna的主链的核苷酸间键,可选为磷酸二酯或优选为硫代磷酸酯核苷酸间键。
在另一实施方式中,抗mir-1983寡核苷酸是包含序列5’-3’,a*c*a*t*g*c*t*c*c*a*g*g*t*g*a(seqidno:2)的lna或其包含至少核苷酸9至15的一部分。在另一实施方式中,寡核苷酸仅包含dna和lna单体。
在另一实施方式中,抗mir-1983寡核苷酸是包含序列5’-3’,a*c*a*u*g*c*u*c*c*a*g*g*u*g*a(seqidno:2)的lna或其包含至少核苷酸9至15的一部分。在另一实施方式中,寡核苷酸包含一个、两个,三个或四个变体dna或rna单体a*c*a*t/u*g*c*t/u*c*c*a*g*g*t/u*g*a(seqidno:2)或其包含至少核苷酸9至15的一部分。
在另一实施方式中,抗mir-1983寡核苷酸至少一个错配和至多4个错配。错配是相对于mirna序列的,并且可以例如包含在抗mir寡核苷酸的中间,例如7个单体的中间。错配可以位于3、4、5或6位中的任一个。
在另一实施方式中,抗mir-1983寡核苷酸是双链的。例如,抗mir-1983寡核苷酸可以合成或构建为双链分子,然后变性。在另一实施方式中,将抗mir-1983寡核苷酸合成为单链。
在另一实施方式中,抗mir-1983寡核苷酸是单链的。
在另一实施方式中,抗mir-1983寡核苷酸与seqidno:2的核酸序列具有相同或高达60%、70%、80%或高达90%的序列同一性。在某些实施方式中,抗-mir-1983寡核苷酸与seqidno:2有1、2、3、4、5、6、7、8、9或10个核苷酸差异。在另一个实施方式中,抗mir-1983寡核苷酸具有与mir-1983相同,比mir-1983更长或更短的长度。在某些实施方式中,抗mir-1983寡核苷酸与seqidno:1的5'端的7或8个核苷酸互补。在其他实施方式中,抗mir-1983寡核苷酸的长度为约或至多7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49或50个核苷酸或至多132个核苷酸的任何数量的核苷酸,或其中可衍生的任何范围。在其他实施方式中,抗mir-1983寡核苷酸的长度为7至10、10至20、20至50、50至75、75至100或100至132个核苷酸或其中可衍生的任何范围。
在另一实施方式中,mir-1983抑制剂包含与抗mir-1983寡核苷酸偶联的标志物分子或标记分子,诸如可激发并以uv/vis或红外波长发射的荧光染料,例如fitc、tritc,texas红,cy染料,alexa染料(bioprobes)等。
在一个实施方式中,mir-1983抑制剂包含连接至抗mir-1983寡核苷酸的一个或两个末端的转运部分,以促进转运跨过细胞膜和血脑屏障。例如,短信号肽(也称为蛋白质转导结构域)以及具有c32官能团的亲脂性基团(如胆固醇结构域,月桂酸和邻苯二酚酸衍生物)已与寡核苷酸结合,以利于将其传递到细胞中。与核酸复合并改善细胞内传递的其他化合物包括脂质(诸如dotap(或其他阳离子脂质)、ddab、dhdeab和dope),基于非脂质的聚合物(诸如聚乙烯亚胺、聚酰胺胺)和这些和其他聚合物的枝状聚合物)。也可以使用组合。例如,在某些实施方式中,使用脂质的组合,如dotap和胆固醇或胆固醇衍生物(美国专利第6,770,291号,通过引用并入本文)。
在另一实施方式中,mir-1983抑制剂包含稳定性部分以增加抑制剂的稳定性。
另一方面是包含mir-1983抑制剂和可选地稀释剂的组合物。稀释剂可以是任何和所有溶剂、分散介质、涂料、表面活性剂、抗氧化剂、防腐剂(例如,抗细菌剂、抗真菌剂)、等渗剂、盐、防腐剂、粘合剂、润滑剂等材料及其组合,如本领域普通技术人员已知的(参见,例如,remington'spharmaceuticalsciences,第18版,mackprintingcompany编,1990,第1289-1329页,通过引用并入本文),并且其可以与核酸一起使用。除非任何常规载体与活性成分不相容,否则考虑将其用于药物组合物中。
在另一实施方式中,所述组合物稀释剂包含盐水。
在一个实施方式中,所述组合物配制为鼻内制剂。在一个实施方式中,鼻内制剂可以是粉末、水溶液、滴鼻剂和/或气雾剂的形式。
在另一实施方式中,用于鼻内施用的固体制剂可以包含赋形剂,诸如乳糖、淀粉或葡聚糖。例如,可以配制用于吸入器装置或吹入器中的明胶胶囊和药筒,其含有抗mir-1983寡核苷酸或组合物的粉末混合物和合适的粉末赋形剂(如乳糖、淀粉或葡聚糖)。在另一实施方式中,用于鼻内施用的液体制剂可以是以气雾剂、滴鼻剂和/或定量喷雾剂形式使用的水性或油性溶液。
可将气雾剂制剂置于加压的可接受的推进剂中,诸如二氯二氟甲烷、三氯氟甲烷、二氯四氟乙烷、二氧化碳、丙烷、氮气或其他合适的气体。例如,可以使用合适的推进剂将制剂以气雾剂的形式从加压包装、喷雾器或其他吸入装置中递送。
在某些实施方式中,用于通过鼻内施用的制剂包括但不限于脂质体制剂。
在某些实施方式中,使用吸入装置施用用于鼻内递送的药物组合物。术语“吸入装置”是指能够向受试者的鼻道施用mir-1983抑制剂的任何装置。吸入装置包括计量吸入器(mdi)、干粉吸入器(dpi)、喷雾器(包括射流喷雾器和超声波喷雾器)、热蒸发器、软雾吸入器、热气雾吸入器和基于电动动力的溶液雾化吸入器等设备。吸入装置还包括高效喷雾器。在某些实施方式中,喷雾器是喷射喷雾器、超声喷雾器、脉动膜喷雾器、包括具有多个孔的振动筛网或板的喷雾器、包含振动发生器和水室的喷雾器、或使用受控装置功能协助雾化水溶液向受试者的鼻腔吸气的雾化器。喷雾器,定量吸入器和软雾吸入器通过形成气雾剂(包括易于吸入的液滴大小)来递送药物。
在使用加压气雾剂的情况下,可以通过提供阀门来确定剂量单位,以递送计量数。例如,吸入装置可以是小的硬瓶,在该硬瓶上装有计量喷雾器。
mir-1983抑制剂也可以直接施用给大脑,例如通过直接注射(可选地通过插管)或脑室内(icv)施用。
可以使用其他递送方式,例如,其中mir-1983抑制剂包含转运部分。例如,施用途径的实例包括肠胃外的,例如静脉、皮下、口服和透粘膜。
另一方面是一种检测细胞是否为胰岛素抵抗的方法,其包括测量mir-1983的水平并与阈值或对照进行比较,其中与对照相比,水平的升高表明细胞是胰岛素抵抗的。在一个实施方式中,对照是具有正常胰岛素敏感性的小鼠细胞。在另一实施方式中,对照是具有正常胰岛素敏感性的人类细胞。
在另一实施方式中,细胞是神经元细胞,可选地,是下丘脑细胞。在另一实施方式中,细胞是下丘脑细胞。在另一实施方式中,细胞是表达神经肽y的下丘脑细胞。
另一方面是在受试者中检测糖尿病前期或发展糖尿病的可能性增加的方法,所述方法包括测量来自受试者的生物样品中的mir-1983的水平,其中,与阈值或对照相比升高的水平表示受试者发展糖尿病的可能性增加。在一个实施方式中,对照是具有正常胰岛素敏感性的小鼠细胞。在另一实施方式中,对照是具有正常胰岛素敏感性的人类细胞。
可以使用基于pcr的方法或一种或多种杂交探针检测细胞或样品中mirna的水平。在一个实施方式中,测量mir-1983的水平包括使用微阵列分析。在另一实施方式中,定量pcr测定法用于测量mir-1983的水平。在另一实施方式中,通过分离mirna,并将所述mirna与杂交探针(例如本文所述的反义分子)杂交来检测mirna的水平。
在另一实施方式中,测量mir-1983的水平包括:(a)用atp和聚(a)聚合酶使mirna聚腺苷酸化,以形成具有连续的a残基序列的聚腺苷酸化的mirna;(b)在反应混合物中逆转录聚腺苷酸化的mirna以形成cdna,该反应混合物包括(i)长度不超过40个核苷酸的第一引物,其与mirna的至少两个3'末端核苷酸和聚腺苷酸化的mirna的连续的a残基序列互补,以与其杂交并启动与聚腺苷酸化的mirna互补的cdna的合成,(ii)逆转录酶和(iii)所有四个脱氧核糖核苷三磷酸;(c)扩增反应混合物中包含cdna的dna分子,所述反应混合物包含(i)cdna,(ii)第一引物;(iii)第二引物,其与cdna的3'核苷酸充分互补以与其杂交并启动延伸产物的合成;(iv)dna聚合酶和(v)所有四种脱氧核糖核苷三磷酸;和(d)检测和/或定量扩增的dna分子,其中,扩增的dna的存在和/或数量对应于mirna的存在和/或数量。在另一实施方式中,使用balcells等,2018中描述的方法设计在测量mirna水平期间使用的引物。在另一实施方式中,检测和/或定量扩增的cdna分子包括利用实时rt-pcr。在另一实施方式中,检测和/或定量扩增的cdna包括利用凝胶电泳。
已检测到血清中的mirna水平。在一个实施方式中,生物样品血液、血浆或血清、尿液或者组织中的一种。
在一个实施方式中,生物样品自禁食至少8小时的受试者。在另一实施方式中,所述方法进一步包括测量胰岛素和/或葡萄糖水平。在某些实施方式中,还测量脂质诸如胆固醇和/或ldl。在某些实施方式中,所述方法进一步包括测量mir1983的标准化水平,可选地相对于内部对照(如snorna202)的水平标准化。
在一个实施方式中,受试者是bmi至少为25的受试者。在一个实施方式中,所述受试者是糖尿病前期的,例如不服用胰岛素或胰岛素增敏剂。
如实施例中所示,相对于预定标准或先前水平检测mir-1983或其升高的水平可预测胰岛素抵抗的发展。它是早期标志物,并在胰岛素抵抗发展之前。它提供了鉴定可能会出现胰岛素抵抗的受试者的机会,从而提供更早的干预。
在一个实施方式中,生物样品是血清样品。在另一实施方式中,使用标准的人类血液回收程序来回收血清样品。
在另一实施方式中,通过定量pcr测定法测量mir-1983的水平。在另一实施方式中,使用了除pcr方法以外的其他扩增程序,如除了别的之外,在美国专利第5,830,711号、美国专利第6,027,889号、美国专利第5,686,243号、公开的p.c.t.申请w00056927a3和公开的p.c.t.申请wo9803673a1中所述。
在另一实施方式中,定量pcr方法包括从获自细胞或生物样品的rna产生cdna,使用序列特异性引物扩增cdna并定量mir-1983的水平。
可以监测mir-1983水平升高的受试者的进一步变化,并通过饮食调整或适合于糖尿病前期或糖尿病的治疗进行治疗。具有增加的mir-1983水平的受试者可以例如用本文所述的抗mir抑制剂治疗。
另一方面是一种改善受试者的胰岛素敏感性的方法,所述方法包括向有需要的受试者施用mir-1983抑制剂或包含所述抑制剂的组合物。在一个实施方式中,有需要的受试者是患有胰岛素抵抗或糖尿病前期的动物。在另一实施方式中,有需要的受试者是患有胰岛素抵抗或糖尿病前期的人。在另一实施方式中,有需要的受试者已经被诊断患有糖尿病。可以施用本文所述的任何mir-1983抑制剂,包括其组合。
在一个实施方式中,当受试者表现出一种或多种胰岛素抵抗的迹象时,例如在通常开出任何其他抗糖尿病药物的处方之前,向受试者施用mir-1983抑制剂或包含所述抑制剂的组合物。在另一实施方式中,向受试者施用mir-1983抑制剂与用于胰岛素抵抗或糖尿病的另一种治疗的组合。
可以在施用mir-1983之前测量mir1983的水平,以确认其水平升高。因此,一方面,向mir-1983水平升高的受试者施用mir-1983抑制剂。
在一个实施方式中,改善胰岛素敏感性是指与施用治疗之前的基线水平相比,达到至少10%,至少20%,至少30%以上的胰岛素敏感性水平,例如通过腹膜内胰岛素耐受性测试(ipitt)测量的,可选地,相对于未诊断出胰岛素抵抗的受试者,其敏感性增加相似。
在另一实施方式中,以允许与神经元接触和/或靶向神经元的方式施用mir-1983抑制剂或包含所述抑制剂的组合物。在另一实施方式中,以允许接触下丘脑神经元和/或靶向下丘脑神经元的方式施用mir-1983抑制剂或包含所述抑制剂的组合物。例如,在某些实施方式中,mir-1983抑制剂或包含所述抑制剂的组合物经鼻内施用。
在一个实施方式中,施用的mir-1983抑制剂是抗mir-1983寡核苷酸。在另一实施方式中,施用的抗mir-1983寡核苷酸包含在包含一种或多种可选地具有稀释剂的组分的组合物中。在另一实施方式中,稀释剂是盐水或本文描述的其他稀释剂。
在另一实施方式中,包含所述抑制剂的mir-1983抑制剂或组合物鼻内、通过贴剂或静脉内施用。
mir-1983抑制剂可以与另一种治疗剂一起施用。特别是,mir-1983抑制剂可以与鼻内施用的另一种治疗剂一起施用。在一个实施方式中,另一种治疗剂是瘦素。
如本文所述,还提供了本文所述的mir-1983抑制剂和组合物在治疗有需要的受试者中的用途。
还提供了包含本文所述的mirna1983抑制剂的鼻腔递送装置。在一个实施方式中,所述装置是鼻喷雾泵或加压递送装置。
以上公开大体上描述了本申请。通过参考以下具体实施例,可以获得更完整的理解。仅出于说明的目的描述了这些实施例,而不意在限制本公开的范围。视情况提示或权宜之计,可以预期形式的改变和等同物的替换。尽管本文使用了特定术语,但是这些术语仅是描述性的,并不用于限制目的。
实施例
以下非限制性实施例是本公开的说明:
实施例1:在表达下丘脑npy的神经元细胞系中胰岛素抵抗的诱导,诱导mir-1983和mir-20a-3p水平的变化
表达和分泌npy和agrp的mhypoe-46细胞系因其对胰岛素的响应而得到了很好的表征,并且已经鉴定了在细胞水平上发展胰岛素抵抗的机制(mayer和belsham,2009,2010a)。特别令人感兴趣的是,胰岛素抵抗发展后,insr或insr底物1(irs-1)蛋白的损失并不涉及mrna转录水平上的任何变化。这一发现表明,可能存在另一层调节。据推测,这可能是通过mirna。mirna是小的rna分子,其可以隔离mrna的p体,从而阻止蛋白质翻译(bhattacharyya等,2006;liu等,2005)。为了检验这一假设,并在更大范围内确定mirna是否因胰岛素诱导的胰岛素抵抗状态而改变,将mhypoe-46细胞用载体或100nm胰岛素处理4小时和24小时,对总rna进行微阵列分析。选择这些时间点是为了区分那些在正常信号传导条件下(4小时后)对胰岛素响应的mirna与一旦建立胰岛素抵抗后的那些mirna(在24小时后)。微阵列数据显示,所有组中都有60个mirna发生了显著变化(图8)。然后对数据进行分类以确定仅在处理4小时至24小时的胰岛素处理组中发生变化的那些mirna(图9)。与载体相比,发现十二个mirna在24小时的时间点具有显著差异,并根据最大倍数变化程度进行排名。选择在24小时时间点增加的前四个mirna,以通过qpcr进行验证。为了确认通过微阵列分析鉴定的胰岛素抵抗的标志,将mhypoe-46细胞用载体或100nm胰岛素处理4小时和24小时,然后进行qpcr和western印迹以定量insrmrna和蛋白质(图10)。mirna的qpcr验证显示,与mir-296-5p(图1c)或let-7d-5p(图1d)不同,只有mir-1983(图1a)和mir-20a-3p(图1b)在使用100nm胰岛素处理后24小时时继续表现出上调。基于这些发现,然后选择了mir-1983和mir-20a-3p进行进一步的实验分析。该实施例的方法可以在实施例10中找到。
实施例2:在表达npy和pomc的细胞系中,在胰岛素抵抗期间mir-1983受到差异调节,并且在诱导胰岛素抵抗后在npy神经元中并未永久升高
为了确定mir-1983和mir-20a-3p的变化对于mhypoe-46(雄性)细胞系是否是独特的,成年来源的非克隆mhypoa-npy/gfp(雌性)和mhypoa-pomc/gfp-2(雄性)细胞系用100nm胰岛素或载体处理16小时,其足以引起胰岛素抵抗(wellhauser等,2016)(分别见图2a和2b)。发现在mhypoa-npy/gfp细胞系mir-1983中mir-1983的水平上调,这表明不管来源(胚胎还是成人)或性别(雄性还是雌性),响应于延长的胰岛素治疗的mir-1983的升高在npy细胞中消息是保守的。
根据arc中神经元群体之间mirna也可能差异性表达的发现(herzer等,2012),测试了暴露于胰岛素后,mhypoa-pomc/gfp-2细胞系的mir-1983是否也会出现相同的变化。与npy/agrp细胞模型不同,100nm胰岛素暴露16小时导致mir-1983水平降低。因此,尽管在给定的范例下npy/agrp和pomc模型都变得具有胰岛素抵抗性,但在这些神经元群体中特定的mirna可能有所不同。在这两种神经元细胞系中看到胰岛素的相反作用的观察结果是在这两个细胞群体中普遍观察到的响应于胰岛素的差异信号的特征(belgardt等,2009),并提出胰岛素抵抗对mir-1983的影响是表达npy的神经元所特有的可能性。在mhypoa-npy/gfp或mhypoa-pomc/gfp-2细胞中,mir-20a-3p均未发生变化,这表明在mhypoe-46细胞系之外,胰岛素对这一特定mirna的作用可能不是保守的。
一旦在mhypoe-46模型中将mir-1983定义为胰岛素抵抗的标志物,下一步就是确定胰岛素抵抗的发作是否引起mir-1983水平的短暂或永久性改变。qpcr和western印迹数据显示,在100nm胰岛素处理48小时后,尽管mir-1983恢复到正常水平,insrβ-亚基的水平仍然很低(图2c)。因此,在该细胞模型中,mir-1983的变化实际上是短暂的,在特定时间范围内响应胰岛素暴露而发生,提示mir-1983可能是胰岛素抵抗进展或发展的原因,而不仅仅是神经元不再保持感知胰岛素能力的最终状态的标志物。本实施例中使用的方法可以在实施例10中找到。
实施例3:mir-1983在体外靶向insr蛋白
已经建立了通过胰岛素调节mir-1983的时间表,阐明了最终的mrna靶标并鉴定了这些靶标如何可能促进mhypoe-46模型中抗性的发展。计算机模拟生物信息学分析中揭示,insr的3'非翻译区(utr)是mir-1983的可能靶标,并且所述mrna的这一种子序列区在包括人类在内的所有物种中也都是保守性的(图3a,3b)。注意的是,尽管mir-1983本身尚未在在线数据库资源中被认为在人类中是保守的,但已发现该mirna相比于正常皮肤,在银屑病人皮肤中存在并升高(edinger等,2014)。最近,已经描述了mir-1983独特地衍生自异亮氨酸(ile)预转移rna(trna)成为人类细胞中完全功能的mirna的过程(hasler等,2016)。
为了评估mir-1983是否可以与insr的3'utr中的预期位点结合,在存在mir-1983模拟物或阴性对照的情况下,对包含该区域的pmirglo载体进行了瞬时转染。萤光素酶检测结果表明,mir-1983模拟物在48小时后将产生的光单位降低了约15%(图1f)。
用mir-1983的模拟物转染mhypoe-46神经元导致24小时时mir-1983水平的增加。24小时后观察到的mir-1983的增加不影响胰岛素敏感性或insrmrna的水平(图3c)。尽管如此,发现仅mir-1983水平的上调足以使insr的蛋白水平在24小时后降低20%(图3d),在48小时后降低35%(图3e),而无论何时,insrmrna水平在24或48小时后均保持不变(图3d和e)。这表明mir-1983可能有助于胰岛素抵抗的发展过程。总的来说,这些数据与先前的观察结果相结合,表明mir-1983是由暴露于高水平胰岛素驱动的神经元中早期胰岛素抵抗的标志物。最终,下一个要解决的问题是在体内胰岛素抵抗下在下丘脑中是否可以调节mir-1983。因此,评估了能够在小鼠中诱导胰岛素抵抗的hfd方案对下丘脑mir-1983水平的影响。本实施例中使用的方法可以在实施例10中找到。
实施例4:在60%hfd的7-9周后,雄性c57bl/6小鼠表现出下丘脑mir-1983的增加
将雄性和雌性c57bl/6小鼠都放置于60%hfd7-9周,在此期间,两种性别都显著增加了体重并显示出升高的血糖水平(图4a和4b)。这些周围的变化在雄性和雌性中均伴随着npy和agrp的mrna水平的降低,以及雌性小鼠中pomc的升高(图4c和4d)。雄性c57bl/6小鼠在7周时表现出循环胰岛素水平升高,其表明胰岛素抵抗,并且其下丘脑中mir-1983随之升高(图4e和4f)。因此,在c57bl/6小鼠品系中,似乎仅在伴随高循环胰岛素水平的情况下暴露于60%hfd会导致下丘脑mir-1983水平升高。由于雌性小鼠在第7周的胰岛素仍未增加,因此未检测到mir-1983的变化。在任一性别中均未观察到insr的mrna水平发生变化(图4c和4d)。鉴于在雄性小鼠体内观察到了mir-1983的变化,因此下一个目标是确定这是否对一种小鼠品系而言是唯一的,并确定与该调控相关的时间表是否是保守的。实施例中使用的方法可以在实施例10中找到。
实施例5:在饲喂60%hfd的雄性cd-1小鼠的下丘脑中的观察到的mir-1983的升高取决于hfd暴露的时间长度
已知雄性cd-1小鼠在60%的hfd5-6周之间产生周围胰岛素抵抗(liu等,2016),与c57bl/6品系相比,其时间更短;因此在此时间点采集空腹血糖。发现5周的hfd增加下丘脑的空腹血糖(图5a)、循环胰岛素(图5b)和mir-1983水平(图5c),而不会引起下丘脑npy、agrp、pomc或insrmrna的变化(图5d)。当将hfd的时间延长至10周时,实验组之间在下丘脑中观察到的mir-1983水平不再存在差异(图5e)。然而,在10周后,与食物相比,在饲喂hfd的小鼠中,npy和agrp的mrna水平降低,而pomc升高(图5f)。再次,没有观察到insrmrna水平的变化(图5f)。这些数据表明在雄性cd-1小鼠中,mir-1983的上调先于神经肽水平的改变。在雄性cd-1小鼠中创建了mir-1983调控的时机后,然后研究了在饮食的5周时间内在雌性小鼠中的响应。实施例中使用的方法可以在实施例10中找到。
实施例6:cd-1雄性小鼠在其下丘脑和血清中表现出mir-1983增加,但是在60%hfd下5周后在肾脏中减少
在经过5周食物或60%hfd后,雄性(图6a)和雌性(图6b)cd-1小鼠均表现出的体重显著增加(图11a),而雌性和雌性的总体重增加约为60%hfd组(图6a和c)的两倍。雌性小鼠中空腹血糖升高,但雄性小鼠中却没有升高(图6d),并且雄性小鼠表现出空腹血胰岛素水平和mir-1983的升高(分别为图6e和6f)。对神经肽mrna水平的检查表明,雄性和雌性的npy降低(图6g),仅雌性中的insrmrna升高了7%(图6g和6h)。为了确定在雄性和雌性小鼠中高胰岛素血症是否均可引起下丘脑中mir-1983的增加,而与hfd无关,收集了来自雄性和雌性mkr小鼠的下丘脑。当与没有hfd暴露的cd-1小鼠相比时,这些小鼠显示了在其下丘脑中mir-1983明显升高,与循环血浆胰岛素的相当水平一致(图6e和f)。
在雄性小鼠的下丘脑水平发生变化后,评估了其他代谢相关组织(即肌肉和肝脏)中mir-1983的变化。此外,还对肾脏中的水平进行了分析,因为先前已在小鼠的该组织中发现了mir-1983,其受醛固酮调节(edinger等,2014)。在比目鱼肌中未观察到mir-1983的变化,在肾脏中减少(图6i),并且在肝脏中未检测到该mirna的水平。
在小鼠和人类的血浆和血清中可检测到mirna,并且可能是代谢疾病的推测生物标志物。由于先前已在饲喂hfd的小鼠的循环中检测到mir-1983,因此评估了雄性cd-1小鼠的血清水平。在饲喂hfd的小鼠血清中,mir-1983的水平增加了近三倍(图6i),并且在血清中的胰岛素水平与mir-1983之间检测到显著相关性(p=0.03,图11b)。
mirna不仅位于体内的各种组织中,而且还在小鼠和人类的血浆和血清中发现,由于它们被包裹在外泌体中或与蛋白质(如argonaute2,(ago2))结合,因此它们具有很高的稳定性(turchinovich等,2011;valadi等,2007)。由于先前的mirna研究还显示可能在循环中检测到mir-1983,因此评估了雄性cd-1小鼠血清中的mir-1983水平(hsieh等,2015)。在hfd5周后,发现雄性cd-1小鼠血清中的mir-1983水平增加了近三倍(图12b)。因此,下丘脑中mir-1983升高的时机与5周时间点循环水平的可检测到的增加相一致。
鉴于该发现,研究了mir-1983的血清水平是否可以与血液中的其他参数相关。已经发现,血清中的葡萄糖水平与血清中的mir-1983之间具有相关性,并且胰岛素水平与mir-1983之间具有显著的相关性。这导致我们评估了mir-1983是否也可以与人类患者的此类参数相关,如实施例7所述。
实施例7:mir-1983血清水平是人类胰岛素抵抗的生物标志物
本文显示小鼠中的血清mir-1983升高与下丘脑中的升高一致,表明血清水平可以用作下丘脑变化的替代物。
mir-1983已在人类中描述(edinger等,2014;hasler等,2016)。因此,决定在人类受试者的血清中测定该mirna。
mir-1983与循环胰岛素水平和homa-ir评分升高有关。
在体重指数(bmi)大于25(那些被归类为超重和肥胖)的所有人类受试者中检测到mir-1983(图11c)。如线性回归分析所显示的,mir-1983的血清水平与各种血液参数显著相关(图11d,表1),与增加的胰岛素和homa-ir评分最正相关。这些数据补充了在小鼠中的发现,并支持了mir-1983可以用作胰岛素抵抗的循环指标的发现。数据表明,可以检测到mir-1983并在发展胰岛素抵抗的不同阶段。实施例10中提供了所使用方法的详细信息。
表1:人类血清mir-1983相关性分析
实施例8:在饲喂60%hfd的cd-1雄性小鼠中mir-1983的抑制可改善胰岛素敏感性、空腹血糖并恢复下丘脑中的insr蛋白水平。
认识到mir-1983在雄性cd-1小鼠中响应60%hfd挑战而在体内多个位置发生变化后,着手使用将mir-1983抑制剂的直接鼻内暴露的方法,确定mir-1983水平升高特别在下丘脑中的影响,。实施例中使用的方法可以在实施例10中找到。
设计了一个实验范例来测试mir-1983在大脑中的特异性抑制是否会在暴露于60%hfd时改变全身胰岛素敏感性。两组雄性cd-1小鼠均饲喂60%的hfd,与给予体内lnamirna阴性对照的食物组相比,以单剂量在体内鼻内施用lnamirnamir-1983抑制剂或体内施用lnamirna阴性对照(图7a)在5周的实验中获得相同的体重(图7b和图7c)。为了将体内研究与通过mir-1983进行的insr调节的体外发现联系起来,在鼻内暴露于mir-1983抑制剂或阴性对照的一周后检查了小鼠的胰岛素敏感性。观察到在腹膜内(ip)胰岛素施用后10分钟和20分钟后,用抑制剂治疗的60%hfd饲喂小鼠与给予阴性对照的其对应小鼠相比,胰岛素敏感性显著改善(图7d)。鼻内治疗两周后对所有小鼠的空腹血糖进行分析后揭示,与hfd饲喂的阴性对照组相比,给予抑制剂的hfd饲喂组显示出显著更低的值,但相对于食物饲喂动物,仍然升高(图7e)。在5周的实验完成后,收集了小鼠的下丘脑,发现与阴性对照组相比,抑制剂治疗组中的mir-1983仍然降低,并且与食物对照相比,这两个组表现出agrp、pomc和insr的相似mrna水平,并且npymrna的下降相似(图7f)。如图7g所示,与对照动物相比,hfd的mir-1983抑制剂治疗的动物的下丘脑中insr蛋白的水平相似,而接受阴性对照的动物的insr水平显著降低,这与mir-1983的水平升高相对应。总体而言,这些发现表明,在接受mir-1983抑制剂的hfd饲喂的动物中观察到的胰岛素敏感性提高是由于下丘脑中insr蛋白信号传导的保留导致的。实施例中使用的方法可以在实施例10中找到。
实施例9:通过鼻内施用mir-1983抑制剂的使用对人中的mir-1983抑制作用
由exiqon(seqidno:2)产生的体内准备好的mir-1983lna抑制剂的鼻内递送,一般来说,研究人员已经通过立体定向插管法将这些抑制剂施用至大脑,并且注射或脑室内(icv)可能通过绕过血脑或csf脑屏障而在人类中选择性地靶向下丘脑(born等,2002;dhuria等,2010)。有证据表明,鼻内施用多种肽可以高渗透率地到达下丘脑,因此在本研究中,弓状核可被抗mirna靶向(meredith等,2015)。虽然与icv施用相比不是常用的,但鼻内施用方法已成功确定mir-206在调节小鼠脑中脑源性神经营养因子(bdnf)中的作用(lee等,2012)。结果表明,暴露于hfd两周后给予单剂量的mir-1983抑制剂足以提高胰岛素敏感性,如一周后通过腹膜内胰岛素耐受性测试(ipitt)测试所测量的,并且与接受对照lna的组相比,足以降低两周后的空腹血糖(图7d)。仅暴露于mir-1983抑制剂即可观察到胰岛素敏感性的显著改善。与未修饰的寡核苷酸相比,lnamir抑制剂具有更高的稳定性,这可以解释这些药物在体内的持久作用。抗mir-1983寡核苷酸的施用还阻止了雄性hfd-饲喂cd-1小鼠下丘脑中insr蛋白水平的降低(图7g),进一步支持涉及将insr作为此mirna下游靶标的体外数据。
实施例10:方法
细胞模型和培养条件
如前所述,将鼠类mhypoe-46(雄性)、mhypoa-npy/gfp(雌性)、mhypoa-pomc/gfp-2(雄性)细胞系在补充有5%胎牛血清(fbs;(gibco,thermofisherscientific,waltham,ma),和1%青霉素-链霉素(gibco)的5.5mm葡萄糖dulbecco改良的eagles培养基(dmem;sigma-aldrich,st.louismo,usa)中在37℃5%co2下培养(belsham等,2004;dhillon等,2011;mayer和belsham,2009,2010a;nazarians-armavil等,2014;wellhauser等,2016)。
动物模型
c57bl/6、cd-1和mkr品系的雄性和雌性小鼠在7周龄时从charlesriverlaboratories(montreal,qc,加拿大)购买,并在实验开始前适应了一周。将小鼠保持12:12的明暗时间表(上午7点至下午7点)养。所有动物实验均在多伦多大学动物委员会的批准下进行。从脂肪饮食(60%hfd)或对照(7%蔗糖匹配)饮食(分别为d12492和d12450j,researchdiets,inc,newbrunswick,nj,美国)饲喂cd-1和c57bl/6小鼠60%卡路里。用tekladdiet8664(envigo,madison,wi,美国)饲喂mkr小鼠。指示时,每周称重一次,以跟踪5周饮食暴露中的体重增加。在禁食4小时至16小时后(如图例所示),用异氟烷对所有动物进行安乐死,然后收集组织和血液,并储存在-80℃直至进行分析。使用肝素包被的注射器收集用于测定胰岛素水平的血液,然后将样品以6,000×g在4℃离心10分钟,其中血浆被分离并储存在-80℃。在麻醉下通过心脏穿刺取出血清样品,并在冰上凝结1小时,然后以3,000×g离心10分钟(hsieh等,2015),然后将样品也保存在-80℃。获得了来自最初cd-1组(雄性)和c57组(雄性和雌性)小鼠的下丘脑组织,并公布了这些小鼠的代谢参数的更多详细信息(liu等,2016;prentice等,2014)。
人类患者研究参与者
人体患者研究已获得大学健康网络研究伦理委员会的批准,并根据赫尔辛基宣言进行。招募了具有不同体重指数(bmi)和胰岛素敏感性的受试者。该研究总共包括24名受试者。根据bmi分组,总共有11位男性(m)和13位女性(f),小于25(m=2,f=4),介于25-29(m=6,f=3)和大于30(m=3,f=6)。在未包被的管中收集过夜禁食状态的血清,使其凝结,并在以3,000×g在4℃旋转15分钟。然后立即将血清冷冻在-20℃,然后移至-80℃以长期保存。用葡萄糖计测量循环葡萄糖。循环胰岛素的测量是通过多伦多总医院(tgh)核心实验室的abbottarchitecti2000系统上的elisa或根据制造商的指示(etobicoke,on,加拿大),采用来自emdmillipore的hi-14k|人体胰岛素特异性放射免疫分析法。tgh的核心实验室分析了所列的所有其他循环血液参数。研究中没有受试者服用胰岛素或胰岛素增敏剂。使用homa2模型确定胰岛素抵抗的稳态模型评估(homa-ir)评分(levy等,1998)。
在体内鼻内施用mir-1983抑制剂或对照
在适应期后,将小鼠置于对照饮食或60%hfd中两周。在两周的最后一天,小鼠体内施用定制的锁核酸(lna)mir-1983抑制剂(5’-3’,a*c*a*t*g*c*t*c*c*a*g*g*t*g*a)(seqidno:2)或阴性对照(5’-3’,a*c*g*t*c*t*a*t*a*c*g*c*c*c*a)(seqidno:3),其由exiqon(woburn,ma,美国)合成,并溶解在无菌的0.9%盐水中至1nmol/μl的浓度。如上所述,通过异氟烷将小鼠麻醉,置于胸骨位置,并以总计10μl体积鼻内施用总计10nmol的mir-1983抑制剂或阴性对照。为了使对小鼠呼吸的影响最小化,同时使治疗在鼻腔中的滞留最大化,在每对小鼠中每剂量施用2.5μl抑制剂或阴性对照,每滴之间间隔两分钟。在麻醉下将小鼠置于胸骨位置。然后从麻醉中恢复小鼠,并放回到干净的居住笼中,继续其饮食方案。处理后一周和两周后,对小鼠进行腹膜内胰岛素耐受性测试(ipitt)。在饮食方案上总共5周后,对小鼠实施安乐死(如上所述),并收集组织和血清并将其储存在-80℃。
腹膜内胰岛素耐受性测试(ipitt)
在胰岛素耐受性测试和血糖测量之前,将小鼠禁食4小时。将emla乳霜(2.5%利多卡因和2.5%丙胺卡因,aztrazeneca)涂在其尾部的末端,并在从其尾部剪断≤1mm之前生效,以收集液滴。通过挤压尾部,并向插入血糖仪(bayercontourusb)的葡萄糖试纸上滴一滴血,获得基础血糖样品。然后给小鼠注射1国际单位(iu)的胰岛素/kg体重(humulinr,eli-lily)。此外,在随后的时间点(10、20、30、60和120分钟)进行血糖测量。胰岛素通过将humulin原液稀释在0.9%盐水中的过滤无菌的(0.22μm)0.1%无脂肪酸无牛血清白蛋白(bsa)中制备。对于所有实验,在大约一天的同一时间开始对小鼠的空腹血糖测量和胰岛素耐受性测试。
微阵列实验:分析与验证
将mhypoe-46细胞系用无菌1×磷酸盐缓冲盐水(pbs,载体对照)或100nm胰岛素(novorapidinsulin,novonordisk,mississauga,on,加拿大)处理4和24小时,之后,根据制造商的说明,用mirvanamirna分离试剂盒(ambion,thermofisherscientific,waltham,ma)收获并分离mirna。还使用100nm胰岛素对mhypoa-npy/gfp和mhypoa-pomc/gfp-2细胞系进行了16小时的处理,以检查mir-1983的表达,根据先前的工作这一时间点足以在这些细胞系中引起胰岛素抵抗。提交给在将以基于包含620个探针的mirbasev15对小鼠mirna和与小鼠相关的病毒mirna的nanostringncounter分析系统进行的mirna分析princessmargaret基因组学中心之前,在nanodrop2000c分光光度计(thermofisherscientific)上评估了总rna的数量和质量。在microarray中心使用nanostringtechnologies(seattle,wa)的nsolver分析软件nanostringv1.0进行了数据质量和标准化的初步评估。使用几何均值方法将代码计数标准化为阳性对照,以计算标准化因子。接下来,使用排名前100位的基因和几何均值方法对codeset内容进行标准化,以计算标准化因子。标准化的代码计数转换为log2值以进行统计分析。收到微阵列结果后,重复在mhypoe-46细胞中进行的实验,并使用purelinkminirna分离柱(ambion,thermofisherscientific,waltham,ma)进行rna分离。按照制造商(thermofisherscientific)的说明,使用柱上purelinkdnase在rna分离过程中进行dna酶处理。定量rna,并在nanodrop2000c分光光度计(thermofisherscientific)上评估纯度。
mirna靶标鉴定
使用mirwalk2.0软件鉴定mirna靶标(dweep和gretz,2015;dweep等,2011)。选择了在所有四种算法(mirwalk,miranda(betel等,2010;betel等,2008),rna22(miranda等,2006)和targetscan(lewis等,2005))中出现的潜在靶标,使用pantherdb.org将其分类为感兴趣的途径(mi等,2013;mi等,2016)。选择被发现参与胰岛素信号传导途径的mrna靶标用于进一步研究。根据公认的文献(betel等,2010),证实mirna靶标具有良好的mirsvr得分,并且使用targetscan小鼠鉴定了人和小鼠mrna序列之间相似的靶标位点(agarwal等,2015;lewis等,2005)。
逆转录(rt),定量pcr(qpcr)和用于mrna分析的引物设计
对于mrna分析,按照制造商的说明,通过abi高容量cdna库试剂盒使用500ng至1μg的总rna用于第一链cdna合成。使用
表2:定量pcr,克隆和测序引物序列
用于mirna分析的rt-pcr
对于mirna分析,使用用于mir-1983(id:121204_mat),mir-20a-3p(id:002491),mir-296-5p(id:000527)和let-7d-5p(id:002283)的rt和qpcr反应的
从小鼠血清rt-qpcr中分离mirna
使用
表3:小鼠mir-1983和人类mir-1983的核苷酸序列
发现人类mir-1983位于6号染色体上,小鼠mir-1983位于13号染色体上。恒河猴具有相同的mirna-1983序列(位于4号染色体上)。
mihypoe-46细胞中的mirna-1983模拟物转染
mhypoe-46细胞在100mm组织培养皿中培养至70%至75%的汇合度,根据制造商的说明,使用5μg/ml的dharmafect3(dharmacon,ge,location)将25nm的mirna-1983模拟物或阴性对照(thermofisherscientific)转染到mhypoe-46细胞中24小时或48小时,然后将细胞用1×pbs洗涤,并置于无血清的5.5mmdmem(sigma-aldrich,st.louismo,美国)中1小时,然后用10nm胰岛素再激发15分钟。用冰冷的1×pbs洗涤细胞,然后根据制造商的说明,使用
将insr3’utr克隆到pmirglo中,并在mhypoe-46细胞中共转染构建体和mirna-1983模拟物。
使用设计成添加与pmirglo载体(promegacorporation,madisonwi,美国)的多克隆位点互补的限制性酶切位点的引物从小鼠基因组dna(基因组dna纯化试剂盒,thermofisherscientific)扩增insr基因的3'utr。限制性消化载体和3’utr后,将两者连接并转化到hb101感受态细胞中。选择具有氨苄青霉素抗性的细菌菌落,使其生长以分离dna,以确认插入物以正确的方向存在。限制性内切酶和缓冲液购自cellsignaltechnologytechnologyinc.(danvers,ma,美国)。含有3’utrinsr插入片段的载体dna被送至应用基因组学中心(tcag,toronto,on,加拿大)进行测序(引物参见表2)。对于转染,将mhypoe-46细胞在6孔板(sarstedt,montreal,qc,加拿大)中培养至70%至75%的汇合度。然后根据制造商的说明书,每孔使用6μlturbofect转染25nmmirna-1983模拟物或阴性对照(如上所用)和5μgpmirglo-empty或3'utrinsr48小时。用冰冷的1×pbs洗涤细胞一次,然后根据制造商的说明在每孔450μl被动裂解缓冲液(biotiuminc.,fremont,ca,美国)中裂解细胞。
蛋白质分离、sds-page和western印迹
如先前所述(nazarians-armavil等,2014)或根据制造商的说明使用
棕榈酸酯处理mhypoe-46细胞系
用无菌水或50μm棕榈酸钠(sigma-aldrich)处理mhypoe-46细胞系24小时。通过将粉末和无菌水混合并将混合物加热至65℃直至完全溶解来制备棕榈酸钠溶液。然后将溶液直接添加到温热的(37℃)细胞培养基中,并立即处理细胞。如上所述完成了mir-1983水平的评估。
统计分析
使用graphpadprism6.0c版分析所有数据。如上所述,进行了学生t检验,单因素或双因素方差分析,然后进行bonferroni或fisher的lsd事后分析。在发现p值为p<0.05的情况下,认为数据具有统计学意义。所有数据以±sem表示。
实施例11
潜在应用的重要考虑因素是它们是否同时适用于两种性别。暴露于hfd后,c57bl/6雌性小鼠的神经肽水平呈现出相似的模式,其方向和大小与饲喂35%hfd12周的雌性c56bl/6小鼠观察到的相似(lee等,2010)。在研究中,5周后,与其雄性对应相比,cd-1雌性小鼠的npy表现出相同的变化,其与暴露于hfd7-9周后,雌性c56bl/6小鼠中观察到的结果相匹配。总体而言,在这些研究中观察到的npy和pomcmrna水平的代偿性变化与先前报道的相似(bergen等,1999)。虽然观察到在胰岛素暴露后的成年雌性mhypoa-npy/gfp细胞模型中mir-1983的明显上调,但在饲喂hfd后体内未观察到相同的变化。这一发现支持以下观点:如果雌性下丘脑神经元长时间暴露于较高的胰岛素水平,则mir-1983可能上调,但由于两种品系的雌性小鼠在所分析的时间内均未发生高胰岛素血症,因此没有观察到mir-1983水平的变化。尽管食用hfd和其他生理变化可能会改变mir-1983,但事实并非如此。
进一步支持以下观点:改变下丘脑mir-1983需要体内循环胰岛素的变化,发现用最富含含有hfd的饱和脂肪酸(50μm棕榈酸酯)处理mhypoe-46细胞24小时会导致mir-1983水平显著下降,其与用胰岛素观察到的相反。这一发现也与先前使用类似的npy/agrp神经元细胞模型的数据相吻合,与胰岛素相比,确定棕榈酸酯是通过其他机制诱导胰岛素抵抗的(mayer和belsham,2010b)。这可能与肥胖和与hfd相关的t2dm以及消瘦和超重人群中疾病的与肥胖无关的、高胰岛素血症发展有关。
c57bl/6和cd-1背景的雌性小鼠食用hfd导致体重明显增加和空腹血糖水平升高。尽管通常公认的是,雌性小鼠直到接受hfd饮食更长的时间后才会在外周产生胰岛素抵抗(medrikova等,2012),但令人惊讶的是,观察到主要神经肽npy、agrp和pomc的mrna水平出现了这种平行变化,像研究时间范围内与雄性小鼠一样。这一发现暗示着,尽管大多数在研究中使用hfd的研究都将雌性小鼠排除在外,因为它们没有像雄性那样早期显示出预期的周围变化,但实际上可能在大脑水平上神经肽含量发生了重大变化,因此在代谢研究中应与雄性对应进行平行检查。
mir-1983的变化出现在cd-1雄性小鼠中神经肽水平失调之前,而当小鼠置于hfd10周后消失。该体内数据与mir-1983在24小时后升高但不在48小时后升高的体外观察结果一致。先前的一项研究在置于58%hfd8周后接受恢复性低脂饮食时,在c57bl/6小鼠中检测到在诸多其他mirna中,mir-1983下降了近3倍,但没有观察到它们的hfd组在12周后mir-1983的表达增加(hsieh等,2015)。先前还显示,在饲喂45%hfd2周的大鼠中,时间对下丘脑中mirna表达的影响,然后在一个月和三个月后对mirna的水平进行检查,发现了对所检查的每种mirna的差异作用(sangiao-alvarellos等,2014)。
参考文献
agarwal,v.,bell,g.w.,nam,j.w.,和bartel,d.p.(2015).predictingeffectivemicrornatargetsitesinmammalianmrnas.elife4.
arroyo,j.d.,chevillet,j.r.,kroh,e.m.,ruf,i.k.,pritchard,c.c.,gibson,d.f.,mitchell,p.s.,bennett,c.f.,pogosova-agadjanyan,e.l.,stirewalt,d.l.等.(2011).argonaute2complexescarryapopulationofcirculatingmicrornasindependentofvesiclesinhumanplasma.proceedingsofthenationalacademyofsciencesoftheunitedstatesofamerica108,5003-5008.
balcells,ingrid,susannacirera,和peterkbusk.“specificandsensitivequantitativert-pcrofmirnaswithdnaprimers.”bmcbiotechnology11(2011):70.pmc.web.2018年7月24日.
belgardt,b.f.,okamura,t.,和bruning,j.c.(2009).hormoneandglucosesignallinginpomcandagrpneurons.thejournalofphysiology587,5305-5314.
belsham,d.d.,cai,f.,cui,h.,smukler,s.r.,salapatek,a.m.,和shkreta,l.(2004).generationofaphenotypicarrayofhypothalamicneuronalcellmodelstostudycomplexneuroendocrinedisorders.endocrinology145,393-400.
bergen,h.t.,mizuno,t.,taylor,j.,和mobbs,c.v.(1999).resistancetodiet-inducedobesityisassociatedwithincreasedproopiomelanocortinmrnaanddecreasedneuropeptideymrnainthehypothalamus.brainresearch851,198-203.
betel,d.,koppal,a.,agius,p.,sander,c.,和leslie,c.(2010).comprehensivemodelingofmicrornatargetspredictsfunctionalnon-conservedandnon-canonicalsites.genomebiology11,r90.
betel,d.,wilson,m.,gabow,a.,marks,d.s.,和sander,c.(2008).themicrorna.orgresource:targetsandexpression.nucleicacidsresearch36,d149-153.
bhattacharyya,s.n.,habermacher,r.,martine,u.,closs,e.i.,和filipowicz,w.(2006).reliefofmicrorna-mediatedtranslationalrepressioninhumancellssubjectedtostress.cell125,1111-1124.
born,j.,lange,t.,kern,w.,mcgregor,g.p.,bickel,u.,和fehm,h.l.(2002).sniffingneuropeptides:atransnasalapproachtothehumanbrain.natureneuroscience5,514-516.bruning,j.c.,gautam,d.,burks,d.j.,gillette,j.,schubert,m.,orban,p.c.,klein,r.,krone,w.,muller-wieland,d.,和kahn,c.r.(2000).roleofbraininsulinreceptorincontrolofbodyweightandreproduction.science289,2122-2125.
clegg,d.j.,gotoh,k.,kemp,c.,wortman,m.d.,benoit,s.c.,brown,l.m.,d'alessio,d.,tso,p.,seeley,r.j.,和woods,s.c.(2011).consumptionofahigh-fatdietinducescentralinsulinresistanceindependentofadiposity.physiology&behavior103,10-16.
crepin,d.,benomar,y.,riffault,l.,amine,h.,gertler,a.,和taouis,m.(2014).theover-expressionofmir-200ainthehypothalamusofob/obmiceislinkedtoleptinandinsulinsignalingimpairment.molecularandcellularendocrinology384,1-11.
dhillon,s.s.,mcfadden,s.a.,chalmers,j.a.,centeno,m.l.,kim,g.l.,和belsham,d.d.(2011).cellularleptinresistanceimpairstheleptin-mediatedsuppressionofneuropeptideysecretioninhypothalamicneurons.endocrinology152,4138-4147.
dhuria,s.v.,hanson,l.r.,和frey,w.h.,2nd(2010).intranasaldeliverytothecentralnervoussystem:mechanismsandexperimentalconsiderations.journalofpharmaceuticalsciences99,1654-1673.
dweep,h.,和gretz,n.(2015).mirwalk2.0:acomprehensiveatlasofmicrorna-targetinteractions.naturemethods12,697.
dweep,h.,sticht,c.,pandey,p.,和gretz,n.(2011).mirwalk--database:predictionofpossiblemirnabindingsitesby"walking"thegenesofthreegenomes.journalofbiomedicalinformatics44,839-847.
edinger,r.s.,coronnello,c.,bodnar,a.j.,labarca,m.,bhalla,v.,laframboise,w.a.,benos,p.v.,ho,j.,johnson,j.p.,和butterworth,m.b.(2014).aldosteroneregulatesmicrornasinthecorticalcollectingducttoaltersodiumtransport.journaloftheamericansocietyofnephrology:jasn25,2445-2457.
feng,j.,xing,w.,和xie,l.(2016).regulatoryrolesofmicrornasindiabetes.internationaljournalofmolecularsciences17.
gao,q.,和horvath,t.l.(2008).neuronalcontrolofenergyhomeostasis.febsletters582,132-141.
hashimoto,n.,和tanaka,t.(2016).roleofmirnasinthepathogenesisandsusceptibilityofdiabetesmellitus.journalofhumangenetics.
hasler,d.,lehmann,g.,murakawa,y.,klironomos,f.,jakob,l.,grasser,f.a.,rajewsky,n.,landthaler,m.,和meister,g.(2016).thelupusautoantigenlapreventsmis-channelingoftrnafragmentsintothehumanmicrornapathway.molecularcell63,110124.
herzer,s.,silahtaroglu,a.,和meister,b.(2012).lockednucleicacid-basedinsituhybridisationrevealsmir-7aasahypothalamus-enrichedmicrornawithadistinctexpressionpattern.journalofneuroendocrinology24,1492-1504.
hollander,j.a.,im,h.i.,amelio,a.l.,kocerha,j.,bali,p.,lu,q.,willoughby,d.,wahlestedt,c.,conkright,m.d.,和kenny,p.j.(2010).striatalmicrornacontrolscocaineintakethroughcrebsignalling.nature466,197-202.
hsieh,c.h.,rau,c.s.,wu,s.c.,yang,j.c.,wu,y.c.,lu,t.h.,tzeng,s.l.,wu,c.j.,和lin,c.w.(2015).weight-reductionthroughalow-fatdietcausesdifferentialexpressionofcirculatingmicrornasinobesec57bl/6mice.bmcgenomics16,699.
janssen,h.l.,reesink,h.w.,lawitz,e.j.,zeuzem,s.,rodriguez-torres,m.,patel,k.,vandermeer,a.j.,patick,a.k.,chen,a.,zhou,y.等.(2013).treatmentofhcvinfectionbytargetingmicrorna.thenewenglandjournalofmedicine368,1685-1694.
kaushik,s.,arias,e.,kwon,h.,lopez,n.m.,athonvarangkul,d.,sahu,s.,schwartz,g.j.,pessin,j.e.,和singh,r.(2012).lossofautophagyinhypothalamicpomcneuronsimpairslipolysis.emboreports13,258-265.
kim,j.m.,lee,s.t.,chu,k.,jung,k.h.,kim,j.h.,yu,j.s.,kim,s.,kim,s.h.,park,d.k.,moon,j.等.(2014).inhibitionoflet7cmicrornaisneuroprotectiveinaratintracerebralhemorrhagemodel.plosone9,e97946.
kleinridders,a.,ferris,h.a.,cai,w.,和kahn,c.r.(2014).insulinactioninbrainregulatessystemicmetabolismandbrainfunction.diabetes63,2232-2243.
konner,a.c.,和bruning,j.c.(2012).selectiveinsulinandleptinresistanceinmetabolicdisorders.cellmetabolism16,144-152.
kornfeld,j.w.,baitzel,c.,konner,a.c.,nicholls,h.t.,vogt,m.c.,herrmanns,k.,scheja,l.,haumaitre,c.,wolf,a.m.,knippschild,u.等.(2013).obesity-inducedoverexpressionofmir-802impairsglucosemetabolismthroughsilencingofhnf1b.nature494,111-115.
kuehnert,j等.novelrnachaperonedomainofrna-bindingproteinlaisregulatedbyaktphosphorylation.nucleicacidsresearch43,581-594,doi:10.1093/nar/gku1309(2015).
lee,a.k.,mojtahed-jaberi,m.,kyriakou,t.,astarloa,e.a.,arno,m.,marshall,n.j.,brain,s.d.,和o'dell,s.d.(2010).effectofhigh-fatfeedingonexpressionofgenescontrollingavailabilityofdopamineinmousehypothalamus.nutrition26,411-422.
lee,s.t.,chu,k.,jung,k.h.,kim,j.h.,huh,j.y.,yoon,h.,park,d.k.,lim,j.y.,kim,j.m.,jeon,d.等.(2012).mir-206regulatesbrain-derivedneurotrophicfactorinalzheimerdiseasemodel.annalsofneurology72,269-277.
levy,j.c.,matthews,d.r.&hermans,m.p.correcthomeostasismodelassessment(homa)evaluationusesthecomputerprogram.diabetescare21,2191-2192(1998).
lewis,b.p.,burge,c.b.,和bartel,d.p.(2005).conservedseedpairing,oftenflankedbyadenosines,indicatesthatthousandsofhumangenesaremicrornatargets.cell120,1520.
li,y.,和kowdley,k.v.(2012).methodformicrornaisolationfromclinicalserumsamples.analyticalbiochemistry431,69-75.
liu,j.,valencia-sanchez,m.a.,hannon,g.j.,和parker,r.(2005).microrna-dependentlocalizationoftargetedmrnastomammalianp-bodies.naturecellbiology7,719-723.
liu,y.,prentice,k.j.,eversley,j.a.,hu,c.,batchuluun,b.,leavey,k.,hansen,j.b.,wei,d.w.,cox,b.,dai,f.f.等.(2016).rapidelevationincmpfmayactasatippingpointindiabetesdevelopment.cellreports14,2889-2900.
livak,k.j.,和schmittgen,t.d.(2001).analysisofrelativegeneexpressiondatausingreal-timequantitativepcrandthe2(-deltadeltac(t))method.methods25,402-408.mayer,c.m.,和belsham,d.d.(2009).insulindirectlyregulatesnpyandagrpgeneexpressionviathemapkmek/erksignaltransductionpathwayinmhypoe-46hypothalamicneurons.molecularandcellularendocrinology307,99-108.
loh,k等.insulincontrolsfoodintakeandenergybalancevianpyneurons.molecularmetabolism6,574-584,doi:10.1016/j.molmet.2017.03.013(2017).
mayer,c.m.,和belsham,d.d.(2010a).centralinsulinsignalingisattenuatedbylong-terminsulinexposureviainsulinreceptorsubstrate-1serinephosphorylation,proteasomaldegradation,和lysosomalinsulinreceptordegradation.endocrinology151,75-84.
mayer,c.m.,和belsham,d.d.(2010b).palmitateattenuatesinsulinsignalingandinducesendoplasmicreticulumstressandapoptosisinhypothalamicneurons:rescueofresistanceandapoptosisthroughadenosine5'monophosphate-activatedproteinkinaseactivation.endocrinology151,576-585.
medrikova,d.,jilkova,z.m.,bardova,k.,janovska,p.,rossmeisl,m.,和kopecky,j.(2012).sexdifferencesduringthecourseofdiet-inducedobesityinmice:adiposetissueexpandabilityandglycemiccontrol.internationaljournalofobesity36,262-272.
meister,b.,herzer,s.,和silahtaroglu,a.(2013).micrornasinthehypothalamus.neuroendocrinology98,243-253.
meredith,m.e.,salameh,t.s.,和banks,w.a.(2015).intranasaldeliveryofproteinsandpeptidesinthetreatmentofneurodegenerativediseases.aapsj17,780-787.
mi,h.,muruganujan,a.,casagrande,j.t.,和thomas,p.d.(2013).large-scalegenefunctionanalysiswiththepantherclassificationsystem.natureprotocols8,1551-1566.mi,h.,poudel,s.,muruganujan,a.,casagrande,j.t.,和thomas,p.d.(2016).panther第10版:expandedproteinfamiliesandfunctions,andanalysistools.nucleicacidsresearch44,d336-342.
miranda,k.c.,huynh,t.,tay,y.,ang,y.s.,tam,w.l.,thomson,a.m.,lim,b.,和rigoutsos,i.(2006).apattern-basedmethodfortheidentificationofmicrornabindingsitesandtheircorrespondingheteroduplexes.cell126,1203-1217.
nazarians-armavil,a.,chalmers,j.a.,lee,c.b.,ye,w.,和belsham,d.d.(2014).cellularinsulinresistancedisruptshypothalamicmhypoa-pomc/gfpneuronalsignalingpathways.thejournalofendocrinology220,13-24.
obici,s.,feng,z.,karkanias,g.,baskin,d.g.,和rossetti,l.(2002a).decreasinghypothalamicinsulinreceptorscauseshyperphagiaandinsulinresistanceinrats.natureneuroscience5,566-572.
obici,s.,zhang,b.b.,karkanias,g.,和rossetti,l.(2002b).hypothalamicinsulinsignalingisrequiredforinhibitionofglucoseproduction.naturemedicine8,1376-1382.
parrizas,m.,和novials,a.(2016).circulatingmicrornasasbiomarkersformetabolicdisease.bestpractice&research.clinicalendocrinology&metabolism30,591-601.
prentice,k.j.,luu,l.,allister,e.m.,liu,y.,jun,l.s.,sloop,k.w.,hardy,a.b.,wei,l.,jia,w.,fantus,i.g.等.(2014).thefuranfattyacidmetabolitecmpfiselevatedindiabetesandinducesbetacelldysfunction.cellmetabolism19,653-666.
rizza,r.a.,mandarino,l.j.,genest,j.,baker,b.a.,和gerich,j.e.(1985).productionofinsulinresistancebyhyperinsulinaemiainman.diabetologia28,70-75.
roh,e.,songdo,k.,和kim,m.s.(2016).emergingroleofthebraininthehomeostaticregulationofenergyandglucosemetabolism.experimental&molecularmedicine48,e216.
sangiao-alvarellos,s.,pena-bello,l.,manfredi-lozano,m.,tena-sempere,m.,和cordido,f.(2014).perturbationofhypothalamicmicrornaexpressionpatternsinmaleratsaftermetabolicdistress:impactofobesityandconditionsofnegativeenergybalance.endocrinology155,1838-1850.
scherer,t.,o'hare,j.,diggs-andrews,k.,schweiger,m.,cheng,b.,lindtner,c.,zielinski,e.,vempati,p.,su,k.,dighe,s.等.(2011).braininsulincontrolsadiposetissuelipolysisandlipogenesis.cellmetabolism13,183-194.
seeger,t.,fischer,a.,muhly-reinholz,m.,zeiher,a.m.,和dimmeler,s.(2014).longterminhibitionofmir-21leadstoreductionofobesityindb/dbmice.obesity22,2352-2360.
thaler,j.p.,yi,c.x.,schur,e.a.,guyenet,s.j.,hwang,b.h.,dietrich,m.o.,zhao,x.,sarruf,d.a.,izgur,v.,maravilla,k.r.等.(2012).obesityisassociatedwithhypothalamicinjuryinrodentsandhumans.thejournalofclinicalinvestigation122,153-162.
trajkovski,m.,hausser,j.,soutschek,j.,bhat,b.,akin,a.,zavolan,m.,heim,m.h.,和stoffel,m.(2011).micrornas103and107regulateinsulinsensitivity.nature474,649-653.
turchinovich,a.,weiz,l.,langheinz,a.,和burwinkel,b.(2011).characterizationofextracellularcirculatingmicrorna.nucleicacidsresearch39,7223-7233.
valadi,h.,ekstrom,k.,bossios,a.,sjostrand,m.,lee,j.j.,和lotvall,j.o.(2007).exosome-mediatedtransferofmrnasandmicrornasisanovelmechanismofgeneticexchangebetweencells.naturecellbiology9,654-659.
varela,l.,和horvath,t.l.(2012).leptinandinsulinpathwaysinpomcandagrpneuronsthatmodulateenergybalanceandglucosehomeostasis.emboreports13,1079-1086.
vienberg,s.,geiger,j.,madsen,s.,和dalgaard,l.t.(2016).micrornasinmetabolism.actaphysiologica.
vinnikov,i.a.,hajdukiewicz,k.,reymann,j.,beneke,j.,czajkowski,r.,roth,l.c.,novak,m.,roller,a.,dorner,n.,starkuviene,v.等.(2014).hypothalamicmir-103protectsfromhyperphagicobesityinmice.thejournalofneuroscience:theofficialjournalofthesocietyforneuroscience34,10659-10674.
wellhauser,l.,chalmers,j.a.,和belsham,d.d.(2016).nitricoxideexertsbasalandinsulin-dependentanorexigenicactionsinpomchypothalamicneurons.molecularendocrinology30,402-416.
序列表
<110>多伦多大学理事会
<120>使用mirna抑制剂治疗胰岛素抵抗的组合物和方法
<130>2223-p53603pc00
<150>62/550,233
<151>2017-08-25
<150>62/697,935
<151>2018-07-13
<160>37
<170>patentinversion3.5
<210>1
<211>21
<212>dna
<213>智人(homosapiens)
<220>
<221>misc_feature
<222>(2)..(2)
<223>n是t或u
<220>
<221>misc_feature
<222>(7)..(7)
<223>n是t或u
<220>
<221>misc_feature
<222>(14)..(14)
<223>n是t或u
<220>
<221>misc_feature
<222>(16)..(19)
<223>n是t或u
<220>
<221>misc_feature
<222>(21)..(21)
<223>n是t或u
<400>1
cncaccnggagcangnnnncn21
<210>2
<211>15
<212>dna
<213>人工序列(artificialsequence)
<220>
<223>合成的
<220>
<221>misc_feature
<222>(4)..(4)
<223>n是t或u
<220>
<221>misc_feature
<222>(7)..(7)
<223>n是t或u
<220>
<221>misc_feature
<222>(13)..(13)
<223>n是t或u
<400>2
acangcnccaggnga15
<210>3
<211>15
<212>dna
<213>人工序列(artificialsequence)
<220>
<223>合成的
<220>
<221>misc_feature
<222>(4)..(4)
<223>n是t或u
<220>
<221>misc_feature
<222>(6)..(6)
<223>n是t或u
<220>
<221>misc_feature
<222>(8)..(8)
<223>n是t或u
<400>3
acgncnanacgccca15
<210>4
<211>15
<212>dna
<213>智人(homosapiens)
<220>
<221>misc_feature
<222>(1)..(1)
<223>n是t或u
<220>
<221>misc_feature
<222>(6)..(6)
<223>n是t或u
<220>
<221>misc_feature
<222>(13)..(13)
<223>n是t或u
<220>
<221>misc_feature
<222>(15)..(15)
<223>n是t或u
<400>4
ncaccnggagcangn15
<210>5
<211>14
<212>dna
<213>智人(homosapiens)
<220>
<221>misc_feature
<222>(1)..(1)
<223>n是t或u
<220>
<221>misc_feature
<222>(6)..(6)
<223>n是t或u
<220>
<221>misc_feature
<222>(13)..(13)
<223>n是t或u
<400>5
ncaccnggagcang14
<210>6
<211>13
<212>dna
<213>智人(homosapiens)
<220>
<221>misc_feature
<222>(1)..(1)
<223>n是t或u
<220>
<221>misc_feature
<222>(6)..(6)
<223>n是t或u
<220>
<221>misc_feature
<222>(13)..(13)
<223>n是t或u
<400>6
ncaccnggagcan13
<210>7
<211>12
<212>dna
<213>智人(homosapiens)
<220>
<221>misc_feature
<222>(1)..(1)
<223>n是t或u
<220>
<221>misc_feature
<222>(6)..(6)
<223>n是t或u
<400>7
ncaccnggagca12
<210>8
<211>11
<212>dna
<213>智人(homosapiens)
<220>
<221>misc_feature
<222>(1)..(1)
<223>n是t或u
<220>
<221>misc_feature
<222>(6)..(6)
<223>n是t或u
<400>8
ncaccnggagc11
<210>9
<211>10
<212>dna
<213>智人(homosapiens)
<220>
<221>misc_feature
<222>(1)..(1)
<223>n是t或u
<220>
<221>misc_feature
<222>(6)..(6)
<223>n是t或u
<400>9
ncaccnggag10
<210>10
<211>9
<212>dna
<213>智人(homosapiens)
<220>
<221>misc_feature
<222>(1)..(1)
<223>n是t或u
<220>
<221>misc_feature
<222>(6)..(6)
<223>n是t或u
<400>10
ncaccngga9
<210>11
<211>8
<212>dna
<213>智人(homosapiens)
<220>
<221>misc_feature
<222>(1)..(1)
<223>n是t或u
<220>
<221>misc_feature
<222>(6)..(6)
<223>n是t或u
<400>11
ncaccngg8
<210>12
<211>7
<212>dna
<213>智人(homosapiens)
<220>
<221>misc_feature
<222>(1)..(1)
<223>n是t或u
<220>
<221>misc_feature
<222>(6)..(6)
<223>n是t或u
<400>12
ncaccng7
<210>13
<211>16
<212>dna
<213>智人(homosapiens)
<220>
<221>misc_feature
<222>(2)..(2)
<223>n是t或u
<220>
<221>misc_feature
<222>(7)..(7)
<223>n是t或u
<220>
<221>misc_feature
<222>(14)..(14)
<223>n是t或u
<220>
<221>misc_feature
<222>(16)..(16)
<223>n是t或u
<400>13
cncaccnggagcangn16
<210>14
<211>15
<212>dna
<213>智人(homosapiens)
<220>
<221>misc_feature
<222>(2)..(2)
<223>n是t或u
<220>
<221>misc_feature
<222>(7)..(7)
<223>n是t或u
<220>
<221>misc_feature
<222>(14)..(14)
<223>n是t或u
<400>14
cncaccnggagcang15
<210>15
<211>14
<212>dna
<213>智人(homosapiens)
<220>
<221>misc_feature
<222>(2)..(2)
<223>n是t或u
<220>
<221>misc_feature
<222>(7)..(7)
<223>n是t或u
<220>
<221>misc_feature
<222>(14)..(14)
<223>n是t或u
<400>15
cncaccnggagcan14
<210>16
<211>13
<212>dna
<213>智人(homosapiens)
<220>
<221>misc_feature
<222>(2)..(2)
<223>n是t或u
<220>
<221>misc_feature
<222>(7)..(7)
<223>n是t或u
<400>16
cncaccnggagca13
<210>17
<211>12
<212>dna
<213>智人(homosapiens)
<220>
<221>misc_feature
<222>(2)..(2)
<223>n是t或u
<220>
<221>misc_feature
<222>(7)..(7)
<223>n是t或u
<400>17
cncaccnggagc12
<210>18
<211>11
<212>dna
<213>智人(homosapiens)
<220>
<221>misc_feature
<222>(2)..(2)
<223>n是t或u
<220>
<221>misc_feature
<222>(7)..(7)
<223>n是t或u
<400>18
cncaccnggag11
<210>19
<211>10
<212>dna
<213>智人(homosapiens)
<220>
<221>misc_feature
<222>(2)..(2)
<223>n是t或u
<220>
<221>misc_feature
<222>(7)..(7)
<223>n是t或u
<400>19
cncaccngga10
<210>20
<211>9
<212>dna
<213>智人(homosapiens)
<220>
<221>misc_feature
<222>(2)..(2)
<223>n是t或u
<220>
<221>misc_feature
<222>(7)..(7)
<223>n是t或u
<400>20
cncaccngg9
<210>21
<211>8
<212>dna
<213>智人(homosapiens)
<220>
<221>misc_feature
<222>(2)..(2)
<223>n是t或u
<220>
<221>misc_feature
<222>(7)..(7)
<223>n是t或u
<400>21
cncaccng8
<210>22
<211>21
<212>dna
<213>小家鼠(musmusculus)
<400>22
cggaggtgctagatccacaga21
<210>23
<211>21
<212>dna
<213>小家鼠(musmusculus)
<400>23
aggactcgtgcagccttacac21
<210>24
<211>21
<212>dna
<213>小家鼠(musmusculus)
<400>24
cgcttccagagtgcagctatt21
<210>25
<211>23
<212>dna
<213>小家鼠(musmusculus)
<400>25
atcttcaaaaaggccaaccagat23
<210>26
<211>23
<212>dna
<213>小家鼠(musmusculus)
<400>26
tcccatcaaatattgccaaaatt23
<210>27
<211>27
<212>dna
<213>小家鼠(musmusculus)
<400>27
cagaaatagataaatacttccaatcac27
<210>28
<211>18
<212>dna
<213>小家鼠(musmusculus)
<400>28
cagaaaacgcccccagaa18
<210>29
<211>25
<212>dna
<213>小家鼠(musmusculus)
<400>29
aaaagtcgggagaacaagtttcatt25
<210>30
<211>20
<212>dna
<213>小家鼠(musmusculus)
<400>30
cccgcccaaggacaagcgtt20
<210>31
<211>20
<212>dna
<213>小家鼠(musmusculus)
<400>31
ctggcccttcttgtgcgcgt20
<210>32
<211>36
<212>dna
<213>人工序列(artificialsequence)
<220>
<223>合成的
<400>32
ttgtctgcatgagctctcagaagtcttgctcaggtg36
<210>33
<211>37
<212>dna
<213>人工序列(artificialsequence)
<220>
<223>合成的
<400>33
cggtctgacccgtcgacaccatcattcatttaccaag37
<210>34
<211>20
<212>dna
<213>人工序列(artificialsequence)
<220>
<223>合成的
<400>34
gatcgccgtgtaattctagt20
<210>35
<211>20
<212>dna
<213>人工序列(artificialsequence)
<220>
<223>合成的
<400>35
atgttgccagggaactcaat20
<210>36
<211>20
<212>dna
<213>小家鼠(musmusculus)
<400>36
ccaactcagcttcctttcgg20
<210>37
<211>22
<212>rna
<213>秀丽线虫(caenorhabditiselegans)
<400>37
ucaccggguguaaaucagcuug22
- 还没有人留言评论。精彩留言会获得点赞!