基于点击的连接的制作方法
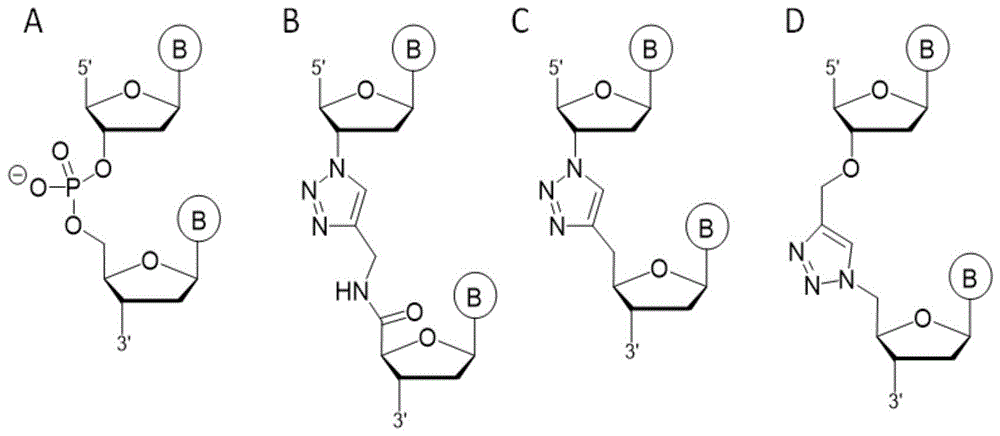
说明本发明涉及在合适的催化剂存在的情况下通过所谓的点击反应偶联分子的新方法和试剂。另外,本发明涉及用于这种点击连接反应的活化剂组合物、点击连接试剂盒、用于进行这种点击连接反应的装置,以及这种方法、组合物、试剂盒和装置用于提高(尤其是在下一代核酸测序方法的背景下)通过点击反应偶联分子的效率。发明背景在2001/2002年,sharpless和meldal小组独立地定义了“点击化学”的概念以及被视为“点击”反应的转化标准(sharpless,k.b.等,angew.chem.2002,114,2708;angew.chem.int.ed.2002,41,2596,meldal,m.等,j.org.chem.,2002,67,3057)。从那时起,铜催化的由叠氮化物与炔烃得到1,2,3-三唑的反应(一种1,3-偶极huisgen环加成反应的变型)(r.huisgen,1,3-dipolarcycloadditionchemistry(ed.:a.padwa),wiley,newyork,1984)已成为使用最广泛的点击反应。由于其温和条件和高效率,该反应在生物学和材料科学中得到非常广泛的应用,诸如目的为dna标记方面的应用(gramlich,p.m.a.等,postsyntheticdnamodificationthroughthecopper-catalyzedazide-alkynecycloadditionreaction.angew.chem.int.ed.2008,47,8350)。针对特定靶序列的遗传物质,例如针对单核苷酸多态性的存在、某种基因(如抗性基因)或mrna的存在的快速分析需要易于使用、有效且可靠的工具。一个主要问题是需要直接在小型生物样品(诸如患者血液或植物)中检测目标dna或rna。这些仅提供微量的分析物。为了达到所需的灵敏度,通常需要扩增步骤,其中在分析之前先扩增核酸分析物,或者使用其中扩增从dna/rna分析物直接获得的微小检测信号的检测方法。核酸分析物的扩增方法包括pcr和其它核酸扩增方案。pcr扩增的主要优势在于,在从生物材料获得的不同dna链库中,仅扩增目标dna序列。这是对复杂生物样品中单个基因进行可靠分析的基础。寡核苷酸的酶促连接是众多寡核苷酸操作方案中的标准程序,是测序、克隆和许多其它基于dna和rna的技术所必需的。参与连接反应催化的酶在dna或rna的5'-磷酸末端与3'-羟基末端之间形成磷酸二酯键。为了高效地工作,连接酶需要双链寡核苷酸,其优选通过使用粘性末端或甚至夹板链来预先化组织。由于该反应不是序列特异性的,因此该连接反应可将任何5'-磷酸与任何3'-羟基末端连接。该底物特异性的缺乏是酶促连接广泛应用的主要有利方面,并有助于其广泛应用。具有讽刺意味的是,对于许多测序应用而言,这同时是一个巨大的挑战,因为从寡核苷酸片段的非特异性连接会产生序列伪像(sequenceartifacts)。由于几乎每种测序技术都需要连接已知的寡核苷酸序列(即所谓的衔接子)以在测序和pcr扩增过程中进行引物结合,因此这可产生人工重组、序列嵌合体形成(用于rna测序的dna-rna链)和衔接子二聚体(一对没有插入序列的连接的衔接子)形成。为了减少序列伪像,通过插入2',3'双脱氧核苷酸(ddntp)封闭引物的3'-末端。这避免了衔接子二聚体的形成并确保正确的定向连接,但其不会影响人工重组和嵌合体的形成。在从序列读取生成重叠群(一组重叠序列数据读取)的过程中,需要使用特殊算法来修正这些影响。本发明涉及用于分子(尤其是寡核苷酸)的非酶连接的基于点击化学的方法,该方法是高效的、位点特异性的并且与pcr扩增相容。两个非模板化单链寡核苷酸之间的反应比类似的酶促连接反应更高效,因此例如,消除了rna测序中对第二链合成的需要。含炔或叠氮化物的寡核苷酸的制备已被广泛研究。对于本发明特别感兴趣的是“主链模拟物”,即磷酸二酯键的非天然替代物,其可通过铜催化的叠氮化物炔烃环加成(cuaac)生成。一些所得的含三唑的寡核苷酸可通过聚合酶转化为天然磷酸二酯主链,而无序列突变,从而具有完全的生物相容性(shivalingam等,molecularrequirementsofhigh-fidelityreplication-competentdnabackbonesfororthogonalchemicalligation(2017)doi:10.1021/jacs.6b11530)。图1显示通过点击化学反应生成的一些三唑主链模拟物的结构,左侧显示用于比较的天然磷酸二酯。这种人工dna主链技术的特别有用的应用在于用于下一代测序的样品制备。routh等(clickseq:fragmentation-freenext-generationsequencingviaclickligationofadaptorstostochasticallyterminated3’-azidocdnas.j.mol.biol.427,2610-2616(2015))提供了用于rna测序的方案,其中将衔接子寡核苷酸点击至3'-叠氮基终止的cdna片段,并进行pcr反应以提供cdna文库。与涉及衔接子寡核苷酸的酶促连接的标准文库制备方案相比,该方法可以大大降低人工重组和嵌合序列形成的速率。迄今为止,对点击技术在dna或rna测序中的这种应用的严重限制是效率不令人满意,在该方法中,在文库生成过程中捕获了该方法中使用的3'-叠氮基封闭的cdna片段。仅约10%的3'-叠氮基封闭的cdna片段可与炔烃修饰的寡核苷酸连接。此外,通过三唑键联的通读将该三唑键连接的单链dna转化成双链dna效率不高。虽然对于标准rnaseq实验而言,这样的效率可能已经足够,但对于需要彻底捕获原始rna或dna的其它应用程序,则认为点击反应和后续的通读效率不足,必须加以改进。因此,本发明的目的是提供实现更高的效率(特别是点击连接的)以便建立点击技术在下一代测序中的成功应用的手段。技术实现要素:本发明涉及在使用炔基和叠氮基作为点击官能团的点击反应中提高连接两个反应配偶体的效率。下文中所期望的点击反应是由金属催化剂(尤其是非均相cu(i)或另一种合适的金属催化剂)催化的1,3-偶极部分(其为叠氮化物)与不饱和部分(其为末端炔烃)之间的反应。催化形式的点击反应是非协调离子机制(non-concertedionicmechanism),其导致5元杂环1,2,3-三唑部分。根据第一方面,本发明涉及在点击反应中将第一分子与第二分子偶联的方法,其中所述第一分子包含为炔基的第一点击官能团,并且所述第二分子包含为叠氮基的第二点击官能团,该方法包括在催化剂,优选非均相cu催化剂存在的情况下使第一分子与第二分子在反应混合物中接触,该方法的特征在于在其它金属阳离子存在下的情况下在所述反应混合物中进行点击反应。本发明的又一方面是用于点击反应的活化剂组合物,其中在催化剂,优选非均相cu催化剂存在的情况下,将包含第一点击官能团(其为炔基)的第一分子与包含第二点击官能团(其为叠氮基)的第二分子偶联,所述活化剂混合物包含金属阳离子、cu(i)稳定配体和任选地有机溶剂。本发明的又一另外的方面是点击连接试剂盒,其至少包含根据本发明的催化剂(作为一种组分)和活化剂组合物(作为第二组分)。本发明的又一另外的方面涉及具有至少一个反应室的装置,其中该反应室包含用于在两个反应配偶体之间进行点击反应的催化剂,优选为非均相铜催化剂,并且另外地其中本发明的金属阳离子或活化剂组合物包含在反应室中。本发明的又一另外的方面涉及上述方法、组合物、试剂盒和装置用于通过点击反应高效地连接分子(优选寡核苷酸)的用途。此类分子的实例具有可检测的标记物。因此,该连接可提供可用于检测样品中的分析物(例如核酸)的标记的生物分子,特别地涉及通过点击反应标记的化合物的用途,所述标记的化合物与待检测的分析物的形成缔合产物。特别地,所述方法、组合物、试剂盒和装置可用于下一代测序技术,包括rna文库制备、dna文库制备,以及用于复杂寡核苷酸混合物的分析,尤其是并行和多重rt-qpcr应用。另外,本发明的方法、组合物、试剂盒和装置可用于通过连接方法进行测序(例如来自abi的“固体测序”),用于连接酶链反应、克隆方法(特别地以形成重组dna)以及基因合成。导致本发明的研究表明,令人惊奇地将金属离子添加到通过催化剂,特别是通过cu(i)催化剂,尤其是非均相cu(i)催化剂催化的点击反应中,尤其可大大改善寡聚物-寡聚物点击反应动力学。这种改善允许在主链官能化的寡核苷酸之间进行高效的点击反应,其产量直到现在才通过反应配偶体的预先化组织(通过夹板寡核苷酸)而获得。优选实施方案的详细描述本发明提供了用于点击反应的改进的条件,所述条件避免了对夹板寡核苷酸的需要并且仍然导致点击连接反应的高产率。在可在本发明的条件下进行的这种点击反应的实例中,可以例如通过在反转录(rna测序)或平端化(将单链dna悬突用核苷酸填充和/或去除)过程中通过酶促掺入和/或通过da拖尾(将核苷酸,最常见地datp非模板化地添加到平端双链dna的3'末端)将3’-炔烃或3’-叠氮化物修饰的核苷酸引入寡核苷酸。对于rna测序,可使用部分随机化的引物引入第一衔接子(衔接子是包含用于引物结合和杂交的互补序列的短寡核苷酸)。将第二衔接子点击连接至3'-炔烃或3'-叠氮化物终止的cdna。clickadaptrna文库制备过程中这种反应的示意图如图2a所示。特别地在这种rna测序的背景下,可能以及可以甚至更优选采用含有用于引物结合和杂交的互补序列的衔接子,其在其5'末端包含与衔接子更下游区域互补的序列。反向回文序列或反向重复序列是此类互补序列的实例。衔接子序列的这些互补部分的杂交导致在衔接子的5'末端形成双链环。使用在5'末端包含双链环的这种衔接子可确保该衔接子不会与反应混合物中包含的相同或另一种rna上的其它序列杂交。在图2b)中示意性地显示了相应的反应。尽管除了双链部分以外,在这种衔接子中还存在单链环区域,但这种环区域不太可能引起非特异性杂交。然而,优选的是使这种环区保持很小。优选地,环区域包含少于10个核苷酸,更优选地少于6个核苷酸,最优选地,环区域仅包含3个核苷酸。对于dna测序,将第一衔接子点击连接至酶促3'-炔烃或3'-叠氮化物修饰的寡核苷酸(在pcr过程中通过衔接子的部分互补结合区(例如12聚体)引入第二衔接子)。相应的clickadaptdna文库制备流程如图3所示。引物或衔接子可包含索引序列,以允许与不同样品之一的后续关联。这允许将点击测序技术应用于几种酶促产生的样品的混合物。去除多余的经修饰的核苷酸后,在酶促产生并纯化的经修饰的寡核苷酸与充当衔接子的5'-炔烃或5'-叠氮化物修饰的寡核苷酸之间进行点击连接,以引入进行pcr扩增和/或测序所必需的核苷酸序列。为此,衔接子寡核苷酸优选通过固相合成并在这种合成结束时引入5’-修饰而产生。作为炔烃修饰元件,可使用例如具有以下结构的商购可得的构件块:作为点击反应的催化剂,使用金属催化剂,优选cu催化剂,最优选非均相cu(i)催化剂。特别地,如ep2416878b1中所述的催化剂可用作点击反应的催化活性铜种类的来源。特别参考ep2416878b1的第[0029]至[0031]段,其中提供了此类催化剂的详细描述。在本发明中,点击反应包括铜催化的叠氮化物与末端炔基之间的形式(3+2)环加成的变型。由于生物体中缺乏叠氮化物和末端炔烃,叠氮化物/炔烃反应导致的1,2,3-三唑不可逆形成是双正交的,所需的化学基团很小(对生物分子环境的破坏最小的掺入)并且反应是位置选择性的,导致1,4位置异构体的排他性形成。以下反应是叠氮化物与末端炔炔的点击连接的基础其中r1和r2是点击反应中的第一和第二配偶体。根据上文引用的现有技术文件,本领域技术人员了解点击反应条件。通常,在室温或略微升高的温度下(优选在20℃与60℃之间,更优选在30℃-50℃下,最优选在40℃-45℃下)在水性反应混合物中进行点击反应。取决于温度,该反应通常需要10分钟至数小时,优选30分钟至3小时,最优选40分钟至90分钟的孵育时间。反应条件不是关键的,并且可调整为用于制备点击产物的试剂的量和体积。催化剂优选为非均相cu催化剂,更优选为非均相cu(i)催化剂。然而,应该指出的是,其它金属催化剂,尤其是其它非均相金属催化剂,诸如zr,w,fe,ru,co,th,ir;ni,pd,pt,ag,au,zn,cd,hg和其它金属离子据报道直接或间接地有助于点击反应连接的催化,并且也可在本发明的背景中使用。或者,也可将均相cu(i)催化剂用于点击连接。非均相cu催化剂是元素铜或金属-c催化剂,即,包含碳基载体(诸如其中结合有cu离子的炭)或元素铜的固体cu催化剂,其可生成cu(i)离子。在特别优选的实施方案中,非均相cu催化剂是cu(i)-c催化剂,可如h.lipshutz和b.r.taft在angew.chem.int.ed.,2006,45,8235-8238中所述制备所述cu(i)-c催化剂。所述非均相催化剂可以是颗粒催化剂,例如由尺寸为10nm至1000μm,优选为100μm至800μm的颗粒组成的非均相催化剂。或者,催化剂也可以是多孔的非颗粒催化剂,例如在其中嵌入了催化活性颗粒的固体基质。在又一个实施方案中,包括另外的固体载体材料,该材料是与非均相cu催化剂不同的材料,例如可在其上固定生物分子(诸如优选地核酸或核酸类似物)的色谱材料。优选地,包括允许将点击反应产物与反应混合物的另外成分分离和纯化的材料。分离和纯化的示例性机制是尺寸排阻色谱或亲和色谱。合适的色谱材料的实例是离子交换材料、亲水材料或疏水材料。在优选实施方案中,可将亲水性材料例如硅胶与非均相催化剂结合使用。在另一个优选实施方案中,疏水性材料例如二氧化硅c18或c4或离子交换树脂可与非均相催化剂结合使用。在又一另外的优选实施方案中,固体载体材料可以是用于生物分子,尤其是核酸和核酸类似物的固相合成的树脂。已经发现,固定化反应配偶体与游离存在于溶液中的反应配偶体之间的点击反应可通过非均相cu催化剂体系有效地催化。该策略可允许同时实现点击反应以及产物的纯化,和/或,产物与杂质和/或与最终存在于反应混合物中的过量试剂或盐的分离。在根据本发明的方法中,反应混合物包含另外的金属阳离子。在本发明的说明书中,术语“另外的金属阳离子”是指与在点击反应过程中以阳离子形式存在或出现的与催化剂金属不同的金属阳离子。优选金属阳离子是碱金属和碱土金属,或其它具有类似性质的二价金属离子例如锌(zn2+)。在本发明的优选实施方案中,要添加到反应混合物中的此类金属阳离子,尤其是此类碱金属与na+不同。在本发明的特别优选的实施方案中,反应混合物中存在二价碱土金属离子,尤其是mg2+离子,已证明它们是用于阴离子dna磷酸酯主链的特别合适的阳离子。用于本发明目的的其它优选的另外的阳离子是li+、k+和zn2+。为此目的,在反应混合物中包含相应的金属盐,尤其是mg2+盐。在导致本发明的研究过程中,令人惊讶地发现,向反应混合物中添加金属阳离子,尤其是mg2+,导致寡核苷酸的磷酸主链上的铜种类的所有可能的配体位置被另外的阳离子占据。结果,在反应混合物中可用作催化剂的铜种类的量增加,并且反应动力学得到显著改善。点击反应混合物中合适的其它金属阳离子的量为1至200mmol/l,优选5至25mmol/l,特别优选10至20mmol/l。在根据本发明的点击反应中,优选地还包括催化剂金属稳定配体,尤其是cu,优选cu(i)稳定配体。可将已知的金属配体,如胺和聚三唑(t.r.chan,r.hilgraf,k.b.sharpless和v.v.fokin,organicletters,2004,6,2853-2855.),尤其是三(3-羟基丙基三唑基甲基)胺(thpta)、2-(4-((双((1-(1-叔丁基)-1h-1,2,3,-三唑-4-基)甲基)氨基)甲基)-1h-1,2,3-三唑基-1-基)乙酸(bttaa)、2-(4-((双(((1-(叔丁基)-1h-1,2,3,-三唑-4-基)甲基)氨基)甲基)-1h-1,2,3-三唑基-1-基)硫酸乙酯(bttes)(d.sorianodelamo,w.wang,h.jiang,c.besanceney,a.c.yan,m.levy,y.liu,f.l.marlow,p.wu,j.am.chem.soc.2010,132,16893–16899.)、三((1-苄基-4-三唑基)甲基)胺(tbta)或其具有类似金属稳定特性的类似物作为这样的配体用于反应混合物中。这些金属配体稳定例如cu(i)催化剂并保护寡核苷酸免于形成活性氧种类。另外,添加此类金属配体还改善了点击反应的反应动力学。金属配体以10至4000μmol/l,优选500至1000μmol/l,特别优选700至900μmol/l的量包含在点击反应混合物中。在本发明的另一优选实施方案中,将有机溶剂加入反应混合物中。特别地,可以有利地将二甲基亚砜(dmso)包含在反应混合物中。这种有机溶剂,尤其是dmso的添加对两个寡核苷酸之间的点击反应的效率具有进一步的积极影响。假定dmso干扰寡核苷酸的二级结构,从而改善了反应配偶体的官能团(例如叠氮基和炔基)的可及性。在本发明的说明书中,向反应混合物中添加最终含量为1至10%(v/v)的这种有机溶剂,尤其是dmso被认为是有用的,向反应混合物中添加最终含量为2至8%(v/v),尤其是4-6%(v/v)的有机溶剂是优选的。令人惊讶地观察到,点击反应的反应混合物中存在另外的金属离子,特别是存在相应的二价金属离子导致点击反应产物形成的显著增加。与在不添加此类二价金属阳离子的情况下进行的反应相比,点击产物的产率可以至少加倍或甚至增至三倍。使用还包含有机溶剂,尤其是dmso和cu稳定配体的反应混合物,可以进一步改善性能,并导致令人高度满意的结果和点击产物的产率。根据本发明的方法的这些效果促进了此类击产物的后续反应和用途,尤其促进了用于dna或rna文库制备和下一代测序的pcr反应。考虑到另外的金属离子,尤其是二价金属离子和一些其它物质与点击连接效率的相关性,本发明的另一主题是用于点击反应的活化剂组合物,其用于在cu催化的反应,优选在非均相铜催化剂存在的情况下进行的反应中偶联一方面被末端炔基官能化,另一方面被叠氮基官能化的分子。这种活化剂组合物包含金属阳离子。如上所述,向点击反应混合物中添加此类另外的金属阳离子,优选碱土金属阳离子,尤其优选地mg2+离子,可防止铜催化剂种类与dna或rna磷酸酯主链结合。这种结合在不添加此类金属阳离子的情况下发生,并减少了可用于点击反应的催化剂的量。由于金属阳离子,尤其是二价金属阳离子的存在,封闭了磷酸盐主链上结合位点,可用作催化剂的铜的量增加,反应动力学得到改善。如以上已经详细解释的,cu(i)稳定配体和/或有机溶剂,尤其是dmso的存在进一步提高了两个寡核苷酸之间的点击反应的效率。因此,根据本发明的优选活化剂组合物包含二价金属阳离子和有机溶剂以及至少一种铜稳定配体。尤其优选的是包含作为二价金属阳离子的mg2+和dmso以及选自thpta、bttaa、其类似物或此类配体的任何混合物的至少一种铜稳定配体的活化剂组合物。活化剂组合物以这样的量包含这些效应分子,所述量使得在点击反应混合物中二价金属阳离子的浓度为1至200mmol/l,优选为5至25mmol/l,特别优选为10至20mmol/l;cu稳定配体的浓度为10至4000μmol/l,优选为500至1000μmol/l,尤其优选为700至900μmol/l;和/或有机溶剂,优选dmso的浓度为1至10%(v/v),优选为2至8%(v/v),特别优选为4至6%(v/v)。根据本发明的活化剂组合物可以以包含一种、两种或所有上述物质的组合物的形式提供,所述物质有助于提高点击反应的效率。活化剂组合物可以是包含效应物物质的预稀释液的水性组合物。活化剂组合物还可包含其它溶剂,尤其是另外的有机溶剂或水和有机溶剂的组合。活化剂组合物还可包含其它物质,如缓冲物质或可包括在点击反应的执行中的任何其它物质。本发明的另一主题是点击连接试剂盒,这种试剂盒至少包含如上定义的催化剂,特别是cu催化剂,优选非均相cu(i)催化剂(作为一种组分)和根据本发明的活化剂组合物(作为第二组分)。如上更详细地公开的,在连接炔烃和叠氮化物官能化的寡核苷酸中,活化剂组合物对铜催化的点击反应具有明显的积极作用。因此,本发明的试剂盒一起提供了催化剂以及活化剂组合物的一种或多种效应分子,以便于实施本文公开的点击连接方法。在优选实施方案中,点击连接试剂盒包含相应的优选实施方案作为活化剂组合物,其中不仅存在金属阳离子,优选碱土金属阳离子,尤其是mg2+,而且还存在有机溶剂和cu稳定配体中的至少一种。为了获得最佳结果,将包含所有三种效应物的活化剂组合物包含在本发明的点击连接试剂盒中。包含在本发明的点击连接试剂盒中的cu(i)催化剂优选是如ep2416878b1中所述的非均相催化剂。任选地,这种试剂盒包含其它物质,所述物质是必需的组分和/或在执行点击反应中是有利的。除了上述组分以外,本发明的点击连接试剂盒还可包含至少一种可用作衔接子或引物的叠氮化物官能化或炔烃官能化的寡核苷酸。试剂盒可以包含标记物、标记的点击官能基团,经修饰的和未修饰的核苷酸、酶、缓冲液、衔接子和/或引物、用于纯化的其它载体材料或色谱材料,优选地根据本发明的点击连接方法的背景中的上述材料及其各种应用。如上所述,取决于要进行的反应,可包括包含互补序列的衔接子和引物,所述互补序列在所述衔接子或引物的5'末端形成环和双链区域。在本发明的另一优选实施方案中,点击连接试剂盒可另外地包含随后的pcr反应所需的物质和组分,以扩增点击连接的产物,以用于dna或rna文库的制备和测序,例如。用于分析复杂的寡核苷酸混合物。本发明的另一主题是有助于进行本发明的点击反应的装置。这种装置包括至少一个反应室,该反应室包含用于在两个反应配偶体之间进行点击反应的催化剂,优选为非均相cu催化剂。在ep2416878b1中公开并进一步描述了相应的装置,并且该公开也适用于可根据本发明使用的装置。根据本发明,除了存在点击连接反应的一种或多种组分以外,该装置还在此装置的一个或多个反应室内包含如上定义的另外的金属阳离子,尤其是二价金属阳离子,优选碱土金属阳离子,尤其是mg2+,或上述活化剂组合物。由于点击反应可以以非常小的体积进行,因此装置可以例如是微量滴定板孔、移液器吸头或旋转柱,包括一个或多个可用作至少一个反应室的区室。根据本发明采用的金属离子或其它优选组分存在于此类装置中的至少一个进行点击反应的反应室中。应当理解,在本发明的说明书中,包括进行这种点击反应的所有不同方式和为此类目的描述的所有物质,只要满足如上所述的本发明的方法、组合物、试剂盒和装置的基本要求即可。因此,只要用于催化点击连接的点击反应混合物包括已被本发明证明对点击反应的效率和产率具有相当大的有利影响的另外的金属离子,尤其是二价金属阳离子,在本文中未明确描述的点击反应的所有其它形式、变型和改进被认为在本发明的说明书中也是适用的。虽然这些阳离子,尤其是碱土金属阳离子,最优选mg2+离子的存在对于解决本发明的目的是强制性的,但点击反应的其它方面可根据例如通过点击连接来偶联的分子的性质、实际使用的催化剂、反应条件和预期用途以及将要进行的相应后续反应而变化。所述方法、组合物、试剂盒和装置用于通过点击反应高效连接分子(优选寡核苷酸)以提供可用于后续反应的点击反应产物的用途是本发明的另一主题。作为这样的后续反应,包括所有需要连接产物的反应,所述连接产物具有高纯度并且以足够的主(lead)高产率提供以进行这种后续反应。此类后续反应的实例是用于分析dna或rna样品的pcr或其它扩增方法,尤其是在要分析复杂的寡核苷酸混合物的情况下。本发明允许获得构成“主链模拟物”的寡核苷酸,即在其主链中含有磷酸二酯键的非天然替代物的rna或dna分子的点击连接产物。由于发现此类物质是完全生物相容的,因此此类分子的扩增非常有效且有用,特别是对于所谓的下一代测序应用而言。当将上述条件应用于例如rna测序文库制备时,可将第二衔接子高效地点击连接至单链cdna,从而无需合成第二链,从而减少了总制备时间。由于点击反应的生物正交性和特异性,点击连接仅在叠氮化物官能化和炔烃官能化分子之间发生。叠氮化物在cdna合成过程中被随机引入的,因为给天然脱氧核苷酸补充了低浓度的例如3’-叠氮基-2’,3’-二脱氧核苷酸,其终止片段。5'-炔烃修饰的衔接子寡核苷酸可通过固相dna合成制备,并且无需纯化即可使用,因为5'-炔烃最后被掺入固相合成中,且较短的未标记链无法被点击连接。本发明的点击概念可以特别优选地应用于dna文库制备方案。通常通过平端化和da拖尾(通过dna聚合酶)来修饰dna片段,以实现更高效的后续酶促连接。在平端化和da加尾期间用例如替换天然的dntp。钝化或da拖尾过程中用例如3'-叠氮基-2',3'-二脱氧核苷酸置换天然dntp引入叠氮基,以用于点击连接至5'-炔烃衔接子寡核苷酸。如果要分析复杂的寡核苷酸混合物,则本发明在例如由routh等(见上文)描述的clickseq技术的背景下的应用也非常有价值。可通过执行多重rt-qpcr对rna病毒rna进行并行分析。为此,将几种病毒rna特异性引物(例如hiv、丙型肝炎等)添加到样品中的rna中,以进行逆转录反应(例如clickadapt反应)。点击叠氮基终止的cdna片段,使其与衔接子连接,以提供用于后续pcr扩增的引物结合位点。由于点击连接的特异性,因此不会发生人工重组,从而无需为每次检测使用单独的qpcr设置。因此,该技术为大规模并行rt-qpcr应用提供了基础。该技术的应用领域包括用于测序、通过连接方法测序(例如,来自abi的“固相测序”)、连接酶链反应、克隆方法(以完成重组dna)和基因合成的所有其它dna和rna文库制备试剂盒,其中通过点击官能团连接两个或更多个分子。尽管上文中已部分一般性地和部分更详细地描述了可能的应用,但应该理解,包括在本文所述条件下进行的点击连接的其它反应也包括在本发明的范围内。附图说明图1显示与天然磷酸二酯(a)相比,通过点击化学(b-d)产生的一些三唑主链模拟物的结构。仅对三唑主链拟物(b)显示了pcr扩增背景中的后续实验实例;图2a)显示示意性clickadaptrna文库制备,其为本发明中的目标方法之一。其涉及组合的cdna合成、片段化和衔接子点击连接。第二链合成已过时,因为ssdna可被高效点击连接。图2b)显示与图2a)相同的clickadaptrna文库制备,但对于该方法,包括了在5'末端包含双链环的第一衔接子。图3显示示意性clickadaptdna文库制备工作流程,其为本发明中的另一目标方法。将样品双链(ds)dna片段化,然后如在标准dna文库制备中一样通过平端化和da拖尾操作片段。通过使用3'-叠氮基-2',3'-二脱氧核苷酸替代天然dntp,获得3'叠氮化物终止的dsdna。通过片段纯化(包括大小选择)去除多余的核苷酸后,将第一衔接子通过其5'-炔基点击至片段。在pcr扩增过程中,通过短的(约12bp)3'序列引入第二衔接子,所述3'序列是第一衔接子的5'末端的反向互补序列,并用作扩增的第一引物。图4显示各种阳离子对点击反应效率和产率的影响。图5显示使用不同的dna聚合酶从3’-叠氮化物终止的cdna和5’-炔烃衔接子产生的cdna的pcr产物;图6显示图5的扩增的点击连接产物的sanger测序结果。图7显示用于酶掺入并随后用于点击连接的叠氮化物和炔烃修饰的核苷酸的示例性结构。图8显示用于点击连接(a-c)的衔接子寡核苷酸的示例性5’-末端的结构。5’-炔烃修饰的寡核苷酸(其中碱基b=结构a的胸腺嘧啶)用于实施例1、2和4(在图4、图5和图9中)。结构b用于随后的实施例1和4(在图4和9中)。图9显示低寡聚物浓度下的寡聚物-寡聚物点击反应的产率。图10显示pcr样品的溴化乙锭染色的琼脂糖凝胶(3%,于tae中),其中模板来自egfpmrna的点击文库制备。在反转录(rt)过程中,使用不同的核苷酸混合物(dntp恒定为500μm,各种azddntp)生成模板cdna。m=低分子量dna标志物(neb),1=仅dntp,2=100μmazddntp,3=50μmazddntp,4=25μmazddntp,5=10μmazddntp。图11显示使用mgso4作为添加剂的寡聚物染料(oligo-dye)cuaac反应的分析hpl色谱图。图12显示在260nm下检测来自寡聚物-寡聚物点击粗反应混合物的分析hplc结果。5.6分钟时的峰对应于炔烃修饰的寡聚物,7.6分钟时的峰对应于叠氮化物修饰的寡聚物。点击反应后新出现的两个峰(6.1分钟和6.4分钟)具有正确的点击产物质量。约80%的积分峰具有esi-ms确认的点击产物的质量。提供以下实施例用于说明目的。实施例1:点击产物的准备在200μl反应小瓶中,将单个反应器沉淀物(600-800μm,包含元素铜)与12.5μl反应混合物组合,并在45℃下孵育60分钟。反应混合物由4mmthpta、55μm炔烃寡聚物1、55μm叠氮化物寡聚物1以及在研究阳离子影响时16mm单价阳离子(或8mm二价阳离子)组成。如有必要,可使用dh2o将体积调整至最终12.5μl。孵育后,将样品短暂离心沉淀,并将上清液转移至新的小瓶中以终止反应。在于tae缓冲液(20mmtris,10mm乙酸,0.5mmedta)中制备的2.5%琼脂糖凝胶(10x15cm)上分析样品。用20%紫色加样染料(neb,newenglandbiolabsinc.)制备样品,并相应地制备低分子量dna梯状条带(25-766bp,neb,n3233);通常在5μl加样体积中使用0.5μl标志物。通过施加恒定功率(10w,最大500v,最大100ma)将凝胶在tae缓冲液中运行60分钟。然后,将凝胶在新鲜制备的1:10000溴化乙锭稀释液中孵育15分钟,然后在dh2o中脱色15分钟。为了可视化,使用了geldocezimager(biorad)。寡核苷酸炔烃寡聚物1:5'-taatgatacggcgaccaccgagatctacactctttccctacacgacgctcttccgatct-3't=5'-炔烃dt叠氮化物寡聚物1:5‘-n3-tggagttcgtgaccgccgccgggatcactctcggcatggacgagctgtacaagtaaagc-3‘由于此实验中使用的非理想点击条件(thpta过多,铜源不足),碱(土)金属添加的阳离子添加的影响变得明显。图4显示寡核苷酸点击反应的2.5%凝胶,以及不同阳离子对点击效率和产物产率的影响。在没有其它阳离子的情况下(槽1),在上述条件下观察到少于5%的点击产物的产率。通过添加mg2+离子(8mm),产率提高到约30%。作为比较,还分析了浓度为16mm的单价阳离子,然而仅观察到了收率的略微提高(槽3-5,产率为5至10%)。实施例2:点击产物的pcr扩增clickadapt方案的可行性以模型rna序列为例。将rna与引物1杂交,然后在200μmdttp、dgtp、dctp和3'-叠氮基-ddatp存在的情况下使用mulv逆转录酶进行反转录。按照制造商的说明,使用核苷酸去除试剂盒(qiagen)纯化cdna,从而去除核苷酸和酶。在200μl反应小瓶中,在总共12.5μl反应混合物中,用单个反应器沉淀物(600-800μm,包含元素铜)将炔烃寡聚物1点击至纯化的cdna,并在45℃下孵育60分钟。反应混合物由800μmthpta、20mmmgcl2、5%(v/v)dmso、7μm炔烃寡聚物1和约4μm纯化的cdna组成。如有必要,可使用dh2o将体积调整至最终12.5μl。孵育后,将样品短暂离心沉淀,并将上清液转移至新的小瓶中以终止反应。将粗点击反应物以1:1000、1:5000和1:10000(最大4nm、0.8nm和0.4nm)的比例稀释,无需进一步纯化即可进行pcr扩增。在200μl反应小瓶中,以20μl的总体积中制备pcr扩增物。将点击反应稀释液与200μmdntp、10pmol引物2和引物3以及1个单位的聚合酶混合。对于各种聚合酶,按照制造商的推荐使用pfu、phusion、q5、onetaq和dreamtaq缓冲液。将样品在热循环仪(biorad)中进行热循环程序。作为标准循环条件,使用以下条件:对于pfu聚合酶,研究了不同的模板稀释度和替代循环条件:孵育后,将样品短暂离心沉淀,并在tae缓冲液(20mmtris,10mm乙酸,0.5mmedta)中制备的3%琼脂糖凝胶(10x15cm)上分析等分试样。用20%的紫色加样染料(neb)制备样品,并相应地制备低分子量dna梯状条带(25-766bp,neb,n3233);通常在5μl加样体积中使用0.5μl标志物。通过施加恒定功率(10w,最大500v,最大100ma)将凝胶在tae缓冲液中运行60分钟。然后,将凝胶在新鲜制备的1:10000溴化乙锭稀释液中孵育15分钟,然后在dh2o中脱色15分钟。为了可视化,使用了geldocezimager(biorad)。图5a显示针对所示模型rna完成的clickadapt工作流程(如上所述)。该工作流程涉及将rna反转录为cdna。通过用3'-azddatp替代天然datp,cdna会以3'-叠氮化物终止。通过纯化cdna去除多余的核苷酸后,将5'-炔烃衔接子点击连接到3'-叠氮化物上,并将粗反应混合物用作pcr模板。图5b是根据5a中模型rna的clickadapt工作流程测试的pcr样品的溴化乙锭染色的琼脂糖凝胶。由于在不同条件下通过各种聚合酶扩增了含三唑的模板,因此这说明非天然主链模拟物的生物相容性。寡核苷酸炔烃寡聚物1(参见实施例1)rna模板:5’-uucgacaaacgaaaacacaaacacaaaccaaacagaaaacaguacauguaaucgacca-3’引物1(用于反转录)5′-fam-tggtcgattacatgtac-3‘;fam=荧光素引物25’-tggtcgattacatgtactgtttt-3’引物35’-agatcggaagagcgtcg-3’反转录后所得的cdna:5’-fam-tggtcgattacatgtactgttttctgtttggtttgtgtttgtgttttcgtttgtcga-n3所得的点击产物:5’-fam-tggtcgattacatgtactgttttctgtttggtttgtgtttgtgttttcgtttgtcgataatgatacggcgaccaccgagatctacactctttccctacacgacgctcttccgatct-3’at=通过主链模拟物b连接的a和t所得pcr产物:5’-tggtcgattacatgtactgttttctgtttggtttgtgtttgtgttttcgtttgtcgataatgatacggcgaccaccgagatctacactctttccctacacgacgctcttccgatct-3’实施例3:扩增产物的序列测定通过测定核酸中核碱基序列的方法来分析扩增产物。已包括的衔接子序列允许互补寡核苷酸的杂交,允许固定、序列测定或进一步扩增。图6显示实施例2的扩增产物之一的sanger测序结果。确定了通过使用phusiondna聚合酶,在三唑主链修饰的位置处或附近没有观察到突变。结果表明,点击反应产物可以成功地包含在pcr反应中,从而以高效率和准确度提供大量pcr产物。实施例4:在反应器(铜源)存在和不存在的情况下的低浓度点击连接在200μl反应小瓶中,将两个反应器沉淀物(600-800μm,包含元素铜;样品1)与12.5μl反应混合物组合或不将反应器沉淀物(样品2)与12.5μl反应混合物组合,并在45℃下孵育60分钟。反应混合物由dh2o中的800μmthpta、20mmmgcl2、7μm炔烃寡聚物1和7μm叠氮化物寡聚物1组成。孵育后,将样品短暂离心沉淀,并将上清液转移至新的小瓶中以终止反应。在于tae缓冲液(20mmtris,10mm乙酸,0.5mmedta)中制备的3%琼脂糖凝胶(10x15cm)上分析样品。用20%的紫色加样染料(neb)制备样品,并相应地制备低分子量dna梯状条带(25-766bp,neb,n3233);通常在5μl加样体积中使用0.5μl标志物。通过施加恒定功率(10w,最大500v,最大100ma)将凝胶在tae缓冲液中运行60分钟。然后,将凝胶在新鲜制备的1:10000溴化乙锭稀释液中孵育15分钟,然后在dh2o中脱色15分钟。为了可视化,使用了geldocezimager(biorad)。寡核苷酸炔烃寡聚物1:5'-taatgatacggcgaccaccgagatctacactctttccctacacgacgctcttccgatct-3't=5'-炔烃dt叠氮化物寡聚物1:5‘-n3-tggagttcgtgaccgccgccgggatcactctcggcatggacgagctgtacaagtaaagc-3‘该实施例的结果如图9所示。当存在反应器时,使用该实施例的点击条件在60分钟后获得36%的产率。当省略反应器时,未观察到产物。实施例5:使用ivtmrna的rna文库制备方案我们在此处描述了使用纯化的体外转录的(ivt)编码egfp基因的mrna在方案开发期间获得的文库制备方案的详细实验条件。反转录在200μl不含rna酶的试管中,将250ngivtmrna与100pmol部分随机化引物、1x反应缓冲液、10mmdtt(二硫苏糖醇)、500μmdntp、0-100μmazddntp和200单位逆转录酶组合。a)引物杂交移液方案:设置/组分12345h2o7.35μl6.95μl6.55μl5.35μl6.35μlillumina_n6引物(100μm)1μl1μl1μl1μl1μlegfpmrna(382ng/μl)0.65μl0.65μl0.65μl0.65μl0.65μldntp(10mm)1μl1μl1μl1μl1μlazddntp(2mm)----1.0μlazddntp(500μm)-0.4μl0.8μl2.0μl-通过轻轻移液将各组分混合,然后在热循环仪中加热至65℃持续3分钟,然后冷却至4℃。为了添加其余成分,制备了预混液(用于6种设置):组分体积h2o18μl反转录缓冲液1(5x)24μldtt1(100mm=10x)12μlprotoscriptii逆转录酶1(200u/μl)6μl1=来自neb(产品编号m0368l)在室温(23℃)下,向每个杂交设置(1-5)中添加10μl预混液,通过移液混合后,将反转录物在25℃下于热循环仪中孵育10分钟,在42℃(protoscriptii逆转录酶的最适温度)下孵育50分钟,在65℃(变性)下孵育20分钟。冷却至4℃后,向每个设置中添加5μlnaoh(aq)(1m),然后在95℃下孵育15分钟,然后在4℃下孵育。通过添加5μlhcl(aq)(1m)来中和混合物,然后按照制造商的建议使用qiagenpcr纯化试剂盒(添加150μlpb缓冲液,使用30μlh2o进行最终的洗脱步骤)进行纯化。nanodrop测量设置a260β[ng/μl]注释总体积估计的cdna量10.05821.1强dna光谱29μl610ng20.05419.6强dna光谱28μl540ng30.04415.9典型的dna光谱29μl460ng40.03311.1典型的dna光谱29μl320ng50.03713.3典型的dna光谱28μl370ng逆转录过程中增加的azddntp浓度会降低cdna产率。我们假设增加的azddntp量会减小片段大小,并且在cdna纯化过程中会移除<100聚体的片段。点击连接在200μl的反应管中,将两个反应器沉淀物与1.25μl的活化剂(200mmmgcl2,在50%(v/v)dmso水溶液中的8mmthpta)(10x)、1μl炔烃衔接子寡聚物(100μm)和10.25μl纯化的cdna组合用于每种设置(1-5)。将混合物在热混合器中在45℃,600rpm下孵育60分钟。然后将每个样品短暂离心沉淀,并将上清液转移至新鲜的小瓶中。pcr在200μl的反应管中,通过将0.5μl点击反应物、10pmol引物、0.5单位的phusion缓冲液中的phusiondna聚合酶和200μmdntp组合制备了20μlpcr。制备了用于6种设置的预混液:3=来自thermofisherscientific,产品编号f530l通过轻轻移液将各组分混合,然后将19.5μl预混液添加到0.5μl完成的点击反应中(无沉淀物!)并混合。使用以下温度程序,将所得混合物在热循环仪中孵育:通过琼脂糖凝胶电泳分析5μl的每个pcr样品。在初始反转录过程中,仅用dntp(设置1,泳道1)开始加载样品,这提供了180bp的单一弱条带和引物(约35bp)。在反转录过程中通过azddntp掺入产生的来自cdna的pcr样品导致100bp(纯化方法的大小截断值)至700bp(泳道5)的拖尾(泳道2-5,设置5-2)。片段大小分布似乎从泳道3-5开始增加,因为在反转录过程中azddntp的量从50μm减少至10μm(脉道3至5)。溴化乙锭染色的琼脂糖凝胶如图10所示。寡核苷酸t=5’-炔烃dt实施例6:使用金属阳离子的寡聚物染料点击反应在1500μl反应小瓶中,将四个反应器沉淀物与27.9μl反应混合物混合,并在45℃下孵育60分钟。反应混合物由2.9μldmso和25μlh2o中的2.9μlmgso4、71.7μmsp2-寡聚物、1.8mmthpta、144μmeterneon-red645叠氮化物组成。孵育后,将样品短暂离心沉淀,并将上清液转移至新的小瓶中。将样品的等分试样在dh2o中稀释,然后通过分析型hplc分析。在配备有waters的xbridgetmostc18柱(2.5μm,4.6x50mm)的分析型hplcwatersalliance(e2695分离模块,2998光二极管阵列检测器)上进行分析型rp-hplc。使用1.5ml/分钟的流速和40℃的柱温,将以下梯度用于点击反应物的分离:8分钟内0-30%b,10分钟后85%b,11分钟内100%b。缓冲液a:0.1m乙酸三乙铵水溶液,ph=7,缓冲液b:0.1m乙酸三乙铵在80%(v/v)乙腈中的溶液,ph=7。将220-680nm的检测范围用于运行。除了来自反应器沉淀物的铜和配体以外,还使用mgso4作为添加剂,将炔烃修饰的寡核苷酸与eterneonred645叠氮化物反应。在mgso4存在的情况下,孵育60分钟后,对于粗反应物获得85%的点击产物转化率(图11)。实施例7.寡聚物-寡聚物点击的hpl色谱图在200μl反应小瓶中,将一个反应器沉淀物(600-800μm,包含元素铜)与6.7μl反应混合物组混合,并在45℃下孵育60分钟。反应混合物由dh2o和5%dmso(v/v)中的800μmthpta、20mmmgcl2、455μm烷烃寡聚物生物素和450μm叠氮化物寡聚物生物素组成。孵育后,将样品短暂离心沉淀,并将上清液转移至新的小瓶中。将样品的等分试样在dh2o中稀释,然后通过分析型hplc分析。在配备有来自waters的xbridgetmostc18柱(2.5μm,4.6x50mm)的分析型hplchplcwatersalliance(e2695分离模块,2998光二极管阵列检测器)上进行分析型rp-hplc。使用1.5ml/分钟的流速和40℃的柱温,将以下梯度用于点击反应物的分离:8分钟内0-30%b,10分钟后85%b,11分钟内100%b。缓冲液a:0.1m乙酸三乙铵水溶液,ph=7,缓冲液b:0.1m乙酸三乙铵在80%(v/v)乙腈中的溶液,ph=7。将220-680nm的检测范围用于运行。通过分析型hplc研究了单炔烃修饰的寡核苷酸与单叠氮化物修饰的寡核苷酸之间的反应,因为炔烃修饰的寡核苷酸的保留时间(rt=5.62分钟)与叠氮化物修饰的寡核苷酸(rt=7.64分钟)有很大差异。在45℃下60分钟后,几乎消耗了全部叠氮化物修饰的寡核苷酸。在6.10分钟和6.38分钟处观察到两个主要的新峰,其具有在随后的esi-ms中所需的点击产物的质量。由于稍微过量的炔烃修饰的寡核苷酸,在反应结束时(5.618分钟)观察到炔烃寡聚物的明显的峰。图12所示的积分峰的峰表。寡核苷酸y=peg12叠氮氨基dtx=c8-炔烃du生物素-teg序列表<110>贝瑟克里科有限公司<120>基于点击的连接<130>0048-pa-001cn<150>ep17194093<151>2017-09-29<150>ep18164071<151>2018-03-26<160>18<170>patentinversion3.5<210>1<211>59<212>dna<213>人工序列(artificialsequence)<220><223>炔烃寡聚物1<220><221>modified_base<222>(1)..(1)<223>5'-炔烃dt<400>1taatgatacggcgaccaccgagatctacactctttccctacacgacgctcttccgatct59<210>2<211>59<212>dna<213>人工序列(artificialsequence)<220><223>叠氮化物寡聚物1<220><221>modified_base<222>(1)..(1)<223>5'-叠氮化物dt<400>2tggagttcgtgaccgccgccgggatcactctcggcatggacgagctgtacaagtaaagc59<210>3<211>58<212>rna<213>人工序列(artificialsequence)<220><223>模型rna<400>3uucgacaaacgaaaacacaaacacaaaccaaacagaaaacaguacauguaaucgacca58<210>4<211>17<212>dna<213>人工序列(artificialsequence)<220><223>反向转录引物1<220><221>modified_base<222>(1)..(1)<223>5'-fam-dt(fam=荧光素)<400>4tggtcgattacatgtac17<210>5<211>23<212>dna<213>人工序列(artificialsequence)<220><223>反向转录引物2<400>5tggtcgattacatgtactgtttt23<210>6<211>17<212>dna<213>人工序列(artificialsequence)<220><223>反向转录引物3<400>6agatcggaagagcgtcg17<210>7<211>57<212>dna<213>人工序列(artificialsequence)<220><223>反向转录后的cdna<220><221>modified_base<222>(1)..(1)<223>5'-fam-dt(fam=荧光素)<220><221>modified_base<222>(57)..(57)<223>3'-叠氮化物da<400>7tggtcgattacatgtactgttttctgtttggtttgtgtttgtgttttcgtttgtcga57<210>8<211>116<212>dna<213>人工序列(artificialsequence)<220><223>点击产物<220><221>modified_base<222>(1)..(1)<223>5'-famdt(fam=荧光素)<220><221>misc_feature<222>(62)..(63)<223>通过主链模拟物b连接的a和t<400>8tggtcgattacatgtactgttttctgtttggtttgtgtttgtgttttcgtttgtcgataa60tgatacggcgaccaccgagatctacactctttccctacacgacgctcttccgatct116<210>9<211>116<212>dna<213>人工序列(artificialsequence)<220><223>pcr产物<400>9tggtcgattacatgtactgttttctgtttggtttgtgtttgtgttttcgtttgtcgataa60tgatacggcgaccaccgagatctacactctttccctacacgacgctcttccgatct116<210>10<211>51<212>dna<213>人工序列(artificialsequence)<220><223>图6预期的序列<400>10gttttctgtttggtttgtgtttgtgttttcgtttgtcgataatgatacggc51<210>11<211>51<212>dna<213>人工序列(artificialsequence)<220><223>图6实测的序列<400>11gttttctgtttggtttgtgtttgtgttttcgtttgtcgataatgatacggc51<210>12<211>21<212>dna<213>人工序列(artificialsequence)<220><223>图6正向序列,平均qv=43<400>12gtttgtcgataatgatacggc21<210>13<211>40<212>dna<213>人工序列(artificialsequence)<220><223>illuminan6引物<220><221>misc_feature<222>(35)..(40)<223>n为a、c、g或t<400>13gtgactggagttcagacgtgtgctcttccgatctnnnnnn40<210>14<211>59<212>dna<213>人工序列(artificialsequence)<220><223>炔烃适体寡核苷酸<220><221>modified_base<222>(1)..(1)<223>5'-炔烃dt<400>14taatgatacggcgaccaccgagatctacactctttccctacacgacgctcttccgatct59<210>15<211>20<212>dna<213>人工序列(artificialsequence)<220><223>p17-006正向引物<400>15gtgactggagttcagacgtg20<210>16<211>21<212>dna<213>人工序列(artificialsequence)<220><223>p17-006反向引物<400>16gtcgtgtagggaaagagtgta21<210>17<211>30<212>dna<213>人工序列(artificialsequence)<220><223>炔低聚生物素<220><221>modified_base<222>(1)..(1)<223>5?biotin-teg-dg<220><221>modified_base<222>(26)..(26)<223>c8-alkynedu<400>17gttctagaaggctaagaaaaatctcuacca30<210>18<211>30<212>dna<213>人工序列(artificialsequence)<220><223>叠氮化物寡生物素<220><221>modified_base<222>(1)..(1)<223>5biotin-teg-dg<220><221>modified_base<222>(26)..(26)<223>peg12叠氮化物氨基dt<400>18gttctagaaggctaagaaaaatctctacca30当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1